
The Role of CC Chemokine Receptor 5 (CCR5) in Islet Allograft Rejection · CC chemokine receptor 5...
7
The Role of CC Chemokine Receptor 5 (CCR5) in Islet Allograft Rejection Reza Abdi, 1 R. Neal Smith, 2 Leila Makhlouf, 1 Nader Najafian, 1 Andrew D. Luster, 3 Hugh Auchincloss, Jr., 4 and Mohamed H. Sayegh 1 Chemokines are important regulators in the develop- ment, differentiation, and anatomic location of leuko- cytes. CC chemokine receptor 5 (CCR5) is expressed preferentially by CD4 T helper 1 (Th1) cells. We sought to determine the role of CCR5 in islet allograft rejection in a streptozotocin-induced diabetic mouse model. BALB/c islet allografts transplanted into CCR5 –/– (C57BL/6) recipients survived significantly longer (mean survival time, 38 8 days) compared with those transplanted into wild-type control mice (10 2 days; P < 0.0001). Twenty percent of islet allografts in CCR5 –/– animals without other treatment survived >90 days. In CCR5 –/– mice, intragraft mRNA expression of interleukin-4 and -5 was increased, whereas that of interferon- was decreased, corresponding to a Th2 pattern of T-cell activation in the target tissues com- pared with a Th1 pattern observed in controls. A similar Th2 response pattern was also observed in the periph- ery (splenocytes responding to donor cells) by enzyme- linked immunosorbent spot assay. We conclude that CCR5 plays an important role in orchestrating the Th1 immune response leading to islet allograft rejection. Targeting this chemokine receptor, therefore, may pro- vide a clinically useful strategy to prevent islet allograft rejection. Diabetes 51:2489 –2495, 2002 T hrough activation of the G-protein– coupled cell- surface receptor on target cells, chemokines and their receptors play a major role in the process by which leukocytes are recruited from the bloodstream into sites of inflammation, and several have been implicated in allograft rejection (1). CC chemokine receptor 5 (CCR5) is the receptor for the proinflammatory chemokines: RANTES (regulated on activation normal T-cell expressed and secreted) (CC chemokine ligand 5 [CCL5]), macrophage inflammatory protein (MIP)-1 (CCL3), and MIP-1 (CCL4) (1). Th1 cells express CCR5 and CXC chemokine receptor 3 (CXCR3) following activa- tion, whereas activated T helper 2 (Th2) cells express CCR3, CCR4, and CCR8 (2,3). Synthesis of the chemokines MIP-1, MIP-1, and RANTES has been shown to be associated with a Th1 response (4). In vitro chemotaxis assays have shown that, whereas MIP-1, MIP-1, and RANTES were efficient chemoattractants for Th1 cells to induce a dose-dependent transmigration, Th2 cells were not attracted by these chemokines (5). In heart allografts, the early expression of some chemokines, including MIP-1 and MIP-1, subsides by day 7–9 posttransplant and is replaced by a late expression of other chemokines such as inducible protein (IP)-10 (CXCL10), monokine induced by interferon- (Mig) (CXCL9) (ligands for CXCR3), and RANTES (a ligand for CCR5) (6). Met- RANTES, a CCR5 antagonist, can reduce the severity of chronic renal allograft rejection in the Lewis3 Fisher model. This effect has been attributed to blocking RANTES-induced firm adhesion of monocytes, monocyte arrest, and recruitment (7). It has recently been demon- strated that targeting CCR5 prolongs vascularized cardiac allograft survival in a mouse transplant model (8). Studies on human renal biopsies have revealed that the expression of CCR5 and its ligands is elevated in acutely rejecting human renal allografts (9). We have recently shown that susceptibility of human renal allograft recipients to acute rejection episodes is influenced by their CCR5 and CCR2 receptor genotypes (10). We now provide evidence that CCR5 plays an important role in mediating islet allograft rejection and that the blockade of CCR5 would be of value in achieving long-term islet allograft survival. RESEARCH DESIGN AND METHODS Mice. CCR5 / and CCR5 –/– mice (B6129F2/J, H-2 b ) (The Jackson Laboratory, Bar Harbor, ME) were housed under specific pathogen–free conditions in enclosed filter-top cages. The recipients had been rendered diabetic by intraperitoneal injection of 250 mg/kg streptozotocin (Upjohn, Kalamazoo, MI) and were considered diabetic when the tail vein blood glucose concentration was 250 mg/dl for 3 consecutive days. Islet isolation, transplantation, and graft removal. Islets were isolated from major histocompatibility complex (MHC)-mismatched male BALB/c (H-2 d ) donors. Pancreata were infused via the common bile duct with rodent Liberase RI (Sigma-Aldrich, St. Louis, MO) and digested for 30 min at 37°C. Islets were purified on a discontinuous Ficoll gradient (Sigma) and hand- picked under a stereomicroscope. A minimum of 500 islets was transplanted beneath the capsule of the left kidney. Daily monitoring of tail vein blood glucose was used to assess islet graft function. Rejection was defined as the return of hyperglycemia (250 mg/dl on two consecutive measurements). From the 1 Laboratory of Immunogenetics and Transplantation, Renal Divi- sion, Brigham and Women’s Hospital, Harvard Medical School, Boston, Massachusetts; the 2 Department of Pathology, Massachusetts General Hospi- tal, Harvard Medical School, Boston, Massachusetts; the 3 Division of Rheu- matology, Allergy and Immunology, Massachusetts General Hospital, Harvard Medical School, Boston, Massachusetts; and the 4 Department of Surgery, Massachusetts General Hospital, Harvard Medical School, Boston, Massachu- setts. Address correspondence and reprint requests to Mohamed H. Sayegh, Laboratory of Immunogenetics and Transplantation, Brigham and Women’s Hospital, 75 Francis St., Boston, MA 02115. E-mail: msayegh@rics. bwh.harvard.edu. Received for publication 19 December 2001 and accepted in revised form 24 April 2002. CCL, CC chemokine ligand; CCR, CC chemokine receptor; CXCR, CXC chemokine receptor; ELISPOT, enzyme-linked immunosorbent spot; IFN, interferon; IL, interleukin; IP, inducible protein; MHC, major histocompatibil- ity complex; Mig, monokine induced by interferon-; MIP, macrophage inflammatory protein; MLR, mixed lymphocyte response; RANTES, regulated on activation normal T-cell expressed and secreted; RPA, ribonuclease protection assay; Th, T helper. DIABETES, VOL. 51, AUGUST 2002 2489
Transcript of The Role of CC Chemokine Receptor 5 (CCR5) in Islet Allograft Rejection · CC chemokine receptor 5...

