The site of action of the ichthyosis locus (ic) in the mouse, as ...
7
/. Embryol. exp. Morph. Vol. 32, 3, pp. 715-721, 1974 715 Printed in Great Britain The site of action of the ichthyosis locus (ic) in the mouse, as determined by dermal-epidermal recombinations BY MARGARET C. GREEN, 1 BARBARA N. ALPERT 1 AND THOMAS C. MAYER 2 From the Jackson Laboratory, Bar Harbor, and Department of Biology, Rider College This paper is dedicated to Professor H. Griineberg, F.R.S., on his retire- ment from the Chair of Animal Genetics at University College London. SUMMARY Skin of 14-day icjic and + / + embryos was separated into dermal and epidermal layers using trypsin, and recombined in the combinations + / + epidermis with + / + dermis, +/ + epidermis with icfic dermis, icjic epidermis with + / + dermis, and icjic epidermis with ic/ic dermis. The recombined tissues were implanted in adult histocompatible testes and allowed to grow for 2 weeks. The type of hair that developed in the grafts corresponded to the geno- type of the epidermis, and was not influenced by the genotype of the dermis. We conclude that the ic locus acts in the epidermis. INTRODUCTION The recessive mutant gene ichthyosis (ic) in the mouse causes a short sparse coat (Carter & Phillips, 1950). Spearman (1960) found no reduction in number of hair follicles in icjic mice, but the hairs were much thinner and shorter than normal and the air spaces were irregular in shape. All hairs were wavy and tended to vary in thickness along their length because of alterations in size of the medulla. In skin from the back both the hair follicles and the epidermis appeared normal, even though the hairs were abnormal, but on the tail the epidermis was thicker than normal and showed hyperkeratinization. The ichthyosis locus apparently controls some process necessary for the formation of normal hair. Previous results give no evidence on the question of the site of action of this locus, whether in the epidermis, the dermis, or possibly both or neither. The technique for separating skin from mouse embryos into dermal and epidermal components and making recombinations of components of different genotypes (Mayer & Fishbane, 1972; Mayer, 1973) allowed us to 1 Authors' address: The Jackson Laboratory, Bar Harbor, Maine 04609, U.S.A. 2 Author's address: Department of Biology, Rider College, Trenton, N.J. 08602, U.S.A.
Transcript of The site of action of the ichthyosis locus (ic) in the mouse, as ...

/ . Embryol. exp. Morph. Vol. 32, 3, pp. 715-721, 1974 7 1 5
Printed in Great Britain
The site of action of theichthyosis locus (ic) in the mouse, as determined
by dermal-epidermal recombinations
BY MARGARET C. GREEN,1 BARBARA N. ALPERT1
AND THOMAS C. MAYER2
From the Jackson Laboratory, Bar Harbor, andDepartment of Biology, Rider College
This paper is dedicated to Professor H. Griineberg, F.R.S., on his retire-ment from the Chair of Animal Genetics at University College London.
SUMMARY
Skin of 14-day icjic and + / + embryos was separated into dermal and epidermal layersusing trypsin, and recombined in the combinations + /+ epidermis with + /+ dermis, +/ +epidermis with icfic dermis, icjic epidermis with + /+ dermis, and icjic epidermis with ic/icdermis. The recombined tissues were implanted in adult histocompatible testes and allowedto grow for 2 weeks. The type of hair that developed in the grafts corresponded to the geno-type of the epidermis, and was not influenced by the genotype of the dermis. We conclude thatthe ic locus acts in the epidermis.
INTRODUCTION
The recessive mutant gene ichthyosis (ic) in the mouse causes a short sparsecoat (Carter & Phillips, 1950). Spearman (1960) found no reduction in numberof hair follicles in icjic mice, but the hairs were much thinner and shorter thannormal and the air spaces were irregular in shape. All hairs were wavy andtended to vary in thickness along their length because of alterations in size ofthe medulla. In skin from the back both the hair follicles and the epidermisappeared normal, even though the hairs were abnormal, but on the tail theepidermis was thicker than normal and showed hyperkeratinization.
The ichthyosis locus apparently controls some process necessary for theformation of normal hair. Previous results give no evidence on the question ofthe site of action of this locus, whether in the epidermis, the dermis, or possiblyboth or neither. The technique for separating skin from mouse embryos intodermal and epidermal components and making recombinations of componentsof different genotypes (Mayer & Fishbane, 1972; Mayer, 1973) allowed us to
1 Authors' address: The Jackson Laboratory, Bar Harbor, Maine 04609, U.S.A.2 Author's address: Department of Biology, Rider College, Trenton, N.J. 08602, U.S.A.

