Teaching Argumentative Writing Using the Common Core State Standards David Pook Susan Pimentel.

The Reverse Transcriptase/RNA Maturase Protein MatR IsRequired for the Splicing of Various Group II Introns inBrassicaceae Mitochondria
Laure D. Sultan,a,1 Daria Mileshina,b,1 Felix Grewe,c,2 Katarzyna Rolle,d Sivan Abudraham,a Paweł Głodowicz,d
Adnan Khan Niazi,b Ido Keren,a Sofia Shevtsov,a Liron Klipcan,e,3 Jan Barciszewski,d Jeffrey P. Mower,c
André Dietrich,b and Oren Ostersetzer-Birana,4
a Department of Plant and Environmental Sciences, The Alexander Silberman Institute of Life Sciences, The Hebrew University ofJerusalem, Givat-Ram, Jerusalem 91904, Israelb Institut de Biologie Moléculaire des Plantes, CNRS and Université de Strasbourg, 67084 Strasbourg, FrancecDepartment of Agronomy and Horticulture, University of Nebraska, Lincoln, Nebraska 68588d Institute of Bioorganic Chemistry, Polish Academy of Sciences, 61-704 Poznan, PolandeDepartment of Structural Biology, Weizmann Institute of Science, Rehovot 7610001, Israel
ORCID IDs: 0000-0002-6090-519X (D.M.); 0000-0003-1140-7095 (P.G.); 0000-0002-9006-7377 (A.K.N.); 0000-0002-1872-8337 (I.K.);0000-0002-1501-5809 (J.P.M.); 0000-0003-2197-7729 (O.O.-B.)
Group II introns are large catalytic RNAs that are ancestrally related to nuclear spliceosomal introns. Sequencescorresponding to group II RNAs are found in many prokaryotes and are particularly prevalent within plants organellargenomes. Proteins encoded within the introns themselves (maturases) facilitate the splicing of their own host pre-RNAs.Mitochondrial introns in plants have diverged considerably in sequence and have lost their maturases. In angiosperms, onlya single maturase has been retained in the mitochondrial DNA: the matR gene found within NADH dehydrogenase 1 (nad1)intron 4. Its conservation across land plants and RNA editing events, which restore conserved amino acids, indicates thatmatR encodes a functional protein. However, the biological role of MatR remains unclear. Here, we performed an in vivoinvestigation of the roles of MatR in Brassicaceae. Directed knockdown of matR expression via synthetically designedribozymes altered the processing of various introns, including nad1 i4. Pull-down experiments further indicated that MatR isassociated with nad1 i4 and several other intron-containing pre-mRNAs. MatR may thus represent an intermediate link in thegradual evolutionary transition from the intron-specific maturases in bacteria into their versatile spliceosomal descendants inthe nucleus. The similarity between maturases and the core spliceosomal Prp8 protein further supports this intriguing theory.
INTRODUCTION
Mitochondria play central roles in cellular energy production andmetabolism. As progenies from a free-living prokaryotic sym-biont, mitochondria contain their own genomes (mtDNAs),ribosomes, and proteins. While the mtDNAs in animal cells aretypically small (16 to 19 kb), encoding 37 or fewer tightly packedgenes, themtDNAs in plants are notably larger (100 to 11,300 kb)and variable in their structure (Kubo andNewton, 2008;Maréchaland Brisson, 2010; Arrieta-Montiel et al., 2009; Knoop, 2012;Sloan et al., 2012; Small, 2013; Gualberto et al., 2014). In an-giosperms, the mtDNAs contain ;60 identifiable genes en-coding tRNAs, rRNAs, ribosomal proteins, and various subunits
of the energy transduction pathway, but they also harbor nu-merous open reading frames (ORFs), many of which are notconserved between different species and whose functions arecurrently unknown (reviewed in Mower et al., 2012). Our workfocuses on the analysis of amaturase-related (MatR) ORF,whichis encoded within the fourth intron in NADH dehydrogenase 1(nad1) (Wahleithner et al., 1990).The expression of the mitochondrial genomes in plants is
complex, particularly at the posttranscriptional level (Liere et al.,2011; Small et al., 2013; Hammani and Giegé, 2014). To becomefunctional, the primary organellar transcripts undergo extensiveprocessing events, including RNA editing (commonly C-to-Uconversions) and the splicing of numerous introns that residewithin the coding regions of many essential genes (Bonen, 2008;Brown et al., 2014; Schmitz-Linneweber et al., 2015). These es-sential processing steps are regulated by various protein co-factors, which may link the respiratory-mediated functions withenvironmental or developmental signals (Li-Pook-Than et al.,2004; Dalby and Bonen, 2013). Several of the factors required formitochondrial introns splicing have been identified, but there arecertainly many more to be discovered (Brown et al., 2014;Schmitz-Linneweber et al., 2015).With the exception of the horizontally acquired group I intron in
the cytochrome c oxidase subunit 1 gene of some angiosperms
1 These authors contributed equally to this work.2 Current address: Integrative Research Center, The Field Museum,1400 S. Lake Shore Drive, Chicago, IL 60605.3 Current address: Institute of Plant Sciences, The Gilat Research Center,Agricultural Research Organization, Rural Delivery Negev, Israel.4 Address correspondence to [email protected] author responsible for the distribution of materials integral to thefindings of this article in accordance with the policy described in theInstructions for Authors (www.plantcell.org) is: Oren Ostersetzer-Biran([email protected]).www.plantcell.org/cgi/doi/10.1105/tpc.16.00398
The Plant Cell, Vol. 28: 2805–2829, November 2016, www.plantcell.org ã 2016 American Society of Plant Biologists. All rights reserved.

(Vaughn et al., 1995; Sanchez-Puerta et al., 2008), all other mi-tochondrial introns in angiosperms are classified as group II type(Bonen, 2008). Canonical introns belonging to this class are self-catalytic RNAs (i.e., ribozymes) and mobile genetic elements, whichare defined by a highly conserved secondary structure of six stem-loop domains (DI to DVI) radiating from a central RNA core (Michelet al., 1989; Ferat and Michel, 1993). Based on their structuralfeatures and splicing chemistry (i.e., two consecutive trans-esterification steps, with a bulged A from the intron acting as theinitiating nucleophile), group II introns are proposed to be theprogenitors of spliceosomal RNAs (reviewed in Cech, 1986). Al-though some model group II introns are able to catalyze their ownexcision invitro, independentlyofproteins (Michel et al., 1989; Feratand Michel, 1993; Lambowitz and Belfort, 1993; Saldanha et al.,1993; Michel and Ferat, 1995), the conditions for self-splicing aregenerally nonphysiological (i.e., high temperatures and salt con-ditions), and for their efficient splicing in vivo, the group II-encodingpre-RNAs depend upon interactions with various proteinaceouscofactors (Lambowitz and Belfort, 2015). In bacteria and yeastmitochondria,proteins that function in thesplicingofgroup II intronstypically involve maturases (MATs), which are encoded within thefourth stem-loopdomain (DIV) of the introns themselves. TheMATswere shown to bind with high affinity and specificity to their owncognate intron-RNAs and are postulated to facilitate intron splicingby assisting the folding of these highly structured RNAs into theircatalytically active forms under physiological conditions (Cousineauet al., 1998;Wank et al., 1999; Singh et al., 2002; Noah andLambowitz,2003; Cui et al., 2004; Blocker et al., 2005; Huang et al., 2005;Ostersetzer et al., 2005).
Group II MATs contain several conserved motifs that are re-quired for both splicing and intron mobility (Mohr et al., 1993;Michel and Ferat, 1995; Wank et al., 1999; Matsuura et al., 2001;Aizawa et al., 2003; Cui et al., 2004; Lambowitz and Zimmerly,2011; Lambowitz andBelfort, 2015; Zimmerly andSemper, 2015).These include a region with sequence similarity to retroviral-typereverse transcriptases (i.e., the reverse transcriptase [RT]domain),with conserved sequence blocks that are present in the fingersand palm regions of retroviral RTs, and a conserved sequencemotif similar to the thumbdomainof retroviralRTs (alsodenotedasdomain X), which is associated with RNA binding and splicing(Mohr et al., 1993). In addition to the RT domain, these proteinsmay harbor C-terminal DNA binding (D) and endonuclease (En)domains that are found in some, but not all, of the MATs.
Phylogenetic studies show that the organellar introns in plantshave all evolved from MAT-containing introns (Bonen and Vogel,2001; Toor et al., 2001; Zimmerly et al., 2001; Ahlert et al., 2006;Lambowitz and Belfort, 2015; Zimmerly and Semper, 2015). Yet,only a single mat-related gene has been retained in each of theorganelles in angiosperms, namely,matK found in the trnK intronin the plastids (Mohr et al., 1993; Hausner et al., 2006) and a mi-tochondrial maturase-related ORF (matR), encoded within thefourth intron of the nad1 gene (i.e., nad1 i4), encoding subunit 1 ofrespiratory complex I (Wahleithner et al., 1990). Both MatK andMatR are well conserved between different monocot and dicotspecies and are thus expected to retain similar functions in allangiosperms.
Several lines of evidence suggest that MatK is involved in thesplicing of plastidial group II introns in plants (Zoschke et al., 2009;
Schmitz-Linneweber et al., 2015). Plantswith defective plastidialribosomes are unable to splice the subclass group IIA introns inthe chloroplast (Jenkins et al., 1997). Also, several parasiticplants that lack matK have also lost the plastid-encoded groupIIA introns from their plastid DNAs (Funk et al., 2007;Wicke et al.,2013), while plants that containmatK as a stand-aloneORF haveretained some subgroup IIA introns in their reduced plastidgenomes (Wolfe et al., 1992; Ems et al., 1995). Recently, usingtransplastomic lines in tobacco (Nicotiana tabacum), Zoschkeet al. (2010) showed that MatK is associated in vivo with many ofthe subgroup IIA introns in the chloroplasts. These data stronglysupport the notion that MatK plays a prominent role in plastidRNA metabolism.Likewise, the roles of MatR are also expected to be essential
for mitochondrial biogenesis in plants. MatR is closely related tomaturases encoded by group II introns in bacteria (Wahleithneret al., 1990) and has been retained as a conserved ORF in themtDNAs in nearly all angiosperms (Adams et al., 2002). Differ-ent RNA analyses show that the matR transcripts undergo sev-eral RNA-editing events, which restore conserved amino acids(Thomson et al., 1994; Bégu et al., 1998). Yet, as a transformationsystem has not been established for mitochondria in plants, noloss- or gain-of-function studies are currently available to test thistheory. In this report, we use Arabidopsis thaliana (Col-0) andcauliflower (Brassica oleracea var botrytis), two keymembers of theBrassicaceae family, to study the functions ofMatR in plants.WhileArabidopsis serves as a prime model system for plant moleculargenetics, cauliflower is employed for biochemical analysis of plantmitochondrial RNA metabolism (Neuwirt et al., 2005). We presentseveral lines of evidence indicating that the matR locus encodesamitochondrial protein,which is associatedwith various pre-RNAsin Brassicales. We further implemented an innovative system forgenetic manipulation of mitochondrial gene expression in Arabi-dopsis using synthetic ribozymes (Val et al., 2011) and show thata reduction inMatR expression affects the processing of a numberof group II introns in Arabidopsis mitochondria.
RESULTS
matR Gene Structure in Brassicales
In Brassicaceae, as in many plant species, the nad1 gene isfragmented into five exons in mtDNA (Grewe et al., 2014). Theseare typically expressed as three individual transcription units,which are fragmented in two of the introns (i.e., nad1 i1 and nad1i3). For its maturation, nad1 undergoes the splicing of two trans-splicedpre-RNAs (i.e.,nad1 i1 andnad1 i3) and theexcisionof twointrons (nad1 i2 and nad1 i4) that are found in a classical cisconfiguration. The fourth intron in nad1 (i.e., nad1 i4) encodesa well conserved maturase-related protein, denoted as MatR(Wahleithner et al., 1990;ChapdelaineandBonen,1991;Thomsonet al., 1994). Figure 1A shows a schematic secondary structuremodel of nad1 i4 and its cognate maturase-related (matR) gene inArabidopsis. Notably, sequencing data indicate that while nad1 i4is found ina standardcisconfiguration inBrassicales (Greweet al.,2014), in some plant species, such as petunia (Petunia hybrida),tobacco,wheat (Triticumaestivum), and rice (Oryza sativa),nad1 i4
2806 The Plant Cell

is fragmented in DIV either upstream or downstream of the matRreading frame (Figure 1B; Malek et al., 1997; Bonen, 2008).
The nature of the complex biochemical interactions betweengroup II introns and their protein partners is under investigation.The best characterizedmaturase to date is LtrAmaturase from the
bacterium Lactococcus lactis, which binds with high affinity andspecificity to its host intron, the Ll.LtrB pre-RNA. The LtrB group IIintron is able to self-splice in the presence of high Mg2+ con-centrations in vitro, but its splicing at physiological conditionsin vivo requires the LtrA protein (Saldanha et al., 1999). The
Figure 1. Schematic Representation of nad1 Intron 4 Predicted Secondary Structure and Its Maturase-Related (matR) Protein.
(A) Secondary structure model of Arabidopsis nad1 intron 4. The conserved domains in the fourth intron of Arabidopsis nad1 (nad1 i4) are illustrated inasecondarystructuremodel. EachsubdomainofDI andDII,DIII,DIV,DV, andDVI isoutlinedwithin thestructure.Theconservedbulged-A residue inDVI, theexon-intronbindingsites (i.e.,EBS1/IBS1andEBS2/IBS2), and tertiary interactionsbetweendifferent intron regions (indicatedby roman letters) areshown inthemodel structure. The ORF encoding the MatR protein in nad1 i4 is encoded by domain IV of the intron. The putative secondary structure of nad1 i4 wasgeneratedby thepredictionalgorithmsAlifold-ViennaRNAWebserver (Hofacker et al., 2002) and theMfoldWebserver (Zuker, 2003), basedonknownnad1i4 sequences from different angiosperms available on the NCBI Organelle Genome Resources server.(B) The gene structures of nad1 intron 4 loci in different plant species. Schematic representation of the nad1 intron 4 andmatR gene locus, including nad1exons 4 and 5, nad1 intron 4 (nad1 i4), and thematR reading frame, in Arabidopsis, cauliflower, tobacco, and rice. While the nad1 i4 sequence is found ina classical cis configuration in Brassicales species, the intron is found in a trans configuration in the mtDNAs of some other plant species, where thebreakpoint occurs in the intron’s DIV region, either upstream (rice) or downstream (tobacco) of the matR ORF.
MatR Role in Mitochondria Group II Intron Splicing 2807