The Role of CC Chemokine Receptor 5 (CCR5) in IsletAllograft RejectionReza Abdi,
1R. Neal Smith,
2Leila Makhlouf,
1Nader Najafian,
1Andrew D. Luster,
3
Hugh Auchincloss, Jr.,4
and Mohamed H. Sayegh1
Chemokines are important regulators in the develop-ment, differentiation, and anatomic location of leuko-cytes. CC chemokine receptor 5 (CCR5) is expressedpreferentially by CD4� T helper 1 (Th1) cells. Wesought to determine the role of CCR5 in islet allograftrejection in a streptozotocin-induced diabetic mousemodel. BALB/c islet allografts transplanted into CCR5–/–
(C57BL/6) recipients survived significantly longer(mean survival time, 38 � 8 days) compared with thosetransplanted into wild-type control mice (10 � 2 days;P < 0.0001). Twenty percent of islet allografts inCCR5–/– animals without other treatment survived >90days. In CCR5–/– mice, intragraft mRNA expression ofinterleukin-4 and -5 was increased, whereas that ofinterferon-� was decreased, corresponding to a Th2pattern of T-cell activation in the target tissues com-pared with a Th1 pattern observed in controls. A similarTh2 response pattern was also observed in the periph-ery (splenocytes responding to donor cells) by enzyme-linked immunosorbent spot assay. We conclude thatCCR5 plays an important role in orchestrating the Th1immune response leading to islet allograft rejection.Targeting this chemokine receptor, therefore, may pro-vide a clinically useful strategy to prevent islet allograftrejection. Diabetes 51:2489–2495, 2002
Through activation of the G-protein–coupled cell-surface receptor on target cells, chemokines andtheir receptors play a major role in the processby which leukocytes are recruited from the
bloodstream into sites of inflammation, and several havebeen implicated in allograft rejection (1). CC chemokinereceptor 5 (CCR5) is the receptor for the proinflammatory
chemokines: RANTES (regulated on activation normalT-cell expressed and secreted) (CC chemokine ligand 5[CCL5]), macrophage inflammatory protein (MIP)-1�(CCL3), and MIP-1� (CCL4) (1). Th1 cells express CCR5and CXC chemokine receptor 3 (CXCR3) following activa-tion, whereas activated T helper 2 (Th2) cells expressCCR3, CCR4, and CCR8 (2,3). Synthesis of the chemokinesMIP-1�, MIP-1�, and RANTES has been shown to beassociated with a Th1 response (4). In vitro chemotaxisassays have shown that, whereas MIP-1�, MIP-1�, andRANTES were efficient chemoattractants for Th1 cells toinduce a dose-dependent transmigration, Th2 cells werenot attracted by these chemokines (5). In heart allografts,the early expression of some chemokines, includingMIP-1� and MIP-1�, subsides by day 7–9 posttransplantand is replaced by a late expression of other chemokinessuch as inducible protein (IP)-10 (CXCL10), monokineinduced by interferon-� (Mig) (CXCL9) (ligands forCXCR3), and RANTES (a ligand for CCR5) (6). Met-RANTES, a CCR5 antagonist, can reduce the severity ofchronic renal allograft rejection in the Lewis3Fishermodel. This effect has been attributed to blockingRANTES-induced firm adhesion of monocytes, monocytearrest, and recruitment (7). It has recently been demon-strated that targeting CCR5 prolongs vascularized cardiacallograft survival in a mouse transplant model (8). Studieson human renal biopsies have revealed that the expressionof CCR5 and its ligands is elevated in acutely rejectinghuman renal allografts (9). We have recently shown thatsusceptibility of human renal allograft recipients to acuterejection episodes is influenced by their CCR5 and CCR2receptor genotypes (10). We now provide evidence thatCCR5 plays an important role in mediating islet allograftrejection and that the blockade of CCR5 would be of valuein achieving long-term islet allograft survival.
RESEARCH DESIGN AND METHODS
Mice. CCR5�/� and CCR5–/– mice (B6129F2/J, H-2b) (The Jackson Laboratory,Bar Harbor, ME) were housed under specific pathogen–free conditions inenclosed filter-top cages. The recipients had been rendered diabetic byintraperitoneal injection of 250 mg/kg streptozotocin (Upjohn, Kalamazoo, MI)and were considered diabetic when the tail vein blood glucose concentrationwas �250 mg/dl for 3 consecutive days.Islet isolation, transplantation, and graft removal. Islets were isolatedfrom major histocompatibility complex (MHC)-mismatched male BALB/c(H-2d) donors. Pancreata were infused via the common bile duct with rodentLiberase RI (Sigma-Aldrich, St. Louis, MO) and digested for 30 min at 37°C.Islets were purified on a discontinuous Ficoll gradient (Sigma) and hand-picked under a stereomicroscope. A minimum of 500 islets was transplantedbeneath the capsule of the left kidney. Daily monitoring of tail vein bloodglucose was used to assess islet graft function. Rejection was defined as thereturn of hyperglycemia (�250 mg/dl on two consecutive measurements).
From the 1Laboratory of Immunogenetics and Transplantation, Renal Divi-sion, Brigham and Women’s Hospital, Harvard Medical School, Boston,Massachusetts; the 2Department of Pathology, Massachusetts General Hospi-tal, Harvard Medical School, Boston, Massachusetts; the 3Division of Rheu-matology, Allergy and Immunology, Massachusetts General Hospital, HarvardMedical School, Boston, Massachusetts; and the 4Department of Surgery,Massachusetts General Hospital, Harvard Medical School, Boston, Massachu-setts.
Address correspondence and reprint requests to Mohamed H. Sayegh,Laboratory of Immunogenetics and Transplantation, Brigham and Women’sHospital, 75 Francis St., Boston, MA 02115. E-mail: [email protected].
Received for publication 19 December 2001 and accepted in revised form 24April 2002.
