
Mutations in the BRCA1 and BRCA2 breast cancer susceptibility genes

ABCDEFG
UNIVERS ITY OF OULU P.O.B . 7500 F I -90014 UNIVERS ITY OF OULU F INLAND
A C T A U N I V E R S I T A T I S O U L U E N S I S
S E R I E S E D I T O R S
SCIENTIAE RERUM NATURALIUM
HUMANIORA
TECHNICA
MEDICA
SCIENTIAE RERUM SOCIALIUM
SCRIPTA ACADEMICA
OECONOMICA
EDITOR IN CHIEF
PUBLICATIONS EDITOR
Senior Assistant Jorma Arhippainen
University Lecturer Santeri Palviainen
Professor Hannu Heusala
Professor Olli Vuolteenaho
University Lecturer Hannu Heikkinen
Director Sinikka Eskelinen
Professor Jari Juga
Professor Olli Vuolteenaho
Publications Editor Kirsti Nurkkala
ISBN 978-951-42-9991-9 (Paperback)ISBN 978-951-42-9992-6 (PDF)ISSN 0355-3221 (Print)ISSN 1796-2234 (Online)
U N I V E R S I TAT I S O U L U E N S I S
MEDICA
ACTAD
D 1184
ACTA
Pasi Eskola
OULU 2012
D 1184
Pasi Eskola
THE SEARCH FOR SUSCEPTIBILITY GENESIN LUMBAR DISC DEGENERATIONFOCUS ON YOUNG INDIVIDUALS
UNIVERSITY OF OULU GRADUATE SCHOOL;UNIVERSITY OF OULU, FACULTY OF MEDICINE,INSTITUTE OF BIOMEDICINE,DEPARTMENT OF MEDICAL BIOCHEMISTRY AND MOLECULAR BIOLOGY;INSTITUTE OF CLINICAL MEDICINE,DEPARTMENT OF PHYSICAL MEDICINE AND REHABILITATION;BIOCENTER OULU;CENTER FOR CELL-MATRIX RESEARCH


A C T A U N I V E R S I T A T I S O U L U E N S I SD M e d i c a 1 1 8 4
PASI ESKOLA
THE SEARCH FOR SUSCEPTIBILITY GENES IN LUMBAR DISC DEGENERATIONFocus on young individuals
Academic dissertation to be presented with the assentof the Doctoral Training Committee of Health andBiosciences of the University of Oulu for public defencein Auditorium F101 of the Department of Physiology(Aapistie 7), on 30 November 2012, at 12 noon
UNIVERSITY OF OULU, OULU 2012

Copyright © 2012Acta Univ. Oul. D 1184, 2012
Supervised byDocent Minna MännikköProfessor Jaro Karppinen
Reviewed byDocent Elisabeth WidénSenior Lecturer Frances M. K. Williams
ISBN 978-951-42-9991-9 (Paperback)ISBN 978-951-42-9992-6 (PDF)
ISSN 0355-3221 (Printed)ISSN 1796-2234 (Online)
Cover DesignRaimo Ahonen
JUVENES PRINTTAMPERE 2012

Eskola, Pasi, The search for susceptibility genes in lumbar disc degeneration. Focuson young individualsUniversity of Oulu Graduate School; University of Oulu, Faculty of Medicine, Institute ofBiomedicine, Department of Medical Biochemistry and Molecular Biology; Institute of ClinicalMedicine, Department of Physical Medicine and Rehabilitation; Biocenter Oulu; Center forCell-Matrix Research, P.O. Box 5000, FI-90014 University of Oulu, FinlandActa Univ. Oul. D 1184, 2012Oulu, Finland
Abstract
Low back pain (LBP) is a truly enervating condition, presenting with considerable negativesocioeconomic and health impacts on many levels. Although LBP may be attributed to manyfactors, there is increasing evidence that disc degeneration (DD) of the lumbar spine is a strongcontributing factor, especially among young individuals. Understanding the aetiopathogenesis ofDD has changed over the past few decades as numerous studies have indicated that inheritedfactors are largely responsible for the development of DD. Despite the many genetic associationsin DD that have been reported, these associations have proven difficult to validate. The geneticcomponent of DD is still unexplained for the most part. Previous studies have focused on adults,who have been exposed to environmental risk factors of DD, which may mask geneticassociations. Thus, investigations among young individuals are well justified.
The purpose of this study was to validate the associations between DD defined by magneticresonance imaging (MRI) and genetic polymorphisms in two study populations of Finnish andDanish young individuals. New polymorphisms that have not been associated with DD were alsoincluded in the study. Associations with progression of DD were also investigated among Danishindividuals. Furthermore, this study aimed to clarify the level of evidence of previously identifiedassociations.
Among Finnish individuals, polymorphisms in IL6, SKT and CILP were associated withmoderate DD, but the association in CILP was significant only in women. Among Danishindividuals, polymorphisms in IL6 and IL1A were associated with early DD and progression ofDD. The genetic associations among the Danish teenagers were gender-specific as they weremainly observed in girls. Among both populations, the individuals with DD were significantlytaller than individuals without DD. In the systematic analysis of previous reports, polymorphismsin GDF5, THBS2, MMP9, COL11A1, SKT and ASPN were found to have a moderate level ofassociation evidence.
The results mostly support earlier findings in adults. However, unexpected differences betweengenders were observed. In conclusion, this study increased the knowledge of genetics in DD butmore investigations are needed to draw any solid conclusions.
Keywords: adolescent, child, genetics, intervertebral disc, intervertebral discdegeneration, sex


Eskola, Pasi, Lannerangan varhaisen välilevyrappeuman alttiusgeenit. Oulun yliopiston tutkijakoulu; Oulun yliopisto, Lääketieteellinen tiedekunta, Biolääketieteenlaitos, Lääketieteellinen biokemia ja molekyylibiologia; Kliinisen lääketieteen laitos, Fysiatria;Biocenter Oulu; Center for Cell-Matrix Research, PL 5000, 90014 Oulun yliopistoActa Univ. Oul. D 1184, 2012Oulu
Tiivistelmä
Alaselkäkipu on yksi merkittävimmistä toimintakykyyn vaikuttavista sairauksista. Alaselkäki-vun riskitekijöitä tunnetaan useita, mutta näyttö siitä, että selän välilevyrappeuma on yksi tär-keimmistä riskitekijöistä, erityisesti nuorilla henkilöillä, lisääntyy jatkuvasti. Aiemmat tutkimuk-set ovat osoittaneet, että perintötekijöillä on merkittävä osuus välilevyjen rappeutumisessa. Usei-den geenien on raportoitu vaikuttavan välilevyrappeumaan. Positiivisten tulosten toistaminen onkuitenkin osoittautunut vaikeaksi, ja kokonaiskuva on vielä suurelta osin epäselvä. Aikaisem-mat tutkimukset aiheesta ovat keskittyneet aikuisiin, joilla nuoria pidempi altistus ympäristöteki-jöille saattaa peittää geneettisten tekijöiden yhteyttä.
Tämän tutkimuksen tavoitteena oli varmentaa yhteyksiä magneettikuvauksella (MRI) määri-tetyn välilevyrappeuman ja geneettisten monimuotoisuuskohtien (polymorfioiden) välillä käyttä-en yhtä suomalaisista ja yhtä tanskalaisista nuorista koostuvaa aineistoa. Työssä analysoitiinmyös polymorfioita, joita ei ole aiemmin yhdistetty välilevyrappeumaan. Yhteyttä polymorfioi-den ja välilevyrappeuman etenemisen välillä tutkittiin tanskalaisessa aineistossa. Lisäksi tutki-muksen tavoitteena oli selvittää aiemmin tunnistettujen geneettisten yhteyksien näytön aste.
Suomalaisten nuorten aineistossa polymorfiat IL6-, SKT- ja CILP-geeneissä olivat yhteydes-sä kohtalaista astetta olevaan välilevyrappeumaan. Tanskalaisten nuorten aineistossa polymorfi-at IL6- ja IL1A-geeneissä olivat yhteydessä varhaiseen välilevyrappeumaan ja sen etenemiseen.Tanskalaisten nuorten aineistossa nämä yhteydet olivat sukupuolesta riippuvaisia, koska yhteyk-siä havaittiin pääasiassa tytöillä. Suomalaisten nuoren aineistossa yhteys CILP-geenissä havait-tiin ainoastaan nuorilla naisilla. Kummassakin tutkimusaineistossa nuoret, joilla oli välilevyrap-peuma, olivat pidempiä, kuin nuoret, joilla ei ollut välilevyrappeumaa. Aikaisempien tutkimus-ten systemaattinen analyysi osoitti, että tunnistetun geneettisen yhteyden näytön aste on kohta-lainen GDF5-, THBS2-, MMP9-, COL11A1-, SKT- ja ASPN-geenien polymorfioissa.
Tutkimusten tulokset ovat pääosin samansuuntaisia aiempien tutkimuksien kanssa, mutta nytnähty eroavaisuus sukupuolten välillä on uusi havainto. Kokonaisuudessaan tämä väitöstutki-mus kasvattaa ymmärtämystä välilevyrappeumasta, mutta lisätutkimuksia aiheesta tarvitaan.
Asiasanat: lapset, nikamavälilevy, nikamavälilevyn rappeuma, nuoret,perinnöllisyystiede, sukupuoli


To my family

8

9
Acknowledgements
This thesis was carried out at the Oulu Center for Cell-Matrix Research, at the
Department of Medical Biochemistry and Molecular Biology, Institute of
Biomedicine, University of Oulu, at the Department of Physical Medicine and
Rehabilitation, Institute of Clinical Medicine, University of Oulu and at the
Institute of Sports Science and Clinical Biomechanics, Part of Clinical
Locomotion Network, University of Southern Denmark, during the years 2006–
2012.
I owe deep gratitude to my principal supervisor, Docent Minna Männikkö for
her guidance and support. This work would not have been possible without her
sound knowledge of human genetics. I´m thankful for all those important issues
of scientific work she has taught me. Her pleasant way of leading and her care for
the well being of our research group has created an enjoyable and stimulating
atmosphere for this research.
I could not have had a better place to observe a truly dedicated scientist at
work than I have had observing my other supervisor, Professor Jaro Karppinen.
He has shown how hard work pays off and made me again remember that setting
an example is the most important way of leading. His continuous support,
optimism and 24/7 availability have made my job a lot easier.
The excellent research infrastructure and high-standard scientific
environment are an outgrowth of persevering efforts. I am grateful that I have had
the opportunity to carry out my thesis at the Department of Medical Biochemistry
and Molecular Biology. It is not a coincidence that our department has educated
more scientists than any other department in our university. I deeply respect
Research Professor Emeritus Ilmo Hassinen, Research Professor Emeritus Kari
Kivirikko, Professor Taina Pihlajaniemi, Professor Johanna Myllyharju, Professor
Seppo Vainio and Professor Peppi Karppinen for their scientific work.
I am grateful to Docent Elisabeth Widén and Senior Lecturer Frances M.K.
Williams for their excellent and accurate comments on this thesis. PhD Deborah
Kaska is acknowledged for her careful language revision of the manuscript.
Professor Antero Kesäniemi, Professor Peppi Karppinen, PhD Tomi Hillukkala,
PhD Antti Salo, PhD Pekka Kilpeläinen and PhD Anthony Heape are thanked for
participating in the thesis progress follow-up group.
The original articles in this thesis have required the knowledge and skills of
many investigators. I wish to thank all my co-authors and collaborators for their
contributions to the original articles. All the members of our research group, both

10
past and present, are warmly thanked. There has not been a single day without a
smile with you guys! Time spent in the lab would have been exponentially higher
without the expert assistance of Aira Erkkilä, Helena Satulehto, Niina Heikkinen,
Minta Lumme and Irma Vuoti. Helena and Niina deserve my special thanks.
Other members of the Department have also contributed to the very pleasant
working atmosphere. Auli Kinnunen, Pertti Vuokila, Seppo Lähdesmäki, Risto
Helminen and Marja-Leena Karjalainen are thanked for all the help in practical
matters. Jani Takatalo and Markus Paananen are also acknowledged for their
thoughts and opinions on the research work. Also Mira Pesälä and Petri Toivola
are thanked for their flexibility at a critical moment.
I owe my gratitude to the colleagues Iita Daavittila and Niko Paalanne for
their company. Iita deserves the credit for teaching me key skills at the beginning
the thesis work. The friendship of colleagues Juha Auvinen, Rami Kankaanpää
and Jussi Mäkelä have made all the challenges during the thesis process a lot
easier to endure. I am also indebted to every brother in "Kartat Taskuun”,
everybody in “SK Mortis” and to all my other friends and colleagues for their
support and company during the too few recreational and relaxing moments. Raija
and Veikko Nissi as well as all the members of your family are warmly thanked.
I wish to express my warmest gratitude to my father Timo and mother Raili,
for all the things you have taught me. You have always encouraged and supported
me in every challenge I have chosen to take. My brother Matti has also always
been there for me at any hour. I am grateful also for all the other close members
of my family.
Finally, I have the deepest and most personal gratitude towards my lovely
companion in life. Johanna, without you this work would not have been
completed. Thank you for bearing all the time I have needed to take away from
you for the research. Without you, the radiant smile on our lively Tytti´s face
would be something totally different. You remind me every day about the most
important things in life.
This work was supported by grants from the University of Oulu Scholarship
Foundation, the University of Oulu Research Council, the Oulu University
Pharmacy Foundation, the Päivikki and Sakari Sohlberg Foundation, the Emil
Aaltonen Foundation, the Oskar Öflunds Stiftelse, the Finnish Medical
Foundation Duodecim, the Finnish-Norwegian Medical Foundation and the
AOSpine Europe.
Oulu, October 2012 Pasi Eskola

11
Abbreviations
A adenine
ACAN aggrecan
ADAMTS a disintegrin and metalloprotease with thrombospondin type 1
motifs
AF annulus fibrosus
ALL anterior longitudinal ligament
An allele representing with n number of repeats
ASPN asporin
bFGF basic fibroblast growth factor
BMD bone mineral density
BMI body mass index
BMP bone morphogenetic protein
bp basepair
C cytosine
CI 95% confidence interval
CILP cartilage intermediate layer protein
CNV copy number variation
COL collagen
CS chondroitin sulphate
CT computed tomography
DD lumbar intervertebral disc degeneration
DDP lumbar intervertebral disc degeneration progression
DNA deoxyribonucleic acid
DZ dizygotic
ECM extracellular matrix
EGF epidermal growth factor
EP vertebral endplate
FAS tumour necrosis factor receptor superfamily member 6
FASLG tumour necrosis factor ligand superfamily member 6
FDH Finnish disease heritage
FokI FokI restriction enzyme site polymorphism in VDR
G guanine
GDF5 growth differentiation factor 5
GWAS genome-wide association study
HA hyaluronan

12
HIZ high intensity zone
HWE Hardy-Weinberg equilibrium
IGF insulin-like growth factor
IL1A interleukin-1 alpha
IL6 interleukin-6
IVD intervertebral disc
KS keratan sulphate
L lumbar vertebra
LBP low back pain
LD linkage disequilibrium
LDD lumbar disc disease characterized by sciatica
LDH lumbar disc herniation; including different subtypes
LP link protein
M million
MAF minor allele frequency
MMP matrix metalloproteinase
MRI magnetic resonance imaging
MZ monozygotic
N number of individuals
NOS nitric oxide synthase
NP nucleus pulposus
OA osteoarthritis
OR odds ratio
PCR polymerase chain reaction
PDGF platelet-derived growth factor
PG proteoglycan
PLL posterior longitudinal ligament
S sacral vertebra
SKT human homolog KIAA1217 to the Skt
Skt small with kinky tail-gene in mouse
SLRP small-leucine-rich-proteoglycan
SNP single nucleotide polymorphism
STR short tandem repeat
T thymine
TaqI TaqI restriction enzyme site polymorphism in VDR
TGF transforming growth factor
Th thoracic vertebra

13
THBS2 thrombospondin 2
TIMP tissue inhibitor of metalloproteinase
TNF tumour necrosis factor
VB vertebral body
VC vertebral column
VNTR variable number of tandem repeats
-n molecule specification where n refers to receptor and/or subtype

14

15
List of original articles
This thesis is based in the following articles, which are referred to in the text by
their Roman numerals:
I Eskola PJ, Lemmelä S, Kjaer P, Solovieva S, Männikkö M, Tommerup N, Lind-Thomsen A, Husgafvel-Pursiainen K, Cheung KMC, Chan D, Samartzis D*, Karppinen J* (2012) Genetic association studies in lumbar disc degeneration: a systematic review. PLOS ONE. In press.
II Kelempisioti A*, Eskola PJ*, Okuloff A, Karjalainen U, Takatalo J, Daavittila I, Niinimäki J, Sequeiros RB, Tervonen O, Solovieva S, Kao PY, Song YQ, Cheung KM, Chan D, Ala-Kokko L, Järvelin MR, Karppinen J, Männikkö M (2011) Genetic susceptibility of intervertebral disc degeneration among young Finnish adults. BMC Med Genet 22(12): 153
III Eskola PJ, Kjaer P, Daavittila IM, Solovieva S, Okuloff A, Sorensen JS, Wedderkopp N, Ala-Kokko L, Männikkö M & Karppinen JI (2010) Genetic risk factors of disc degeneration among 12–14-year-old Danish children: a population study. Int J Mol Epidemiol Genet 1(2): 158–65
IV Eskola PJ, Kjaer P, Sorensen JS, Okuloff A, Wedderkopp N, Daavittila I, Ala-Kokko L, Männikkö M, Karppinen J (2012) Gender difference in genetic association between IL1A variant and early lumbar disc degeneration: a three-year follow-up. Int J Mol Epidemiol Genet 3(3): 195–204
* These authors contributed equally.

16

17
Contents
Abstract
Tiivistelmä
Acknowledgements 9 Abbreviations 11 List of original articles 15 Contents 17 1 Introduction 19 2 Review of the literature 21
2.1 Scope of the literature review ................................................................. 21 2.2 Human spine ........................................................................................... 21
2.2.1 Development ................................................................................ 21 2.2.2 Structure and function .................................................................. 22 2.2.3 Structure of healthy intervertebral disc ........................................ 23
2.3 Disc degeneration (DD) .......................................................................... 29 2.3.1 Structure of degenerated intervertebral disc ................................. 34 2.3.2 Changing the view of aetiology .................................................... 39 2.3.3 Clinical relevance and treatment .................................................. 44
2.4 Genetic studies of complex disorders ..................................................... 46 2.4.1 Variation in the human genome .................................................... 47 2.4.2 Approaches to genetic studies of complex disorders .................... 48
2.5 Finnish disease heritage .......................................................................... 56 2.6 Genetic studies in disc degeneration ....................................................... 56 2.7 Candidate gene studies in disc degeneration ........................................... 57
2.7.1 Vitamin D receptor ....................................................................... 58 2.7.2 Collagen IX genes ........................................................................ 60 2.7.3 Other collagen genes .................................................................... 63 2.7.4 Aggrecan ...................................................................................... 64 2.7.5 Cartilage intermediate layer protein ............................................. 65 2.7.6 Asporin ......................................................................................... 67 2.7.7 Matrix metalloproteinase genes .................................................... 68 2.7.8 Interleukin genes .......................................................................... 70 2.7.9 Findings in other genes ................................................................. 72
2.8 Linkage analysis studies in disc degeneration ......................................... 74 2.9 Genome-wide association studies in disc degeneration .......................... 75 2.10 Summary and research problem .............................................................. 76

18
3 Outlines of the present study 77 4 Materials and methods 79
4.1 Study populations .................................................................................... 80 4.1.1 Finnish study population (II) ........................................................ 80 4.1.2 Danish study population (III, IV) ................................................. 80 4.1.3 Study ethics .................................................................................. 81
4.2 MRI methodology ................................................................................... 81 4.2.1 Finnish young adult study (II) ...................................................... 81 4.2.2 Danish teenager studies (III, IV) .................................................. 82
4.3 Genetic analyses ...................................................................................... 83 4.4 Systematic review methods (I) ................................................................ 86
5 Results 91 5.1 Genetic association studies on MRI defined lumbar DD (I) ................... 91
5.1.1 Characteristics of studies and level of evidence ........................... 91 5.2 Associations between SNPs and lumbar DD on MRI among
Finnish young adults (II) ......................................................................... 95 5.2.1 IL6, SKT, CILP and DD................................................................ 96 5.2.2 Height, BMI and DD .................................................................... 98
5.3 Associations between SNPs and early lumbar DD and DDP
among Danish teenagers (III, IV) ............................................................ 99 5.3.1 IL1A, IL6 and early DD in girls .................................................... 99 5.3.2 IL1A and early DDP in girls ....................................................... 102 5.3.3 Height, weight and early DD (III, IV) ........................................ 102
6 Discussion 105 6.1 The amount of cumulative evidence is currently modest (I) ................. 105 6.2 IL6 SNPs and DD among young individuals (II, III, IV) ...................... 107 6.3 IL1A SNPs and DD among young individuals (II, III, IV) ................... 108 6.4 SKT and CILP SNPs and DD among Finnish young adults (II) ............ 110 6.5 Height, weight and DD among young individuals (II, III, IV) ............... 111 6.6 Future possibilities ................................................................................ 112
7 Conclusions 115 References 117 Appendices 151 List of original articles 187

19
1 Introduction
Low back pain (LBP) is a truly enervating condition presenting with considerable
socioeconomic and health impacts on many levels. LBP can lead to decreased
physical activity, lost wages and a decrease in quality of life. Epidemiological
studies have shown, in contrast to conventional wisdom, that LBP is common
already during childhood, not to speak of adolescence. Although LBP may be
attributed to many factors, there is increasing evidence that disc degeneration (DD)
of the lumbar spine is a strong contributing factor, especially among young
individuals.
Understanding the aetiopathogenesis of DD has changed over the past few
decades. Numerous studies have suggested that genetic factors are largely
responsible for the development of lumbar DD and that environmental factors
play a smaller role than previously believed. Based on twin studies, the
heritability of DD is estimated to 80%. This has led to the well-justified search for
specific genetic risk factors. However, similar to other complex disorders, the
genetic associations found in DD have proved difficult to validate. Furthermore,
the true natural course of DD is not fully elucidated.
Different changes in the intervertebral disc (IVD) have been used as markers
for DD. Early changes of DD may appear already during the teenage years while
more severe clinical phenotypes, such as sciatica and chronic LBP, often appear in
mature adulthood. It has previously been suggested that more efforts should be
concentrated on the study of early DD in order to increase our understanding of
the whole condition. It would also be highly beneficial to be able to screen for
individuals susceptible for symptomatic DD in a subclinical phase in order to
study possible interventions to prevent the painful and crippling clinical phases of
DD. However, previous information about the effects of genetic variations in DD
among young individuals is very limited and substantially more information is
required before any screening protocols are reasonable.
Information obtained from previous genetic studies is somewhat scattered and
the cumulative association evidence has not been reviewed previously. In the
present study the first extensive systematic review of these previously published
genetic association studies in DD was performed along with level of evidence
analyses. Furthermore, the roles of specific genetic markers in DD among Finnish
young adults and in DD and its progression (DDP) among Danish teenagers were
investigated. The purpose of this study was to increase information about DD,

20
focusing especially on young individuals, in order to facilitate future research
efforts.

21
2 Review of the literature
2.1 Scope of the literature review
The scope of this thesis includes basics of 1) the anatomy and development of the
human spine, 2) the concept of disc degeneration (DD), 3) the structure of healthy
and degenerated intervertebral discs, 4) the clinical relevance and treatment of
DD while 5) noting the studies indicating a heritable component in DD and
associated conditions. Additionally, 6) the special aspects of genetic studies in
complex disorders are also introduced, but the closest scrutiny is given to genetic
association studies in DD. Disc degeneration among young individuals is
emphasized when possible. Future possibilities are itemised. Animal studies with
intervertebral disc degeneration are generally excluded. This thesis focuses on the
lumbar intervertebral discs.
2.2 Human spine
The human spine is usually understood as the vertebral column. The back is
understood as the posterior part of the trunk between the neck and buttocks and it
includes also skin, subcutaneous tissue, ligaments, spinal cord and meninges, ribs
as well as various nerves and vessels (Moore 1999).
2.2.1 Development
The axial skeleton (the articulated bones of the skull, vertebral column, ribs and
sternum) is derived from the somites, structures of the developing embryo located
on both sides of the neural tube. These blocks of epithelial cells originate from the
paraxial mesoderm (Kornak & Mundlos 2003). During early development, a
flexible supportive structure is found in all chordate embryos. This notochord,
which is localized ventral to the neural tube and between the paraxial somites, is
vital to the development of intervertebral discs (IVDs) (Fleming et al. 2001). The
notochord induces paraxial somite cell differentiation into sclerotome. Sclerotome
cells then migrate to form a perinotochordal sheath that develops condensed and
uncondensed regions of mesenchyme (Aszodi et al. 1998). These regions adjacent
to the notochord act as the source of IVDs and vertebrae. A remnant of the
notochord is found in the central part of the IVDs, but not normally in the bony

22
vertebrae (Chan et al. 2011, Risbud & Shapiro 2011). Currently, there is no cell-
tracking data to confirm the origin of the outer parts of the IVD. However, it is
currently thought to originate from somites. Additionally, because some of the
data on IVD development are nearly two decades old and are based on anatomic
analyses, it has been recently suggested that these data of IVD development
should be confirmed using modern tools of molecular genetics (Chan et al. 2011).
2.2.2 Structure and function
The human spine is a movable complex structure that bears a great amount of the
body’s weight. The spine consists of vertebral bodies (VB), IVDs, ligaments and
adjacent muscles. Anatomically, a normal adult spine has a movable column
composed of 24 VB (Prescher 1998). Five sacral vertebrae are normally fused to
form the sacrum and another four form the coccyx hence a total of 33 VBs are
usually present. The 24 movable VBs in a typical adult VC can be further divided
based on location and anatomy; the first seven from the skull down are called
cervical (C), next twelve thoracic (Th) and the five above the sacrum are lumbar
(L). The VC extends from the skull to the tip of the coccyx and forms the skeleton
of the neck and back and most of the axial supportive structures. The VC protects
the spinal cord and nerves, supports the body’s weight, provides flexible yet
partly rigid axis for the body and a pivot for the head and also has an important
role in posture and locomotion (Moore 1999).
Approximately 25% of the vertebral column consists of fibrocartilaginous
IVDs that are located between the bony VBs. The VBs and IVDs are also
connected with each other by stabilizing ligaments (Figure 1). The main function
of the IVD is to work as a shock absorber for axial compressive loads. The
anterior longitudinal ligament (ALL) is a strong structure that maintains stability
between VC joints and takes part in preventing hyperextension of the VC. The
posterior longitudinal ligament (PLL) is a weaker and narrower structure
compared to the ALL. The PLL takes part in preventing hyperflexion of the VC
and herniation or posterior protrusion of the IVD (Moore 1999).
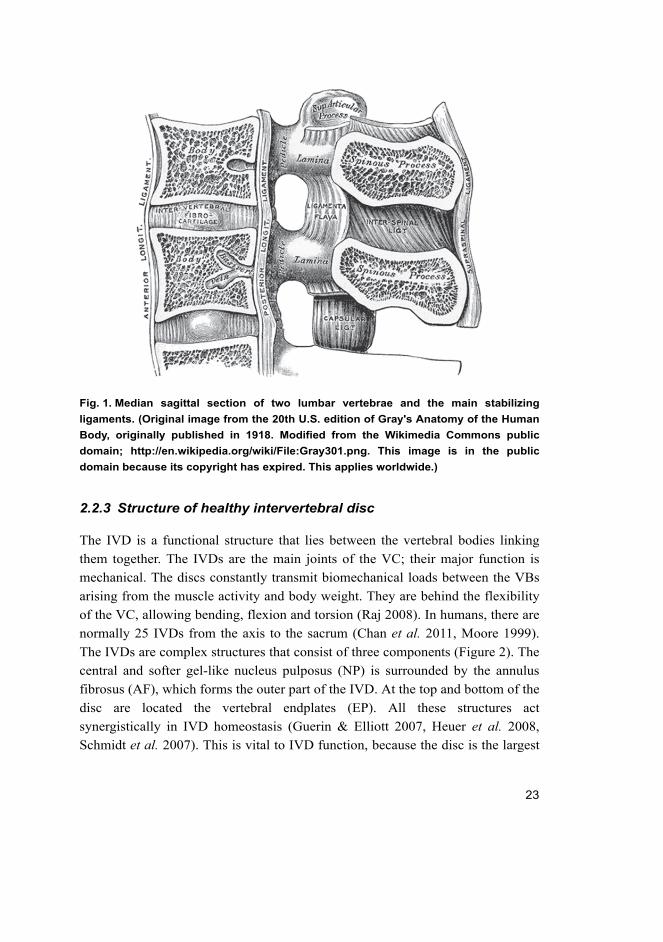
23
Fig. 1. Median sagittal section of two lumbar vertebrae and the main stabilizing
ligaments. (Original image from the 20th U.S. edition of Gray's Anatomy of the Human
Body, originally published in 1918. Modified from the Wikimedia Commons public
domain; http://en.wikipedia.org/wiki/File:Gray301.png. This image is in the public
domain because its copyright has expired. This applies worldwide.)
2.2.3 Structure of healthy intervertebral disc
The IVD is a functional structure that lies between the vertebral bodies linking
them together. The IVDs are the main joints of the VC; their major function is
mechanical. The discs constantly transmit biomechanical loads between the VBs
arising from the muscle activity and body weight. They are behind the flexibility
of the VC, allowing bending, flexion and torsion (Raj 2008). In humans, there are
normally 25 IVDs from the axis to the sacrum (Chan et al. 2011, Moore 1999).
The IVDs are complex structures that consist of three components (Figure 2). The
central and softer gel-like nucleus pulposus (NP) is surrounded by the annulus
fibrosus (AF), which forms the outer part of the IVD. At the top and bottom of the
disc are located the vertebral endplates (EP). All these structures act
synergistically in IVD homeostasis (Guerin & Elliott 2007, Heuer et al. 2008,
Schmidt et al. 2007). This is vital to IVD function, because the disc is the largest

24
avascular structure in the human body (Mirza & White 1995). The IVD is
composed mainly of collagens and proteoglycans (Table 1).
Fig. 2. Cut out portion of a normal disc. (Modified from Raj 2008. Reprinted with
permission from John Wiley & Sons, Inc.)
Annulus fibrosus
The AF is made of ca. 15–25 lamellae that construct the discs circumference
(Marchand & Ahmed 1990, Moore 1999). The lamellae are arranged in a specific
criss-cross pattern to resist mechanical pressure and loading. They are subdivided
into inner fibres, which are connected to the cartilaginous EP and outer fibres
(Sharpey’s Fibres), which link the disc to the vertebral body (Martin et al. 2002).
The outer part of the annulus is made of mainly type I collagen together with
types III, V and VI (Eyre & Muir 1977, Nerlich et al. 1998). The amount of type
II collagen increases towards the inner parts of the AF where the structure
becomes less organized (Humzah & Soames 1988). Different collagen types work
cooperatively with each other forming a dynamic network (Engvall et al. 1986,
Schollum et al. 2009). Elastin fibres are also found in the AF. The outer part has a
higher elastin density and it is thought that elastin might function in protecting the

25
laminated structure and in recovery after structure deformation caused by
compressive loads (Yu et al. 2007). The boundaries between the inner and outer
AF as well as inner AF and NP become less clear with aging (Boos et al. 2002).
Nucleus pulposus
The soft gel-like NP fills the centre of a healthy IVD. From an in vitro study in
humans it is known that in the newborn, the NP is translucent and composed of ca.
87% water (Antoniou et al. 1996). The hydration level begins to fall thereafter
and it is ca. 80% in young human adults according to a histological study in
humans (Boos et al. 2002). The high water content is mainly due to hydrophilic
proteoglycans (PG), of which aggrecan is the most abundant (Roughley 2004).
The PGs are supported by randomly oriented type II collagen fibres, but also
smaller amounts of type XI and XI collagen are included in the NP structure.
Aggrecan has a bottle brush-like structure (Figure 3), composed of a core protein
and glycosaminoglycan side chains, more specifically chondroitin sulphate (CS)
and keratan sulphate (KS), side chains. Aggrecan is non-covalently bound to
hyaluronan (HA), and the complex is further stabilized by the link protein (LP).
The negatively charged side chains draw water via osmosis thus increasing the
internal IVD pressure, which makes the NP tolerant of compression from axial
loading (Whatley & Wen 2012). In addition to aggrecan and collagens, small-
leucine-rich-proteoglycans (SLRP), such as fibromobulin, decorin, lumican,
asporin and chondroadherin are also found in the NP. These components can
interact with collagen fibres and also may have an effect on extracellular matrix
(ECM) homeostasis via growth factors (Chan et al. 2011).
Endplate
The third clearly distinct component of the disc is the vertebral endplate. The
adult EP is a hyaline cartilage layer usually not more than 1 mm thick, but it
provides a vital borderline between the IVD and the VB. In very young
individuals, the EP is thicker and penetrated by vessels (Figure 4). During growth
the EP thins and normal adult endplate is avascular, aneural and partly calcified at
the VB interface (Prescher 1998, Urban & Winlove 2007). Adjacent to this
calcified portion of the endplate are localized specialized VB blood capillaries,
which help to feed the IVDs. In fact, according to our current understanding, the
main IVD nutrient supply as well as a route for metabolites to exit the IVD goes
26
through the EPs (Roberts et al. 1989, Urban & Winlove 2007). Unlike many
animal species, the human EP also functions as an epiphyseal growth plate for the
VBs before maturity (Bernick & Cailliet 1982). For reviews, see (Chan et al.
2011, Raj 2008, Roughley 2004, Urban & Winlove 2007, Whatley & Wen 2012).
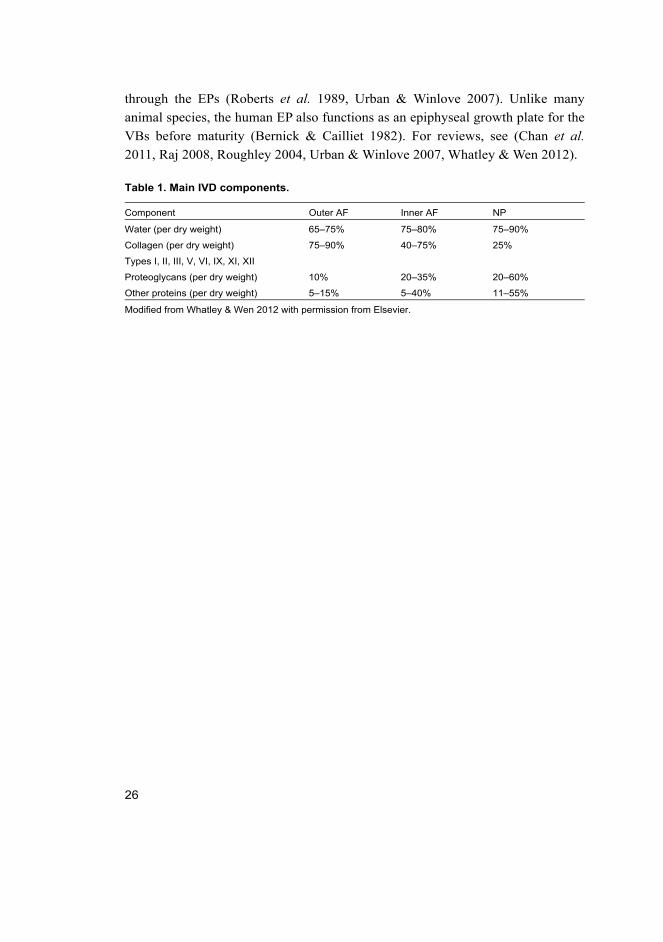
Table 1. Main IVD components.
Component Outer AF Inner AF NP
Water (per dry weight) 65–75% 75–80% 75–90%
Collagen (per dry weight) 75–90% 40–75% 25%
Types I, II, III, V, VI, IX, XI, XII
Proteoglycans (per dry weight) 10% 20–35% 20–60%
Other proteins (per dry weight) 5–15% 5–40% 11–55%
Modified from Whatley & Wen 2012 with permission from Elsevier.

27
Fig. 3. Variation in the composition of the NP with age. The figure illustrates the
collagen fibrils and aggrecan PGs in the NP of fetal, young juvenile, adolescent/young
adult, and mature adult/degenerated IVDs. In the fetus, there is little collagen and the
aggrecan is rich in CS. In the young juvenile, the collagen fibril content is increased,
the aggrecan contains both CS and keratin sulphate (KS), and proteolytic processing
of the proteoglycan aggregates exists. In the young adult, the amount of collagen
peaks, the aggrecan and link protein (LP) have undergone proteolytic processing,
aggrecan is present as both proteoglycan aggregates and nonaggregated fragments,
and the CS chains of aggrecan are of decreased size, whereas its KS chains are of
increased size. In the mature adult, the collagen fibrils show enhanced proteolytic
processing, the aggrecan and LP have undergone extensive proteolytic processing,
the proportion of aggrecan in a nonaggregated form and the hyaluronan content have
increased. (Modified from Roughley 2004. Reprinted with permission from Wolters
Kluwer Health.)

28
Fig. 4. Variation in healthy IVD structure with age. In the fetus, the EP is wide, both it
and the AF are vascularized, and the NP has a fluid consistency rich in proteoglycan
and notochordal cells. In the juvenile, the EP decreases in width, the vascularity of the
AF declines, and the NP becomes more gelatinous because of increased collagen
content. There is a decrease in the proportion of notochordal cells in the NP and
replacement by mesenchymal cells. In the adult, the EP is narrow, no longer covers
the entire nucleus and may be calcified. Inner and outer AF becomes less distinct. The
NP is populated only by mesenchymal cells. (Modified from Roughley 2004. Reprinted
with permission from Wolters Kluwer Health.)

29
2.3 Disc degeneration (DD)
Human disc degeneration (DD) is a complex phenomenon that occurs as a part of
normal aging. However, the largest avascular structure in the body may start to
show signs putative to aging very early — actually before the individual even
reaches maturity (Boos et al. 2002). It is currently not well known which changes
in the IVD should be truly considered abnormal. Even clear pathology, such as
disc herniations that inflict clinical symptoms occur in asymptomatic subjects
(Bradford & Garcia 1971).
Definition of DD
To assess the structure of degenerated IVDs we must first define what DD is. The
term degeneration may include one or more of the following: real or apparent
desiccation, fibrosis, narrowing of the disc space, diffuse or dynamic bulging of
the AF beyond the disc space, AF tears, mucinous AF degeneration, defects and
sclerosis of the endplates, and bony osteophytes of the adjacent structures. The
different degenerative phenotypes (i.e. DD traits) in magnetic resonance imaging
(MRI) include IVD space narrowing, T2-weighted signal intensity loss, IVD
fissures (annular tears), fluid, vacuum changes and calcification within the IVD,
VB changes (Modic changes), ligamentous signal changes, osteophytosis, disc
herniations (LDH), IVD bulges, alignment changes and spinal stenosis (Modic &
Ross 2007, Zou et al. 2009). Before efforts were made to standardize definitions
of DD, it was stated that the term is “only a symbol of our ignorance” (Pritzker
1977). Fortunately the current definition of DD is not that chaotic, but still some
experts disparage the use of DD as a term (An et al. 2004). Histological
definitions are possible, but they are impractical for large-scale studies in vivo
(Roberts et al. 2006), hence the IVD changes must be defined by imaging. Today,
in some cases, DD may be understood simply as desiccation of the NP (Endean et
al. 2011).
As of yet MRI is the gold standard in DD assessment (Haughton 2006). It is
currently the only ethical way to image degenerative changes within the IVD,
because it does not expose the study subjects to potentially harmful radiation. In
patients, plain radiographs or computed tomography (CT) can provide some
information on the IVD status, but especially in the X-ray acquired data is too
limited. This is highlighted in young individuals with early degenerative changes
that are too subtle to be currently detected with modalities other than MRI

30
(Berlemann et al. 1998, Boos et al. 2002). The CT has been used, and is still used
in clinical practice with success when MRI is not available (Thornbury et al.
1993). However, in population-based research the only choice is MRI (Battie et al.
2004, Haughton 2006, Modic & Ross 2007, Urban & Winlove 2007). It has
evolved considerably since its introduction into clinical use in 1980s, and has
almost completely replaced the more invasive imaging modalities (Emch &
Modic 2011), some of which may even predispose to DD (Carragee et al. 2009).
Definitions of DD in MRI are still not uniform, at least in part because the
degenerative cascade is not completely understood. However, during the past
couple of decades there have been multiple justified suggestions of standardized
DD classification schemes. Thompson et al. made one of the first suggestions to
grade DD (Thompson et al. 1990). This MRI scheme of gross disc morphology
was based on 15 human cadaveric spines. It assumes that the process of DD
advances at a similar pace in all IVD structures, which is not always the case
(Battie et al. 2004). Other classifications such as Schneiderman´s (Schneiderman
et al. 1987), Pfirrmann´s (Pfirrmann et al. 2001) and the modified versions of the
latter designed for elderly subjects (Griffith et al. 2007), and young subjects
(Takatalo et al. 2011) provide also discontinuous scales that sum different
measurements of the IVD together. These grades, or other classifications, in
single IVDs have also been used to calculate a summary score of DD in a subject
(Solovieva et al. 2004, Takatalo et al. 2011). The Pfirrmann´s classification is
based on disc signal intensity, differentiation of AF/NP and disc space height
(Pfirrmann et al. 2001).
Other classifications have been developed for more specific changes
including VB and the adjacent EP changes (Modic et al. 1988), LDH (Fardon et
al. 2001), annular tears (AT) (Yu et al. 1988) and high intensity zones (HIZ)
(Aprill & Bogduk 1992). However, all pooled scores that combine different
measurements of IVD characteristics may hide important features and mix
correlations that would shed light on the mechanism of DD. Additionally, the DD
changes are not biologically discontinuous and include multiple different
morphological changes (Urban & Winlove 2007). It has recently been suggested
that different DD phenotypes should be “split” rather than “lumped” (Williams et
al. 2011b).
Despite the lack of solid standards of DD in MRI imaging, it is to be noted
that the T2-weighted image is a valid tool to assess the degeneration of the IVD.
This is especially important in young individuals, since the low signal intensity in
T2-weighted scans presents NP desiccation, which happens in early DD (Emch &

31
Modic 2011, Tertti et al. 1991a). The IVD height is reduced in more severe DD
(Frobin et al. 2001), but again this can be misleading in young individuals, since
disc height has been found to possibly increase in early DD (Boos et al. 1996,
Shao et al. 2002, Twomey & Taylor 1985).
Prevalence of DD
The prevalence of MRI evaluated DD in asymptomatic subjects is an issue of
controversy. In a PubMed-based review the range of different degenerative
findings was: 10–81% for IVD bulges, 3–63% for protrusion, 0–24% for
extrusion, 20–83% for IVD signal changes, 3–56% for decrease of disc height, 6–
56% for AT or HIZ and 8–19% for Schmorl’s nodes. One third of the identified
studies did not specify the subjects’ symptom status, thus these studies were
examined separately (Battie et al. 2004). A systematic review on the topic
reported the ranges among asymptomatic subjects as follows: 0–76% for LDH, 3–
36% for nerve root impingement, 6–56% for HIZ, 7–85% for DD characterized as
decreased signal intensity with or without decreased disc height (Endean et al.
2011). Pooled prevalences were: 27% for LDH, 4% for nerve root impingement,
54% for IVD signal changes and 28% for HIZ/AT. The EP defects are also quite
common, as they have been reported in 6–30% of adults (Videman et al. 2008,
Williams et al. 2007).
A recent study with a large dataset of patients with LBP (N=4322) from
Denmark reported prevalences of: 85.7% for DD (data derived from MRI
narrative reports, possibly IVD signal changes), 65.2% for IVD bulge, 50.8% for
LDH, 13.6% for HIZ and 28.7% for nerve root compromise (Albert et al. 2011),
the prevalence of DD increasing with age (Figure 5, Figure 6). Additionally,
multiple studies have found that MRI defined DD is common already at a young
age (Kjaer et al. 2005, Salminen et al. 1999, Samartzis et al. 2011, Takatalo et al.
2009, Tertti et al. 1991b), but not as common as in older subjects. For reviews,
see (Battie et al. 2004, Endean et al. 2011).
Progression of DD
The IVDs are vascularized and have remnants of the notochord during early life
(Risbud & Shapiro 2011). The EPs are similarly in a premature form thus
providing good nutrition to the IVDs (Figure 4). Hence, the IVD understandably
presents very few histological DD changes during the first two years of life while
32
the fetal IVDs are intact. However, different DD changes become more common
quite early. The prevalence of DD increases after the first 10-year milestone has
been reached (Boos et al. 2002).
Investigations on the DD progression (DDP) are infrequent compared to
cross-sectional studies. Among adults, longitudinal studies have reported annual
progression of DD in ca. 5–10% of subjects (Borenstein et al. 2001, Elfering et al.
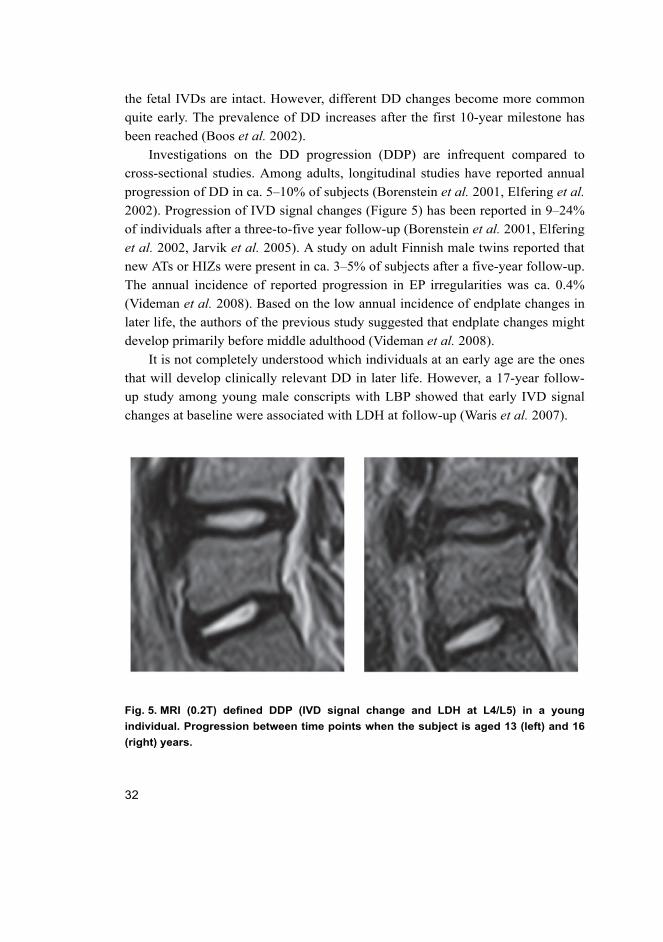
2002). Progression of IVD signal changes (Figure 5) has been reported in 9–24%
of individuals after a three-to-five year follow-up (Borenstein et al. 2001, Elfering
et al. 2002, Jarvik et al. 2005). A study on adult Finnish male twins reported that
new ATs or HIZs were present in ca. 3–5% of subjects after a five-year follow-up.
The annual incidence of reported progression in EP irregularities was ca. 0.4%
(Videman et al. 2008). Based on the low annual incidence of endplate changes in
later life, the authors of the previous study suggested that endplate changes might
develop primarily before middle adulthood (Videman et al. 2008).
It is not completely understood which individuals at an early age are the ones
that will develop clinically relevant DD in later life. However, a 17-year follow-
up study among young male conscripts with LBP showed that early IVD signal
changes at baseline were associated with LDH at follow-up (Waris et al. 2007).
Fig. 5. MRI (0.2T) defined DDP (IVD signal change and LDH at L4/L5) in a young
individual. Progression between time points when the subject is aged 13 (left) and 16
(right) years.

33
Regression of DD
While the prevalence of DD increases with age, there are differences between
individuals in the progression rate of degenerative changes and occasionally the
DD changes may even disappear. In comparison to cross-sectional studies, DD
has been studied considerably less in a longitudinal setting even though
longitudinal studies would be valuable in increasing our understanding of the
natural course of DD and to sharpen our ability to distinguish pathological DD
changes from normal aging. Optimally, the natural course of DD should be
studied in a long-term serial follow-up setting with high-field MRI. The imaging
phenotypes should then be combined with data on clinical symptoms and need of
care in order to be able to discriminate the most important clinical phenotypes in
early DD.
In the light of previous studies, it is generally considered that DD only
progresses. Still, in the studies evaluating DD progression it has been found that
some of the DD traits can also regress. In a study based on adult Finnish male
twins aged 49 years (mean, range 35–69 at baseline) the narrowing of IVD space
(disc height) in the lumbar IVDs (L1–L4) was seen in 69% of subjects at the 5-
year follow-up assessment. However, the changes in disc height ranged from a
41% decrease to a 13% increase in disc height. Similarly, both worsening and
improvement was observed in IVD bulges although the more common trend was
worsening (Videman et al. 2008).
In a 7-year follow-up study of adults aged 52 years (mean, range 26–68 at
follow-up), the investigators observed radiographic improvement in some of the
intervertebral disc abnormalities. However, the three evaluators of both baseline
and follow-up MRIs were able to reach a concensus of IVB bulge improvement in
only one of the 31 subjects (Borenstein et al. 2001). In a 5-year MRI follow-up
study of 41 subjects, the course of DD was characterized by slow progression; no
regression in DD was seen (Elfvering et al. 2002). Another study of 131 subjects
aged 52 years (mean, range 35–70 at baseline) found that 60% of subjects had
some new DD change after 3 years while ca. 5% had regression of DD — mostly
in IVD contour (Jarvik et al. 2005). A 10-year follow-up of 234 twin pairs aged
54 years (mean, range 40–69 at baseline) reported that only one individual (0.2%)
had an improved DD summary score at follow-up (Williams et al. 2011b).
Furthermore, it is well known that LDHs often heal spontaneously and surgical
treatments are required only for a minority of patients (Guinto et al. 1984, Rapan
et al. 2011, Ryu & Kim 2010).

34
In all, it is to be concluded that occasional regression of DD in MRI may
occur, but this issue is not fully elucidated. Furthermore, as mentioned above,
differences in spontaneous regression of changes have been reported between DD
traits.
Fig. 6. Prevalence of IVD degeneration, bulges and herniations according to age
category among 4322 Danish LBP patients. The prevalences increase steeply between
the first two age-categories. The prevalence information is based on MRI narrative
reports dictated by radiologists and specific definitions for these changes are not
available. (Adapted from Albert et al. 2011. Reprinted with permission from Springer.)
2.3.1 Structure of degenerated intervertebral disc
In addition to MRI studies of DD in vivo, the structure of degenerated IVD has
been investigated in biochemical and histological studies ex vivo and in vitro
using also animal models. This approach is justified to increase our understanding
of the DD cascade.

35
The composition of a healthy IVD (Table 1) deteriorates significantly in DD.
The distributions of the main IVD components, collagens and proteoglycans,
change during DD. The amount of type I collagen increases in the NP while the
amount of type II collagen decreases. The structures between collagens can also
change (Pokharna & Phillips 1998, Schollmeier et al. 2000). The PG content
(mainly aggrecan) of the NP decreases (Roughley 2004), and because of this the
amount of water within the IVDs is reduced (Antoniou et al. 1996). These
changes lead to NP fibrosis while the macroscopic IVD tissue colouration also
changes from pale to brownish (DeGroot et al. 2004). The borderlines between
inner and outer AF and between AF and NP become less distinct (Figure 4) while
radial tears in the AF may appear (Figure 7). Furthermore, the internal formation
of AF lamellae may be altered already in early DD (Figure 8). IVD bulging may
appear due to loss of PGs and water (Brinckmann & Horst 1985, Heuer et al.
2007). Structural changes to the IVD emerge not only because of DD but also as a
part of normal aging. It is not easy to differentiate changes that occur solely due
to aging from those in DD (Raj 2008). While discrimination of the IVD changes
is not straightforward, it has been suggested that DD may mimic premature aging
of the IVD (Adams & Roughley 2006, Le Maitre et al. 2007).
The IVD’s well-being is roughly dependent on two factors, ECM synthesis
and breakdown, which are in part reliant on cell type, cell density and metabolite
transport, all of which may be changed in DD or aging (Urban et al. 2001, Zhao
et al. 2007). Aging leads to loss of metabolically active notochordal cells from the
NP, while only less active chondrocyte-like cells remain (Guehring et al. 2008,
Kim et al. 2003, Kim et al. 2005). In addition, IVD cell death rate increases
during aging and DD (Boos et al. 2002, Gruber & Hanley 1998, Trout et al. 1982).
Furthermore, at the time toddlers start to walk, the vascularization of their IVDs
decreases rapidly (Figure 4). In middle age, vascularity of the IVD increases from
the outer parts, which may be related to DD, but also the EP sector is likely to be
affected (Repanti et al. 1998, Yasuma et al. 1990).
In addition to changes in the IVD cell types it has been shown that cell
senescence may contribute to DD (Le Maitre et al. 2007). It has been considered a
separate entity from change in cell phenotype, since phenotypic changes do not
always result from cell senescence (Zhao et al. 2007). All in all, as a result of DD,
the IVD cells seem to lose the ability to produce the correct ECM components.
Multiple such changes have been identified, which, along with possible
implications, are summarized in Table 2.

