
The Role of Mercenaria mercenaria and Mytilus edulis and ... · Ream 1!!!!! The Role of Mercenaria...
31
Ream 1 The Role of Me r cenaria me r cenaria and Myt ilus eduli s in Coupling Benthic and Pelagic Nutrient Cycles Kelsey A. Ream 1 Mentor: Anne Giblin 2 1 Departments of Biology and Environmental Science, Allegheny College, Meadville, PA 16335 2 The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA 02543
Transcript of The Role of Mercenaria mercenaria and Mytilus edulis and ... · Ream 1!!!!! The Role of Mercenaria...

Ream 1
The Role of Mercenaria mercenaria and Mytilus edulis in Coupling Benthic and Pelagic Nutrient Cycles
Kelsey A. Ream1 Mentor: Anne Giblin2
1Departments of Biology and Environmental Science, Allegheny College, Meadville, PA 16335 2 The Ecosystems Center, Marine Biological Laboratory, Woods Hole, MA 02543

Ream 2
Abstract
Bivalves have well-documented bioremediation capabilities, however, these capabilities
differ depending on the preferred lifestyle of a given bivalve species. The Blue Mussel, Mytilus
edulis, prefers to live in aggregate colonies, attached to a hard substrate, suspended slightly
above the benthos. The Hard Shelled Clam, Mercenaria mercenaria, buries itself in the
sediments at the bottom of coastal waters. I chose to study these two species in a microcosm
experiment including micrcosms containing Mytilus alone, Mytilus and sediment, Mercenaria
alone, Mercenaria and sediment, and sediment alone over the course of approximately 3 weeks. I
took water samples in order to measure clearance rates of phytoplankton after a feeding, and to
measure changes in nutrient concentrations over time. Mytilus feed more rapidly, respire more,
and have higher daily fluxes of nitrate, ammonium, and phosphate than do Mercenaria. Mytilus
appear to recycle nutrients much faster than Mercenaria, while simultaneously promoting
denitrification. While claims are often made regarding the abilities of bivalves to remove
phytoplankton growth from excess nutrients and prevent eutrophication events, it appears that
these abilities are highly dependent on living habits and other differences between species, and
that further investigation is warranted into the specifics of these differences.
K ey Words: bivalve, Mercenaria mercenaria, Mytilus edulis, nutrient cycling, phytoplankton
clearance, respiration
Introduction
Knowledge of species with bioremediation capabilities is becoming increasingly
necessary in estuarine coastal ecosystems. These systems tend to be overloaded with nutrients

Ream 3
from waste water inputs, septic plumes, precipitation, fertilizer usage, and other anthropogenic
sources (Valiela et al., 1997). Understanding the capabilities of native species to affect nutrient
levels in the water column can be important for ensuring proper protection and valuation of such
species.
One group species with well-studied bioremediation capabilities is bivalves. Bivalve
species in the North Atlantic are highly varied in their habitat preferences and living habits.
Some species, such as the Hard Shell Clam (Mercenaria mercenaria), prefer to bury themselves
in the sediments of coastal and estuarine environments (Lorio & Malone, 1995). Others like the
Blue Mussel (Mytilus edulis) prefer to form aggregates on a solid substrate, suspending
themselves above the sediment (Bayne, 1964). Both species are filter-feeders, removing
phytoplankton and suspended organic matter from the water column as a food source. Both
species excrete digested matter onto the sediment in the form of inorganic nutrient-rich
biodeposits.
By removing nutrient-rich phytoplankton and other suspended organic material from the
water column while depositing fecal matter in the sediment and stimulating microbial activity
there, bivalves serve as an important link in the nutrient cycles of both of these locations within
an ecosystem. The ability of these animals to stimulate denitrification through their additions of
inorganic nitrogen to the sediments is relatively well known (Cornwell, Kemp, & Kana, 1999).
Bivalves are particularly good at stabilizing ecosystem nutrient fluxes because of the fact that
they turn over very slowly (compared to phytoplankton) (Dame, 1996). Their slow turnover rate
means that a large bivalve population represents a substantial nutrient-sink in coastal ecosystems.
Bivalve growth is limited by food availability, and bivalves can continue to filter feed
increasingly rapidly as their food sources become increasingly available. Their ability to exert

Ream 4
top-down control on phytoplankton populations, thereby helping to control algal blooms, has
often been noted. M. mercenaria, in particular, has a well known ability to exert such control
(Officer, Smayda, & Mann, 1982); (Dame, 1996) during eutrophication events that can
sometimes result in hypoxia (Altieri, 2008). M. edulis appears to be less tolerant in that it
decreases in survivorship under hypoxic conditions (Altieri, 2008). While both of these
organisms play a role in how coastal ecosystems respond to nutrient loading, the unique
lifestyles of M. mercenaria and M. edulis cause important differences in the way they influence
nutrient cycling.
The purpose of this experiment was to investigate how bivalve lifestyle habits (burrowing
vs. non-burrowing) affect cycling of ammonium, nitrate, and phosphate in the water column and
sediment of experimental microcosms over the course of approximately 3 weeks.
Methods
Microcosm Set-up
My experiment included five treatments, Mytilus alone, Mytilus and sediment,
Mercenaria alone, Mercenaria and sediment, and sediment alone, with three replicates of each
treatment. I set up fifteen cylindrical plastic cores to contain the various treatments, each of
which was kept in an incubation tank filled with water at approximately 20°C to maintain a
constant temperature within the microcosms themselves. Each microcosm was sealed to prevent
movement of water, and outfitted with a magnetic stir bar to maintain water circulation.
Sediment for use in the microcosms was collected at the Sippewissett Marsh in Falmouth MA.
Microcosms containing sediment were filled with sediment to a height of approximately 10 cm.
All microcosms were filled with seawater.

Ream 5
Using a living biomass: living biomass+shell ratio, I determined that one Mercenaria
weighing approximately 64 g had a similar biomass to 13 small mussels weighing approximately
42 g. Therefore, in order to achieve approximately equal living biomasses (not including the
shell) in all microcosms containing animals, microcosms containing Mercenaria contained 1
clam, while microcosms containing Mytilus contained 13 mussels. I recorded exact weights of
the bivalve biomass in each microcosm both before the start of the experiment, and after the end
of the experiment. Mytilus were added to the microcosms in mesh bags with large holes in order
to provide them with some substrate to aggregate on while still allowing them to move around
the microcosm. Mercenaria were added to the microcosms without any confinement in order to
allow them to bury themselves in the sediment, where possible. In between periods of water
sampling, the microcosms were bubbled with air-stones continuously in order to ensure proper
oxygenation.
Phytoplankton Clearance
I added a phytoplankton paste, Reed Mariculture Algae Paste Shellfish Diet 1800, to each
microcosm 3 times throughout the course of the experiment in order to provide nutrition for the
clams and measured clearance rates of phytoplankton from the water column. Enough paste was
added to achieve an approximate concentration of 400 cells/ µL in the water column of each
microcosm, and the water in each microcosm was stirred continuously using magnetic stirrers
throughout the duration of water sampling. During the first two feeding trials, water was sampled
0, 15, 30, and 60 minutes after the paste was added to the microcosms. During the third feeding
trial, water was sampled 0, 125, 240, 380, 450, and 480 minutes after the paste was added. Water
samples were analyzed for in-vivo fluorescence using a Turner 450 Fluorometer.

