
Archaeal virus with exceptional virion architecture and the largest

1
The mature virion of ectromelia virus, a pathogenic 1
poxvirus, is capable of intrahepatic spread and can 2
serve as a target for delayed therapy 3
Xueying Ma1,5, Ren-Huan Xu1,5, Felicia Roscoe1, J Charles Whitbeck2, Roselyn 4
J. Eisenberg3, Gary H. Cohen2 and Luis J. Sigal1,4 5
1Immune Cell Development and Host Defense Program, Fox Chase Cancer 6
Center, Philadelphia, PA 19111. 7
2Department of Microbiology, School of Dental Medicine and 3Laboratories of 8
Microbiology and Immunology, School of Veterinary Medicine, University of 9
Pennsylvania, Philadelphia, PA 10
Short title: The mature virion in spread and therapy of an OPV 11
4Correspondence to: Dr. Luis J. Sigal, email: [email protected], Phone: 215-12
728-7061, FAX: 215-728-2409 13
5XM and RX contributed equally to this work 14
15
16
Copyright © 2013, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.03158-12 JVI Accepts, published online ahead of print on 17 April 2013
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

2
17
Abstract 18
Orthopoxviruses (OPV) which include the agent of smallpox variola virus, the 19
zoonotic monkeypox virus, the vaccine and zoonotic species vaccinia virus and the 20
mouse pathogen ectromelia virus (ECTV), form two types of infectious viral particles. 21
The mature virus (MV) which is cytosolic and the enveloped virus (EV) which is 22
extracellular. It is believed that MVs are required for viral entry into the host while EVs 23
are responsible for spread within the host. Following footpad infection of susceptible 24
mice, ECTV spreads lympho-hematogenously entering the liver 3-4 days post-infection 25
(dpi). Afterwards, ECTV spreads intra-hepatically killing the host. We found that 26
antibodies to an MV protein were highly effective at curing mice from ECTV infection 27
when administered after the virus reached the liver. Moreover, a mutant ECTV that does 28
not make EV was able to spread intra-hepatically and kill immunodeficient mice. 29
Together, these findings indicate that MVs are sufficient for the spread of ECTV within 30
the liver and could have implications regarding the pathogenesis of other OPVs, the 31
treatment of emerging OPV infections, as well as for strategies of preparedness in case 32
of accidental or intentional release of pathogenic OPVs. 33
34
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

3
Introduction 35
Orthopoxviruses (OPVs) penetrate their natural hosts through epithelial surfaces 36
and disseminate stepwise to distant organs through the regional draining lymph node (D-37
LN) and then the blood to cause systemic disease (21, 35). For instance, the human 38
pathogen variola virus (VARV) penetrated through the respiratory epithelium to spread 39
lympho-hematogenously through the mediastinal lymph nodes. Thus, smallpox was 40
chiefly a systemic and not a respiratory disease (10, 19). Similarly, some of the gravest 41
complications of the smallpox vaccine, which is made with live vaccinia virus (VACV) are 42
due to lympho-hematogenous (LH) dissemination (7, 8, 12). The OPV ectromelia virus 43
(ECTV), the agent of mousepox, is a mouse pathogen that serves as an excellent model 44
for OPV pathogenesis and as the textbook paradigm for LH spread (21, 35). ECTV 45
penetrates through the skin of the footpad and spreads lympho-hematogenously through 46
the popliteal D-LN to seed the liver and spleen. Susceptible strains of mice such as 47
BALB/c usually die 7-12 days post infection (dpi) with extensive liver and splenic 48
necrosis due to massive viral replication. In resistant strains of mice such as C57BL/6 49
(B6), LH dissemination and viral replication are considerably controlled by the action of 50
the innate and adaptive immune responses and mousepox does not occur (13, 18, 37). 51
During the replication of VACV and likely all other OPVs, the first infectious 52
particle formed is the intracellular mature virus (MV) which consists of a core surrounded 53
by a single membrane bi-layer. While most MVs remain within the cytosol and only 54
released to the extracellular milieu by cell lysis, some MVs become wrapped by a double 55
membrane, transported to the plasma membrane through microtubules and exocytosed, 56
losing the outer membrane in the process. Most of the resulting enveloped virus (EV) 57
remains attached to the plasma membrane as cell-associated enveloped virus (CEV) 58
while some are released as extracellular enveloped virus (EEV). CEVs are important for 59
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

4
cell-to-cell spread and EEV for long-range spread of VACV and probably other OPVs in 60
tissue culture. In addition, EV are thought to be essential for OPV spread within the host 61
(5, 6, 20, 31, 33). The MV and EV membranes each have a characteristic set of proteins 62
that play various roles in the virus life cycle and some of them have been shown to be 63
effective targets for vaccination in several OPV infection models (9, 14, 22-28, 33, 39). 64
However, it remains to be determined which of these proteins can serve as targets for 65
late therapy in a systemic model of OPV infection. 66
While prophylactic immunization with VACV is highly effective, treatment of 67
individuals exposed to pathogenic OPVs or with vaccine complications is less advanced. 68
In the US, vaccinia immunoglobulin (VIG) obtained from vaccinees is the only anti-OPV 69
treatment approved by the Food and Drug Administration (38, 40). However, VIG has 70
limited efficacy and, due to its nature, scarce. Still, it is not yet possible to supplant it with 71
or improve it with a cocktail of monoclonal antibodies (mAbs) because it is unknown 72
which specificities can protect and/or cure OPV infections (40). Of note, VIG can cure 73
ECTV infection when given to immunocompetent mice at 3 dpi but cannot cure severe 74
combined immunodeficient (SCID) mice from VACV infection (30). Here we demonstrate 75
that IgG1 mouse mAbs recognizing the MV protein L1R/EVM072 (VACV/ECTV) and the 76
EV protein A33R/EVM135 but not the EV protein B5R/EVM155 are effective at 77
preventing mousepox when administered immediately after infection. Of interest, the 78
L1R/EVM072 mAb as well as L1R/EVM072 polyclonal rabbit antiserum were also very 79
effective at preventing spread within the liver and curing ECTV infection when 80
administered after the virus reached the liver. Moreover, we show that an ECTV mutant 81
lacking a gene essential for EV formation (F13L/EVM036) can efficiently spread 82
intrahepatically. 83
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