36
It has been argued previously (Adams & Roughley 2006) that true DD should
be considered when structural changes lead to significant weakening of IVD
function, causing severe NP decompression, increased local AF stress, reduced
stability or increased load to the vertebral arch (Adams et al. 1996, Pollintine et al.
2004, Zhao et al. 2005). In any case, the reference state of a healthy IVD is a
moving target. Furthermore, the emerging field of mechanobiology may help to
increase our understanding of cascades behind the changing IVD structure in DD.
For reviews, see (Adams et al. 2010, Hsieh & Twomey 2010, Raj 2008, Smith et
al. 2011, Whatley & Wen 2012, Zhao et al. 2007)
Fig. 7. Histological section of the outer AF of a disrupted middle-aged IVD. A blood
vessel (arrow) has grown into a posterolateral radial fissure (*) close to the disc
periphery. (Adapted from Adams et al. 2010. Reprinted with permission from Elsevier.)

37
Fig. 8. MRIs illustrating different stages of DD. (A) Healthy IVD exhibiting distinct AF
lamellae (AF) and central NP region (NP). (B) IVD exhibiting early DD, including
moderate height reduction, decreased NP signal intensity and inward bulging of AF
lamellae (*). (C) IVD exhibiting advanced stage of degeneration, including severely
reduced height, large fissure (*) and generalized structural deterioration. Images
obtained using a 7T Siemens MRI scanner. (Adapted from Smith et al. 2011 under
Creative Commons Attribution Non-Commercial Share Alike License
[http://creativecommons.org/licenses/by-nc-sa/3.0]. This adaptation is subject to the
same licence.)
38
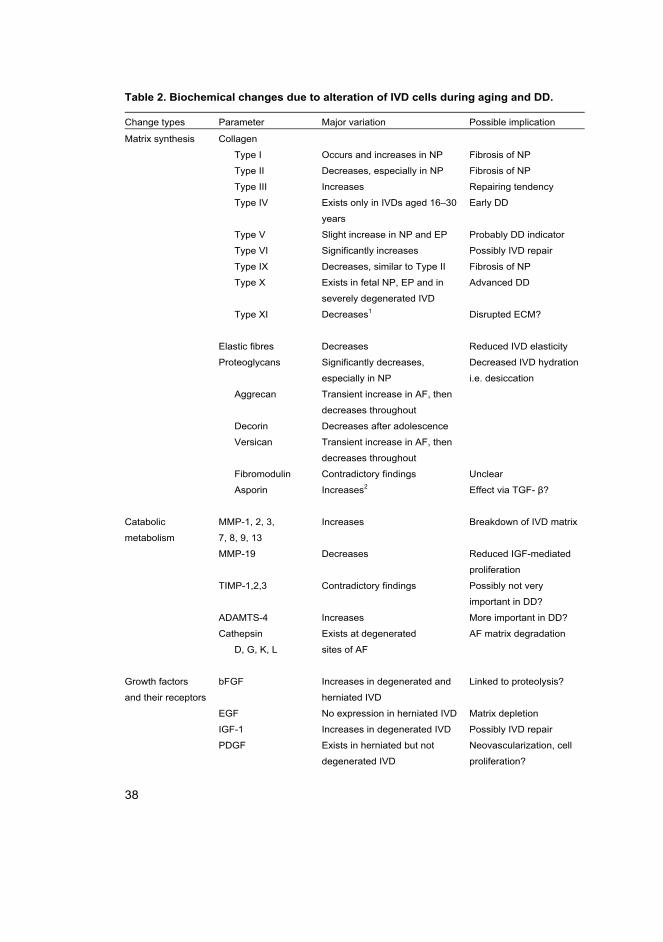
Table 2. Biochemical changes due to alteration of IVD cells during aging and DD.
Change types Parameter Major variation Possible implication
Matrix synthesis Collagen
Type I Occurs and increases in NP Fibrosis of NP
Type II Decreases, especially in NP Fibrosis of NP
Type III Increases Repairing tendency
Type IV Exists only in IVDs aged 16–30
years
Early DD
Type V Slight increase in NP and EP Probably DD indicator
Type VI Significantly increases Possibly IVD repair
Type IX Decreases, similar to Type II Fibrosis of NP
Type X Exists in fetal NP, EP and in
severely degenerated IVD
Advanced DD
Type XI Decreases1 Disrupted ECM?
Elastic fibres Decreases Reduced IVD elasticity
Proteoglycans Significantly decreases,
especially in NP
Decreased IVD hydration
i.e. desiccation
Aggrecan Transient increase in AF, then
decreases throughout
Decorin Decreases after adolescence
Versican Transient increase in AF, then
decreases throughout
Fibromodulin Contradictory findings Unclear
Asporin Increases2 Effect via TGF- β?
Catabolic
metabolism
MMP-1, 2, 3,
7, 8, 9, 13
Increases Breakdown of IVD matrix
MMP-19 Decreases Reduced IGF-mediated
proliferation
TIMP-1,2,3 Contradictory findings Possibly not very
important in DD?
ADAMTS-4 Increases More important in DD?
Cathepsin
D, G, K, L
Exists at degenerated
sites of AF
AF matrix degradation
Growth factors
and their receptors
bFGF Increases in degenerated and
herniated IVD
Linked to proteolysis?
EGF No expression in herniated IVD Matrix depletion
IGF-1 Increases in degenerated IVD Possibly IVD repair
PDGF Exists in herniated but not
degenerated IVD
Neovascularization, cell
proliferation?

39
Change types Parameter Major variation Possible implication
TGF-α Occasionally expressed in
herniated IVD
Matrix depletion
TGF-β Contradictory findings Matrix remodelling?
BMP-RII Expressed in both DD and
healthy IVD
Matrix remodelling?
FGF-R3
IGF-RI
TGF-β-RII Contradictory findings Matrix remodelling?
EGF-R No expression in herniated IVD Matrix depletion
Pro-inflammatory
cytokines
IL1-α Increases in degenerated and
herniated IVD
Enhancement of
catabolic metabolism
IL1-β Increases in degenerated and
herniated IVD
IL-6 Increases in degenerated and
herniated IVD
IL-1-RI Increases in degenerated and
herniated IVD
IL-1-Ra Decreases in degenerated and
herniated IVD
IL-6-R Exists in herniated IVD
TNF-α Increases in degenerated and
herniated IVD
ADAMTS: a disintegrin and metalloprotease with thrombospondin type 1 motifs; AF: annulus fibrosus;
EP: cartilaginous endplate; IVD: intervertebral disc; MMP: matrix metalloproteinase; NP: nucleus
pulposus; PDGF: platelet-derived growth factor; TGF: transforming growth factor; TIMP: tissue inhibitor of
metalloproteinase. bFGF: basic fibroblast growth factor; BMP-R: bone morphogenetic protein receptor;
EGF: epidermal growth factor; IGF: insulin-like growth factor; IL: interleukin; IVD: intervertebral disc;
PDGF: platelet-derived growth factor; TGF: transforming growth factor; TNF: tumour necrosis factor.
(Majority of the data from Zhao 2007 et al. with permission from Elsevier.) 1 Mio et al. 2007, 2Song et al.
2008.
2.3.2 Changing the view of aetiology
Initiating event
The initiating event in DD is not currently known, although decreased nutrition of
the IVD via impaired EP function might be one of the first mechanisms in the
degenerative cascade (Adams et al. 2000, An et al. 2004, Martin et al. 2002,

40
Rajasekaran et al. 2008). However, this theory has been tested in a dog model, in
which the EP nutrition route was blocked by injecting cement into the VB. This
procedure did not lead into DD during the 1-year follow-up (Hutton et al. 2004).
Nevertheless, different kinds of direct traumas to the IVD or EP have been shown
to lead to DD in animal studies (Holm et al. 2004, Moore et al. 1996, Osti et al.
1990). Similarly, trauma caused to human IVDs by invasive diagnostics can result
in DD (Carragee et al. 2009). Among very young children falls and other high-
energy traumas may be the only cause of LDH requiring surgical intervention
(Cahill et al. 2011), but in older subjects it is likely that the IVD function is
compromised by DD before LDH (Dang & Liu 2010, Kumar et al. 2007,
Papagelopoulos et al. 1998, Parisini et al. 2001, Waris et al. 2007).
Risk factors
The traditional view that DD is caused mainly by mechanical factors (wear and
tear), has dominated most of the last century. Conventional risk factors of DD
include age, gender, occupation, obesity, cigarette smoking and exposure to
vehicular vibration (Frymoyer 1992, Hangai et al. 2008, Liuke et al. 2005).
Regardless of what is the exact definition of DD, it increases with age and is
most common in the lower lumbar spine (Cheung & Al Ghazi 2008, Kalichman et
al. 2009, Videman et al. 1995). This supports the biomechanical theory since the
lower lumbar IVDs at levels L4–S1 carry most of the mechanical load (Jensen et
al. 1994). Occupation may predispose to DD via increased axial loading and
torsion movements in physically demanding jobs, which may contribute to
decreased EP function via traumatic events as well (Adams et al. 2000).
Smoking has also been found to be a risk factor for DD (Battie et al. 1991,
Livshits et al. 2001). These findings are supported by recent studies that have
found smoking to be a risk factor to LBP and sciatica (Rivinoja et al. 2011, Shiri
et al. 2010b). However, it is to be noted that LBP, sciatica and LDH are not
independent of one another. Smoking might have an effect to DD via IVD
nutrition, as disc cell viability depends on diffusion of nutrients into the IVD
(Grunhagen et al. 2006, Urban et al. 1977). From animal studies it is known that
smoking affects endplate blood microcirculation and IVD gene expression (Holm
& Nachemson 1988, Uei et al. 2006). Smoking has been also shown to have
affects on type I and III collagen synthesis and MMP-8 levels in human skin
(Knuutinen et al. 2002). Lumbar artery narrowing caused by smoking is a
biologically plausible risk factor for DD. Furthermore, obesity has been reported

41
as a risk factor for DD in multiple studies (Liuke et al. 2005, Samartzis et al. 2011,
Solovieva et al. 2002). Yet, it is not clear whether obesity affects via increasing
the IVD mechanical load or by some other way.
A back trauma reported by the patient does not seem to predict future DD
(Hancock et al. 2010), and neither does a stabile vertebral fracture (Moller et al.
2007). Participation in competitive sports, especially swimming and baseball,
might also be a risk factor for DD (Hangai et al. 2009, Kaneoka et al. 2007).
Familial predisposition
In addition to mechanical and environmental factors that contribute to the DD risk,
it has long been known that some spine problems may be inherited (Bull et al.
1969). During the 1970’s two reports gave some attention to positive family
history in relation of disc failure (Grobler et al. 1979, Wiltse 1971). At the end of
the 1980’s, 13-year-old identical twin girls in Australia suffered from strikingly
similar LDHs at L5–S1 within months of each other. Both girls had a LDH on the
right side of their body and they were also both active in competitive sports. This
sparked Gunzburg et al. to write a case report and review of the literature in 1990
(Gunzburg et al. 1990). An American case-control study reported the next year
that subjects aged less than 21 years had approximately five times greater risk for
LDH if they had positive family history (Varlotta et al. 1991). A larger Japanese
study reported the next year that LDH had a familial predisposition among
individuals aged 18 years or less (Matsui et al. 1992). One year later, an Italian
author reported familial aggregation of LDH often requiring surgery and also
noted that in the large family many individuals also had chronic low back pain
syndrome. The author was convinced that there has to be a genetic predisposition
to early DD to explain such a large aggregation of individuals with lumbar disc
disorders (Scapinelli 1993). It is possible that the underlying aetiology of these
early-onset more severe phenotypes differs from that of the sporadic, age-related
DD. In any case, the subsequent studies have indicated familial predisposition in
DD (Bhardwaj & Midha 2004, Matsui et al. 1998, Richardson et al. 1997, Saftic
et al. 2006, Simmons et al. 1996).

42
Heritability estimation in DD
Familial aggregation of a trait per se does not prove that the trait has a genetic
component; family members normally also share the same environment. In twin
studies the phenotypes of monozygotic (MZ) and dizygotic (DZ) twins are
compared. MZ twins develop from a single ovum fertilized by a single sperm and
thereby share many characteristics; hence they are called identical twins. In
contrast to the MZ twins that are genetically identical, DZ twins share only about
half of their genetic variations, similar to “normal siblings”. The heritability,
which is an estimate of the contribution of genetic effects to variation within a
trait, can be derived by using the data acquired from twin studies. Heritability can
be calculated using the Falconer's formula, hb2=2(rmz - rdz), where hb
2 is the broad
sense heritability, rmz is the MZ twin correlation, and rdz is the DZ twin correlation
(Falconer & MacKay 1996). The results of twin studies are applicable especially
to the population from which the twins have been selected.
The traditional view that DD is mainly caused by environmental factors
began to change at the latest in 1995 when the Twin Spine Study was published
(Battie et al. 1995b). Only about three years before this study was published a
review on aetiology of “degenerative disc disease” stated that ‘‘Among the factors
associated with its occurrence are age, gender, occupation, cigarette smoking,
and exposure to vehicular vibration. The contribution of other factors such as
height, weight, and genetics is less certain’’ (Frymoyer 1992). The Twin Spine
Study, which started in 1991, was a collaboration between researchers mainly
from Canada, Finland and the United States that used a population-based Finnish
Twin Cohort. In this study with 115 MZ twin pairs the familial aggregation,
representing both genetic and early shared environmental factors, explained 61%
of the variance in DD. The knowledge gained from this study and from another
report published the same year (Battie et al. 1995a) has had a very important
input to the present conception that DD is largely caused by genetic factors
(Battie et al. 2009). Subsequent studies have supported this view. A classic twin
study with 172 MZ and 154 DZ twins (mean age 51.7 and 54.4 years, respectively)
used a total DD summary score. After adjustments for age, weight, smoking,
occupational manual work and exercise this score showed a heritability of 0.74
(95% CI 0.64–0.81) (Sambrook et al. 1999). Another study of six Arabic
pedigrees, which included a total of 221 individuals, analysed the heritability of
LDHs. After adjustments for age, weight, smoking and gender, the heritability
estimate for multiple LDHs was 0.73 (Livshits et al. 2001). A study that recruited

43
a total of 257 siblings evaluated DD using radiographs. They adjusted their
analyses for age, sex, BMI and bone mineral density (BMD), after which the
heritability estimate was 0.75 (95% CI 0.30–1.00) (Bijkerk et al. 1999).
Heritability estimates have varied a little between studies and according to the DD
traits used, but still the heritability estimates are significant and high enough to
justify the study of the genetic component in DD.
Heritability estimation in disc degeneration progression (DDP)
A recent twin study with a 10-year follow-up period found that, similar to DD,
disc degeneration progression (DDP) heritability varies between different DD
traits (Williams et al. 2011b). However, at the moment this is the only study to
estimate the heritability of DDP. The study population consisted of 90 MZ and
144 DZ twin pairs. Of all the subjects, 95% were female. IVD height change was
not heritable at all, while posterior IVD bulge was heritable in all age categories.
However, the influence of age varied between different DD traits. The heritability
estimates for IVD bulge progression were 0.28 (95% CI 0–0.65) for the age group
under 50 years, 0.51 (95% CI 0.28–0.74) for 50–60-year-olds and 0.53 (95% CI
0.13–0.94) for those over 60 years. The authors suggested that in the genetic
studies of DDP, at least among women, the focus should be on individuals less
than 50 years of age. This is well justified because disc signal changes (the most
commonly used DD trait) were heritable only among those under 50 years old.
For that age group the heritability estimate for IVD signal changes was 0.76 (95%
CI 0.44–1.00) for disc signal changes and 0.74 (95% CI 0.42–1.00) for anterior
osteophytes (Williams et al. 2011b).
In a summary of the aetiology of DD, it has to be noted that while genes play an
important role in DD including LDH, these conditions are believed to have a
multifactorial aetiology (Figure 9). For reviews, see (Adams et al. 2000, Adams &
Roughley 2006, Ala-Kokko 2002, Battie et al. 2009, Kalichman & Hunter 2008a,
Masuda & Lotz 2010).

44
Fig. 9. A model showing multiple risk factors of DD. Interactions including additive
effects and epistasis between genes are possible.
Associating conditions
Depending on the definition the prevalence of DD can vary, but still it is a
common condition (Battie et al. 2004). As such, associations with other common
conditions or measurements have been reported. Atopic dermatitis, developmental
hip dysplasia, osteoarthritis (OA) and BMD have been previously related to DD
in one way or another (Gruber et al. 2009, Ito et al. 2003, Livshits et al. 2010,
Loughlin 2011, Näkki et al. 2011). These associations do not imply causality, but
are rather suggestions of possible shared underlying aetiology, that require more
rigorous investigation.
2.3.3 Clinical relevance and treatment
According to the latest research, DD is associated with LBP and other clinical
symptoms (Adams et al. 2010, DePalma et al. 2011, Endean et al. 2011),
especially among young individuals (Paajanen et al. 1997, Samartzis et al. 2011,
Takatalo et al. 2011). LBP itself is a common condition already during

45
adolescence (Auvinen 2010, Kjaer et al. 2011, Leboeuf-Yde & Kyvik 1998), and
it decreases the quality of life when untreated (Fontecha et al. 2011).
LBP has been defined as pain between the lower rib cage and gluteal folds
often radiating to the thighs (Frymoyer 1988). The definitions of LBP vary in
spine research and it has been noted that the country where the study has been
done should be considered (Auvinen 2010, Paalanne 2011). Low back stiffness
and other phenotypes with minor symptoms should be avoided, unless it has been
shown that these phenotypes have some clinical relevance. Duration and severity
of the pain should be also evaluated (Dionne et al. 2008). LBP has often been
classified in three categories based on duration: acute (less than 6 weeks),
subacute (between 6 and 12 weeks) and chronic (more than 12 weeks) (van
Tulder et al. 2006). The majority of LBP is non-specific (Nachemson et al. 1985,
Paalanne 2011, Koes et al. 2006), in which imaging at the acute phase rarely leads
to any changes in treatment, and is therefore usually to be avoided (Chou et al.
2009). Furthermore, LBP is often episodic (Cheung & Ghazi 2008) while DD is
not. This issue has not been fully elucidated, but there is evidence that genes play
a role also in the variation of LBP perception (Tegeder & Lotsch 2009). In all, it
has been estimated that LBP is one of the great banes at the individual, society
and health-care system levels (Andersson 1999, Dagenais et al. 2008, Deyo &
Tsui-Wu 1987, Ekman et al. 2005, Katz 2006, Wieser et al. 2011).
Among adults, DD is often seen as the source of LBP, but the association of
DD with pain is more complicated than in young individuals (Cheung et al. 2012,
Chou et al. 2011, de Schepper et al. 2010, Paajanen et al. 1989, Visuri et al. 2005).
According to a recent systematic review, multiple DD traits are associated with
LBP (Endean et al. 2011). In the meta-analyses the odds ratios were; 3.6 for LDH,
2.5 for DD (signal changes) and 2.5 for HIZ/AT. However, it was also noted in
this review that every second (54%) asymptomatic individual has DD (signal
changes) (Endean et al. 2011). Clearly, the evidence for the relationship between
DD (as defined by current imaging modalities) and LBP is still controversial.
A specific entity related to both DD and LBP is lumbar facet joint
osteoarthritis (LFJOA). LFJOA is considered to be one of the factors affecting
LBP, but the role of different imaging abnormalities seen in patients with LBP is
still controversial (Kalichman & Hunter 2007). LFJOA is associated with DD and
it has been suggested that DD might precede LFJOA but also contrary evidence
exists (Kalichman & Hunter 2007). However, LFJOA is not considered to be
relevant in the very early DD as is it not seen before maturity (Stelzeneder et al.

46
2011). In fact, there is even evidence that LFJOA does not occur before the age of
45 years (Carrino et al. 2009, Fujiwara et al. 1999).
Clinical manifestations of DD include LBP as well as sciatica often caused by
LDH. The treatment of very severe LDH is straightforward and surgical
procedures are performed in order to avoid permanent damage from e.g. cauda
equina syndrome (Gitelman et al. 2008, Olivero et al. 2009). However, in more
mild clinical phenotypes surgery (discectomy, fusion of motion segments,
artificial IVD etc.) is an option only for a few patients thus the current treatments
of choice are mainly conservative. Specific exercises do not seem to be effective
in acute LBP but staying active is beneficial (van Tulder et al. 2006). Informing
the patient about the natural course of acute LBP is very important, and also
pain/relaxant medication is often used with some success (van Tulder et al. 2006).
Early physical therapy after acute LBP may reduce subsequent medical service
usage (Gellhorn et al. 2010). In chronic LBP the situation is increasingly
multimodal, while patient education, physical therapy and cognitive behavioural
therapy have some proven effect (Karppinen et al. 2011). Patients also tend to
engage in treatments outside conventional medicine (Brinkhaus et al. 2011).
It is to be noted that while most patients with symptomatic DD are treated
conservatively, young individuals with LDH and clear indication for surgery seem
to respond relatively well to operative treatment (Dang & Liu 2010). For reviews
on current and emerging treatments of DD-related LBP, see (Adams et al. 2010,
Clouet et al. 2009, Jacobs et al. 2011, Karppinen et al. 2011, Kepler et al. 2011,
Raj 2008, Whatley & Wen 2012).
2.4 Genetic studies of complex disorders
The history of genetics dates back to the 19th century. The efforts of friar Gregor
Mendel and Thomas Morgan led to the foundation of genetics and to Mendel’s
laws of genetics. These laws are not valid as such for the study of heritable
diseases with polygenic and multifactorial aetiology. However, there might be
rare monogenic forms of complex disorders (Peltonen et al. 2006). The
contribution of genes compared to environment can be investigated in twin
studies (see chapter 2.3.2).

47
2.4.1 Variation in the human genome
The haploid human genome consists of ca. 3.2 billion base pairs and codes for
about 20,000–25,000 genes (Lander et al. 2001). Individuals differ from each
other for only 0.1% of their genomes, but still each genome contains millions of
variations (Marian 2012). The prevalence of the different variation type varies.
Microsatellites (or STR, short tandem repeats) are repetitive sections of DNA
consisting of a few base pairs (bp) while minisatellites (or VNTR, for variable
number of tandem repeats) are a bit longer; dozens of bp that are similarly
repeated. Copy number variations (CNV) are large regions of the genome that
have been deleted or replicated. The importance of common CNVs in the studies
of complex disorder genetics has been recently questioned as the majority of them
are tagged by single nucleotide polymorphisms (SNP) that are responsible for
only one bp change in the DNA. However, the importance of rare CNVs requires
further investigations (Wellcome Trust Case Control Consortium et al. 2010).
Recent findings have also indicated that it may be too early to neglect the CNVs
in the study of complex diseases as they have not been exhaustively studied
(Gamazon et al. 2011).
The initial whole genome sequencing project identified ca. 1.2 M SNPs
(Lander et al. 2001), but more SNPs have been discovered since. According to a
recent update, the human genome includes ca. 15 M SNPs, 1 M insertions and
deletions and 20,000 structural variants. It was also estimated that de novo
germline base substitution mutations occur in ca. 1 out of 100 Mbp (The 1000
Genomes Project Consortium 2010). In genetic studies with specific diseases the
linkage analyses are largely based on the STRs while candidate gene studies are
driven by hypotheses that have been often tested using SNPs. The SNPs can be
either selected to cover a certain part of a locus/gene (tagSNPs), or to include
variations that have previously shown associations with gene function or related
phenotypes of interest (International Hap Map Consortium et al. 2007). Today,
SNPs can be genotyped using multiple different methods and commercial kits,
and chips are often used so that the number of included variations may exceed
1,000,000 (Haberlandt et al. 2012).

48
2.4.2 Approaches to genetic studies of complex disorders
According to the current knowledge the risk of many complex disorders is
affected by the environment and multiple genes (Bodmer & Bonilla 2008). A
common variation is unlikely to cause a significant increase in an individuals risk
for disease (Figure 10). However, there is evidence suggesting that common
variants do have a role in common diseases (Lohmueller et al. 2003), and that the
more prevalent the disorder the greater the number of alleles predisposing to the
disease (Figure 11) (Marian 2012). Despite the possible rare monogenic forms
(Peltonen et al. 2006), it is not very likely that in a common disease a single
disease causing variation, that is replicable between populations of different
ethnicity, would be found. Different genes may be involved in different phases in
complex cascades and they may have nonadditive interactions with other genes
(epistasis) and the environment (Khoury et al. 2007).
Fig. 10. Rare and common variants and their relation to putative effect sizes. (Adapted
from Manolio et al. 2009, originally from McCarthy et al. 2008. Reprinted by permission
from Macmillan Publishers Ltd: Nature, copyright 2009 and Nature Reviews Genetics,
copyright 2008.) GWA=GWAS.

49
Fig. 11. Schematic presentation of disease prevalence and the effect sizes of the
causative alleles. (Adapted from Marian 2012 with permission from Elsevier.)
Furthermore, it may also be that the disorder consists of a spectrum of phenotypes.
This phenotypic heterogeneity may lower the robustness of the data obtained
from each study as well as decrease applicability of the findings to an individual
(Ioannidis et al. 2008, Marian 2012).
The rare alleles causing rare monogenic forms of complex diseases with large
effect sizes are presumably easier to identify than the common variations and can
increase knowledge of the complex trait, but in general the mode of inheritance is
not known in complex disorders (Peltonen et al. 2006). The smaller the effect the
larger the study population needs to be in order to result in true positive
association, and thus the required study population may need to include tens of
thousand of subjects (Ioannidis et al. 2008, Marian 2012). It may even be that a
single predisposing genetic variation yields so small an effect on the phenotype
that it is not possible to identify by any current investigation approach.
The genome-wide association studies (GWAS) studies are based on the
“common disease-common variation” (CD-CV) theory (Altshuler et al. 2008).
This theory proposed that multiple common polymorphisms (minor allele

50
frequency, MAF; >0.5–1%) with small effect sizes might predispose to common
disorders (Figure 10). According to this theory the common predisposing alleles
have also formed so early in history that different groups of individuals can be
combined and compared in large consortiums. The usage of this approach enables
the identification of low-effect variations. Indeed, many low-effect common
variants have been identified while bigger effects are usually caused by rare
variants (Bodmer & Bonilla 2008). However, it has been suggested that these rare
variants might also cause common disease (the RV-CD -theory, explained in more
detail subsequently).
Genetic studies of complex disorders can be based on a prori knowledge of
underlying biology or gene location that suggests a potential involvement in the
disease cascade for the gene that is investigated. On the other hand, this candidate
approach is also limited to the previous knowledge, unlike the more recent
methods, such as GWAS, that are hypothesis free, and in that sense unbiased.
Different approaches are presented in Table 3.
Table 3. Current approaches to study complex disorder genetics.
Candidate approach Unbiased approach
Candidate genes Genome-wide association studies (GWAS)
Biologic plausibility1 Expression quantitative trait loci analysis
Chromosomal position mapped through linkage
or GWAS
Candidate SNPs Next generation sequencing
Biologic function1 Targeted subgenomic sequencing
Locus position mapped through
linkage or GWAS
Next generation sequencing in combination
with linkage in families2
Linkage disequilibrium structure Whole-genome sequencing
Novel and known SNPs Whole-exome sequencing
Direct sequencing of the candidate gene
Modified from Marian 2012 with permission from Elsevier. 1Approach used in this thesis (II, III, IV). 2For
more information, see Bailey-Wilson et al. 2011.
Candidate approach
It is possible to search for disease causing genes via association analysis using the
candidate gene approach. In this method, previous knowledge of the disease
cascade or the components of target tissues are utilized. Variation distributions in
the genetic regions, or loci, of interest are compared between affected and

51
unaffected individuals most often in a cross-sectional case-control setting. If allele
and/or genotype frequencies are significantly higher or lower in either group, a
plausible link for the disease has been established. It is also possible to investigate
only certain candidate SNPs based on their known or presumed function, and
previously identified locus (Virtanen et al. 2007b) or they may be selected based
on the linkage disequilibrium (LD) structure (explained in more detail
subsequently). While these kinds of association analysis are among the most
frequent genetic studies to date (Little et al. 2009), the validity of the results
obtained from these studies has been criticized as many identified associations
have not been replicated in subsequent studies (Ott 2004, Sullivan 2007).
However, they are still useful especially for validating previously identified
associations. Furthermore, it is to be noted that some of the disease associating
genes have later withstood validation studies in larger datasets (Kere 2010).
While most of the association studies have been performed using populations or
case-control settings, additional approaches with simultaneous usage of family
and association data are also possible (Ott et al. 2011).
Linkage analysis
Prior to genome-wide association studies, the genetic component of complex
disorders has been investigated using a linkage analysis approach. Linkage
analysis may be a powerful tool if the inheritance pattern of a disease is known,
which is not the case in complex disorders. In this method multiple variations
sites, usually microsatellites and SNPs, spanning the whole genome are compared
between unaffected and affected members of preferably large families to detect
disease loci. Two loci in the genome are in linkage if they are inherited together
from a parent to offspring more often than what would be likely with independent
inheritance (Schork 1997). If a marker, either microsatellite or SNP, is adjacent or
close enough to a locus that contains a disease associated variant, it is likely that
these two are inherited together without a recombination taking place between
them during meiosis and are thereby linked. This method may enable the
identification of a chromosomal region in which the gene affecting the phenotype
may lie when the genetic markers are genotyped from each individual of the
family and the inheritance of the marker (co-segregation) is followed through the
pedigree (Dawn Teare & Barrett 2005). The identified regions are nevertheless
large, possibly spanning over millions of bp. Additionally, the reduced effect size
(or penetrance), phenocopies and variable expressivity are factors that increase

52
complexity in common disorders and complicate the usage of linkage analysis
(Kere 2010). Linkage studies have been successful with monogenic diseases, but
in complex disorders the mapping has not proved to be a very effective approach
(Altmuller et al. 2001, Altshuler et al. 2008). Often studies on the complex
disorders have been reported to map to one chromosome after another but the
earlier linkage results have been rarely replicated (Kere 2010). However, meta-
analyses on multiple linkage studies might provide additional guidance on where
to direct the next efforts (Bouzigon et al. 2010). Additionally, it is possible that
with advancing computational approaches the application of linkage analysis in
complex disorders may become more successful in the future. Furthermore, it
may be that the linkage studies in large families may detect variants of rare
Mendelian diseases that mimic complex disorders. These variants may have an
infinitesimal effect at the population level, but the same cascades may be
associated with complex disorder susceptibility in a wider scale (Peltonen et al.
2006). Phenotype definition is also an important issue in studies based on families.
Linkage versus association
The main difference between association and linkage analysis is that in
association analyses the study population is unrelated whereas linkage analysis
utilizes the available information about the families. This leads to the situation in
which the association setting is better suited to investigations focused on finding
common predisposing variants with low effects, whereas linkage analysis is
applied to discover wider genetic regions in which variants leading to greater
effect may lie (Hirschhorn & Gajdos 2011). An additional possibility is to
combine these analyses (Ott et al. 2011).
Genome-wide studies
One of the most significant technical achievements in the study of complex
disorders has been the development of the genome-wide association approach
(GWAS). These studies do not require previous knowledge of the specific genes
behind the studied complex disorder and are therefore unbiased compared to
candidate approaches (Table 3). The shift to this technology from the earlier
candidate association approach required four achievements (Hirschhorn & Gajdos
2011): the discovery of linkage disequilibrium (LD) between groups of common
variants close to each other (Gabriel et al. 2002), databases of common variants

53
and correlations between them, i.e. haploblocks (International Hap Map
Consortium 2005, International Hap Map Consortium et al. 2007), high-
throughput genotyping methods (Ragoussis 2009) and statistical methods to
artificially calculate (impute) missing information from the genotyped variations
(Li et al. 2009, Marchini & Howie 2010). LD is the non-random correlation
between polymorphisms at two or more loci that may or may not be proximal to
each other. The known LD structure and specific tagSNPs can be used to
mathematically fill in genotypes that are not in fact genotyped. This method is
called imputation, and it understandably requires that the LD structure of the
investigated locus has been studied earlier (Gabriel et al. 2002). Additionally, it is
to be noted that the LD structure is representative for the ethnic population it has
been derived from. It has also been noted that the use of sufficiently large and
bias-free populations has been crucial for the GWAS studies (Kere 2010).
Because the number of genotyped variations is high, the “new” p-value to
mark genetic association in these GWAS studies has been lowered to p=5 x 10-8
(McCarthy et al. 2008) This has resulted in a situation where many of the
previously identified associations do not reach genome-wide significance (Kere
2010), but it is to be noted that this "new" threshold applies only if the whole
genome is tested. It is very reasonable that the positive association threshold has
been adjusted when these GWAS studies have been taken into broad use.
Otherwise there would be a significant possibility for false association because of
multiple testing (Gao et al. 2010, McCarthy et al. 2008).
To date hundreds of GWAS studies have been performed, many of them in
large international collaboration groups. More than 1000 associations between
polygenic disorders and genetic markers were identified by the year 2010
(Hirschhorn & Gajdos 2011), and the number is growing rapidly (Zuk et al. 2012).
These associations do not necessarily identify the exact location of a disease
causing mutation, but may point out markers in important regions, which help
researchers decide where to direct their next efforts. Although it is now accepted
that the GWAS studies are usable in gathering evidence, the results have been
often quite surprising (Kere 2010). They may have pointed to genetic regions that
no one would have examined based solely on the previous information of genes
and disease cascades (i.e. it would have been unlikely that any candidate gene
association analysis would have been done) (Kere 2010). One such discovery is
the association between FTO and obesity (Frayling et al. 2007).
The trend has been to increase the sample size in the GWAS studies in order
to find alleles with moderate effect sizes (Figure 10). However, to detect effect

54
sizes smaller than 2-fold, typically more than 10,000 subjects in both case and
control groups are necessary requiring truly large scale collaborative GWAS
(Newton-Cheh et al. 2009, Teslovich et al. 2010, Sotoodehnia et al. 2010). It is to
be noted that while the GWAS studies have provided us with considerable
information, a big part of the variability between individuals in most cases
remains to be explained. In contrast to the CD-CV hypothesis, the "rare variant-
common disease" (RV-CD) theory that suggests that heritability of the complex
phenotypes is mainly a consequence of numerous rare variants with larger effect
sizes (Bodmer & Bonilla 2008). The recent findings have led to a concept referred
to as “missing inheritance”, the still unexplained inheritance that might be partly
explained by these rare variants (Eichler et al. 2010, Kere 2010, Manolio et al.
2009, van et al. 2010, Zuk et al. 2012). The rapid technologic advantages and
significant drop in DNA sequencing cost have now brought us into the post-
GWAS era meaning the possibility to feasibly sequence the entire genome or
exome in order to investigate the aetiology of complex disorders (Marian 2012).
Expression quantitative trait loci analysis
Technical advancements have generated another unbiased method for the study of
complex disorders, the expression quantitative trait loci (eQTL) analysis. In the
eQTL, the mRNA or protein levels in a specific tissue sample are compared for
example with SNPs or microsatellites using linkage or association analysis
(Goring et al. 2007, Kwan et al. 2008). The eQTLs may lie close to the regulated
genes (i.e. cis-acting) or they may have affect to the gene from a distance (i.e.
trans-acting) (Michaelson et al. 2009). In most cases the specific mechanism by
which the eQTLs have an effect on the gene expression is not yet elucidated.
Additionally, it is to be noted that many eQTLs may be specific to the
investigated tissue or cell type (Gerrits et al. 2009). To date, thousands of eQTLs
and their respective target genes have been identified and this method is likely to
continue increasing our understanding on complex disorder genetics (Cookson et
al. 2009). However, this method has not yet been used for any DD trait.
Next generation sequencing
The significant reduction in the cost of DNA sequencing per base pair has
improved the possibilities to obtain more and more sequencing data on subjects in
studies of complex disorders. Next generation sequencing (NGS) makes it

55
possible to identify all the structural variations in an individual (large CNVs may
be possible exceptions). The NGS methods have improved considerably since the
introduction of the first commercial pyrosequencing platform in 2005 (Margulies
et al. 2005). The different approaches in NGS can be divided into three categories:
targeted subgenomic sequencing, whole-genome sequencing and whole exome
sequencing (Table 3).
The targeted subgenomic sequencing (i.e. targeted resequencing) enables the
capture of multiple selected sequence regions of the genome that have been
previously linked to the phenotype of interest. The selected regions may include
exons, splice sites, regulatory regions or the whole genomic area of interest. This
kind of method has been used in mutation screening in genes coding for
sarcomere proteins in patients with cardiomyopathies, autosomal-recessive
cerebellar ataxia and nonsyndromic deafness (Meder et al. 2011, Vermeer et al.
2010, Rehman et al. 2010). However, due to higher sequencing output and lower
cost per bp this method is likely to be replaced by whole-exome and whole-
genome approaches.
At the moment, the whole exome sequencing is likely to be the most widely
used method to investigate the genetic causes of complex disorders. This is
because of practical reasons such as cost, data handling and biostatistical analyses.
The whole-exome sequencing has been used successfully in multiple Mendelian
disorders (Choi et al. 2009, Choi et al. 2011, Comino-Mendez et al. 2011), but
this approach has also faced challenges in complex disorders (Marian 2012).
The NGS method with the most complete output of the DNA primary
structure, whole-genome sequencing, has been used successfully in both rare
Mendelian disorders and sick sinus syndrome (Roach et al. 2010, Sobeira et al.
2010, Holm et al. 2011). The costs of the whole-genome sequencing are
presumed to decrease in the future. After development of new bioinformatics
solutions to handle the enormous amount of data obtained from this approach, it is
likely that this method will be the next widely used tool to investigate complex
disorder genetics. However, a large number of individuals are still likely to be
needed to identify the causal genetic components from this whole-genome data
(Marian 2012).
For reviews on the history and recent advances in complex disorder genetics,
see (Hirschhorn & Gajdos 2011, Kere 2010, Zuk et al. 2012, Marian 2012).

56
2.5 Finnish disease heritage
Finland is a good location to perform genetic studies. The Finnish population
originates from small founder populations, but due to the geographical structure
of the country, the structure of the society in previous centuries along with other
factors have led to the formation of internal genetic isolates within Finland. This
has made it possible for certain diseases to enrich in these isolates and later in
Finland compared to other countries. This is today known as the Finnish disease
heritage (FDH) (Perheentupa 1972, Norio et al. 1973). The FDH includes rare
monogenic disorders, mostly autosomal recessive conditions that are
overrepresented in Finland. The diseases affect many organ systems and require
the attention of many medical specialities, but paediatricians are the first to
encounter them in most cases (Norio 2003c). Quite often FDH diseases affect
cognition and they may be lethal as well. Currently there are nearly 40 identified
individual diseases in the FHD and it is possible that more will be found in the
future. The definition of FDH does not currently include complex disorders,
despite the fact that these may also have some specific aspects among the Finns
(Sarantaus et al. 2000). In addition to the rare monogenic FDH diseases, such as
aspartylglucosaminuria, we may have heritable forms of common disorders, such
as familial hypercholesterolemia (Vuorio et al. 2001). For more information on
the FDH, see reviews by Norio (Norio 2003a, Norio 2003b, Norio 2003c).
Because of the origin of the population the Finns are genetically more
homogenous compared to Central Europeans. Still, due to the internal isolates it
may be useful to include study subjects only from one geographic region of the
country (Salmela et al. 2008, Salmela et al. 2011). This is because genetic
heterogeneity might hinder efforts to identify genetic variants that have only a
modest effect on a phenotype. To reduce this effect, one of the studies included in
this thesis has focused on a Finnish subpopulation living geographically in a
relatively small area, within a 100 km radius of Oulu, a city in Northern Finland
(II).
2.6 Genetic studies in disc degeneration
The exploration of the genetic factors that predispose to disc degeneration (DD) is
in its infancy compared to many other complex disorders. One of the most
important factors explaining this situation is the fact that the clinical
manifestations of DD, low back pain (LBP) and neurologic deficits, are often self-

57
limiting and curable. Even though the accumulated impact on society is
significant, other complex disorders with apparent resultant mortality or more
severe morbidity have understandably been investigated more rigorously.
Furthermore, the phenotyping problems in DD may have hindered efforts to
produce results with strong credibility.
However, the situation is similar in many other non-fatal or less common
complex disorders. For most diseases, the number of published genetic
association studies is less than 100, mostly on single gene/polymorphisms, only
few meta-analyses (if any), and no strong international network or consortia
between investigators (Ioannidis et al. 2008).
The stage of accumulation of evidence for DD is contrasted against five other
selected diseases in Table 4. It is clearly indicated that the stage of accumulation
of evidence is not high for DD at the moment.
Table 4. Variation in the volume of genome epidemiology evidence for selected
diseases.
Disease Published
articles
Genes
studied
Meta-analyses
(HuGE reviews)
GWAS1 Consortia
Type 2 diabetes 2246 555 56 (7) 35 Established
Breast cancer 1020 275 24 (3) 23 Established
Osteoporosis 503 128 11 (2) 5 Established
Pre-term birth 176 112 1 (1) 0 Emerging
Childhood leukaemia 102 76 1 (1) 6 Emerging
Disc degeneration 52 51a 0 (0) 1 Suggested a Dependent on the DD phenotype definition. 1 Number of published GWAS studies: Hindorff LA et al. A
Catalog of Published Genome-Wide Association Studies. Available at: www.genome.gov/gwastudies.
Accessed October 6th 2012. (Table modified from Ioannidis et al. 2008, reprinted here with permission
from Oxford University Press.)
2.7 Candidate gene studies in disc degeneration
The majority of published genetic studies in DD are candidate gene studies. These
studies include the hypothesis that the investigated gene(s) may predispose to DD
based on what is previously known about the genes. For example, vitamin D is
required for normal bone homeostasis, calcium absorption, phosphate and
calcium homeostasis and parathyroid hormone secretion control. Vitamin D
receptors (VDR) bind the hormonally active form of vitamin D (1,25(OH)2D3) that
produces multiple biologic effects. Bone, cartilage and IVD share similar connective

58
tissue characteristics. Therefore the VDR has been considered to be a good candidate
for DD (Videman et al. 1998).
Other studied candidate genes express proteins that are similarly biologically
plausible to have effect in DD due to their known functions. These groups of
genes include the structural components of IVD (collagens, proteoglycans,
cartilage intermediate layer protein), catabolic proteins that may unbalance
connective tissue turnover in IVD (matrix metalloproteinases) and genes affecting
to the regulation of inflammation (interleukins) and other genes that have been
studied because of know association to another complex disorder, such as
osteoarthritis (OA).
A systematic literature review was conducted as part of this thesis (I). The
systematic approach identified 52 association studies reporting on DD defined
using MRI imaging. During the review process it became obvious that there is
gold standard in the coding of MRI for degenerative IVD changes. In almost
every study the specific DD phenotype is different and there is no reliable data
available on how these different phenotypes could be combined or how the
heterogeneity between the studies could be corrected in statistical analyses.
Therefore, in order to present all the available data without inflating of deflating
the available evidence, the identified studies in each gene (or gene group) are
presented here according to the specific DD phenotype. The approach that focuses
on separate phenotypes has been previously suggested to be adapted in genetic
studies with DD (Williams et al. 2011b). However, this results in a rather catalog-
like presentation. For more condensed results of the systematic review, see
Appendix 1 and the article (I).
2.7.1 Vitamin D receptor
The polymorphisms in the Vitamin D receptor gene were the first ones associated
with disc degeneration. A quantitatively assessed decrease in disc signal was
originally reported to associate with the Ff, ff, Tt and tt genotypes (Videman et al.
1998). The original study along with six other studies was eligible for review
(Cheung et al. 2006, Eser et al. 2010, Eskola et al. 2010, Kawaguchi et al. 2002,
Nunes et al. 2007, Videman et al. 2001). The reports focus on two distinct exon
single nucleotide polymorphisms (SNP), FokI (rs10735810) and TaqI (rs731236),
the latter of which has been more extensively studied as described in more detail
below. One study was found where an ApaI polymorphism had been studied
without any positive associations (Kawaguchi et al. 2002).

59
Intervertebral disc (IVD) signal changes
Initially the FokI minor allele f was associated with decreased disc signal among
middle-aged Finnish male twins (Videman et al. 1998). Data in this early genetic
study is not presented in a way that easily allowed for post hoc OR calculations
nor are allele frequencies given. The TaqI minor allele t was similarly originally
associated with decreased disc signal and reported in a similar way (Videman et
al. 1998). The minor allele frequency (MAF) was close to 0.34 calculated from
another study, which likely used the same population of origin (Videman et al.
2001). Subsequent Asian studies have reported positive associations between the
TaqI t allele and disc signal summary scores among Chinese (OR 2.61 [CI 1.15–
5.90]) and Japanese populations (Cheung et al. 2006, Kawaguchi et al. 2002).
The subjects of the former study were mostly females aged 20–29 years, while
age structure of the latter population was broader, 18–55 years, and gender not
reported. A trend to greater effect was seen among the Chinese when using a
younger 18–40-year-old subgroup (OR 5.97 [CI 1.69–21.15]). However, no such
effect for the t allele or the f was seen among Danish schoolchildren (III, IV). In
both Asian studies the MAF was substantially smaller (ca. 0.13) than that among
Caucasians (ca. 0.34) (Videman et al. 2001). In another Japanese study of elderly
women (N=60), semiquantitatively assessed DD was not associated with either of
the TaqI alleles (Oishi et al. 2003). A recent study of a clinical population of fairly
young age (mixed gender; aged 20–30 years) from Turkey reported no significant
differences in allele frequencies of the total population (Eser et al. 2010).
Statistically significant differences were seen, however, among small subgroup
analyses: the FF genotype was associated with subjects having Grade 0 or 1 discs
(i.e. normal discs) using Schneiderman´s classification (Schneiderman et al.
1987). Lastly, the Turkish study reported no significant association in the total
population while the TT genotype was more common among LBP subjects with
mild signal changes (Eser et al. 2010).
IVD contour
The original study (Videman et al. 1998) reported an association between a sum
score of degenerative findings (bulges, signal changes and height) and the FokI f
allele as well as an association between the TaqI t allele and disc bulges in T6–
T12 levels in a Finnish population. The association between lumbar level IVD
bulges and the t –allele was strengthened with the Japanese and the Chinese

60
studies (OR 2.85 [CI 1.08–6.14]) (Cheung et al. 2006, Kawaguchi et al. 2002).
The same allele was controversially associated with reduced risk for disc bulges
in levels T12–S1 and L1–L4 in the Finnish population (Videman et al. 1998,
Videman et al. 2001). The Turkish study again reported a significant association
with herniations based only on a subgroup analysis (Eser et al. 2010).
Other findings
A Brazilian study with clinical patients (N=66) from Sao Paólo characterized with
discopathia and unspecified degenerative findings in MRI reported a positive
association with the FokI Ff genotype. The controls (N=88) for the study were
only clinically assessed and recruited from a different geographical area some 400
km away (Nunes et al. 2007). ATs were significantly associated with the TaqI t -
allele among the Finnish male twins, but the result was non-significant after
multiple testing corrections (Videman et al. 2001). Neither TaqI nor FokI were
associated to Modic changes among Finnish males (Karppinen et al. 2008). Four
papers were excluded from the review either because of different phenotype
(spondylosis, spinal stenosis) or phenotyping method (x-ray) (Jones et al. 1998,
Jordan et al. 2005, Koshizuka et al. 2006, Noponen-Hietala et al. 2003).
2.7.2 Collagen IX genes
Experiments with transgenic mice led to two collagen IX gene examinations with
Finnish sciatica patients (Annunen et al. 1999, Paassilta et al. 2001). In addition
to these, seven more studies were identified and found eligible for the review
(Higashino et al. 2007, Jim et al. 2005, Kales et al. 2004, Seki et al. 2006,
Solovieva et al. 2002, Videman et al. 2009). Collagen IX gene polymorphisms
were also investigated in the present studies (II, III, IV).
Sciatica
The first examinations were made on Finnish sciatica patients with unilateral
discogenic pain radiating from the back to below the knee that lasted at least one
month. The majority of the patients had a confirmed disc herniation mainly at the
two lowest levels. In the first study a Trp to Gln substitution was identified in the
α2 chain of collagen IX (Trp2 allele) in six out of 157 Finnish patients aged 19–
78 years, but in none of the 174 mixed (population and other disease patients)

61
controls (Annunen et al. 1999, Paassilta et al. 2001). A family based approach
was also used in the primary study and the results showed that all the family
members with the allele had LDD. This finding was supported by a family-based
approach and by another report from the same subject group reviewing
degenerative phenotypes of the cases (Karppinen et al. 2002). However, a Greek
study found that the Trp2 polymorphism did not exist in their population at all
(Kales et al. 2004). Seki et al. (Seki et al. 2006) further studied the Trp2 allele in
a larger sample (N=1128). Their results showed that Trp2 was common (MAF >
0.10) in the general Japanese population, but that it was under-represented among
the Japanese sciatica patients. Instead, they found a positive association between
severe disc degeneration and a different haplotype (“221” from –2066C>A,
c.977A>G, IVS19+1147G>C) not in linkage disequilibrium (LD) with the Trp2
(Seki et al. 2006). Even after phenotypic sub-grouping, the Trp2 did not show any
association (Seki et al. 2006).
Besides the COL9A2 gene, the two other type IX collagen genes, COL9A1
and COL9A3, were analysed in a slightly expanded set of 171 Finnish patients
with sciatica and 321 mixed unrelated controls (healthy N=186, other
musculoskeletal disease N=135) aged 21–75 years (Paassilta et al. 2001). This
resulted in the identification of another Trp allele (Trp3, rs61734651) in the
COL9A3 gene. Trp3 was found in 12.2% of the patients aged 19–78 years while
only 4.7% of the controls carried the allele (p=0.000013). The risk for the disease
was increased nearly 3-fold in individuals with at least one Trp3 allele. Similar to
the prevalence differences of the Trp2 allele, the Trp3 prevalence is different
between populations. Trp3 was absent among subjects from Southern China and
Japan (Higashino et al. 2007, Jim et al. 2005) and not as common as in Finland
among cases (8.6%) and controls (4.9%) from Greece (Kales et al. 2004).
IVD signal changes
The first independent association study between IVD signal changes and Trp2
was reported from a Chinese volunteer population (N=804) aged 18–55 years that
consisted of mostly males (60%). However, there was no significant difference
between the Trp2 alleles or haplotypes (constructed of Trp2, E21 + 9 G>A, IVS30
+ 17 g>a and IVS27-61 c>a) in the total population (Jim et al. 2005). Allele
frequency was again different from other populations as the Trp2 was present in
20% of the population and approximately the same in the controls. However, a
sub-group analysis of individuals aged 40–49 (N=436) found an association

62
between disc degeneration and Trp2 (OR 2.38 [CI >1.00–<5.00]). A trend towards
more severe degeneration in Trp2 carriers was also seen. A similar trend was seen
in a sub-group of a small-scale study comparing the disc signals of 37 Japanese
surgery cases aged less than 40 years (Higashino et al. 2007). The Trp2 allele was
seen in three out of 135 Finnish middle-aged men, and all three had dark nucleus
pulposus and two had single level bulges. In the same study the Trp3 allele was
not an independent risk factor for decreased disc signal (Solovieva et al. 2006).
The present studies among Danish children (III, IV) found no significant
differences between subject groups mainly divided according to IVD signal
changes. The Trp3 allele was a bit more common among subjects without DD, but
not significantly (III). In the present study of Finnish young adults (II) neither
Trp2 nor Trp3 associated with IVD signals. A positive association from a SNP
(rs696990) upstream of the COL9A1 was reported in one Finnish study. The
rs696990 associated with decreased IVD signal at L1–L4 (not in L4–S1) and with
bulges at L4–S1 (not in L1–L4) (Videman et al. 2009).
IVD contour
In a recent study with Finnish subjects (Videman et al. 2009), bulges were
significantly associated with the COL9A2 rs7533552 G-allele at L1–L4 levels
(p=0.036), but not at L4–S1. However, a significant deviation from the Hardy-
Weinberg equilibrium was seen (p=0.0286). The study’s subjects consisted of 588
Finnish male MZ and DZ twins (35–70 years old).
Other findings
Solovieva et al. have also studied the additive effects of gene variants and
environmental factors among 135 occupationally active men. Trp3 was found to
be a risk factor for dark NP (OR 7.0 [CI 1.3–38.8]) and joint occurrence of
degenerative changes (OR 8.0 [CI 1.4–44.7]), but only in the absence of the IL1B
rs1143634 T-allele (Solovieva et al. 2006). Furthermore, in the same sample, the
Trp3 allele and persistent obesity (BMI ≥ 25 kg/m2 from 25 years to 40–45 years)
acted synergistically to increase the risk of dark NP, posterior IVD bulge, and
decreased IVD height (Solovieva et al. 2002). These findings have not been
replicated since.