Ream 6
Nutrient Analysis and Respiration Measurements
In order to measure differences in respiration between Mercenaria, Mytilus, and the
sediment, I sealed each of the microcosms to prevent oxygen exchange with the atmosphere and
took approximately five water samples over the course of the dissolved oxygen (DO)
concentration in the microcosms dropping 2 ppm. DO was measured a WTW oxygen electrode
system. Water samples were then analyzed for nitrate using an automated Lachat Flow Injection
Analyzer and a variation of the method by (Wood, Armstrong, & Richards, 1967), phosphate
using a Shimadzu Spectrophotometer and an adaptation of(Murphy & Riley, 1962), and
ammonium using a Shimadzu spectrophotometer and an adaptation of (Solorzano, 1969).
Sediment Analysis
After the completion of the experiment, sediment samples were taken from the top 2 cm
of the sediment layer in each microcosm containing sediment. Sediments were then dried,
acidified to remove carbonates, and analyzed for carbon and nitrogen using a Perkin-Elmer
Series 2 CHNS/O Analyzer 2400.
Results
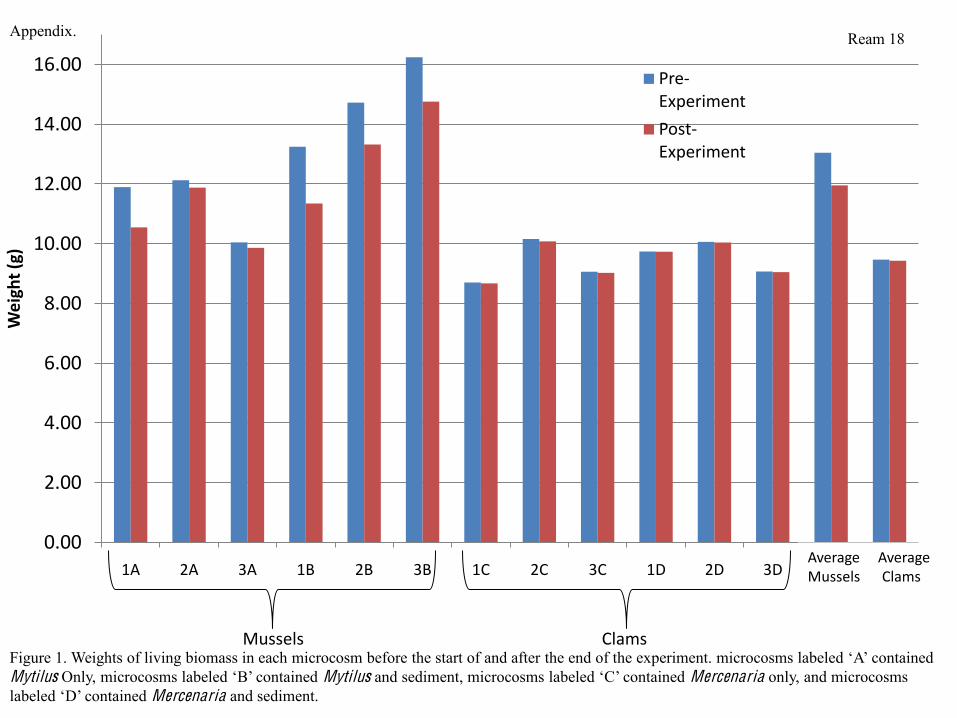
I weighed the bivalves in each microcosm to determine whether they gained or lost
weight over the course of the experiment. Microcosms containing mussels lost, on average,
approximately 1 g of living biomass per microcosm (8.4 % of the total microcosm biomass),
while microcosms containing clams lost an average of 0.033 g per microcosm (0.35% of the total

Ream 7
microcosm biomass) (Figure 1). There were significant differences between the amount of
weight lost by each type of bivalve in sediment and alone. Mercenaria alone lost more weight
than Mercenaria in sediment, while Mytilus in sediment lost more weight than Mytilus alone
(Table 1). No clams died over the course of the experiment, and while 5 mussels total died, no
more than two died in anyone microcosm.
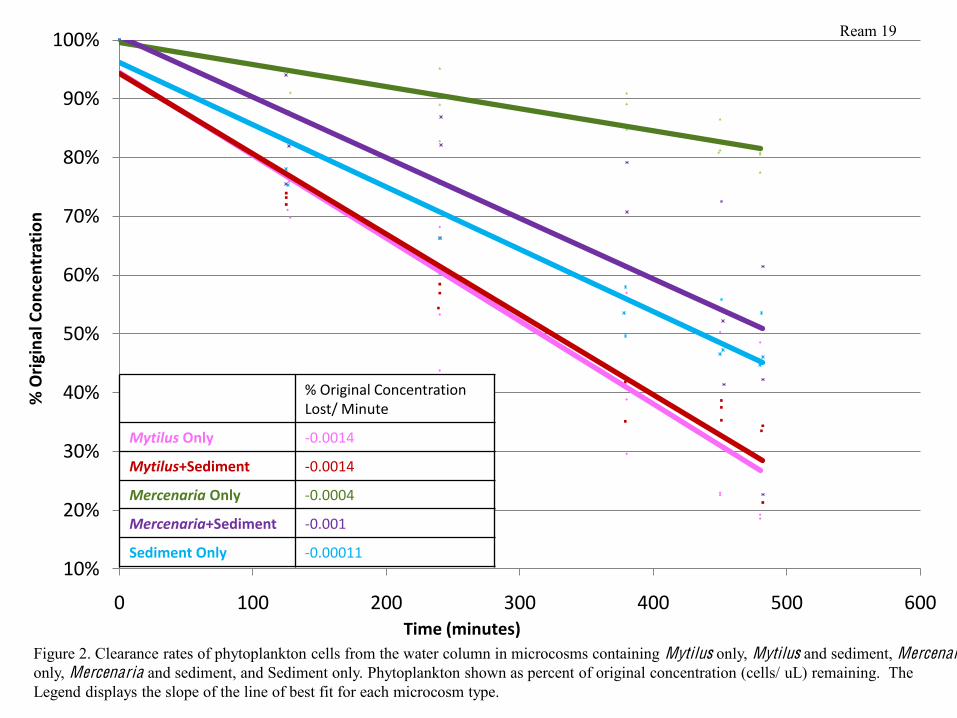
Phytoplankton Clearance
In microcosms containing Mytilus only and Mytilus and sediment, phytoplankton was
removed from the water column at approximately the same rate. In all 6 microcosms containing
Mytilus, the concentration of phytoplankton cells decreased by approximately 0.084% of the
original concentration per hour. Microcosms containing Mercenaria and sediment removed
phytoplankton more slowly; the original concentration decreased by 0.06% per hour.
Microcosms containing Mercenaria only removed phytoplankton much more slowly, at a rate of
only 0.024% of the original concentration removed per hour. Phytoplankton concentrations
decreased in the sediment at the slowest rate; 0.0066% of the original concentration settled out of
solution per hour (Figure 2).
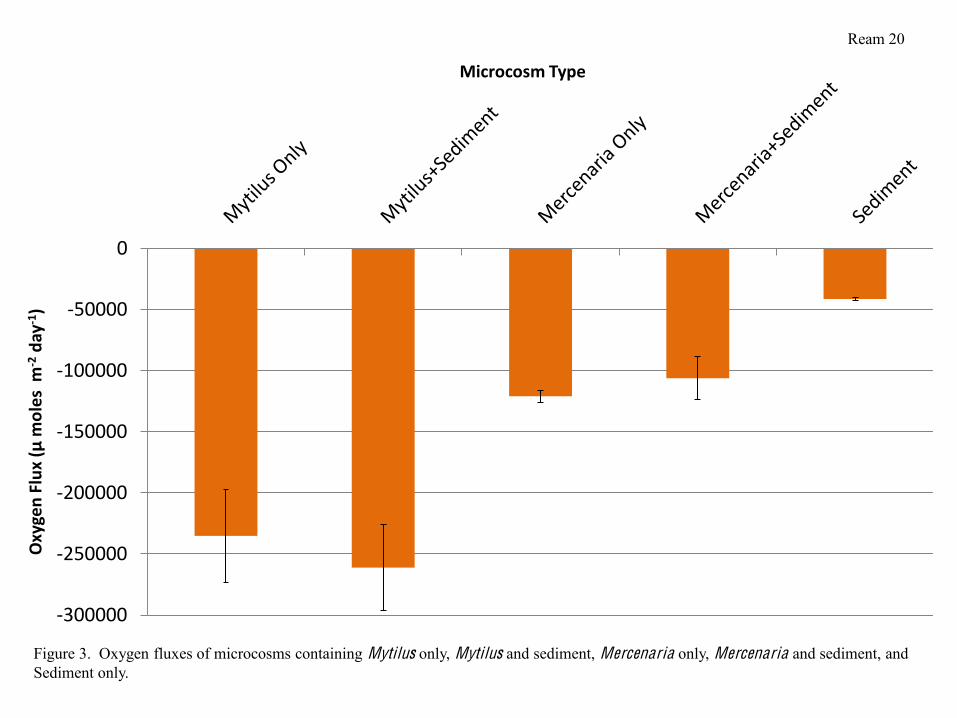
Nutrient Analysis and Respiration Measurements
Microcosms containing Mytilus respired nearly twice as quickly as microcosms
containing Mercenaria, with microcosms containing Mytilus and sediment respiring slightly
faster than those containing Mytilus alone. Microcosms containing Mercenaria and sediment
actually respired slightly more slowly than those containing Mercenaria alone, however in both
cases, the differences between microcosms containing bivalves alone and bivalves and sediment
were not significant. The Sediment alone respired at the lowest rate of all the microcosms, and
was significantly less than microcosms containing both Mercenaria and Mytilus (Figure 3).