5
Materials and Methods 84
Ethics Statement 85
All animal work has been conducted according to relevant national and 86
international guidelines and with protocols approved by The Fox Chase Cancer Center 87
Institutional Animal Care and Use Committee. 88
Cells, viruses and recombinant proteins 89
Media and cells were as previously described (16, 17, 41). Stocks of ECTV 90
Moscow strain (ATCC VR-1374) were propagated in tissue culture as previously 91
described (41). ECTV deficient in EVM036 has been described (32). Production of 92
recombinant A33R, EVM135, B5R and EVM155 was as previously described (2, 22). 93
Production of polyclonal rabbit Abs was also as described (43). mAbs VMC-2, VMC-14 94
and VMC-78 (al IgG1) have been described previously (1, 2) and were either obtained 95
from BEI Resources (Manassas, VA) or produced and purified as described previously 96
(1, 2). 97
Mice and infections 98
BALB/c and C57BL/6 mice were purchased from Taconic Farms. Severe 99
combined immunodeficient (SCID) mice in a BALB/c background were bred at FCCC. 100
B6.129P2-Fcer1gtm1Rav N12 (Taconic Farms) were bred at FCCC with mousepox 101
susceptible B6.D2-(D6Mit149-D6Mit15)/LusJ (B6.D2-D6, Jackson) to generate B6.D2-102
D6-Fcer1gtm1Rav. Unless indicated, mice were infected with ECTV in the left footpad with 103
30 μl PBS containing 3×102 pfu. For the determination of survival, the mice were 104
monitored daily. To avoid unnecessary suffering, mice were euthanized and counted as 105
dead if imminent death was certain. For virus titers and histopathology, mice were 106
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

6
infected with pfu PFU ECTV and euthanized when indicated and whole LNs or 100 mg 107
of liver were homogenized in PBS using a Tissue Lyser homogenizer (Qiagen). Virus 108
titers were determined on BS-C-1 cells in 6-well-plates as before (16, 17, 41). 109
Construction of recombinant baculovirus expressing soluble VACV and ECTV 110
proteins 111
The DNA sequence of the different proteins was amplified by PCR without a 112
transmembrane domain and cloned into the baculovirus transfer vector, pVT-Bac 113
downstream of, and in frame with, the mellitin signal sequence as previously described 114
for B5R (2). Two additional amino acid residues (DP) are present at the N-terminus of 115
the mature (signal-less) recombinant proteins. These are left over following cleavage of 116
the melittin signal sequence. The proteins were constructed with 6 histidine residues at 117
the C-terminus to allow for purification via nickel-NTA affinity chromatography. 118
Hybridoma selection and IgG purification 119
Murine hybridomas secreting antibodies against A33R were generated as 120
previously described (1). 121
ELISA for viral proteins 122
High-binding 96-well ELISA plates (Corning) were coated overnight at 4°C with 123
50μl recombinant A33R, EVM135, B5R or EVM155 protein (50μg/ml) or, for L1R and 124
EVM072, with cell lysates from VACV or ECTV infected cells respectively (2 ×107 pfu/ml) 125
in Phosphate-Buffered Saline (PBS), pH 7.0. Plates were washed twice with PBS, and 126
then blocked for 2 h at 37°C with PBS containing 0.05% Tween-20 (PBST) and 3% BSA. 127
Antibodies were serially diluted in PBST, 1% BSA, and 0.1 ml was added to each well. 128
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

7
The plates were then incubated for 1 h at 37°C, washed four times with PBST and 0.1 ml 129
of horseradish peroxidase (HRP)-conjugated affinity purified goat anti-mouse IgGγ (KPL) 130
was added to each well at a dilution of 1:3,000 in PBST. The plates were incubated for 1 131
h at 37 °C, washed six times with PBST and 100μl Sure Blue TMB (KPL) was added to 132
each well. The plates were incubated at room temperature for 5–20 min. The reactions 133
were stopped by addition of 20μl 3M HCl. The OD was determined at 450 nm using a 134
microplate spectrophotometer (µQuant, Bio-Tek). 135
Plaque reduction assay 136
ECTV stocks were incubated for 1 h at RT with 100μg/ml of the indicating Abs. 137
The virus-antibody mixture (100 pfu/well) were added to confluent BS-C-1 cells (ATCC # 138
CL-26) in 24-well plates with 0.25 ml and the plates were incubated for 2 hours at 37°C. 139
Viral inoculums were removed after incubation, and cells were overlaid with 1ml fresh 140
DMEM media containing 2.5% FBS and 1% CMC. Cells were incubated for 5-7 days at 141
37°C in a 5% CO2 incubator. The cells were fixed with formaldehyde, and stained with 142
crystal violet. 143
Comet inhibition assay 144
Monolayers of BSC-1 cells in 6 well plates were infected with 60 pfu ECTV in 0.5 145
ml DMEM containing 2.5% FBS. After 2 h incubation at 37 °C, the media containing virus 146
was aspirated and fresh 2 ml DMEM containing 2.5% FBS and 50μg/ml of the 147
corresponding antibody was added. Cells were incubated for 5-7 days at 37 °C in a 5% 148
CO2 incubator and stained with crystal violet. 149
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

8
Complement depletion 150
BALB/c mice were inoculated ip with 15 μg purified CVF Cobra Venom Factor 151
(CVF) from najia najia kaouthia (CompTech) at 4, 5 and 8 dpi. 10 µg CVF have been 152
shown to fully inhibit complement activity in mouse sera (11). Mice were bled at 5, 6 and 153
9 dpi and C3 depletion was assessed in serum by WB and using a commercial ELISA kit 154
for C3 (Immunology Consultants Lab) used according to manufacturer’s instructions. 155
RNA isolation 156
D-LNs were collected from infected or naïve mice and immediately placed in 157
RNAse-free tubes containing RNAlater (Ambion). RNA was extracted using RNAeasy kit 158
(Qiagen) as described and the DNA was digested during the process with DNAse 159
(Qiagen). 1 μl of the RNA was analyzed in a nanodrop 2000C (Thermo Scientific). 160
Reverse transcription 161
1 μg of RNA was retrotranscribed to cDNA using the High Capacity cDNA 162
Reverse Transcription kit (Applied Biosystems) according to manufacturer’s instructions. 163
Quantitative PCR (qPCR) 164
We used the Roche Universal library probe #7 and EVM166-specific 165
oligonucleotides gtgcaaagtgtccgcctatt and tctattaagaggtcgtctagtctttcc as indicated by the 166
manufacturer. Briefly, 1 μl of the cDNA from the reverse transcription reactions was used 167
as template. The PCR reactions were performed in a MX3005P (Agilent Technologies), 168
or a Mastercycler ep realplex2 (Eppendorf). The expression was normalized by GAPDH 169
expression and quantified using a standard curve generated with a plasmid containing 170
the EVM166 gene. 171
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