CCL, CC chemokine ligand; CCR, CC chemokine receptor; CXCR, CXCchemokine receptor; ELISPOT, enzyme-linked immunosorbent spot; IFN,interferon; IL, interleukin; IP, inducible protein; MHC, major histocompatibil-ity complex; Mig, monokine induced by interferon-�; MIP, macrophageinflammatory protein; MLR, mixed lymphocyte response; RANTES, regulatedon activation normal T-cell expressed and secreted; RPA, ribonucleaseprotection assay; Th, T helper.
DIABETES, VOL. 51, AUGUST 2002 2489
Occurrence of hyperglycemia after removal of the graft was used to confirmlong-term allograft function (�90 days). The grafts were removed for patho-logical examination under a surgical microscope by excising the edge of thecapsule containing the islets.Mixed lymphocyte response. In mixed lymphocyte response (MLR) assays,2 � 105 of each responder and irradiated stimulator splenocytes were culturedin complete RPMI medium (90% RPMI 1640, 10% FCS, [Sigma] with L-glutamine and penicillin/streptomycin [BioWhittaker, Walkersville, MD], and50 mmol/l 2-mercaptoethanol [Sigma]) in 96-well U-bottom plates (Corning-Costar, Cambridge, MA) (six wells per experimental group). Backgroundproliferation was measured by incubating cells with culture medium alone.Cultures were pulsed with 1 �Ci of tritiated thymidine after 72 h of incubation.The samples were harvested after 12 h (11).ELISPOT assay. The enzyme-linked immunosorbent spot (ELISPOT) assayhas been recently described by us (11,12). Briefly, ELISAspot plates (CellularTechnology, Cleveland, OH) were coated with capture antibodies againstinterleukin (IL)-4, IL-5, IL-10, or interferon (IFN)-� (PharMingen, San Diego,CA) in PBS and left overnight at 4°C. The plates were blocked for 1 h with PBScontaining 1% BSA and were washed with PBS. A total of 1 � 106 splenocyteswere added to each well in 100 �l complete RPMI medium in the presence ofthe same number of irradiated syngeneic or allogeneic splenocytes. After 48 h,the plates were washed, biotinylated detection antibodies were added, and theplates were maintained at 4°C for an additional overnight incubation. Afteradditional washing, horseradish peroxidase conjugate (Dako, Glostrup, Den-mark) was added for 2 h at room temperature. To develop the spots,aminoethylcarbazole (10 mg/ml in N,N-dimethylformamide; Pierre Chemicals,Rockland, IL) freshly prepared in 0.1 mol/l sodium acetate buffer (pH 5.0) andmixed with 30% H2O2 was added to each well (200 �l per well). The resultingspots were counted on a computer-assisted ELISAspot image analyzer (Cel-lular Technology). The results were calculated as cytokine-producing cells permillion splenocytes.Cytokine, chemokine, and chemokine receptor expression. The ribonu-clease protection assay (RPA) was performed as follows. Islet RNA wasextracted in guanidine-thiocyanate with acid phenol/chloroform extractionand alcohol precipitation. RNA integrity was confirmed by agarose gelelectrophoresis and quantitated by optical density measurement (260 nm).Intragraft expression of chemokines, chemokine receptors, and cytokines wasassessed using the custom-designed Riboquant system (PharMingen). Sam-ples of total RNA (5 �g) were hybridized with 3 � 105 cpm/�l of probe labeledwith [32P]UTP (3,000 Ci/mmol; NEN Life Science Products) at 56°C overnight.RNase A and RNase Ti (PharMingen) digestion were carried out at 37°C for 45min. RNA bands were quantitated by densitometric analysis with NIH Imagesoftware (National Institutes of Health, Bethesda, MD), PC environment byScion Corp. (Windows 95 and NT). The results were normalized for L32 andGAPDH gene expressions.Immunopathology. One-half of each islet graft was embedded in OCT, frozenat �20°C, and kept at –70°C until sectioning. Cryostat sections (2 �m) werefixed with acetone and dried. Indirect frozen section immunohistochemistrywas performed using primary antibodies for T cells (CD4, GK1.5, and CD8,53–6.7; PharMingen) and macrophages (F4/80). One-half of each graft wasalso fixed and paraffin embedded. Islets were stained for insulin and glucagonusing guinea pig anti-insulin (1:10 dilution; Dako) and biotin-labeled goatanti-guinea pig Ig (1:200; Vector) and developed with Vector ABC (13). Eachspecimen was evaluated at a minimum of three different levels of sectioning.Images were taken with a RT Spot Camera (Diagnostic Instruments) andimported into Adobe Photoshop at 40� magnification.Statistical analyses. Data were compared by nonparametric analysis (Instatsoftware; GraphPad, San Diego, CA) using the log-rank test for graft survivaldata and Mann-Whitney test for data from proliferation assays and expressionstudies. Differences in graft survival were analyzed by a Kaplan-Meier test.