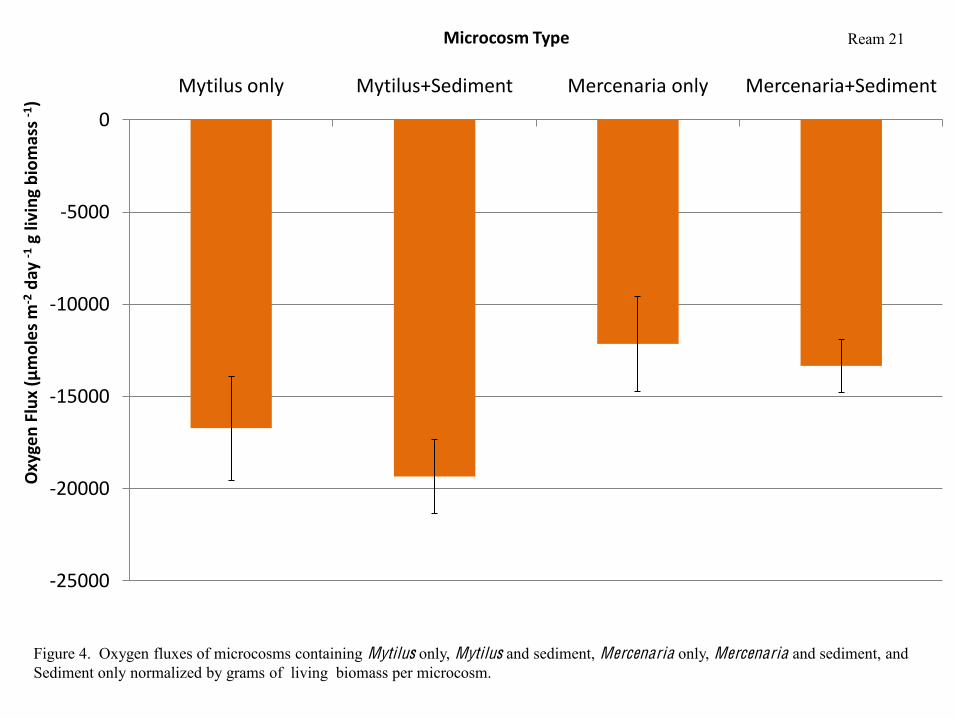
Ream 8
When respiration rates were normalized by grams of living biomass per microcosm, the same
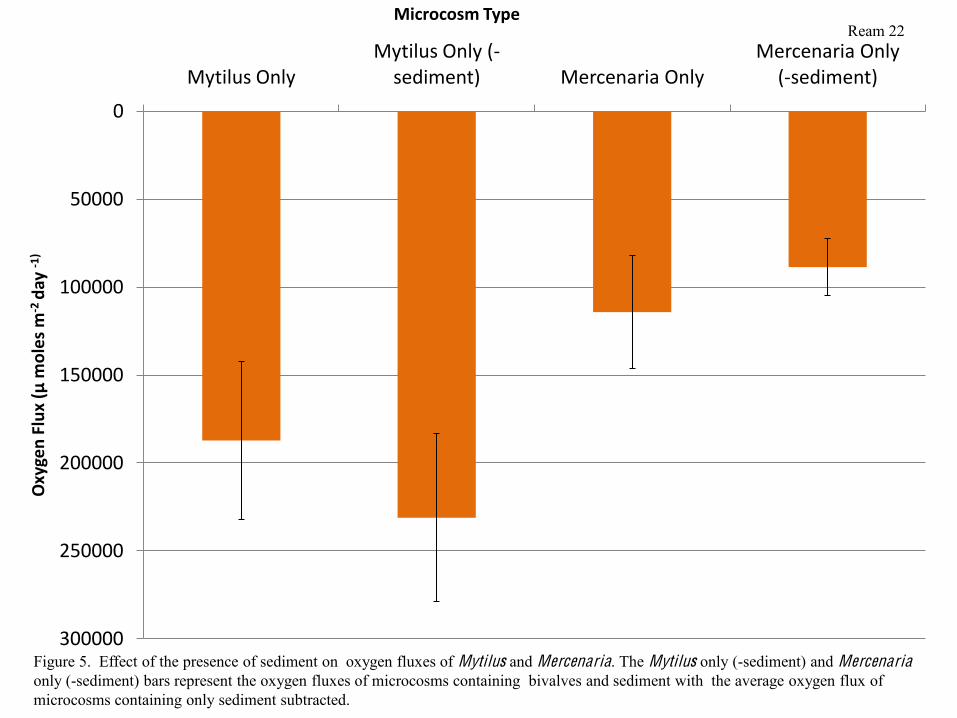
trends were apparent (Figure 4). Subtracting the average respiration rate of microcosms
containing Sediment only from respiration rates of microcosms containing sediment and bivalves
shows that Mytilus in microcosms containing sediment respire slightly more quickly than those
in microcosms containing more sediment. In contrast, Mercenaria in microcosms containing
sediment respire slightly more slowly than those in microcosms containing no sediment (Figure
5), however, the difference was not significant.
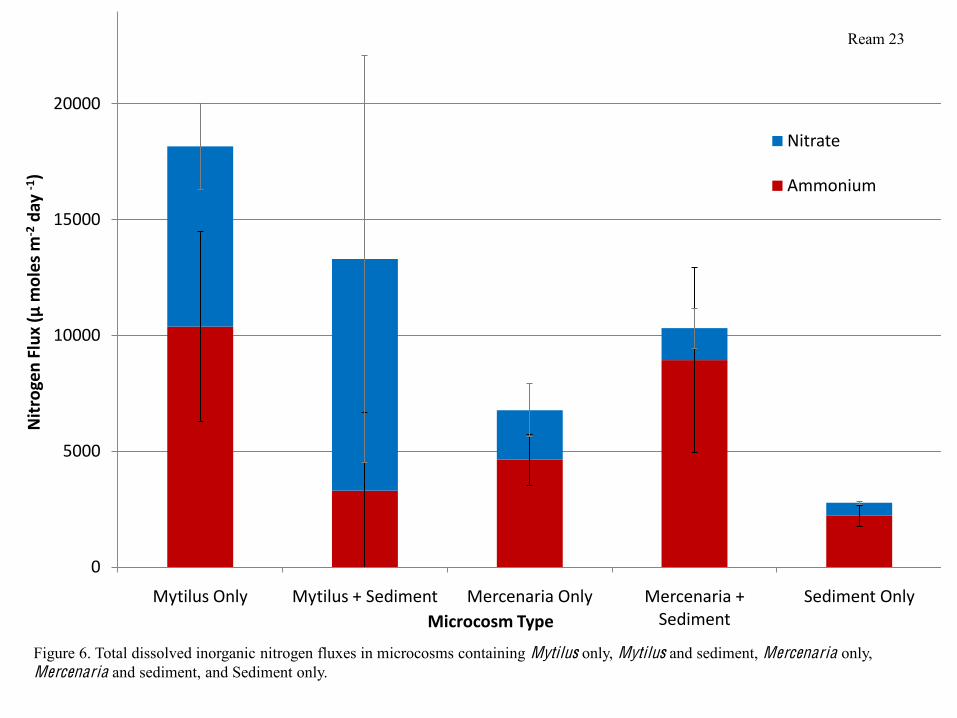
Ammonia fluxes were higher in microcosms containing Mytilus only than in microcosms
containing Mytilus and sediment, while they were slightly higher in microcosms containing
Mercenaria and sediment than in microcosms containing Mercenaria only. Nitrate fluxes were
similar in all microcosms containing Mytilus, and similar in all microcosms containing
Mercenaria, but were considerably higher in microcosms containing Mytilus than in microcosms
containing Mercenaria. Both ammonium and nitrate fluxes were lowest in microcosms
containing Sediment only. Microcosms containing Mytilus typically had higher fluxes of
dissolved inorganic nitrogen (DIN) than microcosms containing Mercenaria, which in turn had
higher fluxes than microcosms containing sediment alone. Additionally, DIN fluxes were most
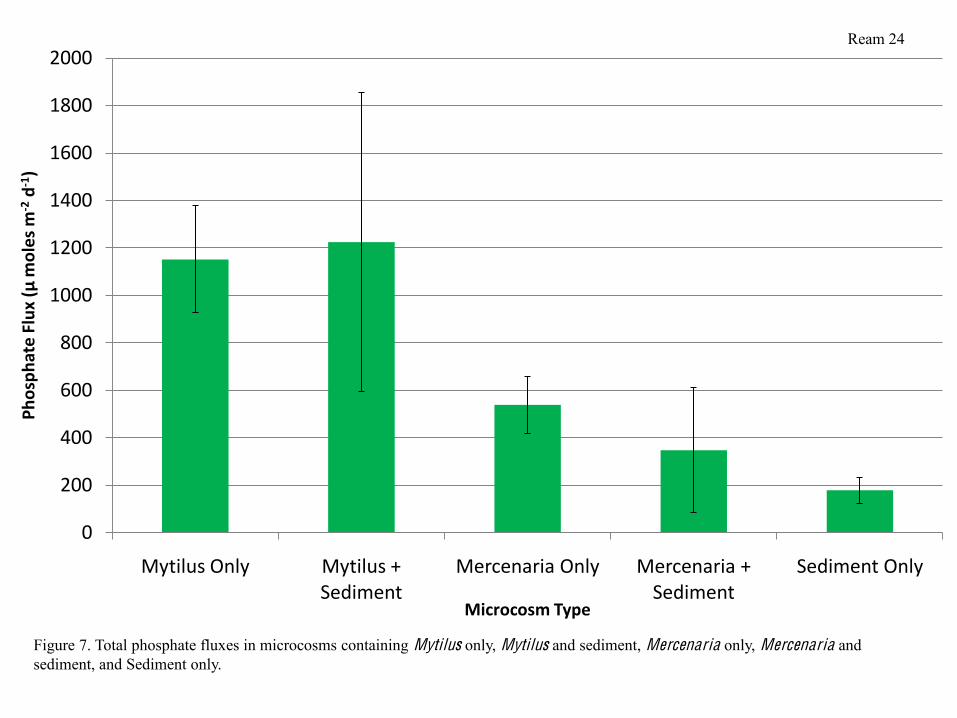
variable in microcosms containing Mytilus and sediment (Figure 6). Phosphate fluxes followed a
similar trend to that of the nitrate fluxes in that they were similar in all microcosms containing
Mytilus, similar in all microcosms containing Mercenaria, and significantly higher in
microcosms containing Mytilus than in microcosms containing Mercenaria. Microcosms
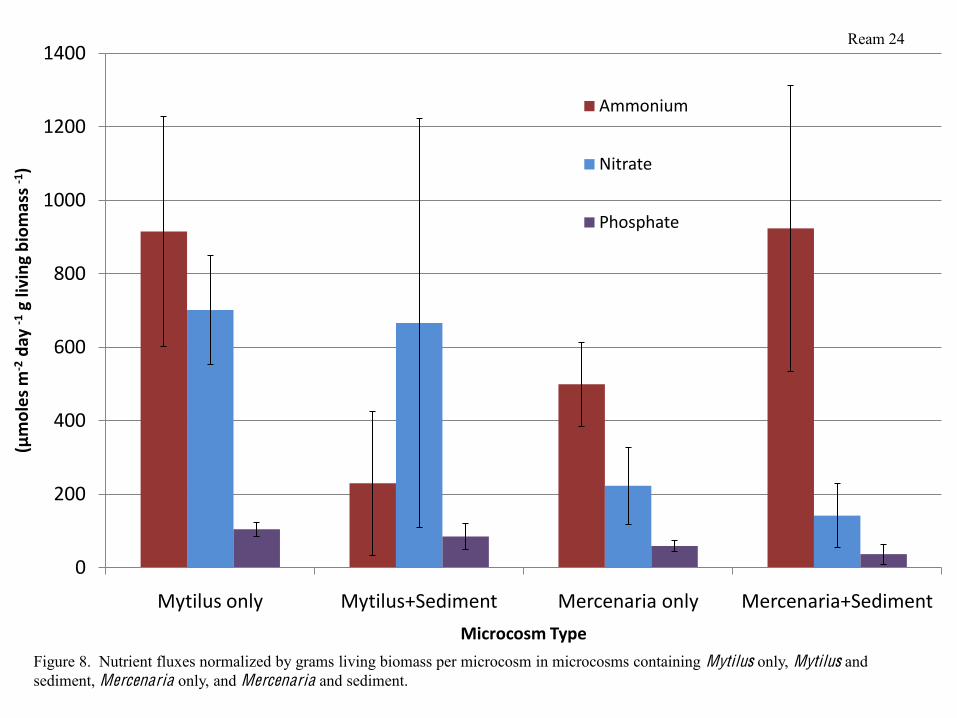
containing sediment only had the lowest phosphate fluxes (Figure 7). When nutrient fluxes were
normalized by grams living biomass per microcosm, the same trends were still apparent (Figure
8).