63
2.7.3 Other collagen genes
The associations between IVD collagen gene polymorphisms and DD phenotypes
have been studied beyond collagen IX SNPs. The search identified five such
studies (Bei et al. 2008, Eskola et al. 2010, Mio et al. 2007, Solovieva et al. 2006,
Videman et al. 2009). Additionally, some collagen gene SNPs were included in
the present investigations (II, III, IV).
Sciatica
The T-allele of rs1676486 in COL11A1 was associated with unilateral sciatica in
Japanese individuals (OR 1.42 [1.23–1.65], p=0.0000033) (Mio et al. 2007). All
the cases (N=823) had LDH. The finding has not been replicated, but the original
study consisted of three groups of Japanese subjects, and a significant difference
in the gene expression was also reported. Furthermore, no difference was seen in
allele frequencies of 188 cases with IVD signal changes and 179 controls (Mio et
al. 2007). A Greek study (N=90) investigated COL1A1 by comparing a 4-base
insertion in the gene. Subjects consisted of 24 army recruits with unilateral leg
symptoms below the knee with a compatible disc prolapse whose allele carriage
was compared to 66 healthy individuals. No association was observed (Bei et al.
2008).
IVD signal changes
A variation (not well specified) in the COL2A1 gene, coding for a major
component of the IVD, was reported in one Finnish study without any significant
difference (Solovieva et al. 2006). The present studies with Danish children (III,
IV) analysed a variation in COL11A2 (rs1799907) and reported no association,
which is in contrast to results of a study of elderly people in Finland with spinal
stenosis (Noponen-Hietala et al. 2003). Among Finnish twin males aged 35–70
years, a different polymorphism (rs2076311) in COL11A2 was associated with
disc signal decrease in L4–S1, as was rs2075555 in COL1A1 (Videman et al.
2009).

64
IVD contour
A study with occupationally active middle-aged Finnish males (N=135) included
also a SNP in intron 9 of COL11A2, and a borderline association between IVD
bulges and the A allele was seen (OR 2.1 [CI 1.0–4.2], p=0.04) (Solovieva et al.
2006). The study using Finnish twin males reported associations between
COL11A1 gene polymorphisms (rs1463035 and rs1337185, in the same linkage
disequilibrium block) and disc bulges (Videman et al. 2009).
2.7.4 Aggrecan
The first report identified, in which the association of a polymorphism in the
aggrecan gene (ACAN) with DD was analysed, was done in Japan (Kawaguchi et
al. 1999). Eight other studies were identified (Cong et al. 2010a, Cong et al.
2010b, Eser et al. 2010, Mashayekhi et al. 2010, Roughley et al. 2006, Solovieva
et al. 2007, Videman et al. 2009).
IVD signal changes
A study on multilevel DD carried out in Japan (N=64) investigated a genetic
polymorphism in a variable number of tandem repeats (VNTR), with an
overrepresentation of rare shorter alleles (A18 and A21) in cases with multilevel
DD (Kawaguchi et al. 1999). The finding was not replicated in a Canadian study
(N=102) (Roughley et al. 2006). However, findings of a very recent Korean study
(N=104) partially agree with the initial findings. The A21 allele was significantly
overrepresented in subjects with multilevel disc degeneration (p<0.006) (Kim et
al. 2011). Solovieva et al. studied the aggrecan VNTR among the Finnish
working males (N=135). Carrying two copies of the A26 allele increased the risk
of dark NP (OR 2.77 [CI 1.24–6.16]) (Solovieva et al. 2007). Cong et al. studied
Chinese individuals, a cohort subgroup with chronic LBP including also trauma
patients. Inclusion was restricted to males aged 14–49 years. Disc degeneration
was determined using Schneiderman’s classification, where Grade 1 indicates
normal discs and Grades 2 and 3 different degrees of degeneration. Subjects who
smoked more than one pack-year and carried one or two alleles with ≤25 repeats
were found to have an increased risk for symptomatic DD (OR 4.5 [1.589–
12.743]) (Cong et al. 2010b). In addition to the aforementioned findings, the
associations between IVD signal decrease and different allele lengths were

65
somewhat contradictory in a Turkish population aged 20–30 years (N=300) with
the statistically most significant association between multilevel DD and the A27
allele (Eser et al. 2010). Videman et al. reported in the aforementioned twin study
(N=558) significant associations between ACAN rs1042631 and quantitative IVD
signal intensity decrease (at L1–L4) (Videman et al. 2009).
IVD contour
Associations between ACAN alleles and LDH were also investigated in the
Turkish study (N=300). However, no clear association was identified, even
though the authors saw some statistically significant allele distribution
characteristics in a sub-group of protrusion type LDHs (Eser et al. 2010). In a
previous Finnish study, the carpenters and machine drivers with the A26 allele
had a statistically significantly higher risk of degenerative changes (disc bulge
and decreased disc height) than non-carrier office workers (OR 2.91 [CI 1.25–
6.77] and OR 3.38 [CI 1.29–8.87], respectively) (Solovieva et al. 2007).
Additionally, the rs1042631 and rs1516797 in ACAN were associated with
qualitative disc bulging (at L4–S1) and qualitative disc height reduction,
respectively, among the Finnish male twins (Videman et al. 2009).
Other findings
In a recent Iranian study, IVDs at L4/L5 and L5/S1 were evaluated using
decreased signal intensity of NP, posterior IVD bulges and decreased IVD height
as a pooled definition of DD. A total of 71 mixed-gender adult patients and 108
controls were analysed. Carriers of shorter VNTR alleles (24 or fewer repeats)
were more frequently seen among patients than among controls (OR 3.28 [CI
1.62–6.65]) (Mashayekhi et al. 2010). Noponen-Hietala et al. found no
association between ACAN alleles and spinal stenosis in a Finnish study (N=85)
with elderly subjects (Noponen-Hietala et al. 2003).
2.7.5 Cartilage intermediate layer protein
Human osteoarthritic articular cartilage from the hip was used in the
characterization of cartilage intermediate layer protein in 1997 (Lorenzo et al.
1998). The first genetic association study between polymorphisms in the
corresponding gene, CILP, and DD was published in 2005 (Seki et al. 2005).

66
Three other studies reporting on the same gene were found (Min et al. 2009, Min
et al. 2010, Virtanen et al. 2007a). The CILP rs2073711 was also investigated in
the present study with Finnish young adults (III).
Sciatica
The first association study included Japanese individuals with symptomatic disc
disease (N=467, about 85% underwent spinal surgery) and controls (N=654). Of
the 20 SNPs studied, the + 1184T>C (rs2073711) was shown to be the most
significantly associated with the disease (OR 1.61 [CI 1.31–1.98], p=0.00020
after correction for multiple testing) (Seki et al. 2005). Functional analysis
showed that the C allele was associated with increased binding and inhibition of
TGF-β1, suggesting that regulation of TGF-β1 signalling by CILP has an
important role in symptomatic disc disease (Seki et al. 2005). However, two
subsequent studies in Finnish and Chinese samples (N=502 and N=691,
respectively) failed to replicate the association (OR 1.35 [CI 0.97–1.87] and OR
1.05 [CI 0.77–1.43], respectively) (Virtanen et al. 2007a).
IVD signal changes
The aforementioned negative replication study of rs2073711 from China (N=691)
was population based and DD was defined using Schneiderman’s classification
(Jim et al. 2005, Virtanen et al. 2007a). However, another Japanese research
group studied the CILP rs2073711 and DD defined using Pfirrmann grading
(Pfirrmann et al. 2001). The subjects were Japanese collegiate high-level judo
athletes (N=89, all young males). Despite the small sample size, analysis resulted
in a significant association between DD and the rs2073711 minor allele C (OR
4.1 [CI 1.57–10.71]) (Min et al. 2009). The same authors studied the SNP in a
larger sample (N=601) of collegiate athletes again using Pfirrmann grading, and
used grades 1 and 2 as the control group while grades 3, 4 and 5 were defined as
DD, as previously. The results indicated an increased risk for DD with the C allele,
but with a smaller effect size (OR 1.4 [CI 1.05–1.86]) (Min et al. 2010). However,
a clear difference between genders was seen (males OR 1.8 [CI 1.27–2.59],
females OR 0.9 [CI 0.57–1.52]). In the present study with Finnish young adults
(II), the CT/TT genotypes, compared to the CC genotype, increased risk for DD,
but only among women (OR 2.04 [CI 1.07–3.89], p=0.025 (corrected p=0.05)
(Kelempisioti et al. 2011). This finding is in contrast to the Japanese findings

67
(Min et al. 2009, Min et al. 2010, Seki et al. 2005), but also the minor allele is
different; T in Europeans and C in Asians.
2.7.6 Asporin
Findings with CILP led to new searches of DD candidates especially on the TGF-
β signalling pathway. Asporin, another extracellular protein, is one of the small
leucine-rich proteoglycans of IVD playing an important role in cartilage
homeostasis. A unique aspartate (D) repeat in the asporin gene (ASPN) was earlier
found to associate with OA of the knee (Kizawa et al. 2005). OA and DD are both
degenerative diseases of skeletal joints, and many of the genes expressed in
cartilage are also expressed in the IVD.
Sciatica
The search identified only one association study with ASPN D-repeats (Song et al.
2008a). However, the initial study consisted of two independent cohorts of
Japanese and Chinese origin. The Japanese cohort was an extension of the
previous study with CILP (Seki et al. 2005). Moreover, in this study all cases
(N=745) had a history of sciatica for longer than three months and disc herniation
confirmed by MRI. The association between the presence of at least one D14
allele and disease was found to be significant (OR 1.78 [CI 1.26–2.51],
p=0.00087). The D14 allele frequency was 7.8% in cases and 4.8% in controls
(Song et al. 2008a).
IVD signal changes
The same report included also a Chinese population (N=1055) in which the IVDs
were graded according to Schneidermann’s classification. The subjects were then
divided into two groups (cases and controls) using adjustment for age. The
association between IVD signal changes and the presence of at least one D14
repeat was found to be significant (OR 1.66 [1.17–2.35], p=0.0039). The D14
allele was also infrequent among the Chinese, with a frequency of 9.3% in cases
and 6.4% in controls (Song et al. 2008a).

68
Other findings
The authors also did a meta-analysis using the abovementioned phenotypes and
cohorts. The meta-analysis showed significant associations both for the allele
frequency (OR 1.58 [CI 1.26–1.99], p=0.00081) and “D14/D14+D14/other vs.
other/other” genotype comparison (OR 1.70 [CI 1.35–2.20], p=0.000013) (Song
et al. 2008a).
2.7.7 Matrix metalloproteinase genes
The enzymes linked with remodelling of the extracellular matrix have also been
investigated in association studies with disc degeneration. Matrix
metalloproteinase 3, also known as stromelysin-1, was previously associated with
cardiovascular problems (Ye et al. 1996), and a few years later with DD
(Takahashi et al. 2001). The search identified a total of five eligible studies
focusing on matrix metalloproteinase (MMP) gene polymorphisms (Dong et al.
2007, Karppinen et al. 2008, Song et al. 2008b, Sun et al. 2009, Takahashi et al.
2001). The present study with Finnish young adults also investigated MMP
polymorphisms in MMP9 and MMP10 (II).
Sciatica
Dong et al. reported a positive association between MMP2 -1306C/T (rs243865)
and sciatica/discogenic LBP in a case (N=162) vs. control (N=318) setting. The
participants of the study were Chinese young adults. Subjects with the CC
genotype had nearly threefold increased risk for sciatica/discogenic LBP (OR
3.08 [CI 1.84–5.16]) compared to subjects carrying at least one variant T allele
(Dong et al. 2007). A Japanese study included subjects (N=847) with lumbar disc
herniation and positive sciatica history and compared them with controls (N=896).
After genotyping 15 tagSNPs and resequencing exons and exon borders of MMP9,
the rs17576 showed the most significant association with LDH (OR 1.29 [CI
1.12–1.48]) (Hirose et al. 2008). The finding has not yet been replicated. Another
Chinese study compared the MMP9 -1562C/T (rs3918242) polymorphism
between cases (N=458) and controls (N=451). The cases had herniation/bulge and
discogenic LBP/sciatica while the controls were not examined by MRI and
included both patients with different diseases and healthy individuals. All subjects
were Hans from Northern China. The subjects with at least one minor allele (TT +

69
CT) had a significantly increased risk of LDD (OR 2.14 [CI 1.55–2.96])
compared with the CC genotype subjects (Sun et al. 2009).
IVD signal changes
The first study on MMP2 in Japan consisted of 103 individuals (Takahashi et al.
2001). The total disc degeneration score was defined by MRI among young
individuals but only x-ray was used in elderly subjects. The 5A genotype (a run of
only five instead of six adenosines in the promoter region of the gene) was
associated with the number of degenerated discs; however this finding was only
significant among the elderly (Takahashi et al. 2001). The aforementioned
Chinese study by Dong et al. analysed also IVD signal changes and the MMP2 -
1306C/T (rs243865) in 133 patients. The patients with a CC genotype had more
severe DD (Dong et al. 2007). A polymorphism at the MMP1 promoter -1607
rs1799750 was examined in another Chinese population of volunteers (N=600)
aged 18–55 years (Song et al. 2008b). The deletion allele resulted in a
significantly higher risk for degeneration (OR 1.41 [CI 1.04–1.90]) defined also
by Schneiderman's classification (Song et al. 2008b). The study with North
Chinese Hans (N=809) by Sun et al. examined also the association between IVD
signal changes and the MMP9 -1562C/T (rs3918242). The total degenerative
score using Scheiderman’s classification (0–15) was ca. 1.7 points higher among
patients carrying at least one T allele compared to the patients with the
homozygous CC genotype (Sun et al. 2009). The MMP9 rs17576 T-allele shown
to associate with sciatica in the Japanese study did not associate positively with
IVD signal changes in young Finnish adults (II) (N=396) (Kelempisioti et al.
2011).
Other findings
Ten polymorphisms from the genes MMP3 (rs645419, rs646910), MMP8
(rs1940475, rs2509013, rs1276283), MMP9 (rs3918241, rs2664538, rs20544) and
MMP13 (rs2252070, rs3819089) were studied among middle-aged Finnish twin
males (Videman et al. 2009) while the 5A/6A polymorphism of the MMP3 was
studied in subjects with spinal stenosis (Noponen-Hietala et al. 2003). No positive
association with decreased disc signal, disc bulging or decreased disc height
(Videman et al. 2009) or spinal stenosis (Noponen-Hietala et al. 2003) was found.

70
The results of MMP3 analysed together with another polymorphism (Karppinen et
al. 2008) are presented in the following chapter on interleukins.
2.7.8 Interleukin genes
Interleukin-1 (IL1) was the first interleukin cloned in the 1980s after which it was
shown to be an important element in the regulation of inflammation (Gabay et al.
2010). Inflammation is considered a key factor in pain – probably because it has
been implicated also in degenerative processes (DeLeo 2006, Goldring et al. 1994,
Loeser 2006, Shinmei et al. 1989). Polymorphisms in the two distinct IL1 genes
were the first interleukin SNPs associated with DD (Solovieva et al. 2004). The
systematic search identified also nine other studies analysing interleukin
polymorphisms in disc degeneration phenotypes (Eskola et al. 2010, Karppinen et
al. 2008, Karppinen et al. 2009, Kim et al. 2010, Lin et al. 2011, Noponen-
Hietala et al. 2005, Paz Aparicio et al. 2010, Solovieva et al. 2006, Videman et al.
2009).
Sciatica
In a Finnish study by Noponen-Hietala et al. ten cytokine candidate genes were
analysed in 155 unrelated sciatica patients (males and females, aged 19–78 years).
The polymorphism distributions were compared to 179 sedentary workers and
students (males and females, aged 20–69 years) (Noponen-Hietala et al. 2005).
The allele frequencies of interleukin-6 gene (IL6) promoter polymorphisms G-
597A (rs1800797), G-174C (rs1800795), T15A (rs13306435) differed
significantly between the two groups. Furthermore, a relatively rare haplotype
GGGA (derived from G-597A (rs1800797), G-572C (rs1800796), G-174C
(rs1800795) and T15A (rs13306435) polymorphisms) was significantly
associated with the disease (OR 4.80 [CI 1.59–14.45]). The other polymorphisms
in the genes IL1A, IL1B, IL1RN, IL2, IL4, IL4R, IL10, IFNG, and TNFA did not
differ significantly between the two groups (Noponen-Hietala et al. 2005). A
recent study with Chinese subjects (N=281) of both genders evaluated the
relationship between an 86-bp VNTR polymorphism in the interleukin 1 receptor
antagonist gene (IL1RN) and the DD phenotype. All cases had single level
paracentral and/or lateral recess subligamentous LDH with definite neural
compression on T2-weighted MRI (Kim et al. 2010). The prevalence of the A3
allele was significantly higher among the patients (N=54) compared to controls

71
(N=227) found healthy without MRI scans (OR 3.86 [CI 1.37–10.90]) (Kim et al.
2010). The same polymorphism presented only alleles A1 and A2 (which were
evenly distributed) among Finnish cases and controls (Noponen-Hietala et al.
2005).
IVD signal changes
Multiple interleukin SNPs were investigated in the Finnish study with male twins
(Videman et al. 2009). The rs2071375 in IL1A (L1–L4 levels) and rs1420100 in
IL18RAP (both upper and lower lumbar spine) were associated with quantitative
IVD signal intensity (Videman et al. 2009). Among the 12–14-year-old Danish
schoolchildren (III) the rs1800587 in IL1A was associated with subjects with
early DD characterized mainly by a decreased disc signal, but only among girls.
The CT/TT genotypes, compared to CC, were more common among subjects with
DD (OR 2.85 [CI 1.19–6.83]). The IL6 promoter polymorphisms showed results
similar to that of the relatively rare haplotype GCG (derived from G-597A
(rs1800797), G-572C (rs1800796), G-174C (rs1800795), which associated with
degeneration, but only among girls (OR 6.46 [CI 1.61–26.0]) (III). In the present
study with Finnish young adults (II) the G allele of both IL6 rs1800797 and IL6
rs1800795 was the risk allele for decreased IVD signal (OR 1.37 [CI 1.02–1.85]
and OR 1.45 [CI 1.07–1.96], respectively. In another Finnish report, the absence
of the IL1B T-allele with carriage of COL9A3 Trp3 was associated with increased
risk for DD (Solovieva et al. 2006). A recent Chinese study reported significant
differences in IL10 -592A>C (rs1800872) and -1082G>A (rs1800896) between
cases (N=320) and controls (N=269). The risk allele A for both (OR 1.68 [CI
1.23–3.46] and OR 1.34 [CI 1.12–3.03], respectively) was associated also with
decreased IL-10 mRNA expression (p<0.05) (Lin et al. 2011).
IVD contour changes
The initial association study with the interleukins done using Finnish subjects
reported an association with IL1 polymorphisms (Solovieva et al. 2004).
Occupationally active males with disc bulges were more often carriers of the T
alleles of rs1800587 and rs1143634 in IL1A (OR 2.4 [CI 1.2–4.8]) and IL1B (OR
1.9 [CI 1.0–3.7]), respectively (Solovieva et al. 2004). Furthermore, the joint
effect of being a carpenter instead of machine driver or office worker increased
the risk even further. The joint effect was seen also for the IL-1RNC1887

72
polymorphism (Solovieva et al. 2004). A recent Spanish study analysed the
genetic association of polymorphisms in the cytokine and nitric oxide synthase
genes in relation to LDH. The study included LDH patients (N=50). The controls
(N=129) were patients admitted for primary lower limb arthroplasty. The IL1B
(rs1143634) minor allele C was detected more frequently in the group of patients
with LDH than in controls (OR 1.7 [CI 0.98–2.93], non significant) (Paz Aparicio
et al. 2010). The risk allele in the Spanish study (C) was the opposite of the
Finnish (T) (Solovieva et al. 2004). However, in another report of the same
Finnish cohort, the absence of the IL1B T allele with carriage of COL9A3 Trp3
was associated with increased risk for DD (Solovieva et al. 2006).
Other findings
The first study to evaluate genetic factors in relation to Modic changes was
published only a few years ago (Karppinen et al. 2008). The study with male train
engineers exposed to whole body vibration (N=159) and male paper mill workers
(N=69) included thirteen SNPs from eight previously identified putative risk
genes (COL9A2, COL9A3, COL11A2, IL1A, IL1B, IL6, MMP3 and VDR), but
found no significant association. However, when the rs1800587 in IL1A and the
5A/6A polymorphism in MMP3 were analysed together, a significant interaction
risk for type II Modic changes was observed (OR 3.2 [CI 1.2–8.5]) (Karppinen et
al. 2008). The haplotype T-C of IL1A rs1800587 and IL1B rs1143634 combined
with MMP3 5A (rs3025058) synergistically increased the risk of Modic type II
(OR 8.1 [CI 1.7–38.4]) (Karppinen et al. 2008). In another Finnish study, six
single SNPs in the IL1 gene cluster were analysed further in the occupational
cohort of Finnish middle-aged males studied earlier (Karppinen et al. 2009,
Solovieva et al. 2004). The minor T allele of rs1800587 in IL1A was more
frequent among subjects with any type of Modic change (OR 2.50 [CI 1.09–
5.71]), the majority (N=39, 87%) of which had type II Modic changes (Karppinen
et al. 2009). A relatively rare (frequencies 0.8% vs. 7.5%) haplotype constructed
from the six SNPs was also associated with the Modic changes (Karppinen et al.
2009).
2.7.9 Findings in other genes
The systematic search and reference tracking identified also six includable recent
studies on genetic associations in DD (Hirose et al. 2008, Karasugi et al. 2009,

73
Paz Aparicio et al. 2010, Sun et al. 2011, Williams et al. 2011a, Zhu et al. 2011).
These studies have focused on variations in GDF5, THBS2, SKT, CASP9, FAS,
FASLG, NOS2 and NOS3.
Sciatica
The aforementioned Japanese study with MMP9 first examined two
thrombospondin genes (THBS1 and THBS2) as candidate genes for LDH (Hirose
et al. 2008). Multiple polymorphisms of the THBS2 were associated with LDH
further characterized by a history of unilateral leg pain. The study size was
sufficient (N=847 cases and N=896 controls) and supportive evidence for the
involvement of THBS2 in LDH was found through biological studies (Hirose et
al. 2008). This very interesting and remarkable finding has not yet been replicated.
The present study (II) with Finnish young adults (N=396) did not find any
significant difference in allele frequencies or genotypes between groups defined
according to the presence or absence of moderate DD using Scheiderman´s
classification (Kelempisioti et al. 2011). Another very recent study reported an
association between yet another gene, SKT (KIAA1217), and LDH (Karasugi et al.
2009). Of the 68 SNPs studied, the rs16924573 was the most significantly
associated with LDH among Japanese subjects (OR 1.31 [CI 1.11–1.55]) while
the finding was also replicated among Finnish subjects (OR 2.81 [CI 1.09–7.24])
(Karasugi et al. 2009). The allele frequencies were quite different between
Finnish and Japanese populations, but nevertheless the meta-analysis of over
2200 subjects supported the association (OR 1.34 [CI 1.14–1.58]) (Karasugi et al.
2009). SKT rs16924573 is also included in the present study with Finnish young
adults (II).
IVD contour
Among the most recent candidate genes for disc degeneration phenotypes is
Caspase 9 (CASP9), which has been suggested to have a central role in initiating
the intrinsic apoptotic pathway (Li et al. 1997). The polymorphism Ex5+32 G>A
(rs1052576) was analysed in Chinese subjects with LDH and unspecified clinical
symptoms (N=387) and compared to matched healthy controls (N=412) (Sun et al.
2011). The minor allele A was associated with LDH (OR 1.91 [1.29–2.81]).
Furthermore, the patients with the AA genotype had a significantly more severe
DD compared to subjects with the GG genotype (Sun et al. 2011). A Spanish

74
study reported an association between a NOS2 polymorphism (exon22 G>A,
rs1060826) and LDH. The AA genotype was absent in cases (N=50) while the
frequency was 14.7% in controls (N=129). Additionally, the CC genotype of a
polymorphism in NOS3 (–786 T/C, rs2070744) was overrepresented in controls
(OR 0.12 [0.03–0.43]) (Paz Aparicio et al. 2010).
Other findings
A recent study included Chinese Hans patients with chronic LBP (N=348) and
healthy controls (N=215). Minor alleles A and T in FAS −1377G/A (rs2234767)
and FASL (rs763110) −844C/T associated with the disease (OR 1.411 [CI 1.096–
1.816], p=0.007 and OR 2.773 [2.104–3.654], p<0.001), respectively (Zhu et al.
2011). Another recent study investigating a SNP (rs143383) in GDF5 was found
by reference tracking (Williams et al. 2011a). Out of the total population
(N=5269), one cohort (N=613) was examined using MRI therefore making the
study eligible for review. The rs143383 showed a significant association among
female subjects for a combined phenotype of disc space narrowing and
osteophytes (OR 1.72 [CI 1.15–2.57]) (Williams et al. 2011a). However, the result
seems to be dependent mainly on the osteophytes in radiographs; a positive
association was not observed in the cohort examined using MRI. Still, the large
number of subjects increases the credibility of association for this phenotype.
2.8 Linkage analysis studies in disc degeneration
To date three linkage analysis studies have been reported in DD (Table 5). An
Italian group reported a preliminary linkage analysis in 2002. The study was
performed with subjects from only one family and consisted of eight siblings in
total. The phenotype in this possibly rare Mendelian form of DD included LDH at
multiple levels and paraplegia while varying among the affected four individuals.
It is possible that the underlying aetiology of this specific phenotype is different
from that of the sporadic, age-related DD. Nevertheless, the analysis reported an
association on chromosome 6 (Zortea et al. 2002).
The first multifamily linkage analysis was performed in Finnish families with
a total of 189 individuals. The study included families with increased prevalence
of lumbar disc disease characterized by sciatica (LDD). Suggestive linkage was
obtained on chromosomes 4, 6 and 21, 21 being the most promising (Virtanen et
al. 2007b).

75
The most recent linkage analysis on DD has been performed in the UK. Only
female DZ twins were included in the study for a total of 348 individuals. A total
DD score was calculated for each individual after evaluation of each IVD using
MRI. The most significant linkage resulted from chromosome 19, while
chromosomes 1 and 5 showed also some interesting results (Williams et al. 2008).
In all these published linkage studies in DD the specific phenotype is
different. Therefore, when attempting to sum up everything that is known about
DD genetics, this fact must be noted. There is no validated method to adjust for
this phenotypic heterogeneity between the definitions of DD because the
pathogenic cascade and it´s relation to normal aging is not understood well
enough (see chapter 2.4).
Table 5. Linkage analysis studies in disc degeneration.
Ethnicity Study size Phenotype Chromosomes Reference
Italian 8 LDH + paraplegia 6 Zortea et al. 2002
Finnish 189 LDD + sciatica 4, 6, 21 Virtanen et al. 2007
UK 384 DD summary score 1, 5, 19 Williams et al. 2008
2.9 Genome-wide association studies in disc degeneration
At the time of preparation of this thesis (October 2012) the first GWAS study in
DD had just been published (Williams et al. 2012). The authors of this study
developed a continuous DD trait based on IVD space narrowing and formation of
osteophytes. Notably, this specific phenotype can be defined on multiple imaging
modalities (plain X-ray, CT and MRI). The different cohorts of this GWAS with
meta-analysis included 4683 individuals with European ancestries and the
genotypes for 2.5 M SNPs (using imputation based on 100K–500K SNP data).
The mean age was 57.7 years and the majority (67%) of the subjects were women.
Genome-wide significance (unadjusted) resulted for four markers (rs17034687,
rs2187689, rs7767277, rs926849) in chromosomes (Chr) 3 and 6. The
rs17034687 on Chr 3 is an intergenic marker that is not in LD with any known
gene-based marker. The rs926849 is in intronic region of the Parkinson protein 2,
E3 ubiquitin protein ligase (PARK2) gene on Chr 6. The rs2187689 and
rs7767277 lie also on Chr 6 and are in perfect LD. Both of these SNPs are in
strong LD with a proteasome subunit, β type 9, large multifunctional peptidase 2
gene (PSMB9) that is located in the class II region of the major histocompatibility

76
complex (MHC). None of these four SNPs are in LD with known functional SNPs
in either PARK2 or PSMB9 (Williams et al. 2012).
The previously identified associations in studies using candidate approaches
were not confirmed in this first unbiased GWAS approach. However, the
phenotypes between the studies are different. Notably Chr 6 has been indicated
previously in studies with the linkage approach (Table 5).
2.10 Summary and research problem
Previous knowledge of genetic risk factors of DD is limited and scattered while
there is a lack of a gold standard in the coding of MRI images for degenerative
change. Nevertheless, DD is shown to be clinically important. The cascade of DD
is not completely understood, but it is known that DD begins already at a young
age. Despite the fact that DD has multifactorial aetiology, the inherited
component seems to be predominant. However, also in heritability studies the
importance of rigorous phenotyping has appeared as heritability estimates for
IVD signal intensity have varied from 0 to 50% when different phenotyping
methods have been used. Studies in DD have focused on adults while genetic
association studies on disc degeneration progression (DDP) have not been
published before. Previous literature suggests that early DD represents a
particularly important phenotype; with aging the prevalence of age-related IVD
changes increases as well as cumulative effects of predisposing environmental
factors e.g. physical workload and smoking (pack years) thus investigations in
young individuals are needed. By identifying new genetic associations with
different DD traits potential new pathogenic pathways might be found. This can
lead to efficient identification of individuals predisposed for back diseases or new
treatments for these diseases in the future. Additionally, systematic analyses of
previously published information are crucial in order to direct future efforts.

77
3 Outlines of the present study
Despite the fact that genetic analyses have gained a more central role in the
exploration of the underlying causes of lumbar disc degeneration (DD), its
aetiology and pathogenesis have not been elucidated. The present study aimed to
refine the knowledge on DD by systematically analysing all known genetic
association studies in DD defined by MRI and to increase the knowledge of the
genetic component in early DD by investigating relevant candidate gene
polymorphisms in two different study cohorts consisting of young individuals.
The included polymorphisms and genes have been earlier found to associate with
some DD trait or an associated condition in adults.
The more specific aims of the present study were:
– to systematically review previously published genetic association studies in
human lumbar disc degeneration (DD) and analyse the evidence level of these
associations
– to investigate the possible associations between genetic polymorphisms in
ADIPOQ, CILP, COL9A2, COL9A3, COL11A2, COL22A1, IL1A, IL1B,
IL1F10, IL6, LEPR, MMP9, MMP10, SKT, THBS2, VDR and moderate DD in
a well-defined and characterized cohort of young Finnish adults
– to investigate the possible associations between genetic polymorphisms in
COL9A3, COL11A2, IL1A, IL1B, IL6 and early DD and progression of early
DD (DDP) over a 3-year period in a Danish teenager population
Based on the aims, specific research questions (Q) were formed. These questions
were investigated and answered in original articles as defined in Table 6. The
questions were:
– Q1: What is the current cumulative evidence level and current clinical
relevance of previously identified associations between genetic markers and
lumbar DD on MRI?
– Q2: Are there associations between selected candidate gene polymorphisms
and lumbar DD on MRI among Finnish adolescents?
– Q3: Are there associations between selected candidate gene polymorphisms
and early lumbar DD or progression of DD on MRI among Danish teenagers?

78
Table 6. Research question investigations in the original articles.
Question I II III IV
Q1 X
Q2 X
Q3 X X
Research hypotheses
Previous literature suggests that early DD represents a particularly important
phenotype. It was hypothesised that at least some of the candidate genes
identified earlier are important in the DD cascade and polymorphisms in these
genes may be associated with DD among young individuals. However, it was also
hypothesised that some of the previously identified associations are likely to
possess only modest cumulative association credibility. It was further
hypothesised that systematic analysis of previously identified genetic association
studies can refine our understanding on the credibility of these associations.

79
4 Materials and methods
The study materials and methods are described in more detail in the original
articles (I–IV). Article I is a systematic review in which the data acquisition and
analysis differ substantially from the other articles. The key methods of original
articles I–IV are summarized in Table 7.
Table 7. Summary of key methods used in the studies.
Method Original article
Study design
Systematic literature search and analysis I
Conventional designing methods II, III, IV
muPlex oligo designer II
Laboratory
DNA acquisition
venous whole blood sample II, III, IV
DNA extraction II, III, IV
Polymerase Chain Reaction (PCR) II, III, IV
Multiplex PCR II, III, IV
Sequencing II, III, IV
Restriction enzyme digestion III, IV
SNaPshotTM II, III, IV
Agarose gel electrophoresis II, III, IV
Ultraviolet light II, III, IV
MRI imaging
1.5T unit II
0.2T open unit III, IV
Data handling software
OsiriX IV
ABI Seq Scanner & Peak Scanner & GeneMapper II, III, IV
Statistics
Chi-square test II, III, IV
SAS II, III, IV
Logistic regression II, III, IV
Linear regression II, III
SNPStats II, III, IV
PHASE II, III, IV
Markov-chain II, III, IV
Likelihood ratio test III
Bonferroni-Holm II, III, IV
Mann-Whitney U-test IV

80
4.1 Study populations
4.1.1 Finnish study population (II)
The study population consisted of members of the 1986 Northern Finland Birth
Cohort with an expected date of birth between July 1, 1985 and June 30, 1986
(N=9479) in the two northernmost provinces of Finland; Oulu and Lapland. The
2969 that lived within 100 km of the city of Oulu received a postal questionnaire
in 2003. Altogether 67% responded (N=1987, with a mean age of 18 years) and
were invited to participate in a physical examination, which took place in 2005–
2006 at a mean age of 19 years. The individuals who participated in the
examination (N=874) were invited to participate in MRI (N=563 participated).
Subjects with claustrophobia were excluded (N=5). Individuals with a DNA
sample and MRI data were included in the final genetic study (N=538). To
evaluate selection bias the MRI participants were compared with the rest of the
postal questionnaire respondents (N=1424) and the rest of individuals invited to
participate in MRI (N=311). The results of these comparisons have been
published earlier; there was some selection bias. In short, the MRI participants
were mostly females, physically more active and more likely to be non-smokers
than non-participants, and a higher proportion of them suffered from LBP
(Takatalo et al. 2009).
4.1.2 Danish study population (III, IV)
The subjects for this study were a subgroup of a Danish cohort of 771 children
sampled for the European Youth Heart Study in 1997 (Andersen et al. 2003). In
2001, the children who had previously participated in that study (N=589) and who
were still living in the Odense municipality (N=552) were invited to take part in
MRI at the age of 12 to 14 years (Kjaer et al. 2011). In all, 439 (80%) children
were scanned at the baseline time point. The whole cohort (N=771) was re-invited
in 2003 and interviewed on average 2.7 (range 2.2–3.1) years after the MRI study
baseline (Kjaer et al. 2011). As many as 443 individuals participated in the
follow-up interview about back pain and underwent new MRI scans (N=439)
within the same day. A total of 318 subjects participated in both baseline and
follow-up MRI.

81
4.1.3 Study ethics
The Ethics Committee of the Northern Ostrobothnia Hospital District approved
the study in Finland (II), and the ethics committee of the Vejle and Funen counties
approved the studies in Denmark (III, IV). Written informed consent was obtained
from all individuals (II) and also from their parents when the study subjects were
minors (III, IV). Participation has been voluntary, and the studies have been
performed according to the Declaration of Helsinki.
4.2 MRI methodology
4.2.1 Finnish young adult study (II)
MRI scans were performed using a 1.5 tesla unit (Signa, General Electric,
Milwaukee, WI) and phased array spine coil (USA Instruments, Aurora, OH,
USA) with the imaging protocol of a sagittal T1-weighted (440/14 [repetition
time msec/echo time msec]) spin echo, a sagittal T2-weighted (3960/116) fast
spin echo (FSE), and the axial T2-weighted FSE (5160/107) of the entire lumbar
spine (Takatalo et al. 2009).
Definition of DD in the study (II)
The degree of DD was graded from T2-weighted images using a modified
Pfirrmann classification: Grade 1 (normal shape, no horizontal bands, distinction
of nucleus, and annulus is clear), Grade 2 (non-homogeneous shape with
horizontal bands, some blurring between nucleus and annulus), Grade 3 (non-
homogeneous shape with blurring between nucleus and annulus, annulus shape
still recognizable), Grade 4 (non-homogeneous shape with hypointensity, annulus
shape not intact and distinction between nucleus and annulus impossible, disc
height usually decreased), and Grade 5 (same as Grade 4 but collapsed disc space)
(Pfirrmann et al. 2001, Takatalo et al. 2009). In the modified Pfirrmann
classification, hyperintensity or isointensity of the intervertebral disc to
cerebrospinal fluid (CSF) ratios were not used as criteria for Grades 1 through 3,
due to the fact that CSF was hyperintense compared to the discs in all the MR
sequences. Moderate DD was defined as Grade 4 degeneration at one or more
lumbar levels, or Grade 3 degeneration at three or more levels. Subjects with
Grade 1 and Grade 2 findings and no other pathology in MRI were defined as the

82
reference group, i.e. subjects without DD. Information on inter-rater reliability
and comparison of the MRI study population and the rest of the survey
respondents have been published elsewhere (Takatalo et al. 2009).
4.2.2 Danish teenager studies (III, IV)
MRI was performed using an open low-field 0.2 tesla MRI unit (Magnetom Open
Viva, Siemens AG, Erlangen, Germany). Axial and sagittal T2-weighted turbo
spin echo sequences were used to obtain images of the lumbar spine as described
elsewhere (Kjaer et al. 2005). All images were evaluated by an experienced
radiologist who was blinded to the gender and all health data of the subjects. The
latest international nomenclature for describing disc pathology was generally used
in the definitions (Fardon et al. 2001). The signal intensity changes of the disc in
sagittal sections on T2-weighted images was graded using a scale from 0 to 3
where 0 = homogeneous hyper-intense (white), 1 = hyper-intense with visible
intranuclear cleft (white with a dark band in the equator plane of the disc), 2 =
intermediate signal intensity (all colours between white and black), and 3 = hypo-
intense (dark disc without visible nuclear complex) (Eyre et al. 1989, Weishaupt
et al. 1998). Changes in the disc contour were described on a nominal scale: 0 =
normal, 1 = bulge, 2 = focal protrusion, 3 = broad based protrusion, 4 = extrusion
and 5 = sequestration (Fardon et al. 2001, Milette 1997). Defects in endplates
were graded: 0 = normal endplates, 1 = defects and 2 = large defects and
Schmorl’s nodes (Videman et al. 1995). ATs including radial tears and HIZ
lesions were analysed according to the existing definition (Aprill & Bogduk 1992,
Yu et al. 1988).
Of the baseline genetic study subjects (N=220, 66 with DD and 154 without),
a total of 166 participated in the follow-up. Children both with and without DD at
follow-up were defined and the follow-up status was compared to the baseline
DD status. We identified different patterns (eg. no lumbar DD – lumbar DD, no
lumbar DD – no lumbar DD, no lumbar DD – not eligible, lumbar DD – no
lumbar DD and lumbar DD – lumbar DD). Because some of the patterns seemed
strange (lumbar DD to no lumbar DD) and because there was 2.7-years between
the radiologist ratings, we compared baseline and follow-up images in 2011 to
ensure correct classification of children with and without DD. All other patterns
except for no lumbar DD – no lumbar DD (N=78) were re-evaluated in consensus
sessions by two of the authors (PK and PE) using the software OsiriX v.3.9.1 32-
bit (Pixmeo SARL, Geneva, Switzerland). We evaluated sagittal MRI images

83
from both time points simultaneously side-by-side while axial images were used
to confirm herniations, bulges, HIZ lesions, annular tears or unclear signal
changes. Previous MRI ratings (JSS) were available during the sessions. No more
than 25 sets of images were analysed per day to minimize the fatigue effect.
Definition of early DD in the studies (III, IV)
At both baseline and follow-up, the DD was defined the same way. Early lumbar
DD was defined if there was either a signal intensity change (Grade 2 or 3) or a
change in disc contour (Grade 2 or higher) at one or more lumbar levels. Those
with normal signal intensity (Grade 0 and 1), normal disc contour (Grade 0 or 1),
no AT, normal endplates and no other pathology in MRI were classified as
subjects without DD.
Definition of early DDP (IV)
Progression was defined separately at follow-up (N=166). Progression in DD
status was defined if a subject had either a worsened or new decrease in disc
signal intensity, a new bulge or a herniation, new endplate changes, or a new
Modic change at one or multiple lumbar levels, compared to baseline (Figure 13).
We also rated significant new annular tears (AT) and significant high intensity
zone lesions (HIZ) as progression (DDP). Individuals without these findings or
the same degenerative findings as at baseline were considered non-progressive
(no-DDP). Image sets without unequivocal ratings in lumbar DD or progression
after joint sessions (PK and PE) were re-evaluated and confirmed by JSS in 2011.
4.3 Genetic analyses
Candidate SNPs selection
At the time the studies (II, III, IV) were initiated we listed the known DD
candidate genes and polymorphisms. Most of the previous approaches as well as
the present studies in this thesis focus on the genes that code for known
functioning molecules of the IVD/ECM, including the components of collagen IX
(COL9A2, COL9A3), collagen XI (COL11A2), collagen XXII (COL22A1),
cartilage intermediate layer protein (CILP), skt protein homolog KIAA1217
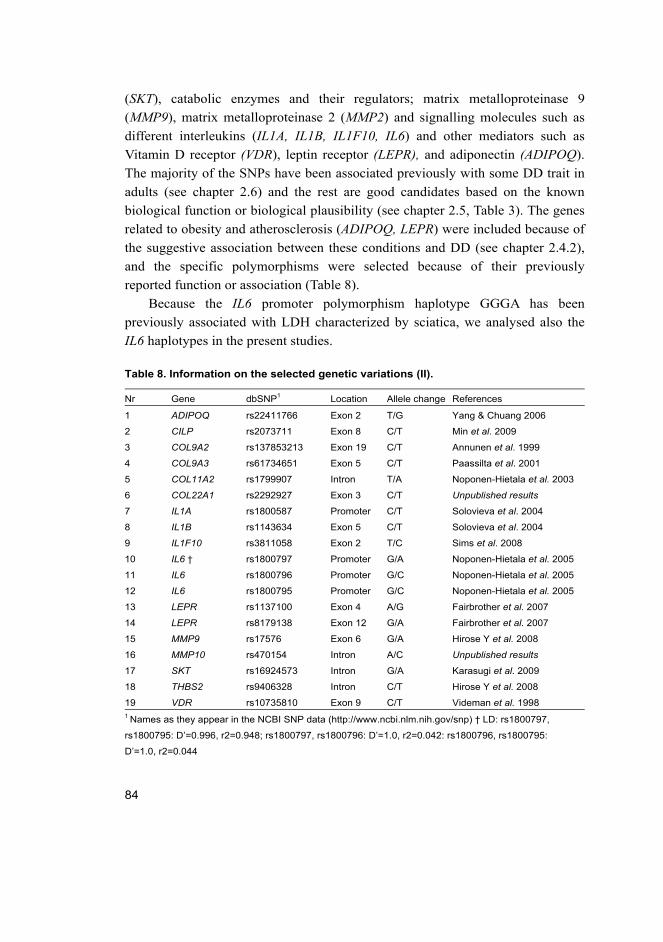
84
(SKT), catabolic enzymes and their regulators; matrix metalloproteinase 9
(MMP9), matrix metalloproteinase 2 (MMP2) and signalling molecules such as
different interleukins (IL1A, IL1B, IL1F10, IL6) and other mediators such as
Vitamin D receptor (VDR), leptin receptor (LEPR), and adiponectin (ADIPOQ).
The majority of the SNPs have been associated previously with some DD trait in
adults (see chapter 2.6) and the rest are good candidates based on the known
biological function or biological plausibility (see chapter 2.5, Table 3). The genes
related to obesity and atherosclerosis (ADIPOQ, LEPR) were included because of
the suggestive association between these conditions and DD (see chapter 2.4.2),
and the specific polymorphisms were selected because of their previously
reported function or association (Table 8).
Because the IL6 promoter polymorphism haplotype GGGA has been
previously associated with LDH characterized by sciatica, we analysed also the
IL6 haplotypes in the present studies.
Table 8. Information on the selected genetic variations (II).
Nr Gene dbSNP1 Location Allele change References
1 ADIPOQ rs22411766 Exon 2 T/G Yang & Chuang 2006
2 CILP rs2073711 Exon 8 C/T Min et al. 2009
3 COL9A2 rs137853213 Exon 19 C/T Annunen et al. 1999
4 COL9A3 rs61734651 Exon 5 C/T Paassilta et al. 2001
5 COL11A2 rs1799907 Intron T/A Noponen-Hietala et al. 2003
6 COL22A1 rs2292927 Exon 3 C/T Unpublished results
7 IL1A rs1800587 Promoter C/T Solovieva et al. 2004
8 IL1B rs1143634 Exon 5 C/T Solovieva et al. 2004
9 IL1F10 rs3811058 Exon 2 T/C Sims et al. 2008
10 IL6 † rs1800797 Promoter G/A Noponen-Hietala et al. 2005
11 IL6 rs1800796 Promoter G/C Noponen-Hietala et al. 2005
12 IL6 rs1800795 Promoter G/C Noponen-Hietala et al. 2005
13 LEPR rs1137100 Exon 4 A/G Fairbrother et al. 2007
14 LEPR rs8179138 Exon 12 G/A Fairbrother et al. 2007
15 MMP9 rs17576 Exon 6 G/A Hirose Y et al. 2008
16 MMP10 rs470154 Intron A/C Unpublished results
17 SKT rs16924573 Intron G/A Karasugi et al. 2009
18 THBS2 rs9406328 Intron C/T Hirose Y et al. 2008
19 VDR rs10735810 Exon 9 C/T Videman et al. 1998 1 Names as they appear in the NCBI SNP data (http://www.ncbi.nlm.nih.gov/snp) † LD: rs1800797,
rs1800795: D’=0.996, r2=0.948; rs1800797, rs1800796: D’=1.0, r2=0.042: rs1800796, rs1800795:
D’=1.0, r2=0.044

85
The SNaPshot (II) assay was designed for SNP genotyping thus excluding other
types of genetic variation. After testing different polymerase chain reaction (PCR)
multiplexes we were able to include nineteen SNPs in the study with Finnish
young adults (II). These SNPs were distributed in 16 different genes (Table 8).
Due to the structure of the DNA sequences at the SNP sites we were not able to
include all possible polymorphisms in the analyses.
Genotyping methods
The DNA was isolated from frozen venous blood samples. After PCR and/or
Multiplex PCR, SNaPshotTM Multiplex Kit (Applied Biosystems), direct
sequencing and different restriction enzyme digestions were used in genotyping
(Table 9). The quality of DNA was not acceptable in all samples and therefore
several genotyping methods were used for some SNPs (III, IV, Table 9). A
negative control in the PCR was used as a standard laboratory procedure.
Concordance between different genotyping methods was verified by re-
sequencing 3–7% of the samples for each SNP (III, IV). In the SNaPshot, the
purified PCR product was processed according to the manufacturer’s protocol
with the exception of the halved amount (2.5 μl) of SNaPshot Ready Reaction
Mix. Analysis was carried out using the ABI3031xl Genetic Analyzer (Applied
Biosystems). In the SNaPshot, the genotyping was validated from a randomly
chosen 10% of samples for each SNP by direct sequencing, using the
abovementioned genetic analyzer with BigDye chemistry from the same
manufacturer. The genotypes were generally scored using the GeneMapper
software (Applied Biosystems). The genotyping was blinded towards MRI status.
Table 9. Genetic variations analysed in the Danish studies (III, IV).
Gene dbSNP1 Risk allele Genotype call rate Detection methods
COL9A3 rs61734651 (C52T) T 1.000 Sequencing, SNaPshot TM
COL11A2 rs1799907 (IVS6-4A>T) T 0.964 BsmFI, Sequencing, SNaPshot TM
IL1A rs1800587 (C-889T) T 0.977 NcoI, Sequencing, SNaPshot TM
IL1B rs1143634 (C3954T) T 0.995 TaqI, SNaPshot TM
IL6 rs1800797 (G-597A) G 0.991 Sequencing
IL6 rs1800796 (G-572C) G 0.991 Sequencing
IL6 rs1800795 (G-174C) G 0.995 Sequencing
IL6 rs13306435 (T15A) A 1.000 Sequencing
IL6 rs2069849 (132 C>T) T 0.995 Sequencing
VDR rs2228570 (T2C) T 0.936 FokI, SNaPshot TM
VDR rs731236 (T352C) C 0.991 TaqI, SNaPshot TM 1SNP numbers as they appear in NCBI database and (prior literature).

86
Statistical analyses in the Finnish young adult study and Danish teenager studies (II, III, IV)
We tested the potential deviation from the Hardy-Weinberg equilibrium (HWE)
using the chi-square test. Prior to polymorphism association, the possible effect of
gender, BMI and height on the DD status was analysed using the SAS program
(V9.1 and V 9.2). Crude and adjusted ORs and their 95% confidence intervals
(CIs) were calculated using logistic regression with SAS. The dependent variable
was DD phenotype and independent variables were genotype, allele or haplotype.
Haplotypes were first estimated by SNPStats (Sole et al. 2006) and then
statistically reconstructed from population genotype data, using the PHASE
program and the Markov-chain method for hapotype assignments (Stephens et al.
2001). When testing for associations between the genotype and polymorphisms,
the SNPStats tests automatically for five different inheritance models:
codominant, dominant, recessive, overdominant and log-additive. One mode of
inheritance is tested for alleles (Sole et al. 2006). In candidate gene association
studies, the questions are more specific than in whole genome association studies.
Therefore, in the setting of existing a priori hypotheses, corrections for multiple
testing were restricted to genes with multiple SNPs. The Bonferroni-Holm
method was used for multiple testing corrections (Holm 1979). Possible gender-
genotype interactions were analysed using SNPStats.
The effect of cluster (school) sampling was tested by the likelihood ratio test
(III). The null hypothesis was that the variance of random effect school is zero
and the test statistic (LRT) is assumed to follow chi-square distribution with one
degree of freedom. The resulting p-value is conservative.
4.4 Systematic review methods (I)
Data Sources and Searches
A systematic search was conducted in MEDLINE, MEDLINE In-Process, ISI
Web Of Science and SCOPUS from 1990 through to August 2011. On-line
association databases, the Genetic Association Database and the Human Genome
Epidemiology Network were consulted after a search for any missing studies. The
SCI-EXPANDED of ISI Web Of Science was searched from 1990 through to
August 2011. Utilizing Boolean operators, different forms (truncation) of the
keywords allele, polymorphism and genotype in either title or topic were

87
combined with the words disc, disk, endplate, lumbar, modic, spondyl(o)arthrosis
similarly in either title or topic, and terms noting macular, retinal and ocular were
used to exclude conditions not related to the intervertebral disc. A search in
SCOPUS was performed in a parallel way using formulations of the words allele,
polymorphism and genotype in either title, abstract or article keyword. This search
was then combined with the words disc, disk, endplate, lumbar, modic and
spondyl(o)arthrosis similarly in either title, abstract or article keyword with AND
Boolean. The NOT operator was used with words macular, retinal, ocular and
optic. The MEDLINE and MEDLINE In-Process were searched using the MeSH
terms intervertebral disk degeneration and intervertebral disk displacement prior
to combining the word endplate and formulations of spondyloarthrosis to the
search in all fields -manner. Different formulations of the words allele,
polymorphism and genotype were then combined with AND Boolean. Finally the
results were limited to humans. The Genetic Association Database
(http://geneticassociationdb.nih.gov/) was searched using the words disc and disk
in all fields-manner. The Human Genome Epidemiology Network (HuGENet™)
was also searched via the HuGE Literature Finder (http://hugenavigator.net/)
using the words disc and disk. Reference tracking of included studies was
performed after retrieving the full text articles. The citations were handled using
the RefWorks–software (ProQuest LLC, both Classic and 2.0 versions were used).
Study Selection
The criteria for considering studies for this review were formalized in an
inclusion criteria form (Appendix 2), which was piloted to minimize human error.
Two investigators (PE and SL) independently examined the titles and abstracts of
the identified studies. If study eligibility was unclear from the abstract, the full
text of the article was retrieved and independently assessed by the assessors
(Figure 12). Any disagreement was resolved by discussion. Eligible studies
included in this review had the following criteria: relevant outcome or disease
(intervertebral disc changes, vertebral endplate changes, spondyl(o)arthrosis),
reliable definition of outcome (MRI), study subjects not less than fifty, human
subjects, and description of specific genetic variant(s). Studies that did not meet
one or more of the eligibility criteria were excluded. The studies were not limited
to any language.