RESULTS
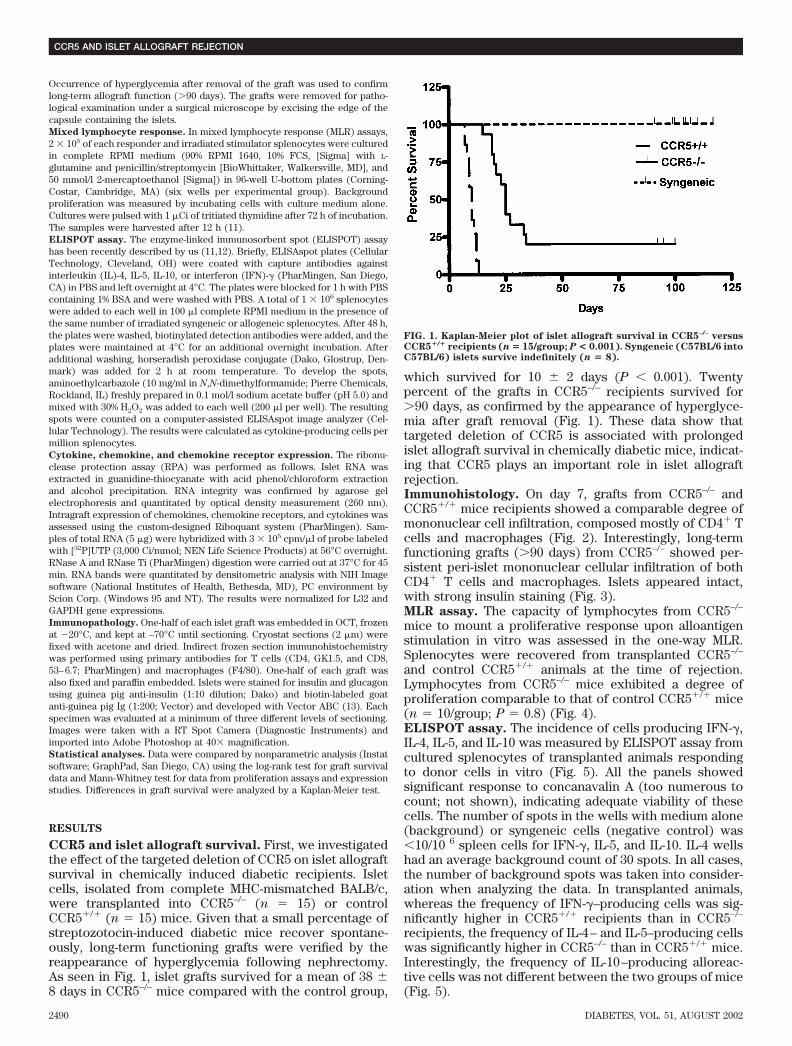
CCR5 and islet allograft survival. First, we investigatedthe effect of the targeted deletion of CCR5 on islet allograftsurvival in chemically induced diabetic recipients. Isletcells, isolated from complete MHC-mismatched BALB/c,were transplanted into CCR5–/– (n 15) or controlCCR5�/� (n 15) mice. Given that a small percentage ofstreptozotocin-induced diabetic mice recover spontane-ously, long-term functioning grafts were verified by thereappearance of hyperglycemia following nephrectomy.As seen in Fig. 1, islet grafts survived for a mean of 38 8 days in CCR5–/– mice compared with the control group,
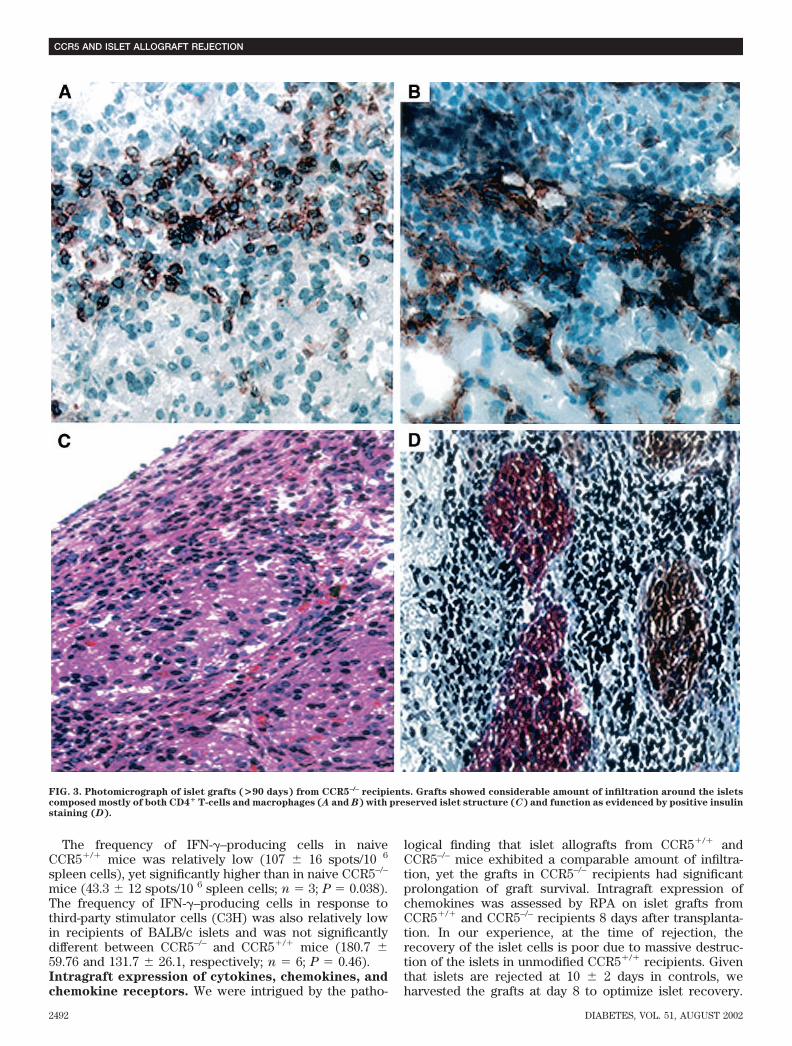
which survived for 10 2 days (P � 0.001). Twentypercent of the grafts in CCR5–/– recipients survived for�90 days, as confirmed by the appearance of hyperglyce-mia after graft removal (Fig. 1). These data show thattargeted deletion of CCR5 is associated with prolongedislet allograft survival in chemically diabetic mice, indicat-ing that CCR5 plays an important role in islet allograftrejection.Immunohistology. On day 7, grafts from CCR5–/– andCCR5�/� mice recipients showed a comparable degree ofmononuclear cell infiltration, composed mostly of CD4� Tcells and macrophages (Fig. 2). Interestingly, long-termfunctioning grafts (�90 days) from CCR5–/– showed per-sistent peri-islet mononuclear cellular infiltration of bothCD4� T cells and macrophages. Islets appeared intact,with strong insulin staining (Fig. 3).MLR assay. The capacity of lymphocytes from CCR5–/–
mice to mount a proliferative response upon alloantigenstimulation in vitro was assessed in the one-way MLR.Splenocytes were recovered from transplanted CCR5–/–
and control CCR5�/� animals at the time of rejection.Lymphocytes from CCR5–/– mice exhibited a degree ofproliferation comparable to that of control CCR5�/� mice(n 10/group; P 0.8) (Fig. 4).ELISPOT assay. The incidence of cells producing IFN-�,IL-4, IL-5, and IL-10 was measured by ELISPOT assay fromcultured splenocytes of transplanted animals respondingto donor cells in vitro (Fig. 5). All the panels showedsignificant response to concanavalin A (too numerous tocount; not shown), indicating adequate viability of thesecells. The number of spots in the wells with medium alone(background) or syngeneic cells (negative control) was�10/10 6 spleen cells for IFN-�, IL-5, and IL-10. IL-4 wellshad an average background count of 30 spots. In all cases,the number of background spots was taken into consider-ation when analyzing the data. In transplanted animals,whereas the frequency of IFN-�–producing cells was sig-nificantly higher in CCR5�/� recipients than in CCR5–/–
recipients, the frequency of IL-4– and IL-5–producing cellswas significantly higher in CCR5–/– than in CCR5�/� mice.Interestingly, the frequency of IL-10–producing alloreac-tive cells was not different between the two groups of mice(Fig. 5).