association of LtrA with the LtrB intron is facilitated primarilythrough its association with regions found within DIV of the intron(Wank et al., 1999). The binding of LtrA with regions in LtrB intronDIV may enable its association with core intron elements, pro-moting short- and long-range tertiary RNA base interactions thatare necessary forRNA folding andcatalysis (Matsuura et al., 2001;Noah and Lambowitz, 2003).
BasedonthePHYRE (KelleyandSternberg,2009)andROBETTA(Kim et al., 2004) servers, the MatR protein in Brassicales sharessimilarity with the RT domains of various proteins, including telo-merases (c3du6A; confidence 99.6), HIV-1 reverse transcriptases(c1rthA; confidence 97.3), and DNA/RNA polymerases (d1ztwa1;confidence96.2).TheRTdomainsofmodelgroupII-encodedMATscontain several subdomains, which are required for RNA recog-nition and intron splicing (Lambowitz and Belfort, 2015; Zimmerlyand Semper, 2015; Piccirilli and Staley, 2016; Qu et al., 2016; ZhaoandPyle, 2016). These include the fingers and palm subdomains ofa polymeraseand the lessconserveddomainX that is analogous toa polymerase thumb domain. In addition, model maturases alsocontain in the RT region an N-terminal subdomain (NTD), which inthecaseof thebacterial LtrAprotein is required for thebindingof thematurase with regions in DIV of its host ltrB intron RNA that includethe LtrA-ORF’s Shine-Dalgarno site (Qu et al., 2016). This suggeststhat canonicalMATsmay autoregulate their own expression in vivothroughbinding totheirownribosomebindingsites (Matsuuraetal.,2001; Singh et al., 2002).
Analysis of conserved domains, implemented in the NCBIserver (Marchler-Bauer et al., 2015), showed that the MatR-ORFin Arabidopsis harbors only some parts of the consensus RTregion, i.e., sequence blocks RT 3 to 7 of the fingers-palm and anintact thumb (X) motif, but does not contain an N terminus NTDsubdomain (Figure 2). Like the nucleus-encoded type I ma-turases (Mohr and Lambowitz, 2003; Keren et al., 2009), MatRalso lacks the C-terminal DNA binding and endonuclease (D/En)domains. It is therefore anticipated that the intron mobilityfunctions associated with some canonical group II intron-en-coded MATs has been lost, while splicing activity is retained formitochondrial MatR in plants (Brown et al., 2014; Schmitz-Linneweber et al., 2015). Interestingly, MatR harbors two largeinsertions in the fingers-palm region, between the conservedsequence blocks RT-4 and RT-5 (amino acids 153 to 317) andbetween the RT-7 and the thumb motif (amino acids 385 to 533),which may correspond to additional RT subdomains, which areunique to the MatR protein (Figure 2, labeled by dotted boxes;Supplemental Figure 1).
RNA-Seq Data Indicate That the matR Gene Is Transcribedas Part of a Polycistronic Unit in Brassicales mtDNAs
To analyze the expression of MatR in Brassicaceae, we useddifferent RNA methodologies, i.e., RNA-seq (high-throughputRNA sequencing), RACE (rapid amplification of cDNA ends), andRT-PCR analyses, with purified mitochondrial RNA (mt-RNA)preparations. For this purpose, we used mitochondria isolatedfrom cauliflower inflorescences, which allows the purification oflarge quantities of highly enriched organellar preparations froma plant that is also closely related to Arabidopsis (Neuwirt et al.,2005; Keren et al., 2009). Previously, we established the genome
sequence of B. oleracea mitochondria (Grewe et al., 2014).Comparisons of the mtDNA sequences of Arabidopsis andcauliflowershowthatnad1 i4, and its cognatematR reading frame,are nearly identical between the two plant species (SupplementalFigure 1).RNA-seq of total cauliflower mt-RNA suggested that matR is
foundona largepolycistronic transcript unit containing thenad1 i4sequence together with nad1 exons 4 and 5, a (pseudo) genefragmentofnad5 (i.e.,cnad5-2), andpossibly alsoseveralORFsofunknown functions (i.e.,orf159,orf161, andorf287) (SupplementalFigure2).RNA-seqalso indicated thepresenceofUbasesatmanypositions in the RNA sequence where the DNA sequence has Cbases (Supplemental Figure 2, labeled in green). These likelycorrespond to RNA editing sites, which seem to be particularlyprevalent in the coding regions of nad1 exons 4 and 5, but alsowithin the sequence corresponding to the cnad5-2 gene. Theputative RNA editing sites in the nad1 pre-RNA are also shown inSupplemental Figure 3.We further performed RACE to characterize the putative po-
sition of the 59 and 39 ends of the nad1 i4 pre-RNA transcripts.Using circularized mt-RNAs as templates for the RACE analyses(Eyal et al., 1999; Forner et al., 2007), the sequencing data of theRACE products identified only mitochondrial transcripts thatharbored the matR gene together with the upstream regions ofnad1 i4, the fourth exon in nad1 (i.e., nad1 ex4), and a stretch of120 additional nucleotides (i.e., the longest RACE sequenceidentified) upstream to nad1 ex4 belonging to the 39-end of thetrans-spliced nad1 i3 (Supplemental Figure 3). No transcriptscontaining only the matR-ORF could be identified by the RACEanalyses. The 39-end of the transcript included;240 nucleotidesdownstream of nad1 ex5.Comparisons of the deduced MatR protein sequences from
various plants indicate a high flexibility of the N-terminal domains(Supplemental Figure 1). The upstream region of the canonicalstart site of theMatRsequencescontainsputativecodons that arealso recognizedasapartof theconservedRT-2sequenceblockofgroup II intron-encoded MATs (Figure 2; Supplemental Figure 1).Interestingly, different editing sites identified by the RNA-seq andRACE data also included a C-to-U substitution 28 nucleotidesupstreamof thefirst commonAUG, found inside thisputativeRT-2motif of MatR-ORF (Supplemental Figure 3). It will therefore beinteresting to investigate whether the translation of MatR mayinitiate upstream to the first common AUG site. Although trans-lation from non-AUG codons has not yet been proven for mito-chondria in plants, there is some indirect evidence indicating thatGTG and AUA can initiate translation for several mitochondrialgenes in different plant species (Bock et al., 1994; Siculella et al.,1996; Zhu et al., 2014).The maturation of pre-RNAs in plant organelles involves a
complex series of posttranscriptional maturation events, whichinclude 59- and 39-end processing and numerous nucleationevents that lead to the release of the mRNAs from their corre-sponding polycistronic transcripts (Germain et al., 2013; Smallet al., 2013;Brownet al., 2014;Hammani andGiegé, 2014).Whilethe putative 59 and 39 terminal regions of nad1 i4 are likelygenerated posttranscriptionally, no RNAs corresponding to theMatR coding region alone, nor to any alternativematR transcriptsother than the nad1 ex4-i4-ex5 pre-RNA, were supported by the
2808 The Plant Cell

RNA-seq and RACE data. These observations may imply that,similar to the bacterial maturases (reviewed in Lambowitz andZimmerly, 2011; Zimmerly and Semper, 2015), MatR translationis facilitated by mitoribosomes that are directly associated withthe pre-RNA of nad1 i4. Yet, the data are not sufficient to assesswhether translation is initiated by non-AUG start codons.Studies are underway to test this intriguing possibility.
matR Encodes an ∼70-kD Protein That Is Associated withMitochondrial Membranes
We anticipate that the matR locus encodes a functional protein,but this notion was not yet tested experimentally. MatR was notfound among the different proteins identified in global massspectrometry (liquid chromatography-tandem mass spectrome-try [LC-MS/MS]) analysesofplant organellar preparations (Jacobyet al., 2010; Taylor et al., 2011; Lee et al., 2012; Huang et al., 2013;Nelson et al., 2013, 2014; Shingaki-Wells et al., 2014). However,this may be expected due to its predicted low expression level inthe mitochondria, as was also evident in the cases of MatK(Zoschke et al., 2010) and thenucleus-encoded (nMAT) homologs(Keren et al., 2009, 2012; Cohen et al., 2014), as well as variousother RNA-processing enzymes in the mitochondria in angio-sperms (Law et al., 2012).
As no antibodies to MatR protein were available prior to thisstudy, a polyclonal antibody was raised against a syntheticpeptide corresponding to a unique region in the RT domain, whichis conserved among different Brassicales (i.e., NH3
+-RRIDD-QENPGEEASFNA) (Supplemental Figure 1). The affinity andspecificity of the antibody were confirmed in protein gel blots usingtotal protein and enriched organelle preparations (i.e., mito-chondria, chloroplasts, and peroxisomes) obtained from 3-week-oldArabidopsisvarColumbia (Figure3). The immunoblotswith theanti-MatR antibodies showed a single band corresponding to
a protein of ;70 kD, which is also the expected size for a nativeMatR protein (calculated mass 73.65 kD, assuming that MatRtranslation initiates at the first common AUG site; SupplementalFigure 1); this was observed only in the mitochondria subfraction(Figure 3A). The use of antibodies to the mitochondrial proteinsserine-hydroxymethyltransferase (SHMT) and cytochrome oxi-dase subunit 2 (COX2), the plastid-encoded 33-kD subunit of theoxygen evolving complex of PSII (PsbO), and the peroxisomalmarker catalase protein confirmed that the organelle preparationswere highly enriched (Figure 3A).We further investigated whether MatR is found in the soluble
(matrix) or membranous fractions in the mitochondria. For thispurpose, intact mitochondria (Figure 3B) were disrupted byseveral freeze and thaw cycles followed by sonication and thenseparated by centrifugation into the pellet, which contains themembranes and the supernatant containing the matrix compo-nents. The fractionation procedure was validated by protein gelblot analyses with antibodies against SHMT (as an indicator ofmatrix proteins), while COX2 was used as a marker of organellar(inner) membrane proteins (Figure 3B). Unexpectedly, the im-munoblot assays show that the MatR protein is enriched in themitochondrial membranous fraction (Figure 3B), even though thededuced protein sequence of MatR lacks any obvious membranetargeting motifs.To investigate the topology of MatR in the mitochondria, the
membrane fraction was prewashed with 10mMTris-HCl, pH 8.0(to remove contaminating proteins), and then treated with 1 MNaCl (removes loosely attached membrane proteins), 2 M NaBr(a chaotropic agent used for the removal of some peripherallybound proteins), or 0.05% (v/v) Triton X-100 + 1MNaCl (this lowconcentration of the detergent will not completely solubilize themembranes but can wash peripheral proteins attached by sur-face hydrophobic interactions) (Ostersetzer et al., 2007). Thedata indicate thatMatR is likely tobeperipherally associatedwith
Figure 2. Schematic Representation of Plant Mitochondrial Maturase-Related Proteins.
Schematic representation of maturase proteins found in bacteria and different plant species. Shaded boxes represent different domains associated withmodel group II intron-encodedmaturases. These include the reverse transcriptase (RNA-directed DNA polymerase) domain (pfam-00078), with its intrinsicNTD, thefingers-palm (RT I toVIImotifs), and theRNAbindingandsplicing (i.e., domainXor thumb; pfam-08388) subdomains. Somemembersof this familyharbor an additional C-terminal DNA endonuclease (D/En) domain (pfam-01348).
MatR Role in Mitochondria Group II Intron Splicing 2809

the membranes, as neither the 10 mM Tris-HCl nor the 1 M NaCltreatment released the protein from the membranes, whereastreating theorganellarmembraneswith 2MNaBr or 0.05%TritonX-100 + 1MNaCl removed theMatR signal from themembranes
(Figure 3C). Similarly to MatR, the peripheral AtpB subunit of themitochondrial ATP-synthase coupling factor-1 (CF1) was re-moved by either 2MNaBr or 0.05%Triton X-100 + 1MNaCl, butnot by the 10 mM Tris-HCl or 1 M NaCl washes. In contrast toMatR and AtpB, the 2MNaBr wash did not remove the signals ofthe Nad9 subunit of the soluble arm of complex I or the COX2protein of complex IV, which is integrally associated with themitochondrial inner membranes. While the Nad9 subunit waslargely removed from the membranes by 0.05% Triton X-100 +1MNaCl, the integral COX2proteinwas only partially affectedbythe detergent treatment (Figure 3C). Taken together, these re-sults indicate that the MatR protein associates peripherally withmembranes. As MatR is synthesized inside the organelles, wespeculate that the protein is associated with the organellar innermembranes.
MatR Expression Profiles in Arabidopsis
The relativeaccumulationof transcriptscorresponding tomatRandnad1 in Arabidopsis was analyzed by RT-qPCR in different planttissuesandduringdifferentdevelopmentstages(Figure4).Thedataindicate that the steady state levels of transcripts corresponding tomatR/nad1-i4 and to the mature nad1 exons 4-5 were all belowdetectable levels in dry (desiccated) seeds. Notably, accumulationof transcripts corresponding to matR/nad1 i4 pre-mRNAs wasevident as early as 2 h after initial seed imbibition (Figure 4). Thesteadystate levelsofmatRpre-RNAtranscriptsgradually increasedduringthe imbibitionstage, reachingthehighest levelbetween12to48hafter initialwateruptakebythedryseed.BasedontheRNA-seqand RACE data, it is anticipated that matR is cotranscribed withnad1 i4. The subtle differences in the accumulation of transcriptscorresponding to matR and nad1 i4 pre-RNA may be due to thedifferences caused by the RT-qPCR analyses, i.e., the use of dif-ferent sets of oligonucleotides or the differences in the sizes of thePCR products (153 and 64 nucleotides, respectively) (Figure 4).Following seed initiation, where the radicle protrudes through theseed coat (i.e., ;48 h), the levels of matR transcripts steadily de-creased throughout the plant’s development. In contrast to thematR expression profiles, accumulation of mature nad1 RNAtranscriptscorresponding toex4-5wasapparentonlyafter;24hofseed imbibition (Figure 4). Following this step, the steady statelevels of nad1 ex4-5 mRNA gradually increased, peaking around10 d after seed initiation (Figure 4).To analyze the accumulation of MatR and various other mito-
chondrial proteinsduringearly seedgermination, total proteinwasextracted fromdryand imbibedArabidopsis seeds following1and2 d of imbibition, and the accumulation of mitochondrial proteinswas examined by SDS-PAGE and protein gel blotting analyses(Figure 5). The immunoblots indicated that the ;70-kD band,corresponding to the MatR protein, was hardly detectable in thedry seeds,whereas it was clearly visible in the imbibed seeds after24 h and further increased in its levels after 48 h of imbibition.However, theMatRprotein signalwasnoticeably reduced (;2.53lower) in mature Arabidopsis seedlings (i.e., at 1.03 dilution)compared with the levels of MatR in germinating seeds 2 d fol-lowing the initial imbibitionstage (Figure5A).The levelsof theoutermitochondrial membrane protein porin-1 (VDAC1) were alsohigher (1.3 to ;1.83) in seeds than in mature (i.e., 3-week-old)
Figure 3. The matR Locus in Arabidopsis Encodes an ;70-kD ProteinLocalized to Mitochondrial Membranes.
(A) Apolyclonal antibodywas raised against a synthetic peptide correspondingto a unique region in the RT domain of MatR, which is conserved amongdifferent Brassicales (i.e., NH3
+-RRIDDQENPGEEASFNA; SupplementalFigure 1). The affinity and specificity of the antibody were confirmed inprotein gel blots with total protein and enriched organelle preparations (i.e.,mitochondria, chloroplasts, and peroxisomes) obtained from 3-week-oldArabidopsis plants. COX2 (a mitochondrial protein), PsbO (a plastidialprotein), and Cat (a peroxisomal marker) were used as controls.(B)Mitochondria isolated fromArabidopsiswere subjected to several freezeand thaw cycles, followed by a brief sonication, and centrifuged to separatethe soluble matrix fraction and the membrane fraction. Intact mitochondria(Mit), soluble fractions (S), and membrane fractions (M) were subjectedto immunoblot analysis using anti-MatR antibody. Mitochondrial SHMT(a matrix protein) and COX2 (a membranous protein) were used as controls.(C) Todeterminewhethermembranousproteins areperipherally or integrallybound to themembranes, themembrane fractionwas treatedwith 1MNaCl(removes looselyattachedmembraneproteins),2MNaBr(achaotropicagentused for the removal of peripherally bound proteins), or 0.05% Triton X-100,and 1 M NaCl (wash some peripheral proteins attached by surface hydro-phobic interactions). The location of each protein (i.e., peripherally or in-tegrally associated) was determined by centrifugation following immunoblotanalysis of the washed membranes pellet and supernatant fractions.
2810 The Plant Cell