716 M. C. GREEN, B. N. ALPERT AND T. C. MAYER
investigate this question. We found that the primary effect of the ic locus occursin the epidermis, and that icjic dermis is indistinguishable from normal dermisin its effect on hair development.
MATERIALS AND METHODS
Mice homozygous for ichthyosis {icjic) were obtained from an inbred linemaintained by brother-sister matings of icj-\- x icjic mice for approximately22 generations. Mutant embryos were obtained from icjic x icjic matings.Homozygous normal mice were derived from the same inbred line by inter-crossing icj + mice and testing the + / ? offspring for absence of ic. In order toproduce enough + / + mice for the experiment the homozygotes ( + / + ) wereoutcrossed to mice of the C57BL/6J strain, producing + / + Fx hybrids. Normalembryos were obtained from Fx x Fx matings. Male Fx mice served as histo-compatible hosts for tissue from both types of embryos.
Skin was obtained from 14-day icjic and + / + embryos, separated into dermaland epidermal layers, and recombined in all four possible combinations accord-ing to the methods described by Mayer, Mittelberger & Green (1974). Therecombined tissues were grafted into the testes of adult males and allowed todevelop for 2 weeks, after which the testes were removed, fixed in 10 % formalin,and cleared in oil of wintergreen. The skin grafts were removed and examinedfor hair type and structure.
Two types of control procedures were carried out in order to determine theeffectiveness of the separation procedures. First, pieces of dermis and epidermiswere fixed immediately after separation, sectioned, and examined for presenceof contaminating cells. No contaminating cells were found. Secondly, isolateddermis and epidermis were grafted alone into adult testes and allowed todevelop for 14 days. No hair follicles developed in six such grafts of epidermisalone or in four of dermis alone.
RESULTS
From a total of 121 original grafts, 94 were recovered for study. Two of thesehad sharply contrasting sections of hair types and apparently were mosaicsattributable to incomplete separation of embryonic skin layers. They were notincluded in the results. The observations on the 92 remaining grafts are shown inTable 1.
All the grafts possessed numerous hairs arranged either parallel or in a ball.Since the inbred ichthyosis line was homozygous for agouti many of the hairscontained much yellow pigment which made them a little more difficult toclassify than hairs containing only black pigment. In order to eliminate sub-jective judgements as much as possible, we scored the grafts on three morpho-logical criteria: (1) overall appearance of the graft, including the degree ofcrowding as well as the alignment of hair shafts; (2) appearance of the most

Tab
le 1
. D
istr
ibut
ion
of g
raft
s sc
ored
on
a sc
ale
of 1
-4 f
or
thre
e m
orph
olog
ical
cri
teri
a
See
text
for
def
initi
on o
f sc
ores
.
Scor
est
Sign
ifica
nce
of d
iffer
ence
s
Com
bina
tion
s*
No.
of
graf
tsC
ateg
orie
sco
mbi
ned
x-O
vera
ll ap
pear
ance
A.
B.
C.
D.
+ / +
+ / +
iciic
ic/ic
+ 1
+ic
iicic
iic
16 26 36 14
App
eara
nce
of in
divi
dual
hai
rA
.
B.
C.
D.
+ / +
+ / +
iciic
iciic
+ 1
+ic
iicic
iic
16 26 36 14
Pigm
ent
dist
ribu
tion
A.
B.
C.
D.
* i
+ /
+
+ /
+ic
iicic
iic+
1 +
iciic
iciic
Epi
derm
is
16 26 35 14
0 0 19 11 0 0 6 4 0 0 0 0
0 0 14 3 A +
B v
s.
0 0 28 9 A +
B v
s.
0 0 4 0 A +
B v
s.
0C
+ D 2 5 0 1
C +
D 1 0 12 6
C +
D
2, 2 0 15 26 19
1,2,
3, v
s. 4
1,2,
3 v
s. 4
0-32
1 vs
. 2, 3
, 4
2-79
1,2,
3 v
s. 4
56
-31
1,2
, 3
vs.
4
0-3
2
1 v.
2, 3
, 4
0-89
1,2,
3 v
s. 4
59
-75
1-66
1,2,
3 v
s. 4
0
03
1,2,
3 v
s. 4
21
-64
> 0
05
> 0
05
> 00
1
>
005
>
005
> 001
> 00
5
>
005
> 001
=5- -I- S'
t Se
e R
esul
ts,
p.
718.
Der
mis