FIG. 1. Kaplan-Meier plot of islet allograft survival in CCR5–/– versusCCR5�/� recipients (n � 15/group; P < 0.001). Syngeneic (C57BL/6 intoC57BL/6) islets survive indefinitely (n � 8).
CCR5 AND ISLET ALLOGRAFT REJECTION
2490 DIABETES, VOL. 51, AUGUST 2002
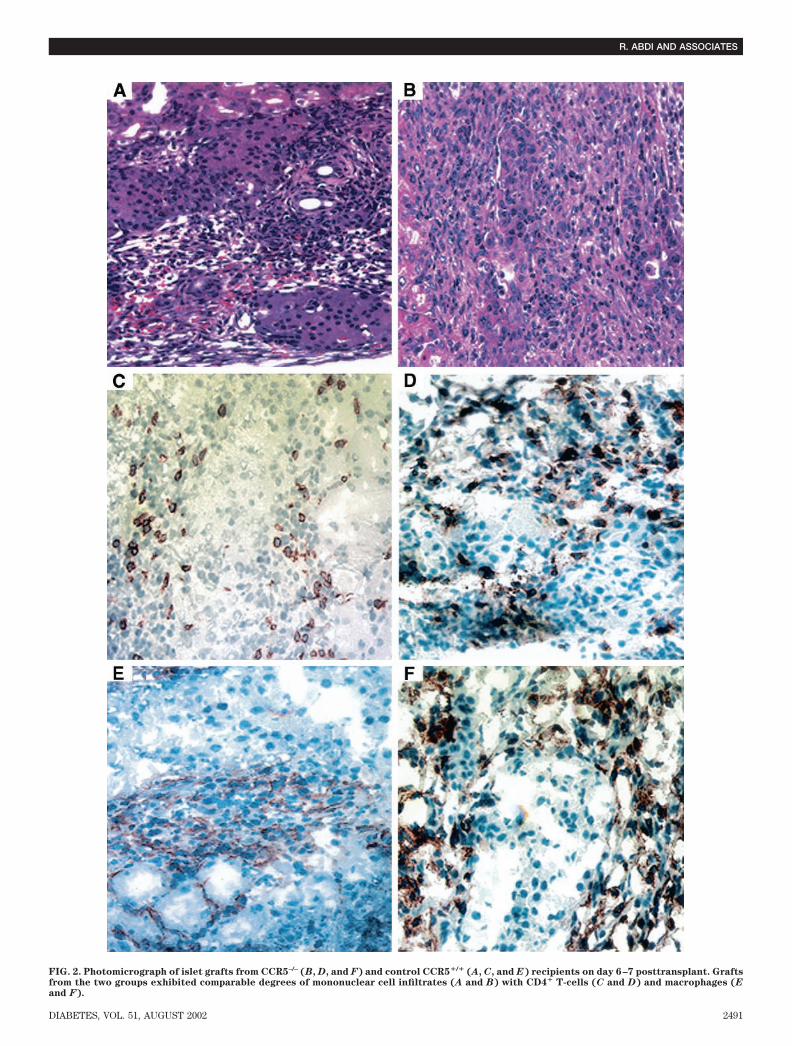
FIG. 2. Photomicrograph of islet grafts from CCR5–/– (B, D, and F) and control CCR5�/� (A, C, and E) recipients on day 6–7 posttransplant. Graftsfrom the two groups exhibited comparable degrees of mononuclear cell infiltrates (A and B) with CD4� T-cells (C and D) and macrophages (E
and F).
R. ABDI AND ASSOCIATES
DIABETES, VOL. 51, AUGUST 2002 2491
The frequency of IFN-�–producing cells in naiveCCR5�/� mice was relatively low (107 16 spots/10 6
spleen cells), yet significantly higher than in naive CCR5–/–
mice (43.3 12 spots/10 6 spleen cells; n 3; P 0.038).The frequency of IFN-�–producing cells in response tothird-party stimulator cells (C3H) was also relatively lowin recipients of BALB/c islets and was not significantlydifferent between CCR5–/– and CCR5�/� mice (180.7 59.76 and 131.7 26.1, respectively; n 6; P 0.46).Intragraft expression of cytokines, chemokines, and
chemokine receptors. We were intrigued by the patho-
logical finding that islet allografts from CCR5�/� andCCR5–/– mice exhibited a comparable amount of infiltra-tion, yet the grafts in CCR5–/– recipients had significantprolongation of graft survival. Intragraft expression ofchemokines was assessed by RPA on islet grafts fromCCR5�/� and CCR5–/– recipients 8 days after transplanta-tion. In our experience, at the time of rejection, therecovery of the islet cells is poor due to massive destruc-tion of the islets in unmodified CCR5�/� recipients. Giventhat islets are rejected at 10 2 days in controls, weharvested the grafts at day 8 to optimize islet recovery.
FIG. 3. Photomicrograph of islet grafts (>90 days) from CCR5–/– recipients. Grafts showed considerable amount of infiltration around the isletscomposed mostly of both CD4� T-cells and macrophages (A and B) with preserved islet structure (C) and function as evidenced by positive insulinstaining (D).