seedlings. In Arabidopsis, VDAC1 has a ubiquitous expressionpattern but shows particularly high expression levels in re-productive organs and during germination and early seedlingestablishment (Pan et al., 2014).
The levels of several other mitochondrial proteins, includingNad9 and the nucleus-encoded CA2 (carbonic anhydrase likesubunit 2) of complex I (CI), theRieske Iron-Sulfur protein (RISP) ofCIII, the COX2 subunit of CIV, and the AtpB subunit of the ATPsynthase enzyme (CV) also increased during the imbibition stage(Figure 5A). Yet, unlike the patterns of MatR and VDAC1 accu-mulation, the steady state levels of these proteins were not sig-nificantly altered during the first 24 h of seed imbibition andincreased only 48 h following initial seed imbibition. Unlike MatRand VDAC1, the highest steady state levels of the respiratorysubunits Nad9, CA2, RISP, COX2, and AtpB were observed inmature plants rather than seeds (Figure 5A). Blue-native (BN)-PAGE gel electrophoresis, followed by protein gel blot analyses,corroborated the immunoblot results. The mitochondrial holo-complexes corresponding to CI, CIII, CIV, and CV had the lowestabundances in dry seeds, but then gradually increased in theirlevels during the 48 h of imbibition, showing the highest abun-dances in mature (i.e., 3-week-old) seedlings (Figure 5B). Thenumbers below the blots in Figure 5 show the relative accumu-lation (quantifiedusing ImageJsoftware; Jensen,2013)ofdifferentorganellar proteins during the imbibition stage and in mature,3-week-old Arabidopsis seedlings.
MatR Is Associated with Various Pre-RNAs inBrassicales Mitochondria
It was previously noted that group II intron-containing pre-RNAsare associated with their corresponding splicing cofactors inbacteria and chloroplasts in plants (Stern et al., 2010; Barkan,2011; Lambowitz and Zimmerly, 2011; Germain et al., 2013).Likewise, the mitochondrial maturase-related nMAT2 protein isbound to several group II RNAs, which were also identified asthe native intron targets of the protein in vivo (Keren et al., 2009).These data suggest that splicing cofactors of group II introns areassociated specifically with those introns whose splicing theyfacilitate in bacteria and in plant organelles. Accordingly, coim-munoprecipitation (co-IP) analyseswith antibodies raised againstdifferent organellar splicing factors indicated that the plant factorstypically co-precipitated with their genetically defined intron tar-gets (Ostheimer et al., 2003; Schmitz-Linneweber et al., 2005,2006; Asakura andBarkan, 2007;Watkins et al., 2007;Kerenet al.,2009; Zoschkeet al., 2010).Here,we further applied this approachto define and catalog the native RNA targets of MatR protein. Forthis purpose, anti-MatR antibodies were used in pull-down ex-periments to pellet the associated MatR-RNP (ribonucleoprotein)complexes from solubilized (1% Nonidet P-40, v/v) mitochondriapreparations obtained from cauliflower inflorescences. cDNAsprepared from the RNA recovered by the co-IP were applied tocustom arrays and analyzed by hybridization to specific intron
Figure 4. Analysis of matR and nad1 Transcript Profiles during Germination, Initiation, and Early Developmental Stages.
The expression and accumulation of transcripts corresponding to nad1 ex4-5 mRNA, matR, and nad1 i4 in Arabidopsis mitochondria during imbibition,initiation (i.e., when the radicle protruded through the seed coat, ;48 h), and at later developmental stages were examined by RT-qPCR analysis withspecifically designed primers. The qPCR data were normalized against the nuclear gene 18S rRNA (At3g41768) (the value 1.0 represents the maximumexpression level during the time course). The values are means of three biological replicates (error bars indicate one SD). Oligonucleotides used for theanalysis of nad1 expression during imbibition include AATTTTTGCGAGAGCTGGAA and TTGAACCCCGTCCTGTAGAC for matR; AGCCCGGGATC-TTCTTGA and ACGGAGCTGCATCCCTACT for nad1 i4; AGCCCGGGATCTTCTTGA and TCTTCAATGGGGTCTGCTC for nad1 exons 4 and 5; andAAACGGCTACCACATCCAAG and ACTCGAAAGAGCCCGGTATT for 18S rRNA.
MatR Role in Mitochondria Group II Intron Splicing 2811
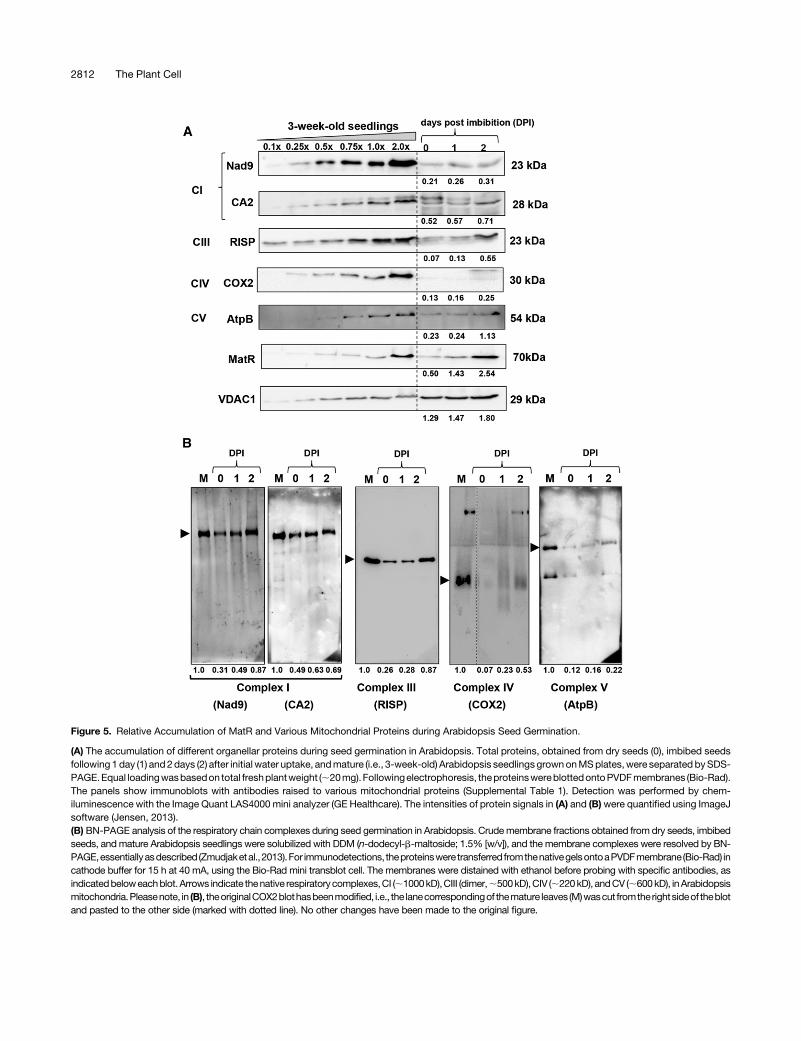
Figure 5. Relative Accumulation of MatR and Various Mitochondrial Proteins during Arabidopsis Seed Germination.
(A) The accumulation of different organellar proteins during seed germination in Arabidopsis. Total proteins, obtained from dry seeds (0), imbibed seedsfollowing1 day (1) and2days (2) after initial water uptake, andmature (i.e., 3-week-old) Arabidopsis seedlingsgrownonMSplates,were separated bySDS-PAGE.Equal loadingwasbasedon total freshplantweight (;20mg).Followingelectrophoresis, theproteinswereblottedontoPVDFmembranes (Bio-Rad).The panels show immunoblots with antibodies raised to various mitochondrial proteins (Supplemental Table 1). Detection was performed by chem-iluminescence with the Image Quant LAS4000 mini analyzer (GE Healthcare). The intensities of protein signals in (A) and (B)were quantified using ImageJsoftware (Jensen, 2013).(B) BN-PAGE analysis of the respiratory chain complexes during seed germination in Arabidopsis. Crudemembrane fractions obtained from dry seeds, imbibedseeds, andmature Arabidopsis seedlings were solubilized with DDM (n-dodecyl-b-maltoside; 1.5% [w/v]), and the membrane complexes were resolved by BN-PAGE,essentiallyasdescribed (Zmudjaketal.,2013).For immunodetections, theproteinswere transferred fromthenativegelsontoaPVDFmembrane(Bio-Rad) incathode buffer for 15 h at 40 mA, using the Bio-Rad mini transblot cell. The membranes were distained with ethanol before probing with specific antibodies, asindicatedbeloweachblot. Arrows indicate thenative respiratorycomplexes,CI (;1000kD),CIII (dimer,;500kD),CIV (;220kD), andCV (;600kD), inArabidopsismitochondria.Pleasenote, in (B), theoriginalCOX2blothasbeenmodified, i.e., the lanecorrespondingof themature leaves (M)wascut fromtherightsideof theblotand pasted to the other side (marked with dotted line). No other changes have been made to the original figure.
2812 The Plant Cell

probes (see Methods; Supplemental Figure 4). Commercial an-tibodies against the mitochondrial SHMT protein (SupplementalTable 1) were used as a control for the RNA coimmunoprecipitation/microarray (RIP)-chip analysis. RNAs identified by the MatR RIP-chip assay included nad1 i4, the host intron of matR, as well asseveral other organellar transcripts, includingnad1 i1,nad1 i3,nad4i1, nad5 i4, nad7 i2, rpl2 i1, and rps3 i1 (Figure 6; SupplementalFigure 4). In addition to these RNAs, weaker signals were seen inthe cases of nad2 i2, i3, and i4 and nad4 i2 (Supplemental Figure 4).The signals of abundant organellar transcripts, such as tRNAs,rRNAs, and many of the organellar mRNAs, were all below de-tectable levels, strongly supporting the specificity of the RIP-chipanalyses.
The association of MatR with pre-RNAs corresponding to nad1,nad4, nad5, rpl2, and rps3 was also supported by sequencinganalyses of cDNA libraries prepared from the pelleted RNAsobtained from the co-IPs with MatR antibodies (SupplementalFile 1). Therefore,weassume that thenad1,nad5,nad7, rpl2, andrps3 pre-RNAs represent the native intron targets of the MatRprotein in Brassicales mitochondria. Unlike MatR, no RNAs wereidentified in the SHMT-related RIP-chip analysis (SupplementalFigure 4).
In addition to identifying and characterizingmt-RNAs that areassociated with the MatR protein, we also characterized dif-ferent proteins that are found together with MatR in mito-chondrial ribonucleoprotein particle(s) in vivo (i.e., proteins thathave been coprecipitated with MatR in the RIP-chip assays).LC-MS/MS analyses of MatR indicated the presence of sev-eral mitochondrial proteins that coprecipitated with MatR(Supplemental Table 2). Several proteins, such as heat shockproteins, prohibitins, andATP-synthase subunits, are abundantin the mitochondria and may therefore be nonspecificallyprecipitated by the MatR antibodies (some were also identifiedby the co-IP with SHMT antibodies). Yet, in addition to MatR,other proteins identified by the co-IP may correspond to splicingcofactors that are associated together with MatR in spliceosomal-like RNP particles in the mitochondria (Schmitz-Linneweberet al., 2015). Such proteins may include the splicing cofactorDEAD-box ATP-dependent RNA helicase 53 (PMH2) (Köhleret al., 2010) and a few other RNA binding cofactors belongingto the large family of the pentatricopeptide repeat-containingproteins inplants (Small andPeeters, 2000) (Supplemental Table 2).Importantly, these data indicate that polypeptides corre-sponding to the MatR protein are indeed present in angiospermmitochondria.
Downregulation of MatR Expression by a PioneeringRibozyme-Based Strategy in Arabidopsis Mitochondria
The RIP-chip data indicate that MatR is associated with manymitochondrial pre-RNAs (Figure 6; Supplemental Figure 4).However, these data are not sufficient to indicate a direct role forMatR in the processing of group II introns in angiosperms mi-tochondria. As no methods are currently available to modifymtDNA in plants, we sought to utilize a modified hammerheadribozyme system, which was recently shown to cleave andthereby strongly downregulate the accumulation of mitochon-drial atp9 transcripts in vivo (Val et al., 2011). To investigate the
roles of MatR in the processing of group II intron-containingtranscripts in plant mitochondria, synthetically engineeredtrans-cleaving hammerhead ribozymes (matRz1 and matRz2),designed to cleave inside the matR reading frame, were driveninto the organelles as “passenger” molecules conjugated toa tRNAVal-like shuttle RNA vector (Figure 7; Val et al., 2011).Because the functions of MatRmay be essential to the plant, thechimeric constructs, comprising the matR-ribozyme compo-nents fused to the tRNA-like shuttle vector, were cloned under anestradiol-inducible promoter (Zuo et al., 2000). To assess pos-sible side effects caused by the expression of mitochondria-localized ribozymes, we further designed a ribozyme against thetobaccomitochondrial sdh3 gene (Figure 7), which is not presentin the mitochondria of Brassicaceae, including Arabidopsis.The transgenic lines matRz1 and matRz2 represent individualtransformants expressing different synthetic ribozymes thatwere designed to affect the expression of the ArabidopsismatRgene in two separate gene loci (Figure 7).Upon induction of the matRz ribozymes, the levels of matR
transcripts were reduced by 50 to 60% in the matRz1 and thematRz2 lines, as determined by RT-qPCR analyses (Figure 8;Supplemental Figure 5). The steady state levels of the matRtranscripts remained low for at least 5 d following estrogen in-duction of the transgenes. Accordingly, the steady state levels ofMatR protein also decreased and was reduced by ;50% inmatRz1 and by ;20% in matRz2 5 d after the induction of thematRz ribozymes (Figure 8; Supplemental Figure 5). No signifi-cant changes inMatR accumulation and expressionwere seen inthe control sdhRz lines (Figure 8). Ribozyme-mediated knock-down ofmatR had no significant effect on the steady state levelsof various intronless mitochondrial RNAs (Supplemental Figure5). Under normal growth conditions (seeMethods), expression ofthe ribozymes generated no obvious phenotypic differencesbetween the transgenic lines andwild-type plants (i.e., within thetime frame considered) (Supplemental Figure 6). This may beexpected due to only partial reductions inMatR expression in thetwo matRz lines (Figure 8; Supplemental Figure 5), while thecomplete loss of matR gene function is expected to be lethal tothe plant.The accumulation of each of the 23 group II introns in Arabi-
dopsis mitochondria (Unseld et al., 1997), including nad1 i1-i4,nad2 i1-i4, nad4 i1-i3, nad5 i1-i4, nad7 i1-i4, cox2 i1, ccmFc i1,rpl2 i1, rps3 i1, and their flanking exons, was analyzed in thematRz and sdhRz lines at the time of induction (Figure 9A) and5 d (Figure 9B) following the addition of estrogen to the growthmedium. The expression profiling of the Arabidopsis mt-RNAsby RT-qPCR was performed generally as described previously(Francs-Small et al., 2012; Keren et al., 2012; Zmudjak et al.,2013; Cohen et al., 2014). Oligonucleotides designed forthe different exon and intron sequences are indicated inSupplemental Table 3A. Introns thatwere notably affected by thereduction in MatR expression (i.e., at day 5) included the soleintrons within rpl2, rps3, cox2, and ccmFc (all in a cis configu-ration) as well as nad1 i3 (trans-spliced), nad1 i4 (cis-spliced),nad5 i4 (cis-spliced), and nad7 i2 (cis-spliced) (Figure 9). Toa lesser extent, reducedsplicingefficiencies (i.e., higherpre-RNAto mRNA ratios) were also seen in the cases of nad2 i4 (cis-spliced), nad4 i3 (cis-spliced), andnad5 i3 (trans-spliced). In each
MatR Role in Mitochondria Group II Intron Splicing 2813

Figure 6. MatR Is Associated with Various Mitochondrial Pre-RNAs in Vivo.
Identification of RNAs associated with MatR in cauliflower mitochondria by RIP-chip analysis (see also Supplemental Figure 4). The enrichment ratios werecalculated in two independent assays, using anti-MatR and anti-SHMT antibodies, based on the signals in dot-blot hybridizations with radiolabeled cDNAsgenerated from the RNAs recovered in the respective immunoprecipitates. Coimmunoprecipitated mt-RNAs were reverse-transcribed in the presence ofradiolabeled C and U nucleotides and random primers and hybridized to DNA probes blotted onto nylon filters, essentially as described previously (Zoschkeet al., 2010). For each experiment, signalswere normalized to thenad1 i4 signal. The corresponding data canbe found inSupplemental Figure 4. TheRIP-chipdata were further validated through sequencing of the cDNAs obtained from the pelleted mt-RNAs in the co-IP (i.e., cDNA-seq; Supplemental File 1).
2814 The Plant Cell
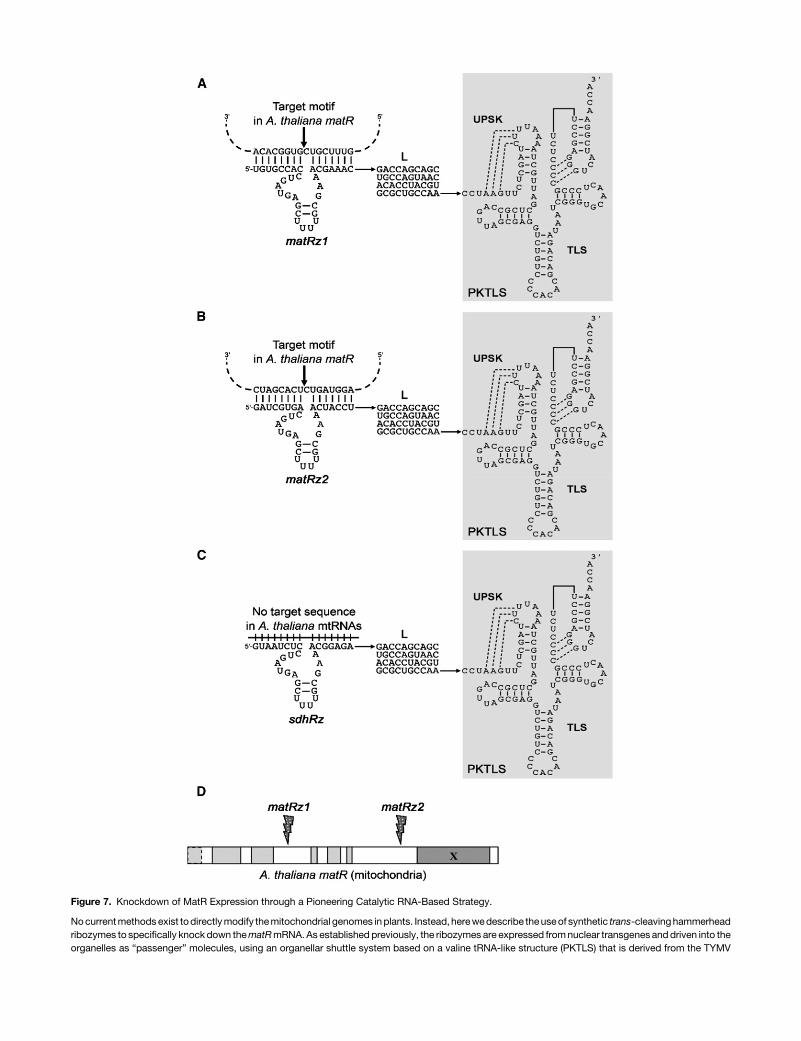
Figure 7. Knockdown of MatR Expression through a Pioneering Catalytic RNA-Based Strategy.
Nocurrentmethodsexist todirectlymodify themitochondrial genomes inplants. Instead, herewedescribe theuseof synthetic trans-cleavinghammerheadribozymes to specifically knock down thematRmRNA. As established previously, the ribozymes are expressed fromnuclear transgenes anddriven into theorganelles as “passenger” molecules, using an organellar shuttle system based on a valine tRNA-like structure (PKTLS) that is derived from the TYMV