Ream 9
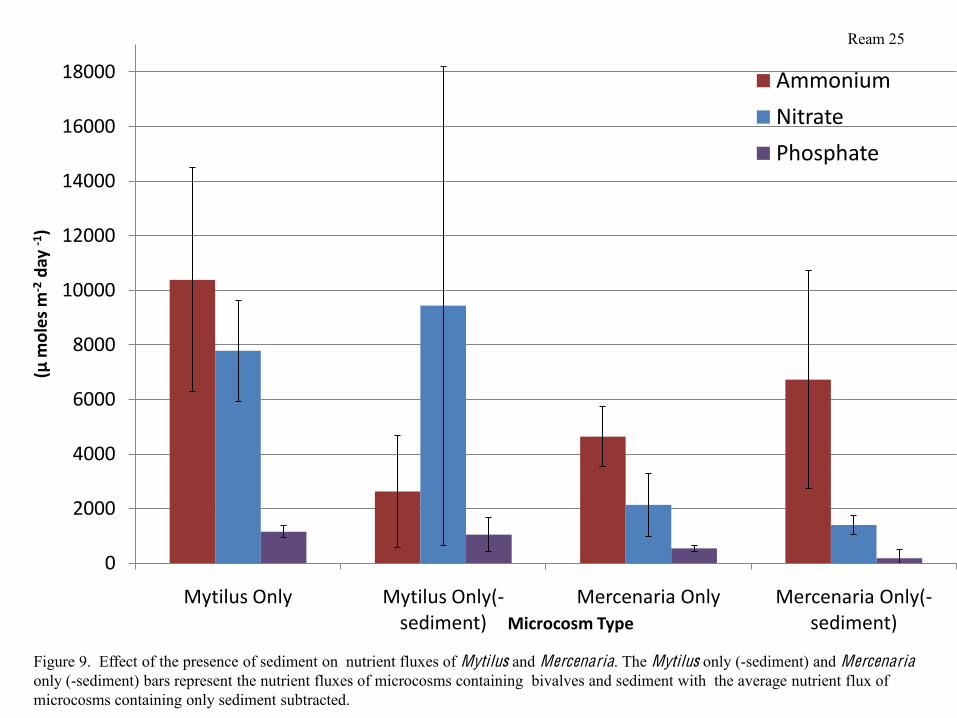
In order to examine how the presence of the sediment affects nutrient fluxes, I subtracted
the average nutrient fluxes of the sediment from the nutrient fluxes of microcosms containing
sediment and animals. In comparing microcosms containing Mytilus, those containing sediment
had significantly higher ammonium fluxes than those without sediment. While nitrate
concentrations were slightly higher in microcosms containing sediment than microcosms
containing Mytilus only, there were no other significant differences in nutrient fluxes between
Mytilus only and Mytilus and sediment. In microcosms containing Mercenaria, there were no
significant differences between those that contained sediment and those that did not. Ammonia,
however, was slightly higher in microcosms containing Mercenaria and sediment, while
phosphate and nitrate fluxes were lower in microcosms containing Mercenaria and sediment
than in microcosms containing only Mercenaria (Figure 9).
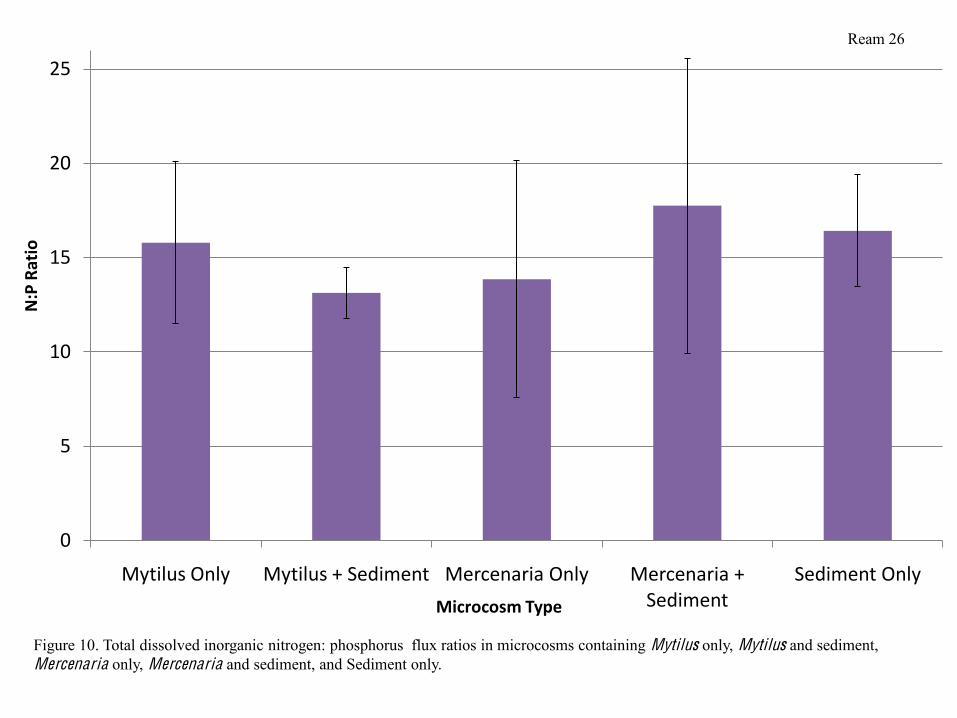
Dissolved inorganic nitrogen: phosphorus (N:P) flux ratios were not significantly
different from eachother in any of the 5 treatments, however, were all relatively close to a ratio
of 16:1, with microcosms containing Mytilus and sediment alone having the lowest N:P ratio,
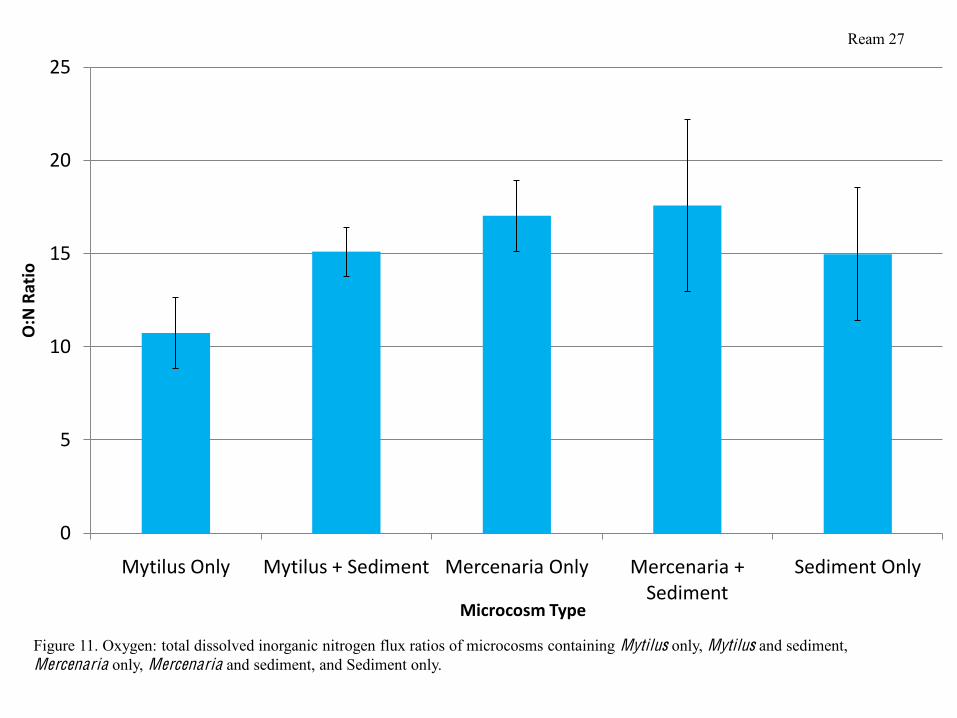
and microcosms containing Mercenaria and sediment having the highest (Figure 10). Oxygen
(O2): nitrogen ratios (O:N) were not significantly different in microcosms containing Mytilus and
sediment, Mercenaria only, Mercenaria and sediment, and Sediment only. Microcosms
containing Mytilus only had significantly lower O:N ratios than any other treatments (Figure 11).
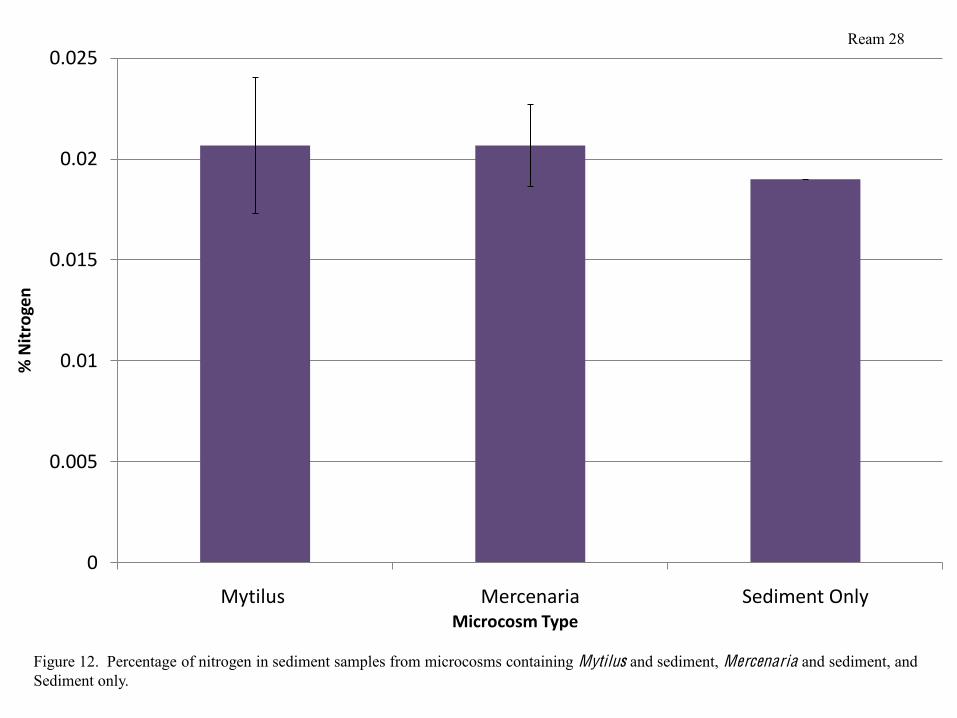
Sediment Analysis
CHN analysis of sediment from each type of microcosm did not yield any significantly
different results. Sediment from microcosms containing Mytilus and Mercenaria did, however,
have a slightly a slightly higher percentage of nitrogen than sediment from microcosms
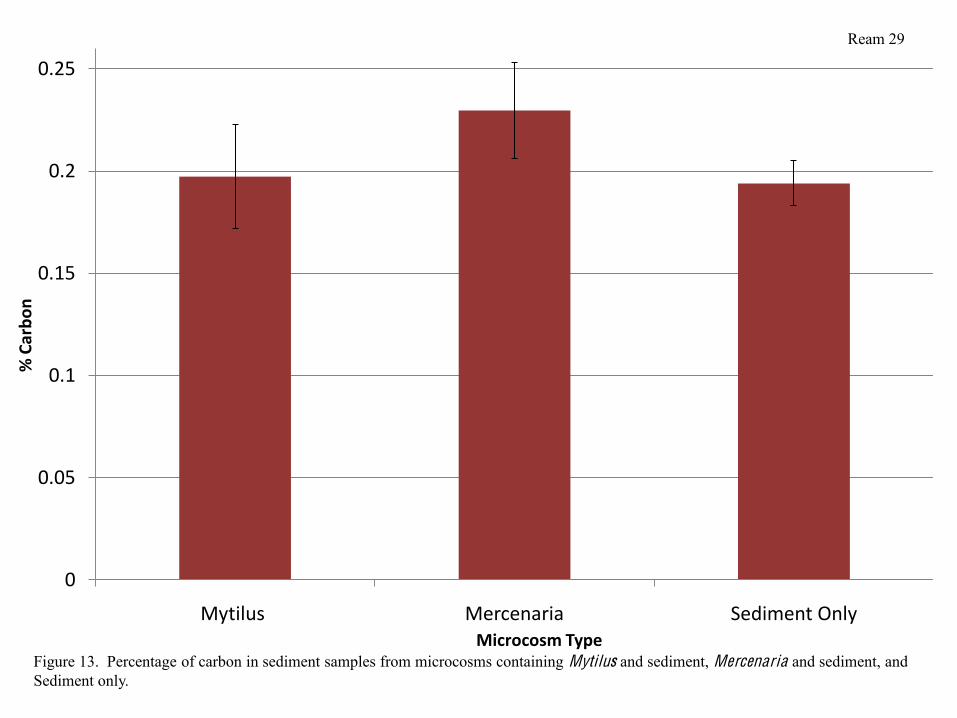
containing sediment only (Figure 12). Percentage of carbon also did not differ significantly