CCR5 AND ISLET ALLOGRAFT REJECTION
2492 DIABETES, VOL. 51, AUGUST 2002
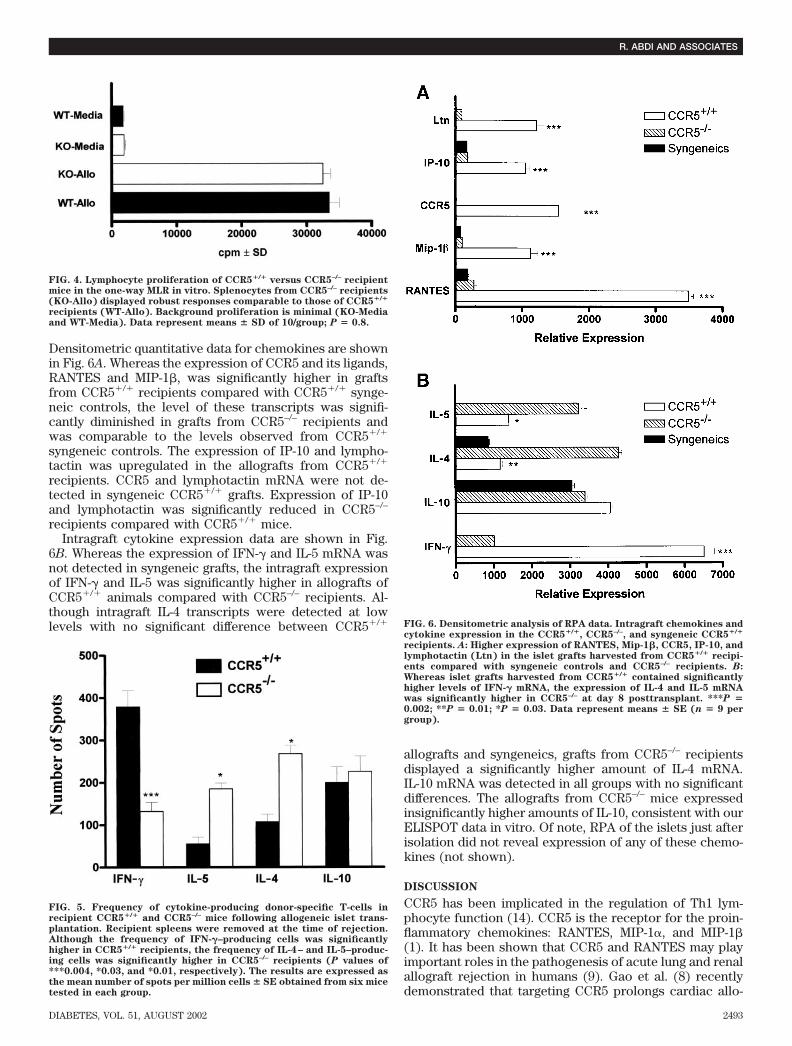
Densitometric quantitative data for chemokines are shownin Fig. 6A. Whereas the expression of CCR5 and its ligands,RANTES and MIP-1�, was significantly higher in graftsfrom CCR5�/� recipients compared with CCR5�/� synge-neic controls, the level of these transcripts was signifi-cantly diminished in grafts from CCR5–/– recipients andwas comparable to the levels observed from CCR5�/�
syngeneic controls. The expression of IP-10 and lympho-tactin was upregulated in the allografts from CCR5�/�
recipients. CCR5 and lymphotactin mRNA were not de-tected in syngeneic CCR5�/� grafts. Expression of IP-10and lymphotactin was significantly reduced in CCR5–/–
recipients compared with CCR5�/� mice.Intragraft cytokine expression data are shown in Fig.
6B. Whereas the expression of IFN-� and IL-5 mRNA wasnot detected in syngeneic grafts, the intragraft expressionof IFN-� and IL-5 was significantly higher in allografts ofCCR5�/� animals compared with CCR5–/– recipients. Al-though intragraft IL-4 transcripts were detected at lowlevels with no significant difference between CCR5�/�
allografts and syngeneics, grafts from CCR5–/– recipientsdisplayed a significantly higher amount of IL-4 mRNA.IL-10 mRNA was detected in all groups with no significantdifferences. The allografts from CCR5–/– mice expressedinsignificantly higher amounts of IL-10, consistent with ourELISPOT data in vitro. Of note, RPA of the islets just afterisolation did not reveal expression of any of these chemo-kines (not shown).
DISCUSSION
CCR5 has been implicated in the regulation of Th1 lym-phocyte function (14). CCR5 is the receptor for the proin-flammatory chemokines: RANTES, MIP-1�, and MIP-1�(1). It has been shown that CCR5 and RANTES may playimportant roles in the pathogenesis of acute lung and renalallograft rejection in humans (9). Gao et al. (8) recentlydemonstrated that targeting CCR5 prolongs cardiac allo-
FIG. 4. Lymphocyte proliferation of CCR5�/� versus CCR5–/– recipientmice in the one-way MLR in vitro. Splenocytes from CCR5–/– recipients(KO-Allo) displayed robust responses comparable to those of CCR5�/�
recipients (WT-Allo). Background proliferation is minimal (KO-Mediaand WT-Media). Data represent means � SD of 10/group; P � 0.8.
FIG. 5. Frequency of cytokine-producing donor-specific T-cells inrecipient CCR5�/� and CCR5–/– mice following allogeneic islet trans-plantation. Recipient spleens were removed at the time of rejection.Although the frequency of IFN-�–producing cells was significantlyhigher in CCR5�/� recipients, the frequency of IL-4– and IL-5–produc-ing cells was significantly higher in CCR5–/– recipients (P values of***0.004, *0.03, and *0.01, respectively). The results are expressed asthe mean number of spots per million cells � SE obtained from six micetested in each group.
FIG. 6. Densitometric analysis of RPA data. Intragraft chemokines andcytokine expression in the CCR5�/�, CCR5–/–, and syngeneic CCR5�/�
recipients. A: Higher expression of RANTES, Mip-1�, CCR5, IP-10, andlymphotactin (Ltn) in the islet grafts harvested from CCR5�/� recipi-ents compared with syngeneic controls and CCR5–/– recipients. B:Whereas islet grafts harvested from CCR5�/� contained significantlyhigher levels of IFN-� mRNA, the expression of IL-4 and IL-5 mRNAwas significantly higher in CCR5–/– at day 8 posttransplant. ***P �0.002; **P � 0.01; *P � 0.03. Data represent means � SE (n � 9 pergroup).