case, the accumulation of unspliced pre-mRNAs was correlatedwith decreased transcript levels in the correspondingmRNAs, inboth matRz1 and matRz2 lines, compared with those of thecontrol sdhRz plants (Figure 9; Supplemental Figure 7). Accu-mulation of pre-mRNAs in the matR-reduced lines was alsoapparent for nad2 i4, nad4 i3, and nad5 i3 (Figure 9B). However,as thecorrespondingmRNA levels (i.e., spliced exons)were notsignificantly affected in the matRz lines (Supplemental Figure7), we could not draw any firm conclusions regarding the pu-tative roles ofMatR in the splicing of these introns. The levels ofmt-RNAs corresponding to nad2 exons 2-3, exons 3-4 (and tosome extent nad2 exons 4-5, aswell), and nad5 exons 1-2weresomewhat higher in the matRz lines, while no significant dif-ferences in the transcript levels of nad1 i1 and i2, nad2 i1, nad4i1, nad7 i1, i3, and i4, or their corresponding mRNAs (i.e.,spliced exons), were seen between the sdhRz andmatRz lines(Figure 9; Supplemental Figure 7). Thus, we conclude that thesplicing of this entire subset of mitochondrial RNAs does notrequire MatR.
DISCUSSION
Angiosperm Genomes Encode Several MAT-RelatedProteins, Which Function in the Splicing of Different Subsetsof Organellar Group II Introns
Sequences similar to retroviral-type RTs are widely distributedamong different organisms. In bacteria, such genes includevarious ORFs encoded within many of the group II intronstermed as MATs (reviewed in Lambowitz and Zimmerly, 2011;Lambowitz and Belfort, 2015; Zimmerly and Semper, 2015).These were shown to act specifically in the splicing and intronmobility (i.e., retrohoming) activities of their own cognate intronRNAs.While the chemistry of the splicing reaction ismediated bythe introns themselves, mounting evidence indicates that theMATs are required to facilitate or to stabilize the folding of thehighly structured group II RNAs into their catalytically activeforms.
Plantgenomesencodeseveralproteins thatareclosely related tobacterial group II reverse transcriptase/maturaseproteins (Figure 3)(Mohr and Lambowitz, 2003; Knoop, 2012; Brown et al., 2014;Schmitz-Linneweber et al., 2015). In angiosperms, these includeMatKencodedby thegroup II intronof trnK (i.e., tRNALysUUU) in theplastids (Mohr et al., 1993), four nucleus-encoded maturases (i.e.,nMAT1 to 4) (Mohr and Lambowitz, 2003) that were all shown toreside in the mitochondria in Arabidopsis (Keren et al., 2009), andthe mitochondrial MatR-ORF found inside nad1 i4 (Wahleithneret al., 1990) that is in the focus of this work.
The sixmaturases in angiospermsareall expected to function inthe splicing of different subsets of group II introns in plantorganelles (Schmitz-Linneweber et al., 2015). Genetic and bio-chemical studies indicate a role for MatK in the processing ofabout half of the plastidial introns, all belonging to subgroup IIA(Zoschke et al., 2010). Likewise, the functions of the nucleus-encodednMAThomologs, includingnMAT1,nMAT2, andnMAT4,are required for the splicing of various group II introns in the mi-tochondria,manyofwhich residewithinRNAsencodingcomplex Isubunits (Nakagawa and Sakurai, 2006; Keren et al., 2009, 2012;Cohen et al., 2014). It is therefore anticipated that other plantmaturases, including the nucleus-encoded nMAT3 and the mi-tochondria-encoded MatR proteins, will also function in thesplicing of mitochondrial pre-RNAs in plants. Further inves-tigations of the roles of nMAT3 are underway in our laboratories.ThematR sequence has remained intact in the mtDNAs of nearlyall angiosperms and gymnosperms (Guo and Mower, 2013).Furthermore, analyses of the transcription profiles of many plantspecies indicate that matR transcripts undergo various RNA-editing events that restore conserved amino acids (SupplementalFigure 3). Here, we provide multiple lines of evidence indicatingthat the matR locus encodes a protein that functions in the pro-cessing of manymitochondrial group II intron-containing RNAs inBrassicales.
The 3D Homology Model of MatR Reveals Similarities withCanonical Bacterial Maturases
A major advance in the understanding of the functionality ofMATs in group II intron splicing has been recently accomplishedby structural analyses of bacterial MATs bound to their cognategroup II intron RNAs (Piccirilli and Staley, 2016; Qu et al., 2016;Zhao and Pyle, 2016). These including the crystal structures ofthe RT domains of MATs from the bacteria Roseburia intestinalisand Eubacterium rectale (Zhao and Pyle, 2016) and a cryo-electron microscopy analysis of the ribonucleoprotein complexof the L. lactis intron-encoded LtrA maturase bound to its hostltrB intron RNA (Qu et al., 2016). Analyses of the structures of thespliced ltrB intron (at 4.5 Å resolution) and of the ltrB intron in itsribonucleoprotein complex with LtrA (at 3.8 Å resolution) arefurther revealing functional coordination between the intronRNAwith its cognate MAT protein. Remarkably, these structuresreveal close relationships between the RT catalytic domain andtelomerases, whereas the active splicing centers resemble thatof the Prp8 protein (Dlakic and Mushegian, 2011; Galej et al.,2013; Yan et al., 2015), which also resides at the core of thespliceosome.We tested whether MatR in plants may adopt a structural fold
similar to thatofbacterialmaturases (e.g., LtrAprotein).Using the
Figure 7. (continued).
genomic RNA (Val et al., 2011). The synthetic ribozymes, denoted here asmatRz1 (A) andmatRz2 (B), are designed to specifically cleave the ArabidopsismatRRNA (ArthMp045) in two different gene loci (D). The specific recognition sequences for the cleavage sites of thematR ribozymes are presentedwithin(A) and (B). The sdhRz ribozyme construct (C), designed against themitochondrial sdh3mRNA in tobacco, was used as a control for thematR knockdownexperiments, since it has no corresponding targets in Arabidopsis that has lost the sdh3 gene from the mtDNA and thus should have no effect on mi-tochondriageneexpression inArabidopsis. For their localization to themitochondria inArabidopsis, eachof the ribozymeswasattached to the59-endof thePKTLS shuttle sequence via a 40-nucleotide linker (L) region (Val et al., 2011).
2816 The Plant Cell

RaptorX Web server (Källberg et al., 2012), we constructeda homology 3D model of the Arabidopsis MatR protein(Supplemental Figure 8) in an attempt to map functional do-mains in MatR and to address its putative roles in mitochondriagroup II intron RNA recognition and splicing. The predictedstructure of MatR includes most of the regions of the fingers-palm motif and the thumb (X) domain (Supplemental Figure 8).
As MatR shows only limited homology in the C-terminal regiontomodel maturases, this region could not be fully modeled. Wewere also unable to provide a structural prediction of the twoinsertions in MatR found inside the fingers-palm (Figure 2;Supplemental Figure 8, labeled with dotted lines) due to thelack of homology of these regions with known protein se-quences. Nonetheless, the superimposed 3D model of MatR
Figure 8. Downregulation of MatR Expression by Synthetic trans-Cleaving Hammerhead Ribozymes.
The chimeric RNAs, comprising the matRz ribozymes attached to the PKTLS shuttle through a linker sequence (Figure 7), were expressed from nucleartransgenes under an estradiol-inducible promoter (see Methods). Accumulation ofmatR transcripts in transgenic plants expressing the matRz1 or matRz2ribozymewasanalyzedbyRT-qPCRwithspecificprimersdesigned toamplify regionsofmatRspanningover the ribozymecleavagesites (A). The resultswerenormalized againstmatR transcript levels in control transgenic plants expressing the sdhRz ribozyme. The real-time qPCR data were normalized against thenucleargenesACTIN2,GAPDH, andEXPRESSED. ThecorrespondingMatRprotein levels inmatRz1-,matRz2-andsdhRz-expressingplantsweredeterminedby protein gel blot analyses with anti-MatR antibodies (B). Themeans of three experiments are shown as a graph on the left, whereas a representative blot isshown on the right; normalization of the data was performed with commercial antibodies raised against actin-8 (see Supplemental Table 1).
MatR Role in Mitochondria Group II Intron Splicing 2817

protein fits quite well with the established structures of the bac-terial LtrA maturase (Supplemental Figure 8).
We further speculate that predicted basic surfaces in MatR (i.e.,positively charged regions; Supplemental Figure 8, regions labeledin blue) may correspond to RNA binding sites required for the
association of the protein with its target pre-RNA molecules, inasimilarmanner tothebacterialmaturases(Quetal.,2016;ZhaoandPyle, 2016). However, additional structural studies andbiochemicalassays would be required to confirm the hypothetical 3D model ofMatR protein structure and to analyze its binding characteristics.
Figure 9. Knockdown of MatR Affects the Splicing of Multiple Mitochondrial Group II Introns.
The splicing efficiencies of the 23 intron-containing mitochondrial transcripts in Arabidopsis (Unseld et al., 1997) were analyzed in ribozyme-expressing transformants. Total RNAs extracted from Arabidopsis lines expressing the sdhRz, matRz1, and matRz2 ribozymes were analyzed byRT-qPCR, with primers designed to specifically amplify regions of mitochondrial introns, exons, and intron-exon or exon-exon junctions in rpl2,rps3, cox2, ccmFc, nad1, nad2, nad4, nad5, and nad7 pre-mRNAs/mRNAs (see Supplemental Table 3A). The histograms show the log2-transformedratios of transcript splicing efficiencies in thematR-reduced linesmatRz1 andmatRz2 compared with those of the sdhRz control siblings. Sampleswere taken at both day 0 (A), just before onset of thematRz1,matRz2, and sdhRz ribozymes expression and after 5 d following ribozymes induction ofexpression (B). The values are means of three biological replicates (error bars indicate one SD).
2818 The Plant Cell

The N-Terminal Region of MatR: Potential Use of AlternativeTranslation Initiation Sites for MatR Proteins inAngiosperms Mitochondria
Comparison of the deduced amino acid sequences ofMatR-ORFsfrom different angiosperms indicates a promiscuous mode oftranslational initiation. Some, including MATs in dicots, mostmonocots, and a few rosids, begin with an “AGA” triplet (for argi-nine) followedby 18putative codons upstreamof the first commoninitiation (AUG) site (i.e., RKKEGLKFRLTVVLPIEKIM), while otherMatR-ORFs in eudicots (e.g., tobacco) beginwith the “GGG” triplet(for glycine) (i.e., RKK*GSKFRPLTVVLPIEKIM), and a stopcodon isfound at the 4th codon position in the matR’s coding region(Supplemental Figure 1). Interestingly, the codons upstream of thecanonical start site are recognized as part of the conserved RT-2sequence block of group II maturases (Figure 2; SupplementalFigure 1). The translation of some nuclear genes in eukaryotes caninitiate at codons that differ from the consensus site in one base(e.g.,CUG,ACG,AUU,andGUG) (Kozak,1989,1997;Gordonetal.,1992; Hann, 1994). Prokaryotes and mitochondria in animals usealternative initiation codons more frequently (commonly U/GUG inbacteria and AUA/U in animalmitochondria) (Lobanov et al., 2010).Although the translation from non-AUG codons has not yet beenproven for mitochondria in plants, there is some indirect evidencestrongly indicating that GUG and AUA can initiate translation forseveral mitochondrial genes in different plant species (Bock et al.,1994; Siculella et al., 1996; Zhu et al., 2014).
The Expression of MatR Is Developmentally Regulated
Gene expression mechanisms that occur in plant mitochondriainclude RNA editing, splicing of many group II introns, and nu-merous nucleation events that lead to the release of mRNAs fromtheir organellar-encoded polycistronic transcripts (Germain et al.,2013; Small et al., 2013; Brown et al., 2014; Hammani and Giegé,2014). The MatR ORF in angiosperm mitochondria is found withinthe fourth intron innad1. Anuncertainty regardingMatRexpressionin angiosperms involves the transcription of the gene by an alter-native promoter site found inside nad1 i4, upstream from thematRlocus. Inwheat, thismay lead to theexpressionofa truncatedMatRproduct, which lacks most of the N-terminal RT domain (Farré andAraya, 1999). The expression of MatR in Arabidopsis and cauli-flower was studied here by different RNA and protein methodol-ogies.RNA-seqandRACEanalyses (SupplementalFigures2and3)indicated thatMatR is cotranscribed with its host nad1 i4 but failedto reveal the presence of any transcripts containing only the MatRORF, arguing against the presence of any internal transcriptionalstart sites in cauliflower. As MatR is most likely required for thematuration of its own transcript, we speculate that MatR is trans-lated directly from the unspliced precursor nad1 ex4-i4-ex5 tran-script, in a similar manner to the canonical maturases encoded bygroup II introns in bacterial genomes (Lambowitz and Zimmerly,2011; Zimmerly and Semper, 2015).
The immunoblot assays with anti-MatR antibodies identified asingle protein of;70 kD in themitochondria, which corresponds toan intact MatR protein (Figure 3A). No additional protein bandscorresponding to polypeptides of either lower or higher molecu-lar weight were seen in the protein gel blot analyses. In the
mitochondria, MatR was found to be associated mainly with themembranes (Figure 3B). Analysis ofMatR topology by different saltand detergent treatments indicated that the protein is most likelyperipherallyboundtothemembranes (Figure3C).TheassociationofMatR with the membranes is surprising, especially as its plastidialhomolog,MatK, is foundinthestromalchloroplast fraction(Zoschkeet al., 2010). Thus, it remains possible that under the low saltconditions used for mitochondrial subfractionation (see Methods),MatR interacts nonspecifically with the negatively charged organ-ellar membranes. Alternatively, the binding of MatR with themembranes might be facilitated by its association with membrane-bound ribonucleoprotein particle(s) (Uyttewaal et al., 2008).The accumulation of MatR and several other mitochondrial
proteins was studied during early developmental stages in Ara-bidopsis (Figures 4 and 5). The data indicate that the expressionand accumulation of MatR, at both the RNA and protein level, isupregulated during early seed imbibition but declines pro-gressively after the initiation stage as plant development pro-ceeds. These data suggest that the MatR protein is unstable andundergoes degradation via the proteolytic machinery in the mi-tochondria. The levels of VDAC1 were also higher in germinatingseeds compared mature (i.e., 3-week-old) seedlings. In Arabi-dopsis, VDAC1 is required for maintaining the steady state ofmitochondrial transmembrane potential (DC) and hence for op-timal respiratory activities, and its functions were found to becritical during the reproductive stage andduring early germination(Pan et al., 2014). In contrast to MatR and Porin, the steady-statelevels of various other mitochondrial proteins, including Nad9,CA2, RISP, COX2, and AtpB, all increased in during plant de-velopment, with the highest accumulation levels seen in matureplants. In accordance with these data, faster turnover rates werelinked toproteins involved inDNA/RNAmetabolismand thestressresponse in themitochondria, whereas subunits of the respiratorymachinery are generally more stable in Arabidopsis cells (Nelsonet al., 2013, 2014).
Unlike Canonical Maturases, MatR Is Involved in theSplicing of Many Group II-Containing Pre-RNAs inBrassicales Mitochondria
Weexpected thatMatRwouldbe involved in thesplicingofgroup IIintrons in the plant mitochondria. To establish the roles of MatR inmitochondrial RNA metabolism, we used two complementaryapproaches. The first strategy involved the identification of mt-RNAs that are stably associated with MatR in vivo using RIP-chipassays (Schmitz-Linneweber et al., 2005; Keene et al., 2006;Townley-Tilson et al., 2006). RNAs identified by the RIP-chipmethod involved different transcripts that all contained group IIintrons, including nad1 i1, i3, and i4, nad4 i1, nad5 i3 and i4, nad7i2, rpl2 i1, and rps3 i1 (Figure 6; Supplemental Figure 4). Se-quencingdata frommt-RNAs,whichcoprecipitatedwith theMatRprotein (Supplemental File 1), indicated that the signals detectedby the microarrays in the MatR co-IPs likely corresponded tounsplicedprecursorRNAsor thatMatR is also associatedwith thespliced exons in vivo. Accordingly, the 3D structure of the ribo-nucleoprotein complex of LtrA bound to its cognate ltrB intronRNA indicated that the spliced exons remain associated with theLtrAprotein,while the LtrA-depleted ltrB intron structures lack any
MatR Role in Mitochondria Group II Intron Splicing 2819