9
Immunohistochemistry 172
Viral foci were detected using EVM135 rabbit antisera as described previously 173
(43). 174
Statistics 175
We used Prism software to determine significance of the differences between 176
groups. For survival experiments each group consisted of five mice. We determined 177
significant differences using the Log-rank test. For dot plots, each point represents an 178
individual mouse and differences were determined using the Mann-Whitney test or a 179
two-tailed unpaired t test as applicable. In all experiments P ≤ 0.05 = *, P ≤ 0.01=**, P ≤ 180
0.001=***, P ≤ 0.0001=****. All experiments were repeated a minimum of two but in most 181
cases three times. 182
183
184
Results 185
L1R A33R and B5R monoclonal antibodies (mAbs) recognize the ECTV ortholog 186
proteins and block their biological function 187
Previous work in several laboratories including ours has shown that immunization 188
against the EV proteins A33R/EVM135 and/or B5R as well as the MV protein L1R alone 189
or in combination can protect mice against intranasal VACV and/or ECTV and primates 190
against MPXV infection (9, 14, 22-28, 39) . Additional work showed that polyclonal Abs 191
(pAbs) or mAbs to these proteins alone or in combination can protect from intranasal 192
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

10
VACV before or soon after infection (3, 11, 29). However, whether Ab treatment can 193
cure natural OPV infections after becoming systemic has not been explored. Given that 194
OPVs are antigenically similar, we tested a panel of VACV L1R, A33R and B5R mAbs 195
for reactivity to their respective ECTV orthologs EVM072, EVM135 and EVM155. The 196
L1R mAb VMC-2 reacted similarly in ELISA assays with plate bound VACV and ECTV 197
particles suggesting identical binding to L1R and EVM072 (Figure 1a). Plate-bound 198
recombinant A33R and EVM135 were recognized similarly by the A33R mAb VMC-78 199
(Figure 1b), and plate-bound recombinant B5R and EVM072 were also similarly 200
recognized by the B5R mAb VMC-14 (Figure 1c). 201
We have previously shown that VMC-2 (anti-L1R/EVM072) neutralized VACV 202
stocks which contain mostly MV (2). We now found that, as compared with no Ab VMC-2 203
also neutralized ECTV stocks in plaque reduction assays while the anti-A33R/EVM135 204
VMC-78 or the anti-B5R/EVM155 VMC-14 did not. Control polyclonal rabbit anti-L1R 205
(rL1R) also neutralized ECTV while rabbit EVM155 antisera (rEVM155) did not (Figure 206
1d). As expected, the A33R/EVM135 VMC-78 mAb and the B5R/EVM155 VMC14 mAb 207
inhibited comet formation in liquid media (Figure 1e top), a sign of distant EEV-208
dependent spread. Notably, not only control rEVM155 but also VMC-2 and rL1R 209
inhibited comet formation (Figure 1 e, bottom). In addition, VMC-14 partially neutralized 210
EV but VMC-78 or VMC-2 did not (reported as text). That anti-L1R Abs can inhibit 211
ECTV comet formation is somewhat surprising as it does not inhibit VACV comets. This 212
suggests differences in the spread of VACV and ECTV in tissue culture . The reasons 213
for this difference are unknown and grant future comparative studies. Nevertheless, from 214
these experiments we concluded that in addition to recognizing their respective VACV 215
targets; VMC-2, VMC-78 and VMC-14 also recognize and inhibit the biological function 216
of the respective ECTV orthologs EVM072, EVM135 and EVM155. 217
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

11
Prophylactic treatment with mAbs to L1R/EVM072 and A33R/EVM135 but not 218
B5R/EVM155 protect mice from mousepox 219
We tested whether VMC-2, VMC-78 and VMC-14 could be used to prevent 220
mousepox. BALB/c mice were infected with 300 plaque forming units (pfu) ECTV in the 221
footpad, its natural route (18), and a few minutes later inoculated with 200 μg of the 222
different mAbs intraperitoneally (ip). We found that 200 µg VMC-2 and VMC-78 but not 223
VMC-14 significantly protected BALB/c mice from lethal mousepox (P=**, Figure 2a). 224
Protection with VMC-2 was more effective because the mice treated with this mAb did 225
not lose weight while those inoculated with VMC-78 did (Figure 2b). 226
Treatment with anti-L1R cures mice from mousepox when administered after 227
ECTV dissemination to the liver 228
We next determined when after infection ECTV becomes clearly established in 229
the liver of BALB/c mice infected with 300 pfu in the footpad. Immunohistochemical 230
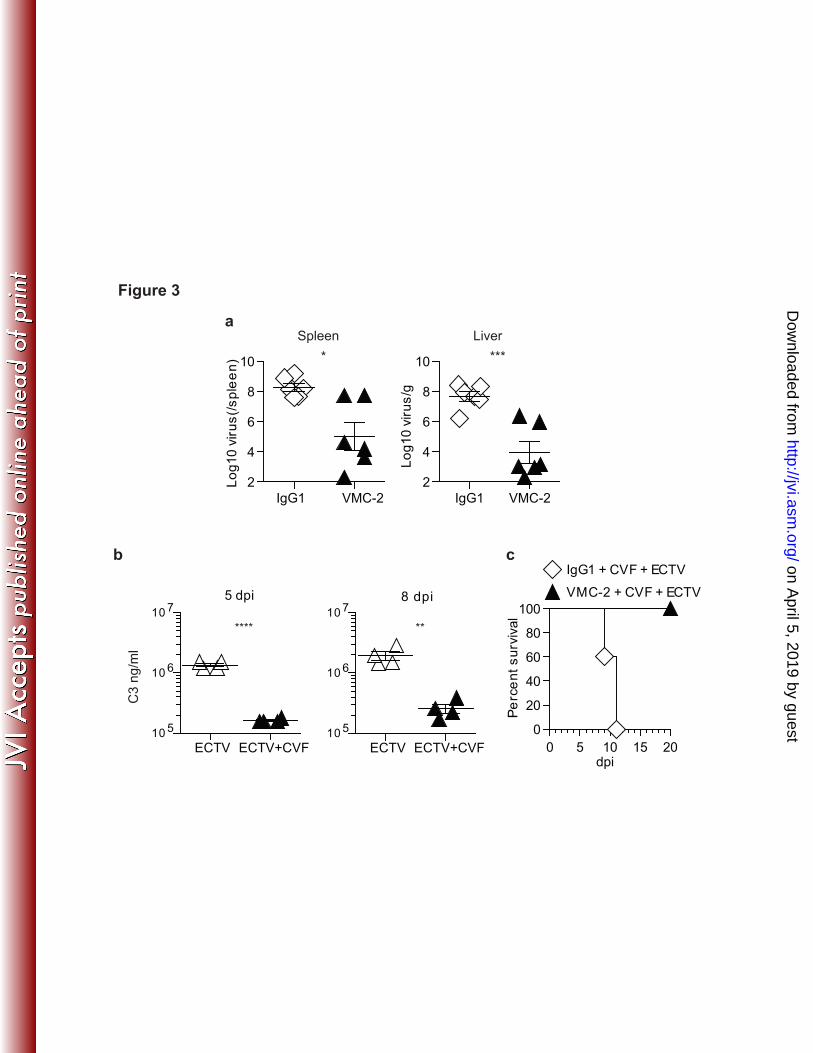
analysis of liver sections showed few infection foci in the liver at 4 dpi and most were 231
comprised of a single cell. At 5 dpi, most foci were multicellular. During the following 232
days, the size of the individual foci gradually increased and finally coalesced to cover 233
most of the liver at 8-10 dpi. The increase in foci size was reminiscent of the growth of 234
viral plaques in semi-solid media and suggested that ECTV spread centrifugally to 235
nearby cells (Figure 2c). From these results we concluded that that, following infection 236
with 300 pfu in the footpad, ECTV is well established in the liver at 4-5 dpi. Thus, we 237
tested whether the different mAbs could cure mousepox when administered at these dpi. 238
BALB/c mice were treated with VMC-2, VMC-78 or mouse IgG1 at 4 or 5 dpi with 300 239
pfu in the footpad. All the mice treated with 200 µg VMC-2 at 4 or 5 dpi survived while all 240
those treated with 500 µg IgG1 succumbed. Mice treated with either 200 or 500 µg 241
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