88
Data Extraction
Two investigators (PE and SL) independently extracted the data using a
standardized form (Appendix 2). The form was pilot-tested on three studies to
identify and reduce any potential for misinterpretation (PE, SL and PK). The
following topics were recorded from the included studies: study details and
sponsorship, population structure, phenotypes and details of the MRI, genotyping
details as well as possible biases in selection, performance, detection, attrition and
statistical analyses.
Quality Assessment and Data Synthesis
We developed an instrument for methodological quality assessment (Appendix 2).
The study quality was based on the information reported in the articles and was
simultaneously analyzed with the data extraction phase by two investigators (PE
and SL) independently. However, during the data analysis phase, it was noted
that the formalized summary score did not fully serve all the needs of the current
review (Juni et al. 1999). This discrepancy was resolved through discussion (PE
and SL), after which the synthesized study quality assessment was noted when
estimating protection from bias at the level of evidence analyses. The level of
evidence in each genetic variation was analysed according to the Venice interim
guidelines by The HuGENet Working Group (Ioannidis et al. 2008). These
current guidelines suggest that the level of evidence for genetic association should
be assessed at three main levels: amount of evidence, replication and protection
from bias. The amount of evidence was graded strong (A) in the case of >1000,
moderate (B) in the case of 100–1000 and weak (C) in the case of <100
individuals evaluated in the smallest genetic group of interest. The level of
replication was graded strong (A): extensive replication including at least one
well-conducted meta-analysis with little between-study inconsistency; moderate
(B): well-conducted meta-analysis with some methodological limitations or
moderate between-study inconsistency; and weak (C): no association, no
independent replication, failed replication, scattered studies, flawed meta-analysis
or large inconsistency. Similar tripartite grading was used to analyze the
protection from bias: strong (A): bias, if at all present, could affect the magnitude
but probably not the presence of the association; moderate (B): no obvious bias
that may affect the presence of the association but there is considerable missing
information on the generation of evidence; and weak (C): considerable potential

89
for, or demonstrable, bias that can affect even the presence or absence of the
association (Ioannidis et al. 2008, Munafo 2010). For a positive replication, both
the same phenotype and the same genetic variation were required. The possible
additional biological evidence reported in the studies was also acknowledged
(Ioannidis et al. 2008). Credibility of cumulative epidemiological evidence was
recorded for each variation as described in the guidelines (Appendix 2). The level
of evidence is graded strong when the evidence is strong in all three levels (i.e.
AAA), and weak when any of the three is graded weak (i.e. ACA, BBC, CCC,
etc.). All the other gradings lead to a moderate level of evidence. Two
investigators (PE and SL) confirmed the credibility assessments, which were then
further reviewed by the other investigators. We examined the studies and
extracted data with close scrutiny in order to identify possible multiple
association reports from a single study or clear double publications. Multiple
association reports from a single study or population were included if they
reported on different genetic variations or different disc degeneration phenotypes
than those contained in the first report. Multiple association reports were not
allowed to inflate the level of association evidence.

90

91
5 Results
5.1 Genetic association studies on MRI defined lumbar DD (I)
5.1.1 Characteristics of studies and level of evidence
The systematic search resulted in 1,356 citations (Figure 12). Duplications and
clearly unrelated titles (N=1240) were removed and the full-text articles of the
remaining titles were obtained. Reference tracking, independent evaluation and
reviewer discussions found 52 studies eligible for inclusion (Appendix 1). The list
of studies that were excluded after evaluation (N=72) can be found on Appendix 2.
One double publication was identified (Paz Aparicio et al. 2010, Paz Aparicio et
al. 2011), and the more recent report was excluded.
Fig. 12. Study flow of the systematic review (I) on genetic association studies in DD.

92
Methods and Phenotypes in Studies
The genetic association studies identified in this review were published from the
year 1998 onwards. All 52 studies included in this review used a candidate gene
approach. The number of studied polymorphisms in each study varied between
one and 163 (Bei et al. 2008, Videman et al. 2009). The accuracy of the study
methodology and reporting improved from the early to the more recent studies.
However, many items still related to error and bias were not consistently reported.
Genotyping methodology was generally found to be suitable for each study
performed, although methods to validate genotyping, as well as blinding of
genotyping towards phenotype or vice versa, were rarely reported. In many cases,
the phenotype of disc degeneration varied between the initial and replication
studies (Appendix 1). The phenotype of disc herniation characterized by sciatica
showed the most convincing evidence for association as it was the phenotype in
80% of the studies with moderate evidence of association in the current review
(Table 10). The other phenotypes used in the original studies included decrease in
disc signal intensity or disc height, disc bulges, disc herniations without
specification of symptoms, Modic changes, osteophytes and lumbar spinal
stenosis. Different modifications and combinations of these were also used.
Despite several studies investigating the same genetic variation, meta-analysis
was not feasible due to the clinical and overall heterogeneity of the studies.
Additional biological evidence was reported in six studies (Hirose et al. 2008,
Karasugi et al. 2009, Mio et al. 2007, Seki et al. 2005, Song et al. 2008a, Zhu et
al. 2011). References for equivalent rs-numbers (Sherry et al. 2001) for the
identified polymorphisms are available on-line as supplementary data (Appendix
2).
Level of Association Evidence
None of the genetic variations reached the level of strong evidence for association
in the current review. We found a moderate level of evidence for variations from
studies investigating asporin (ASPN), collagen XI alpha 1 (COL11A1), growth
differentiation factor 5 (GDF5), Sickle tail (SKT); thrombospondin 2 (THBS2)
and matrix metallopeptidase 9 (MMP9) genes (Table 10). These studies had at
least a moderate amount of evidence, replication in an independent sample (same
or independent report), meta-analysis or a combined analysis performed in the
initial study, sufficient protection from bias, and a high statistical significance

93
level in the initial study. Furthermore, additional biological evidence was reported
in the original papers for all these polymorphisms (Table 10). Inadequacy in the
number of subjects, lack of an independent replication report or some
inconsistency in replications, phenotyping problems or missing information in the
report to evaluate protection from bias hindered these associations from reaching
the level of strong association evidence (Appendix 1).
The association study of ASPN consisted of two independent Asian cohorts of
Japanese (N=1353) and Chinese (N=1055) origin as reported by Song et al (Song
et al. 2008a). All Japanese cases had a lumbar disc herniation characterized by
sciatica (LDH) confirmed by MRI, while disc signal decreases were also recorded.
Association between the presence of at least one D14 allele and LDH was found
to be significant, while in the Chinese population the presence of at least one D14
repeat was associated with lumbar disc degeneration. A meta-analysis using the
above-mentioned phenotypes showed that individuals carrying the D14 allele had
increased odds of LDH or disc degeneration 1.7-fold (Song et al. 2008a).
A Japanese study found an association between COL11A1 rs1676486 T-allele
and LDH characterized by sciatica. The original study consisted of three case-
control populations (N=367, N=645, N=710), each independently showing a
significant association. When the populations were combined for meta-analysis
(N=1661), the minor allele T was more prevalent among cases compared to
controls (Appendix 1). Additional biological evidence was also reported (Mio et
al. 2007). A recent multicohort study with Northern European subjects
investigated the rs143383 of the GDF5 gene. Out of the total population
(N=5259), one cohort (N=613) was scanned with MRI, therefore making the
study eligible for our review. In the meta-analysis rs143383, a significant
association was found among women for combined phenotype of disc space
narrowing and osteophytes (Table 10, Appendix 1). When only the MRI cohort
was investigated, the association was not statistically significant, thus generating
some inconsistency in this association (Williams et al. 2011). However, as this
specific disc degeneration phenotype can be obtained reliably on multiple
imaging modalities, such as radiographs, computed tomography or MRI
(Williams et al. 2012), we included the results of the meta-analysis.
Another recent study analyzed the Sickle tail (SKT) gene polymorphisms. Of
the 68 SNPs studied, the rs16924573 was the most strongly associated with LDH
among Japanese subjects (N=1758) and the finding was replicated among Finnish
subjects (N=506) (Table 10, Appendix 1). Allele frequencies were different
between Finnish and Japanese populations, but the meta-analysis of over 2200

94
subjects supported the association (Karasugi et al. 2009). A replication study
between disc signal decrease and SKT rs16924573 has been recently published
(OR 0.27 [95% CI 0.07–0.96], p=0.024) (Kelempisioti et al. 2011). The G-allele
frequency was higher in the case group of both studies, thus indicating an
increased risk. However, the A-allele was rarer in the more recent study and the
association was only seen when the GA-genotype was compared to GG-genotype.
Therefore the OR in the more recent study is protective (Kelempisioti et al. 2011).
Two thrombospondin genes (thrombospondin-1 and THBS2) were examined
in two independent Japanese populations (N=1089 and N=654) as candidate
genes for LDH. Multiple polymorphisms of the THBS2 were associated with
LDH. The polymorphism rs9406328 showed significant association in both
populations independently as well as when populations were combined (Hirose et
al. 2008). The same study also reported a significant association between the
rs17576 of the MMP9 gene and LDH (Hirose et al. 2008).
In this review, the most studied candidate genes for LDD were vitamin-D
receptor (VDR), aggrecan (ACAN), interleukin-1 alpha (IL1A), interleukin-1 beta
(IL1B), collagen IX alpha 2 (COL9A2) and collagen IX alpha 3 (COL9A3)
(Appendix 1). However, a large proportion of the association studies investigating
disc degeneration had some faults, which weakened the evidence for association.
The most common weaknesses were the relatively low number of study subjects
and difficulty in replicating the previous association signal. In general, there was
a lack of replication and where replication did exist, studies were often too
heterogeneous, leading to inconsistencies and differences in the final phenotype.
In some studies, protection from bias seemed to be insufficient. However, due to
failure to report the study details properly, it was often very difficult to adequately
assess studies. In summary, due to general heterogeneity of studies, replications
were inconclusive and meta-analyses were not feasible, thus leading to a weak
level of association evidence in many cases.

95
Table 10. Variations with moderate association credibility1
Gene Variation Amount
of
evidence
Replication
level
Protection
from bias
Additional
biological
evidence2
Ethnicity Phenotype Reference
ASPN Allele D14 B B B yes Japanese
Chinese
sciatica,
signal
Song
et al
2008a
COL11A1 rs1676486 B B B yes Japanese sciatica Mio
et al 2007
GDF5 rs143383 A B A - Northern
European
X-ray3
MRI
Williams
et al 2011
SKT rs16924573 B B A yes Japanese
Finnish
sciatica Karasugi
et al 2009
THBS24 rs9406328 B B B yes Japanese sciatica Hirose
et al 2008
MMP94 rs17576 B B B yes Japanese sciatica Hirose
et al 2008 1Based on Venice interim guidelines (Ioannidis et al. 2008), statistical significance level (p-value) of
original association and replication level including also the absence of inconsistent replications. The
amount of evidence increases when alleles are contrasted 2Reported in the included studies 3Combined
phenotype of disc space narrowing and presence of osteophytes 4One negative replication report in disc
signal phenotype and in different ethnicity.
5.2 Associations between SNPs and lumbar DD on MRI among Finnish young adults (II)
The polymorphisms in SKT, CILP and the IL6 promoter have been associated
before with lumbar disc herniation characterized by sciatica. Therefore, we
analysed these polymorphisms for possible associations with moderate DD in
Finnish young adults (Table 11). We analysed the data also separately by gender.
We hypothesized that as there are differences between genders in back disorders
and many other diseases, there might be differences also in the genetic
associations of DD.
Of all the young adults included in this study (N=538), 45.7% (N=246) did
not have DD, whereas 54.3% (N=292) had DD. Of the 292 individuals with DD,
51.4% (N=150) had moderate DD (Grade 4 DD at one or more lumbar levels, or
Grade 3 DD at three of more lumbar levels). The remaining 142 individuals had
degenerative findings but they were not included in the statistical analysis.

96
The mean genotype call rate was 0.98 in the total population (N=538). Two
markers (COL11A1; rs1676486 and VDR; rs731236) did not produce technically
high quality genotyping results (data not shown), thus they were excluded
resulting in a SNP call rate of 0.90. No deviations from the Hardy-Weinberg
Equilibrium (HWE) were observed. Significant associations with DD were
observed in three (IL6, SKT, CILP) of the 16 genes tested. Differences between
genders in the genetic associations with DD were not observed in IL6 and SKT,
but the association in CILP was seen only among women.
5.2.1 IL6, SKT, CILP and DD
Of all the 19 SNPs analysed in Finnish young adults, significant associations with
DD were observed for polymorphisms in IL6, SKT and CILP (Table 11). The IL6
rs1800795 G-allele was associated with the degenerative phenotype in an additive
manner; GC-genotype, OR 1.51 [CI 0.92–2.47] and GG-genotype, OR 2.09 [CI
1.14–3.85]. The G-allele frequencies were 0.490 and 0.398 in subjects with and
without DD, respectively (OR 1.45 [CI 1.07–1.96]).
The rs1800797 was similarly associated with the DD phenotype, suggesting
an additive model; GA: OR 1.55 [CI 0.95–2.54] and GG: OR 1.85 [CI 1.01–3.39],
compared to the AA-genotype. G-allele frequencies were 0.490 and 0.409 among
subjects with and without DD, respectively (OR 1.37 [CI 1.02–1.85]). The
rs1800795 and rs1800797 are in almost complete LD (Table 8), therefore they are
redundant.
The GA genotype of the SKT (KIAA1217) polymorphism rs16924573 was
associated with a reduced risk of DD (OR 0.27 [CI 0.07–0.96], p=0.024). The
frequency of the GA genotype was lower among subjects with DD (2.0%),
compared to subjects without DD (6.5%). The CILP SNP rs2073711 was
associated with DD, but only among women. The CT/TT genotypes, compared to
the CC genotype, resulted in an OR of 2.04 [CI 1.07–3.89], and p=0.025
(corrected p=0.05, correction made for two tests). We did not observe statistically
significant associations between the other SNPs and DD. Table 11 presents the
genotype frequencies and respective ORs for an individual SNP.
Prior studies indicate an association between a particular IL6 haplotype and
LDH. Therefore, we subsequently tested if IL6 haplotypes similarly associate
with moderate DD. The haplotype analyses resulted in an association between the
GGG haplotype (from rs1800797, rs1800796 and rs1800795, respectively) and
DD (OR 1.48 [CI 1.09–2.01]) (Table 12). From this haplotype, only one SNP
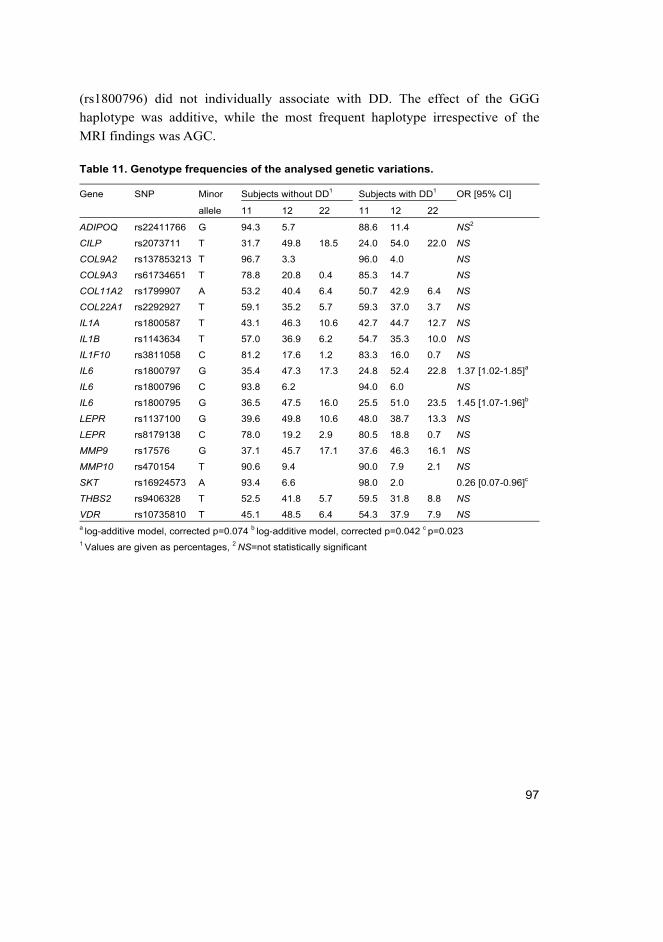
97
(rs1800796) did not individually associate with DD. The effect of the GGG
haplotype was additive, while the most frequent haplotype irrespective of the
MRI findings was AGC.
Table 11. Genotype frequencies of the analysed genetic variations.
Gene SNP Minor Subjects without DD1 Subjects with DD1 OR [95% CI]
allele 11 12 22 11 12 22
ADIPOQ rs22411766 G 94.3 5.7 88.6 11.4 NS2
CILP rs2073711 T 31.7 49.8 18.5 24.0 54.0 22.0 NS
COL9A2 rs137853213 T 96.7 3.3 96.0 4.0 NS
COL9A3 rs61734651 T 78.8 20.8 0.4 85.3 14.7 NS
COL11A2 rs1799907 A 53.2 40.4 6.4 50.7 42.9 6.4 NS
COL22A1 rs2292927 T 59.1 35.2 5.7 59.3 37.0 3.7 NS
IL1A rs1800587 T 43.1 46.3 10.6 42.7 44.7 12.7 NS
IL1B rs1143634 T 57.0 36.9 6.2 54.7 35.3 10.0 NS
IL1F10 rs3811058 C 81.2 17.6 1.2 83.3 16.0 0.7 NS
IL6 rs1800797 G 35.4 47.3 17.3 24.8 52.4 22.8 1.37 [1.02-1.85]a
IL6 rs1800796 C 93.8 6.2 94.0 6.0 NS
IL6 rs1800795 G 36.5 47.5 16.0 25.5 51.0 23.5 1.45 [1.07-1.96]b
LEPR rs1137100 G 39.6 49.8 10.6 48.0 38.7 13.3 NS
LEPR rs8179138 C 78.0 19.2 2.9 80.5 18.8 0.7 NS
MMP9 rs17576 G 37.1 45.7 17.1 37.6 46.3 16.1 NS
MMP10 rs470154 T 90.6 9.4 90.0 7.9 2.1 NS
SKT rs16924573 A 93.4 6.6 98.0 2.0 0.26 [0.07-0.96]c
THBS2 rs9406328 T 52.5 41.8 5.7 59.5 31.8 8.8 NS
VDR rs10735810 T 45.1 48.5 6.4 54.3 37.9 7.9 NS a log-additive model, corrected p=0.074 b log-additive model, corrected p=0.042 c p=0.023 1 Values are given as percentages, 2 NS=not statistically significant

98
Table 12. Frequencies of IL6 haplotypes and diplotypes.
Haplotype/Diplotype Subjects without DD Subjects with DD
n % n % OR [95% CI] p-value
AGC1 287 59.1 151 50.7 1.00
GGG 177 36.4 136 45.6 1.48 [1.09-2.01] 0.0122
GCG 15 3.1 9 3.0 0.88 [0.37-2.12]
Other 7 1.4 2 0.6 -
AGC/AGC 86 35.0 37 24.7 1.00
GGG/AGC 102 41.5 71 47.3 1.61 [0.97-2.67] 0.0650
GGG/GGG 33 13.4 30 20.0 2.22 [1.17-4.22] 0.0152
GCG/AGC 10 4.1 5 3.3 1.01 [0.31-3.23]
GGG/GCG 5 2.0 4 2.7 1.17 [0.28-4.80]
GGC/AGC 3 1.2 1 0.7 0.70 [0.07-7.28]
other 7 2.9 2 1.3 - 1 The predisposition (for DD) of haplotypes GGG and GCG is tested vs. the most common haplotype
AGC that is assumed to have OR 1.00
In summary, five variations of the 19 selected candidate SNP s in 16 genes were
associated with susceptibility to moderate DD in our study. The present results
support the role of IL6, SKT and CILP in DD. We oberved significant differences
between genders only in CILP in the Finnish study (II).
5.2.2 Height, BMI and DD
Obesity and height have been associated before with DD and LBP. Therefore, we
analysed also height and BMI for possible association with DD in our study
population of Finnish young adults.
Height and BMI were found to be associated with DD (Table 13), but after
adjustment for height, BMI was no longer associated with DD. DD was more
frequent among men (32.1%) than women (24.9%), but the difference was not
statistically significant when adjusted for height.
Table 13. Anthropometric data of the study population at the age of 19 years
Anthropometric Subjects
without DD
Subjects
with moderate DD
N 246 150
Gender (females, %) 66.3 52.0
Height (mean; cm)1 167.8 172.3
BMI (mean; kg/m2)2 22.6 23.4 1 p<0.0001, 2 p=0.0384

99
5.3 Associations between SNPs and early lumbar DD and DDP among Danish teenagers (III, IV)
The polymorphisms in IL1A and the IL6 promoter have been associated before
with disc degeneration (IL1A) and lumbar disc herniation characterized by
sciatica (IL6). Therefore, we analysed these polymorphisms for possible
associations with moderate DD in Danish teenagers. We hypothesized that as
there are differences between genders in back disorders and many other diseases,
there might be differences also in the genetic associations with early DD. After
observing differences between genders in DD, we investigated also whether there
are gender differences in the genetic association with DDP.
5.3.1 IL1A, IL6 and early DD in girls
Baseline study (III)
At baseline, we investigated the possible associations between the selected 11
SNPs (Table 15) with DD at among the Danish teenagers at 13 years of age.
Of the 352 children studied, 73 boys and 81 girls had no MRI changes, while
30 boys and 36 girls were classified as having lumbar DD (Table 14). Subjects
with other specific findings (N=132, 37.5%) were excluded from the genetic
analyses thus leaving 220 individuals to be included at baseline.
We were able to successfully genotype all the SNPs (i.e. SNP call rate is 1.00)
using different genotyping methods. The quality of DNA was low in some of the
samples requiring the use of multiple methods to obtain reliable genotyping
results (Table 9). Genotype frequencies for all variations were in HWE. We also
achieved a high sample call rate in the Danish study (Table 15). Among girls, a
positive association between the degenerative phenotype and IL1A rs1800587 was
observed in a dominant genotype model, where CT/TT genotypes were compared
to the CC genotype, OR 2.85 [CI 1.19–6.83], Pcorr=0.028, correction made for
two tests. In the IL6 promoter polymorphism rs1800796, the C-allele was more
frequent among the subjects with DD than the subjects without DD. The OR from
the log-additive model for the C allele was 6.71 [CI 1.71–26.3], Pcorr=0.0105,
correction made for five tests. No association between the studied polymorphisms
and DD was observed among boys.
Subsequent haplotype analysis of IL6 resulted in a haplotype GCG (of
promoter polymorphisms rs1800797, rs1800796 and rs1800795, respectively),
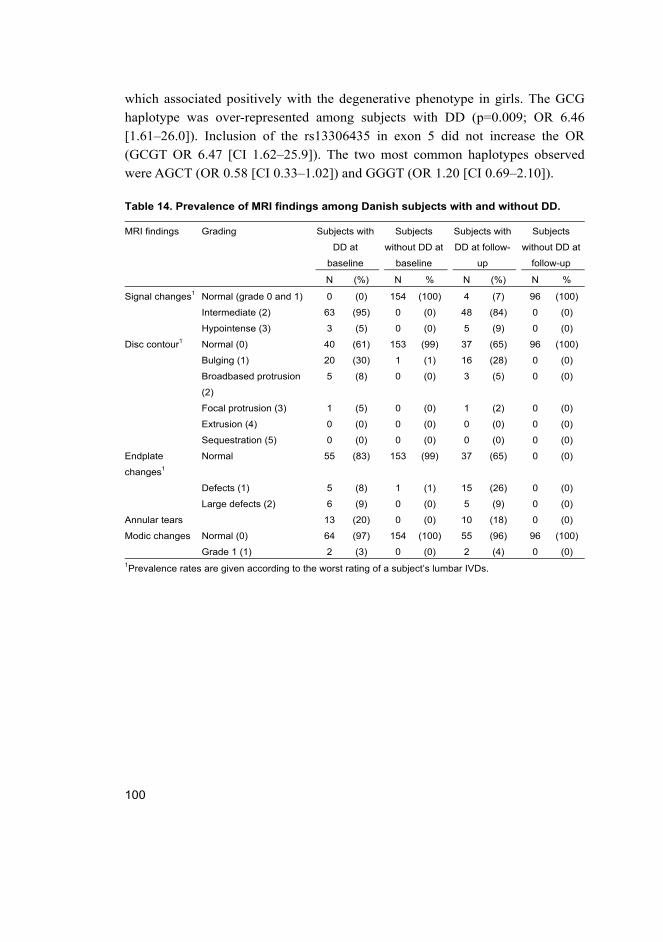
100
which associated positively with the degenerative phenotype in girls. The GCG
haplotype was over-represented among subjects with DD (p=0.009; OR 6.46
[1.61–26.0]). Inclusion of the rs13306435 in exon 5 did not increase the OR
(GCGT OR 6.47 [CI 1.62–25.9]). The two most common haplotypes observed
were AGCT (OR 0.58 [CI 0.33–1.02]) and GGGT (OR 1.20 [CI 0.69–2.10]).
Table 14. Prevalence of MRI findings among Danish subjects with and without DD.
MRI findings Grading Subjects with
DD at
baseline
Subjects
without DD at
baseline
Subjects with
DD at follow-
up
Subjects
without DD at
follow-up
N (%) N % N (%) N %
Signal changes1 Normal (grade 0 and 1) 0 (0) 154 (100) 4 (7) 96 (100)
Intermediate (2) 63 (95) 0 (0) 48 (84) 0 (0)
Hypointense (3) 3 (5) 0 (0) 5 (9) 0 (0)
Disc contour1 Normal (0) 40 (61) 153 (99) 37 (65) 96 (100)
Bulging (1) 20 (30) 1 (1) 16 (28) 0 (0)
Broadbased protrusion
(2)
5 (8) 0 (0) 3 (5) 0 (0)
Focal protrusion (3) 1 (5) 0 (0) 1 (2) 0 (0)
Extrusion (4) 0 (0) 0 (0) 0 (0) 0 (0)
Sequestration (5) 0 (0) 0 (0) 0 (0) 0 (0)
Endplate
changes1
Normal 55 (83) 153 (99) 37 (65) 0 (0)
Defects (1) 5 (8) 1 (1) 15 (26) 0 (0)
Large defects (2) 6 (9) 0 (0) 5 (9) 0 (0)
Annular tears 13 (20) 0 (0) 10 (18) 0 (0)
Modic changes Normal (0) 64 (97) 154 (100) 55 (96) 96 (100)
Grade 1 (1) 2 (3) 0 (0) 2 (4) 0 (0) 1Prevalence rates are given according to the worst rating of a subject’s lumbar IVDs.

101
Table 15. Genetic variations analysed.
Gene dbSNP1 Risk
allele3
Sample
call rate
Detection methods References
COL9A3 rs61734651
(C52T)
T 1.000 Sequencing,
SNaPshotTM
Paassilta et al. 2001
COL11A2 rs1799907
(IVS6-4A>T)
T 0.964 BsmFI, Sequencing,
SNaPshotTM
Noponen-Hietala et al. 2003
IL1A rs1800587
(C-889T)2
T 0.977 NcoI, Sequencing,
SNaPshotTM
Solovieva et al. 2004
IL1B rs1143634
(C3954T)
T 0.995 TaqI, SNaPshotTM Solovieva et al. 2004
IL6 rs1800797
(G-597A)
G 0.991 Sequencing Noponen-Hietala et al. 2005
IL6 rs1800796
(G-572C)
G 0.991 Sequencing Noponen-Hietala et al. 2005
IL6 rs1800795
(G-174C)
G 0.995 Sequencing Noponen-Hietala et al. 2005
IL6 rs13306435
(T15A)
A 1.000 Sequencing Noponen-Hietala et al. 2005
IL6 rs2069849
(132 C>T)
T 0.995 Sequencing Noponen-Hietala et al. 2005
VDR rs2228570
(T2C)
T 0.936 FokI, SNaPshotTM Videman et al. 1998
VDR rs731236
(T352C)
C 0.991 TaqI,SNaPshotTM Videman et al. 1998
1 Names as they appear in the NCBI SNP database; http://www.ncbi.nlm.nih.gov/snp and (prior literature). 2 A priori genetic model was dominant. 3 Putative risk allele according to previous studies.
In summary, the baseline Danish study (III) was the first to examine the genetic
determinants of DD in childhood. Our results suggest possible roles for IL1A and
IL6 in early DD among girls and support in part the earlier findings in adult
populations.
Follow-up study (IV)
We investigated the possible associations between the selected 11 SNPs (Table 15)
with DD in the Danish teenagers also at follow-up, at the age of 16 years. Out of
220 individuals included at baseline, a total of 166 (75%) were available for the
analyses at the follow-up. Of these 166 subjects, 57.8% (N=96) were defined as

102
subjects without DD while 34.3% (N=57) had DD (Table 18). Subjects with
intermediate findings at follow-up (N=13) were not eligible for either group.
Among girls, the T-allele of IL1A rs1800587 was associated in an additive manner
with DD. The resultant OR was 2.82 [CI 1.29–6.16] in the additive genotype
model (p=0.0064). Gender-genotype interaction for DD was significant; the p-
value of the test for interaction in the trend was 0.024.
5.3.2 IL1A and early DDP in girls
We investigated the possible associations between the selected 11 SNPs (Table 15)
and disc degeneration progression (DDP) in the Danish teenagers at 16 years of
age. DDP was defined based on morphological IVD changes in MRIs taken at 13
and 16 years of age.
Unexpectedly, follow-up improvement in MRI findings was seen in 12
individuals classified as subjects with DD at baseline. After a consensus session
and re-evaluation by the radiologist (JSS), eight individuals were rated as subjects
without DD, and three subjects with intermediate findings were not eligible for
either group (Figure 13).
DDP was seen in 33.8% of the boys (N=25/74) and in 17.4% (N=16/92) of
the girls. Out of the subjects with DDP (N=41), it was defined based on a
decrease in disc signal intensity in 15.7% (N=26), a new or clearly worsened
endplate change in 15.1% (N=25), a new disc herniation in 7.2% (N=12), a new
AT in 2.4% (N=4), a new bulge in 1.8% (N=3), and a new HIZ in 1.2% (N=2).
DDP definition was based on one change in 36.6% (N=15), two changes in 51.2%
(N=21) and three changes in 12.2% (N=5) of the DDP subjects. Multiple level
DDP was present in 56.1% (N=23) of the subjects while the two lowest levels
(L4/L5 and L5/S1) were the most often affected (68.3%, N=28).
Among girls, the T-allele of IL1A rs1800587 was associated in an additive
manner with DDP (OR 2.45 [95% CI 1.03-5.82] in the additive genotype model,
p=0.037. However, the gender-genotype interaction for IL1A was not significant
in DDP.
5.3.3 Height, weight and early DD (III, IV)
Obesity and height have been associated before with DD and LBP. Therefore, we
analysed height, weight and BMI for possible association with DD in our studies
including the population of Danish teenagers at both baseline and follow-up.

103
At baseline, the subjects with DD were on average 3 cm taller that those
without DD (Table 16). The effect size of height was the same, but not significant,
when analysed among boys and girls separately. There was no association
between BMI and DD.
At follow-up, the subjects with DD were on average 5 cm taller than subjects
without DD. They also tended to weigh more than subjects without DD (Table 16).
Table 16. a) Gender distribution among study subjects with and without DD.
Gender Subjects
with DD
at baseline
Subjects
without DD
at baseline
Subjects
with DD
at follow-up
Subjects
without DD
at follow-up
N (%) N (%) N (%) N (%)
Boys 30 (45) 73 (47) 30 (53) 36 (38)
Girls 36 (55) 81 (53) 27 (47) 60 (62)
Table 16. b) Anthropometric data of study subjects with and without DD.
Anthropometric Subjects
with DD
at baseline
Subjects
without DD
at baseline
Subjects
with DD
at follow-up
Subjects
without DD
at follow-up
Mean SD Mean SD Mean SD Mean SD
Age (years) 13.1 0.4 13.1 0.4 15.7 0.3 15.7 0.3
Height (cm) 162 7.7 159a 7.3 173 9.3 168b 7.9
Weight (kg) 51 8.5 49c 8.7 63 10.4 59 9.1
BMI (kg/m2) 19.5 2.3 19.2 2.8 20.9 2.7 20.8 2.6 ap=0.029, bp=0.008, cp=0.042 (Mann-Whitney U-test)

104
Fig. 13. Transitions of study subjects between groups. DD=subjects with DD
nDD=subjects without DD, nElig=subjects with other specific MRI findings,
DDP=progression of DD, nDDP=no progression of DD. *Additional definition of
phenotype (DDP+nDPP=DD+nDD+nElig at follow up). Dashed line indicates illogical
pathways, in which subjects were re-evaluated three times.

105
6 Discussion
6.1 The amount of cumulative evidence is currently modest (I)
In the systematic review 52 candidate gene association studies were identified. All
these studies used MRI for DD definition and defined a specific genetic
polymorphism. The phenotype with the most convincing evidence was LDH
characterized by sciatica.
Based on the present systematic review (I), the quality of the evaluated
studies varied considerably and some recurrent weaknesses were identified.
Definitions of MRI phenotypes were not clearly reported and there was variability
in the selection of subjects; some studies used clinical patients while other studies
were population-based. There is an important difference between subjects
belonging to a population-based cohort and cases recruited from a clinic
specialized in treatment of LBP patients. Results from studies with this
discrepancy alone are not comparable. Further, there were a few studies, where
the control group was not evaluated using MRI or subjects with LBP and/or
sciatica were included as controls. As such, differences in subject selection and
phenotype definition hinder efforts to produce reasonable meta-analyses.
Population based studies with large study samples with adequate statistical power
were rare; in fact, only six studies possessed a sample size greater than 1000
individuals (Appendix 1). Quality control steps, such as control of population
stratification, Hardy-Weinberg equilibrium testing, and statistical power
calculations, were often not consistently reported, even though reporting
improved in more recent publications. An improvement in the quality of reporting
of genetic studies has been observed in the most recent publications (Yesupriya et
al. 2008). Moreover, the publication bias (i.e. negative results are not published)
was clearly visible in the published data under review, whereby nearly 90% of the
included studies reported positive associations between lumbar disc degeneration
and specific polymorphisms.
The earlier non-systematic reviews have suggested that various genes (ACAN,
VDR, COL9A2, COL9A3, COL11A2, IL1, IL6, MMP-3, MMP-2, TIMP-1,
COL1A1, ESR1, MATN3, CILP, COX2, THSD2, NCOR2, CD36, TNA and IGF-1R)
may be related to the development of lumbar disc degeneration (Ala-Kokko 2002,
Battie et al. 2009, Battie & Videman 2006, Chan et al. 2006, Kalb et al. 2011,
Kalichman & Hunter 2008b, Song et al. 2008, Zhang et al. 2008). However,

106
based on the present systematic review, these genes have a weak level of
cumulative association evidence. This is probably due to more strict inclusion
criteria applied in the present review; the earlier reviews included, for instance,
studies without MRI evaluation as well as studies with success of surgical
treatments as the phenotype. Moreover, studies with less than 100 subjects, no
replication/meta-analysis or with some bias were considered as having weak
evidence. Furthermore, this first extensive systematic review is the only one that
has used the guidelines by the HuGE Working Group (Ioannidis et al. 2008). The
main difference between the present review compared to the earlier ones is that in
the present review the heterogeneous replication reports between different DD
phenotypes were not considered to be replications at all. This is justified, because
there is no data at the moment on how different DD traits could be combined for
statistical analyses. No adjustment for this heterogeneity can be made because the
interrelations between these traits are not yet elucidated.
There have been reports of clearly different risk allele frequencies between
different ethnic populations (e.g. Caucasians, Asians) in the genetic association
studies on DD (Appendix 1). This suggests that different risk alleles may be
involved in lumbar DD in different ethnic groups. Thus, replication studies are
needed in study populations of different ethnic/geographic origin to provide a
more comprehensive understanding. In the present systematic review, the most
consistent evidence (i.e. moderate evidence) was based on studies in Asian
populations. However, in one study, the association with moderate evidence (SKT
gene and sciatica) was originally reported in Japanese and replicated in Finnish
populations (Karasugi et al. 2009, Kelempisioti et al. 2011). Alternatively, in
many cases, replications were inconsistent, either due to different phenotypes or
different genetic variations examined in the replication study.
The present analysis of the previously published associations indicate that the
level of association evidence remains weak for multiple genetic polymorphisms.
Current results stress the limitations of the present status of genetic association
studies in relation to DD. The main problem in these studies is the lack of a gold
standard in the coding of MRI images for degenerative changes. Collaborative
studies with large population-based cohorts and well-defined phenotypes and
genotypes are necessary for major advances in understanding the genetic
component of DD.

107
6.2 IL6 SNPs and DD among young individuals (II, III, IV)
The role of IL6 and in particular the effect of three common promoter
polymorphisms (rs1800797, rs1800796, rs1800795) have been studied previously
for the presence of sciatica (Noponen-Hietala et al. 2005); hence we wanted to
investigate the IL6 haplotypes for possible associations with DD in young
individuals. Furthermore, the haplotype analysis is a way to simultaneously study
the effects of multiple polymorphisms on a dependent variable. In the study with
Finnish young adults imaged with MRI at the age of 21 (II), polymorphisms in
IL6 (rs1800797 and rs1800795) were associated with moderate DD and the G
allele in both of these was the risk allele.
The IL6 haplotype GGG (derived from (rs1800797, rs1800796 and
rs1800795, respectively) was further associated with moderate DD. This finding
is supported by an earlier observation that, among sciatica patients, GGGA was
the risk haplotype (Noponen-Hietala et al. 2005). The two most common
haplotypes (AGC and GGG) observed in the Finnish study (II) were in
accordance with the ones we observed in the study with Danish children (III),
indicating that there are no major differences in allele frequencies of these
polymorphisms between the two study populations.
The baseline study among Danish children (III) found an association between
rs1800796 and early DD in girls. Furthermore, a three SNP GCG haplotype
(derived from (rs1800797, rs1800796 and rs1800795, respectively) was
associated with early DD. The GGGA haplotype that was earlier found to
associate with LDD characterized by sciatica had a frequency less than 0.02 and
was not associated with early DD (III). The lack of association may be related to
sample origin, sample size or different phenotypes; early DD vs. sciatica. The
most significant weakness of the studies (II, III, IV) is the small sample size. With
a haplotype frequency of 0.02 significantly more subjects would be required. To
reach 80% power and 5% significance with the haplotype frequencies observed in
this study (III), more than 30,000 subjects would be needed assuming a 1:1 case-
control ratio (OSSE, accessed October 2012: http://osse.bii.a-star.edu.sg/). Still,
these retrospective power calculations are a little controversial (Thomas 1997),
but they can give a good estimate of whether lack of association might be due to
lack of power. However, the two most common haplotypes in the present study
were again in accordance with earlier findings (Noponen-Hietala et al. 2005). In
the follow-up study (IV) with the Danish children the association with IL6 among

108
boys was refuted after correction for multiple testing and while no associations
were observed among girls.
Biologically, IL6 is a good candidate gene for DD. The pleiotropic cytokine
IL6 is one of the most important mediators of inflammatory reactions (Smith &
Humphries 2009) and it has also been reported that IL-6 is produced at the site of
lumbar disc herniation (Burke et al. 2002). Previous reports of protective effects
of heterozygotes of either IL6 rs1800797 or 1800795 in DD or DDP are not
available. Functional investigations have shown that the production of IL6 is
higher in individuals carrying the same alleles (in the haplotypes, however) that
were here associated with an increased risk for DD (Rivera-Chavez et al. 2003,
Terry et al. 2000). However, based on recent research, the effects of IL6 promoter
polymorphisms on gene expression are likely to be more complex than what has
been initially expected (Smith & Humphries 2009). A recent in vitro study using
blood cells from prepubescent children reported gender differences in IL6
production (Casimir et al. 2010). Furthermore, increased production of IL6 has
been previously associated with a wide range of conditions related to aging
(Omoigui 2007). It is also interesting to note that the same IL6 promoter variant
associated with hand OA (Kämäräinen et al. 2008). DD and OA have a few
similar features and may share some common etiological factors, but this issue is
not well understood (Ryder et al. 2008).
In conclusion, the present findings are parallel to earlier findings among adult
sciatica patients and bring up the importance of IL6 promoter region in DD
among young individuals. However, in the analyses with IL6, as well as with all
the other included genes (II, III, IV), we corrected the p-values only when the
analysis included multiple SNPs in the same gene, and in the corrections we noted
only the markers within each gene for each correction. We did not take into
account all the SNPs that are independent from each other, and for this reason the
statistical significance of the results in IL6 as well as in all the other genetic
associations in young individuals reported in this thesis (II, III, IV) are to be
interpreted with caution.
6.3 IL1A SNPs and DD among young individuals (II, III, IV)
The IL1A rs1800587 T-allele was associated with early DD in both baseline and
follow-up time-points among Danish girls. Such associations were not observed
in boys. Further, there were no associations among the Finnish young adults (II),

109
but the TT genotype was bit more common in subjects with DD (non significant
difference).
Additionally, the same allele was associated with progression of DD. Similar
to DD, the association between DDP and IL1A rs1800587 T-allele was significant
only among girls (IV). There is some overlap between DD in baseline, DD in
follow-up and DDP (Figure 13), but separate positive associations in all these
groups increase the value of these findings.
The IL1A rs1800587 T-allele has been associated previously with IVD bulges and
Modic changes in an occupational cohort of Finnish men (Karppinen et al. 2009,
Solovieva et al. 2004). Based on animal studies it is also known that IL-1 is the
dominant cartilage-destructive cytokine (van & Bresnihan 1999). Furthermore,
additional biological evidence is available that makes IL1A also a good candidate for
DD (see III and IV for more details). The present results are in line with the earlier
findings with the exception of gender difference, which is a novel finding.
The sample size was limited at both baseline (III) and follow-up (IV).
However, it is to be noted that the sample size was larger than in the majority of
the previous studies focusing on DDP (Borenstein et al. 2001, Elfering et al. 2002,
Jarvik et al. 2005, Videman et al. 2006). It is possible that the present findings
indicate true sex differences in genetic effects on DD, but these results are to be
interpreted with caution (Patsopoulos et al. 2007). However, in osteoporosis and
osteoarthritis more evidence is available on the gender-specificity of the effects of
genetic variants (Karasik & Ferrari 2008, Valdes et al. 2007). Furthermore, the
gender specific effects in complex disorders are also recognized in the expanding
field of epigenetics (Kaminsky et al. 2006, Ordovas 2007). It is possible that
epigenetic modifications may in part explain gender effects of DNA variations
identified in gender-stratified genetic association studies (Kaminsky et al. 2006).
In sciatica, the prevalence in men has long been considered to be 1.5–3 times
higher compared to women (Frymoyer 1992). This has been conventionally
explained by the fact that men tend to work in more physically demanding
occupations. However, in childhood and adolescence the situation might be even
vice versa, at least when it comes to LBP, as girls have been reported to have a
higher prevalence of pain (Diepenmaat et al. 2006). Similarly, gender differences
in LBP reporting were seen in a study with partly the same population as in the
present study (Kjaer et al. 2011). These dissimilarities on the other hand have
been suggested to be due to hormonal differences and the onset of menstruation-
related pains in girls (Auvinen 2010, Wedderkopp et al. 2005). Interestingly, a
recent very large-scale study among individuals serving in the United States

110
Military during the years 1999–2008 (N=14,071,570) found that the incidence of
lumbar degenerative disc disease was higher among women compared to men
(adjusted incidence rate ratio 1.21 [CI 1.18–1.23]) (Schoenfeld et al. 2011).
However, the disease status was based on a previously assigned disease
classification code, a diagnosis, which may have some variability thus presenting
a clear limitation of the study. Based on the current results, it seems that gender
specific effects of IL1A polymorphisms in DD and DDP are possible. However,
further evaluations in larger populations are needed.
6.4 SKT and CILP SNPs and DD among Finnish young adults (II)
The association of the SKT rs16924573 variant with DD (defined using disc
signal changes) has not been previously observed neither has a negative
association been published. It is suggested that Skt acts at the later stage of NP
growth and hypertrophy in mice but the function of its human homolog
(KIAA1217 or SKT) is not well known. However, the SKT is expressed in the
human IVD (Karasugi et al. 2009). In the present study, carriers of the minor
allele A were at a reduced risk of DD (OR 0.26 [CI 0.07–0.96]). The importance
of Skt in the development of caudal intervertebral discs (IVD) in mice has been
established previously (Semba et al. 2006). The mice with homozygously trapped
SKT gene produce a deformed “kinky tail” phenotype – seemingly due to NP
changes – during adulthood (Semba et al. 2006). These observations led to the
first study with human LDH patients characterized by sciatica (Karasugi et al.
2009). In the study an association of SKT rs16924573 G allele with LDHs was
found (OR 1.34 [CI 1.14–1.58]) (Karasugi et al. 2009). DD, the phenotype of the
present study, is believed to be an important intermediate stage in symptomatic
disc disease (Videman et al. 2006), which is often characterized by LDH either
with or without sciatic pain (Frymoyer 1988). The current finding is in line with
the earlier findings, but the differences between the phenotypes in these studies
need to be noted. The earlier study has investigated a clinical phenotype, LDH
with sciatica while our study focuses on moderate DD. The interrelation between
these two phenotypes is far from elucidated. These findings provide some support
to the hypothesis that SKT could have long-term importance in the onset of
symptomatic disc disease by making the discs more vulnerable for DD.
In addition, an association between CILP rs2073711 and DD was observed.
In the present study, the T allele was associated with an increased risk of DD. The
TT/CT genotype compared to the CC genotype showed an OR of 2.04 [1.07–

111
3.89]. This association was, however, only found in females. This finding is in
contrast with previously reported associations among Japanese subjects, which
indicated the C allele as the risk allele for symptomatic lumbar disc disease (Seki
et al. 2005) as well as for lumbar DD (Min et al. 2009, Min et al. 2010). However,
a replication study in Finnish and Chinese populations did not find an association
between CILP and symptomatic disc disease or MRI-defined DD (Virtanen et al.
2007a). Interestingly, the C allele was associated with reduced risk of knee OA
(Valdes et al. 2004), which is in accordance with our findings that the T allele,
and not the C allele, is the risk allele. Although functional studies in Japanese
white rabbits (sex not reported) suggest that the C allele is responsible for
increased binding and inhibition of TGF-β1 (Seki et al. 2005), the discrepancies
in the association studies could simply be due to the differences between
European and Japanese populations. It may be possible, that all the Caucasian risk
polymorphisms are not significant in Asians and vice versa. Among Finns, the
minor allele is different and minor allele frequency is over 0.40. Furthermore, it is
to be noted that while in the Japanese collegiate athletes (N=601, aged ca. 20
years) the C was more common among males with DD, the T allele was more
common in females with DD (non-significant difference) (Min et al. 2010).
Therefore, it can be suggested that a true gender difference may exist also in the
association between CILP rs2073711 and DD. These findings highlight the
importance of well-defined phenotypes and support the concept that early DD is a
particularly important phenotype to study.
6.5 Height, weight and DD among young individuals (II, III, IV)
In the present studies, subjects with DD were significantly taller than those
without DD (II, III, IV). Furthermore, the Danish teenagers with DD tended to
weigh more than those without DD (IV). The association between IVD signal
changes and height has not been observed before among young individuals.
However, it is to be noted that there are very few MRI studies on young
individuals in relation to DD in total. Regarding other DD phenotypes, the
association between body height and LDH has previously been contradictory
(Heliövaara 1989, Kelsey & Hardy 1975). In another study among middle-aged
women living in the UK (N=796), individuals with radiographically defined DD
progression over a 9-year period were taller than individuals without DD
progression, but the difference (0.7 cm) was not statistically significant (Hassett
et al. 2003). However, in a recent large cohort study (N=13,282) body height

112
recorded at baseline was associated with higher likelihood of back surgery and
low back pain during the 12-year follow-up. Although the associations reached
statistical significance only among males, a similar trend was observed among
females as well (Coeuret-Pellicer et al. 2010).
One possible mechanism for the role of body height as a risk factor is lumbar
disc size. Larger discs might be predisposed to DD due to greater diffusion
distances of nutrients into the centre of the nucleus pulposus (Urban et al. 2004).
Furthermore, higher discs might be at an increased risk of mechanical failure
under external loading (Natarajan & Andersson 1999). Finally, facet joint
alterations have been found to be more frequent in taller individuals with LDH
(Karacan et al. 2004). The effects of body height and weight on LBP have also
been reviewed and discussed previously (Coeuret-Pellicer et al. 2010, Shiri et al.
2010a). The associations between height/weight and DD among young
individuals observed in the present studies are other interesting findings and
support the earlier results that these factors are associated with LBP.
6.6 Future possibilities
Rapid advancements in genetics and bioinformatics have led to the situation
where the amount of data under analysis has increased substantially providing
new opportunities to reveal the genetic background of complex traits (Kere 2010,
Mechanic et al. 2011, Montgomery & Dermitzakis 2011, Niedringhaus et al. 2011,
Pattin & Moore 2009, Sethumadhavan et al. 2011). Over the last five years
genome-wide association studies (GWAS) have become a powerful tool for
identifying common genetic variants, which have led to the discovery of common
risk loci for several complex disorders (Day-Williams et al. 2011, Parkes et al.
2007, Sladek et al. 2007, Wellcome Trust Case Control Consortium 2007).
However, at the moment (October 2012) only one GWAS has been published to
address low back disorders (Williams et al. 2012). For further refined discovery
of risk variants for several complex disorders, efforts towards incorporation of
whole-exome or whole-genome sequencing approaches, due to increased capacity
and accuracy of next-generation sequencing, are possible today. While these new
approaches are also possible tools to be used in future studies to increase our
understanding of the cascades of DD, the amount of sequence information
obtained from these approaches is huge, and robust methods of bioinformatics are
required to handle the raw data. Furthermore, new possibilities to define DD with
specific MRI techniques have emerged (Blumenkrantz et al. 2010, Borthakur et al.