Ream 10
between the 3 types of microcosms, however, it was slightly higher in microcosms containing
Mercenaria than in microcosms containing either Mytilus or sediment alone (Figure 13).
Discussion
Over the course of my experiment, the weight of the total living biomass in each
microcosm decreased, with significant differences between microcosms containing bivalves
alone and bivalves in sediment (Figures 1, Table 1). Microcosms containing Mytilus lost, on
average, much more weight than microcosms containing Mercenaria (Figure 2) due to the fact
that several Mytilus died over the course of the experiment, while no Mercenaria died. Mytilus
are known to be sensitive to hypoxic conditions (Altieri & Witman, 2006), so this result is to be
expected. The more extensive weight loss in microcosms containing Mytilus and sediment may
Tbe related to stress from higher nutrient concentrations, such as ammonium, in microcosms
with this treatment type. It is likely that Mercenaria may have lost weight for other reasons;
because they appeared to be respiring more and feeding less, it is likely that they were somewhat
more stressed than the Mytilus. Mercenaria in microcosms without sediment were particularly
stressed, undoubtedly because living in an exposed environment without being buried in
sediment is unnatural, and something that would not occur in the wild. Additionally, it is possible
that I did not feed the bivalves enough over the course of the experiment; because the Mytilus
used for the experiment were younger than the Mercenaria, a lack of proper nutrition may
explain why they were more sensitive, lost more weight, and had more deaths.