exon sequences (Qu et al., 2016). However, LtrA binds less tightlyto the spliced ltrB intron RNA than to the exon-containing pre-cursor RNA, and the binding of LtrA to the intron lariat can beenhanced in the presence of oligoribonucleotides, correspondingto sequences of intron-exon binding sites. The RIP-chip datasuggest that MATs in plant organelles also demonstrate highaffinities in their binding to precursor RNAs.
It was anticipated that MatR would associate in vivo with thoseintrons whose splicing is facilitated by the protein. However, thisassumption needed to be supported experimentally. Accordingly,the second approach involved a pioneering strategy for the down-regulation of the expression of mitochondrial genes by syntheticallydesignedribozymesthatareconjugated toa tRNA-likeshuttlevectorRNA. Using a similar approach, the expression of the mitochondrialatp9 mRNA and Atp9 subunit was reduced by ;80 and 50%, re-spectively (Val et al., 2011). As the functions ofMatRwere expectedtobeessential in theplant, theexpressionof thematRz1andmatRz2ribozymes (Figure 7) was regulated by the inducible b-estradiol-mediated gene expression system (Zuo et al., 2000).
The expression of matRz1 and matRz2 ribozymes resulted inreductions in MatR gene expression (up to 50% decrease in tran-script levels and 20 to 50% reductions in MatR protein levels inmatRz2andmatRz1, respectively) (Figure8;Supplemental Figure5),leading to splicing deficiencies in many organellar pre-mRNAs inboth transformant lines (Figure9B). Theseparticularly includednad1i3and i4, aswell as thesplicingofefficienciesofnad5 i4,nad7 i2, rpl2i1, and rps3 i1 pre-RNAs (Figure 9B). A reduction innad1 i3 pre-RNAmay correspond to an indirect effect, since the 39 part of this trans-spliced intron is cotranscribed with nad1 i4 and the matR ORF(Supplemental Figure 2). Thus, the cleavage ofmatR reading framemay also lead to RNA instability of nad1 i3. On the other hand, themajorityof the intronswhosesplicingwasaffectedbytheexpressionof thematR ribozymes were also found to be associated with MatRin vivo by the RIP-chip analysis (Figure 6; Supplemental Figure 4),including nad1 intron 3. It therefore anticipated that this intron, to-gether with multiple other group II introns highlighted in our ex-periments, is a processing substrate of MatR. Yet, some pre-RNAsthat were identified in the RIP-chip assay (Figure 6; SupplementalFigure 4) were not significantly affected in their splicing upondownregulatingmatRexpression (i.e.,nad1 i1 andnad4 i1; Figure9).Such differences between the RIP-chip data and the analysis ofsplicing efficiencies in matR knockdown lines by RT-qPCR maycorrespond to the limited resolution of the RNA footprints, but theymore likelycorrespond toonlyapartial reduction in theexpressionofMatRupon the induction ofmatRz1ormatRz2 ribozymeexpression(Figure 8; Supplemental Figure 5). Likewise, the nucleus-encodednMAT1, nMAT2, and nMAT4 proteins also facilitate the splicing ofdifferent subsets of mitochondrial introns (Keren et al., 2009, 2012;Cohen et al., 2014), while the plastidial homolog, MatK, was shownto be associated withmany of the subgroup-IIA intron RNAs in vivo(Zoschke et al., 2010). Table 1 summarizes the putativeRNA targetsof the MatR protein in Brassicales mitochondria.
MatR Functions Are Likely to Be Critical during Early Stagesin Plant Development
As indicated in Figure 8, splicing efficiencies were only partiallyaffected in matRz1 and matRz2 lines, and significant levels of
mature transcripts (mRNAs) corresponding to nad1, nad4, nad5,nad7, cox2, ccmFc, rpl2, and rps3 were seen in both transgeniclines. It is possible that in addition toMatR, the splicing of some ofthese mt-RNAs also involves additional nucleus-encoded factors(reviewed in Brown et al., 2014; Schmitz-Linneweber et al., 2015).For example, both nMAT4 and mCSF1 are involved in the mat-uration of nad1 i3, while the splicing of nad1 i4 requires at leastnMAT4 andMatR (Zmudjak et al., 2013; Brown et al., 2014;Cohenet al., 2014; Schmitz-Linneweber et al., 2015). Yet, it is also an-ticipated that theRNAphenotypesweobserve correspond toonlya partial silencing of MatR expression in thematRz lines (Figure 8;Supplemental Figure 5). We assume that the complete loss ofMatRwould lead to farmore profounddefects in the processingofthese RNAs, leading to embryo-lethal developmental defects.Accordingly,while acomplex I knockout is expected tobeviable inplants (Kühn et al., 2015; Fromm et al., 2016; Ostersetzer-Biran,2016), the functions of other respiratory complexes, includingCIII,CIV, and the translation machinery are considered essential fornormal embryo development (Gu et al., 1994; Berg et al., 2005;Meyer et al., 2005; Francs-Small et al., 2012; Dahan et al., 2014).While the splicing efficiencies of various transcripts were af-
fected upon matR knockdown, and their corresponding mRNAlevels were reduced in the transformants, several mt-mRNAsaccumulated to rather higher levels in both the matRz1 andmatRz2 lines (Figure 8B; Supplemental Figures 5 and 7). Theseinclude transcripts corresponding to nad2 exons 3-4 and nad5exons 1-2 in thematRz1 andmatRz2 lines. An upregulation in thesteady state levels of nad2 and nad5 RNAs in matR knockdownplants may correspond to the accumulation of precursor tran-scripts that are correctly processed by various other splicingcofactors, but cannot be spliced together with the other exons,of which the maturation relies on MatR. Analogously to inducedmatRz1 and matRz2 lines, increases in the levels of variousmRNAs were also apparent in nmat1 (Keren et al., 2012) andnmat4 (Cohen et al., 2014) mutants, which are also affected inthe maturation of nad1 and show reduced complex I levels. Inthe matR knockdown lines, these may involve small increases inlevels of the ATP-synthase subunits atp6 (in both matRz1 andmatRz2 lines) and atp8 (only in matRz1, also annotated as orfB)(Supplemental Figure 6). An upregulation in the steady state levelsof mRNAs in mutants affected in mitochondrial RNA metabolismand complex I biogenesis may correspond to compensatory ef-fects due to the altered organellar functions, involving reducedrespiration andaltered cellularmetabolism (Keren et al., 2012).Weanticipate that a more profound effect on MatR expression wouldresult innotable changes in the levelsof variousothermt-RNAs, asseen in the cases of nmat1 and nmat4 mutants.
Seed Plants with Unusual matR Genes Have Also Lost ManyGroup II Introns from Their Mitochondrial Genomes
The matR ORF has been retained as a putatively functional genewithin the nad1 i4 intron in the mitochondrial genomes of nu-merous plant species, and until recently it was believed to beinvariantly present at this position in all seed plants. In fact, recentsequencing has demonstrated that thematR gene can exist in themtDNAs of several species (most Geraniaceae, the mistle-toe Viscum album, the gnetophyte Welwitschia mirabilis) as
2820 The Plant Cell

a freestanding gene (Park et al., 2015; Petersen et al., 2015; Guoet al., 2016), while in some plants, including Viscum scurruloi-deum, Pelargonium x hortorum, Croizatia brevipetiolata, andLachnostylis bilocularis, the matR gene has been lost from theirmtDNAs (Wurdack and Davis, 2009; Skippington et al., 2015;Supplemental Table 4.
Except for Croizatia and Lachnostylis, for which mitochondrialintron content is not known, all of the other species with unusualmatR genes have also experienced substantial loss of theirmitochondrial group II introns (Supplemental Table 4). InV. scurruloideum, no nuclear copy of matR was detected in thesequencing data (Skippington et al., 2015), and all mitochondrialintrons spliced by MatR have been lost, suggesting that MatRfunction has been lost completely from this species. In P. hor-torum, although the mitochondrial matR gene was lost from themtDNA, copies of the gene were found in the nuclear genome byDNA-seq analysis of P. hortorum plants (Supplemental Table 4).Comparisons of the deduced amino acid sequences between thenuclear genes and canonical (i.e., mitochondria-encoded) MatRproteins suggest that the original Ph.matR gene has been split inthe nucleus into two isoforms, denoted here as nMAT-X andnMAT-RT, both of which are predicted by their N-terminal se-quences to be targeted to the mitochondria (Figure 2). ThenMATR-X isoform contains an intact domain X and parts of thefingers-palm RT sequence blocks (5 to 7), whereas the nMatR-RTisoform harbors the upstream region with the remaining RT 2 to4motifs, which are conserved in themitochondria-encodedmatRgenes. In other Geraniaceae species, thematR gene was moved
from its ancestral position in the nad1 i4 intron to becomea freestanding gene in themitochondrial genome. Althoughmanyintrons were lost from the mitochondrial genomes of all Ger-aniaceae species, the nad7 i2 gene is still present, consistent witha need for retained function for MatR and thus explaining theretention of matR. It is also possible that MatR may function inthese species on additional intron targets. In W. mirabilis, nu-merous introns have been lost from the mitochondrial genome,including most, but not all, of the splicing targets of MatR inBrassicales. Here again, the matR gene is in an unusual config-uration, but it has not been lost from the genome due to thepresence of nad1 i3, nad1 i4, and nad7 i2, which require MatRactivity to be spliced out (Supplemental Table 4).Combined, theseobservationsprovide furtherevidence that the
matR locus encodes a functional protein, which is required for theprocessing of many group II introns. Only after all intron targets ofMatR have been lost can the matR gene be fully lost from thegenome, as seen in V. scurruloideum (Supplemental Table 4)(Skippingtonet al., 2015). TheV. albummitochondrial genomehasalso lost all introns that require MatR activity for removal, yeta nearly completematR gene has still been retained. However, the39 truncation of ;150 bp suggests that the gene may be in theinitial stages of degradation, and it is possible that the gene is nolonger transcribed, although transcriptional activity was not as-sayed in the previous study (Petersen et al. 2015). In Geraniaceaeand Welwitschia, the loss of most introns processed by MatRcorrelates with amodified configuration for thematR gene. Thesefindings suggest that the loss of several intron targetsmay reducefunctional constraints on localization or cotranscription with thenad1 gene, enablingmatR to persist as a freestanding gene in themitochondrial or nuclear genome and be transcribed indepen-dently from nad1. Nevertheless, because at least one intron splicedout by MatR remains, thematR gene has still been retained in someform. These scenarios could signify an intermediate stage of evo-lution where matR activity is reduced and may eventually be lostupon loss of the remaining matR-processed introns.
MatR: An Evolutionary Step between the Highly“Specialized” Bacteria Maturases and the GeneralSpliceosomal Machinery in Eukaryotes?
Group II introns are catalytic RNAs that exist in the genetic sys-tems of the threemajor domains of life (i.e., archaea, bacteria, andeukaryotes). Based on structural features, the similarities of exon-intron boundaries and a common splicing mechanism, group IIintrons are proposed to be the progenitors of the eukaryoticspliceosome system (Schmitz-Linneweber et al., 2015). It hasbeenpostulated thatgroup II introns invaded thenucleargenomesof eukaryotes early during the evolutionary transition from an(a-proteo)-bacterial endosymbiont to the mitochondrion. Insupport of this idea, some bacterial maturases have the ability tosupport retrohoming, and recent studies have shown homologybetweengroup IImaturasesand thecorespliceosomalPrp8 factor(Dlakic and Mushegian, 2011). The structures of bacterial ma-turases bound to their host RNAs (Qu et al., 2016; Zhao and Pyle,2016) further support this intriguing theory. Yet, it still remainsunclear how the universal spliceosomal splicing machineryevolved from the bacterial group II introns and their related
Table 1. List of Identified Group II Intron Targets of MatR Protein inBrassicales
Gene/Intron Configuration RIP-Chip matR Knockdowns
matRrpl2 i1 cis + +rps3 i1 cis + +cox2 i1 cis 2 +ccmFc i1 cis 2 +nad1 i1 trans + 2
nad1 i2 cis (+) 2
nad1 i3 trans + +nad1 i4 cis + +nad2 i1 cis (+) 2
nad2 i2 trans (+) 2
nad2 i3 cis 2 2
nad2 i4 cis 2 (+)nad4 i1 cis (+) 2
nad4 i2 cis (+) 2
nad4 i3 cis 2 (+)nad5 i1 cis 2 2
nad5 i2 trans 2 2
nad5 i3 trans + (+)nad5 i4 cis + +nad7 i1 cis 2 2
nad7 i2 cis + +nad7 i3 cis 2 2
nad7 i4 cis 2 2
Parentheses indicate the data that could not be fully supported by theRIP-chip assays or the matR-knockdown experiments.
MatR Role in Mitochondria Group II Intron Splicing 2821

monospecific maturase factors. Throughout their evolution, or-ganellar introns inplantshavedivergedconsiderably fromtheir self-splicing group II RNAs (Bonen, 2008). This divergence in sequenceand structure has been accompanied by the acquisition of a largenumber of accessory protein cofactors. All of the plant maturases(i.e., MatR, MatK, and the nMATs) act on multiple pre-RNA targetsand function in the splicing of group II RNAs. Also required area number of additional (nucleus-encoded) splicing cofactors,possibly acting as an “organellar proto-spliceosome” (Schmitz-Linneweber et al., 2015). It is possible that in parallel to the invasionof group II introns into the nucleus of eukaryotes, a general ma-turase has evolved early during evolution to function in the splicingofmanyRNAtargets inasimilarmanner to theacquisitionofseveralorganellar intron targets by the organellar maturases in plants.
Our hypothesis is that the MatR protein is found in the mito-chondria in RNP particles containing different group II introns andvarious splicing cofactors. The generalization of the intron targetsofMatRmakes thisproteinan interestingmodel for theevolutionofthe nuclear spliceosomal machinery, which also underwenta process of splicing-target generalization. Interestingly, MatRlacks theNTD,which in thecaseof thebacterial LtrA is required forits associationwith theDIV domain of the ltrB intron (or possibly tocontrol its own translation, in vivo) (Qu et al., 2016), and alsoharbors two amino acids sequence insertions within the fingers-palm region (Figure 1B). The lack of a NTD subdomain and thepresence of additional motifs within the RT (i.e., fingers-palm)domainmay coincidewith the loss of a specific target in parallel tothe acquisition of multiple intron RNA targets. Likewise, the Prp8protein also harbors additional sequence blocks within the fingers-palm region of the RT domain, which functions in intron binding(Galej et al., 2013; Yan et al., 2015; Qu et al., 2016).
METHODS
Plant Material and Growth Conditions
Arabidopsis thaliana ecotype Columbia (Col-0) seeds were obtained fromthe ARBCatOhio StateUniversity. Prior to germination, seeds ofwild-typeormatRz/sdhRz transformantswere surface-sterilizedwith bleach (sodiumhypochlorite) solution and sown on MS-agar plates containing 1% (w/v)sucrose. The plates were kept in the dark for a few days at 4°C and thentransferred to controlled temperature and light conditions in growthchambers. Wild-type plants were grown in a controlled environment plantgrowth chamber (Arabidopsis Chamber AR-41L3; Percival Scientific) at22°Cwith 50% relative humidity, under short-day conditions (i.e., 16 hdarkand 8 h light), at ;150 mE m22 s21 (Philips bulbs type F17T8/L841).Cauliflower (Brassica oleracea var botrytis) inflorescenceswere purchasedfresh at local markets.
Gene Constructs for Ribozyme-Mediated Knockdown of matR
Constructs for the expression of the matRz1, matRz2, and sdhRz ribo-zymes associated with the PKTLS shuttle were prepared by PCR ampli-fication with the large direct primers dirRz1matrXho, dirRz3matrXho, anddirRzsdhXho, respectively (Supplemental Table 3). These primers in-cluded, from 59 to 39, an XhoI site, the complete sequence of the relevantribozyme, the linker sequence, and nucleotides 100 to 120 from the 39-endof the Turnip yellow mosaic virus (TYMV) genomic sequence. The re-vHDVSpe reverse primer (Supplemental Table 3B) included, from 59 to 39,an SpeI site and the reverse complement to nucleotides 1 to 20 from the
39-end of the Hepatitis delta virus antigenomic cis-cleaving ribozyme (cHDV)(Perrotta and Been, 1991). The pBI-PSTYPKTLScHDV plasmid (Val et al.,2011), containing the last 186 nucleotides from the 39-end of the TYMVgenomic sequence fused to the sequence of the cHDV cis-ribozyme, wasused as a template. The resulting PCR products, encoding the trans-ribozymes combined with the linker, the PKTLS, and the cHDV cis-ribozyme,were directly cloned into the pGEM-T vector (Promega) for sequencing,re-excised with the XhoI and SpeI restriction endonucleases, and clonedinto the XhoI and SpeI sites of the estradiol-inducible transcription unit ofthepER8vector (Zuoet al., 2000). Transcriptionof theseconstructsbyRNApolymerase II yields 59-capped transcripts comprising the matRz1,matRz2, or sdhRz ribozyme, the linker, the PKTLS, the cHDV, the termi-nationsequenceandapoly(A) tail. Self-cleavageof thecHDVeliminates thetermination sequence and the poly(A) tail and releases the regular 39-CCAend of the PKTLS. The resulting ribozyme-linker-PKTLS RNA is exportedfrom the nucleus to the cytosol, recognized by the mitochondrial tRNAimport machinery, and imported into the organelles (Val et al., 2011).
Nuclear Transformation and Chimeric Ribozyme Expression
The recombinant pER8 plasmids carrying the ribozyme-linker-PKTLS-cHDV constructs (Figure 7; Supplemental Table 3B) served to transformArabidopsis (Col-0) plants via Agrobacterium tumefaciens through floraldip (Clough andBent, 1998). Primary transformantswere selected on solidagar medium containing hygromycin. Resistant plants were grown in thegreenhouse to the next generations and homozygous transformants wereselected for further studies. To induce ribozyme expression and matRmRNA knockdown, seedlings were grown on solid agar medium incomplete darkness or in long daylight conditions (16 h light/8 h dark). Incomplete dark conditions, the agar medium was overlaid with 5 mMb-estradiol-supplemented liquid medium for transgene induction. In lightconditions, the seedlings were transferred to 6-well culture plates con-taining liquid medium supplemented with estradiol, with the roots dippinginto the medium. The matRz1 and matRz2 lines represent two individualtransformant lines, expressing the trans-ribozymes designed to specifi-cally cleave the ArabidopsismatR transcript (ArthMp045) in different geneloci (Figure 7). The sdh3 line expresses a ribozyme (sdhRz) designedagainst the mitochondrial sdh3 in tobacco (Nicotiana tabacum; NitaMp030),which was used as a control for the matR knockdown experiments, since ithas no corresponding targets in the Arabidopsis mtDNA (NC_001284).Positive transformant lines (i.e., sdh3, matRz1, and matRz2) were furtherscreened based on the lack of obvious phenotypes in the uninduced state,while showing a notable expression levels upon estradiol induction. Forthe analysis ofmatR knockdown effects on mitochondrial gene expressionand biogenesis, we selectedmatRz plants, which showed equivalent levelsof transgene (i.e., matRz1 or matRz2) expression to those seen for thesdhRz construct in the sdh3 line. Full biological replicates (i.e., separateexperiments) were then run with each selected transformant. We useda transient induction (i.e., 5 to ;6 d) for the expression of each ribozymetransgene, as estradiol may be metabolized by the plant cells (Janeczkoand Skoczowski, 2005).
RNA Extraction and Analysis
RNA extraction was performed essentially as described previously (Kerenet al., 2011; Cohen et al., 2014). In brief, RNA was prepared followingstandard TRI reagent protocols (Molecular Research Center) with addi-tional phenol/chloroformextraction. TheRNAwas treatedwithRNase-freeDNase (Ambion)prior to itsuse in theassays.RT-qPCRwasperformedwithspecific oligonucleotides designed to intron-exon regions (pre-mRNAs)andexon-exon (mRNAs) regionscorresponding to the23 intron-containingmitochondrial transcripts in wild-type and matR knockdown (matRz)plants, generally as previously described (Koprivova et al., 2010; Kühnet al., 2011). Reverse transcription was performed with the Superscript III
2822 The Plant Cell