113
2011, Johannessen et al. 2006, Zou et al. 2009, Zuo et al. 2009). The new ultra-
high-field MRIs (7 tesla and above), that are still rare even in scientific use today,
will likely provide interesting new opportunities to investigate DD in vivo
(Madelin et al. 2012, Regatte & Schweitzer 2007). The present results of the
systematic review (I) emphasize the importance of well-defined phenotypes of
DD, which should be taken into consideration in future studies. Furthermore, the
gender differences observed here (II, III, IV) in associations between genetic
polymorphisms and DD/DDP should be noted and analyses should be made also
for possible gender-genotype interactions.

114

115
7 Conclusions
In conclusion, the results of this thesis indicate that DD defined by MRI is
common already at an early age and suggest that polymorphisms in IL6, SKT,
CILP and IL1A may be important in the development of DD. These results mostly
support earlier findings in adults. However, the population sizes in our studies
were limited, and thus the small effect sizes may not have been robustly detected.
Therefore, it is possible that some of the associations we were not able to observe
in this study could be significant in a larger population. When the cumulative
evidence of previous studies was taken into account, the association in SKT
reached the level of moderate association evidence. However, we investigated for
the first time associations between genetic polymorphisms and DD or progression
of DD in teenagers. Thus, this thesis was the first building block in the “house” of
cumulative evidence of genetics of DD among teenagers, adolescents or young
adults. The findings of this thesis are, however, preliminary, and require further
investigations in a larger setting.
In the systematic analysis of previous reports the polymorphisms in GDF5,
THBS2, MMP9, COL11A1, SKT and ASPN were found to have a moderate level
of association evidence. The majority of these associations were seen in cases of a
clinical DD phenotype; LDH characterized by sciatica. Multiple other
polymorphisms have also been previously reported to associate with the
development of DD. However, due to variations between study designs, sampling
methods, populations, and phenotype definitions, the level of cumulative evidence
is currently modest for the other polymorphims. In order to decrease the
discrepancy between studies, collaboration between investigators and large
population-based cohorts and well-defined phenotypes are required.
Body height was associated with DD in both of the current study populations.
While height has been reported to be associated with LDH, LBP and risk for back
surgery, no reports on the association between height and DD has been published
previously. The mechanism beyond the observed association is not known but it
may relate to early growth pattern.
We observed gender differences in the genetic associations with DD among
Danish teenagers in IL1A and IL6 but not among young Finnish adults, where the
association-difference between genders was seen only in CILP. The gender-
specific effects of genetic risk factors are not well known in DD. These
preliminary results suggest possible gender-genotype interaction in IL1A, IL6 and

116
CILP but a replication in a larger setting is needed before any solid conclusions
can be made.
This thesis succeeded in increasing the knowledge of the hereditary
component in lumbar DD. However, the majority of the genetic component in DD
is still unexplained. The more advanced genotyping methods, new imaging
modalities and robust methods in bioinformatics are likely to provide us more
information about this interesting and important condition.

117
References
A Cecile J W Janssens, John P A Ioannidis, Cornelia M van Duijn, Julian Little & Muin J Khoury (2011) Strengthening the reporting of genetic risk prediction studies: the GRIPS statement. BMJ 342.
Adams MA, Freeman BJ, Morrison HP, Nelson IW & Dolan P (2000) Mechanical initiation of intervertebral disc degeneration. Spine (Phila Pa 1976) 25(13): 1625–1636.
Adams MA, McNally DS & Dolan P (1996) 'Stress' distributions inside intervertebral discs. The effects of age and degeneration. J Bone Joint Surg Br 78(6): 965–972.
Adams MA & Roughley PJ (2006) What is intervertebral disc degeneration, and what causes it? Spine (Phila Pa 1976) 31(18): 2151–2161.
Adams MA, Stefanakis M & Dolan P (2010) Healing of a painful intervertebral disc should not be confused with reversing disc degeneration: implications for physical therapies for discogenic back pain. Clin Biomech (Bristol, Avon) 25(10): 961–971.
Ala-Kokko L (2002) Genetic risk factors for lumbar disc disease. Ann Med 34(1): 42–47. Albert HB, Briggs AM, Kent P, Byrhagen A, Hansen C & Kjaergaard K (2011) The
prevalence of MRI-defined spinal pathoanatomies and their association with Modic changes in individuals seeking care for low back pain. Eur Spine J 20(8): 1355–1362.
Altmuller J, Palmer LJ, Fischer G, Scherb H & Wjst M (2001) Genomewide scans of complex human diseases: true linkage is hard to find. Am J Hum Genet 69(5): 936–950.
Altshuler D, Daly MJ & Lander ES (2008) Genetic Mapping in Human Disease. Science 322(5903): 881–888.
An HS, Anderson PA, Haughton VM, Iatridis JC, Kang JD, Lotz JC, Natarajan RN, Oegema TR,Jr, Roughley P, Setton LA, Urban JP, Videman T, Andersson GB & Weinstein JN (2004) Introduction: disc degeneration: summary. Spine (Phila Pa 1976) 29(23): 2677–2678.
Andersen LB, Wedderkopp N, Hansen HS, Cooper AR & Froberg K (2003) Biological cardiovascular risk factors cluster in Danish children and adolescents: the European Youth Heart Study. Prev Med 37(4): 363–367.
Andersson GB (1999) Epidemiological features of chronic low-back pain. Lancet 354(9178): 581–585.
Annunen S, Paassilta P, Lohiniva J, Perala M, Pihlajamaa T, Karppinen J, Tervonen O, Kroger H, Lahde S, Vanharanta H, Ryhanen L, Goring HH, Ott J, Prockop DJ & Ala-Kokko L (1999) An allele of COL9A2 associated with intervertebral disc disease. Science 285(5426): 409–412.
Antoniou J, Steffen T, Nelson F, Winterbottom N, Hollander AP, Poole RA, Aebi M & Alini M (1996) The human lumbar intervertebral disc: evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J Clin Invest 98(4): 996–1003.
Aprill C & Bogduk N (1992) High-intensity zone: a diagnostic sign of painful lumbar disc on magnetic resonance imaging. Br J Radiol 65(773): 361–369.

118
Aszodi A, Chan D, Hunziker E, Bateman JF & Fassler R (1998) Collagen II is essential for the removal of the notochord and the formation of intervertebral discs. J Cell Biol 143(5): 1399–1412.
Auvinen J (2010) Neck, shoulder, and low back pain adolescence. Acta Univ Oul D 1052 Bailey-Wilson JE & Wilson AF (2011) Linkage analysis in the next-generation sequencing
era. Hum Hered 72(4): 228–236. Battie MC, Haynor DR, Fisher LD, Gill K, Gibbons LE & Videman T (1995a) Similarities
in degenerative findings on magnetic resonance images of the lumbar spines of identical twins. J Bone Joint Surg Am 77(11): 1662–1670.
Battie MC, Videman T, Gibbons LE, Fisher LD, Manninen H & Gill K (1995b) 1995 Volvo Award in clinical sciences. Determinants of lumbar disc degeneration. A study relating lifetime exposures and magnetic resonance imaging findings in identical twins. Spine (Phila Pa 1976) 20(24): 2601–2612.
Battie MC, Videman T, Gill K, Moneta GB, Nyman R, Kaprio J & Koskenvuo M (1991) 1991 Volvo Award in clinical sciences. Smoking and lumbar intervertebral disc degeneration: an MRI study of identical twins. Spine (Phila Pa 1976) 16(9): 1015–1021.
Battie MC, Videman T, Kaprio J, Gibbons LE, Gill K, Manninen H, Saarela J & Peltonen L (2009) The Twin Spine Study: contributions to a changing view of disc degeneration. Spine J 9(1): 47–59.
Battie MC & Videman T (2006) Lumbar disc degeneration: epidemiology and genetics. J Bone Joint Surg Am 88 Suppl 2: 3–9.
Battie MC, Videman T & Parent E (2004) Lumbar disc degeneration: epidemiology and genetic influences. Spine (Phila Pa 1976) 29(23): 2679–2690.
Bei T, Tilkeridis C, Garantziotis S, Boikos S, Kazakos K, Simopoulos C & Stratakis C (2008) A novel, non-functional, COL1A1 polymorphism is not associated with lumbar disk disease in young male Greek subjects unlike that of the Sp1 site. Hormones (Athens) 7(3): 251–254.
Berlemann U, Gries NC & Moore RJ (1998) The relationship between height, shape and histological changes in early degeneration of the lower lumbar discs. Eur Spine J 7(3): 212–217.
Bernick S & Cailliet R (1982) Vertebral end-plate changes with aging of human vertebrae. Spine (Phila Pa 1976) 7(2): 97–9102.
Bhardwaj R & Midha R (2004) Synchronous lumbar disc herniation in adult twins. Case report. Can J Neurol Sci 31(4): 554–557.
Bijkerk C, Houwing-Duistermaat JJ, Valkenburg HA, Meulenbelt I, Hofman A, Breedveld FC, Pols HA, van Duijn CM & Slagboom PE (1999) Heritabilities of radiologic osteoarthritis in peripheral joints and of disc degeneration of the spine. Arthritis Rheum 42(8): 1729–1735.
Blumenkrantz G, Zuo J, Li X, Kornak J, Link TM & Majumdar S (2010) In vivo 3.0-tesla magnetic resonance T1rho and T2 relaxation mapping in subjects with intervertebral disc degeneration and clinical symptoms. Magn Reson Med 63(5): 1193–1200.

119
Bodmer W & Bonilla C (2008) Common and rare variants in multifactorial susceptibility to common diseases. Nat Genet 40(6): 695–701.
Boos N, Wallin A, Aebi M & Boesch C (1996) A new magnetic resonance imaging analysis method for the measurement of disc height variations. Spine (Phila Pa 1976) 21(5): 563–570.
Boos N, Weissbach S, Rohrbach H, Weiler C, Spratt KF & Nerlich AG (2002) Classification of age-related changes in lumbar intervertebral discs: 2002 Volvo Award in basic science. Spine (Phila Pa 1976) 27(23): 2631–2644.
Borenstein DG, O'Mara JW, Boden SD, Lauerman WC, Jacobson A, Platenberg C, Schellinger D & Wiesel SW (2001) The value of magnetic resonance imaging of the lumbar spine to predict low-back pain in asymptomatic subjects : a seven-year follow-up study. J Bone Joint Surg Am 83-A(9): 1306–1311.
Borthakur A, Maurer PM, Fenty M, Wang C, Berger R, Yoder J, Balderston RA & Elliott DM (2011) T1ρ magnetic resonance imaging and discography pressure as novel biomarkers for disc degeneration and low back pain. Spine (Phila Pa 1976) 36(25): 2190–2196.
Bouzigon E, Forabosco P, Koppelman GH, Cookson WOCM, Dizier M, Duffy DL, Evans DM, Ferreira MAR, Kere J, Laitinen T, Malerba G, Meyers DA, Moffatt M, Martin NG, Ng MY, Pignatti PF, Wjst M, Kauffmann F, Demenais F & Lewis CM (2010) Meta-analysis of 20 genome-wide linkage studies evidenced new regions linked to asthma and atopy. Eur J Hum Genet 18(6): 700–706.
Bracken MB (2005) Genomic epidemiology of complex disease: the need for an electronic evidence-based approach to research synthesis. Am J Epidemiol 162(4): 297–301.
Bradford DS & Garcia A (1971) Lumbar intervertebral disk herniations in children and adolescents. Orthop Clin North Am 2(2): 583–592.
Brinckmann P & Horst M (1985) The influence of vertebral body fracture, intradiscal injection, and partial discectomy on the radial bulge and height of human lumbar discs. Spine (Phila Pa 1976) 10(2): 138–145.
Brinkhaus B, Lewith G, Rehberg B, Heusser P, Cummings M, Michalsen A, Teut M, Willich SN & Irnich D (2011) How to treat a patient with chronic low back pain - methodology and results of the first international case conference of integrative medicine. Complement Ther Med 19(1): 54–62.
Bull J, el Gammal T & Popham M (1969) A possible genetic factor in cervical spondylosis. Br J Radiol 42(493): 9–16.
Burke JG, Watson RW, McCormack D, Dowling FE, Walsh MG & Fitzpatrick JM (2002) Intervertebral discs which cause low back pain secrete high levels of proinflammatory mediators. J Bone Joint Surg Br 84(2): 196–201.
Cahill J, Frost G & Solanki GA (2011) Paediatric lumbar disc herniation in the very young: a case-based update. Childs Nerv Syst 27(5): 687–691.
Carragee EJ, Don AS, Hurwitz EL, Cuellar JM, Carrino JA, Carrino J & Herzog R (2009) 2009 ISSLS Prize Winner: Does discography cause accelerated progression of degeneration changes in the lumbar disc: a ten-year matched cohort study. Spine (Phila Pa 1976) 34(21): 2338–2345.

120
Carrino JA, Lurie JD, Tosteson AN, Tosteson TD, Carragee EJ, Kaiser J, Grove MR, Blood E, Pearson LH, Weinstein JN & Herzog R (2009) Lumbar spine: reliability of MR imaging findings. Radiology 250(1): 161–170.
Casimir GJ, Heldenbergh F, Hanssens L, Mulier S, Heinrichs C, Lefevre N, Desir J, Corazza F & Duchateau J (2010) Gender differences and inflammation: an in vitro model of blood cells stimulation in prepubescent children. J Inflamm (Lond) 7: 28–28.
Chan D, Song Y, Sham P & Cheung KMC (2006) Genetics of disc degeneration. Eur Spine J 15 Suppl 3: 317–325.
Chan WCW, Sze KL, Samartzis D, Leung VYL & Chan D (2011) Structure and biology of the intervertebral disk in health and disease. Orthop Clin North Am 42(4): 447–464.
Chambers JC, Zhang W, Sehmi J, Li X, Wass MN, Van der Harst P, Holm H, Sanna S, Kavousi M, Baumeister SE, Coin LJ, Deng G, Gieger C, Heard-Costa NL, Hottenga JJ, Kuhnel B, Kumar V, Lagou V, Liang L, Luan J, Vidal PM, Mateo Leach I, O'Reilly PF, Peden JF, Rahmioglu N, Soininen P, Speliotes EK, Yuan X, Thorleifsson G, Alizadeh BZ, Atwood LD, Borecki IB, Brown MJ, Charoen P, Cucca F, Das D, de Geus EJ, Dixon AL, Doring A, Ehret G, Eyjolfsson GI, Farrall M, Forouhi NG, Friedrich N, Goessling W, Gudbjartsson DF, Harris TB, Hartikainen AL, Heath S, Hirschfield GM, Hofman A, Homuth G, Hypp\onen E, Janssen HL, Johnson T, Kangas AJ, Kema IP, Kuhn JP, Lai S, Lathrop M, Lerch MM, Li Y, Liang TJ, Lin JP, Loos RJ, Martin NG, Moffatt MF, Montgomery GW, Munroe PB, Musunuru K, Nakamura Y, O'Donnell CJ, Olafsson I, Penninx BW, Pouta A, Prins BP, Prokopenko I, Puls R, Ruokonen A, Savolainen MJ, Schlessinger D, Schouten JN, Seedorf U, Sen-Chowdhry S, Siminovitch KA, Smit JH, Spector TD, Tan W, Teslovich TM, Tukiainen T, Uitterlinden AG, Van der Klauw MM, Vasan RS, Wallace C, Wallaschofski H, Wichmann HE, Willemsen G, Wurtz P, Xu C, Yerges-Armstrong LM, Alcohol Genome-wide Association (AlcGen) Consortium, Diabetes Genetics Replication, Meta-analyses (DIAGRAM+) Study, Genetic Investigation of Anthropometric Traits (GIANT) Consortium, Global Lipids Genetics Consortium, Genetics of Liver Disease (GOLD) Consortium, International Consortium for Blood Pressure, Meta-analyses of Glucose, Insulin-Related Traits Consortium, Abecasis GR, Ahmadi KR, Boomsma DI, Caulfield M, Cookson WO, van Duijn CM, Froguel P, Matsuda K, McCarthy MI, Meisinger C, Mooser V, Pietil\ainen KH, Schumann G, Snieder H, Sternberg MJ, Stolk RP, Thomas HC, Thorsteinsdottir U, Uda M, Waeber G, Wareham NJ, Waterworth DM, Watkins H, Whitfield JB, Witteman JC, Wolffenbuttel BH, Fox CS, Ala-Korpela M, Stefansson K, Vollenweider P, Volzke H, Schadt EE, Scott J, Järvelin MR, Elliott P & Kooner JS (2011) Genome-wide association study identifies loci influencing concentrations of liver enzymes in plasma. Nat Genet 43(11): 1131–1138.
Cheung KMC & Al Ghazi S (2008) (i) Current understanding of low back pain and intervertebral disc degeneration: epidemiological perspectives and phenotypes for genetic studies Current Orthopaedics 22(4): 237–244.

121
Cheung KMC, Chan D, Karppinen J, Chen Y, Jim JJT, Yip S, Ott J, Wong KK, Sham P, Luk KDK, Cheah KSE, Leong JCY & Song Y (2006) Association of the Taq I allele in vitamin D receptor with degenerative disc disease and disc bulge in a Chinese population. Spine (Phila Pa 1976) 31(10): 1143–1148.
Cheung KMC, Samartzis D, Karppinen J & Luk KDK (2012) Are "patterns" of lumbar disc degeneration associated with low back pain?: new insights based on skipped level disc pathology. Spine (Phila Pa 1976) 37(7): 430–438.
Choi M, Scholl UI, Ji W, Liu T, Tikhonova IR, Zumbo P, Nayir A, Bakkalolu A, Ozen S, Sanjad S, Nelson-Williams C, Farhi A, Mane S & Lifton RP (2009) Genetic diagnosis by whole exome capture and massively parallel DNA sequencing. Proc Natl Acad Sci U S A 106(45): 19096–19101.
Choi M, Scholl UI, Yue P, Bjorklund P, Zhao B, Nelson-Williams C, Ji W, Cho Y, Patel A, Men CJ, Lolis E, Wisgerhof MV, Geller DS, Mane S, Hellman P, Westin G, Åkerström G, Wang W, Carling T & Lifton RP (2011) K+ channel mutations in adrenal aldosterone-producing adenomas and hereditary hypertension. Science 331(6018): 768–772.
Chou D, Samartzis D, Bellabarba C, Patel A, Luk KDK, Kisser JMS & Skelly AC (2011) Degenerative magnetic resonance imaging changes in patients with chronic low back pain: a systematic review. Spine (Phila Pa 1976) 36(21): 43–53.
Chou R, Fu R, Carrino JA & Deyo RA (2009) Imaging strategies for low-back pain: systematic review and meta-analysis. Lancet 373(9662): 463–472.
Clouet J, Vinatier C, Merceron C, Pot-Vaucel M, Hamel O, Weiss P, Grimandi G & Guicheux J (2009) The intervertebral disc: from pathophysiology to tissue engineering. Joint Bone Spine 76(6): 614–618.
Coeuret-Pellicer M, Descatha A, Leclerc A & Zins M (2010) Are tall people at higher risk of low back pain surgery? A discussion on the results of a multipurpose cohort. Arthritis Care Res (Hoboken) 62(1): 125–127.
Cong L, Pang H, Xuan D & Tu GJ (2010a) Association between the expression of aggrecan and the distribution of aggrecan gene variable number of tandem repeats with symptomatic lumbar disc herniation in Chinese Han of Northern China. Spine (Phila Pa 1976) 35(14): 1371–1376.
Cong L, Pang H, Xuan D & Tu G (2010b) The interaction between aggrecan gene VNTR polymorphism and cigarette smoking in predicting incident symptomatic intervertebral disc degeneration. Connect Tissue Res 51(5): 397–403.
Comino-Mendez I, Gracia-Aznarez FJ, Schiavi F, Landa I, Leandro-Garcia LJ, Leton R, Honrado E, Ramos-Medina R, Caronia D, Pita G, Gomez-Grana A, de Cubas AA, Inglada-Perez L, Maliszewska A, Taschin E, Bobisse S, Pica G, Loli P, Hernandez-Lavado R, Diaz JA, Gomez-Morales M, Gonzalez-Neira A, Roncador G, Rodriguez-Antona C, Benitez J, Mannelli M, Opocher G, Robledo M & Cascon A (2011) Exome sequencing identifies MAX mutations as a cause of hereditary pheochromocytoma. Nat Genet 43(7): 663–667
Cookson W, Liang L, Abecasis G, Moffatt M, Lathrop M (2009) Mapping complex disease traits with global gene expression. Nat Rev Genet. 2009 Mar;10(3):184–94.

122
Coulombe-Huntington J, Lam KCL, Dias C & Majewski J (2009) Fine-scale variation and genetic determinants of alternative splicing across individuals. PLoS Genet 5(12).
Dagenais S, Caro J & Haldeman S (2008) A systematic review of low back pain cost of illness studies in the United States and internationally. Spine J 8(1): 8–20.
Dang L & Liu Z (2010) A review of current treatment for lumbar disc herniation in children and adolescents. Eur Spine J 19(2): 205–214.
Dawn Teare M & Barrett JH (2005) Genetic linkage studies. Lancet 366(9490): 1036–1044.
Day-Williams AG, Southam L, Panoutsopoulou K, Rayner NW, Esko T, Estrada K, Helgadottir HT, Hofman A, Ingvarsson T, Jonsson H, Keis A, Kerkhof HJM, Thorleifsson G, Arden NK, Carr A, Chapman K, Deloukas P, Loughlin J, McCaskie A, Ollier WER, Ralston SH, Spector TD, Wallis GA, Wilkinson JM, Aslam N, Birell F, Carluke I, Joseph J, Rai A, Reed M, Walker K, , Doherty SA, Jonsdottir I, Maciewicz RA, Muir KR, Metspalu A, Rivadeneira F, Stefansson K, Styrkarsdottir U, Uitterlinden AG, van Meurs JBJ, Zhang W, Valdes AM, Doherty M & Zeggini E (2011) A variant in MCF2L is associated with osteoarthritis. Am J Hum Genet 89(3): 446–450.
de Schepper EIT, Damen J, van Meurs JBJ, Ginai AZ, Popham M, Hofman A, Koes BW & Bierma-Zeinstra SM (2010) The association between lumbar disc degeneration and low back pain: the influence of age, gender, and individual radiographic features. Spine (Phila Pa 1976) 35(5): 531–536.
DeGroot J, Verzijl N, Wenting-van Wijk MJG, Jacobs KMG, Van El B, Van Roermund PM, Bank RA, Bijlsma JWJ, TeKoppele JM & Lafeber FPJG (2004) Accumulation of advanced glycation end products as a molecular mechanism for aging as a risk factor in osteoarthritis. Arthritis Rheum 50(4): 1207–1215.
DeLeo JA (2006) Basic science of pain. J Bone Joint Surg Am 88 Suppl 2: 58–62. DePalma MJ, Ketchum JM & Saullo T (2011) What is the source of chronic low back pain
and does age play a role? Pain Med 12(2): 224–233. Deyo RA & Tsui-Wu YJ (1987) Descriptive epidemiology of low-back pain and its related
medical care in the United States. Spine (Phila Pa 1976) 12(3): 264–268. Diepenmaat ACM, van dW, de Vet HCW & Hirasing RA (2006) Neck/shoulder, low back,
and arm pain in relation to computer use, physical activity, stress, and depression among Dutch adolescents. Pediatrics 117(2): 412–416.
Dimas AS, Deutsch S, Stranger BE, Montgomery SB, Borel C, Attar-Cohen H, Ingle C, Beazley C, Gutierrez Arcelus M, Sekowska M, Gagnebin M, Nisbett J, Deloukas P, Dermitzakis ET & Antonarakis SE (2009) Common regulatory variation impacts gene expression in a cell type-dependent manner. Science 325(5945): 1246–1250.

123
Dionne CE, Dunn KM, Croft PR, Nachemson AL, Buchbinder R, Walker BF, Wyatt M, Cassidy JD, Rossignol M, Leboeuf-Yde C, Hartvigsen J, Leino-Arjas P, Latza U, Reis S, Gil Del Real MT, Kovacs FM, Oberg B, Cedraschi C, Bouter LM, Koes BW, Picavet HS, van Tulder MW, Burton K, Foster NE, Macfarlane GJ, Thomas E, Underwood M, Waddell G, Shekelle P, Volinn E & Von Korff M (2008) A consensus approach toward the standardization of back pain definitions for use in prevalence studies. Spine (Phila Pa 1976) 33(1): 95–103.
Dong DM, Yao M, Liu B, Sun CY, Jiang YQ & Wang YS (2007) Association between the -1306C/T polymorphism of matrix metalloproteinase-2 gene and lumbar disc disease in Chinese young adults. Eur Spine J 16(11): 1958–1961.
Eichler EE, Flint J, Gibson G, Kong A, Leal SM, Moore JH & Nadeau JH (2010) Missing heritability and strategies for finding the underlying causes of complex disease. Nat Rev Genet 11(6): 446–450.
Ekman M, Johnell O & Lidgren L (2005) The economic cost of low back pain in Sweden in 2001. Acta Orthop 76(2): 275–284.
Elfering A, Semmer N, Birkhofer D, Zanetti M, Hodler J & Boos N (2002) Risk factors for lumbar disc degeneration: a 5-year prospective MRI study in asymptomatic individuals. Spine (Phila Pa 1976) 27(2): 125–134.
Emch TM & Modic MT (2011) Imaging of lumbar degenerative disk disease: history and current state. Skeletal Radiol 40(9): 1175–1189.
Endean A, Palmer KT & Coggon D (2011) Potential of magnetic resonance imaging findings to refine case definition for mechanical low back pain in epidemiological studies: a systematic review. Spine (Phila Pa 1976) 36(2): 160–169.
Engvall E, Hessle H & Klier G (1986) Molecular assembly, secretion, and matrix deposition of type VI collagen. J Cell Biol 102(3): 703–710.
Eser B, Cora T, Eser O, Kalkan E, Haktanir A, Erdogan MO & Solak M (2010) Association of the polymorphisms of vitamin D receptor and aggrecan genes with degenerative disc disease. Genet Test Mol Biomarkers 14(3): 313–317.
Eskola PJ, Kjaer P, Daavittila IM, Solovieva S, Okuloff A, Sorensen JS, Wedderkopp N, Ala-Kokko L, Mannikko M & Karppinen JI (2010) Genetic risk factors of disc degeneration among 12–14-year-old Danish children: a population study Int J Mol Epidemiol Genet 1(2): 158–165.
Eyre D, Benya P, Buckwalter J, Caterson B, Heinegård D, Oegema T, Pearce R, Pope M & Urban J (1989) Intervertebral disk: Basic science perspectives. In Frymoyer JW GS (ed) New Perspectives on Low Back Pain. Park Ridge, American Academy of Orthopaedic Surgeons: 147–207.
Eyre DR & Muir H (1977) Quantitative analysis of types I and II collagens in human intervertebral discs at various ages. Biochim Biophys Acta 492(1): 29–42.
Fairbrother UL, Tanko LB, Walley AJ, Christiansen C, Froguel P, Blakemore AI (2007) Leptin receptor genotype at Gln223Arg is associated with body composition, BMD, and vertebral fracture in postmenopausal Danish women. J Bone Miner Res: 22(4):544–550.

124
Falconer, DS, MacKay TFC. Introduction to Quantitative Genetics, 4th Ed. 1996. Longmans Green, Harlow, Essex, UK.
Fardon DF, Milette PC & (2001) Nomenclature and classification of lumbar disc pathology. Recommendations of the Combined task Forces of the North American Spine Society, American Society of Spine Radiology, and American Society of Neuroradiology. Spine (Phila Pa 1976) 26(5): 93–93.
Fleming A, Keynes RJ & Tannahill D (2001) The role of the notochord in vertebral column formation. J Anat 199(1–2): 177–180.
Fontecha CG, Balague F, Pellise F, Rajmil L, Aguirre M, Pasarin M, Cedraschi C & Ferrer M (2011) Low Back Pain in Adolescents: Is Quality of Life Poorer in Those Seeking Medical Attention? Spine (Phila Pa 1976) 36(17): 1154–1154.
Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM, Lindgren CM, Perry JRB, Elliott KS, Lango H, Rayner NW, Shields B, Harries LW, Barrett JC, Ellard S, Groves CJ, Knight B, Patch A, Ness AR, Ebrahim S, Lawlor DA, Ring SM, Ben-Shlomo Y, Jarvelin M, Sovio U, Bennett AJ, Melzer D, Ferrucci L, Loos RJF, Barroso I, Wareham NJ, Karpe F, Owen KR, Cardon LR, Walker M, Hitman GA, Palmer CNA, Doney ASF, Morris AD, Smith GD, Hattersley AT & McCarthy MI (2007) A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 316(5826): 889–894.
Frobin W, Brinckmann P, Kramer M & Hartwig E (2001) Height of lumbar discs measured from radiographs compared with degeneration and height classified from MR images. Eur Radiol 11(2): 263–269.
Frymoyer JW (1992) Lumbar disk disease: epidemiology. Instr Course Lect 41: 217–223. Frymoyer JW (1988) Back pain and sciatica. N Engl J Med 318(5): 291–300. Fujiwara A, Tamai K, Yamato M, An HS, Yoshida H, Saotome K & Kurihashi A (1999)
The relationship between facet joint osteoarthritis and disc degeneration of the lumbar spine: an MRI study. Eur Spine J 8(5): 396–401.
Gabay C, Lamacchia C & Palmer G (2010) IL-1 pathways in inflammation and human diseases. Nat Rev Rheumatol 6(4): 232–241.
Gabriel SB, Schaffner SF, Nguyen H, Moore JM, Roy J, Blumenstiel B, Higgins J, DeFelice M, Lochner A, Faggart M, Liu-Cordero SN, Rotimi C, Adeyemo A, Cooper R, Ward R, Lander ES, Daly MJ & Altshuler D (2002) The structure of haplotype blocks in the human genome. Science 296(5576): 2225–2229.
Gamazon ER, Nicolae DL & Cox NJ (2011) A study of CNVs as trait-associated polymorphisms and as expression quantitative trait loci. PLoS Genet 7(2).
Gao X, Becker LC, Becker DM, Starmer JD & Province MA (2010) Avoiding the high Bonferroni penalty in genome-wide association studies. Genet Epidemiol 34(1): 100–105.
Gellhorn AC, Chan L, Martin B & Friedly J (2010) Management Patterns in Acute Low Back Pain: The Role of Physical Therapy. Spine (Phila Pa 1976).
Gerrits A, Li Y, Tesson BM, Bystrykh LV, Weersing E, Ausema A, Dontje B, Wang X, Breitling R, Jansen RC & de Haan G (2009) Expression quantitative trait loci are highly sensitive to cellular differentiation state. PLoS Genet 5(10).

125
Girard SL, Gauthier J, Noreau A, Xiong L, Zhou S, Jouan L, Dionne-Laporte A, Spiegelman D, Henrion E, Diallo O, Thibodeau P, Bachand I, Bao JYJ, Tong A,Hin Yan, Lin C, Millet B, Jaafari N, Joober R, Dion PA, Lok S, Krebs M & Rouleau GA (2011) Increased exonic de novo mutation rate in individuals with schizophrenia. Nat Genet 43(9): 860–863.
Gitelman A, Hishmeh S, Morelli BN, Joseph SA, Casden A, Kuflik P, Neuwirth M & Stephen M (2008) Cauda equina syndrome: a comprehensive review. Am J Orthop (Belle Mead NJ) 37(11): 556–562.
Goldring MB, Birkhead JR, Suen LF, Yamin R, Mizuno S, Glowacki J, Arbiser JL & Apperley JF (1994) Interleukin-1 beta-modulated gene expression in immortalized human chondrocytes. J Clin Invest 94(6): 2307–2316.
Griffith JF, Wang YX, Antonio GE, Choi KC, Yu A, Ahuja AT & Leung PC (2007) Modified Pfirrmann grading system for lumbar intervertebral disc degeneration. Spine (Phila Pa 1976) 32(24): E708–12.
Grobler LJ, Simmons EH & Barrington TW (1979) Intervertebral disc herniation in the adolescent. Spine (Phila Pa 1976) 4(3): 267–278.
Gruber HE & Hanley EN (1998) Analysis of aging and degeneration of the human intervertebral disc. Comparison of surgical specimens with normal controls. Spine (Phila Pa 1976) 23(7): 751–757.
Gruber HE, Ingram JA, Hoelscher GL, Zinchenko N, Hanley EN & Sun Y (2009) Asporin, a susceptibility gene in osteoarthritis, is expressed at higher levels in the more degenerate human intervertebral disc. Arthritis Res Ther 11(2).
Grunhagen T, Wilde G, Soukane DM, Shirazi-Adl SA & Urban JPG (2006) Nutrient supply and intervertebral disc metabolism. J Bone Joint Surg Am 88 Suppl 2: 30–35.
Guehring T, Urban JP, Cui Z & Tirlapur UK (2008) Noninvasive 3D vital imaging and characterization of notochordal cells of the intervertebral disc by femtosecond near-infrared two-photon laser scanning microscopy and spatial-volume rendering. Microsc Res Tech 71(4): 298–304.
Guerin HL & Elliott DM (2007) Quantifying the contributions of structure to annulus fibrosus mechanical function using a nonlinear, anisotropic, hyperelastic model. J Orthop Res 25(4): 508–516.
Guinto FC, Hashim H & Stumer M (1984) CT demonstration of disk regression after conservative therapy. AJNR Am J Neuroradiol 5(5): 632–633.
Gunzburg R, Fraser RD & Fraser GA (1990) Lumbar intervertebral disc prolapse in teenage twins. A case report and review of the literature. J Bone Joint Surg Br 72(5): 914–916.
Göring HH, Curran JE, Johnson MP, Dyer TD, Charlesworth J, Cole SA, Jowett JB, Abraham LJ, Rainwater DL, Comuzzie AG, Mahaney MC, Almasy L, MacCluer JW, Kissebah AH, Collier GR, Moses EK & Blangero J (2007) Discovery of expression QTLs using large-scale transcriptional profiling in human lymphocytes. Nat Genet 39(10): 1208–1216.

126
Haberlandt E, Spreiz A, Sigl SB, Janetschek C, Rothlisberger B, Zotter S, Rostasy K, Zschocke J & Kotzot D (2012) Microdeletion 19p13.2 in an almost 5-year-old boy. Am J Med Genet A 158A(5): 1190–1194.
Hancock MJ, Battie MC, Videman T & Gibbons L (2010) The role of back injury or trauma in lumbar disc degeneration: an exposure-discordant twin study. Spine (Phila Pa 1976) 35(21): 1925–1929.
Hangai M, Kaneoka K, Kuno S, Hinotsu S, Sakane M, Mamizuka N, Sakai S & Ochiai N (2008) Factors associated with lumbar intervertebral disc degeneration in the elderly. Spine J 8(5): 732–740.
Hangai M, Kaneoka K, Hinotsu S, Shimizu K, Okubo Y, Miyakawa S, Mukai N, Sakane M & Ochiai N (2009) Lumbar intervertebral disk degeneration in athletes. Am J Sports Med 37(1): 149–155.
Hassett G, Hart DJ, Manek NJ, Doyle DV & Spector TD (2003) Risk factors for progression of lumbar spine disc degeneration: the Chingford Study. Arthritis Rheum 48(11): 3112–3117.
Haughton V (2006) Imaging intervertebral disc degeneration. J Bone Joint Surg Am 88 Suppl 2: 15–20.
Heliovaara M (1989) Risk factors for low back pain and sciatica. Ann Med 21(4): 257–264. Heuer F, Schmidt H & Wilke H (2008) The relation between intervertebral disc bulging
and annular fiber associated strains for simple and complex loading. J Biomech 41(5): 1086–1094.
Heuer F, Schmitt H, Schmidt H, Claes L & Wilke H (2007) Creep associated changes in intervertebral disc bulging obtained with a laser scanning device. Clin Biomech (Bristol, Avon) 22(7): 737–744.
Higashino K, Matsui Y, Yagi S, Takata Y, Goto T, Sakai T, Katoh S & Yasui N (2007) The alpha2 type IX collagen tryptophan polymorphism is associated with the severity of disc degeneration in younger patients with herniated nucleus pulposus of the lumbar spine. Int Orthop 31(1): 107–111.
Hirose Y, Chiba K, Karasugi T, Nakajima M, Kawaguchi Y, Mikami Y, Furuichi T, Mio F, Miyake A, Miyamoto T, Ozaki K, Takahashi A, Mizuta H, Kubo T, Kimura T, Tanaka T, Toyama Y & Ikegawa S (2008) A functional polymorphism in THBS2 that affects alternative splicing and MMP binding is associated with lumbar-disc herniation. Am J Hum Genet 82(5): 1122–1129.
Hirschhorn JN & Gajdos ZKZ (2011) Genome-wide association studies: results from the first few years and potential implications for clinical medicine. Annu Rev Med 62: 11–24.
Holm S & Nachemson A (1988) Nutrition of the intervertebral disc: acute effects of cigarette smoking. An experimental animal study. Ups J Med Sci 93(1): 91–99.
Holm S, Holm AK, Ekstrom L, Karladani A & Hansson T (2004) Experimental disc degeneration due to endplate injury. J Spinal Disord Tech 17(1): 64–71.
Hsieh AH & Twomey JD (2010) Cellular mechanobiology of the intervertebral disc: new directions and approaches. J Biomech 43(1): 137–145.

127
Humzah MD & Soames RW (1988) Human intervertebral disc: structure and function. Anat Rec 220(4): 337–356.
Hutton WC, Murakami H, Li J, Elmer WA, Yoon ST, Minamide A, Akamaru T & Tomita K (2004) The effect of blocking a nutritional pathway to the intervertebral disc in the dog model. J Spinal Disord Tech 17(1): 53–63.
International Hap Map Consortium (2005) A haplotype map of the human genome. Nature 437(7063): 1299–1320.
International Hap Map Consortium, Frazer KA, Ballinger DG, Cox DR, Hinds DA, Stuve LL, Gibbs RA, Belmont JW, Boudreau A, Hardenbol P, Leal SM, Pasternak S, Wheeler DA, Willis TD, Yu F, Yang H, Zeng C, Gao Y, Hu H, Hu W, Li C, Lin W, Liu S, Pan H, Tang X, Wang J, Wang W, Yu J, Zhang B, Zhang Q, Zhao H, Zhao H, Zhou J, Gabriel SB, Barry R, Blumenstiel B, Camargo A, Defelice M, Faggart M, Goyette M, Gupta S, Moore J, Nguyen H, Onofrio RC, Parkin M, Roy J, Stahl E, Winchester E, Ziaugra L, Altshuler D, Shen Y, Yao Z, Huang W, Chu X, He Y, Jin L, Liu Y, Shen Y, Sun W, Wang H, Wang Y, Wang Y, Xiong X, Xu L, Waye MMY, Tsui SKW, Xue H, Wong JT, Galver LM, Fan J, Gunderson K, Murray SS, Oliphant AR, Chee MS, Montpetit A, Chagnon F, Ferretti V, Leboeuf M, Olivier J, Phillips MS, Roumy S, Sallee C, Verner A, Hudson TJ, Kwok P, Cai D, Koboldt DC, Miller RD, Pawlikowska L, Taillon-Miller P, Xiao M, Tsui L, Mak W, Song YQ, Tam PKH, Nakamura Y, Kawaguchi T, Kitamoto T, Morizono T, Nagashima A, Ohnishi Y, Sekine A, Tanaka T, Tsunoda T, Deloukas P, Bird CP, Delgado M, Dermitzakis ET, Gwilliam R, Hunt S, Morrison J, Powell D, Stranger BE, Whittaker P, Bentley DR, Daly MJ, de Bakker PIW, Barrett J, Chretien YR, Maller J, McCarroll S, Patterson N, Pe'er I, Price A, Purcell S, Richter DJ, Sabeti P, Saxena R, Schaffner SF, Sham PC, Varilly P, Altshuler D, Stein LD, Krishnan L, Smith AV, Tello-Ruiz MK, Thorisson GA, Chakravarti A, Chen PE, Cutler DJ, Kashuk CS, Lin S, Abecasis GR, Guan W, Li Y, Munro HM, Qin ZS, Thomas DJ, McVean G, Auton A, Bottolo L, Cardin N, Eyheramendy S, Freeman C, Marchini J, Myers S, Spencer C, Stephens M, Donnelly P, Cardon LR, Clarke G, Evans DM, Morris AP, Weir BS, Tsunoda T, Mullikin JC, Sherry ST, Feolo M, Skol A, Zhang H, Zeng C, Zhao H, Matsuda I, Fukushima Y, Macer DR, Suda E, Rotimi CN, Adebamowo CA, Ajayi I, Aniagwu T, Marshall PA, Nkwodimmah C, Royal CDM, Leppert MF, Dixon M, Peiffer A, Qiu R, Kent A, Kato K, Niikawa N, Adewole IF, Knoppers BM, Foster MW, Clayton EW, Watkin J, Gibbs RA, Belmont JW, Muzny D, Nazareth L, Sodergren E, Weinstock GM, Wheeler DA, Yakub I, Gabriel SB, Onofrio RC, Richter DJ, Ziaugra L, Birren BW, Daly MJ, Altshuler D, Wilson RK, Fulton LL, Rogers J, Burton J, Carter NP, Clee CM, Griffiths M, Jones MC, McLay K, Plumb RW, Ross MT, Sims SK, Willey DL, Chen Z, Han H, Kang L, Godbout M, Wallenburg JC, L'Archeveque P, Bellemare G, Saeki K, Wang H, An D, Fu H, Li Q, Wang Z, Wang R, Holden AL, Brooks LD, McEwen JE, Guyer MS, Wang VO, Peterson JL, Shi M, Spiegel J, Sung LM, Zacharia LF, Collins FS, Kennedy K, Jamieson R & Stewart J (2007) A second generation human haplotype map of over 3.1 million SNPs. Nature 449(7164): 851–861.

128
Ioannidis JP, Boffetta P, Little J, O'Brien TR, Uitterlinden AG, Vineis P, Balding DJ, Chokkalingam A, Dolan SM, Flanders WD, Higgins JP, McCarthy MI, McDermott DH, Page GP, Rebbeck TR, Seminara D & Khoury MJ (2008) Assessment of cumulative evidence on genetic associations: interim guidelines. Int J Epidemiol 37(1): 120–132.
Ioannidis JP, Ntzani EE, Trikalinos TA & Contopoulos-Ioannidis DG (2001) Replication validity of genetic association studies. Nat Genet 29(3): 306–309.
Ito S, Hattori T, Fukutake T & Sugimoto K (2003) Is atopic dermatitis a risk factor for intervertebral disc degeneration? A preliminary clinical and MRI study. J Neurol Sci 206(1): 39–42.
Jacobs LJ, Vo N & Kang JD (2011) Identifying inflammatory targets for biologic therapies for spine pain. PM R 3(6): 12–17.
Jakkula E, Rehnstrom K, Varilo T, Pietilainen OP, Paunio T, Pedersen NL, deFaire U, Jarvelin MR, Saharinen J, Freimer N, Ripatti S, Purcell S, Collins A, Daly MJ, Palotie A & Peltonen L (2008) The genome-wide patterns of variation expose significant substructure in a founder population. Am J Hum Genet 83(6): 787–794.
Jarvik JG, Hollingworth W, Heagerty PJ, Haynor DR, Boyko EJ & Deyo RA (2005) Three-year incidence of low back pain in an initially asymptomatic cohort: clinical and imaging risk factors. Spine (Phila Pa 1976) 30(13): 1541–1548.
Jensen MC, Brant-Zawadzki MN, Obuchowski N, Modic MT, Malkasian D & Ross JS (1994) Magnetic resonance imaging of the lumbar spine in people without back pain. N Engl J Med 331(2): 69–73.
Jim JJ, Noponen-Hietala N, Cheung KM, Ott J, Karppinen J, Sahraravand A, Luk KD, Yip SP, Sham PC, Song YQ, Leong JC, Cheah KS, Ala-Kokko L & Chan D (2005) The TRP2 allele of COL9A2 is an age-dependent risk factor for the development and severity of intervertebral disc degeneration. Spine (Phila Pa 1976) 30(24): 2735–2742.
Johannessen W, Auerbach JD, Wheaton AJ, Kurji A, Borthakur A, Reddy R & Elliott DM (2006) Assessment of human disc degeneration and proteoglycan content using T1rho-weighted magnetic resonance imaging. Spine (Phila Pa 1976) 31(11): 1253–1257.
Jones G, White C, Sambrook P & Eisman J (1998) Allelic variation in the vitamin D receptor, lifestyle factors and lumbar spinal degenerative disease. Ann Rheum Dis 57(2): 94–99.
Jordan KM, Syddall H, Dennison EM, Cooper C & Arden NK (2005) Birthweight, vitamin D receptor gene polymorphism, and risk of lumbar spine osteoarthritis. J Rheumatol 32(4): 678–683.
Juni P, Witschi A, Bloch R & Egger M (1999) The hazards of scoring the quality of clinical trials for meta-analysis. JAMA 282(11): 1054–1060.
Kalb S, Martirosyan NL, Kalani MYS, Broc GG & Theodore N (2012) Genetics of the Degenerated Intervertebral Disc. World Neurosurg 77(3-4):491–501.
Kales SN, Linos A, Chatzis C, Sai Y, Halla M, Nasioulas G & Christiani DC (2004) The role of collagen IX tryptophan polymorphisms in symptomatic intervertebral disc disease in Southern European patients. Spine (Phila Pa 1976) 29(11): 1266–1270.

129
Kalichman L, Guermazi A, Li L & Hunter DJ (2009) Association between age, sex, BMI and CT-evaluated spinal degeneration features J Back Musculoskelet Rehabil 22(4): 189–195.
Kalichman L & Hunter DJ (2008a) The genetics of intervertebral disc degeneration. Familial predisposition and heritability estimation Joint Bone Spine 75(4): 383–387.
Kalichman L & Hunter DJ (2008b) The genetics of intervertebral disc degeneration. Associated genes. Joint Bone Spine 75(4): 388–396.
Kalichman L & Hunter DJ (2007) Lumbar facet joint osteoarthritis: a review. Semin Arthritis Rheum 37:69–80
Kamarainen O, Solovieva S, Vehmas T, Luoma K, Riihimaki H, Ala-Kokko L, Mannikko M & Leino-Arjas P (2008) Common interleukin-6 promoter variants associate with the more severe forms of distal interphalangeal osteoarthritis. Arthritis Res Ther 10(1).
Kaminsky Z, Wang S & Petronis A (2006) Complex disease, gender and epigenetics. Ann Med 38(8): 530–544.
Kaneoka K, Shimizu K, Hangai M, Okuwaki T, Mamizuka N, Sakane M & Ochiai N (2007) Lumbar intervertebral disk degeneration in elite competitive swimmers: a case control study. Am J Sports Med 35(8): 1341–1345.
Karacan I, Aydin T, Sahin Z, Cidem M, Koyuncu H, Aktas I & Uludag M (2004) Facet angles in lumbar disc herniation: their relation to anthropometric features. Spine (Phila Pa 1976) 29(10): 1132–1136.
Karasik D & Ferrari SL (2008) Contribution of gender-specific genetic factors to osteoporosis risk. Ann Hum Genet 72: 696–714.
Karasugi T, Semba K, Hirose Y, Kelempisioti A, Nakajima M, Miyake A, Furuichi T, Kawaguchi Y, Mikami Y, Chiba K, Kamata M, Ozaki K, Takahashi A, Makela P, Karppinen J, Kimura T, Kubo T, Toyama Y, Yamamura K, Mannikko M, Mizuta H & Ikegawa S (2009) Association of the tag SNPs in the human SKT gene (KIAA1217) with lumbar disc herniation. J Bone Miner Res 24(9): 1537–1543.
Karppinen J, Daavittila I, Solovieva S, Kuisma M, Taimela S, Natri A, Haapea M, Korpelainen R, Niinimaki J, Tervonen O, Ala-Kokko L & Mannikko M (2008) Genetic factors are associated with modic changes in endplates of lumbar vertebral bodies. Spine (Phila Pa 1976) 33(11): 1236–1241.
Karppinen J, Paakko E, Raina S, Tervonen O, Kurunlahti M, Nieminen P, Ala-Kokko L, Malmivaara A & Vanharanta H (2002) Magnetic resonance imaging findings in relation to the COL9A2 tryptophan allele among patients with sciatica. Spine (Phila Pa 1976) 27(1): 78–83.
Karppinen J, Shen FH, Luk KDK, Andersson GBJ, Cheung KMC & Samartzis D (2011) Management of degenerative disk disease and chronic low back pain. Orthop Clin North Am 42(4): 513–528.
Karppinen J, Solovieva S, Luoma K, Raininko R, Leino-Arjas P & Riihimaki H (2009) Modic changes and interleukin 1 gene locus polymorphisms in occupational cohort of middle-aged men. Eur Spine J 18(12): 1963–1970.
Katz JN (2006) Lumbar disc disorders and low-back pain: socioeconomic factors and consequences. J Bone Joint Surg Am 88 Suppl 2: 21–24.

130
Kawaguchi Y, Osada R, Kanamori M, Ishihara H, Ohmori K, Matsui H & Kimura T (1999) Association between an aggrecan gene polymorphism and lumbar disc degeneration. Spine (Phila Pa 1976) 24(23): 2456–2460.
Kawaguchi Y, Kanamori M, Ishihara H, Ohmori K, Matsui H & Kimura T (2002) The association of lumbar disc disease with vitamin-D receptor gene polymorphism. J Bone Joint Surg Am 84-A(11): 2022–2028.
Kelempisioti A, Eskola PJ, Okuloff A, Karjalainen U, Takatalo J, Daavittila I, Niinimaki J, Sequeiros RB, Tervonen O, Solovieva S, Kao PY, Song Y, Cheung KM, Chan D, Ala-Kokko L, Jarvelin M, Karppinen J & Mannikko M (2011) Genetic susceptibility of intervertebral disc degeneration among young Finnish adults. BMC Med Genet 12(1): 153–153.
Kelsey JL & Hardy RJ (1975) Driving of motor vehicles as a risk factor for acute herniated lumbar intervertebral disc. Am J Epidemiol 102(1): 63–73.
Kepler CK, Anderson DG, Tannoury C & Ponnappan RK (2011) Intervertebral disk degeneration and emerging biologic treatments. J Am Acad Orthop Surg 19(9): 543–553.
Kere J (2010) Genetics of complex disorders. Biochem Biophys Res Commun 396(1): 143–146.
Khoury MJ, Little J, Gwinn M & Ioannidis JPA (2007) On the synthesis and interpretation of consistent but weak gene-disease associations in the era of genome-wide association studies. Int J Epidemiol 36(2): 439–445.
Kiezun A, Garimella K, Do R, Stitziel NO, Neale BM, McLaren PJ, Gupta N, Sklar P, Sullivan PF, Moran JL, Hultman CM, Lichtenstein P, Magnusson P, Lehner T, Shugart YY, Price AL, de Bakker PIW, Purcell SM & Sunyaev SR (2012) Exome sequencing and the genetic basis of complex traits. Nat Genet 44(6): 623–630.
Kim D, Lee S, Kim K & Yu S (2010) Association of interleukin-1 receptor antagonist gene polymorphism with response to conservative treatment of lumbar herniated nucleus pulposus. Spine (Phila Pa 1976) 35(16): 1527–1531.
Kim K, Kim Y, Ha K, Woo Y, Park J, Park W & An HS (2005) An autocrine or paracrine Fas-mediated counterattack: a potential mechanism for apoptosis of notochordal cells in intact rat nucleus pulposus. Spine (Phila Pa 1976) 30(11): 1247–1251.
Kim K, Lim T, Kim JG, Jeong S, Masuda K & An HS (2003) The origin of chondrocytes in the nucleus pulposus and histologic findings associated with the transition of a notochordal nucleus pulposus to a fibrocartilaginous nucleus pulposus in intact rabbit intervertebral discs. Spine (Phila Pa 1976) 28(10): 982–990.
Kim NK, Shin DA, Han IB, Yoo EH, Kim SH & Chung SS (2011) The association of aggrecan gene polymorphism with the risk of intervertebral disc degeneration. Acta Neurochir 153(1): 129–133.
Kizawa H, Kou I, Iida A, Sudo A, Miyamoto Y, Fukuda A, Mabuchi A, Kotani A, Kawakami A, Yamamoto S, Uchida A, Nakamura K, Notoya K, Nakamura Y & Ikegawa S (2005) An aspartic acid repeat polymorphism in asporin inhibits chondrogenesis and increases susceptibility to osteoarthritis. Nat Genet 37(2): 138–144.