Ream 11
Phytoplankton clearance
Mytilus appeared to filter phytoplankton from the water column much more rapidly than
Mercenaria did (Figure 2). This could have occurred for several reasons, one of which is that the
mussels are situated directly in the water column itself, and therefore were much closer to the
location at which phytoplankton additions were made during the feeding trials (the top of the
microcosm). Because of this difference, mussels would have been able to remove phytoplankton
from the water as soon as it was added, while Mercenaria would have had to wait until it settled
towards the bottom of the water column, potentially creating a delay in the Mercenaria
measured feeding rates. In addition, it is also possible that the Mytilus were simply more
metabolically active within the microcosms, causing them to feed more. Similarly, Mercenaria
may have fed less because they were less metabolically active. In particular, it is also possible
that Mercenaria in microcosms without sediment were so stressed to be exposed in open water
that they did not wish to expend energy in order to feed. These observations are particularly
interesting in that mussels are not as well documented as clams in their ability to control
eutrophication events, despite the fact that it appears that mussel populations may be more
effective at controlling algal blooms under certain conditions. It is important, however, to
consider this in conjunction with their limited ability to tolerate hypoxic conditions(Altieri &
Witman, 2006). Phytoplankton concentrations decreased at the slowest rate in microcosms
containing sediment only. This rate of decrease represented the rate at which phytoplankton were
falling out of solution and settling to the bottom of the microcosm.
Respiration Measurements