reverse transcriptase (Invitrogen), using1 to2mgof total RNAand250ngofa random hexanucleotide mixture (Promega) and incubated for 50 min at50°C. Reactions were stopped by 15 min incubation at 70°C and the RTsamplesserveddirectly forPCR.qPCR reactionswere runonaLightCycler480 (Roche), using 2.5mL of LightCycler 480 SYBRGreen IMastermix and2.5 mM forward and reverse primers in a final volume of 5 mL. Reactionswere performed in triplicate in the following conditions: preheating at 95°Cfor 10 min, followed by 40 cycles of 10 s at 95°C, 10 s at 58°C, and 10 s at72°C. For the analysis of MatR expression, the nuclear genes ACTIN2,GAPDH, and EXPRESSED were taken as reference genes (Czechowskiet al., 2005). For the analysis of mitochondrial gene expression and RNAaccumulation, the nucleus-encoded 18S rRNA subunit and the mito-chondrial 26S rRNA (rrn26) subunit were used as reference genes in theqPCR analyses (Koprivova et al., 2010; Kühn et al., 2011).
Transcriptome Mapping by High-Throughput (RNA-Seq) Analysis
Total mt-RNA was extracted from highly enriched mitochondria prepa-rations obtained from cauliflower inflorescence (see below). Sequencingwas performed on an Illumina Genome Analyzer (The Genome High-Throughput Sequencing Laboratory, Tel-Aviv University, Israel) to gen-erate transcriptome profiles and evaluated by mapping of the cauliflowermt-RNA sequences onto the genomic reference sequence using bowtie2.1.0 with default parameters, essentially as described by Grewe et al.(2014). To reduce mapping of chloroplast transcript reads to MIPT (mi-tochondrial DNA of plastid origin) regions or putative paralogous mito-chondrial genes (e.g., plastid atpA and mitochondrial atp1), we added theBrassica napus chloroplast genome sequence (NC_016735) to the mito-chondrial reference sequence as bait for native plastid sequences. Bowtieresults in SAM format were aligned using utilities from SAMtools (Li et al.,2009). A custom Perl script was used to determine the depth and directionof transcriptionateachnucleotide in the referencegenome.Theexpressionlevel of each genewasdeterminedby the average of transcript coverage ateach nucleotide.
Generation of Rabbit Polyclonal Antibodies against the MatR Protein
Since no antibodies to MatR protein were available prior to this study,a polyclonal antibody was raised against a synthetic peptide corre-sponding to a unique region in MatR’s RT domain (i.e., NH3
+-RRIDD-QENPGEEASFNA), which is conserved among different Brassicales(Supplemental Figure 1). The antibody was purified from the serum byaffinity chromatography column and was then tested for its ability torecognize the immunizingpeptide.Theaffinityandspecificityof thepurifiedMatRantibodywere further evaluated inprotein gel blots using total proteinand enriched organelle preparations (i.e., mitochondria, chloroplasts, andperoxisomes) obtained from 3-week-old Arabidopsis var Columbia andpurified mitochondria extracted from cauliflower inflorescences.
RIP-Chip Assays
Mitochondrial preparations from cauliflower inflorescences were solubi-lized with 1%Nonidet P-40 (v/v) in assay buffer (150 mMNaCl and 10mMsodium phosphate buffer, pH 7.2) at a protein concentration of;1mg/mL.After 30 min incubation on ice, the organellar extract was centrifuged for10min at 21,000g. The clear supernatant was then incubated with affinity-purified anti-MatR antibodies conjugated to protein A/G Sepharose beads(Amersham),withgentlemixingat4°C for16h.Thebeadswerecollectedbya brief centrifugation (1 min at 1000g, at 4°C) and washed three times with0.6MNaCl, 0.5%(v/v)NonidetP-40, and50mMTris-HCl (pH8.3), followedby a single wash with 13 PBS buffer. Dot-blot analysis of coimmuno-precipitated RNA was performed essentially as described (Zoschke et al.,2010). Total mt-RNA coprecipitated with the anti-MatR antibodies wasDNase digested and then reverse transcribed using Superscript III reverse
transcriptase (Invitrogen) and a randomhexanucleotidemixture (Promega)in the presence of [a-32P] dCTP and [a-32P]dATP (to increase specificsignals). The identity of the proteins in the co-IPs was established by LC-MS/MS analysis (Smoler Proteomics Center, Technion, Israel). Com-mercial antibodies to SHMT were used as controls for the integrity of theRIP-chip method.
Mitochondria Extraction and Analysis
Preparations of highly enriched mitochondria from cauliflower inflores-cence generally followed the protocol described in detail by Neuwirt et al.(2005). About 1.0 kg fresh weight inflorescences (two to three heads ofcauliflower) were cut off from the stems and kept overnight (;16 h) in coldwater (4°C). On the followingmorning, the inflorescenceswere groundwithice-cold extraction buffer (0.9 M mannitol, 90 mM Na-pyrophosphate,6 mM EDTA, 2.4% PVP25 [w/v], 0.9% BSA [w/v], 9 mM cysteine, 15 mMglycine, and 6 mM b-mercaptoethanol, pH 7.5 ). Mitochondria were re-covered from the extract by differential centrifugations and purification onPercoll gradients. Following fractionation, themitochondria were pelleted,resuspended in a small volume of wash buffer (0.3 M mannitol, 10 mMK-phosphate, 1mMEDTA, pH7.5), aliquoted, and stored frozen at280°C.For the isolation of mitochondria from Arabidopsis, we used 2-week-oldseedlings grown on MS plates. The enriched organelle preparation wasperformed essentially as described (Keren et al., 2012).
Mitochondrial Subfractionation into Matrix and Membranes
Intact mitochondria were disrupted by several freeze-thaw cycles in washbuffer (i.e., 0.3Mmannitol, 10mMKH2PO4, and1mMEDTA, pH7.5), brieflysonicated, and then centrifuged at 25,000g for 20 min to separate thematrix (supernatant) andmembrane (pellet) fractions.Washbuffer, equal tothe initial volume, was added to the pellet and briefly sonicated. Wholemitochondria and the organellar subfractions were diluted in 1:1 ratio with33 Laemmli sample loading buffer (Laemmli, 1970) supplemented with50 mM b-mercaptoethanol, heated at 70°C for 10 min, and cleared bycentrifugation at 25,000g for 5 min. To determine whether membranousproteins are peripherally or integrally bound to the membranes, themembrane fraction (equivalent of 40 mg leaves) was prewashed with200mL 10mMTris-HCl, pH8.0 (for the removal of contaminating proteins),and pelleted by centrifugation for 8 min at 21,000g (4°C). The membraneswere then treated with either 1 M NaCl (removes loosely attached mem-brane proteins), 2 M NaBr (a chaotropic agent used for the removal ofperipherally bound proteins), or 0.05%Triton X-100, 1MNaCl (wash someperipheral proteins attached by surface hydrophobic interactions), gen-erally as described (Ostersetzer et al., 2007). The location of each protein(i.e., peripherally or integrally associated)wasdeterminedbycentrifugation(i.e., 8 min at 21,000g, 4°C) and immunoblot analysis of both the pellet(containing the membranes) and supernatant fractions.
Protein Extraction and Analysis
Protein extraction and analysis were performed generally as described(Keren et al., 2009). Total protein was extracted from 3-week-old Arabi-dopsis leaves or isolated mitochondria obtained from cauliflower in-florescences by the borate/ammonium acetate method (Maayan et al.,2008). For this purpose, frozen plant tissue was homogenized in thepresence of polyvinylpolypyrrolidone. The homogenate was added tomicrofuge tubes containing 400 mL ice-cold protein extraction buffer(50 mM Na-borate, 50 mM ascorbic acid, 1.25% [w/v] SDS, and 12.5 mMb-mercaptoethanol, pH 9.0) and the protease inhibitor cocktail completeMini from Roche Diagnostics. Proteins were recovered by centrifugation(25,000g) in the presence of three volumes of ice-cold 0.1 M ammoniumacetate in methanol buffer (NH4-OAc-MeOH), generally as described(Maayan et al., 2008). Protein concentration was determined according to
MatR Role in Mitochondria Group II Intron Splicing 2823

the Bradford method (Bio-Rad), with BSA used as a standard. Approxi-mately 20 mg total protein was mixed with an equal volume of 33 proteinsample buffer (Laemmli, 1970), supplemented with 50 mM b-mercap-toethanol, and subjected to 12% SDS-PAGE (at a constant 100 V). Fol-lowing electrophoresis, the proteinswere transferred to aPVDFmembrane(Bio-Rad) and blotted overnight at 4°C with specific primary antibodies.Detection was performed by chemiluminescence assays after incubationwith an appropriate horseradish peroxidase-conjugated secondary anti-body (Sigma-Aldrich or Santa Cruz).
For matRz and sdhRz Arabidopsis transformants, plant samples wereweighed, snap frozen in liquid nitrogen, and ground in liquid nitrogen withprechilled mortar and pestle. The fine frozen powder was transferred toa 1.5-mLmicrotube and 10mL extraction buffer (140mMTris base, 105mMTris-HCl, pH 7.0, 0.5 mM EDTA, 2% [w/v] SDS, 10% [v/v] glycerol, and0.1 mg/mL protease inhibitors [Roche]) was added per mg sample wetweight. The suspension was immediately refrozen in liquid nitrogen andsonicated until it was just thawed. The procedure was repeated three timeswith 1-min breaks in liquid nitrogen. The samples were subsequentlycentrifuged for 3 min at 10,000g to remove insoluble material and the su-pernatants were used for protein gel blot analysis. Proteins (50mg and 5mgforMatRandb-actindetection, respectively)were separatedon12.5% (w/v)SDS-PAGEgels andblottedontoPVDFmembranes. Immunodetectionwasperformed with a SNAP 2.0 protein detection system (Merck Milllipore),according tothemanufacturer’s instructions.MatRwasimmunoprobedwitha polyclonal rabbit antiserum raised against the overexpressed and affinity-purified N-terminal half of the protein. b-Actin was detected with a com-mercial mouse monoclonal antibody (Actin MA1-744; ThermoFisherScientific). Bound specific antibodies were revealed with biotinylated poly-clonal anti-rabbit IgG or monoclonal anti-mouse IgG secondary antibodies(Sigma-Aldrich), streptavidin-alkaline phosphatase conjugate (AmershamBiosciences), and FASTTM BCIP/NBT buffer (Sigma-Aldrich). Immunoblotsignals were quantified with the MultiGauge software (Fujifilm).
Seed Protein Extraction and Analysis
Total seed protein extracts were prepared from dry mature and viableseeds at different stages of imbibition and germination. About 100 mgArabidopsis seed (;5000 seeds; Meinke, 1994) was ground with mortarand pestle in the presence of 2 mL extraction buffer (75 mMMOPS-KOH,0.6Msucrose, 4mMEDTA, 0.2%PVP-40, 0.2%BSA, and 8mML-cystein,pH 7.6) and the protease inhibitor cocktail complete Mini from RocheDiagnostics. Crude membrane extracts were prepared essentially asdescribed (Francs-Small et al., 2012). The membranous fraction wasobtained by centrifugation at 22,000g for 10 min at 4°C. The pellet con-taining the crudemembranous fractionwaswashed twicewithwashbuffer(37.5 mM MOPS-KOH, 0.3 M sucrose, and 2 mM EDTA, pH 7.6). Thesamples were kept frozen at 280°C until used in denaturing (i.e., SDS-PAGE) or BN gel electrophoresis.
BN Electrophoresis for Isolation of Native Organellar Complexes
BN-PAGEof crudemembranous fractionswasperformedaccording to themethod described by Zmudjak et al. (2013). An aliquot equivalent to 25mgof crude membrane extracts, obtained from Arabidopsis (Col-0) seeds orleaves from 3-week-old seedlings, was solubilized with n-dodecyl-b-maltoside (1.5% [w/v]) in ACA buffer (750 mM amino-caproic acid,0.5 mM EDTA, and 50mM Tris-HCl, pH 7.0), and then incubated on ice for30 min. The samples were centrifuged 8min at 20,000g, and Serva Blue G(0.2% [v/v]) was added to the supernatant. The samples were then loadedonto a native 4 to 16% gradient gel. For nondenaturing-PAGE-protein gelblotting, thegelwas transferred toaPVDFmembrane (Bio-Rad) inCathodebuffer (50 mM Tricine and 15 mM Bis-Tris-HCl, pH 7.0) for 16 h at 4°C(constant current of 40 mA). The membrane was incubated with variousantibodies, as indicated in each blot, and detection was performed by
chemiluminescence assay after incubation with an appropriate horse-radish peroxidase-conjugated secondary antibody.
Accession Numbers
Sequence data from this article can be found in the GenBank/EMBL li-braries under the following accession numbers: Arabidopsis mitochondriagenome sequence (NC_001284); cauliflower mitochondria genome se-quence (KJ820683); Arabidopsis ACTIN2 (At3g18780); Arabidopsis atp6(ArthMp035); Arabidopsis atp8 (also annotated as orfB, ArthMp040);Arabidopsis EXPRESSED (At4g26410); ArabidopsisGADPH (At1g13440);Arabidopsis matR (ArthMp045); Arabidopsis mitochondrial rRNA subunit18S protein (At3g41768); Arabidopsis mitochondrial 26S rRNA (rrn26)subunit (ArthMr001); tobacco sdh3 (NitaMp030); and TYMV genome se-quence (X16378).
Supplemental Data
Supplemental Figure 1. Alignment of MatR homologs from variousangiosperms.
Supplemental Figure 2. Transcription profile of the nad1 ex4-int4-ex5gene locus in cauliflower mitochondria according to RNA-seq analysis.
Supplemental Figure 3. Analysis of matR transcription in Brassica-ceae.
Supplemental Figure 4. Identification of MatR-associated mt-RNAsby the RIP-chip method.
Supplemental Figure 5. Expression of various mitochondrial RNAs inmatR knockdown lines.
Supplemental Figure 6. Phenotypes associated with Arabidopsisthaliana transgenic lines expressing the matRz and sdh3 ribozymes.
Supplemental Figure 7. Accumulation of mitochondrial mRNAs inmatR knockdown lines.
Supplemental Figure 8. Homology modeling of the Arabidopsisthaliana maturase-related (MatR) protein.
Supplemental Table 1. List of antibodies used for the analysis ofMatR.
Supplemental Table 2. Proteins identified by LC-MS/MS analysis inthe RIP-chip assays.
Supplemental Table 3. Lists of oligonucleotides.
Supplemental Table 4. Comparison ofmatR status and mitochondrialintron content among selected seed plants.
Supplemental File 1. Identification of MatR-associated mitochondrialRNAs by RNA-coimmunoprecipitation/cDNA sequencing.
ACKNOWLEDGMENTS
We thank Christian Schmitz-Linneweber (Humboldt University, Berlin) forhelpwithmitochondriaRNA-seqdata anddot-blot analysis of RNAco-IPs,IanSmall (University ofWesternAustralia) for the assistancewithRT-qPCRanalyses, Yoram Eyal (The Volcani Research Instute) for providing theRACE protocols, Amalia Biran-Ostersetzer for assistance with mitochon-dria RNA-seq analysis, Maciej Szymanski (A. Mickiewicz University,Poznan) for help in ribozyme design, Marta Gabryelska (PAN-IBCH,Poznan) for help in ribozyme testing, Anne Cosset for help in transformantanalysis, and Sam Aldrin for his careful reading of the manuscript andpoints raised. Thisworkwassupportedbygrants toO.O.-B. from the IsraeliScience Foundation (No. 741/15) and in part from the German-Israeli
2824 The Plant Cell