131
Kjaer P, Leboeuf-Yde C, Sorensen JS & Bendix T (2005) An epidemiologic study of MRI and low back pain in 13-year-old children. Spine (Phila Pa 1976) 30(7): 798–806.
Kjaer P, Wedderkopp N, Korsholm L & Leboeuf-Yde C (2011) Prevalence and tracking of back pain from childhood to adolescence. BMC Musculoskelet Disord 12: 98–98.
Knuutinen A, Kokkonen N, Risteli J, Vahakangas K, Kallioinen M, Salo T, Sorsa T & Oikarinen A (2002) Smoking affects collagen synthesis and extracellular matrix turnover in human skin. Br J Dermatol 146(4): 588–594.
Koes BW, van Tulder MW & Thomas S (2006) Diagnosis and treatment of low back pain. BMJ 332(7555): 1430–1434.
Kornak U & Mundlos S (2003) Genetic disorders of the skeleton: a developmental approach. Am J Hum Genet 73(3): 447–474.
Koshizuka Y, Ogata N, Shiraki M, Hosoi T, Seichi A, Takeshita K, Nakamura K & Kawaguchi H (2006) Distinct association of gene polymorphisms of estrogen receptor and vitamin D receptor with lumbar spondylosis in post-menopausal women. Eur Spine J 15(10): 1521–1528.
Kumar R, Kumar V, Das NK, Behari S & Mahapatra AK (2007) Adolescent lumbar disc disease: findings and outcome. Childs Nerv Syst 23(11): 1295–1299.
Kwan T, Benovoy D, Dias C, Gurd S, Provencher C, Beaulieu P, Hudson TJ, Sladek R & Majewski J (2008) Genome-wide analysis of transcript isoform variation in humans. Nat Genet 40(2): 225–231.
Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J, Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, Sougnez C, Stange-Thomann N, Stojanovic N, Subramanian A, Wyman D, Rogers J, Sulston J, Ainscough R, Beck S, Bentley D, Burton J, Clee C, Carter N, Coulson A, Deadman R, Deloukas P, Dunham A, Dunham I, Durbin R, French L, Grafham D, Gregory S, Hubbard T, Humphray S, Hunt A, Jones M, Lloyd C, McMurray A, Matthews L, Mercer S, Milne S, Mullikin JC, Mungall A, Plumb R, Ross M, Shownkeen R, Sims S, Waterston RH, Wilson RK, Hillier LW, McPherson JD, Marra MA, Mardis ER, Fulton LA, Chinwalla AT, Pepin KH, Gish WR, Chissoe SL, Wendl MC, Delehaunty KD, Miner TL, Delehaunty A, Kramer JB, Cook LL, Fulton RS, Johnson DL, Minx PJ, Clifton SW, Hawkins T, Branscomb E, Predki P, Richardson P, Wenning S, Slezak T, Doggett N, Cheng JF, Olsen A, Lucas S, Elkin C, Uberbacher E, Frazier M, Gibbs RA, Muzny DM, Scherer SE, Bouck JB, Sodergren EJ, Worley KC, Rives CM, Gorrell JH, Metzker ML, Naylor SL, Kucherlapati RS, Nelson DL, Weinstock GM, Sakaki Y, Fujiyama A, Hattori M, Yada T, Toyoda A, Itoh T, Kawagoe C, Watanabe H, Totoki Y, Taylor T, Weissenbach J, Heilig R, Saurin W, Artiguenave F, Brottier P, Bruls T, Pelletier E, Robert C, Wincker P, Smith DR, Doucette-Stamm L, Rubenfield M, Weinstock K, Lee HM, Dubois J, Rosenthal A, Platzer M, Nyakatura G, Taudien S, Rump A, Yang H, Yu J, Wang J, Huang G, Gu J, Hood L, Rowen L, Madan A, Qin S, Davis RW, Federspiel NA, Abola AP, Proctor MJ, Myers RM, Schmutz J, Dickson M, Grimwood J, Cox DR, Olson MV, Kaul R, Raymond C, Shimizu N, Kawasaki K,

132
Minoshima S, Evans GA, Athanasiou M, Schultz R, Roe BA, Chen F, Pan H, Ramser J, Lehrach H, Reinhardt R, McCombie WR, de lB, Dedhia N, Blocker H, Hornischer K, Nordsiek G, Agarwala R, Aravind L, Bailey JA, Bateman A, Batzoglou S, Birney E, Bork P, Brown DG, Burge CB, Cerutti L, Chen HC, Church D, Clamp M, Copley RR, Doerks T, Eddy SR, Eichler EE, Furey TS, Galagan J, Gilbert JG, Harmon C, Hayashizaki Y, Haussler D, Hermjakob H, Hokamp K, Jang W, Johnson LS, Jones TA, Kasif S, Kaspryzk A, Kennedy S, Kent WJ, Kitts P, Koonin EV, Korf I, Kulp D, Lancet D, Lowe TM, McLysaght A, Mikkelsen T, Moran JV, Mulder N, Pollara VJ, Ponting CP, Schuler G, Schultz J, Slater G, Smit AF, Stupka E, Szustakowski J, Thierry-Mieg D, Thierry-Mieg J, Wagner L, Wallis J, Wheeler R, Williams A, Wolf YI, Wolfe KH, Yang SP, Yeh RF, Collins F, Guyer MS, Peterson J, Felsenfeld A, Wetterstrand KA, Patrinos A, Morgan MJ, de Jong P, Catanese JJ, Osoegawa K, Shizuya H, Choi S, Chen YJ, Szustakowki J & (2001) Initial sequencing and analysis of the human genome. Nature 409(6822): 860–921.
Le Maitre CL, Freemont AJ & Hoyland JA (2007) Accelerated cellular senescence in degenerate intervertebral discs: a possible role in the pathogenesis of intervertebral disc degeneration. Arthritis Res Ther 9(3).
Leboeuf-Yde C & Kyvik KO (1998) At what age does low back pain become a common problem? A study of 29,424 individuals aged 12–41 years. Spine (Phila Pa 1976) 23(2): 228–234.
Li P, Nijhawan D, Budihardjo I, Srinivasula SM, Ahmad M, Alnemri ES & Wang X (1997) Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 91(4): 479–489.
Li Y, Willer C, Sanna S & Abecasis G (2009) Genotype imputation. Annu Rev Genomics Hum Genet 10: 387–406.
Lin WP, Lin JH, Chen XW, Wu CY, Zhang LQ, Huang ZD & Lai JM (2011) Interleukin-10 promoter polymorphisms associated with susceptibility to lumbar disc degeneration in a Chinese cohort. Genet Mol Res 10(3): 1719–1727.
Little J, Higgins JPT, Ioannidis JPA, Moher D, Gagnon F, von Elm E, Khoury MJ, Cohen B, Davey-Smith G, Grimshaw J, Scheet P, Gwinn M, Williamson RE, Zou GY, Hutchings K, Johnson CY, Tait V, Wiens M, Golding J, van Duijn C, McLaughlin J, Paterson A, Wells G, Fortier I, Freedman M, Zecevic M, King R, Infante-Rivard C, Stewart A & Birkett N (2009) STrengthening the REporting of Genetic Association Studies (STREGA)--an extension of the STROBE statement. Genet Epidemiol 33(7): 581–598.
Liuke M, Solovieva S, Lamminen A, Luoma K, Leino-Arjas P, Luukkonen R & Riihimaki H (2005) Disc degeneration of the lumbar spine in relation to overweight. Int J Obes (Lond) 29(8): 903–908.
Livshits G, Cohen Z, Higla O & Yakovenko K (2001) Familial history, age and smoking are important risk factors for disc degeneration disease in Arabic pedigrees. Eur J Epidemiol 17(7): 643–651.

133
Livshits G, Ermakov S, Popham M, Macgregor AJ, Sambrook PN, Spector TD & Williams FM (2010) Evidence that bone mineral density plays a role in degenerative disc disease: the UK Twin Spine study. Ann Rheum Dis 69(12): 2102–2106.
Loeser RF (2006) Molecular mechanisms of cartilage destruction: mechanics, inflammatory mediators, and aging collide. Arthritis Rheum 54(5): 1357–1360.
Lohmueller KE, Pearce CL, Pike M, Lander ES & Hirschhorn JN (2003) Meta-analysis of genetic association studies supports a contribution of common variants to susceptibility to common disease. Nat Genet 33(2): 177–182.
Lorenzo P, Bayliss MT & Heinegaard D (1998) A novel cartilage protein (CILP) present in the mid-zone of human articular cartilage increases with age. J Biol Chem 273(36): 23463–23468.
Loughlin J (2011) Knee osteoarthritis, lumbar-disc degeneration and developmental dysplasia of the hip - an emerging genetic overlap. Arthritis Res Ther 13(2): 108–108.
Madelin G, Chang G, Otazo R, Jerschow A & Regatte RR (2012) Compressed sensing sodium MRI of cartilage at 7T: preliminary study. J Magn Reson 214(1): 360–365.
Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, Hunter DJ, McCarthy MI, Ramos EM, Cardon LR, Chakravarti A, Cho JH, Guttmacher AE, Kong A, Kruglyak L, Mardis E, Rotimi CN, Slatkin M, Valle D, Whittemore AS, Boehnke M, Clark AG, Eichler EE, Gibson G, Haines JL, Mackay TFC, McCarroll SA & Visscher PM (2009) Finding the missing heritability of complex diseases. Nature 461(7265): 747–753.
Marchand F & Ahmed AM (1990) Investigation of the laminate structure of lumbar disc anulus fibrosus. Spine (Phila Pa 1976) 15(5): 402–410.
Marchini J & Howie B (2010) Genotype imputation for genome-wide association studies. Nat Rev Genet 11(7): 499–511.
Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen YJ, Chen Z, Dewell SB, Du L, Fierro JM, Gomes XV, Godwin BC, He W, Helgesen S, Ho CH, Ho CH, Irzyk GP, Jando SC, Alenquer ML, Jarvie TP, Jirage KB, Kim JB, Knight JR, Lanza JR, Leamon JH, Lefkowitz SM, Lei M, Li J, Lohman KL, Lu H, Makhijani VB, McDade KE, McKenna MP, Myers EW, Nickerson E, Nobile JR, Plant R, Puc BP, Ronan MT, Roth GT, Sarkis GJ, Simons JF, Simpson JW, Srinivasan M, Tartaro KR, Tomasz A, Vogt KA, Volkmer GA, Wang SH, Wang Y, Weiner MP, Yu P, Begley RF & Rothberg JM (2005) Genome sequencing in microfabricated high-density picolitre reactors. Nature 437(7057): 376–380.
Marian AJ (2012) Molecular genetic studies of complex phenotypes. Transl Res 159(2):64–79
Martin MD, Boxell CM & Malone DG (2002) Pathophysiology of lumbar disc degeneration: a review of the literature. Neurosurg Focus 13(2).
Mashayekhi F, Shafiee G, Kazemi M & Dolati P (2010) Lumbar disk degeneration disease and aggrecan gene polymorphism in northern Iran. Biochem Genet 48(7–8): 684–689.
Masuda K & Lotz JC (2010) New challenges for intervertebral disc treatment using regenerative medicine. Tissue Eng Part B Rev 16(1): 147–158.

134
Matsui H, Kanamori M, Ishihara H, Yudoh K, Naruse Y & Tsuji H (1998) Familial predisposition for lumbar degenerative disc disease. A case-control study Spine (Phila Pa 1976) 23(9): 1029–1034.
Matsui H, Terahata N, Tsuji H, Hirano N & Naruse Y (1992) Familial predisposition and clustering for juvenile lumbar disc herniation Spine (Phila Pa 1976) 17(11): 1323–1328.
McCarthy MI, Abecasis GR, Cardon LR, Goldstein DB, Little J, Ioannidis JPA & Hirschhorn JN (2008) Genome-wide association studies for complex traits: consensus, uncertainty and challenges. Nat Rev Genet 9(5): 356–369.
Mechanic LE, Chen H, Amos CI, Chatterjee N, Cox NJ, Divi RL, Fan R, Harris EL, Jacobs K, Kraft P, Leal SM, McAllister K, Moore JH, Paltoo DN, Province MA, Ramos EM, Ritchie MD, Roeder K, Schaid DJ, Stephens M, Thomas DC, Weinberg CR, Witte JS, Zhang S, Zollner S, Feuer EJ & Gillanders EM (2011) Next generation analytic tools for large scale genetic epidemiology studies of complex diseases. Genet Epidemiol.
Meder B, Haas J, Keller A, Heid C, Just S, Borries A, Boisguerin V, Scharfenberger-Schmeer M, Stahler P, Beier M, Weichenhan D, Strom TM, Pfeufer A, Korn B, Katus HA & Rottbauer W (2011) Targeted next-generation sequencing for the molecular genetic diagnostics of cardiomyopathies. Circ Cardiovasc Genet 4(2): 110–122.
Michaelson JJ, Loguercio S & Beyer A (2009) Detection and interpretation of expression quantitative trait loci (eQTL). Methods 48(3): 265–276.
Milette PC (1997) The proper terminology for reporting lumbar intervertebral disk disorders. AJNR Am J Neuroradiol 18(10): 1859–1866.
Min SK, Nakazato K, Okada T, Ochi E & Hiranuma K (2009) The cartilage intermediate layer protein gene is associated with lumbar disc degeneration in collegiate judokas. Int J Sports Med 30(9): 691–694.
Min S, Nakazato K, Yamamoto Y, Gushiken K, Fujimoto H, Fujishiro H, Kobayakawa Y & Hiranuma K (2010) Cartilage intermediate layer protein gene is associated with lumbar disc degeneration in male, but not female, collegiate athletes. Am J Sports Med 38(12): 2552–2557.
Mio F, Chiba K, Hirose Y, Kawaguchi Y, Mikami Y, Oya T, Mori M, Kamata M, Matsumoto M, Ozaki K, Tanaka T, Takahashi A, Kubo T, Kimura T, Toyama Y & Ikegawa S (2007) A functional polymorphism in COL11A1, which encodes the alpha 1 chain of type XI collagen, is associated with susceptibility to lumbar disc herniation. Am J Hum Genet 81(6): 1271–1277.
Mirza SK & White AA (1995) Anatomy of intervertebral disc and pathophysiology of herniated disc disease. J Clin Laser Med Surg 13(3): 131–142.
Modic MT & Ross JS (2007) Lumbar degenerative disk disease. Radiology 245(1): 43–61. Modic MT, Steinberg PM, Ross JS, Masaryk TJ & Carter JR (1988) Degenerative disk
disease: assessment of changes in vertebral body marrow with MR imaging. Radiology 166(1): 193–199.

135
Moller A, Maly P, Besjakov J, Hasserius R, Ohlin A & Karlsson MK (2007) A vertebral fracture in childhood is not a risk factor for disc degeneration but for Schmorl's nodes: a mean 40-year observational study. Spine (Phila Pa 1976) 32(22): 2487–2492.
Montgomery SB & Dermitzakis ET (2011) From expression QTLs to personalized transcriptomics. Nat Rev Genet 12(4): 277–282.
Moore KL (1999) Clinically Oriented Anatomy: Fourth Edition. Baltimore, Lippincott Williams & Wilkins.
Moore RJ, Vernon-Roberts B, Osti OL & Fraser RD (1996) Remodeling of vertebral bone after outer anular injury in sheep. Spine (Phila Pa 1976) 21(8): 936–940.
Munafo MR (2010) Credible genetic associations? Int J Mol Epidemiol Genet 1(1): 31–34. Nachemson AL (1985) Advances in low-back pain. Clin Orthop Relat Res (200): 266–278. Nakki A, Videman T, Kujala UM, Suhonen M, Mannikko M, Peltonen L, Battie MC,
Kaprio J & Saarela J (2011) Candidate gene association study of magnetic resonance imaging-based hip osteoarthritis (OA): evidence for COL9A2 gene as a common predisposing factor for hip OA and lumbar disc degeneration. J Rheumatol 38(4): 747–752.
Natarajan RN & Andersson GB (1999) The influence of lumbar disc height and cross-sectional area on the mechanical response of the disc to physiologic loading. Spine (Phila Pa 1976) 24(18): 1873–1881.
Nelis M, Esko T, Magi R, Zimprich F, Zimprich A, Toncheva D, Karachanak S, Piskackova T, Balascak I, Peltonen L, Jakkula E, Rehnstrom K, Lathrop M, Heath S, Galan P, Schreiber S, Meitinger T, Pfeufer A, Wichmann H, Melegh B, Polgar N, Toniolo D, Gasparini P, D'Adamo P, Klovins J, Nikitina-Zake L, Kucinskas V, Kasnauskiene J, Lubinski J, Debniak T, Limborska S, Khrunin A, Estivill X, Rabionet R, Marsal S, Julià A, Antonarakis SE, Deutsch S, Borel C, Attar H, Gagnebin M, Macek M, Krawczak M, Remm M & Metspalu A (2009) Genetic structure of Europeans: a view from the North-East. PLoS One 4(5).
Nerlich AG, Boos N, Wiest I & Aebi M (1998) Immunolocalization of major interstitial collagen types in human lumbar intervertebral discs of various ages. Virchows Arch 432(1): 67–76.
Newton-Cheh C, Johnson T, Gateva V, Tobin MD, Bochud M, Coin L, Najjar SS, Zhao JH, Heath SC, Eyheramendy S, Papadakis K, Voight BF, Scott LJ, Zhang F, Farrall M, Tanaka T, Wallace C, Chambers JC, Khaw KT, Nilsson P, van der Harst P, Polidoro S, Grobbee DE, Onland-Moret NC, Bots ML, Wain LV, Elliott KS, Teumer A, Luan J, Lucas G, Kuusisto J, Burton PR, Hadley D, McArdle WL, Wellcome Trust Case Control Consortium, Brown M, Dominiczak A, Newhouse SJ, Samani NJ, Webster J, Zeggini E, Beckmann JS, Bergmann S, Lim N, Song K, Vollenweider P, Waeber G, Waterworth DM, Yuan X, Groop L, Orho-Melander M, Allione A, Di Gregorio A, Guarrera S, Panico S, Ricceri F, Romanazzi V, Sacerdote C, Vineis P, Barroso I, Sandhu MS, Luben RN, Crawford GJ, Jousilahti P, Perola M, Boehnke M, Bonnycastle LL, Collins FS, Jackson AU, Mohlke KL, Stringham HM, Valle TT, Willer CJ, Bergman RN, Morken MA, Doring A, Gieger C, Illig T, Meitinger T, Org E, Pfeufer A, Wichmann HE, Kathiresan S, Marrugat J, O'Donnell CJ, Schwartz SM,

136
Siscovick DS, Subirana I, Freimer NB, Hartikainen AL, McCarthy MI, O'Reilly PF, Peltonen L, Pouta A, de Jong PE, Snieder H, van Gilst WH, Clarke R, Goel A, Hamsten A, Peden JF, Seedorf U, Syvänen AC, Tognoni G, Lakatta EG, Sanna S, Scheet P, Schlessinger D, Scuteri A, Dorr M, Ernst F, Felix SB, Homuth G, Lorbeer R, Reffelmann T, Rettig R, Volker U, Galan P, Gut IG, Hercberg S, Lathrop GM, Zelenika D, Deloukas P, Soranzo N, Williams FM, Zhai G, Salomaa V, Laakso M, Elosua R, Forouhi NG, Volzke H, Uiterwaal CS, van der Schouw YT, Numans ME, Matullo G, Navis G, Berglund G, Bingham SA, Kooner JS, Connell JM, Bandinelli S, Ferrucci L, Watkins H, Spector TD, Tuomilehto J, Altshuler D, Strachan DP, Laan M, Meneton P, Wareham NJ, Uda M, Jarvelin MR, Mooser V, Melander O, Loos RJ, Elliott P, Abecasis GR, Caulfield M & Munroe PB (2009) Genome-wide association study identifies eight loci associated with blood pressure. Nat Genet 41(6): 666–676.
Niedringhaus TP, Milanova D, Kerby MB, Snyder MP & Barron AE (2011) Landscape of next-generation sequencing technologies. Anal Chem 83(12): 4327–4341.
Noponen-Hietala N, Kyllonen E, Mannikko M, Ilkko E, Karppinen J, Ott J & Ala-Kokko L (2003) Sequence variations in the collagen IX and XI genes are associated with degenerative lumbar spinal stenosis. Ann Rheum Dis 62(12): 1208–1214.
Noponen-Hietala N, Virtanen I, Karttunen R, Schwenke S, Jakkula E, Li H, Merikivi R, Barral S, Ott J, Karppinen J & Ala-Kokko L (2005) Genetic variations in IL6 associate with intervertebral disc disease characterized by sciatica. Pain 114(1–2): 186–194.
Norio R (2003a) Finnish Disease Heritage I: characteristics, causes, background. Hum Genet 112(5–6): 441–456.
Norio R (2003b) Finnish Disease Heritage II: population prehistory and genetic roots of Finns. Hum Genet 112(5–6): 457–469.
Norio R (2003c) The Finnish Disease Heritage III: the individual diseases. Hum Genet 112(5–6): 470–526.
Norio R, Perheentupa J, Nevanlinna HR (1973) Hereditary diseases in Finland; rare flora in rare soil. Ann Clin Res 5:109–141
Nunes F, Conforti-Froes N, Negrelli W, Souza D (2007) Genetic and environmental factors involved on intervertebral disc degeneration. Acta Ortopedica Brasileira 15(1): 9–13.
Ochagavia ME, Miranda J, Nazabal M, Martin A, Novoa LI, Bringas R, Fernandez-DE-Cossio J & Camacho H (2011) A methodology based on molecular interactions and pathways to find candidate genes associated to diseases: its application to schizophrenia and Alzheimer's disease. J Bioinform Comput Biol 9(4): 541–557.
Oishi Y, Shimizu K, Katoh T, Nakao H, Yamaura M, Furuko T, Narusawa K & Nakamura T (2003) Lack of association between lumbar disc degeneration and osteophyte formation in elderly japanese women with back pain. Bone 32(4): 405–411.
Olivero WC, Wang H, Hanigan WC, Henderson JP, Tracy PT, Elwood PW, Lister JR & Lyle L (2009) Cauda equina syndrome (CES) from lumbar disc herniations. J Spinal Disord Tech 22(3): 202–206.

137
Omoigui S (2007) The Interleukin-6 inflammation pathway from cholesterol to aging--role of statins, bisphosphonates and plant polyphenols in aging and age-related diseases. Immun Ageing 4: 1–1.
Ordovas JM (2007) Gender, a significant factor in the cross talk between genes, environment, and health. Gend Med 4 Suppl B: 111–122.
Osti OL, Vernon-Roberts B & Fraser RD (1990) 1990 Volvo Award in experimental studies. Anulus tears and intervertebral disc degeneration. An experimental study using an animal model. Spine (Phila Pa 1976) 15(8): 762–767.
Ott J (2004) Association of genetic loci: Replication or not, that is the question. Neurology 63(6): 955–958.
Ott J, Kamatani Y & Lathrop M (2011) Family-based designs for genome-wide association studies. Nat Rev Genet 12(7): 465–474.
Paajanen H, Erkintalo M, Kuusela T, Dahlstrom S & Kormano M (1989) Magnetic resonance study of disc degeneration in young low-back pain patients. Spine (Phila Pa 1976) 14(9): 982–985.
Paajanen H, Erkintalo M, Parkkola R, Salminen J & Kormano M (1997) Age-dependent correlation of low-back pain and lumbar disc regeneration. Arch Orthop Trauma Surg 116(1–2): 106–107.
Paalanne N (2011) Postural balance, isometric trunk muscle strength and low back symptoms among young adults. Acta Univ Oul D 1109.
Paassilta P, Lohiniva J, Goring HH, Perala M, Raina SS, Karppinen J, Hakala M, Palm T, Kroger H, Kaitila I, Vanharanta H, Ott J & Ala-Kokko L (2001) Identification of a novel common genetic risk factor for lumbar disk disease. JAMA 285(14): 1843–1849.
Papagelopoulos PJ, Shaughnessy WJ, Ebersold MJ, Bianco AJ & Quast LM (1998) Long-term outcome of lumbar discectomy in children and adolescents sixteen years of age or younger. J Bone Joint Surg Am 80(5): 689–698.
Parisini P, Di Silvestre M, Greggi T, Miglietta A & Paderni S (2001) Lumbar disc excision in children and adolescents. Spine (Phila Pa 1976) 26(18): 1997–2000.
Parkes M, Barrett JC, Prescott NJ, Tremelling M, Anderson CA, Fisher SA, Roberts RG, Nimmo ER, Cummings FR, Soars D, Drummond H, Lees CW, Khawaja SA, Bagnall R, Burke DA, Todhunter CE, Ahmad T, Onnie CM, McArdle W, Strachan D, Bethel G, Bryan C, Lewis CM, Deloukas P, Forbes A, Sanderson J, Jewell DP, Satsangi J, Mansfield JC, , Cardon L & Mathew CG (2007) Sequence variants in the autophagy gene IRGM and multiple other replicating loci contribute to Crohn's disease susceptibility. Nat Genet 39(7): 830–832.
Patsopoulos NA, Tatsioni A & Ioannidis JPA (2007) Claims of sex differences: an empirical assessment in genetic associations. JAMA 298(8): 880–893.
Pattin KA & Moore JH (2009) Role for protein-protein interaction databases in human genetics. Expert Rev Proteomics 6(6): 647–659.

138
Paz Aparicio J, López-Anglada Fernández E, Montes Prieto H, Pena Vázquez J, Fernández Bances I, Folgueras Henriksen V, López Fernández P, López-Fanjúl Menéndez JC, Valle Garay E & Asensi Álvarez V (2010) Association between the genetic polymorphism of interleukin-1β (3953 T/C) and symptomatic lumbar herniated disc. Revista Espanola de Cirugia Ortopedica y Traumatologia 54(4): 227–233.
Pelak K, Shianna KV, Ge D, Maia JM, Zhu M, Smith JP, Cirulli ET, Fellay J, Dickson SP, Gumbs CE, Heinzen EL, Need AC, Ruzzo EK, Singh A, Campbell CR, Hong LK, Lornsen KA, McKenzie AM, Sobreira NL, Hoover-Fong JE, Milner JD, Ottman R, Haynes BF, Goedert JJ & Goldstein DB (2010) The characterization of twenty sequenced human genomes. PLoS Genet 6(9).
Peltonen L (1997) Molecular background of the Finnish disease heritage. Ann Med 29(6): 553–556.
Peltonen L, Perola M, Naukkarinen J & Palotie A (2006) Lessons from studying monogenic disease for common disease. Hum Mol Genet 15 Spec No 1: R67–74.
Perheentupa J (1972) Suomalainen tautiperinto (Symposium on inherited disease in Finland). Duodecim 88:1–166
Pfirrmann CW, Metzdorf A, Zanetti M, Hodler J & Boos N (2001) Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine (Phila Pa 1976) 26(17): 1873–1878.
Pokharna HK & Phillips FM (1998) Collagen crosslinks in human lumbar intervertebral disc aging. Spine (Phila Pa 1976) 23(15): 1645–1648.
Pollintine P, Przybyla AS, Dolan P & Adams MA (2004) Neural arch load-bearing in old and degenerated spines. J Biomech 37(2): 197–204.
Prescher A (1998) Anatomy and pathology of the aging spine. Eur J Radiol 27(3): 181–195. Pritzker KP (1977) Aging and degeneration in the lumbar intervertebral disc. Orthop Clin
North Am 8(1): 66–77. Ragoussis J (2009) Genotyping technologies for genetic research. Annu Rev Genomics
Hum Genet 10: 117–133. Raj PP (2008) Intervertebral disc: anatomy-physiology-pathophysiology-treatment. Pain
Pract 8(1): 18–44. Rajasekaran S, Venkatadass K, Naresh Babu J, Ganesh K & Shetty AP (2008)
Pharmacological enhancement of disc diffusion and differentiation of healthy, ageing and degenerated discs : Results from in-vivo serial post-contrast MRI studies in 365 human lumbar discs. Eur Spine J 17(5): 626–643.
Rapan S, Gulan G, Lovric I & Jovanovic S (2011) Spontaneous regression of intervertebral disc herniation-case reports. Coll Antropol 35(1): 211–215.
Regatte RR & Schweitzer ME (2007) Ultra-high-field MRI of the musculoskeletal system at 7.0T. J Magn Reson Imaging 25(2): 262–269.
Rehman AU, Morell RJ, Belyantseva IA, Khan SY, Boger ET, Shahzad M, Ahmed ZM, Riazuddin S, Khan SN, Riazuddin S & Friedman TB (2010) Targeted capture and next-generation sequencing identifies C9orf75, encoding taperin, as the mutated gene in nonsyndromic deafness DFNB79. Am J Hum Genet 86(3): 378–388.

139
Repanti M, Korovessis PG, Stamatakis MV, Spastris P & Kosti P (1998) Evolution of disc degeneration in lumbar spine: a comparative histological study between herniated and postmortem retrieved disc specimens. J Spinal Disord 11(1): 41–45.
Richardson JK, Chung T, Schultz JS & Hurvitz E (1997) A familial predisposition toward lumbar disc injury. Spine (Phila Pa 1976) 22(13): 1487–1492.
Risbud MV & Shapiro IM (2011) Notochordal cells in the adult intervertebral disc: new perspective on an old question. Crit Rev Eukaryot Gene Expr 21(1): 29–41.
Rivera-Chavez FA, Peters-Hybki DL, Barber RC & O'Keefe GE (2003) Interleukin-6 promoter haplotypes and interleukin-6 cytokine responses. Shock 20(3): 218–223.
Rivinoja AE, Paananen MV, Taimela SP, Solovieva S, Okuloff A, Zitting P, Jarvelin M, Leino-Arjas P & Karppinen JI (2011) Sports, smoking, and overweight during adolescence as predictors of sciatica in adulthood: a 28-year follow-up study of a birth cohort. Am J Epidemiol 173(8): 890–897.
Roach JC, Glusman G, Smit AF, Huff CD, Hubley R, Shannon PT, Rowen L, Pant KP, Goodman N, Bamshad M, Shendure J, Drmanac R, Jorde LB, Hood L & Galas DJ (2010) Analysis of genetic inheritance in a family quartet by whole-genome sequencing. Science 328(5978): 636–639.
Roberts S, Menage J & Urban JP (1989) Biochemical and structural properties of the cartilage end-plate and its relation to the intervertebral disc. Spine (Phila Pa 1976) 14(2): 166–174.
Roberts S, Evans H, Trivedi J & Menage J (2006) Histology and pathology of the human intervertebral disc. J Bone Joint Surg Am 88 Suppl 2: 10–14.
Roughley P, Martens D, Rantakokko J, Alini M, Mwale F & Antoniou J (2006) The involvement of aggrecan polymorphism in degeneration of human intervertebral disc and articular cartilage. Eur Cell Mater 11: 1–7.
Roughley PJ (2004) Biology of intervertebral disc aging and degeneration: involvement of the extracellular matrix. Spine (Phila Pa 1976) 29(23): 2691–2699.
Ryder JJ, Garrison K, Song F, Hooper L, Skinner J, Loke Y, Loughlin J, Higgins JPT & MacGregor AJ (2008) Genetic associations in peripheral joint osteoarthritis and spinal degenerative disease: a systematic review. Ann Rheum Dis 67(5): 584–591.
Ryu S & Kim IS (2010) Spontaneous regression of a large lumbar disc extrusion. J Korean Neurosurg Soc 48(3): 285–287.
Saftic R, Grgic M, Ebling B & Splavski B (2006) Case-control study of risk factors for lumbar intervertebral disc herniation in Croatian island populations. Croat Med J 47(4): 593–600.
Sagoo GS, Little J & Higgins JP (2009) Systematic reviews of genetic association studies. Human Genome Epidemiology Network. PLoS Med 6(3): e28.
Salmela E, Lappalainen T, Fransson I, Andersen PM, Dahlman-Wright K, Fiebig A, Sistonen P, Savontaus M, Schreiber S, Kere J & Lahermo P (2008) Genome-wide analysis of single nucleotide polymorphisms uncovers population structure in Northern Europe. PLoS One 3(10).

140
Salmela E, Lappalainen T, Liu J, Sistonen P, Andersen PM, Schreiber S, Savontaus M, Czene K, Lahermo P, Hall P & Kere J (2011) Swedish population substructure revealed by genome-wide single nucleotide polymorphism data. PLoS One 6(2).
Salminen JJ, Erkintalo MO, Pentti J, Oksanen A & Kormano MJ (1999) Recurrent low back pain and early disc degeneration in the young. Spine (Phila Pa 1976) 24(13): 1316–1321.
Samartzis D, Karppinen J, Mok F, Fong DYT, Luk KDK & Cheung KMC (2011) A population-based study of juvenile disc degeneration and its association with overweight and obesity, low back pain, and diminished functional status. J Bone Joint Surg Am 93(7): 662–670.
Sambrook PN, MacGregor AJ & Spector TD (1999) Genetic influences on cervical and lumbar disc degeneration: a magnetic resonance imaging study in twins. Arthritis Rheum 42(2): 366–372.
Sarantaus L, Huusko P, Eerola H, Launonen V, Vehmanen P, Rapakko K, Gillanders E, Syrjakoski K, Kainu T, Vahteristo P, Krahe R, Paakkonen K, Hartikainen J, Blomqvist C, Lopponen T, Holli K, Ryynanen M, Butzow R, Borg A, Wasteson Arver B, Holmberg E, Mannermaa A, Kere J, Kallioniemi OP, Winqvist R & Nevanlinna H (2000) Multiple founder effects and geographical clustering of BRCA1 and BRCA2 families in Finland. Eur J Hum Genet 8(10): 757–763.
Scapinelli R (1993) Lumbar disc herniation in eight siblings with a positive family history for disc disease. Acta Orthop Belg 59(4): 371–376.
Schork NJ (1997) Genetics of complex disease: approaches, problems, and solutions. Am J Respir Crit Care Med 156(4 Pt 2): 103–109.
Schmidt H, Kettler A, Heuer F, Simon U, Claes L & Wilke H (2007) Intradiscal pressure, shear strain, and fiber strain in the intervertebral disc under combined loading. Spine (Phila Pa 1976) 32(7): 748–755.
Schneiderman G, Flannigan B, Kingston S, Thomas J, Dillin WH & Watkins RG (1987) Magnetic resonance imaging in the diagnosis of disc degeneration: correlation with discography. Spine (Phila Pa 1976) 12(3): 276–281.
Schoenfeld AJ, Nelson JH, Burks R & Belmont PJ (2011) Incidence and risk factors for lumbar degenerative disc disease in the United States military 1999–2008. Mil Med 176(11): 1320–1324.
Schollmeier G, Lahr-Eigen R & Lewandrowski KU (2000) Observations on fiber-forming collagens in the anulus fibrosus. Spine (Phila Pa 1976) 25(21): 2736–2741.
Schollum ML, Robertson PA & Broom ND (2009) A microstructural investigation of intervertebral disc lamellar connectivity: detailed analysis of the translamellar bridges. J Anat 214(6): 805–816.
Seki S, Kawaguchi Y, Chiba K, Mikami Y, Kizawa H, Oya T, Mio F, Mori M, Miyamoto Y, Masuda I, Tsunoda T, Kamata M, Kubo T, Toyama Y, Kimura T, Nakamura Y & Ikegawa S (2005) A functional SNP in CILP, encoding cartilage intermediate layer protein, is associated with susceptibility to lumbar disc disease. Nat Genet 37(6): 607–612.

141
Seki S, Kawaguchi Y, Mori M, Mio F, Chiba K, Mikami Y, Tsunoda T, Kubo T, Toyama Y, Kimura T & Ikegawa S (2006) Association study of COL9A2 with lumbar disc disease in the Japanese population. J Hum Genet 51(12): 1063–1067.
Semba K, Araki K, Li Z, Matsumoto K, Suzuki M, Nakagata N, Takagi K, Takeya M, Yoshinobu K, Araki M, Imai K, Abe K & Yamamura K (2006) A novel murine gene, Sickle tail, linked to the Danforth's short tail locus, is required for normal development of the intervertebral disc. Genetics 172(1): 445–456.
Sethumadhavan R, Doss CGP & Rajasekaran R (2011) In silico searching for disease-associated functional DNA variants. Methods Mol Biol 760: 239–250.
Shao Z, Rompe G & Schiltenwolf M (2002) Radiographic changes in the lumbar intervertebral discs and lumbar vertebrae with age. Spine (Phila Pa 1976) 27(3): 263–268.
Shinmei M, Masuda K, Kikuchi T & Shimomura Y (1989) Interleukin 1, tumor necrosis factor, and interleukin 6 as mediators of cartilage destruction. Semin Arthritis Rheum 18(3): 27–32.
Shiri R, Karppinen J, Leino-Arjas P, Solovieva S & Viikari-Juntura E (2010a) The association between obesity and low back pain: a meta-analysis. Am J Epidemiol 171(2): 135–154.
Shiri R, Karppinen J, Leino-Arjas P, Solovieva S & Viikari-Juntura E (2010b) The association between smoking and low back pain: a meta-analysis. Am J Med 123(1): 7–35.
Simmons ED, Guntupalli M, Kowalski JM, Braun F & Seidel T (1996) Familial predisposition for degenerative disc disease. A case-control study. Spine (Phila Pa 1976) 21(13): 1527–1529.
Sims AM, Timms AE, Bruges-Armas J, Burgos-Vargas R, Chou CT, Doan T, Dowling A, Fialho RN, Gergely P, Gladman DD, Inman R, Kauppi M, Kaarela K, Laiho K, Maksymowych W, Pointon JJ, Rahman P, Reveille JD, Sorrentino R, Tuomilehto J, Vargas-Alarcon G, Wordsworth BP, Xu H, Brown MA (2008) International Genetics of Ankylosing Spondylitis: Prospective meta-analysis of interleukin 1 gene complex polymorphisms confirms associations with ankylosing spondylitis. Ann Rheum Dis: 67(9):1305–1309.
Sladek R, Rocheleau G, Rung J, Dina C, Shen L, Serre D, Boutin P, Vincent D, Belisle A, Hadjadj S, Balkau B, Heude B, Charpentier G, Hudson TJ, Montpetit A, Pshezhetsky AV, Prentki M, Posner BI, Balding DJ, Meyre D, Polychronakos C & Froguel P (2007) A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature 445(7130): 881–885.
Smith AJP & Humphries SE (2009) Cytokine and cytokine receptor gene polymorphisms and their functionality. Cytokine Growth Factor Rev 20(1): 43–59.
Smith LJ, Nerurkar NL, Choi K, Harfe BD & Elliott DM (2011) Degeneration and regeneration of the intervertebral disc: lessons from development. Dis Model Mech 4(1): 31–41.

142
Solovieva S, Kouhia S, Leino-Arjas P, Ala-Kokko L, Luoma K, Raininko R, Saarela J & Riihim\aki H (2004) Interleukin 1 polymorphisms and intervertebral disc degeneration. Epidemiology 15(5): 626–633.
Solovieva S, Lohiniva J, Leino-Arjas P, Raininko R, Luoma K, Ala-Kokko L & Riihimaki H (2002) COL9A3 gene polymorphism and obesity in intervertebral disc degeneration of the lumbar spine: evidence of gene-environment interaction. Spine (Phila Pa 1976) 27(23): 2691–2696.
Solovieva S, Lohiniva J, Leino-Arjas P, Raininko R, Luoma K, Ala-Kokko L & Riihimaki H (2006) Intervertebral disc degeneration in relation to the COL9A3 and the IL-1ss gene polymorphisms. Eur Spine J 15(5): 613–619.
Solovieva S, Noponen N, Mannikko M, Leino-Arjas P, Luoma K, Raininko R, Ala-Kokko L & Riihimaki H (2007) Association between the aggrecan gene variable number of tandem repeats polymorphism and intervertebral disc degeneration. Spine (Phila Pa 1976) 32(16): 1700–1705.
Song YQ, Sham P, Cheung KMC, Chan D (2008) Genetics of disc degeneration. Current Orthopaedics 22(4): 259–266.
Song Y, Cheung KMC, Ho DWH, Poon SCS, Chiba K, Kawaguchi Y, Hirose Y, Alini M, Grad S, Yee AFY, Leong JCY, Luk KDK, Yip S, Karppinen J, Cheah KSE, Sham P, Ikegawa S & Chan D (2008a) Association of the asporin D14 allele with lumbar-disc degeneration in Asians. Am J Hum Genet 82(3): 744–747.
Song Y, Ho DWH, Karppinen J, Kao PYP, Fan B, Luk KDK, Yip S, Leong JCY, Cheah KSE, Sham P, Chan D & Cheung KMC (2008b) Association between promoter -1607 polymorphism of MMP1 and lumbar disc disease in Southern Chinese. BMC Med Genet 9: 38–38.
Sotoodehnia N, Isaacs A, de Bakker PI, Dorr M, Newton-Cheh C, Nolte IM, van der Harst P, Muller M, Eijgelsheim M, Alonso A, Hicks AA, Padmanabhan S, Hayward C, Smith AV, Polasek O, Giovannone S, Fu J, Magnani JW, Marciante KD, Pfeufer A, Gharib SA, Teumer A, Li M, Bis JC, Rivadeneira F, Aspelund T, Kottgen A, Johnson T, Rice K, Sie MP, Wang YA, Klopp N, Fuchsberger C, Wild SH, Mateo Leach I, Estrada K, Volker U, Wright AF, Asselbergs FW, Qu J, Chakravarti A, Sinner MF, Kors JA, Petersmann A, Harris TB, Soliman EZ, Munroe PB, Psaty BM, Oostra BA, Cupples LA, Perz S, de Boer RA, Uitterlinden AG, Volzke H, Spector TD, Liu FY, Boerwinkle E, Dominiczak AF, Rotter JI, van Herpen G, Levy D, Wichmann HE, van Gilst WH, Witteman JC, Kroemer HK, Kao WH, Heckbert SR, Meitinger T, Hofman A, Campbell H, Folsom AR, van Veldhuisen DJ, Schwienbacher C, O'Donnell CJ, Volpato CB, Caulfield MJ, Connell JM, Launer L, Lu X, Franke L, Fehrmann RS, te Meerman G, Groen HJ, Weersma RK, van den Berg LH, Wijmenga C, Ophoff RA, Navis G, Rudan I, Snieder H, Wilson JF, Pramstaller PP, Siscovick DS, Wang TJ, Gudnason V, van Duijn CM, Felix SB, Fishman GI, Jamshidi Y, Stricker BH, Samani NJ, Kaab S & Arking DE (2010) Common variants in 22 loci are associated with QRS duration and cardiac ventricular conduction. Nat Genet 42(12): 1068–1076.
Sullivan PF (2007) Spurious genetic associations. Biol Psychiatry 61(10): 1121–1126.

143
Sun ZM, Miao L, Zhang YG & Ming L (2009) Association between the -1562 C/T polymorphism of matrix metalloproteinase-9 gene and lumbar disc disease in the young adult population in North China. Connect Tissue Res 50(3): 181–185.
Sun Z, Ling M, Huo Y, Chang Y, Li Y, Qin H, Yang G & Lucas R (2011) Caspase 9 gene polymorphism and susceptibility to lumbar disc disease in the Han population in northern China. Connect Tissue Res 52(3): 198–202.
Takahashi M, Haro H, Wakabayashi Y, Kawa-uchi T, Komori H & Shinomiya K (2001) The association of degeneration of the intervertebral disc with 5a/6a polymorphism in the promoter of the human matrix metalloproteinase-3 gene. J Bone Joint Surg Br 83(4): 491–495.
Takatalo J, Karppinen J, Niinimaki J, Taimela S, Nayha S, Mutanen P, Sequeiros RB, Kyllonen E & Tervonen O (2011) Does lumbar disc degeneration on MRI associate with low back symptom severity in young Finnish adults? Spine (Phila Pa 1976) .
Takatalo J, Karppinen J, Niinimaki J, Taimela S, Nayha S, Jarvelin M, Kyllonen E & Tervonen O (2009) Prevalence of degenerative imaging findings in lumbar magnetic resonance imaging among young adults. Spine (Phila Pa 1976) 34(16): 1716–1721.
Taylor JR, Scott JE, Cribb AM & Bosworth TR (1992) Human intervertebral disc acid glycosaminoglycans. J Anat 180 ( Pt 1): 137–141.
Terry CF, Loukaci V & Green FR (2000) Cooperative influence of genetic polymorphisms on interleukin 6 transcriptional regulation. J Biol Chem 275(24): 18138–18144.
Tertti M, Paajanen H, Laato M, Aho H, Komu M & Kormano M (1991a) Disc degeneration in magnetic resonance imaging. A comparative biochemical, histologic, and radiologic study in cadaver spines. Spine (Phila Pa 1976) 16(6): 629–634.
Tertti MO, Salminen JJ, Paajanen HE, Terho PH & Kormano MJ (1991b) Low-back pain and disk degeneration in children: a case-control MR imaging study. Radiology 180(2): 503–507.
Teslovich TM, Musunuru K, Smith AV, Edmondson AC, Stylianou IM, Koseki M, Pirruccello JP, Ripatti S, Chasman DI, Willer CJ, Johansen CT, Fouchier SW, Isaacs A, Peloso GM, Barbalic M, Ricketts SL, Bis JC, Aulchenko YS, Thorleifsson G, Feitosa MF, Chambers J, Orho-Melander M, Melander O, Johnson T, Li X, Guo X, Li M, Shin Cho Y, Jin Go M, Jin Kim Y, Lee JY, Park T, Kim K, Sim X, Twee-Hee Ong R, Croteau-Chonka DC, Lange LA, Smith JD, Song K, Hua Zhao J, Yuan X, Luan J, Lamina C, Ziegler A, Zhang W, Zee RY, Wright AF, Witteman JC, Wilson JF, Willemsen G, Wichmann HE, Whitfield JB, Waterworth DM, Wareham NJ, Waeber G, Vollenweider P, Voight BF, Vitart V, Uitterlinden AG, Uda M, Tuomilehto J, Thompson JR, Tanaka T, Surakka I, Stringham HM, Spector TD, Soranzo N, Smit JH, Sinisalo J, Silander K, Sijbrands EJ, Scuteri A, Scott J, Schlessinger D, Sanna S, Salomaa V, Saharinen J, Sabatti C, Ruokonen A, Rudan I, Rose LM, Roberts R, Rieder M, Psaty BM, Pramstaller PP, Pichler I, Perola M, Penninx BW, Pedersen NL, Pattaro C, Parker AN, Pare G, Oostra BA, O'Donnell CJ, Nieminen MS, Nickerson DA, Montgomery GW, Meitinger T, McPherson R, McCarthy MI, McArdle W, Masson D, Martin NG, Marroni F, Mangino M, Magnusson PK, Lucas G, Luben R, Loos RJ, Lokki ML, Lettre G, Langenberg C, Launer LJ, Lakatta EG, Laaksonen R,

144
Kyvik KO, Kronenberg F, Konig IR, Khaw KT, Kaprio J, Kaplan LM, Johansson A, Jarvelin MR, Janssens AC, Ingelsson E, Igl W, Kees Hovingh G, Hottenga JJ, Hofman A, Hicks AA, Hengstenberg C, Heid IM, Hayward C, Havulinna AS, Hastie ND, Harris TB, Haritunians T, Hall AS, Gyllensten U, Guiducci C, Groop LC, Gonzalez E, Gieger C, Freimer NB, Ferrucci L, Erdmann J, Elliott P, Ejebe KG, Doring A, Dominiczak AF, Demissie S, Deloukas P, de Geus EJ, de Faire U, Crawford G, Collins FS, Chen YD, Caulfield MJ, Campbell H, Burtt NP, Bonnycastle LL, Boomsma DI, Boekholdt SM, Bergman RN, Barroso I, Bandinelli S, Ballantyne CM, Assimes TL, Quertermous T, Altshuler D, Seielstad M, Wong TY, Tai ES, Feranil AB, Kuzawa CW, Adair LS, Taylor HA, Borecki IB, Gabriel SB, Wilson JG, Holm H, Thorsteinsdottir U, Gudnason V, Krauss RM, Mohlke KL, Ordovas JM, Munroe PB, Kooner JS, Tall AR, Hegele RA, Kastelein JJ, Schadt EE, Rotter JI, Boerwinkle E, Strachan DP, Mooser V, Stefansson K, Reilly MP, Samani NJ, Schunkert H, Cupples LA, Sandhu MS, Ridker PM, Rader DJ, van Duijn CM, Peltonen L, Abecasis GR, Boehnke M & Kathiresan S (2010) Biological, clinical and population relevance of 95 loci for blood lipids. Nature 466(7307): 707–713.
The 1000 Genomes Project Consortium (2010) A map of human genome variation from population-scale sequencing. Nature 467(7319): 1061–1073.
Thomas L (1997) Retrospective power analysis. Conservation Biology 11(1):276–280 Thompson JP, Pearce RH, Schechter MT, Adams ME, Tsang IK & Bishop PB (1990)
Preliminary evaluation of a scheme for grading the gross morphology of the human intervertebral disc. Spine (Phila Pa 1976) 15(5): 411–415.
Thornbury JR, Fryback DG, Turski PA, Javid MJ, McDonald JV, Beinlich BR, Gentry LR, Sackett JF, Dasbach EJ & Martin PA (1993) Disk-caused nerve compression in patients with acute low-back pain: diagnosis with MR, CT myelography, and plain CT. Radiology 186(3): 731–738.
Trout JJ, Buckwalter JA & Moore KC (1982) Ultrastructure of the human intervertebral disc: II. Cells of the nucleus pulposus. Anat Rec 204(4): 307–314.
Twomey L & Taylor J (1985) Age changes in lumbar intervertebral discs. Acta Orthop Scand 56(6): 496–499.
Uei H, Matsuzaki H, Oda H, Nakajima S, Tokuhashi Y & Esumi M (2006) Gene expression changes in an early stage of intervertebral disc degeneration induced by passive cigarette smoking. Spine (Phila Pa 1976) 31(5): 510–514.
Urban JP & Winlove CP (2007) Pathophysiology of the intervertebral disc and the challenges for MRI. J Magn Reson Imaging 25(2): 419–432.
Urban JP, Holm S, Maroudas A & Nachemson A (1977) Nutrition of the intervertebral disk. An in vivo study of solute transport. Clin Orthop Relat Res (129): 101–114.
Urban JPG, Smith S & Fairbank JCT (2004) Nutrition of the intervertebral disc. Spine (Phila Pa 1976) 29(23): 2700–2709.
Urban MR, Fairbank JC, Bibby SR & Urban JP (2001) Intervertebral disc composition in neuromuscular scoliosis: changes in cell density and glycosaminoglycan concentration at the curve apex. Spine (Phila Pa 1976) 26(6): 610–617.