718 M. C. GREEN, B. N. ALPERT AND T. C. MAYER
common type of hair in each graft, judging both the size and shape of the hairs;and (3) pigment distribution in the hair shafts. A scale of 1-4 was used asfollows:
Overall appearance of graft:
(1) Few hairs in disorderly arrangement.(2) Relatively crowded hairs in disorderly arrangement.(3) Somewhat crowded hairs in only slightly disordered arrangement.(4) Crowded hairs in parallel arrangement.
Appearance of individual hairs:
(1) Short, narrow, curled.(2) Medium long, narrow, somewhat wavy.(3) Short, narrow, straight, and tapering, or long, broad, slightly bent, and
tapering.(4) Long, broad, straight, and tapering.
Pigment distribution:
(1) No pigment.(2) Granular, unevenly distributed pigment.(3) Slightly more even distribution of pigment.(4) Evenly distributed pigment.
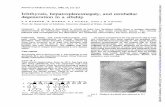
All scoring was done blind. Table 1 gives the frequencies and mean scores ofthe four combinations for the three criteria. Typical grafts of the four combina-tions are shown in Figures 1-4. The recombination grafts with + / + epidermisand icjic dermis resembled the grafts in which both layers were + / + (Figs. 1, 2).The recombination grafts with icjic epidermis and + / + dermis resembledgrafts in which both layers were icjic (Figs. 3, 4).
The significance of the difference in frequency of scores between control andrecombination grafts was tested by x2 (Table 1). There were no significantdifferences between the combinations with the same type of epidermis for allthree criteria. However, when combinations with + / + epidermis were combinedand compared with those with icjic epidermis (A + B vs. C + D, Table 1), thedifferences were highly significant for all three criteria. Thus the genotype of theepidermis determines the type of hair produced.
DISCUSSION
Our results demonstrate that the gene at the ic locus acts in the skin and morespecifically in the epidermal portion of the skin. However, they do not demon-strate conclusively that the activity of the locus is confined to the epidermis.The defect caused by the ic allele is demonstrable in our system only when thatallele is present in the epidermis. But we have not ruled out the possibility that

Ichthyosis locus (ic) in the mouse 719
\
Figs. 1-4. Typical grafts of four combinations of + /+ and icjic epidermis anddermis. (1) + /+ epidermis, + /+ dermis. (2) + /+ epidermis, \c\ic dermis. (3) ic\icepidermis, + /+ dermis. (4) icjic epidermis, ic/ic dermis. x 80.
the wild-type allele has some function in the dermis also, and that the loss ofsuch a function might be revealed by a mutant allele more severely defective thanic. In a similar experiment designed to determine the site of action of the agoutilocus, Mayer & Fishbane (1972) found that the substitution of nonagouti (a)for white bellied agout (AW~J) was effective in determining the colour pattern