12
VMC-78 at 4 or 5 dpi were also significantly protected but in each group some mice 242
succumbed. Thus, together, VMC-78 was significantly less protective than VMC-2 (P=*, 243
Figure 2d). When organs from groups of mice treated with 200 µg mAbs at 5 dpi were 244
analyzed at 7 dpi (2 days after treatment) virus loads in the liver and spleen were 245
significantly lower in VMC-2 as compared to VMC-78 or IgG1 treated mice (Figure 2e). 246
Thus, treatment with anti-L1R/EVM072 mAb, was more effective at curing late 247
mousepox than treatment with anti-A33R/EVM135 mAb. In addition, 248
immunohistochemical analysis of the livers showed that the lesions in VMC-2 treated 249
mice at 7 dpi (2 days post-treatment) were fewer and smaller as compared with VMC-78 250
or IgG1 treated mice (Figure 2f) suggesting less efficient intrahepatic ECTV spread in 251
mice treated with VMC-2 than in mice treated with VMC-78 or IgG1. 252
To investigate whether the differential effects we were seeing were due to the 253
monoclonality of the Abs, we used rabbit antisera against VACV L1R (rL1R), ECTV 254
EVM135 (rEVM135) and EVM155 (rEVM155). Each antiserum reacted with its 255
respective target in ELISA. L1R antiserum was highly effective at reducing plaque and 256
comet formation while EVM-135 and EVM-155 antisera did not reduce plaques but 257
inhibited comets (Figure 1 and data not shown). When given at 0 dpi (Figure 2g, left) 258
L1R and EVM-135 but not EVM-155 antisera significantly protected BALB/c mice from 259
death (P=**). The protection afforded by L1R and EVM-135 antisera were not 260
significantly different. EVM-155 antiserum had significant protection but mostly by 261
delaying the time of death. At 2 dpi (Figure 2g, center), L1R and EVM135 but not 262
EVM155 antisera were protective. When given at 5 dpi, only the L1R antiserum was 263
protective (P=*, Figure 2g, right). In all cases, the positive control antiserum to the T1-264
IFNbp was protective (43) while naïve serum was not. Hence, the data with pAb confirm 265
that the MV protein EVM-072 is a better target for late Ab therapy than the EV proteins 266
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

13
EVM-135 and EVM-155. Moreover, because rabbit Abs can activate mouse complement 267
in vivo to protect from VACV infection (11), these data suggests that the less effective 268
protection of the anti-EV mAbs was not due only to the lack of effector functions of the 269
IgG1 isotype. 270
Late control of ECTV by anti-L1R/EVM072 does not require antibody effector 271
functions 272
While unlikely because it is IgG1, an isotype with poor effector functions, it was 273
possible that mAb VMC-2 protected mice through antibody effector functions such as Fc-274
mediated antibody dependent cytotoxicity (ADCC) or Fc-dependent or -independent 275
complement (C’) activation. To test whether VMC-2 controlled ECTV through a 276
mechanism dependent on Fc receptors, B6.129P2-Fcer1gtm1Rav N12 (Fcer1γ-/-) which are 277
deficient in Fc receptor expression and signaling (34) were backcrossed to B6.D2-278
(D6Mit149-D6Mit15)/LusJ (B6.D2-D6) which are a C57BL/6 congenic strain susceptible 279
to mousepox (15). For unknown reasons, not all B6.D2-D6- Fcer1γ-/- succumbed to 280
mousepox suggesting that they are not as susceptible as the B6.D2-D6 parental strain 281
(reported as text). Still, B6.D2-D6- Fcer1γ -/- mice treated with VMC-2 at 5 dpi had 282
significantly lower ECTV loads in the liver and spleen (Figure 3a) as compared to those 283
treated with IgG1 indicating that VMC-2 reduced virus loads late in infection 284
independently of Fc receptors. To test whether C’ activation was required, ECTV 285
infected BALB/c mice were depleted of the C3 fraction of C’ with cobra venom factor 286
(CVF) administered three times (4, 11) . This treatment eliminated most C3 as 287
determined by ELISA (Figure 3b) and a similar schedule with a 33% lower dose has 288
been shown to fully inhibit C’ activity in mouse serum (11). All C3 depleted mice treated 289
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

14
with VMC-2 survived mousepox while all control mice depleted of C3 but treated with 290
IgG1 succumbed (Figure 3c). 291
ECTV deficient in EV spreads in the liver and is lethal to severe combined 292
immunodeficient (SCID) mice. 293
The data above suggested that following LH spread, ECTV dissemination within 294
the liver is more dependent on MV than on EV. It has previously been shown that VACV 295
deficient in F13L is unable to make EV becoming manifest by its formation of small 296
plaques in tissue culture. It has also been demonstrated that VACV deficient in F13L is 297
highly attenuated (36). Very recently, we reported the generation of ECTV deficient in 298
EVM036 (ECTV-Δ036), the ortholog of VACV F13L. Similar to its VACV counterpart, 299
ECTV-Δ036 has a very small plaque phenotype in tissue culture indicating that EVs are 300
very important for ECTV spread in cultured cells. Furthermore, ECTV-Δ036 is non-301
pathogenic in immunocompetent BALB/c mice even at high doses (32). However, ECTV-302
Δ036 was lethal to SCID mice infected in the footpad (P=**, Figure 4a), but the time of 303
death was highly variable. This suggested that in the absence of adaptive immunity, an 304
OPV deficient in EV can still disseminate lympho-hematogenously albeit inefficiently. To 305
determine whether ECTV-Δ036 kills immunodeficient mice by replicating in the liver, we 306
infected SCID mice ip with a high dose of ECTV-Δ036 which permitted the rapid and 307
synchronized seeding of the liver. Under these conditions, ECTV-Δ036 was rapidly lethal 308
(P=**, Figure 4b) and transcripts of an ECTV gene (EVM166) in the liver and spleen 309
increased 104 fold from day 2 to day 6 suggesting rapid replication in these organs 310
(Figure 4c). Furthermore, immunohistochemical staining of the liver with Abs to EVM135 311
showed few infected cells at 2 dpi but massive infection at 8 dpi (Figure 4d) 312
demonstrating efficient EV-independent intrahepatic ECTV spread. 313
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

15
Discussion 314
Given the threat of intentional release of variola virus, the zoonotic potential of 315
other OPVs, and the possible complications of the live smallpox vaccine, OPVs still 316
constitute a risk to human health. Hence, development of a cocktail of mAbs with the 317
potential of curing disseminated OPV disease and replacing VIG is of interest (40). Our 318
work here demonstrates that as measured by survival, viral loads and liver damage, 319
neutralizing Abs to the MV protein L1R/EVM072 are more effective than comet-inhibiting 320
Abs to the EV proteins A33R/EVM135 and B5R/EVM155 at controlling an otherwise 321
lethal ECTV when given after viral dissemination to susceptible but otherwise 322
immunocompetent mice. We have recently shown that blocking IgG1 mAbs to 323
B18R/EVM166, which encode a secreted, non-structural Type I interferon decoy 324
receptor, also cures mousepox when administered after viral dissemination (43). While 325
mAb VMC-78 to the EV protein A33R/EVM135 promoted survival when given at 5 dpi, it 326
did not significantly decrease virus loads or liver pathology 2 days after treatment. 327
Furthermore, mAb VMC-14 to the EV protein B5R/EVM155 Abs was not protective even 328
when given at the time of infection. These results were surprising because both proteins 329
have been shown to be good targets for vaccines in various ECTV and/or VACV models 330
(9, 11, 14, 22-29, 39). With the cautionary note that our experiment with ECTV do not 331
necessarily extend to every OPV, our results suggest that mAbs to L1R/EVM072 and to 332
B18R/EVM166 are both excellent candidates to be included in mAb cocktails for the late 333
treatment of OPV diseases, the former by controlling virus spread within tissues and the 334
latter by restoring Type I interferon signaling. The results also suggest that neutralizing 335
Abs to other MV proteins and blocking Abs to other secreted virulence factors should be 336
explored as additional components for a VIG replacement. 337
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

16
It has been recently shown that a major mechanism whereby Abs to EV protect 338
from VACV is through the activation of complement (3, 4, 11), and all our mAbs are 339
IgG1, which are known to lack effector functions. Thus, it is possible that the deficient 340
protection by VMC-14 and VMC-78 that we observed is due to their isotype and this 341
could be explored using IgG2 mAbs. However, this doesn’t seem to be the only reason 342
because rabbit anti-L1R was also more potent and rabbit Abs have been shown capable 343
of using complement to help protect from VACV (11). It would also be of interest to test 344
these Abs in the recovery from infections with OPVs other than ECTV and VACV. In 345
addition, it is possible that combinations of specific EV, MV and virulence factor mAbs of 346
various isotypes will be even more effective at curing late mousepox than single Ab 347
therapy or VIG and this could be tested. 348
It is also important to note that we do not think that the L1R Abs can clear ECTV 349
singlehandedly. Rather, we think that passive immunization temporarily reduces virus 350
loads allowing for the development of active immunity. This is supported by our previous 351
reports that passive transfer of Abs or memory T cells protect only immunocompetent 352
hosts from mousepox (42, 43) and a report by Lustig et al showing that VIG does not 353
permanently protect SCID mice from VACV challenge (30). 354
Work with the prototypic OPV VACV established the current model that MVs are 355
important for initial OPV infection while EVs are essential for their spread within the host 356
(31, 33). Our results indicate that, at least for ECTV, this model must be revised. While 357
the possibility that some EVs are produced in the absence of EVM036 cannot be 358
discarded, our finding that ECTV-Δ036 eventually kills SCID mice when inoculated into 359
the footpad strongly suggests that EVs are very important albeit not absolutely essential 360
for LH spread. Unexpectedly, we also showed that L1R/EVM072 mAb protects from 361
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

17
lethal mousepox by curtailing intrahepatic spread and that ECTV deficient in EVM036 362
rapidly disseminates within the liver of immunodeficient mice. Thus, our experiments 363
suggest a model where EVs are important in the initial LH spread while MVs are key for 364
ECTV intrahepatic spread. While the liver is not thought to be a target for VARV or other 365
human OPV infections, our work suggests the chance that other OPVs preferentially use 366
MV to spread within their target organs. The development of effective and reliable anti-367
OPV therapies for late exposure requires testing for this previously unsuspected 368
possibility. 369
Acknowledgements 370
We thank Fox Chase Cancer Center Laboratory Animal and Tissue Culture 371
Facilities for their services and Ms. Holly Gillin for assistance in the preparation of the 372
manuscript. This work was supported by NIAID grant U19AI083008 to LJS, NCI grant 373
P30CA006927 to FCCC and by a generous gift from the Kirby Foundation to the FCCC 374
Inflammation Group. 375
376
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

18
377
References 378
1. Aldaz-Carroll, L., J. C. Whitbeck, M. Ponce de Leon, H. Lou, L. Hirao, S. N. 379 Isaacs, B. Moss, R. J. Eisenberg, and G. H. Cohen. 2005. Epitope-mapping 380 studies define two major neutralization sites on the vaccinia virus extracellular 381 enveloped virus glycoprotein B5R. J Virol 79:6260-6271. 382
2. Aldaz-Carroll, L., J. C. Whitbeck, M. Ponce de Leon, H. Lou, L. K. Pannell, J. 383 Lebowitz, C. Fogg, C. L. White, B. Moss, G. H. Cohen, and R. J. Eisenberg. 384 2005. Physical and immunological characterization of a recombinant secreted 385 form of the membrane protein encoded by the vaccinia virus L1R gene. Virology 386 341:59-71. 387
3. Benhnia, M. R., M. M. McCausland, J. Laudenslager, S. W. Granger, S. Rickert, 388 L. Koriazova, T. Tahara, R. T. Kubo, S. Kato, and S. Crotty. 2009. Heavily 389 isotype-dependent protective activities of human antibodies against vaccinia 390 virus extracellular virion antigen B5. J Virol 83:12355-12367. 391
4. Benhnia, M. R., M. M. McCausland, J. Moyron, J. Laudenslager, S. Granger, S. 392 Rickert, L. Koriazova, R. Kubo, S. Kato, and S. Crotty. 2009. Vaccinia virus 393 extracellular enveloped virion neutralization in vitro and protection in vivo depend 394 on complement. J Virol 83:1201-1215. 395
5. Blasco, R., and B. Moss. 1991. Extracellular vaccinia virus formation and cell-to-396 cell virus transmission are prevented by deletion of the gene encoding the 397 37,000-Dalton outer envelope protein. J Virol 65:5910-5920. 398
6. Blasco, R., and B. Moss. 1992. Role of cell-associated enveloped vaccinia virus 399 in cell-to-cell spread. J Virol 66:4170-4179. 400
7. Bray, M. 2003. Pathogenesis and potential antiviral therapy of complications of 401 smallpox vaccination. Antiviral research 58:101-114. 402
8. Bray, M., and M. E. Wright. 2003. Progressive vaccinia. Clinical infectious 403 diseases : an official publication of the Infectious Diseases Society of America 404 36:766-774. 405
9. Buchman, G. W., M. E. Cohen, Y. Xiao, N. Richardson-Harman, P. Silvera, L. J. 406 DeTolla, H. L. Davis, R. J. Eisenberg, G. H. Cohen, and S. N. Isaacs. 2010. A 407 protein-based smallpox vaccine protects non-human primates from a lethal 408 monkeypox virus challenge. Vaccine 28:6627-6636. 409
10. Chapman, J. L., D. K. Nichols, M. J. Martinez, and J. W. Raymond. 2010. Animal 410 models of orthopoxvirus infection. Vet Pathol 47:852-870. 411
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

19
11. Cohen, M. E., Y. Xiao, R. J. Eisenberg, G. H. Cohen, and S. N. Isaacs. 2011. 412 Antibody against extracellular vaccinia virus (EV) protects mice through 413 complement and Fc receptors. PloS one 6:e20597. 414
12. Cono, J., C. G. Casey, D. M. Bell, C. Centers for Disease, and Prevention. 2003. 415 Smallpox vaccination and adverse reactions. Guidance for clinicians. MMWR. 416 Recommendations and reports : Morbidity and mortality weekly report. 417 Recommendations and reports / Centers for Disease Control 52:1-28. 418
13. Esteban, D. J., and R. M. Buller. 2005. Ectromelia virus: the causative agent of 419 mousepox. J Gen Virol 86:2645-2659. 420
14. Fang, M., H. Cheng, Z. Dai, Z. Bu, and L. J. Sigal. 2006. Immunization with a 421 single extracellular enveloped virus protein produced in bacteria provides partial 422 protection from a lethal orthopoxvirus infection in a natural host. Virology 423 345:231-243. 424
15. Fang, M., M. T. Orr, P. Spee, T. Egebjerg, L. L. Lanier, and L. J. Sigal. 2011. 425 CD94 is essential for NK cell-mediated resistance to a lethal viral disease. 426 Immunity 34:579-589. 427
16. Fang, M., and L. J. Sigal. 2005. Antibodies and CD8+ T Cells Are 428 Complementary and Essential for Natural Resistance to a Highly Lethal 429 Cytopathic Virus. J Immunol 175:6829-6836. 430
17. Fang, M., and L. J. Sigal. 2006. Direct CD28 Costimulation Is Required for CD8+ 431 T Cell-Mediated Resistance to an Acute Viral Disease in a Natural Host. J 432 Immunol 177:8027-8036. 433
18. Fenner, F. 1994. Mousepox (ectromelia), p. 412. In A. Osterhaus (ed.), Virus 434 infections of rodents and lagomorphs, vol. 5. Elsevier Science, Amsterdam ; New 435 York. 436
19. Fenner, F., D. A. Henderson, I. Arita, Z. Jezek, D. Ladnyi, and W. H. 437 Organization. 1988. Smallpox and its eradication. World Health Organization, 438 Geneva. 439
20. Fields, B. N., D. M. Knipe, and P. M. Howley. 2007. Fields' virology, 5th ed. 440 Wolters kluwer/Lippincott Williams & Wilkins, Philadelphia. 441
21. Flint, S. J., and American Society for Microbiology. 2009. Principles of virology, 442 3rd ed. ASM Press, Washington, DC. 443
22. Fogg, C., S. Lustig, J. C. Whitbeck, R. J. Eisenberg, G. H. Cohen, and B. Moss. 444 2004. Protective immunity to vaccinia virus induced by vaccination with multiple 445 recombinant outer membrane proteins of intracellular and extracellular virions. J 446 Virol 78:10230-10237. 447
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

20
23. Golden, J. W., and J. W. Hooper. 2011. The strategic use of novel smallpox 448 vaccines in the post-eradication world. Expert review of vaccines 10:1021-1035. 449
24. Heraud, J. M., Y. Edghill-Smith, V. Ayala, I. Kalisz, J. Parrino, V. S. 450 Kalyanaraman, J. Manischewitz, L. R. King, A. Hryniewicz, C. J. Trindade, M. 451 Hassett, W. P. Tsai, D. Venzon, A. Nalca, M. Vaccari, P. Silvera, M. Bray, B. S. 452 Graham, H. Golding, J. W. Hooper, and G. Franchini. 2006. Subunit recombinant 453 vaccine protects against monkeypox. J. Immunol. 177:2552-2564. 454
25. Hooper, J. W., D. M. Custer, C. S. Schmaljohn, and A. L. Schmaljohn. 2000. 455 DNA vaccination with vaccinia virus L1R and A33R genes protects mice against 456 a lethal poxvirus challenge. Virology 266:329-339. 457
26. Hooper, J. W., D. M. Custer, and E. Thompson. 2003. Four-gene-combination 458 DNA vaccine protects mice against a lethal vaccinia virus challenge and elicits 459 appropriate antibody responses in nonhuman primates. Virology 306:181-195. 460
27. Hooper, J. W., J. W. Golden, A. M. Ferro, and A. D. King. 2007. Smallpox DNA 461 vaccine delivered by novel skin electroporation device protects mice against 462 intranasal poxvirus challenge. Vaccine 25:1814-1823. 463
28. Hooper, J. W., E. Thompson, C. Wilhelmsen, M. Zimmerman, M. A. Ichou, S. E. 464 Steffen, C. S. Schmaljohn, A. L. Schmaljohn, and P. B. Jahrling. 2004. Smallpox 465 DNA Vaccine Protects Nonhuman Primates against Lethal Monkeypox. J Virol 466 78:4433-4443. 467
29. Lustig, S., C. Fogg, J. C. Whitbeck, R. J. Eisenberg, G. H. Cohen, and B. Moss. 468 2005. Combinations of polyclonal or monoclonal antibodies to proteins of the 469 outer membranes of the two infectious forms of vaccinia virus protect mice 470 against a lethal respiratory challenge. J Virol 79:13454-13462. 471
30. Lustig, S., G. Maik-Rachline, N. Paran, S. Melamed, T. Israely, N. Erez, N. Orr, 472 S. Reuveny, A. Ordentlich, O. Laub, A. Shafferman, and B. Velan. 2009. 473 Effective post-exposure protection against lethal orthopoxviruses infection by 474 vaccinia immune globulin involves induction of adaptive immune response. 475 Vaccine 27:1691-1699. 476
31. Payne, L. G. 1980. Significance of extracellular enveloped virus in the in vitro and 477 in vivo dissemination of vaccinia. J Gen Virol 50:89-100. 478
32. Roscoe, F., R. H. Xu, and L. J. Sigal. 2012. Characterization of Ectromelia Virus 479 Deficient in EVM036, the Homolog of Vaccinia virus F13L, and its Application for 480 the Rapid Generation of Recombinant Viruses. J Virol. 481
33. Smith, G. L., A. Vanderplasschen, and M. Law. 2002. The formation and function 482 of extracellular enveloped vaccinia virus. J Gen Virol 83:2915-2931. 483
34. Takai, T., M. Li, D. Sylvestre, R. Clynes, and J. V. Ravetch. 1994. FcR gamma 484 chain deletion results in pleiotrophic effector cell defects. Cell 76:519-529. 485
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

21
35. Virgin, H. W. 2007. Pathogenesis of viral infection, p. 335-336. In B. N. Fields, D. 486 M. Knipe, and P. M. Howley (ed.), Fields' virology, 5th ed, vol. 1. Wolters 487 kluwer/Lippincott Williams & Wilkins, Philadelphia. 488
36. Vliegen, I., G. Yang, D. Hruby, R. Jordan, and J. Neyts. 2012. Deletion of the 489 vaccinia virus F13L gene results in a highly attenuated virus that mounts a 490 protective immune response against subsequent vaccinia virus challenge. 491 Antiviral research 93:160-166. 492
37. Wallace, G. D., and R. M. Buller. 1985. Kinetics of ectromelia virus (mousepox) 493 transmission and clinical response in C57BL/6j, BALB/cByj and AKR/J inbred 494 mice. Laboratory animal science 35:41-46. 495
38. Wittek, R. 2006. Vaccinia immune globulin: current policies, preparedness, and 496 product safety and efficacy. International journal of infectious diseases : IJID : 497 official publication of the International Society for Infectious Diseases 10:193-498 201. 499
39. Xiao, Y., L. Aldaz-Carroll, A. M. Ortiz, J. C. Whitbeck, E. Alexander, H. Lou, H. L. 500 Davis, T. J. Braciale, R. J. Eisenberg, G. H. Cohen, and S. N. Isaacs. 2007. A 501 protein-based smallpox vaccine protects mice from vaccinia and ectromelia virus 502 challenges when given as a prime and single boost. Vaccine 25:1214-1224. 503
40. Xiao, Y., and S. N. Isaacs. 2010. Therapeutic Vaccines and Antibodies for 504 Treatment of Orthopoxvirus Infections. Viruses 2:2381-2403. 505
41. Xu, R. H., M. Cohen, Y. Tang, E. Lazear, J. C. Whitbeck, R. J. Eisenberg, G. H. 506 Cohen, and L. J. Sigal. 2008. The orthopoxvirus type I IFN binding protein is 507 essential for virulence and an effective target for vaccination. J Exp Med 508 205:981-992. 509
42. Xu, R. H., M. Fang, A. Klein-Szanto, and L. J. Sigal. 2007. Memory CD8+ T cells 510 are gatekeepers of the lymph node draining the site of viral infection. 511 Proceedings of the National Academy of Sciences of the United States of 512 America 104:10992-10997. 513
43. Xu, R. H., D. Rubio, F. Roscoe, T. E. Krouse, M. E. Truckenmiller, C. C. Norbury, 514 P. N. Hudson, I. K. Damon, A. Alcami, and L. J. Sigal. 2012. Antibody inhibition 515 of a viral type 1 interferon decoy receptor cures a viral disease by restoring 516 interferon signaling in the liver. PLoS pathogens 8:e1002475. 517
518
519
520
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

22
Figures and legends 521
Figure 1. L1R A33R and B5R mAbs recognize the ECTV ortholog proteins 522
and block their biological function. a) L1R mAb VMC-2 was compared for reactivity 523
with plated VACV and ECTV viral particles. The Kd calculated with Prism software using 524
non-linear fit is indicated. b) As in A but using A33R mAb VMC-78 and purified 525
recombinant EVM135 and A33R in the ELISA. c) As in B but using B5R mAb VMC-14 526
and purified recombinant B5R and EVM155 in the ELISA. d) Plaque reduction assay 527
after treatment of ECTV stocks with mAbs or rabbit antisera as indicated. e) Comet 528
inhibition in the presence of the indicated mAbs or rabbit antisera. The pictures at the top 529
are from one experiment and the pictures at the bottom are from another one. 530
Figure 2. Prophylactic and post-exposure treatment with L1R/EVM72 mAb 531
protects from mousepox. a) BALB/c mice were infected with ECTV and immediately 532
treated with 200 µg of the indicated mAbs. Survival was monitored. b) The mice in A 533
were weighed at the indicated dpi. c) Liver sections from BALB/c mice infected with 534
ECTV were stained with anti-EVM135 at the indicated dpi. d) BALB/c mice were infected 535
with ECTV and treated with the indicated mAbs and doses at the indicated dpi. Survival 536
was monitored. e) BALB/c mice were infected with ECTV, treated with the indicated 537
mAbs at 5 dpi and virus titers determined in the indicated organs at 7 dpi. f) 538
Immunohistochemistry of the livers from the mice in e. g) Mice were infected with ECTV 539
and treated at the indicated dpi with the indicated rabbit sera. Survival was monitored. 540
Figure 3. Late control of ECTV by anti-L1R/EVM072 does not require 541
antibody effector functions. a) Virus titers at 7 dpi in the indicated organs of B6.D2-542
D6- Fcer1γ0/0 mice treated with the indicated mAbs at 5 dpi. b) C3 determined by ELISA 543
at the indicated dpi in the serum of BALB/c mice infected with ECTV and treated with 544
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

23
with cobra venom factor (CVF) at 4, 6 and 8 dpi. c) Survival of BALB/c mice infected 545
with ECTV and treated with the indicated mAbs at 5 dpi and with CVF at 4, 6 and 8 dpi. 546
Figure 4. ECTV unable to make enveloped virus spreads in the liver and is 547
lethal to severe combined immunodeficient (SCID) mice. a) SCID mice were infected 548
with the indicated doses of ECTV-WT or ECTV-D036 in the footpad. Survival was 549
monitored. b) As in a but infected intraperitoneally. c) SCID mice were infected with 106 550
pfu ECTV-Δ36 ip. At 5 dpi copy numbers of transcripts of the ECTV gene EVM166 were 551
determined by qPCR in the indicated organs. d) SCID mice were infected with 106 pfu 552
ECTV-Δ36 and immunohistochemistry with anti-EVM135 sera was performed at the 553
indicated dpi. 554
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

VMC-78
10 - 4 10 - 3 10 - 2 10 - 1 100 101 102 103-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
A33R Kd=0.015 nM
EVM135 Kd=0.013 nM
Antibody concentration(nM)
VMC-2
10 - 4 10 - 3 10 - 2 10 - 1 100 10 1
0.0
0.2
0.4
0.6
VV Kd=0.029 nM
ECTV Kd=0.029 nM
Antibody Concentration (nM)
OD
450nm
a b
VMC-14
10 - 4 10 - 3 10 - 2 10 - 1 100 101 102 103-0.5
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
B5R Kd=0.043 nM
EVM155 Kd=0.056 nM
Antibody concentration(nM)
c
Figure 1
e VMC-78 VMC-14No mAb
rEVM155rL1RNo Ab VMC-2
d
VMC-78 VMC-14 rEVM155rL1RNo Ab VMC-2
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

dpi0 05 10 15 20 25
0
20
40
60
80
100
VMC-2
VMC-78
VMC-14
IgG1
Perc
ent surv
ival
PBS
5 10 15 20 25
80
90
100
110
120
VMC-2
VMC-78
dpi
% in
itia
l w
eig
ht
a
c
f
g
b
0 4 8 12 16
0
20
40
60
80
100 IgG1
200 ug VMC-2 at 4 dpi
200 ug VMC-2 at 5 dpi
200 ug VMC-78 at 4 dpi
500 ug VMC-78 at 4 dpi
500 ug VMC-78 at 5 dpi
dpi
Perc
ent surv
ival
d eSpleen (organ) Liver (gram)
IgG1 VMC-2 VMC-14VMC-78
IgG1 VMC-2 VMC-14 VMC-78
6
7
8
9
**** **
IgG1 VMC-2 VMC-14 VMC-78
0
2
4
6
8
10
Lo
g10 v
iru
s t
ite
r
****
****
****
** *
4 dpi 5 dpi 6 dpi 7 dpi 8 dpi 10 dpi3 dpiUninfected
40X
200X
0 5 10 15 20dpidpi dpi
0 5 10 15 20
Perc
ent sur v
ival
0 5 10 15 20
0
20
40
60
80
1000 dpi 2 dpi 5 dpi
Naïve
T1-IFNbp
EVM135
EVM155
L1R
Antisera
Figure 2
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from
IgG1 VMC-2
2
4
6
8
10
Lo
g10 v
iru
s(/
sp
lee
n) *
Spleena
IgG1 VMC-2
2
4
6
8
10
Lo
g10 v
iru
s/g
***
Liver
C3
ng
/ml
0 5 10 15 20
0
20
40
60
80
100
VMC-2 + CVF + ECTV
IgG1 + CVF + ECTV
dpi
Pe
rce
nt
su
rviv
al
cb
5 dpi
ECTV ECTV+CVF10 5
10 6
10 78 dpi
ECTV ECTV+CVF10 5
10 6
10 7
**** **
Figure 3
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

EV
M166 C
op
ies/n
g R
NA
(L
og
10
)
dpi dpi2 5
0
2
4
6
8
10
2 50
2
4
6
8
10Liver Spleen
0 10 20 30 40 50 600
20
40
60
80
100
dpi
Pe
rce
nt
su
rviv
al
ECTV-WT
ECTV-∆036
0 5 10 15 20 25
ECTV-∆036 (105 pfu)
ECTV-∆036 (106 pfu)
ECTV-WT (104 pfu)
ECTV-WT (105 pfu)
dpi
Footpad (3,000 pfu) Intraperitoneal
0 dpi 2 dpi 5 dpi 7 dpi 8 dpi
40X
200X
a b c
d
*** ***
Figure 4
on April 5, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from


