145
Valdes AM, Hart DJ, Jones KA, Surdulescu G, Swarbrick P, Doyle DV, Schafer AJ & Spector TD (2004) Association study of candidate genes for the prevalence and progression of knee osteoarthritis. Arthritis Rheum 50(8): 2497–2507.
Valdes AM, Loughlin J, Oene MV, Chapman K, Surdulescu GL, Doherty M & Spector TD (2007) Sex and ethnic differences in the association of ASPN, CALM1, COL2A1, COMP, and FRZB with genetic susceptibility to osteoarthritis of the knee. Arthritis Rheum 56(1): 137–146.
van Tulder M, Becker A, Bekkering T, Breen A, del Real M,Teresa Gil, Hutchinson A, Koes B, Laerum E, Malmivaara A & (2006) Chapter 3. European guidelines for the management of acute nonspecific low back pain in primary care. Eur Spine J 15 Suppl 2: 169–191.
van dB & Bresnihan B (1999) Pathogenesis of joint damage in rheumatoid arthritis: evidence of a dominant role for interleukin-I. Baillieres Best Pract Res Clin Rheumatol 13(4): 577–597.
van dS, Verhage M, Posthuma D & Dolan CV (2010) Phenotypic complexity, measurement bias, and poor phenotypic resolution contribute to the missing heritability problem in genetic association studies. PLoS One 5(11).
Varlotta GP, Brown MD, Kelsey JL & Golden AL (1991) Familial predisposition for herniation of a lumbar disc in patients who are less than twenty-one years old J Bone Joint Surg Am 73(1): 124–128.
Vermeer S, Hoischen A, Meijer RP, Gilissen C, Neveling K, Wieskamp N, de Brouwer A, Koenig M, Anheim M, Assoum M, Drouot N, Todorovic S, Milic-Rasic V, Lochm\uller H, Stevanin G, Goizet C, David A, Durr A, Brice A, Kremer B, van de Warrenburg BP, Schijvenaars MM, Heister A, Kwint M, Arts P, van der Wijst J, Veltman J, Kamsteeg EJ, Scheffer H & Knoers N (2010) Targeted next-generation sequencing of a 12.5 Mb homozygous region reveals ANO10 mutations in patients with autosomal-recessive cerebellar ataxia. Am J Hum Genet 87(6): 813–819.
Videman T, Battié M C, Parent E, Gibbons LE, Vainio P & Kaprio J (2008) Progression and determinants of quantitative magnetic resonance imaging measures of lumbar disc degeneration: a five-year follow-up of adult male monozygotic twins. Spine (Phila Pa 1976) 33(13): 1484–1490.
Videman T, Battie MC, Gill K, Manninen H, Gibbons LE & Fisher LD (1995) Magnetic resonance imaging findings and their relationships in the thoracic and lumbar spine. Insights into the etiopathogenesis of spinal degeneration. Spine (Phila Pa 1976) 20(8): 928–935.
Videman T, Gibbons LE, Battie MC, Maravilla K, Vanninen E, Leppavuori J, Kaprio J & Peltonen L (2001) The relative roles of intragenic polymorphisms of the vitamin d receptor gene in lumbar spine degeneration and bone density. Spine (Phila Pa 1976) 26(3): 7–7.
Videman T, Leppavuori J, Kaprio J, Battie MC, Gibbons LE, Peltonen L & Koskenvuo M (1998) Intragenic polymorphisms of the vitamin D receptor gene associated with intervertebral disc degeneration. Spine (Phila Pa 1976) 23(23): 2477–2485.

146
Videman T, Battie MC, Ripatti S, Gill K, Manninen H & Kaprio J (2006) Determinants of the progression in lumbar degeneration: a 5-year follow-up study of adult male monozygotic twins. Spine (Phila Pa 1976) 31(6): 671–678.
Videman T, Saarela J, Kaprio J, Nakki A, Levalahti E, Gill K, Peltonen L & Battie MC (2009) Associations of 25 structural, degradative, and inflammatory candidate genes with lumbar disc desiccation, bulging, and height narrowing. Arthritis Rheum 60(2): 470–481.
Virtanen IM, Song YQ, Cheung KMC, Ala-Kokko L, Karppinen J, Ho DWH, Luk KDK, Yip SP, Leong JCY, Cheah KSE, Sham P & Chan D (2007a) Phenotypic and population differences in the association between CILP and lumbar disc disease. J Med Genet 44(4): 285–288.
Virtanen IM, Noponen N, Barral S, Karppinen J, Li H, Vuoristo M, Niinimaki J, Ott J, Ala-Kokko L & Mannikko M (2007b) Putative susceptibility locus on chromosome 21q for lumbar disc disease (LDD) in the Finnish population. J Bone Miner Res 22(5): 701–707.
Visuri T, Ulaska J, Eskelin M & Pulkkinen P (2005) Narrowing of lumbar spinal canal predicts chronic low back pain more accurately than intervertebral disc degeneration: a magnetic resonance imaging study in young Finnish male conscripts. Mil Med 170(11): 926–930.
Vuorio AF, Aalto-Setala K, Koivisto UM, Turtola H, Nissen H, Kovanen PT, Miettinen TA, Gylling H, Oksanen H, Kontula K & (2001) Familial hypercholesterolaemia in Finland: common, rare and mild mutations of the LDL receptor and their clinical consequences. Finnish FH-group. Ann Med 33(6): 410–421.
Waris E, Eskelin M, Hermunen H, Kiviluoto O & Paajanen H (2007) Disc degeneration in low back pain: a 17-year follow-up study using magnetic resonance imaging Spine (Phila Pa 1976) 32(6): 681–684.
Watanabe H, Cheung SC, Itano N, Kimata K & Yamada Y (1997) Identification of hyaluronan-binding domains of aggrecan. J Biol Chem 272(44): 28057–28065.
Wedderkopp N, Andersen LB, Froberg K & Leboeuf-Yde C (2005) Back pain reporting in young girls appears to be puberty-related. BMC Musculoskelet Disord 6: 52–52.
Weishaupt D, Zanetti M, Hodler J & Boos N (1998) MR imaging of the lumbar spine: prevalence of intervertebral disk extrusion and sequestration, nerve root compression, end plate abnormalities, and osteoarthritis of the facet joints in asymptomatic volunteers. Radiology 209(3): 661–666.
Wellcome Trust Case Control Consortium (2007) Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature 447(7145): 661–678.

147
Wellcome Trust Case Control Consortium, Craddock N, Hurles ME, Cardin N, Pearson RD, Plagnol V, Robson S, Vukcevic D, Barnes C, Conrad DF, Giannoulatou E, Holmes C, Marchini JL, Stirrups K, Tobin MD, Wain LV, Yau C, Aerts J, Ahmad T, Andrews TD, Arbury H, Attwood A, Auton A, Ball SG, Balmforth AJ, Barrett JC, Barroso I, Barton A, Bennett AJ, Bhaskar S, Blaszczyk K, Bowes J, Brand OJ, Braund PS, Bredin F, Breen G, Brown MJ, Bruce IN, Bull J, Burren OS, Burton J, Byrnes J, Caesar S, Clee CM, Coffey AJ, Connell JMC, Cooper JD, Dominiczak AF, Downes K, Drummond HE, Dudakia D, Dunham A, Ebbs B, Eccles D, Edkins S, Edwards C, Elliot A, Emery P, Evans DM, Evans G, Eyre S, Farmer A, Ferrier IN, Feuk L, Fitzgerald T, Flynn E, Forbes A, Forty L, Franklyn JA, Freathy RM, Gibbs P, Gilbert P, Gokumen O, Gordon-Smith K, Gray E, Green E, Groves CJ, Grozeva D, Gwilliam R, Hall A, Hammond N, Hardy M, Harrison P, Hassanali N, Hebaishi H, Hines S, Hinks A, Hitman GA, Hocking L, Howard E, Howard P, Howson JMM, Hughes D, Hunt S, Isaacs JD, Jain M, Jewell DP, Johnson T, Jolley JD, Jones IR, Jones LA, Kirov G, Langford CF, Lango-Allen H, Lathrop GM, Lee J, Lee KL, Lees C, Lewis K, Lindgren CM, Maisuria-Armer M, Maller J, Mansfield J, Martin P, Massey DCO, McArdle WL, McGuffin P, McLay KE, Mentzer A, Mimmack ML, Morgan AE, Morris AP, Mowat C, Myers S, Newman W, Nimmo ER, O'Donovan MC, Onipinla A, Onyiah I, Ovington NR, Owen MJ, Palin K, Parnell K, Pernet D, Perry JRB, Phillips A, Pinto D, Prescott NJ, Prokopenko I, Quail MA, Rafelt S, Rayner NW, Redon R, Reid DM, Renwick , Ring SM, Robertson N, Russell E, St Clair D, Sambrook JG, Sanderson JD, Schuilenburg H, Scott CE, Scott R, Seal S, Shaw-Hawkins S, Shields BM, Simmonds MJ, Smyth DJ, Somaskantharajah E, Spanova K, Steer S, Stephens J, Stevens HE, Stone MA, Su Z, Symmons DPM, Thompson JR, Thomson W, Travers ME, Turnbull C, Valsesia A, Walker M, Walker NM, Wallace C, Warren-Perry M, Watkins NA, Webster J, Weedon MN, Wilson AG, Woodburn M, Wordsworth BP, Young AH, Zeggini E, Carter NP, Frayling TM, Lee C, McVean G, Munroe PB, Palotie A, Sawcer SJ, Scherer SW, Strachan DP, Tyler-Smith C, Brown MA, Burton PR, Caulfield MJ, Compston A, Farrall M, Gough SCL, Hall AS, Hattersley AT, Hill AVS, Mathew CG, Pembrey M, Satsangi J, Stratton MR, Worthington J, Deloukas P, Duncanson A, Kwiatkowski DP, McCarthy MI, Ouwehand W, Parkes M, Rahman N, Todd JA, Samani NJ & Donnelly P (2010) Genome-wide association study of CNVs in 16,000 cases of eight common diseases and 3,000 shared controls. Nature 464(7289): 713–720.
Whatley BR & Wen X (2012) Intervertebral disc (IVD): Structure, degeneration, repair and regeneration. Mater Sci Eng C 32(2): 61–77.
Wieser S, Horisberger B, Schmidhauser S, Eisenring C, Brugger U, Ruckstuhl A, Dietrich J, Mannion AF, Elfering A, Tamcan O & Muller U (2011) Cost of low back pain in Switzerland in 2005. Eur J Health Econ 12(5): 455–467.

148
Williams FMK, Bansal AT, van Meurs JB, Bell JT, Meulenbelt I, Suri P, Rivadeneira F, Sambrook PN, Hofman A, Bierma-Zeinstra S, Menni C, Kloppenburg M, Slagboom PE, Hunter DJ, Macgregor AJ, Uitterlinden AG & Spector TD (2012) Novel genetic variants associated with lumbar disc degeneration in northern Europeans: a meta-analysis of 4600 subjects. Ann Rheum Dis [epub].
Williams FM, Kato BS, Livshits G, Sambrook PN, Spector TD & MacGregor AJ (2008) Lumbar disc disease shows linkage to chromosome 19 overlapping with a QTL for hand OA. Ann Rheum Dis 67(1): 117–119.
Williams FM, Popham M, Hart DJ, de Schepper E, Bierma-Zeinstra S, Hofman A, Uitterlinden AG, Arden NK, Cooper C, Spector TD, Valdes AM & van Meurs J (2011a) GDF5 single-nucleotide polymorphism rs143383 is associated with lumbar disc degeneration in Northern European women. Arthritis Rheum 63(3): 708–712.
Williams FM, Popham M, Sambrook PN, Jones AF, Spector TD & Macgregor AJ (2011b) Progression of lumbar disc degeneration over a decade: a heritability study. Ann Rheum Dis 70(7): 1203–1207.
Williams FMK, Manek NJ, Sambrook PN, Spector TD & Macgregor AJ (2007) Schmorl's nodes: common, highly heritable, and related to lumbar disc disease. Arthritis Rheum 57(5): 855–860.
Wiltse LL (1971) The effect of the common anomalies of the lumbar spine upon disc degeneration and low back pain. Orthop Clin North Am 2(2): 569–582.
Xu B, Roos JL, Dexheimer P, Boone B, Plummer B, Levy S, Gogos JA & Karayiorgou M (2011) Exome sequencing supports a de novo mutational paradigm for schizophrenia. Nat Genet 43(9): 864–868.
Yang WS, Chuang LM (2006) Human genetics of adiponectin in the metabolic syndrome. J Mol Med: 84(2):112–121.
Yasuma T, Koh S, Okamura T & Yamauchi Y (1990) Histological changes in aging lumbar intervertebral discs. Their role in protrusions and prolapses. J Bone Joint Surg Am 72(2): 220–229.
Ye S, Eriksson P, Hamsten A, Kurkinen M, Humphries SE & Henney AM (1996) Progression of coronary atherosclerosis is associated with a common genetic variant of the human stromelysin-1 promoter which results in reduced gene expression. J Biol Chem 271(22): 13055–13060.
Yesupriya A, Evangelou E, Kavvoura FK, Patsopoulos NA, Clyne M, Walsh MC, Lin BK, Yu W, Gwinn M, Ioannidis JPA & Khoury MJ (2008) Reporting of human genome epidemiology (HuGE) association studies: an empirical assessment. BMC Med Res Methodol 8: 31–31.
Yu B (2009) Role of in silico tools in gene discovery. Mol Biotechnol 41(3): 296–306. Yu J, Tirlapur U, Fairbank J, Handford P, Roberts S, Winlove CP, Cui Z & Urban J (2007)
Microfibrils, elastin fibres and collagen fibres in the human intervertebral disc and bovine tail disc. J Anat 210(4): 460–471.
Yu SW, Sether LA, Ho PS, Wagner M & Haughton VM (1988) Tears of the anulus fibrosus: correlation between MR and pathologic findings in cadavers. AJNR Am J Neuroradiol 9(2): 367–370.

149
Zhang Y, Sun Z, Liu J & Guo X (2008) Advances in susceptibility genetics of intervertebral degenerative disc disease. Int J Biol Sci 4(5): 283–290.
Zhao CQ, Wang LM, Jiang LS & Dai LY (2007) The cell biology of intervertebral disc aging and degeneration. Ageing Res Rev 6(3): 247–261.
Zhao F, Pollintine P, Hole BD, Dolan P & Adams MA (2005) Discogenic origins of spinal instability. Spine (Phila Pa 1976) 30(23): 2621–2630.
Zhu G, Jiang X, Xia C, Sun Y, Zeng Q, Wu X & Li X (2011) Association of FAS and FAS ligand polymorphisms with the susceptibility and severity of lumbar disc degeneration in Chinese Han population. Biomarkers .
Zortea M, Vettori A, Trevisan CP, Bellini S, Vazza G, Armani M, Simonati A & Mostacciuolo ML (2002) Genetic mapping of a susceptibility locus for disc herniation and spastic paraplegia on 6q23.3-q24.1. J Med Genet 39(6): 387–390.
Zou J, Yang H, Miyazaki M, Morishita Y, Wei F, McGovern S & Wang JC (2009) Dynamic bulging of intervertebral discs in the degenerative lumbar spine. Spine (Phila Pa 1976) 34(23): 2545–2550.
Zuk O, Hechter E, Sunyaev SR & Lander ES (2012) The mystery of missing heritability: Genetic interactions create phantom heritability. Proc Natl Acad Sci U S A 109(4): 1193–1198.
Zuo J, Saadat E, Romero A, Loo K, Li X, Link TM, Kurhanewicz J & Majumdar S (2009) Assessment of intervertebral disc degeneration with magnetic resonance single-voxel spectroscopy. Magn Reson Med 62(5): 1140–1146.

150
Ap
pe
nd
ix 1
Su
pp
ort
ing
in
form
ati
on
ta
ble
1.
Th
e s
tud
ies in
clu
de
d in
th
e s
ys
tem
ati
c r
ev
iew
(I)
.
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
AC
AN
V
NT
R
D
64
Case
-contr
ol
A18+
A21
- 0.0
08
<
0.0
5
<0.0
5
Japanese
K
aw
aguch
i
et al.
1999
V
NT
R
N,S
100
Clin
ical
popula
tion
A1
3-A
25
-
<0
.00
1
- T
urk
ish
E
ser
et al.
2011
V
NT
R
D(N
),H
300
Clin
ical
popula
tion
A1
3-A
26
A27
- <
0.0
01
0.0
07
0.0
02
Tu
rkis
h
Ese
r
et al.
2010
V
NT
R
D,B
132
Cro
ss-
sect
ional
A26
2.7
7
[1.2
4–6.1
6]
- <
0.0
5
Fin
nis
h
Solo
vieva
et al.
2007
V
NT
R
N
132
Clin
ical
popula
tion
<A
26
4.5
[1.5
9–12.7
]
0.0
05
0.0
11
C
hin
ese
C
ong
et al.
2010a
V
NT
R
S
19
7
Ca
se-c
on
tro
l A
25
2.1
2
[1.1
3–3.9
8]
0.0
19
0.0
05
C
hin
ese
C
ong
et al.
2010b
V
NT
R
U
179
Nest
ed c
ase
-
con
tro
l
<A
25
3.2
8
[1.6
2–6.6
5]
<0.0
01
0.0
17
Ir
anese
M
ash
aye
khi
et al.
2010
V
NT
R
St
85
C
ase
-co
ntr
ol
- -
- -
Fin
nis
h
No
po
ne
n
et al.
2003
V
NT
R
U
102
Nest
ed
case
-contr
ol
- -
- 0
.00
3
- R
ou
gh
ley
et al.
2006
V
NT
R
N,S
104
Clin
ical
popula
tion
A2
1
- <
0.0
06
<
0.0
5
Ko
rea
n
Kim
et al.
2011
rs
1042631
D
B
588
Male
tw
in
popula
tion
- -
0.0
01
0.0
10
0.2
3
Fin
nis
h
Vid
em
an
et al.
2009
151
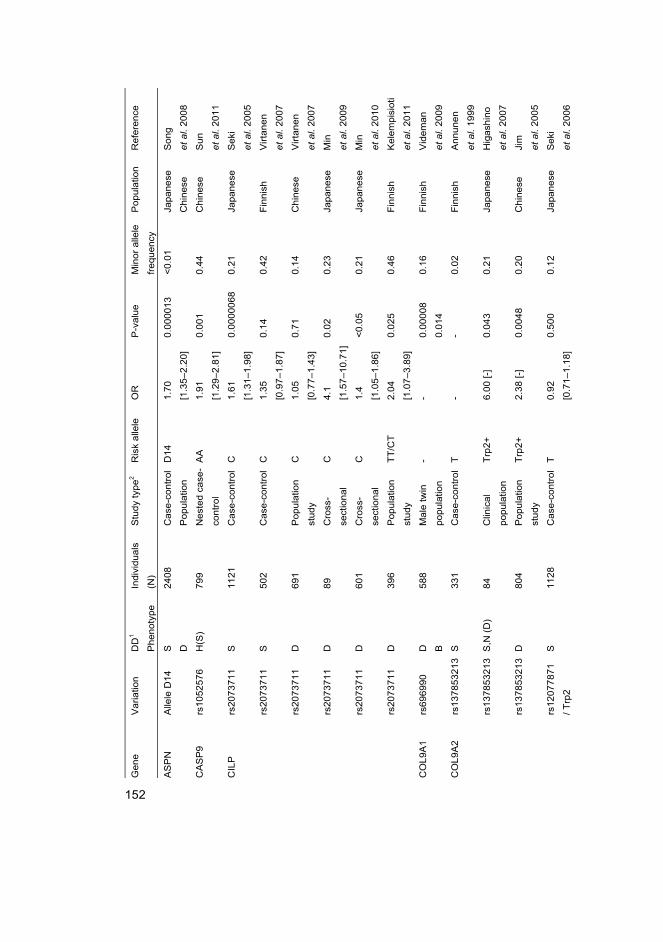
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
AS
PN
A
llele
D14
S
D
2408
C
ase
-contr
ol
Popula
tion
D14
1.7
0
[1.3
5–2.2
0]
0.0
00013
<
0.0
1
Japanese
Chin
ese
So
ng
et al.
2008
CA
SP
9
rs1
05
25
76
H
(S)
79
9
Ne
ste
d c
ase
-
con
tro
l
AA
1.9
1
[1.2
9–2.8
1]
0.0
01
0.4
4
Chin
ese
S
un
et al.
2011
CIL
P
rs2073711
S
1121
C
ase
-contr
ol
C
1.6
1
[1.3
1–1.9
8]
0.0
000068
0.2
1
Japanese
S
eki
et al.
2005
rs
2073711
S
502
Case
-contr
ol
C
1.3
5
[0.9
7–1.8
7]
0.1
4
0.4
2
Fin
nis
h
Virta
nen
et al.
2007
rs
2073711
D
691
Popula
tion
study
C
1.0
5
[0.7
7–1.4
3]
0.7
1
0.1
4
Chin
ese
V
irta
nen
et al.
2007
rs
2073711
D
89
Cro
ss-
sect
ional
C
4.1
[1.5
7–10.7
1]
0.0
2
0.2
3
Japanese
M
in
et al.
2009
rs
2073711
D
601
Cro
ss-
sect
ional
C
1.4
[1.0
5–1.8
6]
<0.0
5
0.2
1
Japanese
M
in
et al.
2010
rs
2073711
D
396
Popula
tion
study
TT
/CT
2
.04
[1.0
7–3.8
9]
0.0
25
0.4
6
Fin
nis
h
Kele
mpis
ioti
et al.
2011
CO
L9A
1
rs696990
D
B
588
Male
tw
in
popula
tion
- -
0.0
00
08
0.0
14
0.1
6
Fin
nis
h
Vid
em
an
et al.
2009
CO
L9
A2
rs
13
78
53
21
3
S
33
1
Ca
se-c
on
tro
l T
-
- 0
.02
F
inn
ish
A
nnunen
et al.
1999
rs
137853213
S
,N (
D)
84
Clin
ical
popula
tion
Trp
2+
6.0
0 [-]
0.0
43
0.2
1
Japanese
H
igash
ino
et al.
2007
rs
137853213
D
804
Popula
tion
study
Trp
2+
2.3
8 [-]
0.0
048
0.2
0
Chin
ese
Ji
m
et al.
2005
rs
12077871
/ T
rp2
S
11
28
C
ase
-co
ntr
ol
T
0.9
2
[0.7
1–1.1
8]
0.5
00
0.1
2
Japanese
S
eki
et al.
2006
152
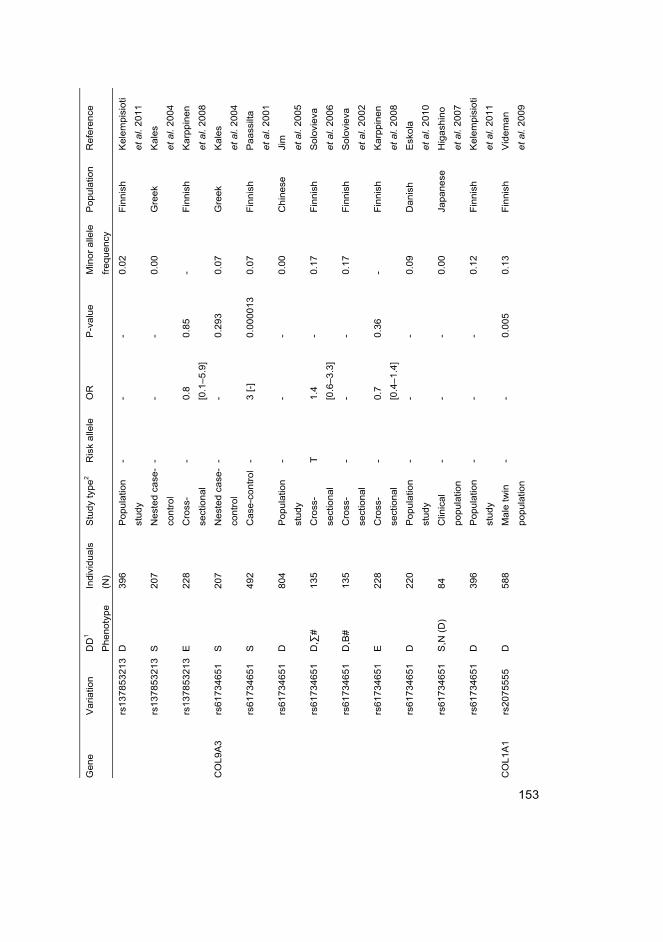
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
rs
137853213
D
396
Popula
tion
study
- -
- 0
.02
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
137853213
S
207
Nest
ed c
ase
-
con
tro
l
- -
- 0
.00
G
ree
k K
ale
s
et al.
2004
rs
137853213
E
228
Cro
ss-
sect
ional
- 0
.8
[0.1
–5.9
]
0.8
5
- F
inn
ish
K
arp
pin
en
et al.
2008
CO
L9A
3
rs61734651
S
207
Nest
ed c
ase
-
con
tro
l
- -
0.2
93
0
.07
G
ree
k K
ale
s
et al.
2004
rs
61734651
S
492
Case
-contr
ol
- 3 [-]
0.0
00013
0.0
7
Fin
nis
h
Paass
ilta
et al.
2001
rs
61734651
D
804
Popula
tion
study
- -
- 0
.00
C
hin
ese
Ji
m
et al.
2005
rs
61734651
D
,∑#
135
Cro
ss-
sect
ional
T
1.4
[0.6
–3.3
]
- 0.1
7
Fin
nis
h
Solo
vieva
et al.
2006
rs
61734651
D
,B#
135
Cro
ss-
sect
ional
- -
- 0
.17
F
inn
ish
S
olo
vie
va
et al.
2002
rs
61734651
E
228
Cro
ss-
sect
ional
- 0
.7
[0.4
–1.4
]
0.3
6
- F
inn
ish
K
arp
pin
en
et al.
2008
rs
61734651
D
220
Popula
tion
study
- -
- 0
.09
D
an
ish
E
sko
la
et al.
2010
rs
61734651
S
,N (
D)
84
Clin
ical
popula
tion
- -
- 0
.00
Ja
pa
ne
se
Hig
ash
ino
et al.
2007
rs
61734651
D
396
Popula
tion
study
- -
- 0
.12
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
CO
L1A
1
rs2075555
D
588
Male
tw
in
popula
tion
- -
0.0
05
0
.13
F
inn
ish
V
ide
ma
n
et al.
2009
153
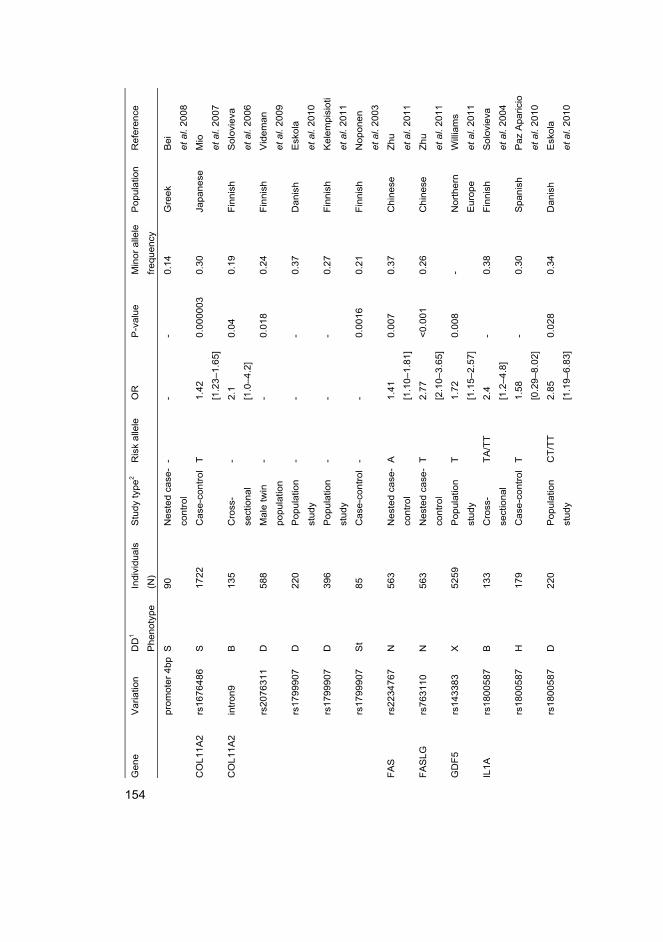
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
p
rom
ote
r 4
bp
S
90
N
est
ed
ca
se-
con
tro
l
- -
- 0
.14
G
ree
k B
ei
et al.
2008
CO
L1
1A
2
rs1
67
64
86
S
1
72
2
Ca
se-c
on
tro
l T
1
.42
[1.2
3–1.6
5]
0.0
00003
0.3
0
Japanese
M
io
et al.
2007
CO
L11A
2
intr
on9
B
135
Cro
ss-
sect
ional
- 2
.1
[1.0
–4.2
]
0.0
4
0.1
9
Fin
nis
h
Solo
vieva
et al.
2006
rs
2076311
D
588
Male
tw
in
popula
tion
- -
0.0
18
0
.24
F
inn
ish
V
ide
ma
n
et al.
2009
rs
1799907
D
220
Popula
tion
study
- -
- 0
.37
D
an
ish
E
sko
la
et al.
2010
rs
1799907
D
396
Popula
tion
study
- -
- 0
.27
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
1799907
S
t 85
Case
-contr
ol
- -
0.0
016
0.2
1
Fin
nis
h
Noponen
et al.
2003
FA
S
rs2234767
N
563
Nest
ed c
ase
-
con
tro
l
A
1.4
1
[1.1
0–1.8
1]
0.0
07
0.3
7
Chin
ese
Z
hu
et al.
2011
FA
SLG
rs
763110
N
563
Nest
ed c
ase
-
con
tro
l
T
2.7
7
[2.1
0–3.6
5]
<0.0
01
0.2
6
Chin
ese
Z
hu
et al.
2011
GD
F5
rs
14
33
83
X
5
25
9
Po
pu
latio
n
study
T
1.7
2
[1.1
5–2.5
7]
0.0
08
-
No
rth
ern
Eu
rop
e
Will
iam
s
et al.
2011
IL1
A
rs1
80
05
87
B
1
33
C
ross
-
sect
ional
TA
/TT
2
.4
[1.2
–4.8
]
- 0.3
8
Fin
nis
h
Solo
vieva
et al.
2004
rs
1800587
H
179
Case
-contr
ol
T
1.5
8
[0.2
9–8.0
2]
- 0
.30
S
pa
nis
h
Pa
z A
pa
rici
o
et al.
2010
rs
1800587
D
220
Popula
tion
study
CT
/TT
2
.85
[1.1
9–6.8
3]
0.0
28
0.3
4
Danis
h
Esk
ola
et al.
2010
154
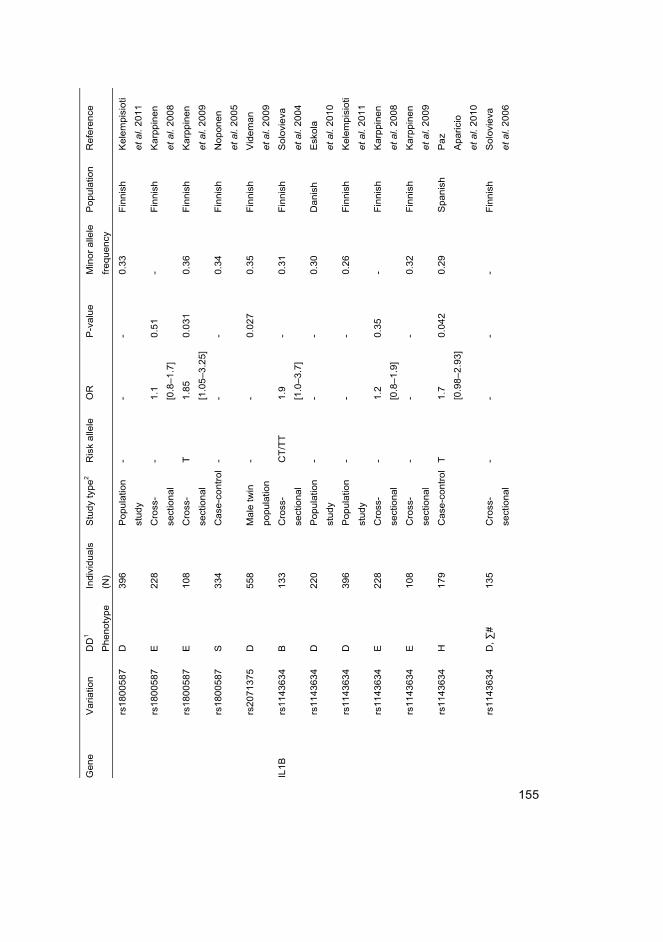
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
rs
1800587
D
396
Popula
tion
study
- -
- 0
.33
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
1800587
E
228
Cro
ss-
sect
ional
- 1
.1
[0.8
–1.7
]
0.5
1
- F
inn
ish
K
arp
pin
en
et al.
2008
rs
1800587
E
108
Cro
ss-
sect
ional
T
1.8
5
[1.0
5–3.2
5]
0.0
31
0.3
6
Fin
nis
h
Karp
pin
en
et al.
2009
rs
18
00
58
7
S
33
4
Ca
se-c
on
tro
l -
- -
0.3
4
Fin
nis
h
No
po
ne
n
et al.
2005
rs
2071375
D
558
Male
tw
in
popula
tion
- -
0.0
27
0
.35
F
inn
ish
V
ide
ma
n
et al.
2009
IL1
B
rs1
14
36
34
B
1
33
C
ross
-
sect
ional
CT
/TT
1
.9
[1.0
–3.7
]
- 0.3
1
Fin
nis
h
Solo
vieva
et al.
2004
rs
1143634
D
220
Popula
tion
study
- -
- 0
.30
D
an
ish
E
sko
la
et al.
2010
rs
1143634
D
396
Popula
tion
study
- -
- 0
.26
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
1143634
E
228
Cro
ss-
sect
ional
- 1
.2
[0.8
–1.9
]
0.3
5
- F
inn
ish
K
arp
pin
en
et al.
2008
rs
1143634
E
108
Cro
ss-
sect
ional
- -
- 0
.32
F
inn
ish
K
arp
pin
en
et al.
2009
rs
1143634
H
179
Case
-contr
ol
T
1.7
[0.9
8–2.9
3]
0.0
42
0.2
9
Spanis
h
Paz
Aparici
o
et al.
2010
rs
1143634
D
, ∑
#
135
Cro
ss-
sect
ional
- -
- -
Fin
nis
h
So
lovi
eva
et al.
2006
155

Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
rs
11
43
63
4
S
33
4
Ca
se-c
on
tro
l -
- -
0.4
3
Fin
nis
h
No
po
ne
n
et al.
2005
IL1R
N
VN
TR
H
(S)
281
Case
-contr
ol
A3
3.8
6
[1.3
7–10.9
0]
0.0
00
6
0.0
0
Ko
rea
n
Kim
et al.
2010
V
NT
R
S
33
4
Ca
se-c
on
tro
l -
- -
0.3
0
Fin
nis
h
No
po
ne
n
et al.
2005
IL6
p
rom
ote
r S
3
34
C
ase
-co
ntr
ol
GG
GA
4
.80
[1.5
9–14.4
5]
0.0
033
0.0
4
Fin
nis
h
Noponen
et al.
2005
pro
mote
r D
220
Popula
tion
study
GC
G
6.4
6
[1.6
2–26.0
]
0.0
09
0.0
5
Danis
h
Esk
ola
et al.
2010
pro
mote
r D
396
Popula
tion
study
GG
G
1.4
8
[1.0
9–2.0
1]
0.0
122
0.0
3
Fin
nis
h
Kele
mpis
ioti
et al.
2011
IL10
rs1800896
D
(N)
589
Nest
ed c
ase
-
con
tro
l
AA
1.3
4
[1.1
2–3.0
3]
0.0
32
0.4
2
Chin
ese
Lin
et al.
2011
rs
18
00
89
6
S
33
4
Ca
se-c
on
tro
l -
- -
0.4
4
Fin
nis
h
No
po
ne
n
et al.
2005
rs
1800872
D
(N)
589
Nest
ed c
ase
-
con
tro
l
AA
1.6
8
[1.2
3–3.4
6]
0.0
26
0.4
3
Chin
ese
Lin
et al.
2011
IL18R
AP
rs
1420100
D
588
Male
tw
in
popula
tion
- -
0.0
05
0
.48
F
inn
ish
V
ide
ma
n
et al.
2009
MM
P1
rs1799750
D
691
Popula
tion
study
dele
tion
1.4
1
[1.0
4–1.9
0]
0.0
27
0.3
6
Chin
ese
S
ong
et al.
2008
MM
P2
rs
24
38
65
S
4
80
N
est
ed
ca
se-
con
tro
l
CC
3.0
8
[1.8
4–5.1
6]
<0.0
01
0.1
4
Chin
ese
D
ong
et al.
2007
MM
P3
rs
30
25
05
8
X
10
3
Po
pu
latio
n
study
5A
/5A
+
5A
/6A
- 0.0
029
0.1
9
Japanese
T
aka
hash
i
et al.
2001
156

Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
rs
30
25
05
8
St
85
C
ase
-co
ntr
ol
- -
- 0
.39
F
inn
ish
N
op
on
en
et al.
2003
rs
3025058
E
#
228
Cro
ss-
sect
ional
5A
1
.1
[0.7
–1.6
]
- 0
.36
F
inn
ish
K
arp
pin
en
et al.
2008
MM
P9
rs17576
D
396
Popula
tion
study
- -
- 0
.40
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
17576
S
1743
C
ase
-contr
ol
G
1.2
9
[1.1
2–1.4
8]
0.0
0049
0.3
4
Japanese
H
irose
et al.
2008
rs
3918242
S
,N
859
Popula
tion
study
CT
+T
T
2.1
4
[1.5
5–2.9
6]
<0.0
1
0.1
2
Chin
ese
S
un
et al.
2009
NO
S2
rs
1060826
H
,N
179
Case
-contr
ol
GA
+G
G
- 0.0
09
0.3
6
Spanis
h
Paz
Aparici
o
et al.
2010
NO
S3
rs
2070744
H
,N
179
Case
-contr
ol
- 0.1
2
[0.0
3–0.4
3]
0.0
002
0.5
1
Spanis
h
Paz
Aparici
o
et al.
2010
SK
T
rs1
69
24
57
3
S
S
1758
506
Ca
se-c
on
tro
l
G
G
1.3
1
[1.1
1–1.5
5]
2.8
1
[1.0
9–7.2
4]
0.0
032
0.0
26
0.2
0
0.0
4
Japanese
Fin
nis
h
Ka
rasu
gi
et al.
2009
rs
16924573
D
396
Popula
tion
study
GA
0
.27
[0.0
7–0.9
6]
0.0
23
0.0
2
Fin
nis
h
Kele
mpis
ioti
et al.
2011
TH
BS
2
rs9
40
63
28
S
1
74
3
Ca
se-c
on
tro
l T
1
.38
[1.2
1–1.5
8]
0.0
000028
0.4
3
Japanese
H
irose
et al.
2008
rs
9406328
D
396
Popula
tion
study
- -
- 0
.26
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
157
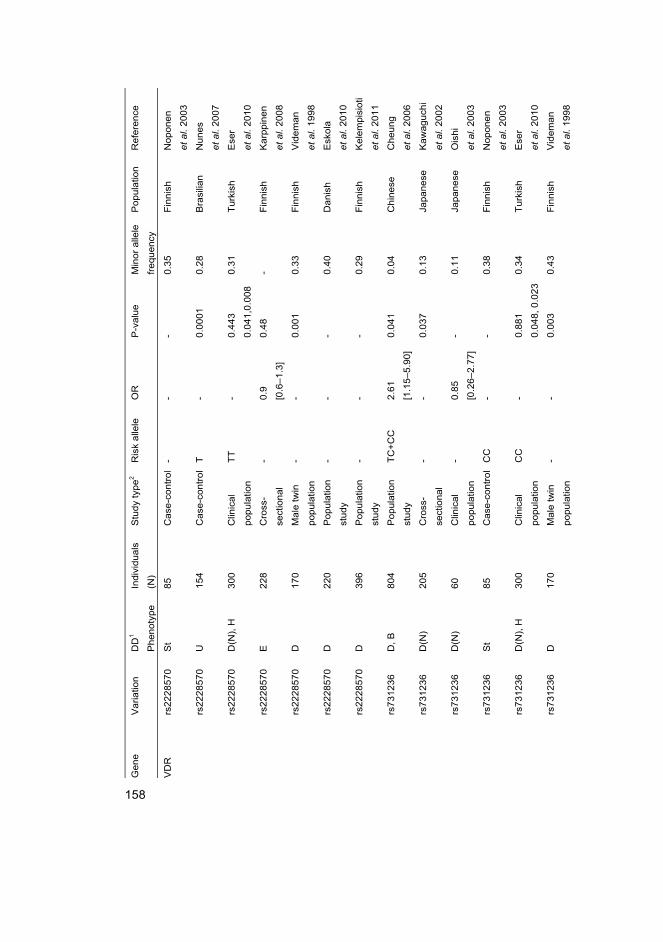
Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
VD
R
rs2
22
85
70
S
t 8
5
Ca
se-c
on
tro
l -
- -
0.3
5
Fin
nis
h
No
po
ne
n
et al.
2003
rs
2228570
U
154
Case
-contr
ol
T
- 0.0
001
0.2
8
Bra
silia
n
Nunes
et al.
2007
rs
2228570
D
(N),
H
300
Clin
ical
popula
tion
TT
-
0.4
43
0.0
41,0
.008
0.3
1
Tu
rkis
h
Ese
r
et al.
2010
rs
2228570
E
228
Cro
ss-
sect
ional
- 0
.9
[0.6
–1.3
]
0.4
8
- F
inn
ish
K
arp
pin
en
et al.
2008
rs
2228570
D
170
Male
tw
in
popula
tion
- -
0.0
01
0
.33
F
inn
ish
V
ide
ma
n
et al.
1998
rs
2228570
D
220
Popula
tion
study
- -
- 0
.40
D
an
ish
E
sko
la
et al.
2010
rs
2228570
D
396
Popula
tion
study
- -
- 0
.29
F
inn
ish
K
ele
mp
isio
ti
et al.
2011
rs
73
12
36
D
, B
8
04
P
op
ula
tion
study
TC
+C
C
2.6
1
[1.1
5–5.9
0]
0.0
41
0.0
4
Chin
ese
C
heung
et al.
2006
rs
731236
D
(N)
205
Cro
ss-
sect
ional
- -
0.0
37
0
.13
Ja
pa
ne
se
Ka
wa
gu
chi
et al.
2002
rs
731236
D
(N)
60
Clin
ical
popula
tion
- 0
.85
[0.2
6–2.7
7]
- 0.1
1
Japanese
O
ishi
et al.
2003
rs
731236
S
t 85
Case
-contr
ol
CC
-
- 0.3
8
Fin
nis
h
Noponen
et al.
2003
rs
731236
D
(N),
H
300
Clin
ical
popula
tion
CC
-
0.8
81
0.0
48, 0.0
23
0.3
4
Tu
rkis
h
Ese
r
et al.
2010
rs
731236
D
170
Male
tw
in
popula
tion
- -
0.0
03
0
.43
F
inn
ish
V
ide
ma
n
et al.
1998
158

Gene
V
ariatio
n
DD
1
Ph
en
oty
pe
Indiv
idua
ls
(N)
Stu
dy
typ
e2
Ris
k a
llele
O
R
P-v
alu
e
Min
or
alle
le
frequency
Popula
tion
R
efe
rence
rs
73
12
36
E
2
28
C
ross
-
sect
ional
- 0
.9
[0.6
–1.4
]
0.7
7
- F
inn
ish
K
arp
pin
en
et al.
2008
rs
731236
D
220
Popula
tion
study
- -
- 0
.38
D
an
ish
E
sko
la
et al.
2010
# A
ssoc
iatio
n po
sitiv
e in
inte
ract
ion
with
ano
ther
gen
e, a
nthr
opom
etric
or
envi
ronm
enta
l fac
tor.
OR
= O
dds
ratio
, 95
% c
onfid
ence
inte
rval
is g
iven
in [s
quar
e br
acke
ts].
1 D
isc
deg
ener
atio
n p
hen
oty
pes
D=
dis
c si
gnal
cha
nges
usi
ng d
iffer
ent
clas
sific
atio
ns,
S=
disc
her
niat
ion
with
sci
atic
a, N
=no
nspe
cific
dis
coge
nic
pain
with
dis
c
dege
nera
tion,
B=
disc
con
tour
cha
nge;
bul
ge,H
=di
sc c
onto
ur c
hang
e; h
erni
atio
n w
ithou
t sp
ecifi
ed c
linic
al s
ympt
oms,
E=
endp
late
cha
nges
incl
udin
g M
odic
cha
nges
,
U=
unsp
ecifi
ed d
egen
erat
ive
phen
otyp
e,X
= p
lain
rad
iogr
aph,
MR
I for
som
e su
bjec
ts,
St=
sten
osis
, di
sc s
igna
l dec
reas
e al
so r
ecor
ded,∑
=co
mbi
natio
n ph
enot
ype
2 S
tud
y ty
pes
Cas
e-co
ntro
l: cl
inic
al c
ases
vs.
con
trol
s, N
este
d ca
se-c
ontr
ol: c
linic
al c
ases
vs.
mat
ched
con
trol
s, P
opul
atio
n st
udy:
a s
ampl
e of
gen
eral
pop
ulat
ion
or a
subp
opul
atio
n w
ith a
com
mon
cha
ract
eris
tic(s
), C
linic
al p
opul
atio
n: s
ubje
cts
recr
uite
d fr
om e
.g. o
utpa
tient
clin
ic,
Cro
ss-s
ectio
nal:
poss
ibly
mul
tiple
sub
grou
ps o
r
subp
opul
atio
ns a
mon
g su
bjec
ts,
Mal
e tw
in p
opul
atio
n: s
ubpo
pula
tion
of t
he F
inni
sh T
win
Coh
ort
159

160

161
Appendix 2
Genetic association studies in lumbar disc degeneration: a
systematic review
Supporting information
1 Supporting method information 162 1.1 Study inclusion criteria ......................................................................... 162 1.2 Study quality assessment essentials ...................................................... 163 1.3 Data extraction form ............................................................................. 165 1.4 Categories for the credibility of cumulative epidemiological
evidence ................................................................................................ 169 1.5 Protein-protein interaction network analysis methods .......................... 169
2 Supporting results information 170 2.1 Equivalent rs-numbers .......................................................................... 170 2.2 Included studies..................................................................................... 171 2.3 Excluded studies ................................................................................... 175
3 References for Appendix 2 184

162
1 Supporting method information
1.1 Study inclusion criteria
Studies that were considered eligible to be included in the review had the
following features:
– Relevant outcome or disease
– Intervertebral disc changes
– Vertebral endplate changes
– Spondylarthrosis
– Reliable definition of outcome (MRI)
– Number of study subjects 50 or higher
– Human study
– Specific genetic variant is described
Studies considered not eligible (and were therefore excluded from the review) had
one or more of the following features:
– Outcome or disease is not relevant
– Definition of outcome is not reliable (i.e. is other than MRI)
– Number of study subjects is under 50
– No specific genetic variant is described
– Animal study

163
1.2 Study quality assessment essentials
Type of bias Criteria Classification
Selection bias
Major
Selection of study population (inclusion and
exclusion criteria well specified)
Representativeness (response rate,
difference between participants and non-
participants, and control for variables in
case difference found between participants
and non-participants)
Population stratification (confounding by
ethnic origin)
Minor
Awareness of study hypothesis
Possibility of change in the status of a risk
factor as a result of low back pain
No or minor: Defined target population
represents the general population or
subgroup of the general population (e.g.
women or men, certain age group,
geographical area, certain occupational
group) and response rate is above 60%.
Ethnic origin is known. Control
population is well defined and is similar
to affected individuals while unaffected.
Moderate: Defined target population
represents a narrow subgroup of the
general population and response rate is
80 -100%. Selection process or proper
characteristics of control population are
not reported.
Severe: Defined target population
represents a narrow subgroup of the
general population and response rate is
60 - 80%. Control population differs from
affected individuals or no information of
controls is available.
Definite: Ethnic origin of the population
is not known. Study population consists
of "self-selected" volunteers if suspicion
of payment to non-patients. Control
population is of different ethnic origin,
nationality or from great geographical
distance. Controls have other
pathological phenotypes.