Foundation (GIF 1213/2012) to A.D., by the French State program “Invest-ments for the future” (LABEX ANR-11-LABX-0057_MITOCROSS), and bythe French National Research Agency (ANR-06-MRAR-037-02 and ANR-09-BLAN-0240-01). This work was also supported by a grant from thePolish National Science Centre (UMO-2013/09/B/NZ1/03359) to J.B. andby grants from the U.S. National Science Foundation (IOS-1027529 andMCB-1125386) to J.P.M. Regular funding to A.D. from the FrenchNationalCenter for Scientific Research (CNRS-UPR2357) and the University ofStrasbourg is acknowledged.
AUTHOR CONTRIBUTIONS
L.D.S. did most of the biochemical analysis of MatR expression andperformed co-IP experiments, analysis of the RNA profiles of cauliflowerand Arabidopsis mitochondria, and the analysis ofmatR-knockdown linesby RT-qPCR. D.M. was involved in the establishment ofmatRz transgeniclines and analysis of the mt-RNA profiles of matR-knockdown lines. F.G.assistedwith the sequencing and analysis of plantmitochondrial genomesand preformed bioinformatics analysis of mitochondria transcriptomes,intron distribution in angiospermmtDNAs, and MatR status in plants. K.R.assistedwith theRNAandprotein analysesofmatR-knockdown lines. S.A.produced antibodies to MatR and assisted in the RIP-chip analyses ofcauliflower mitochondria MatR protein. P.G. did RNA and RT-qPCR anal-yses ofmatR-knockdown lines. A.K.N. participated in the development ofthe sdhRz ribozyme system in Arabidopsis and tobacco. I.K. assisted withthe RIP-chip assays and performed hybridization assays of the RIP-chips.S.S. assisted with plant growth and analysis, protein and RNA extraction,and analysis. L.K. assisted with the 3D modeling of MatR structures. J.B.and A.D. were involved in the development of the mitochondrial ribozymesystem. J.B., J.P.M., A.D., and O.O.-B. supervised the study. O.O.-B.finalized the manuscript.
Received May 19, 2016; revised September 26, 2016; accepted October19, 2016; published October 19, 2016.
REFERENCES
Adams, K.L., Qiu, Y.-L., Stoutemyer, M., and Palmer, J.D. (2002).Punctuated evolution of mitochondrial gene content: high andvariable rates of mitochondrial gene loss and transfer to the nucleusduring angiosperm evolution. Proc. Natl. Acad. Sci. USA 99: 9905–9912.
Ahlert, D., Piepenburg, K., Kudla, J., and Bock, R. (2006). Evolu-tionary origin of a plant mitochondrial group II intron from a reversetranscriptase/maturase-encoding ancestor. J. Plant Res. 119: 363–371.
Aizawa, Y., Xiang, Q., Lambowitz, A.M., and Pyle, A.M. (2003). Thepathway for DNA recognition and RNA integration by a group IIintron retrotransposon. Mol. Cell 11: 795–805.
Arrieta-Montiel, M.P., Shedge, V., Davila, J., Christensen, A.C.,and Mackenzie, S.A. (2009). Diversity of the Arabidopsis mito-chondrial genome occurs via nuclear-controlled recombinationactivity. Genetics 183: 1261–1268.
Asakura, Y., and Barkan, A. (2007). A CRM domain protein functionsdually in group I and group II intron splicing in land plant chloro-plasts. Plant Cell 19: 3864–3875.
Barkan, A. (2011). Expression of plastid genes: organelle-specific elabo-rations on a prokaryotic scaffold. Plant Physiol. 155: 1520–1532.
Bégu, D., Mercado, A., Farré, J.C., Moenne, A., Holuigue, L., Araya, A.,and Jordana, X. (1998). Editing status of mat-r transcripts in
mitochondria from two plant species: C-to-U changes occur in pu-tative functional RT and maturase domains. Curr. Genet. 33: 420–428.
Berg, M., Rogers, R., Muralla, R., and Meinke, D. (2005). Re-quirement of aminoacyl-tRNA synthetases for gametogenesis andembryo development in Arabidopsis. Plant J. 44: 866–878.
Blocker, F.J.H., Mohr, G., Conlan, L.H., Qi, L., Belfort, M., andLambowitz, A.M. (2005). Domain structure and three-dimensionalmodel of a group II intron-encoded reverse transcriptase. RNA 11:14–28.
Bock, H., Brennicke, A., and Schuster, W. (1994). Rps3 and rpl16genes do not overlap in Oenothera mitochondria: GTG as a poten-tial translation initiation codon in plant mitochondria? Plant Mol.Biol. 24: 811–818.
Bonen, L. (2008). Cis- and trans-splicing of group II introns in plantmitochondria. Mitochondrion 8: 26–34.
Bonen, L., and Vogel, J. (2001). The ins and outs of group II introns.Trends Genet. 17: 322–331.
Brown, G.G., Colas des Francs-Small, C., and Ostersetzer-Biran,O. (2014). Group II intron splicing factors in plant mitochondria.Front. Plant Sci. 5: 35.
Cech, T.R. (1986). The generality of self-splicing RNA: relationship tonuclear mRNA splicing. Cell 44: 207–210.
Chapdelaine, Y., and Bonen, L. (1991). The wheat mitochondrialgene for subunit I of the NADH dehydrogenase complex: a trans-splicing model for this gene-in-pieces. Cell 65: 465–472.
Clough, S.J., and Bent, A.F. (1998). Floral dip: a simplified method forAgrobacterium-mediated transformation of Arabidopsis thaliana.Plant J. 16: 735–743.
Cohen, S., Zmudjak, M., Colas des Francs-Small, C., Malik, S.,Shaya, F., Keren, I., Belausov, E., Many, Y., Brown, G.G., Small,I., and Ostersetzer-Biran, O. (2014). nMAT4, a maturase factorrequired for nad1 pre-mRNA processing and maturation, is essen-tial for holocomplex I biogenesis in Arabidopsis mitochondria. PlantJ. 78: 253–268.
Cousineau, B., Smith, D., Lawrence-Cavanagh, S., Mueller, J.E.,Yang, J., Mills, D., Manias, D., Dunny, G., Lambowitz, A.M., andBelfort, M. (1998). Retrohoming of a bacterial group II intron: mo-bility via complete reverse splicing, independent of homologousDNA recombination. Cell 94: 451–462.
Cui, X., Matsuura, M., Wang, Q., Ma, H., and Lambowitz, A.M.(2004). A group II intron-encoded maturase functions preferentiallyin cis and requires both the reverse transcriptase and X domains topromote RNA splicing. J. Mol. Biol. 340: 211–231.
Czechowski, T., Stitt, M., Altmann, T., Udvardi, M.K., and Scheible,W.-R. (2005). Genome-wide identification and testing of superiorreference genes for transcript normalization in Arabidopsis. PlantPhysiol. 139: 5–17.
Dahan, J., Tcherkez, G., Macherel, D., Benamar, A., Belcram, K.,Quadrado, M., Arnal, N., and Mireau, H. (2014). Disruption of theCYTOCHROME C OXIDASE DEFICIENT1 gene leads to cytochromec oxidase depletion and reorchestrated respiratory metabolism inArabidopsis. Plant Physiol. 166: 1788–1802.
Dalby, S.J., and Bonen, L. (2013). Impact of low temperature onsplicing of atypical group II introns in wheat mitochondria. Mito-chondrion 13: 647–655.
Dlakic, M., and Mushegian, A. (2011). Prp8, the pivotal protein of thespliceosomal catalytic center, evolved from a retroelement-encodedreverse transcriptase. RNA 17: 799–808.
Ems, S.C., Morden, C.W., Dixon, C.K., Wolfe, K.H., dePamphilis,C.W., and Palmer, J.D. (1995). Transcription, splicing and editingof plastid RNAs in the nonphotosynthetic plant Epifagus virginiana.Plant Mol. Biol. 29: 721–723.
MatR Role in Mitochondria Group II Intron Splicing 2825

Eyal, Y., Neumann, H., Or, E., and Frydman, A. (1999). Inverse single-strand RACE: an adapter-independent method of 59 RACE. Bio-techniques 27: 656–658.
Farré, J.C., and Araya, A. (1999). The mat-r open reading frame istranscribed from a non-canonical promoter and contains an internalpromoter to co-transcribe exons nad1e and nad5III in wheat mito-chondria. Plant Mol. Biol. 40: 959–967.
Ferat, J.L., and Michel, F. (1993). Group II self-splicing introns inbacteria. Nature 364: 358–361.
Forner, J., Weber, B., Thuss, S., Wildum, S., and Binder, S. (2007).Mapping of mitochondrial mRNA termini in Arabidopsis thaliana:t-elements contribute to 59 and 39 end formation. Nucleic Acids Res.35: 3676–3692.
Francs-Small, C.C., Kroeger, T., Zmudjak, M., Ostersetzer-Biran,O., Rahimi, N., Small, I., and Barkan, A. (2012). A PORR domainprotein required for rpl2 and ccmF(C) intron splicing and for thebiogenesis of c-type cytochromes in Arabidopsis mitochondria.Plant J. 69: 996–1005.
Fromm, S., Senkler, J., Eubel, H., Peterhansel, C., and Braun, H.P.(2016). Life without complex I: proteome analyses of an Arabidopsismutant lacking the mitochondrial NADH dehydrogenase complex.J. Exp. Bot. 67: 3079–3093.
Funk, H.T., Berg, S., Krupinska, K., Maier, U.G., and Krause, K.(2007). Complete DNA sequences of the plastid genomes of twoparasitic flowering plant species, Cuscuta reflexa and Cuscutagronovii. BMC Plant Biol. 7: 45–48.
Galej, W.P., Oubridge, C., Newman, A.J., and Nagai, K. (2013).Crystal structure of Prp8 reveals active site cavity of the spliceo-some. Nature 493: 638–643.
Germain, A., Hotto, A.M., Barkan, A., and Stern, D.B. (2013). RNAprocessing and decay in plastids. Wiley Interdiscip. Rev. RNA 4:295–316.
Gordon, K., Fütterer, J., and Hohn, T. (1992). Efficient initiation oftranslation at non-AUG triplets in plant cells. Plant J. 2: 809–813.
Grewe, F., Edger, P.P., Keren, I., Sultan, L., Pires, J.C.,Ostersetzer-Biran, O., and Mower, J.P. (2014). Comparativeanalysis of 11 Brassicales mitochondrial genomes and the mito-chondrial transcriptome of Brassica oleracea. Mitochondrion 19:135–143.
Gu, J., Dempsey, S., and Newton, K.J. (1994). Rescue of a maizemitochondrial cytochrome oxidase mutant by tissue culture. Plant J.6: 787–794.
Gualberto, J.M., Mileshina, D., Wallet, C., Niazi, A.K., Weber-Lotfi,F., and Dietrich, A. (2014). The plant mitochondrial genome: dy-namics and maintenance. Biochimie 100: 107–120.
Guo, W., Grewe, F., Fan, W., Young, G.J., Knoop, V., Palmer, J.D.,and Mower, J.P. (2016). Ginkgo and Welwitschia mitogenomesreveal extreme contrasts in gymnosperm mitochondrial evolution.Mol. Biol. Evol. 33: 1448–1460.
Guo, W., and Mower, J.P. (2013). Evolution of plant mitochondrialintron-encoded maturases: frequent lineage-specific loss and re-current intracellular transfer to the nucleus. J. Mol. Evol. 77: 43–54.
Hammani, K., and Giegé, P. (2014). RNA metabolism in plant mito-chondria. Trends Plant Sci. 19: 380–389.
Hann, S.R. (1994). Regulation and function of non-AUG-initiatedproto-oncogenes. Biochimie 76: 880–886.
Hausner, G., Olson, R., Simon, D., Johnson, I., Sanders, E.R.,Karol, K.G., McCourt, R.M., and Zimmerly, S. (2006). Origin andevolution of the chloroplast trnK (matK) intron: a model for evolutionof group II intron RNA structures. Mol. Biol. Evol. 23: 380–391.
Hofacker, I.L., Fekete, M., and Stadler, P.F. (2002). Secondarystructure prediction for aligned RNA sequences. J. Mol. Biol. 319:1059–1066.
Huang, H.-R., Rowe, C.E., Mohr, S., Jiang, Y., Lambowitz, A.M.,and Perlman, P.S. (2005). The splicing of yeast mitochondrialgroup I and group II introns requires a DEAD-box protein with RNAchaperone function. Proc. Natl. Acad. Sci. USA 102: 163–168.
Huang, S., Shingaki-Wells, R.N., Taylor, N.L., and Millar, A.H.(2013). The rice mitochondria proteome and its response duringdevelopment and to the environment. Front. Plant Sci. 4: 16.
Jacoby, R.P., Millar, A.H., and Taylor, N.L. (2010). Wheat mito-chondrial proteomes provide new links between antioxidant de-fense and plant salinity tolerance. J. Proteome Res. 9: 6595–6604.
Janeczko, A., and Skoczowski, A. (2005). Mammalian sex hormonesin plants. Folia Histochem. Cytobiol. 43: 71–79.
Jenkins, B.D., Kulhanek, D.J., and Barkan, A. (1997). Nuclear mu-tations that block group II RNA splicing in maize chloroplasts revealseveral intron classes with distinct requirements for splicing factors.Plant Cell 9: 283–296.
Jensen, E.C. (2013). Quantitative analysis of histological staining andfluorescence using ImageJ. Anat. Rec. (Hoboken) 296: 378–381.
Källberg, M., Wang, H., Wang, S., Peng, J., Wang, Z., Lu, H., andXu, J. (2012). Template-based protein structure modeling using theRaptorX web server. Nat. Protoc. 7: 1511–1522.
Keene, J.D., Komisarow, J.M., and Friedersdorf, M.B. (2006). RIP-Chip: the isolation and identification of mRNAs, microRNAs andprotein components of ribonucleoprotein complexes from cell ex-tracts. Nat. Protoc. 1: 302–307.
Kelley, L.A., and Sternberg, M.J. (2009). Protein structure predictionon the Web: a case study using the Phyre server. Nat. Protoc. 4:363–371.
Keren, I., Bezawork-Geleta, A., Kolton, M., Maayan, I., Belausov,E., Levy, M., Mett, A., Gidoni, D., Shaya, F., and Ostersetzer-Biran, O. (2009). AtnMat2, a nuclear-encoded maturase required forsplicing of group-II introns in Arabidopsis mitochondria. RNA 15:2299–2311.
Keren, I., Shaya, F., and Ostersetzer-Biran, O. (2011). An optimizedmethod for the analysis of plant mitochondria RNAs by northern-blotting. Endocytobiosis Cell Res. 1: 34–42.
Keren, I., Tal, L., des Francs-Small, C.C., Araújo, W.L., Shevtsov,S., Shaya, F., Fernie, A.R., Small, I., and Ostersetzer-Biran, O.(2012). nMAT1, a nuclear-encoded maturase involved in the trans-splicing of nad1 intron 1, is essential for mitochondrial complex Iassembly and function. Plant J. 71: 413–426.
Kim, D.E., Chivian, D., and Baker, D. (2004). Protein structure pre-diction and analysis using the Robetta server. Nucleic Acids Res.32: W526–W531.
Knoop, V. (2012). Seed plant mitochondrial genomes: complexity evolv-ing. In Genomics of Chloroplasts and Mitochondria, R. Bock and V.Knoop, eds (East Lansing, MI: Michigan State University), pp. 175–200.
Köhler, D., Schmidt-Gattung, S., and Binder, S. (2010). The DEAD-box protein PMH2 is required for efficient group II intron splicing inmitochondria of Arabidopsis thaliana. Plant Mol. Biol. 72: 459–467.
Koprivova, A., des Francs-Small, C.C., Calder, G., Mugford, S.T.,Tanz, S., Lee, B.-R., Zechmann, B., Small, I., and Kopriva, S.(2010). Identification of a pentatricopeptide repeat protein impli-cated in splicing of intron 1 of mitochondrial nad7 transcripts.J. Biol. Chem. 285: 32192–32199.
Kozak, M. (1989). Context effects and inefficient initiation at non-AUGcodons in eucaryotic cell-free translation systems. Mol. Cell. Biol. 9:5073–5080.
Kozak, M. (1997). Recognition of AUG and alternative initiator codonsis augmented by G in position +4 but is not generally affected by thenucleotides in positions +5 and +6. EMBO J. 16: 2482–2492.
Kubo, T., and Newton, K.J. (2008). Angiosperm mitochondrial ge-nomes and mutations. Mitochondrion 8: 5–14.
2826 The Plant Cell