R. ABDI AND ASSOCIATES
DIABETES, VOL. 51, AUGUST 2002 2493

graft survival in the mouse model. However, given a lackof intragraft IL-4, IL-5, or IL-10 mRNA in CCR5–/– recipi-ents, no evidence for immune deviation toward an intra-graft Th2 regulatory cell population was apparent in theirstudy (8). Studies on murine and human heart and skinmodels indicate that, in the process of acute rejection, thetemporal expression of critical chemokines varies amongdifferent organs or tissues. For instance, CXCR3 and itsligands, IP-10 and Mig, are expressed by the T-cells infil-trating lung and heart allografts and mediate chemotaxisof T-cells at sites of rejection (15,16) (17). TargetingCXCR3 and its ligands has been shown to be associatedwith a significant prolongation of cardiac allograft survival(6,18). Hence, each organ or tissue may require a uniqueset of chemokines to generate acute rejection (6,18).Despite the documented presence of chemokine in exper-imental and clinical allografts during acute rejection, therole of specific chemokines in the rejection process of isletallografts remains unclear. In our study, CCR5 mRNA wasnot detected in the grafts from syngeneic CCR5�/� recip-ients, and RANTES and MIP-1� expression in CCR5�/�
recipients was significantly higher than that noted insyngeneic controls and CCR5–/– recipients. The formerfinding indicates that production of CCR5 and its ligands isdependent on the infiltrating alloantigen-specific T-cells.Grafts from CCR5–/– recipients also exhibited significantlylower expression of lymphotactin and IP-10. Islet graftrejection in wild-type mice was characterized by a Th1-type response with a strong induction of IFN-� mRNAexpression. In contrast, the prolonged islet allograft sur-vival observed in CCR5–/– mice was associated with anincrease in intragraft and peripheral expression of IL-4 andIL-5 and a reduction in expression of IFN-�. CCR5 ligandsinduce selective migration of Th1 cells in chemotaxisassays (5,19). Therefore, absence of CCR5 may explain theshift toward Th2 immune response in our study, and thatthe infiltrating cells in the CCR5–/– grafts represent cells ofa predominantly Th2 phenotype. It is possible that thesmall amount of RANTES expressed in the CCR5–/– recip-ients may signal through CCR3 in the absence of CCR5,preferentially attracting Th2 cells (20). Decreased IFN-�and increased IL-4 expression in turn may have led tosuppressed macrophage activation (21,22). Because IFN-�induces CCR5 ligands, lower expression of IFN-� inCCR5–/– may explain the lower expression of CCR5 ligandsin this model (4). Furthermore, the ability to produceIFN-� has been shown to be critical for efficient CD8�
T-cell–mediated rejection of islet allografts (23). Using adextran sodium sulfate–induced colitis model, CCR5�/�
mice were characterized by a Th1-type response with astrong induction of IFN-� mRNA expression. In contrast,the reduced colonic damage observed in CCR5–/– mice wasassociated with an increase in IL-4 and IL-5 mRNA expres-sion, a reduction in IFN-� mRNA expression, and anincrease in the proportion of CD4� T-cells in the laminapropria (24).
Interestingly, the intragraft mononuclear cell infiltrate inCCR5–/– mice was comparable to that in wild-type recipi-ents. Thus, the immunosuppressive effect of the targeteddeletion of CCR5 is not necessarily related to the general-ized recruitment and migration of CD4� T-cells and mac-rophages into the graft. One could speculate that the Th2
switch observed in the graft and periphery of CCR5-deficient animals may be protective against islet allograftdestruction. However, a regulatory function of Th2 cells inalloimmune responses remains controversial (25–27). An-imals lacking the Th2 cytokine IL-4 can accept allografts(28), although this is not a universal finding in all models(29). In addition, Li et al. (30) showed that a Th2 switch isassociated with graft acceptance in minor but not majormismatched graft recipients. The finding that CCR5–/– miceultimately reject their islet graft indicates that a Th2switch alone is not sufficient for induction of long-termgraft acceptance. Importantly, long-term functioning graftsthat survive in CCR5–/– mice in the presence of cellularinfiltration indicate that not all types of inflammation areidentical and that the amount of inflammation may notpredict outcome. This infiltration appears to resembleother autoimmune settings, such as diabetes, in whichT-cell recruitment does not universally lead to organdamage. Identifying the nature of this nonpathogenicinfiltration may be extremely crucial in understandingsimilar presentations in various autoimmune models suchas diabetes. In conclusion, our results demonstrate thatthe lack of CCR5 signaling results in prolongation of isletallograft survival and a switch to Th2 response. Targetingof CCR5 may have clinical application as a novel approachto clinical management of islet allograft recipients.
ACKNOWLEDGMENTS
This work was supported in part by the Juvenile DiabetesResearch Foundation (JDRF) Center for Islet Transplan-tation at Harvard Medical School.
We thank the Islet Care Laboratory of the JDRF Centerfor Islet Transplantation for providing mouse islets.