164
Type of bias Criteria Classification
Performance bias Major
Validity genetic analyses method
Quality control (genotyping error)
Minor
Blinding of genetic analyses method
towards the outcome
No: Validated method, quality control,
blinding of assessors of exposure
towards the outcome.
Minor: Blinding not reported.
Moderate: Validated method, no quality
control.
Definite: Non-validated genetic analyses
method, no quality control, genotyping
was not blinded towards the outcome.
Detection bias
Major
Clear definition of outcome
Standardized validated method of
assessing outcome
Minor
Blinding of assessors of outcome towards
genetic factors
No or minor: Clear definition of outcome
and standardized method of assessing
outcome.
Moderate: Clear definition of outcome
and not standardized method of
assessing outcome.
Definite: Unclear definition of outcome
or non-systematic assessment of
outcome.
Attrition bias Major
Magnitude of missing data (including
unsuccessful genotyping)
Completeness of follow-up (if applicable)
Where the magnitude of missing data is not
reported, studies are to be considered
having Possible attrition bias.
No: Less than 20% of missing data.
Participation rate 50-100% for follow-up
time < 5 years, or comparison done for
lost to follow-up regarding variables of
interest.
Possible: Missing data between 20%
and 40%. Participation rate 30-50% for
follow-up time < 5 years, and no
comparison done for lost to follow-up
regarding variables of interest and
Definite: More than 40% of missing
data. Participation rate less than 30%
for follow-up time < 5 years or
Statistical analyses -Population stratification has been
addressed
-Hardly-Weinberg equilibrium was
considered
-Methods used for inferring genotypes and
haplotypes are specified and valid
-Multiple comparison has been addressed /
risk of false positive findings was controlled
Each item should be responded as
no/yes/unclear

165
1.3 Data extraction form
Data Extraction Form
DD and Genes – A Systematic
Review
STUDY DETAILS
Title (ID) Scoring (max. 31 points)
First author, year of publication
Year(s) of study
1 point if reported, not the year
of publication
Country of study
Study design
Sponsorship of study
POPULATION
Subjects Clinical General Working
Teenage
1 point if reported
Symptoms (if reported) LBP Sciatica
Asymptomatic
Other;what: Not reported
N of individuals in total N= 1 point if reported and more
than 100 individuals
2 points if reported and more
than 300 individuals
3 points if reported and more
than 600 individuals
4 points if reported and more
than 900 individuals
N of cases Ncase=
N of controls Ncont=
Gender in total population F (%)= M (%)= 1 point if reported in all
subgroups / properly, or if age and
sex matched controls
Gender in cases (fill in the
reported gender)
Fcase (%)= Mcase
(%)=
Gender in controls (fill in the
reported gender)
Fcont (%)= Mcont
(%)=
Age (fill in what is reported) Age range = Mean age
and SD=
Age in cases Age rangecase= Mean age
and SDcase=
1 point if reported in all
subgroups (population study=only
one subgroup)

166
Data Extraction Form
DD and Genes – A Systematic
Review
Age in controls Age rangecont= Mean age
and SDcont=
Ethnicity Caucasian Asian American
African
1 point if reported
Other, specify:
PHENOTYPES
MRI
Field strength (Tesla) Field strength Not reported
1 point if reported
T1/T2 /PD Not reported
Slice orientation Sagittal Axial Coronal
Multi-slice Other Not
reported
Lumbar disk levels imaged L1/L2 L2/L3 L3/L4 L4/L5
L5/S1 Not specified
1 point if all lumbar levels
Other disk levels 1 point if other levels analysed
separately
Slice thickness (mm.) Not reported
Other specific technical data on
MRI
Not reported
Number of MRI readers N=
Qualification of reader(s) Specialist= Specialist
(radiology)=
Specialist(surgery)=
Researcher= Student= Not
reported
Other:
Reader blinded to genetic data? Yes No Not reported 1 point if ”Yes”
Reproductibility tested? Yes No Not reported 1 point if ”Yes”
Intra/inter-tester Intra Inter Not reported
Results of reproductilibity test
(methods, results: Kappa, ICC or
percentage; i.e. results written
open)

167
Data Extraction Form
DD and Genes – A Systematic
Review
Degeneration defined by (mark all
that fit)
Disk space narrowing / Disk height reduction
Signal intensity loss Presence of fissures (AT)
HIZ lesions Bulging Protrusion Ligamentous signal changes
Endplate signal changes Osteophytosis
Vertebral malalignment Foraminal stenosis
Central canal stenosis Nerve root displacement
Swelling Dura impression / spinal cord affected
Standardized definition of
degeneration
GENOTYPES AND OTHER
EXPOSURES
Reference 2 points if reported and
definition is well established
before
Genotyping methods
Blinding of researchers in
genotyping (to degeneration data)
No Yes Not reported
Genes
Polymorphisms (Report number if multiple
variations)
rs numbers (if reported) (Report number if multiple
variations)
Other exposures (e.g. smoking,
BMI, height, physical workload)
QUALITY ASSESMENT
Selection bias No or Minor Moderate
Severe
Definite
(3-0 points) 3 if No or
Minor, 2 Moderate, 1 Severe, 0
Definite
Performance bias No Minor Moderate Definite
(2-0 points) 3 = ”No”,
2=”Minor” 1= ”Moderate”, 0
= ”Definite”
Detection bias No or Minor Moderate
Definite
(2-0 points) 2 = ”No or
Minor”, 1 = ”Moderate”, 0
= ”Definite”
Attrition bias No Possible Definite (2-0 points) 2 = ”No”, 1
= ”Possible” , 0 = ”Definite”
Statistical analyses (1 points each if ”Adequate”, max.
4 points)
Population stratification Adequate Poor Unclear
HWE considered Adequate Poor Unclear

168
Data Extraction Form
DD and Genes – A Systematic
Review
Methods used for inferring
genotypes and haplotypes
specified and valid
Adequate Poor Unclear
Multiple comparison has been
addressed / risk of false positive
findings was controlled
Adequate Poor Unclear
Other exposure issue considered
in analyses
No Yes Not reported 1 point if ”Yes”
FINDINGS Genotype or haplotype or
combined genotypes
Phenotype Genotype (data for cases and
controls, as presented in the
study)
Measures of association (e.g. ORs
and 95% CI)
Interactions
Interacting phenotypes Genotype (data for cases and
controls, as presented in the
study)
and
Measures of association (e.g. ORs
and 95% CI)
SOURCE OF DATA Publication Online database
From correspondence with
authors

169
1.4 Categories for the credibility of cumulative epidemiological evidence
Supporting information Figure 1. Categories for the credibility of cumulative
epidemiological evidence. The three letters correspond (in order) to amount of
evidence, replication and protection from bias. Evidence is categorized as strong,
when there is A for all three items, and is categorized as weak when there is a C for
any of the three items. All other combinations are categorized as moderate. Modified
from Ioannidis 2008 et al. with permission.
1.5 Protein-protein interaction network analysis methods
The PPI-network was created with InWeb 2.0 (Lage et al. 2007) using all genes
with a positive association to lumbar disc degeneration as input.
InWeb is based on mining of online interactiondatabses String (Jensen et al.
2009), MINT (Zanzoni et al. 2002), Bind (Bader et al. 2003), DIP (Xenarios et al.
2002), GRID (Breitkreutz et al. 2003), HPRD (Keshava Prasad et al. 2009), Kegg
(Kanehisa et al. 2010), Reactome (Joshi-Tope et al. 2005) and IntAct (Aranda et
al. 2010). All experimentally based interactions from each was kept and given a
score between 0 and 1 (Lage et al. 2007). We used all interactions with a score on
0.5 or higher. To avoid highly interacting proteins (hubs) we used a network score
on 0.1, which means all linker-protein (protein not in input) at most is allowed to
interact with 9 other linker-proteins for each input protein they interact with (Lage
et al. 2007). P-values was calculated creating 10.000 random networks.
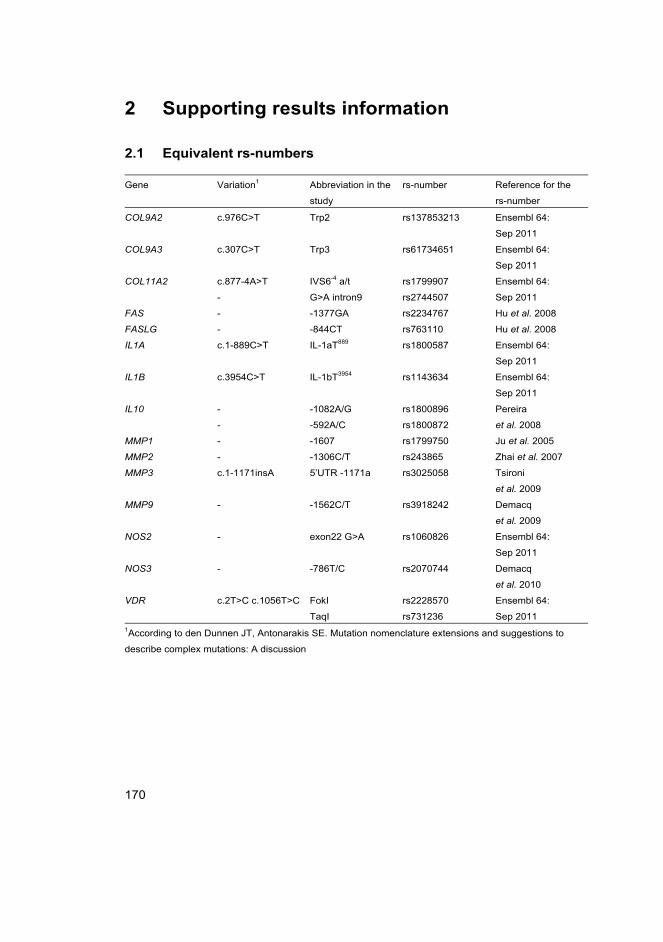
170
2 Supporting results information
2.1 Equivalent rs-numbers
Gene Variation1 Abbreviation in the
study
rs-number Reference for the
rs-number
COL9A2 c.976C>T Trp2 rs137853213 Ensembl 64:
Sep 2011
COL9A3 c.307C>T Trp3 rs61734651 Ensembl 64:
Sep 2011
COL11A2 c.877-4A>T
-
IVS6-4 a/t
G>A intron9
rs1799907
rs2744507
Ensembl 64:
Sep 2011
FAS - -1377GA rs2234767 Hu et al. 2008
FASLG - -844CT rs763110 Hu et al. 2008
IL1A c.1-889C>T IL-1aT889 rs1800587 Ensembl 64:
Sep 2011
IL1B c.3954C>T IL-1bT3954 rs1143634 Ensembl 64:
Sep 2011
IL10 -
-
-1082A/G
-592A/C
rs1800896
rs1800872
Pereira
et al. 2008
MMP1 - -1607 rs1799750 Ju et al. 2005
MMP2 - -1306C/T rs243865 Zhai et al. 2007
MMP3 c.1-1171insA 5’UTR -1171a rs3025058 Tsironi
et al. 2009
MMP9 - -1562C/T rs3918242 Demacq
et al. 2009
NOS2 - exon22 G>A rs1060826 Ensembl 64:
Sep 2011
NOS3 - -786T/C rs2070744 Demacq
et al. 2010
VDR c.2T>C c.1056T>C FokI
TaqI
rs2228570
rs731236
Ensembl 64:
Sep 2011 1According to den Dunnen JT, Antonarakis SE. Mutation nomenclature extensions and suggestions to
describe complex mutations: A discussion

171
2.2 Included studies
Studies included in the systematic review (N=52)
Annunen S, Paassilta P, Lohiniva J, Perala M, Pihlajamaa T, Karppinen J, Tervonen O, Kroger H,
Lahde S, Vanharanta H, et al. 1999. An allele of COL9A2 associated with intervertebral disc disease.
Science 285(5426):409-12.
Bei T, Tilkeridis C, Garantziotis S, Boikos S, Kazakos K, Simopoulos C, Stratakis C. 2008. A novel,
non-functional, COL1A1 polymorphism is not associated with lumbar disk disease in young male Greek
subjects unlike that of the Sp1 site. Hormones (Athens) 7(3):251-4.
Cheung KMC, Chan D, Karppinen J, Chen Y, Jim JJT, Yip S, Ott J, Wong KK, Sham P, Luk KDK, et
al. 2006. Association of the taq I allele in vitamin D receptor with degenerative disc disease and disc bulge
in a Chinese population. Spine (Phila Pa 1976) 31(10):1143-8.
Cong L, Pang H, Xuan D, Tu GJ. 2010a. Association between the expression of aggrecan and the
distribution of aggrecan gene variable number of tandem repeats with symptomatic lumbar disc herniation
in Chinese Han of northern China. Spine (Phila Pa 1976) 35(14):1371-6.
Cong L, Pang H, Xuan D, Tu G. 2010b. The interaction between aggrecan gene VNTR
polymorphism and cigarette smoking in predicting incident symptomatic intervertebral disc degeneration.
Connect Tissue Res 51(5):397-403.
Dong DM, Yao M, Liu B, Sun CY, Jiang YQ, Wang YS. 2007. Association between the -1306C/T
polymorphism of matrix metalloproteinase-2 gene and lumbar disc disease in Chinese young adults. Eur
Spine J 16(11):1958-61.
Eser B, Cora T, Eser O, Kalkan E, Haktanir A, Erdogan MO, Solak M. 2010. Association of the
polymorphisms of vitamin D receptor and aggrecan genes with degenerative disc disease. Genet Test Mol
Biomarkers 14(3):313-7.
Eser O, Eser B, Cosar M, Erdogan MO, Aslan A, Yıldız H, Solak M, Haktanır A. 2011. Short
aggrecan gene repetitive alleles associated with lumbar degenerative disc disease in Turkish patients.
Genet Mol Res 10(3):1923-30.
Eskola PJ, Kjaer P, Daavittila IM, Solovieva S, Okuloff A, Sorensen JS, Wedderkopp N, Ala-Kokko L,
Mannikko M, Karppinen JI. 2010. Genetic risk factors of disc degeneration among 12-14-year-old Danish
children: A population study Int J Mol Epidemiol Genet 1(2):158-65.
Higashino K, Matsui Y, Yagi S, Takata Y, Goto T, Sakai T, Katoh S, Yasui N. 2007. The alpha2 type
IX collagen tryptophan polymorphism is associated with the severity of disc degeneration in younger
patients with herniated nucleus pulposus of the lumbar spine. Int Orthop 31(1):107-11.
Hirose Y, Chiba K, Karasugi T, Nakajima M, Kawaguchi Y, Mikami Y, Furuichi T, Mio F, Miyake A,
Miyamoto T, et al. 2008. A functional polymorphism in THBS2 that affects alternative splicing and MMP
binding is associated with lumbar-disc herniation. Am J Hum Genet 82(5):1122-9.

172
Jim JJ, Noponen-Hietala N, Cheung KM, Ott J, Karppinen J, Sahraravand A, Luk KD, Yip SP, Sham
PC, Song YQ, et al. 2005. The TRP2 allele of COL9A2 is an age-dependent risk factor for the
development and severity of intervertebral disc degeneration. Spine (Phila Pa 1976) 30(24):2735-42.
Kales SN, Linos A, Chatzis C, Sai Y, Halla M, Nasioulas G, Christiani DC. 2004. The role of collagen
IX tryptophan polymorphisms in symptomatic intervertebral disc disease in southern European patients.
Spine (Phila Pa 1976) 29(11):1266-70.
Karasugi T, Semba K, Hirose Y, Kelempisioti A, Nakajima M, Miyake A, Furuichi T, Kawaguchi Y,
Mikami Y, Chiba K, et al. 2009. Association of the tag SNPs in the human SKT gene (KIAA1217) with
lumbar disc herniation. J Bone Miner Res 24(9):1537-43.
Karppinen J, Solovieva S, Luoma K, Raininko R, Leino-Arjas P, Riihimaki H. 2009. Modic changes
and interleukin 1 gene locus polymorphisms in occupational cohort of middle-aged men. Eur Spine J
18(12):1963-70.
Karppinen J, Paakko E, Raina S, Tervonen O, Kurunlahti M, Nieminen P, Ala-Kokko L, Malmivaara A,
Vanharanta H. 2002. Magnetic resonance imaging findings in relation to the COL9A2 tryptophan allele
among patients with sciatica. Spine (Phila Pa 1976) 27(1):78-83.
Karppinen J, Paakko E, Paassilta P, Lohiniva J, Kurunlahti M, Tervonen O, Nieminen P, Goring HHH,
Malmivaara A, Vanharanta H, et al. 2003. Radiologic phenotypes in lumbar MR imaging for a gene defect
in the COL9A3 gene of type IX collagen. Radiology 227(1):143-8.
Karppinen J, Daavittila I, Solovieva S, Kuisma M, Taimela S, Natri A, Haapea M, Korpelainen R,
Niinimaki J, Tervonen O, et al. 2008. Genetic factors are associated with modic changes in endplates of
lumbar vertebral bodies. Spine (Phila Pa 1976) 33(11):1236-41.
Kawaguchi Y, Osada R, Kanamori M, Ishihara H, Ohmori K, Matsui H, Kimura T. 1999. Association
between an aggrecan gene polymorphism and lumbar disc degeneration. Spine (Phila Pa 1976)
24(23):2456-60.
Kawaguchi Y, Kanamori M, Ishihara H, Ohmori K, Matsui H, Kimura T. 2002. The association of
lumbar disc disease with vitamin-D receptor gene polymorphism. J Bone Joint Surg Am 84-A(11):2022-8.
Kelempisioti A, Eskola P et al. 2011. Genetic susceptibility of intervertebral disc degeneration among
young finnish adults . BMC Med Genet 12(1):153.
Kim D, Lee S, Kim K, Yu S. 2010. Association of interleukin-1 receptor antagonist gene
polymorphism with response to conservative treatment of lumbar herniated nucleus pulposus. Spine (Phila
Pa 1976) 35(16):1527-31.
Kim NK, Shin DA, Han IB, Yoo EH, Kim SH, Chung SS. 2011. The association of aggrecan gene
polymorphism with the risk of intervertebral disc degeneration. Acta Neurochir 153(1):129-33.
Lin WP, Lin JH, Chen XW, Wu CY, Zhang LQ, Huang ZD, Lai JM. 2011. Interleukin-10 promoter
polymorphisms associated with susceptibility to lumbar disc degeneration in a chinese cohort. Genet Mol
Res 10(3):1719-27.

173
Mashayekhi F, Shafiee G, Kazemi M, Dolati P. 2010. Lumbar disk degeneration disease and
aggrecan gene polymorphism in northern Iran. Biochem Genet 48(7-8):684-9.
Min SK, Nakazato K, Okada T, Ochi E, Hiranuma K. 2009. The cartilage intermediate layer protein
gene is associated with lumbar disc degeneration in collegiate judokas. Int J Sports Med 30(9):691-4.
Min S, Nakazato K, Yamamoto Y, Gushiken K, Fujimoto H, Fujishiro H, Kobayakawa Y, Hiranuma K.
2010. Cartilage intermediate layer protein gene is associated with lumbar disc degeneration in male, but
not female, collegiate athletes. Am J Sports Med 38(12):2552-7.
Mio F, Chiba K, Hirose Y, Kawaguchi Y, Mikami Y, Oya T, Mori M, Kamata M, Matsumoto M, Ozaki
K, et al. 2007. A functional polymorphism in COL11A1, which encodes the alpha 1 chain of type XI
collagen, is associated with susceptibility to lumbar disc herniation. Am J Hum Genet 81(6):1271-7.
Noponen-Hietala N, Kyllonen E, Mannikko M, Ilkko E, Karppinen J, Ott J, Ala-Kokko L. 2003.
Sequence variations in the collagen IX and XI genes are associated with degenerative lumbar spinal
stenosis. Ann Rheum Dis 62(12):1208-14.
Noponen-Hietala N, Virtanen I, Karttunen R, Schwenke S, Jakkula E, Li H, Merikivi R, Barral S, Ott J,
Karppinen J, et al. 2005. Genetic variations in IL6 associate with intervertebral disc disease characterized
by sciatica. Pain 114(1-2):186-94.
Nunes, F.T.B.b , Conforti-Froes, N.D.T.a b c , Negrelli, W.F.e , Souza,D.R.S.b d. 2007. Genetic and
environmental factors involved on intervertebral disc degeneration. Acta Ortopedica Brasileira 15(1):9-13.
Oishi Y, Shimizu K, Katoh T, Nakao H, Yamaura M, Furuko T, Narusawa K, Nakamura T. 2003. Lack
of association between lumbar disc degeneration and osteophyte formation in elderly Japanese women
with back pain. Bone 32(4):405-11.
Paassilta P, Lohiniva J, Goring HH, Perala M, Raina SS, Karppinen J, Hakala M, Palm T, Kroger H,
Kaitila I, et al. 2001. Identification of a novel common genetic risk factor for lumbar disk disease. JAMA
285(14):1843-9.
Paz Aparicio J, López-Anglada Fernández E, Montes Prieto H, Pena Vázquez J, Fernández Bances
I, Folgueras Henriksen V, López Fernández P, López-Fanjúl Menéndez JC, Valle Garay E, Asensi Álvarez
V. 2010. Association between the genetic polymorphism of interleukin-1β (3953 T/C) and symptomatic
lumbar herniated disc. Revista Espanola De Cirugia Ortopedica y Traumatologia 54(4):227-33.
Roughley P, Martens D, Rantakokko J, Alini M, Mwale F, Antoniou J. 2006. The involvement of
aggrecan polymorphism in degeneration of human intervertebral disc and articular cartilage. Eur Cell
Mater 11:1-7.
Seki S, Kawaguchi Y, Chiba K, Mikami Y, Kizawa H, Oya T, Mio F, Mori M, Miyamoto Y, Masuda I,
et al. 2005. A functional SNP in CILP, encoding cartilage intermediate layer protein, is associated with
susceptibility to lumbar disc disease. Nat Genet 37(6):607-12.
Seki S, Kawaguchi Y, Mori M, Mio F, Chiba K, Mikami Y, Tsunoda T, Kubo T, Toyama Y, Kimura T,
et al. 2006. Association study of COL9A2 with lumbar disc disease in the Japanese population. J Hum
Genet 51(12):1063-7.

174
Solovieva S, Kouhia S, Leino-Arjas P, Ala-Kokko L, Luoma K, Raininko R, Saarela J, Riihimaki H.
2004. Interleukin 1 polymorphisms and intervertebral disc degeneration. Epidemiology 15(5):626-33.
Solovieva S, Lohiniva J, Leino-Arjas P, Raininko R, Luoma K, Ala-Kokko L, Riihimaki H. 2006.
Intervertebral disc degeneration in relation to the COL9A3 and the IL-1ss gene polymorphisms. Eur Spine
J 15(5):613-9.
Solovieva S, Lohiniva J, Leino-Arjas P, Raininko R, Luoma K, Ala-Kokko L, Riihimaki H. 2002.
COL9A3 gene polymorphism and obesity in intervertebral disc degeneration of the lumbar spine: Evidence
of gene-environment interaction. Spine (Phila Pa 1976) 27(23):2691-6.
Solovieva S, Noponen N, Mannikko M, Leino-Arjas P, Luoma K, Raininko R, Ala-Kokko L, Riihimaki
H. 2007. Association between the aggrecan gene variable number of tandem repeats polymorphism and
intervertebral disc degeneration. Spine (Phila Pa 1976) 32(16):1700-5.
Song Y, Ho DWH, Karppinen J, Kao PYP, Fan B, Luk KDK, Yip S, Leong JCY, Cheah KSE, Sham P,
et al. 2008a. Association between promoter -1607 polymorphism of MMP1 and lumbar disc disease in
southern Chinese. BMC Med Genet 9:38-.
Song Y, Cheung KMC, Ho DWH, Poon SCS, Chiba K, Kawaguchi Y, Hirose Y, Alini M, Grad S, Yee
AFY, et al. 2008b. Association of the asporin D14 allele with lumbar-disc degeneration in Asians. Am J
Hum Genet 82(3):744-7.
Sun ZM, Miao L, Zhang YG, Ming L. 2009. Association between the -1562 C/T polymorphism of
matrix metalloproteinase-9 gene and lumbar disc disease in the young adult population in north China.
Connect Tissue Res 50(3):181-5.
Sun Z, Ling M, Huo Y, Chang Y, Li Y, Qin H, Yang G, Lucas R. 2011. Caspase 9 gene
polymorphism and susceptibility to lumbar disc disease in the Han population in northern China. Connect
Tissue Res 52(3):198-202.
Takahashi M, Haro H, Wakabayashi Y, Kawa-uchi T, Komori H, Shinomiya K. 2001. The association
of degeneration of the intervertebral disc with 5a/6a polymorphism in the promoter of the human matrix
metalloproteinase-3 gene. J Bone Joint Surg Br 83(4):491-5.
Videman T, Leppavuori J, Kaprio J, Battie MC, Gibbons LE, Peltonen L, Koskenvuo M. 1998.
Intragenic polymorphisms of the vitamin D receptor gene associated with intervertebral disc degeneration.
Spine (Phila Pa 1976) 23(23):2477-85.
Videman T, Gibbons LE, Battie MC, Maravilla K, Vanninen E, Leppavuori J, Kaprio J, Peltonen L.
2001. The relative roles of intragenic polymorphisms of the vitamin d receptor gene in lumbar spine
degeneration and bone density. Spine (Phila Pa 1976) 26(3):E7-E12.
Videman T, Saarela J, Kaprio J, Nakki A, Levalahti E, Gill K, Peltonen L, Battie MC. 2009.
Associations of 25 structural, degradative, and inflammatory candidate genes with lumbar disc desiccation,
bulging, and height narrowing. Arthritis Rheum 60(2):470-81.

175
Virtanen IM, Song YQ, Cheung KMC, Ala-Kokko L, Karppinen J, Ho DWH, Luk KDK, Yip SP, Leong
JCY, Cheah KSE, et al. 2007. Phenotypic and population differences in the association between CILP and
lumbar disc disease. J Med Genet 44(4):285-8.
Williams FM, Popham M, Hart DJ, de Schepper E, Bierma-Zeinstra S, Hofman A, Uitterlinden AG,
Arden NK, Cooper C, Spector TD, et al. 2011. GDF5 single-nucleotide polymorphism rs143383 is
associated with lumbar disc degeneration in northern European women. Arthritis Rheum 63(3):708-12.
Zhu G, Jiang X, Xia C, Sun Y, Zeng Q, Wu X, Li X. 2011. Association of FAS and FAS ligand
polymorphisms with the susceptibility and severity of lumbar disc degeneration in Chinese Han population.
Biomarkers 16(6):485-90.
2.3 Excluded studies
Systematic search hits excluded after dual examination of abstracts
and/or full text articles (N=72)
-the reason for exclusion
Genetic predispositions to low back pain. (2005) Medecine/Sciences 21(10): 820.
-review article
Aladin DMK, Cheung KMC, Chan D, Yee AFY, Jim JJT, Luk KDK & Lu WW (2007) Expression of the Trp2
allele of COL9A2 is associated with alterations in the mechanical properties of human intervertebral discs.
Spine (Phila Pa 1976) 32(25): 2820-2826.
-number of study subjects is less than 50
Aladin DMK, Cheung KMC, Ngan AHW, Chan D, Leung VYL, Lim CT, Luk KDK & Lu WW (2010)
Nanostructure of Collagen Fibrils in Human Nucleus Pulposus and Its Correlation with Macroscale Tissue
Mechanics. J Orthop Res 28(4): 497-502.
-no genetic variant described
Ala-Kokko L (2002) Genetic risk factors for lumbar disc disease. Ann Med 34(1): 42-7.
-review article
Anderson JA, Ng JJ, Bowe C, Mcdonald C, Richman DP, Wollmann RL & Maselli RA (2008) Variable
phenotypes associated with mutations in DOK7. Muscle Nerve 37(4): 448-456.
-not a relevant outcome
Aulisa L, Papaleo P, Pola E, Angelini F, Aulisa AG, Tamburrelli FC, Pola P & Logroscino CA (2007)
Association between IL-6 and MMP-3 gene polymorphisms and adolescent idiopathic scoliosis: A case-
control study. Spine (Phila Pa 1976) 32(24): 2700-2702.

176
-not a relevant outcome
Battié MC & Videman T (2006) Lumbar disc degeneration: Epidemiology and genetics. J Bone Joint Surg
Am 88: 3-9.
-review article
Battié MC, Videman T, Kaprio J, Gibbons LE, Gill K, Manninen H, Saarela J & Peltonen L (2009) The Twin
Spine Study: Contributions to a changing view of disc degeneration. Spine J 9(1): 47-59.
-review article
Behrens A, Haigh J, Mechta-Grigoriou F, Nagy A, Yaniv M & Wagner EF (2003) Impaired intervertebral
disc formation in the absence of Jun. Development 130(1): 103-109.
-no genetic variant described, no relevant outcome, no MRI
Bei T, Tilkeridis C, Garatziotis S, Kortesas E & Stratakis C (2002) COL1A1 polymorphism (a four-base
pair insertion polymorphism in 3'UTR) in young male Greek army recruits with lumbar disk disease. Am J
Hum Genet (71)(4, Suppl. S): 1739.
-no MRI, only abstract, nro of study subjects is unclear
Bell N, Morrison N, Nguyen T, Eisman J & Hollis B (2001) ApaI polymorphisms of the vitamin D receptor
predict bone density of the lumbar spine and not racial difference in bone density in young men. J Lab Clin
Med 137(2): 133-140.
-not a relevant outcome
Bollettino A, Buzas B, Atlas S, Wu T, Belfer I, Hipp H, McKnight C, Scully M, Kingman A, Keller R,
Goldman D & Max M (2005) Association of interleukin-10 and interleukin-13 gene polymorphisms with
persistent sciatica following lumbar diskectomy. J Pain 6(8): 559.
-not a relevant outcome
Burkemper K & Garris D (2006) Influences of obese (ob/ob) and diabetes (db/db) genotype mutations on
lumber vertebral radiological and morphometric indices: Skeletal deformation associated with
dysregulated systemic glucometabolism. BMC Musc Disord (7): 10
-not a relevant outcome, no MRI
Buttle DJ (2007) Factors controlling matrix turnover in health and disease. Biochem Soc Trans 35(4): 643-
646.
-review article
Chan D, Song Y, Sham P & Cheung KMC (2006) Genetics of disc degeneration. Eur Spine J 15 (Suppl 3):
S317-25.
-review article

177
Cheung KMC & Al Ghazi S (2008) (i) Current understanding of low back pain and intervertebral disc
degeneration: epidemiological perspectives and phenotypes for genetic studies. Curr Orthopaed 22(4):
237-244.
-review article
Cheung KMC, Karppinen J, Chan D, Ho DWH, Song Y, Sham P, Cheah KSE, Leong JCY & Luk KDK
(2009) Prevalence and Pattern of Lumbar Magnetic Resonance Imaging Changes in a Population Study of
One Thousand Forty-Three Individuals. Spine 34(9): 934-940.
-no genetic variant described
Colombini A, Lombardi G, Corsi MM & Banfi G (2008) Pathophysiology of the human intervertebral disc.
Int J Biochem Cell B 40(5): 837-842.
-review article
Eisenstein S & Roberts S (2003) The physiology of the disc and its clinical relevance. J Bone Joint Surg
Br 85B(5): 633-636.
-review article
Eun I, Park WW, Suh KT, Il Kim J & Lee JS (2009) Association between osteoprotegerin gene
polymorphism and bone mineral density in patients with adolescent idiopathic scoliosis. Eur Spine J
18(12): 1936-1940.-not a relevant outcome
Eyre DR, Matsui Y & Wu J- (2002) Collagen polymorphisms of the intervertebral disc. Biochem Soc Trans
30(6): 844-848.
-review article
Fink JK (2006) Genetics of common neurologic diseases. Semin Neurol 26(5): 463.
-review article
Freemont AJ (2009) The cellular pathobiology of the degenerate intervertebral disc and discogenic back
pain. Rheumatology 48(1): 5-10.
-review article
Gruber HE, Hoelscher G, Ingram JA, Chow Y, Loeffler B & Hanley EN (2008) 1,25(OH)2-vitamin D3
inhibits and angiogenin by human annulus cells in vitro. Spine (Phila Pa 1976) 33(7): 755-765.
-no genetic variant described
Gruber HE, Ingram JA, Hoelscher GL, Zinchenko N, Hanley EN,Jr & Sun Y (2009) Asporin, a susceptibility
gene in osteoarthritis, is expressed at higher levels in the more degenerate human intervertebral disc.
Arthritis Res Ther 11(2): R47.
-no genetic variant described

178
Hipp H, Scully M, Belfer I, Atlas S, Wu T, Buzas B, McKnight C, Bollettino A, Kingman A, Keller R,
Goldman D & Max M (2005) Association of interleukin-1 beta polymorphisms with persistent sciatica
following lumbar diskectomy. J Pain 6(8): 559.
-MRI has not been mentioned, phenotype definition is unclear
Jones G, White C, Sambrook P & Eisman J (1998}) Allelic variation in the vitamin D receptor, lifestyle
factors and lumbar spinal degenerative disease. Ann Rheum Dis 57(2): 94-99.
-no MRI, only radiographs
Jordan K, Syddall H, Dennison E, Cooper C & Arden N (2005) Birthweight, vitamin D receptor gene
polymorphism, and risk of lumbar spine osteoarthritis. J Rheumatol 32 (4): 678-683.
-no MRI, only radiographs
Kalichman L & Hunter DJ (2008) The genetics of intervertebral disc degeneration.Associated genes. Joint
Bone Spine 75(4): 388-396.
-review article
Kawaguchi Y, Furushima K, Sugimori K, Inoue I & Kimura T (2003) Association between polymorphism of
the transforming growth factor-beta 1 gene with the radiologic characteristic and ossification of the
posterior longitudinal ligament. Spine (Phila Pa 1976) 28 (13): 1424-1426.
-not a relevant outcome
Kim J, Ku S, Lim K, Jee B, Suh C, Kim S, Choi Y & Moon S (2005) Cytokine production by whole blood
cells: Relationship to interleukin gene polymorphism and bone mass. J Korean Med Sci 20(6): 1017-1022.
-not a relevant outcome
Knoeringer M, Reinke A, Trappe A & Schlegel J (2008) Absence of the mutated Trp2 allele but a common
polymorphism of the COL9A2 collagen gene is associated with early recurrence after lumbar discectomy
in a German population. Eur Spine J 17(3): 463-7.
-not a relevant outcome, success of surgery
Koshizuka Y, Ogata N, Shiraki M, Hosoi T, Seichi A, Takeshita K, Nakamura K & Kawaguchi H (2006)
Distinct association of gene polymorphisms of estrogen receptor and vitamin D receptor with lumbar
spondylosis in post-menopausal women. Eur Spine J 15(10): 1521-1528.
-not a relevant outcome, no MRI
Le Maitre CL, Hoyland JA & Freemont AJ (2007) Interleukin-1 receptor antagonist delivered directly and
by gene therapy inhibits matrix degradation in the intact degenerate human intervertebral disc: an in situ
zymographic and gene therapy study. Arth Res Ther 9(4).
-no genetic variant described, no relevant outcome of the disease, no MRI, nro of
study subjects < 50

179
Liu Z, Tang NLS, Cao X-, Liu W-, Qiu X-, Cheng JCY & Qiu Y (2010) Lack of association between the
promoter polymorphisms of MMP-3 and IL-6 genes and adolescent idiopathic scoliosis: A case-control
study in a Chinese Han population. Spine (Phila Pa 1976) 35(18):1701-5.
-not a relevant outcome
Matsui Y (2006) Type IX collagen diseases. Clinical calcium 16(11): 1894-1898.
-review article
Matsui Y, Mirza SK, Wu J-, Carter B, Bellabarba C, Shaffrey CI, Chapman JR & Eyre DR (2004) The
association of lumbar spondylolisthesis with collagen IX tryptophan alleles. J Bone Joint Surg Br 86(7):
1021-1026.
-not a relevant outcome, unclear whether MRI has been done or not
Matsui Y, Wu J-, Weis MA, Pietka T & Eyre DR (2003) Matrix deposition of tryptophan-containing allelic
variants of type IX collagen in developing human cartilage. Matrix Biol 22(2): 123-129.
-no MRI
Min JL, Meulenbelt I, Riyazi N, Kloppenburg M, Houwing-Duistermaat JJ, Seymour AB, van Duijn CM &
Slagboom PE (2006) Association of matrilin-3 polymorphisms with spinal disc degeneration and
osteoarthritis of the first carpometacarpal joint of the hand. Ann Rheum Dis 65(8): 1060-1066.
-no MRI
Min JL, Meulenbelt I, Kloppenburg M, van Duijn CM & Slagboom PE (2007) Mutation analysis of
candidate genes within the 2q33.3 linkage area for familial early-onset generalised osteoarthritis. Eur J
Hum Genet 15(7): 791-799.
-not a relevant outcome, OA
Modic MT (2007) Degenerative disc disease: genotyping, MR imaging and phenotyping. Skeletal Radiol
36(2): 91-3.
-review article
Nakki A, Videman T, Kujala UM, Suhonen M, Mannikko M, Peltonen L, Battie MC, Kaprio J & Saarela J
(2011) Candidate gene association study of magnetic resonance imaging-based hip osteoarthritis (OA):
evidence for COL9A2 gene as a common predisposing factor for hip OA and lumbar disc degeneration. J
Rheumatol 38(4): 747-752.
-not a relevant outcome, OA
Pluijm S, van Essen H, Bravenboer N, Uitterlinden A, Smit J, Pols H & Lips P (2004) Collagen type I alpha
1 Sp1 polymorphism, osteoporosis, and intervertebral disc degeneration in older men and women. Ann
Rheum Dis 63(1): 71-77.
-no MRI

180
Podichetty VK (2007) The aging spine: The role of inflammatory mediators in intervertebral disc
degeneration. Cell Mol Biol 53(5): 4-18.
-review article
Rannou F, Revel M & Poiraudeau S (2003) Is degenerative disk disease genetically determined? Joint
Bone Spine 70(1): 3-5.
-review article
Reimanna F, Cox JJ, Belfer I, Diatchenko L, Zaykin DV, McHale DP, Drenth JPH, Dai F, Wheeler J,
Sanders F, Wood L, Wu T-, Karppinen J, Nikolajsen L, Männikkö M, Max MB, Kiselyczny C, Poddar M, Te
Morsche RHM, Smith S, Gibson D, Kelempisioti A, Maixner W, Gribble FM & Woods CG (2010) Pain
perception is altered by a nucleotide polymorphism in SCN9A. Proc Natl Acad Sci USA 107(11): 5148-
5153.
-not a relevant outcome, pain
Revel M (2006) The concept of discolysis in intervertebral disk disease. Acta Reumatol Port 31(2): 133-
140.
-review article
Ryder JJ, Garrison K, Song F, Hooper L, Skinner J, Loke Y, Loughlin J, Higgins JPT & MacGregor AJ
(2008) Genetic associations in peripheral joint osteoarthritis and spinal degenerative disease: a systematic
review Ann Rheum Dis 67(5): 584-591.
-review article
Sakai Y, Matsuyama Y, Hasegawa Y, Yoshihara H, Nakamura H, Katayama Y, Imagama S, Ito Z, Ishiguro
N & Hamajima N (2007) Association of gene polymorphisms with intervertebral disc degeneration and
vertebral osteophyte formation. Spine (Phila Pa 1976) 32(12): 1279-86.
-no MRI
Semba K, Araki K, Li Z, Matsumoto K-, Suzuki M, Nakagata N, Takagi K, Takeya M, Yoshinobu K, Araki M,
Imai K, Abe K & Yamamura K- (2006) A novel murine gene, Sickle tail, linked to the Danforth's short tail
locus, is required for normal development of the intervertebral disc. Genetics 172(1): 445-456.
-no genetic variant described, no MRI, animal study
Song HF, Wu ZH, Fei Q, Yan JZ, Liu Z, Zhang JG, Li SG & Qiu GX (2010) Association study of Trp2 allele
polymorphism with degenerative disc disease in a Chinese Han nationality. Chung-Hua I Hsueh Tsa Chih
90(3): 148-152.
-unclear whether MRI has been done or not, only abstract, in Chinese;
translation not available
Tang Y, Yuan H-, Wang Z-, Xu J- & Lei L (2007) The genetic polymorphisms of MMP-3 and VDR on
susceptibility of lumbar disc degeneration. Fudan University Journal of Medical Sciences 34(1): 37-41.

181
-only abstract, in Chinese; translation not available
Tegeder I (2009) Current evidence for a modulation of low back pain by human genetic variants. J Cell
Mol Med 13(8): 1605-1619.
-review article
Tilkeridis C, Bei T, Garantziotis S & Stratakis CA (2005) Association of a COL1A1 polymorphism with
lumbar disc disease in young military recruits. J Med Genet. 42(7).
-number of study subjects under 50, no MRI
Urano T, Narusawa K, Kobayashi S, Shiraki M, Horie-Inoue K, Sasaki N, Hosoi T, Ouchi Y, Nakamura T &
Inoue S (2010) Association of HTRA1 promoter polymorphism with spinal disc degeneration in Japanese
women. J Bone Miner Metab 28(2): 220-6.
-no MRI
Urano T, Narusawa K, Shiraki M, Sasaki N, Hosoi T, Ouchi Y, Nakamura T & Inoue S (2011) Single-
nucleotide polymorphism in the hyaluronan and proteoglycan link protein 1 (HAPLN1) gene is associated
with spinal osteophyte formation and disc degeneration in Japanese women. Eur Spine J 20(4): 572-577.
-no MRI
Urano T, Narusawa K, Shiraki M, Usui T, Sasaki N, Hosoi T, Ouchi Y, Nakamura T & Inoue S (2007)
Association of a single nucleotide polymorphism in the WISP1 gene with spinal osteoarthritis in
postmenopausal Japanese women. J Bone Miner Metab 25(4): 253-8.
-no MRI
Urano T, Narusawa K, Shiraki M, Usui T, Sasaki N, Hosoi T, Ouchi Y, Nakamura T & Inoue S (2008)
Association of a single nucleotide polymorphism in the insulin-like growth factor-1 receptor gene with
spinal disc degeneration in postmenopausal Japanese women. Spine (Phila Pa 1976) 33(11): 1256-61.
-no MRI
Urano T, Shiraki M, Narusawa K, Usui T, Sasaki N, Hosoi T, Ouchi Y, Nakamura T & Inoue S (2007)
Q89R polymorphism in the LDL receptor-related protein 5 gene is associated with spinal osteoarthritis in
postmenopausal Japanese women. Spine (Phila Pa 1976) 32(1): 25-9.
-no MRI
Urano T, Usui T, Shiraki M, Ouchi Y & Inoue S (2009) Association of a single nucleotide polymorphism in
the constitutive androstane receptor gene with bone mineral density. Geriatr Gerontol Int 9(3): 235-241.
-not a relevant outcome
Valdes AM, Hassett G, Hart DJ & Spector TD (2005) Radiographic progression of lumbar spine disc
degeneration is influenced by variation at inflammatory genes: a candidate SNP association study in the
Chingford cohort. Spine (Phila Pa 1976) 30(21): 2445-51
-no MRI

182
Videman T & Battié MC (2002) Point of view. Spine 27(1): 82-83.
-commentary
Virtanen IM, Karppinen J, Taimela S, Ott J, Barral S, Kaikkonen K, Heikkila O, Mutanen P, Noponen N,
Mannikko M, Tervonen O, Natri A & Ala-Kokko L (2007) Occupational and genetic risk factors associated
with intervertebral disc disease. Spine (Phila Pa 1976) 32(10): 1129-34.
-no MRI
Wang ZC, Chen XS, Wang da W, Shi JG, Jia LS, Xu GH, Huang JH & Fan L (2010) The genetic
association of vitamin D receptor polymorphisms and cervical spondylotic myelopathy in Chinese subjects.
Clin Chim Acta 411(11-12): 794-797.
-not a relevant outcome
Williams FMK, Manek NJ, Sambrook PN, Spector TD & Macgregor AJ (2007) Schmorl's nodes: Common,
highly heritable, and related to lumbar disc disease. Arthritis Rheum 57(5): 855-860.
-no specific genetic variant reported
Wrocklage C, Wassmann H & Paulus W (2000) COL9A2 allelotypes in intervertebral disc disease.
Biochem Biophys Res Commun 279(2): 398-400.
-unclear whether MRI has been done or not
Yamada Y (2000) Association of a Leu(10)-> Pro polymorphism of the transforming growth factor-beta 1
with genetic susceptibility to osteoporosis and spinal osteoarthritis. Mech Ageing Dev 116(2-3): 113-123.
-review article
Yamada Y, Okuizumi H, Miyauchi A, Takagi Y, Ikeda K & Harada A (2000) Association of transforming
growth factor beta 1 genotype with spinal osteophytosis in Japanese women. Arthritis Rheum 43(2): 452-
460.
-no MRI for all, very small study
Ye W, Ma RF, Su PQ, Huang DS, Liu SL, Chen WJ & Wang XG (2007a) Association of single nucleotide
polymorphisms of IL-1b with lumbar disc disease. Yi chuan = Hereditas / Zhongguo yi chuan xue hui bian
ji 29(8): 923-928.
-only abstract, in Chinese; translation not available
Ye W, Huang D, Chen W, Li C, Peng Y, Liang A & Liu S (2007b) [Association of 86 bp variable number
tandem repeat polymorphism of interleukin-1 receptor antagonist gene with lumbar disc disease]. Nan
Fang Yi Ke Da Xue Xue Bao 27(10): 1485-8.
-only abstract, in Chinese; translation not available

183
Yuan H- & Lei L (2007) Current research in candidate genes of lumbar disc degeneration. Fudan
University Journal of Medical Sciences 34(3).
-only abstract, in Chinese; translation not available, likely a review article
Yuan H-, Tang Y, Liang Y-, Lei L, Xiao G-, Wang S & Xia Z- (2010) Matrix metalloproteinase-3 and vitamin
D receptor genetic polymorphisms, and their interactions with occupational exposure in lumbar disc
degeneration. J Occup Health 52(1): 23-30.
-no MRI

184
3 References for Appendix 2
Aranda B, Achuthan P, Alam-Faruque Y, Armean I, Bridge A, Derow C, Feuermann M, Ghanbarian AT, Kerrien S, Khadake J, Kerssemakers J, Leroy C, Menden M, Michaut M, Montecchi-Palazzi L, Neuhauser SN, Orchard S, Perreau V, Roechert B, van Eijk K & Hermjakob H (2010) The IntAct molecular interaction database in 2010. Nucleic Acids Res 38: 525–531.
Bader GD, Betel D & Hogue CWV (2003) BIND: the Biomolecular Interaction Network Database. Nucleic Acids Res 31(1): 248–250.
Breitkreutz B, Stark C & Tyers M (2003) The GRID: the General Repository for Interaction Datasets. Genome Biol 4(3).
Demacq C, Vasconcellos VB, Marcaccini AM, Gerlach RF, Machado AA & Tanus-Santos JE (2009) A genetic polymorphism of matrix metalloproteinase 9 (MMP-9) affects the changes in circulating MMP-9 levels induced by highly active antiretroviral therapy in HIV patients. Pharmacogenomics J 9(4): 265–273.
Demacq C, Vasconcellos VB, Izidoro-Toledo TC, da SS, Canalle R, Queiroz RG,de Paula, Tone LG & Tanus-Santos JE (2010) Vascular endothelial growth factor (VEGF) and endothelial nitric oxide synthase (NOS3) polymorphisms are associated with high relapse risk in childhood acute lymphoblastic leukemia (ALL). Clin Chim Acta 411(17–18): 1335–1340.
den Dunnen JT & Antonarakis SE (2000) Mutation nomenclature extensions and suggestions to describe complex mutations: a discussion. Hum Mutat 15(1): 7–12.
Hu Z, Li C, Chen K, Wang L, Sturgis EM, Spitz MR & Wei Q (2008) Single Nucleotide Polymorphisms in Selected Apoptotic Genes and BPDE-Induced Apoptotic Capacity in Apparently Normal Primary Lymphocytes: A Genotype-Phenotype Correlation Analysis. J Cancer Epidemiol 2008: 147905–147905.
Ioannidis JP, Boffetta P, Little J, O'Brien TR, Uitterlinden AG, Vineis P, Balding DJ, Chokkalingam A, Dolan SM, Flanders WD, Higgins JP, McCarthy MI, McDermott DH, Page GP, Rebbeck TR, Seminara D & Khoury MJ (2008) Assessment of cumulative evidence on genetic associations: interim guidelines. Int J Epidemiol 37(1): 120–132.
Jensen LJ, Kuhn M, Stark M, Chaffron S, Creevey C, Muller J, Doerks T, Julien P, Roth A, Simonovic M, Bork P & von Mering C (2009) STRING 8--a global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res 37: 412–416.
Joshi-Tope G, Gillespie M, Vastrik I, D'Eustachio P, Schmidt E, de Bono B, Jassal B, Gopinath GR, Wu GR, Matthews L, Lewis S, Birney E & Stein L (2005) Reactome: a knowledgebase of biological pathways. Nucleic Acids Res 33: 428–432.
Ju W, Kang S, Kim JW, Park NH, Song YS, Kang SB & Lee HP (2005) Promoter polymorphism in the matrix metalloproteinase-1 and risk of cervical cancer in Korean women. Cancer Lett 217(2): 191–196.

185
Kanehisa M, Goto S, Furumichi M, Tanabe M & Hirakawa M (2010) KEGG for representation and analysis of molecular networks involving diseases and drugs. Nucleic Acids Res 38: 355–360.
Keshava Prasad TS, Goel R, Kandasamy K, Keerthikumar S, Kumar S, Mathivanan S, Telikicherla D, Raju R, Shafreen B, Venugopal A, Balakrishnan L, Marimuthu A, Banerjee S, Somanathan DS, Sebastian A, Rani S, Ray S, Harrys Kishore CJ, Kanth S, Ahmed M, Kashyap MK, Mohmood R, Ramachandra YL, Krishna V, Rahiman BA, Mohan S, Ranganathan P, Ramabadran S, Chaerkady R & Pandey A (2009) Human Protein Reference Database--2009 update. Nucleic Acids Res 37: 767–772.
Lage K, Karlberg EO, Storling ZM, Olason PI, Pedersen AG, Rigina O, Hinsby AM, Tumer Z, Pociot F, Tommerup N, Moreau Y & Brunak S (2007) A human phenome-interactome network of protein complexes implicated in genetic disorders. Nat Biotechnol 25(3): 309–316.
Pereira FA, Pinheiro dS, Rodart IF, Carmo T,Mauadie Azevedo, Lemaire DC & Reis MG (2008) Association of TGF-beta1 codon 25 (G915C) polymorphism with hepatitis C virus infection. J Med Virol 80(1): 58–64.
Tsironi EE, Pefkianaki M, Tsezou A, Kotoula MG, Dardiotis E, Almpanidou P, Papathanasiou AA, Rodopoulou P, Chatzoulis DZ & Hadjigeorgiou GM (2009) Evaluation of MMP1 and MMP3 gene polymorphisms in exfoliation syndrome and exfoliation glaucoma. Mol Vis 15: 2890–2895.
Xenarios I, Salwinski L, Duan XJ, Higney P, Kim S & Eisenberg D (2002) DIP, the Database of Interacting Proteins: a research tool for studying cellular networks of protein interactions. Nucleic Acids Res 30(1): 303–305.
Zanzoni A, Montecchi-Palazzi L, Quondam M, Ausiello G, Helmer-Citterich M & Cesareni G (2002) MINT: a Molecular INTeraction database. FEBS Lett 513(1): 135–140.
Zhai Y, Qiu W, Dong X, Zhang X, Xie W, Zhang H, Yuan X, Zhou G & He F (2007) Functional polymorphisms in the promoters of MMP-1, MMP-2, MMP-3, MMP-9, MMP-12 and MMP-13 are not associated with hepatocellular carcinoma risk. Gut 56(3): 445–447.

186

187
List of original articles
I Eskola PJ, Lemmelä S, Kjaer P, Solovieva S, Männikkö M, Tommerup N, Lind-Thomsen A, Husgafvel-Pursiainen K, Cheung KMC, Chan D, Samartzis D*, Karppinen J* (2012) Genetic association studies in lumbar disc degeneration: a systematic review. PLOS ONE. In press.
II Kelempisioti A*, Eskola PJ*, Okuloff A, Karjalainen U, Takatalo J, Daavittila I, Niinimäki J, Sequeiros RB, Tervonen O, Solovieva S, Kao PY, Song YQ, Cheung KM, Chan D, Ala-Kokko L, Järvelin MR, Karppinen J, Männikkö M (2011) Genetic susceptibility of intervertebral disc degeneration among young Finnish adults. BMC Med Genet 22(12): 153
III Eskola PJ, Kjaer P, Daavittila IM, Solovieva S, Okuloff A, Sorensen JS, Wedderkopp N, Ala-Kokko L, Männikkö M & Karppinen JI (2010) Genetic risk factors of disc degeneration among 12–14-year-old Danish children: a population study. Int J Mol Epidemiol Genet 1(2): 158–65
IV Eskola PJ, Kjaer P, Sorensen JS, Okuloff A, Wedderkopp N, Daavittila I, Ala-Kokko L, Männikkö M, Karppinen J (2012) Gender difference in genetic association between IL1A variant and early lumbar disc degeneration: a three-year follow-up. Int J Mol Epidemiol Genet 3(3): 195–204
Reprinted with permission from Public Library of Science (I), BioMed Central (II)
and e-Century Publishing Corporation (III, IV).
Original publications are not included in the electronic version of the dissertation.

188

A C T A U N I V E R S I T A T I S O U L U E N S I S
Book orders:Granum: Virtual book storehttp://granum.uta.fi/granum/
S E R I E S D M E D I C A
1168. Moilanen, Anne-Mari (2012) Identification of novel drug targets for the treatmentof heart failure
1169. Jounio, Ulla (2012) Oropharyngeal carriage of respiratory bacteria among militaryconscripts
1170. Virtanen, Katri (2012) ”Äiti, täällä on toisia samanlaisia, ku mä!” : Voimisteluseuraja kouluterveydenhuolto perheiden tukena lasten painonhallinnassa
1171. Ilomäki, Essi (2012) Conduct disorder among girls: violent behaviour, suicidalityand comorbidity : A study of adolescent inpatients in Northern Finland
1172. Ilomäki, Risto (2012) Substance use disorders in adolescence: Comorbidity,temporality of onset and socio-demographic background : A study of adolescentpsychiatric inpatients in Northern Finland
1173. Saxlin, Tuomas (2012) Periodontal infection and obesity—results of a population-based survey
1174. Lämsä, Virpi (2012) Regulation of murine hepatic Cytochrome P450 2a5expression by transcription factor Nuclear factor (erythroid-derived 2)-like 2
1175. Huhtakangas, Juha (2012) The influence of medication on the incidence, outcome,and recurrence of primary intracerebral hemorrhage
1176. Puljula, Jussi (2012) Alcohol-related traumatic brain injuries before and after thereduction of alcohol prices : Observations from Oulu Province and NorthernOstrobothnia
1177. Kenttä, Tuomas (2012) Dynamics of cardiac repolarization during exercise : Rate-dependence and prognostic significance
1178. Vasunta, Riitta-Liisa (2012) Ambulatory blood pressure : association withmetabolic risk indicators, renal function and carotid artery atherosclerosis
1179. Haapsamo, Helena (2012) A Follow-up study of children's communicativedevelopment : Associations to social-emotional and behavioural problems andcompetences and experienced maternal stress
1180. Norppa, Anna (2012) Association between periodontal and systemicinflammation : A study of pro- and anti-inflammatory mediators
1181. Karjalainen, Teemu (2012) Nitinol shape memory alloy in flexor tendon repair
1182. Saukkonen, Tuula (2012) Prediabetes and associated cardiovascular risk factors :A prospective cohort study among middle-aged and elderly Finns

ABCDEFG
UNIVERS ITY OF OULU P.O.B . 7500 F I -90014 UNIVERS ITY OF OULU F INLAND
A C T A U N I V E R S I T A T I S O U L U E N S I S
S E R I E S E D I T O R S
SCIENTIAE RERUM NATURALIUM
HUMANIORA
TECHNICA
MEDICA
SCIENTIAE RERUM SOCIALIUM
SCRIPTA ACADEMICA
OECONOMICA
EDITOR IN CHIEF
PUBLICATIONS EDITOR
Senior Assistant Jorma Arhippainen
University Lecturer Santeri Palviainen
Professor Hannu Heusala
Professor Olli Vuolteenaho
University Lecturer Hannu Heikkinen
Director Sinikka Eskelinen
Professor Jari Juga
Professor Olli Vuolteenaho
Publications Editor Kirsti Nurkkala
ISBN 978-951-42-9991-9 (Paperback)ISBN 978-951-42-9992-6 (PDF)ISSN 0355-3221 (Print)ISSN 1796-2234 (Online)
U N I V E R S I TAT I S O U L U E N S I S
MEDICA
ACTAD
D 1184
ACTA
Pasi Eskola
OULU 2012
D 1184
Pasi Eskola
THE SEARCH FOR SUSCEPTIBILITY GENESIN LUMBAR DISC DEGENERATIONFOCUS ON YOUNG INDIVIDUALS
UNIVERSITY OF OULU GRADUATE SCHOOL;UNIVERSITY OF OULU, FACULTY OF MEDICINE,INSTITUTE OF BIOMEDICINE,DEPARTMENT OF MEDICAL BIOCHEMISTRY AND MOLECULAR BIOLOGY;INSTITUTE OF CLINICAL MEDICINE,DEPARTMENT OF PHYSICAL MEDICINE AND REHABILITATION;BIOCENTER OULU;CENTER FOR CELL-MATRIX RESEARCH


