Ream 12
Mytilus respired significantly more than Mercenaria, regardless of whether the
microcosm housing the bivalves had sediment or not (Figures 3 and 4). This, in addition to the
phytoplankton clearance rates, alludes to the idea that Mytilus were much more metabolically
active than Mercenaria. Mytilus in sediment did tend to respire slightly more than Mytilus alone
(Figure 5) and although this trend was not significant, it is logical that stressed mussels that lost a
large amount of weight over the course of the experiment would have been respiring more than
the less stressed Mytilus in microcosms without sediment. A similar trend exists in Mercenaria;
microcosms containing Mercenaria alone respired slightly more than microcosms with
Mercenaria and sediment (Figure 5). While the trend is not significant, it correlates to the trend
of higher weight loss in microcosms containing bivalves that were stressed.
Nutrient Analysis
The lower dissolved inorganic nitrogen (DIN) fluxes in microcosms containing Mytilus
and sediment as compared with microcosms containing Mytilus (Figure 6) only indicate that
some coupled nitrification/ denitrification of the DIN in Mytilus waste may be occurring. It is
likely that some ammonium is being converted to nitrate via nitrification, however, some nitrate
must be lost via denitrification in anaerobic areas of the sediment in order to account for the
lower DIN flux in microcosms containing Mytilus and sediment. The tendency of mussels to
attach themselves to a hard substrate above the sediment, rather than burrowing into it, is likely
conducive to the anaerobic sediment conditions that facilitate denitrification. The burrowing
habits of Mercenaria appear to be less conducive to denitrification. As Mercenaria dig their way
into the sediment, they may allow oxygen to enter the sediment where it previously would have
been excluded, effectively slowing denitrification. While some denitrification may have occurred

Ream 13
in microcosms containing Mercenaria and sediment, it probably played a less significant role
than in microcosms with Mytilus and sediment. It is likely that nitrification and ammonification
were the more prominent processes in microcosms containing Mercenaria and sediment. This is
particularly likely given the lower rates of phytoplankton consumption and respiration rates in
microcosms containing Mercenaria. Decomposing phytoplankton that were not consumed by the
less metabolically active Mercenaria may have provided a large base of organic N for
ammonification. Finally, DIN fluxes were lowest in microcosms containing sediment only. This
fits with expectation as sediment alone should not produce nearly as much ammonium and
nitrate as metabolically active animals. Additionally, it should be noted that nutrient fluxes in
microcosms containing Mytilus were usually the most variable and therefore had the largest error
bars. This is likely due to the fact that there were a larger number of individual animals in
microcosms containing Mytilus introducing a larger amount of variability.
Phosphate fluxes were significantly higher in microcosms containing Mytilus than in
microcosms containing Mercenaria, while sediment alone had slightly lower phosphate fluxes
than microcosms containing Mercenaria, but not significantly so (Figure 7). Mytilus phosphate
production is likely due to the increased metabolic activity and corresponding waste production
of these bivalves.
N:P ratios all fluctuated around 16:1 (Figure 10), which is the expected Redfield ratio of
marine aquatic environments(Schlesinger, 1997). The ratio is lowest in microcosms with Mytilus
and sediment because of their high DIN fluxes, and lowest in microcosms with Mercenaria and
sediment because of the low DIN fluxes in those microcosms. The closeness to the expected ratio

Ream 14
serves to validate (to a degree) the phosphate and DIN fluxes I observed. O:N ratios indicated
that were excreting the most inorganic nitrogen per unit respiration and thereby facilitating
denitrification in the presence of sediment.
Sediment Analysis
There were no significant differences in percent nitrogen in sediment between any
treatments, however, the percentage of nitrogen was slightly higher in microcosms containing
bivalves than in microcosms containing sediment (Figure 12). This is because animals inevitably
produce much more waste than bacteria respiring in microcosms containing sediment only. The
percent carbon in sediments was also not significantly different between microcosms containing
bivalves, however, the percentage of sediment C was higher in microcosms containing
Mercenaria than in microcosms containing Mytilus (Figure 13). This is likely due to the fact that
Mercenaria were buried in sediment and depositing any organic material in waste more directly
into the sediment than Mytilus in the water column. More significant differences in % soil C may
have been observable in sediment collected from depths greater than 2 cm as clams have more of
an influence at greater depths than do mussels.
Conclusions
Mercenaria appears to affect benthic nutrient cycling more than pelagic. Hard-shelled
clams seem to become stressed in the absence of sediment as this is a solely experimental
condition that are unlikely to occur in natural settings. Clams appear to add some organic matter
to the sediment, although this requires further exploration of carbon contents of sediments at
various depths to determine to what depth and extent Mercenaria supplement the sediment. In

Ream 15
my experiment, Mercenaria did not promote much denitrification in the sediment and appeared
to be slower at recycling nutrients than Mytilus. The Blue Mussel seems to affect pelagic
nutrient cycling much more than it does benthic. The presence or absence of sediment seems to
have a small effect on the feeding rates of Mytilus, as compared to its great importance for
Mercenaria, and, in general, Mytilus seemed to be more metabolically active per unit biomass
than Mercenaria. Additionally, Mytilus appeared to promote denitrification in the presence of
sediment.
The ability of bivalves to remove nutrients from overloaded estuarine systems is often
touted as a tragically under-utilized way to prevent eutrophication, however, my experiment
suggests that bivalves may accelerate nutrient cycling on short time scales. The comparison of
mussels and clams is especially interesting in that Mytilus increase DIN fluxes more than
mercenaria, but also promote denitrification more than Mercenaria. More research is certainly
warranted in order to compare M. mercenaria and M. edulis to other bivalve species in order to
understand the range of denitrification capabilities of bivalve species in nutrient overloaded
coastal areas such as Cape Cod.
Acknowledgements
This project would not have been possible without the guidance of my mentor Dr. Anne
TAs Rich McHorney, Carrie Harris, Stef Strebel, and Laura Van der Pol, and the clams, mussels,
shellfish paste food, and advice from Emma Green-Beach
Marine Resource Center.

Ream 16
L iterature C ited
Altieri, A. H. (2008). Dead zones enhance key fisheries species by providing predation refuge.
Ecology, 89, 2808-2818.
Altieri, A. H., & Witman, J. D. (2006). Local extinction of a foundation species in a hypoxic
estuary: Integrating individuals to ecosystem. Ecology, 87(3), 717-730.
Bayne, B. L. (1964). Primary and secondary settlement in mytilus edulis L.(mollusca). The
Journal of Animal Ecology, 33(3), 513-523.
Cornwell, J. C., Kemp, W. M., & Kana, T. M. (1999). Denitrification in coastal ecosystems:
Methods, environmental controls, and ecosystem level controls, a review. Aquatic Ecology,
33(1), 41-54.
Dame, R. F. (1996). Ecology of marine bivalves: An ecosystem approach. Boca Raton, FL.: CRC
Press Inc.
Lorio, W. J., & Malone, S. (1995). Biology and culture of the northern quahog clam (mercenaria
mercenaria). Southern Regional Aquaculture Center, SRAC Publication No.433
Murphy, J., & Riley, J. (1962). A modified single solution method for the determination of
phosphate in natural waters. Analytica Chimica Acta, 27, 31-36.
Officer, C. B., Smayda, T. J., & Mann, R. (1982). Benthic filter feeding: A natural eutrophication
control. Marine Ecology Progress Series, 9, 203-210.