REFERENCES
1. Luster AD: Chemokines: chemotactic cytokines that mediate inflammation(Review). N Engl J Med 338:436–445, 1998
2. Sallusto F, Lanzavecchia A, Mackay CR: Chemokines and chemokinereceptors in T-cell priming and Th1/Th2-mediated responses (Review).Immunol Today 19:568–574, 1998
3. Baggiolini M: Chemokines and leukocyte traffic (Review). Nature 392:565–568, 1998
4. Schrum S, Probst P, Fleischer B, Zipfel PF: Synthesis of the CC-chemo-kines MIP-1alpha, MIP-1beta, and RANTES is associated with a type 1immune response. J Immunol 157:3598–3604, 1996
5. Siveke JT, Hamann A: T helper 1 and T helper 2 cells respond differentiallyto chemokines. J Immunol 160:550–554, 1998
6. Miura M, Morita K, Kobayashi H, Hamilton TA, Burdick MD, Strieter RM,Fairchild RL: Monokine induced by IFN-gamma is a dominant factordirecting T cells into murine cardiac allografts during acute rejection.J Immunol 167:3494–3504, 2001
7. Grone HJ, Weber C, Weber KS, Grone EF, Rabelink T, Klier CM, Wells TN,Proudfood AE, Schlondorff D, Nelson PJ: Met-RANTES reduces vascularand tubular damage during acute renal transplant rejection: blockingmonocyte arrest and recruitment. FASEB J 13:1371–1383, 1999
8. Gao W, Faia KL, Csizmadia V, Smiley ST, Soler D, King JA, Danoff TM,Hancock WW: Beneficial effects of targeting CCR5 in allograft recipients.Transplantation 72:1199–1205, 2001
9. Segerer S, Cui Y, Eitner F, Goodpaster T, Hudkins KL, Mack M, Cartron JP,Colin Y, Schlondorff D, Alpers CE: Expression of chemokines and chemo-kine receptors during human renal transplant rejection. Am J Kidney Dis
37:518–531, 200110. Abdi R, Tran TB, Sahagun-Ruiz A, Murphy PM, Brenner BM, Milford EL,
McDermott DH: Chemokine receptor polymorphism and risk of acuterejection in human renal transplantation. J Am Soc Nephrol 13:754–758,2002
11. Yamada A, Kishimoto K, Dong VM, Sho M, Salama AD, Anosova NG,Benichou G, Mandelbrot DA, Sharpe AH, Turka LA, Auchincloss H Jr,
CCR5 AND ISLET ALLOGRAFT REJECTION
2494 DIABETES, VOL. 51, AUGUST 2002

Sayegh MH: CD28-independent costimulation of T cells in alloimmuneresponses. J Immunol 167:140–146, 2001
12. Kishimoto K, Dong VM, Issazadeh S, Fedoseyeva EV, Waaga AM, YamadaA, Sho M, Benichou G, Auchincloss H Jr, Grusby MJ, Khoury SJ, SayeghMH: The role of CD154-CD40 versus CD28–B7 costimulatory pathways inregulating allogeneic Th1 and Th2 responses in vivo. J Clin Invest
106:63–72, 200013. Friedman T, Shimizu A, Smith RN, Colvin RB, Seebach JD, Sachs DH,
Iacomini J: Human CD4� T cells mediate rejection of porcine xenografts.J Immunol 162:5256–5262, 1999
14. Loetscher P, Uguccioni M, Bordoli L, Baggiolini M, Moser B, Chizzolini C,Dayer JM: CCR5 is characteristic of Th1 lymphocytes. Nature 391:344–345,1998
15. Kondo T, Novick AC, Toma H, Fairchild RL: Induction of chemokine geneexpression during allogeneic skin graft rejection. Transplantation 61:1750–1757, 1996
16. Agostini C, Calabrese F, Rea F, Facco M, Tosoni A, Loy M, Binotto G,Valente M, Trentin L, Semenzato G: Cxcr3 and its ligand CXCL10 areexpressed by inflammatory cells infiltrating lung allografts and mediatechemotaxis of T cells at sites of rejection. Am J Pathol 158:1703–1711, 2001
17. Hancock WW, Gao W, Csizmadia V, Faia KL, Shemmeri N, Luster AD:Donor-derived IP-10 initiates development of acute allograft rejection. J
Exp Med 193:975–980, 200118. Hancock WW, Lu B, Gao W, Csizmadia V, Faia K, King JA, Smiley ST, Ling
M, Gerard NP, Gerard C: Requirement of the chemokine receptor CXCR3for acute allograft rejection. J Exp Med 192:1515–1520, 2000
19. Syrbe U, Siveke J, Hamann A: Th1/Th2 subsets: distinct differences inhoming and chemokine receptor expression? (Review). Springer Semin
Immunopathol 21:263–285, 199920. Rollins BJ: Chemokines (Review). Blood 90:909–928, 199721. Bonecchi R, Bianchi G, Bordignon PP, D’Ambrosio D, Lang R, Borsatti A,
Sozzani S, Allavena P, Gray PA, Mantovani A, Sinigaglia F: Differentialexpression of chemokine receptors and chemotactic responsiveness oftype 1 T helper cells (Th1s) and Th2s. J Exp Med 187:129–134, 1998
22. Gerber BO, Zanni MP, Uguccioni M, Loetscher M, Mackay CR, Pichler WJ,
Yawalkar N, Baggiolini M, Moser B: Functional expression of the eotaxinreceptor CCR3 in T lymphocytes co-localizing with eosinophils. Curr Biol
7:836–843, 199723. Diamond AS, Gill RG: An essential contribution by IFN-gamma to CD8� T
cell-mediated rejection of pancreatic islet allografts. J Immunol 165:247–255, 2000
24. Andres PG, Beck PL, Mizoguchi E, Mizoguchi A, Bhan AK, Dawson T,Kuziel WA, Maeda N, MacDermott RP, Podolsky DK, Reinecker HC: Micewith a selective deletion of the CC chemokine receptors 5 or 2 areprotected from dextran sodium sulfate-mediated colitis: lack of CC che-mokine receptor 5 expression results in a NK1.1� lymphocyte-associatedTh2-type immune response in the intestine. J Immunol 164:6303–6312,2000
25. Strom TB, Roy-Chaudhury P, Manfro R, Zheng XX, Nickerson PW, Wood K,Bushell A: The Th1/Th2 paradigm and the allograft response (Review).Curr Opin Immunol 8:688–693, 1996
26. Waaga AM, Gasser M, Kist-van Holthe JE, Najafian N, Muller A, Vella JP,Womer KL, Chandraker A, Khoury SJ, Sayegh MH: Regulatory functions ofself-restricted MHC class II allopeptide-specific Th2 clones in vivo. J Clin
Invest 107:909–916, 200127. Sayegh MH, Akalin E, Hancock WW, Russell ME, Carpenter CB, Linsley PS,
Turka LA: CD28–B7 blockade after alloantigenic challenge in vivo inhibitsTh1 cytokines but spares Th2. J Exp Med 181:1869–1874, 1995
28. Lakkis FG, Konieczny BT, Saleem S, Baddoura FK, Linsley PS, AlexanderDZ, Lowry RP, Pearson TC, Larsen CP: Blocking the CD28–B7 T cellcostimulation pathway induces long term cardiac allograft acceptance inthe absence of IL-4. J Immunol 158:2443–2448, 1997
29. Sirak JH, Orosz CG, Roopenian DC, Wakely E, VanBuskirk AM: Cardiacallograft tolerance: failure to develop in interleukin-4-deficient mice cor-relates with unusual allosensitization patterns. Transplantation 65:1352–1356, 1998
30. Li XC, Zand MS, Li Y, Zheng XX, Strom TB: On histocompatibility barriers,Th1 to Th2 immune deviation, and the nature of the allograft responses.J Immunol 161:2241–2247, 1998
R. ABDI AND ASSOCIATES
DIABETES, VOL. 51, AUGUST 2002 2495


