720 M. C. GREEN, B. N. ALPERT AND T. C. MAYER
of hair only when present in the dermis, but Poole (1974) found that when thealleles compared were lethal yellow (Av) and a, an effect of gene substitution onpigmentation could be demonstrated in both dermis and epidermis. Possiblythe agouti locus is active in both parts of the skin. While some pairs of allelesat this locus differ in their effect on both layers, other pairs differ in effect onlyon dermis.
Mutant genes at well over 30 loci cause abnormal hair form or texture in themouse (Green, 1966). For most of them it is not known whether the site ofaction is in the skin or elsewhere. In principle this information could be rathereasily obtained by reciprocal skin transplantations if suitable congenic strainswere available. We are aware of only two mutants for which this experiment hasbeen performed. Reed (1938) made reciprocal skin transplants between newbornwaved-2 (wa-2/wa-2) mice and their normal sibs and found that the grafts pro-duced hair according to their own genotype, thus demonstrating that the siteof action of the wa-2 locus was in the skin. Fraser (1946) made reciprocal skintransplants between newborn rhino (hrrhlhrrh) and normal litter-mates andsimilarly found that the centres of the grafts behaved according to their owngenotype in all cases. Moreover, in grafts of rhino skin, which is normallyhairless, to normal hosts, some normal hair the colour of the rhino donordeveloped around the edges of the grafts, and in grafts of normal skin to rhinohosts a narrow hairless strip developed around the edges. A possible interpreta-tion of this result is that the hr locus acts in the dermis and that host dermisgrowing in under the graft epidermis supported normal growth of hair whenthe invading dermis was normal and suppressed it when the invading dermiswas rhino. Of the other hair-defect mutants, two in addition to ic have beeninvestigated by the dermal-epidermal recombination technique (downless, dl,Sofaer, 1973; fuzzy, fz, Mayer et al. 1974), and all three have been shown to actin the epidermis.
The use of the dermal-epidermal recombination technique is restricted tothose mutants that produce an abnormality of the hair recognizable in theresulting grafts. Grafts can be grown in the embryonic chick coelom as well asin the adult mouse testis, but in both these sites the conditions of hair growthmay be somewhat abnormal and not suitable for the detection of moderatedifferences in waviness, texture or length of hair, or in abnormalities of themolting cycle. In a small experiment to test the suitability of naked (N), amutation that causes the hairs to break off a little above the surface of the skinbeginning at 10-14 days of age, we grafted intact pieces of skin from eight14-day embryos to the testes of adult males and allowed them to remain for2 weeks. Half of the grafts should have been naked (N/ + ) and half normal( + /+ ) , but we were unable to detect any differences among them. The proba-bility that all would have been Nf + or that all would have been + / + is 1/128.Possibly an effect of N might have been demonstrable if the grafts had beenmaintained longer. Undoubtedly, a more extensive search would reveal mutants

Ichthyosis locus (ic) in the mouse 721other than the three so far studied to which the dermal-epidermal recombinationtechnique could be applied for determining the site of gene action.
For research aimed at discovering how genes act in controlling morphogenesis,genes affecting hair growth may be a favourable system. There are many suchgenes, the skin is readily accessible for experimentation, and the hair folliclesundergo cycles of activity and regression. Determining the site of action of thegenes involved is a necessary step preliminary to further study of this system.
We are grateful to Mrs Hope O. Sweet for her diligence in producing the mice for thisexperiment.
Supported in part by NSF grant GB 27487 and ACS grant VC-17-L, a Florence M.Kerrison Memorial Grant for Cancer Research in Maine to the Jackson Laboratory, andby NSF grant GB 18271 to Rider College. The Jackson Laboratory is fully accredited by theAmerican Association for Accreditation of Laboratory Animal Care.
One of us (B.N. A.) is a participant in the Jackson Laboratory Training Program supportedby training grant GW 8212 from the Student Science Training Program of the NationalScience Foundation.
REFERENCES
CARTER, T. C. & PHILLIPS, R. S. (1950). Ichthyosis, a new recessive mutant in the housemouse. / . Hered. 41, 297-300.
FRASER, F. C. (1946). The expression and interaction of hereditary factors producing hypo-trichosis in the mouse: histology and experimental results. Can. J. Res. 24, 10-25.
GREEN, M. C. (1966). Mutant genes and linkages. In Biology of the Laboratory Mouse (ed.E. L. Green), pp. 87-150. New York: McGraw-Hill.
MAYER, T. C. (1973). The migratory pathway of neural crest cells into the skin of mouseembryos. Devi Biol. 34, 39^6.
MAYER, T. C. & FISHBANE, J. L. (1972). Mesoderm-ectoderm interaction in the productionof the agouti pigmentation pattern in mice. Genetics, Princeton 71, 297-303.
MAYER, T. C, MITTELBERGER, J. A. & GREEN, M. C. (1974). The site of action of the fuzzylocus (fz) in the mouse, as determined by dermal-epidermal recombinations. / . Embryo!.Exp. Morphol. 32, 707-713.
POOLE, T. W. (1974). Dermal-epidermal interactions and the site of action of the yellow{Av) and nonagouti (a) coat color genes in the mouse. Devi Biol. 36, 208-211.
REED, S. C. (1938). Determination of hair structure. I. The production of waved hair fromgenetically non-waved cells. / . exp. Zool. 79, 347-354.
SOFAER, J. A. (1973). Hair follicle initiation in reciprocal recombinations of downless homo-zygote and heterozygote mouse tail epidermis and dermis. Devi Biol. 34, 289-296.
SPEARMAN, R. I. (1960). The skin abnormality of 'ichthyosis', a mutant of the house mouse.J. Embryol. exp. Morph. 8, 387-395.
{Received 12 March 1974)