Ream 17
Schlesinger, W. H. (1997). Biogeochemistry: An analysis of global change. Geochimica Et
Cosmochimica Acta, 56(2)
Solorzano, L. (1969). Determination of ammonia in natural waters by the phenolhypochlorite
method. Limnology and Oceanography, 14(5), 799-801.
Valiela, I., Collins, G., Kremer, J., Lajtha, K., Geist, M., Seely, B., . . . Sham, C. (1997).
Nitrogen loading from coastal watersheds to receiving estuaries: New method and
application. Ecological Applications, 7(2), 358-380.
Wood, E., Armstrong, F., & Richards, F. (1967). Determination of nitrate in sea water by
cadmium-copper reduction to nitrite. J.Mar.Biol.Assoc.UK , 47(1), 23-31.
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
16.00
1A 2A 3A 1B 2B 3B 1C 2C 3C 1D 2D 3D Average -‐
Mussels
Average -‐ Clams
Weight (g)
Pre-‐ExperimentPost-‐Experiment
Mussels Clams
Average AverageMussels Clams
Figure 1 contained Mytilus Mytilus Mercenaria only, and microcosms
Mercenaria and sediment.
Appendix. Ream 18
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 100 200 300 400 500 600
% Orig
inal Con
centratio
n
Time (minutes)Figure 2. Clearance rates of phytoplankton cells from the water column in microcosms containing Mytilus only, Mytilus and sediment, Mercenariaonly, Mercenaria and sediment, and Sediment only. Phytoplankton shown as percent of original concentration (cells/ uL) remaining. The Legend displays the slope of the line of best fit for each microcosm type.
% Original Concentration Lost/ Minute
Mytilus Only -‐0.0014
Mytilus+Sediment -‐0.0014
Mercenaria Only -‐0.0004
Mercenaria+Sediment -‐0.001
Sediment Only -‐0.00011
Ream 19
-‐300000
-‐250000
-‐200000
-‐150000
-‐100000
-‐50000
0
Oxygen Flux (
moles m
-‐2da
y-‐1 )
Microcosm Type
Figure 3. Oxygen fluxes of microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only.
Ream 20
-‐25000
-‐20000
-‐15000
-‐10000
-‐5000
0
Mytilus only Mytilus+Sediment Mercenaria only Mercenaria+Sediment
Oxygen Flux (µ
moles m
-‐2da
y -‐1g living biom
ass -‐
1 )Microcosm Type
Figure 4. Oxygen fluxes of microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only normalized by grams of living biomass per microcosm.
Ream 21
0
50000
100000
150000
200000
250000
300000
Mytilus OnlyMytilus Only (-‐sediment) Mercenaria Only
Mercenaria Only (-‐sediment)
Oxygen Flux (
moles m
-‐2 day -‐1
)Microcosm Type
Figure 5. Effect of the presence of sediment on oxygen fluxes of Mytilus and Mercenaria. The Mytilus only (-sediment) and Mercenariaonly (-sediment) bars represent the oxygen fluxes of microcosms containing bivalves and sediment with the average oxygen flux of microcosms containing only sediment subtracted.
Ream 22
0
5000
10000
15000
20000
Mytilus Only Mytilus + Sediment Mercenaria Only Mercenaria + Sediment
Sediment Only
Nitrogen
Flux (
moles m
-‐2da
y -‐1)
Microcosm Type
Nitrate
Ammonium
Figure 6. Total dissolved inorganic nitrogen fluxes in microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only.
Ream 23
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Mytilus Only Mytilus + Sediment
Mercenaria Only Mercenaria + Sediment
Sediment Only
Phosph
ate Flux (
moles m
-‐2d-‐
1 )
Microcosm Type
Figure 7. Total phosphate fluxes in microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only.
Ream 24
0
200
400
600
800
1000
1200
1400
Mytilus only Mytilus+Sediment Mercenaria only Mercenaria+Sediment
(µmoles m
-‐2da
y -‐1g living biom
ass -‐
1 )
Microcosm Type
Ammonium
Nitrate
Phosphate
Figure 8. Nutrient fluxes normalized by grams living biomass per microcosm in microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, and Mercenaria and sediment.
Ream 24
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
Mytilus Only Mytilus Only(-‐sediment)
Mercenaria Only Mercenaria Only(-‐sediment)
(moles m
-‐2da
y -‐1)
Microcosm Type
AmmoniumNitratePhosphate
Figure 9. Effect of the presence of sediment on nutrient fluxes of Mytilus and Mercenaria. The Mytilus only (-sediment) and Mercenariaonly (-sediment) bars represent the nutrient fluxes of microcosms containing bivalves and sediment with the average nutrient flux of microcosms containing only sediment subtracted.
Ream 25
0
5
10
15
20
25
Mytilus Only Mytilus + Sediment Mercenaria Only Mercenaria + Sediment
Sediment Only
N:P Ratio
Microcosm Type
Figure 10. Total dissolved inorganic nitrogen: phosphorus flux ratios in microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only.
Ream 26
0
5
10
15
20
25
Mytilus Only Mytilus + Sediment Mercenaria Only Mercenaria + Sediment
Sediment Only
O:N Ratio
Microcosm Type
Figure 11. Oxygen: total dissolved inorganic nitrogen flux ratios of microcosms containing Mytilus only, Mytilus and sediment, Mercenaria only, Mercenaria and sediment, and Sediment only.
Ream 27
0
0.005
0.01
0.015
0.02
0.025
Mytilus Mercenaria Sediment Only
% Nitrogen
Microcosm Type
Figure 12. Percentage of nitrogen in sediment samples from microcosms containing Mytilus and sediment, Mercenaria and sediment, and Sediment only.
Ream 28
0
0.05
0.1
0.15
0.2
0.25
Mytilus Mercenaria Sediment Only
% Carbo
n
Microcosm TypeFigure 13. Percentage of carbon in sediment samples from microcosms containing Mytilus and sediment, Mercenaria and sediment, and Sediment only.
Ream 29
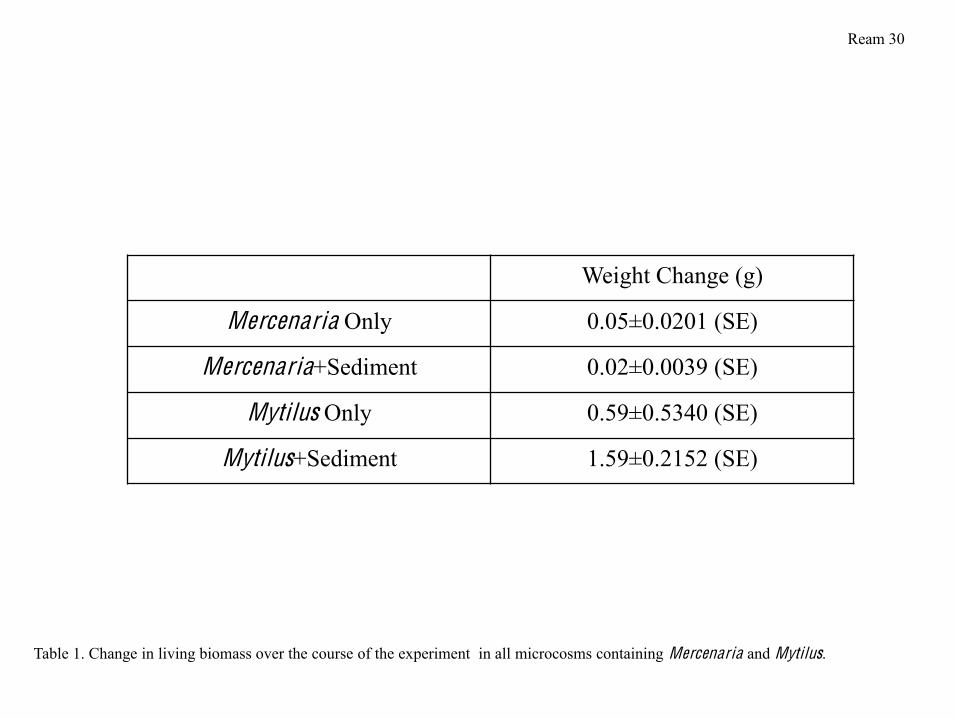
Weight Change (g)
Mercenaria Only 0.05 0.0201 (SE)
Mercenaria+Sediment 0.02 0.0039 (SE)
Mytilus Only 0.59 0.5340 (SE)
Mytilus+Sediment 1.59 0.2152 (SE)
Table 1. Change in living biomass over the course of the experiment in all microcosms containing Mercenaria and Mytilus.
Ream 30










![FeedingBehaviouroftheMussel,Mytilusedulis: NewObservations ...¥rd et al (2011... · Mytilus edulis and other suspension-feeding bivalves by, for example, Jørgensen [29], Riisg˚ard](https://static.fdocuments.in/doc/165x107/605e5acbc20a2c154c4f8c7b/feedingbehaviourofthemusselmytilusedulis-newobservations-rd-et-al-2011.jpg)