Kühn, K., Carrie, C., Giraud, E., Wang, Y., Meyer, E.H., Narsai, R.,des Francs-Small, C.C., Zhang, B., Murcha, M.W., and Whelan,J. (2011). The RCC1 family protein RUG3 is required for splicing ofnad2 and complex I biogenesis in mitochondria of Arabidopsisthaliana. Plant J. 67: 1067–1080.
Kühn, K., Obata, T., Feher, K., Bock, R., Fernie, A.R., and Meyer,E.H. (2015). Complete mitochondrial complex I deficiency inducesan up-regulation of respiratory fluxes that is abolished by traces offunctional complex I. Plant Physiol. 168: 1537–1549.
Laemmli, U.K. (1970). Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227: 680–685.
Lambowitz, A.M., and Belfort, M. (1993). Introns as mobile geneticelements. Annu. Rev. Biochem. 62: 587–622.
Lambowitz, A.M., and Belfort, M. (2015). Mobile bacterial group II intronsat the crux of eukaryotic evolution. Microbiol. Spectr. 3: 1–26.
Lambowitz, A.M., and Zimmerly, S. (2011). Group II introns: mobile ri-bozymes that invade DNA. Cold Spring Harb. Perspect. Biol. 3: a003616.
Law, S.R., Narsai, R., Taylor, N.L., Delannoy, E., Carrie, C., Giraud,E., Millar, A.H., Small, I., and Whelan, J. (2012). Nucleotide andRNA metabolism prime translational initiation in the earliest eventsof mitochondrial biogenesis during Arabidopsis germination. PlantPhysiol. 158: 1610–1627.
Lee, C.P., Eubel, H., Solheim, C., and Millar, A.H. (2012). Mito-chondrial proteome heterogeneity between tissues from the vege-tative and reproductive stages of Arabidopsis thaliana development.J. Proteome Res. 11: 3326–3343.
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N.,Marth, G., Abecasis, G., and Durbin, R.; 1000 Genome ProjectData Processing Subgroup (2009). The Sequence Alignment/Mapformat and SAMtools. Bioinformatics 25: 2078–2079.
Li-Pook-Than, J., Carrillo, C., and Bonen, L. (2004). Variation inmitochondrial transcript profiles of protein-coding genes duringearly germination and seedling development in wheat. Curr. Genet.46: 374–380.
Liere, K., Weihe, A., and Börner, T. (2011). The transcription ma-chineries of plant mitochondria and chloroplasts: Composition,function, and regulation. J. Plant Physiol. 168: 1345–1360.
Lobanov, A.V., Turanov, A.A., Hatfield, D.L., and Gladyshev, V.N.(2010). Dual functions of codons in the genetic code. Crit. Rev.Biochem. Mol. Biol. 45: 257–265.
Maayan, I., Shaya, F., Ratner, K., Mani, Y., Lavee, S., Avidan, B.,Shahak, Y., and Ostersetzer-Biran, O. (2008). Photosynthetic activityduring olive (Olea europaea) leaf development correlates with plastidbiogenesis and Rubisco levels. Physiol. Plant. 134: 547–558.
Malek, O., Brennicke, A., and Knoop, V. (1997). Evolution of trans-splicing plant mitochondrial introns in pre-Permian times. Proc.Natl. Acad. Sci. USA 94: 553–558.
Marchler-Bauer, A., et al. (2015). CDD: NCBI’s conserved domaindatabase. Nucleic Acids Res. 43: D222–D226.
Maréchal, A., and Brisson, N. (2010). Recombination and the mainte-nance of plant organelle genome stability. New Phytol. 186: 299–317.
Matsuura, M., Noah, J.W., and Lambowitz, A.M. (2001). Mechanism ofmaturase-promoted group II intron splicing. EMBO J. 20: 7259–7270.
Meinke, D.W. (1994). Seed development in Arabidopsis thaliana. In Ara-bidopsis, E.M. Meyerowitz and C.R. Somerville, eds (Cold Spring Har-bor, NY: Cold Spring Harbor Laboratory Press), pp. 253–295.
Meyer, E.H., Giegé, P., Gelhaye, E., Rayapuram, N., Ahuja, U.,Thöny-Meyer, L., Grienenberger, J.-M., and Bonnard, G. (2005).AtCCMH, an essential component of the c-type cytochrome mat-uration pathway in Arabidopsis mitochondria, interacts with apoc-ytochrome c. Proc. Natl. Acad. Sci. USA 102: 16113–16118.
Michel, F., and Ferat, J.L. (1995). Structure and activities of group IIintrons. Annu. Rev. Biochem. 64: 435–461.
Michel, F., Umesono, K., and Ozeki, H. (1989). Comparative andfunctional anatomy of group II catalytic introns–a review. Gene 82:5–30.
Mohr, G., and Lambowitz, A.M. (2003). Putative proteins related togroup II intron reverse transcriptase/maturases are encoded by nu-clear genes in higher plants. Nucleic Acids Res. 31: 647–652.
Mohr, G., Perlman, P.S., and Lambowitz, A.M. (1993). Evolutionaryrelationships among group II intron-encoded proteins and identifi-cation of a conserved domain that may be related to maturasefunction. Nucleic Acids Res. 21: 4991–4997.
Mower, J.P., Sloan, D.B., and Alverson, A.J. (2012). Plant mito-chondrial genome diversity: the genomics revolution. In Plant Ge-nome Diversity, Vol. 1: Plant Genomes, their Residents, and theirEvolutionary Dynamics. J.F. Wendel, J. Greilhuber, J. Dolezel, andI.J. Leitch, eds (Dordrecht, The Netherlands: Springer), pp. 123–144.
Nakagawa, N., and Sakurai, N. (2006). A mutation in At-nMat1a,which encodes a nuclear gene having high similarity to group IIintron maturase, causes impaired splicing of mitochondrial NAD4transcript and altered carbon metabolism in Arabidopsis thaliana.Plant Cell Physiol. 47: 772–783.
Nelson, C.J., Li, L., Jacoby, R.P., and Millar, A.H. (2013). Degra-dation rate of mitochondrial proteins in Arabidopsis thaliana cells.J. Proteome Res. 12: 3449–3459.
Nelson, C.J., Li, L., and Millar, A.H. (2014). Quantitative analysis ofprotein turnover in plants. Proteomics 14: 579–592.
Neuwirt, J., Takenaka, M., van der Merwe, J.A., and Brennicke, A.(2005). An in vitro RNA editing system from cauliflower mitochon-dria: editing site recognition parameters can vary in different plantspecies. RNA 11: 1563–1570.
Noah, J.W., and Lambowitz, A.M. (2003). Effects of maturase bindingand Mg2+ concentration on group II intron RNA folding investigatedby UV cross-linking. Biochemistry 42: 12466–12480.
Ostersetzer, O., Cooke, A.M., Watkins, K.P., and Barkan, A. (2005).CRS1, a chloroplast group II intron splicing factor, promotes intronfolding through specific interactions with two intron domains. PlantCell 17: 241–255.
Ostersetzer, O., Kato, Y., Adam, Z., and Sakamoto, W. (2007).Multiple intracellular locations of Lon protease in Arabidopsis: Evi-dence for the localization of AtLon4 to chloroplasts. Plant CellPhysiol. 48: 881–885.
Ostersetzer-Biran, O. (2016). Respiratory complex I and embryodevelopment. J. Exp. Bot. 67: 1205–1207.
Ostheimer, G.J., Williams-Carrier, R., Belcher, S., Osborne, E.,Gierke, J., and Barkan, A. (2003). Group II intron splicing factorsderived by diversification of an ancient RNA-binding domain. EMBOJ. 22: 3919–3929.
Pan, X., Chen, Z., Yang, X., and Liu, G. (2014). Arabidopsis voltage-dependent anion channel 1 (AtVDAC1) is required for female de-velopment and maintenance of mitochondrial functions related toenergy-transaction. PLoS One 9: e106941.
Park, S., Grewe, F., Zhu, A., Ruhlman, T.A., Sabir, J., Mower, J.P.,and Jansen, R.K. (2015). Dynamic evolution of Geranium mito-chondrial genomes through multiple horizontal and intracellulargene transfers. New Phytol. 208: 570–583.
Perrotta, A.T., and Been, M.D. (1991). A pseudoknot-like structurerequired for efficient self-cleavage of hepatitis delta virus RNA.Nature 350: 434–436.
Petersen, G., Cuenca, A., Møller, I.M., and Seberg, O. (2015).Massive gene loss in mistletoe (Viscum, Viscaceae) mitochondria.Sci. Rep. 5: 17588.
Piccirilli, J.A., and Staley, J.P. (2016). Reverse transcriptases lenda hand in splicing catalysis. Nat. Struct. Mol. Biol. 23: 507–509.
MatR Role in Mitochondria Group II Intron Splicing 2827

Qu, G., Kaushal, P.S., Wang, J., Shigematsu, H., Piazza, C.L.,Agrawal, R.K., Belfort, M., and Wang, H.W. (2016). Structure ofa group II intron in complex with its reverse transcriptase. Nat.Struct. Mol. Biol. 23: 549–557.
Saldanha, R., Mohr, G., Belfort, M., and Lambowitz, A.M. (1993).Group I and group II introns. FASEB J. 7: 15–24.
Saldanha, R., Chen, B., Wank, H., Matsuura, M., Edwards, J., andLambowitz, A.M. (1999). RNA and protein catalysis in group II in-tron splicing and mobility reactions using purified components.Biochemistry 38: 9069–9083.
Sanchez-Puerta, M.V., Cho, Y., Mower, J.P., Alverson, A.J., andPalmer, J.D. (2008). Frequent, phylogenetically local horizontaltransfer of the cox1 group I Intron in flowering plant mitochondria.Mol. Biol. Evol. 25: 1762–1777.
Schmitz-Linneweber, C., Williams-Carrier, R., and Barkan, A.(2005). RNA immunoprecipitation and microarray analysis showa chloroplast pentatricopeptide repeat protein to be associated withthe 59 region of mRNAs whose translation it activates. Plant Cell 17:2791–2804.
Schmitz-Linneweber, C., Lampe, M.-K., Sultan, L.D., andOstersetzer-Biran, O. (2015). Organellar maturases: A window in-to the evolution of the spliceosome. Biochim. Biophys. Acta 1847:798–808.
Schmitz-Linneweber, C., Williams-Carrier, R.E., Williams-Voelker,P.M., Kroeger, T.S., Vichas, A., and Barkan, A. (2006). A penta-tricopeptide repeat protein facilitates the trans-splicing of the maizechloroplast rps12 pre-mRNA. Plant Cell 18: 2650–2663.
Shingaki-Wells, R., Millar, A.H., Whelan, J., and Narsai, R. (2014).What happens to plant mitochondria under low oxygen? An omicsreview of the responses to low oxygen and reoxygenation. PlantCell Environ. 37: 2260–2277.
Siculella, L., Pacoda, D., Treglia, S., Gallerani, R., and Ceci, L.R.(1996). GTG as translation initiation codon in the apocytochromeb gene of sunflower mitochondria. DNA Seq. 6: 365–369.
Singh, R.N., Saldanha, R.J., D’Souza, L.M., and Lambowitz, A.M.(2002). Binding of a group II intron-encoded reverse transcriptase/maturase to its high affinity intron RNA binding site involves se-quence-specific recognition and autoregulates translation. J. Mol.Biol. 318: 287–303.
Skippington, E., Barkman, T.J., Rice, D.W., and Palmer, J.D. (2015).Miniaturized mitogenome of the parasitic plant Viscum scurruloi-deum is extremely divergent and dynamic and has lost all nadgenes. Proc. Natl. Acad. Sci. USA 112: E3515–E3524.
Sloan, D.B., Alverson, A.J., Chuckalovcak, J.P., Wu, M., McCauley,D.E., Palmer, J.D., and Taylor, D.R. (2012). Rapid evolution ofenormous, multichromosomal genomes in flowering plant mito-chondria with exceptionally high mutation rates. PLoS Biol. 10:e1001241.
Small, I. (2013). Mitochondrial genomes as living ‘fossils’. BMC Biol.11: 30.
Small, I.D., and Peeters, N. (2000). The PPR motif - a TPR-relatedmotif prevalent in plant organellar proteins. Trends Biochem. Sci.25: 46–47.
Small, I.D., Rackham, O., and Filipovska, A. (2013). Organelletranscriptomes: products of a deconstructed genome. Curr. Opin.Microbiol. 16: 652–658.
Stern, D.B., Goldschmidt-Clermont, M., and Hanson, M.R. (2010).Chloroplast RNA metabolism. Annu. Rev. Plant Biol. 61: 125–155.
Taylor, N.L., Heazlewood, J.L., and Millar, A.H. (2011). The Arabi-dopsis thaliana 2-D gel mitochondrial proteome: Refining the valueof reference maps for assessing protein abundance, contaminantsand post-translational modifications. Proteomics 11: 1720–1733.
Thomson, M.C., Macfarlane, J.L., Beagley, C.T., and Wolstenholme,D.R. (1994). RNA editing of mat-r transcripts in maize and soybeanincreases similarity of the encoded protein to fungal and bryophytegroup II intron maturases: evidence that mat-r encodes a functionalprotein. Nucleic Acids Res. 22: 5745–5752.
Toor, N., Hausner, G., and Zimmerly, S. (2001). Coevolution of groupII intron RNA structures with their intron-encoded reverse tran-scriptases. RNA 7: 1142–1152.
Townley-Tilson, W.H.D., Pendergrass, S.A., Marzluff, W.F., andWhitfield, M.L. (2006). Genome-wide analysis of mRNAs bound tothe histone stem-loop binding protein. RNA 12: 1853–1867.
Unseld, M., Marienfeld, J.R., Brandt, P., and Brennicke, A. (1997).The mitochondrial genome of Arabidopsis thaliana contains 57 genesin 366,924 nucleotides. Nat. Genet. 15: 57–61.
Uyttewaal, M., Mireau, H., Rurek, M., Hammani, K., Arnal, N.,Quadrado, M., and Giegé, P. (2008). PPR336 is associatedwith polysomes in plant mitochondria. J. Mol. Biol. 375: 626–636.
Val, R., Wyszko, E., Valentin, C., Szymanski, M., Cosset, A., Alioua,M., Dreher, T.W., Barciszewski, J., and Dietrich, A. (2011). Or-ganelle trafficking of chimeric ribozymes and genetic manipulationof mitochondria. Nucleic Acids Res. 39: 9262–9274.
Vaughn, J.C., Mason, M.T., Sper-Whitis, G.L., Kuhlman, P., andPalmer, J.D. (1995). Fungal origin by horizontal transfer of a plantmitochondrial group I intron in the chimeric CoxI gene of Pepero-mia. J. Mol. Evol. 41: 563–572.
Wahleithner, J.A., MacFarlane, J.L., and Wolstenholme, D.R.(1990). A sequence encoding a maturase-related protein in a group IIintron of a plant mitochondrial nad1 gene. Proc. Natl. Acad. Sci. USA87: 548–552.
Wank, H., SanFilippo, J., Singh, R.N., Matsuura, M., andLambowitz, A.M. (1999). A reverse transcriptase/maturase pro-motes splicing by binding at its own coding segment in a group IIintron RNA. Mol. Cell 4: 239–250.
Watkins, K.P., Kroeger, T.S., Cooke, A.M., Williams-Carrier, R.E.,Friso, G., Belcher, S.E., van Wijk, K.J., and Barkan, A. (2007). Aribonuclease III domain protein functions in group II intron splicingin maize chloroplasts. Plant Cell 19: 2606–2623.
Wicke, S., Müller, K.F., de Pamphilis, C.W., Quandt, D., Wickett,N.J., Zhang, Y., Renner, S.S., and Schneeweiss, G.M. (2013).Mechanisms of functional and physical genome reduction in pho-tosynthetic and nonphotosynthetic parasitic plants of the broom-rape family. Plant Cell 25: 3711–3725.
Wolfe, K.H., Morden, C.W., and Palmer, J.D. (1992). Function andevolution of a minimal plastid genome from a nonphotosyntheticparasitic plant. Proc. Natl. Acad. Sci. USA 89: 10648–10652.
Wurdack, K.J., and Davis, C.C. (2009). Malpighiales phylogenetics:Gaining ground on one of the most recalcitrant clades in the an-giosperm tree of life. Am. J. Bot. 96: 1551–1570.
Yan, C., Hang, J., Wan, R., Huang, M., Wong, C.C., and Shi, Y.(2015). Structure of a yeast spliceosome at 3.6-angstrom resolution.Science 349: 1182–1191.
Zhao, C., and Pyle, A.M. (2016). Crystal structures of a group II intronmaturase reveal a missing link in spliceosome evolution. Nat.Struct. Mol. Biol. 23: 558–565.
Zhu, A., Guo, W., Jain, K., and Mower, J.P. (2014). Unprecedentedheterogeneity in the synonymous substitution rate within a plantgenome. Mol. Biol. Evol. 31: 1228–1236.
Zimmerly, S., and Semper, C. (2015). Evolution of group II introns.Mob. DNA 6: 7.
Zimmerly, S., Hausner, G., and Wu Xc. (2001). Phylogenetic rela-tionships among group II intron ORFs. Nucleic Acids Res. 29: 1238–1250.
2828 The Plant Cell

Zmudjak, M., Colas des Francs-Small, C., Keren, I., Shaya, F.,Belausov, E., Small, I., and Ostersetzer-Biran, O. (2013). mCSF1,a nucleus-encoded CRM protein required for the processing ofmany mitochondrial introns, is involved in the biogenesis of re-spiratory complexes I and IV in Arabidopsis. New Phytol. 199: 379–394.
Zoschke, R., Ostersetzer-Biran, O., Börner, T., and Schmitz-Linneweber, C. (2009). Analysis of the regulation of MatK geneexpression. Endocytobiosis Cell Res. 19: 127–135.
Zoschke, R., Nakamura, M., Liere, K., Sugiura, M., Börner, T., andSchmitz-Linneweber, C. (2010). An organellar maturase asso-ciates with multiple group II introns. Proc. Natl. Acad. Sci. USA 107:3245–3250.
Zuker, M. (2003). Mfold web server for nucleic acid folding and hy-bridization prediction. Nucleic Acids Res. 31: 3406–3415.
Zuo, J., Niu, Q.-W., and Chua, N.-H. (2000). Technical advance: An es-trogen receptor-based transactivator XVE mediates highly induciblegene expression in transgenic plants. Plant J. 24: 265–273.
MatR Role in Mitochondria Group II Intron Splicing 2829

DOI 10.1105/tpc.16.00398; originally published online October 19, 2016; 2016;28;2805-2829Plant Cell
André Dietrich and Oren Ostersetzer-BiranAdnan Khan Niazi, Ido Keren, Sofia Shevtsov, Liron Klipcan, Jan Barciszewski, Jeffrey P. Mower,
Laure D. Sultan, Daria Mileshina, Felix Grewe, Katarzyna Rolle, Sivan Abudraham, Pawel Glodowicz,Group II Introns in Brassicaceae Mitochondria
The Reverse Transcriptase/RNA Maturase Protein MatR Is Required for the Splicing of Various
This information is current as of March 27, 2019
Supplemental Data /content/suppl/2016/10/19/tpc.16.00398.DC1.html
References /content/28/11/2805.full.html#ref-list-1
This article cites 130 articles, 34 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists