
The Mathematical Epidemiology of Human Babesiosis in the North-Eastern United States - Jessica Dunn,...
41
The Mathematical Epidemiology of Human Babesiosis in the North-Eastern United States Jessica Margaret Dunn, Dr. Stephen Davis (RMIT), Dr. Andrew Stacey (RMIT), Assoc. Prof. Maria Diuk-Wasser (Yale/Columbia) J. M. Dunn (QUT) QUT Seminar 08.08.2014 1 / 41
-
Upload
qutsef -
Category
Data & Analytics
-
view
101 -
download
0
Transcript of The Mathematical Epidemiology of Human Babesiosis in the North-Eastern United States - Jessica Dunn,...

The Mathematical Epidemiology of Human Babesiosis inthe North-Eastern United States
Jessica Margaret Dunn, Dr. Stephen Davis (RMIT), Dr. AndrewStacey (RMIT), Assoc. Prof. Maria Diuk-Wasser (Yale/Columbia)
J. M. Dunn (QUT) QUT Seminar 08.08.2014 1 / 41
J. M. Dunn (QUT) QUT Seminar 08.08.2014 2 / 41

Tick-borne disease in the USA
The geographical range of tick-borne diseases are expanding. There areseven emerging tick diseases:
Lyme disease
Human babesiosis
Human anaplasmosis
Powassan
Deer tick encephalitis
B. miyamotoi borreliosis
Deer tick ehrlichiosis
J. M. Dunn (QUT) QUT Seminar 08.08.2014 3 / 41

Lyme Disease (Borrelia burgdorferi)
J. M. Dunn (QUT) QUT Seminar 08.08.2014 4 / 41

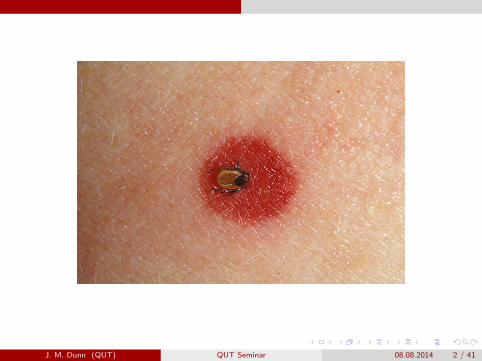
Human Babesiosis (Babesia microti)
Reported cases of Human Babesiosis – United States, 2011
J. M. Dunn (QUT) QUT Seminar 08.08.2014 5 / 41

Hosts
White-footed mice (Peromyscus leucopus) Tick (Ixodes scapularis)
J. M. Dunn (QUT) QUT Seminar 08.08.2014 6 / 41

Research Objective
To identify the key factors driving human babesiosis (B. microti) andLyme disease (B. burgdorferi) in endemic sites, and their expansioninto new areas in the north-eastern United States.
J. M. Dunn (QUT) QUT Seminar 08.08.2014 7 / 41

Mathematical Modelling Challenges
Deriving mathematical models of tick-borne disease transmission isnotoriously difficult!
Multiple hosts (competent and non-competent)
Tick life-cycle (biting rate)
Multiple tranmission routes
Multiple pathogens
J. M. Dunn (QUT) QUT Seminar 08.08.2014 8 / 41

Tick life cycle
J. M. Dunn (QUT) QUT Seminar 08.08.2014 9 / 41

Tick-phenology
Densities-Northeast
Weeks
Density
0 5 10 15 20 25 30 35 40 45 500
50
100
150
200
250
300
350
400
450
500 LarvaeNymphsAdults
J. M. Dunn (QUT) QUT Seminar 08.08.2014 10 / 41

Tick-borne pathogen transmission routes
J. M. Dunn (QUT) QUT Seminar 08.08.2014 11 / 41

Modelling challenges
The modelling challenge then becomes to one of incorporating thesecomplexities whilst maintaining a model that:
1 is representative of the transmission cycle
2 can be used with field data which will provide meaningful estimates ofthe parameters
3 has a minimal number of parameters to ensure the model can beadequately analysed
J. M. Dunn (QUT) QUT Seminar 08.08.2014 12 / 41

Overview
Model emergence- Identify the factors driving emergence- Identify control measures
Model the risk to humans- Incorporate the identified factors- Analyse changes in risk
J. M. Dunn (QUT) QUT Seminar 08.08.2014 13 / 41

Modelling emergence
Modelling emergence
The basic Reproduction number, R0
In single host systems, R0 is the expected number of secondary casesproduced by one infectious individual in a fully susceptible population.
R0 = 1 provides a threshold condition:
pathogen will spread R0 > 1
pathogen will fade out R0 < 1
J. M. Dunn (QUT) QUT Seminar 08.08.2014 14 / 41

Modelling emergence
R0 for multiple hosts
Next generation Matrix (NGM) (Diekmann and Heasterbeek)
Define kij as the expected number of new cases that have state atinfection i caused by one individual at state at infection j , during its wholeinfectious period.
For example given 2 host types i and j there are four possibilities:
K = (kij) =
(k11 k12k21 k22
)R0 is the dominant eigenvalue of the NGM such that
vk+1 = Kvk
J. M. Dunn (QUT) QUT Seminar 08.08.2014 15 / 41

Modelling emergence
NGM for tick-borne pathogens
J. M. Dunn (QUT) QUT Seminar 08.08.2014 16 / 41

Modelling emergence
Reduction for US Lyme and Human Babesiosis
J. M. Dunn (QUT) QUT Seminar 08.08.2014 17 / 41

Modelling emergence
NGM for US Lyme and Human Babesiosis
J. M. Dunn (QUT) QUT Seminar 08.08.2014 18 / 41

Modelling emergence
Quantifying R0
J. M. Dunn (QUT) QUT Seminar 08.08.2014 19 / 41
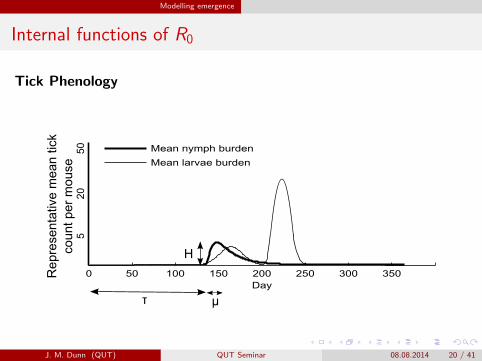
Modelling emergence
Internal functions of R0
Tick Phenology
0 50 100 150 200 250 300 350Day
Mean nymph burden
Mean larvae burden
Rep
rese
ntat
ive
mea
n tic
k co
unt p
er m
ouse
5
20
50
μ
H
τ
J. M. Dunn (QUT) QUT Seminar 08.08.2014 20 / 41

Modelling emergence
Block Island
Connecticut
100 250150 200 100 150 200 250
100 150 200 250100 150 200 250
0
1
5
20
50
150
0
1
5
20
50
150150
50
20
5
1
0
150
50
20
5
1
0
Day of year Day of year
Day of year Day of year
Larv
al ti
ck b
urde
nLa
rval
tick
bur
den
Nym
phal
tick
bur
den
Nym
phal
tick
bur
den
J. M. Dunn (QUT) QUT Seminar 08.08.2014 21 / 41

Modelling emergence
Brunner and Ostfeld (2008)
Z̄N(t) =
{HNe
− 12
[ln(
(t−τN )
µN
)/σN
]2if t ≥ τN ;
0 otherwise
Z̄L(t) =
HEe− 1
2
(t−τEµE
)2if t ≤ τL;
HLe− 1
2
[ln(
(t−τL)µL
)]2+ HEe
− 12
(t−τEµE
)2otherwise
J. M. Dunn (QUT) QUT Seminar 08.08.2014 22 / 41
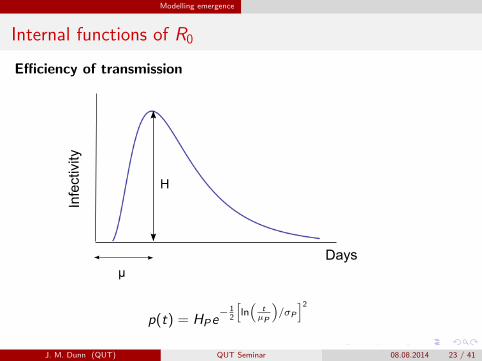
Modelling emergence
Internal functions of R0
Efficiency of transmissionInfectivity
Days
H
μ
p(t) = HPe− 1
2
[ln(
tµP
)/σP
]2
J. M. Dunn (QUT) QUT Seminar 08.08.2014 23 / 41

Modelling emergence
Global Sensitivity Analysis of R0
Ranks the parameters by their contribution to the variation of R0 usingSobol’s indices:
Main effect: calculates the effect of parameter xi on R0 fixing allother variables
Total effect: includes the main effect for xi plus all other interactioninvolving xi .
J. M. Dunn (QUT) QUT Seminar 08.08.2014 24 / 41

Modelling emergence
Global Sensitivity Results
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Parameter
Sob
ol’s
fIndi
ces
MainfEffect
TotalfEffect
H τ μ σ τ H τ μ H μ σ H Dq ρ σμ s cN N N N L L L L P P PLE E E NN
J. M. Dunn (QUT) QUT Seminar 08.08.2014 25 / 41
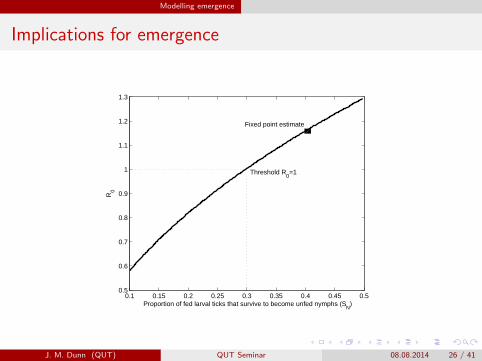
Modelling emergence
Implications for emergence
0.1 0.15 0.2 0.25 0.3 0.35 0.4 0.45 0.50.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
Proportion of fed larval ticks that survive to become unfed nymphs (SN
)
R0
Threshold R0=1
Fixed point estimate
J. M. Dunn (QUT) QUT Seminar 08.08.2014 26 / 41

Modelling emergence
Implications for control
Given, R̄0 = 1.57
Vaccination requirements (Roberts, 2003)
V = 1 − 1
R20
≈ 60%
J. M. Dunn (QUT) QUT Seminar 08.08.2014 27 / 41

Modelling emergence
The Coinfection Story
J.M. Dunn et al. Borrelia burgdorferi enhances the enzootic establishment ofBabesia microti in the northeastern United States, PLOS ONE (2014).
J. M. Dunn (QUT) QUT Seminar 08.08.2014 28 / 41

Modelling emergence
Modification of R0
k13
k31
k13
k31
k32
k23
k32
k23
White-footedmouse-Bb free,infectedawithBm1:
White-footedmouse-infectedawith Bb & Bm2:
TickainfectedawithaBmaduringa1stabloodmeal3:
Ka= 0 0
0 0
0
k13
k31
k23
k32R =0 +
1 2
3
R0 =√k13k31 + k23k32√√√√. . .
∫ t=365
t=0
. . .
(ψ
∫ t′=365−t
t′=0
p1(t ′) . . . dt ′ + (1 − ψ)
∫ t′=365−t
t′=0
p2(t ′) . . . dt ′
)dt
J. M. Dunn (QUT) QUT Seminar 08.08.2014 29 / 41
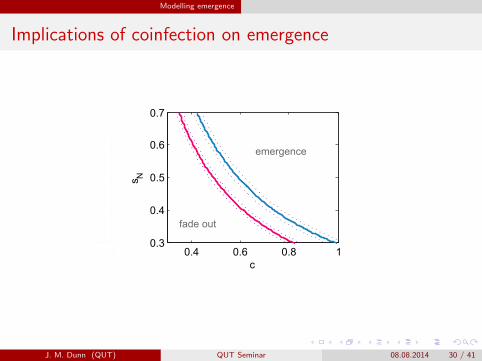
Modelling emergence
Implications of coinfection on emergence
0.4 0.6 0.8 10.3
0.4
0.5
0.6
Low8prevalence
c
s N
0.4 0.6 0.8 10.3
0.4
0.5
0.6
High8prevalence
c
s N
0.4 0.6 0.8 10.3
0.4
0.5
0.6
0.7
c
s N
0.4 0.6 0.8 10.3
0.4
0.5
0.6
0.7
c
s N
Babesia8microti
Coinfection
Block8Island
Connecticut
30w8B88 b8prevalence8in8mice 80w8Bb8prevalence8in8mice
Student8Version8of8MATLAB
B. microti
B. microti C8B. Burgdorferi BL2068
fade8out fade8out
fade8outfade8out
emergence emergence
emergenceemergence
30w8B. burgdorferi8BL2068prevalencein8mice
80w8B. burgdorferi8BL2068prevalencein8mice
J. M. Dunn (QUT) QUT Seminar 08.08.2014 30 / 41

Modelling emergence
Timing is everything!120 140 160 180 200 220 240 260 280 300
0
5
10
15
20
25
30
35
120 140 160 180 200 220 240 260 280 3000
5
10
15
Rep
rese
ntat
ive3
mea
n3tic
k3co
unt3p
er3m
ous
e Prop
ortion3of3infected3larval3ticks3per3m
ouseP
roportion3of3infected
3larval3ticks3per3mouseR
epre
sent
ativ
e3m
ean3
tick3
coun
t3per
3mo
use
Block3Island
Connecticut
3333333333333333333333333333330.93333330.83333330.73333330.63333330.53333330.43333330.333333330.233333330.1
Mean nymph burden
Mean larvae burden
Babesia3+3Borrelia
Babesia
3333333333333333333333333333330.93333330.83333330.73333330.63333330.53333330.43333330.333333330.233333330.1
J. M. Dunn (QUT) QUT Seminar 08.08.2014 31 / 41

Modelling emergence
Coinfection is not the whole story!
Accounting for aggregation on hosts
k13
k13
32
1
54
2
k51
k15
k12
k21
k14
k41
2: High aggregation white footed mouse - infected with Bb
4: Low aggregation white footed mouse - infected with Bb
3: High aggregation white footed mouse - infected with Bm
5: Low aggregation white footed mouse - infected with Bm
R0 =√k12k21 + k12k21 + k13k31 + k14k41 + k15k51
J. M. Dunn (QUT) QUT Seminar 08.08.2014 32 / 41

Modelling emergence
Scenario Estimated R0
No co-aggregation; no coinfection 0.70 (0.62,0.78)
Low co-aggregation; no coinfection 0.80 (0.71,0.86)
Moderate co-aggregation; no coinfection 0.97 (0.81,1.04)
High co-aggregation; no coinfection 1.13 (1.00, 1.21)
High co-aggregation; coinfection 1.78 (1.64, 1.91)
J. M. Dunn (QUT) QUT Seminar 08.08.2014 33 / 41

Modelling emergence
Conclusions
Epidemiological:
Values of R0 are consistently low 1 < R0 < 3
Transmission efficiency drives emergence
Timing is everything!
Mathematical:
Models are mechanistic, transparent, linked directly with field data
Step towards a model for more complicated tick-borne pathogens
First such model that that assesses the importance of (i) coinfectionand (ii) aggregation
J. M. Dunn (QUT) QUT Seminar 08.08.2014 34 / 41
Modelling emergence
Questions?
J. M. Dunn (QUT) QUT Seminar 08.08.2014 35 / 41

Modelling risk
Modelling risk to humans
Risk is directly proportional to the infection prevalence in nymphal ticks.
Compartment type SIR Model: (S)usceptibles to (I)nfectives to(R)ecovered
J. M. Dunn (QUT) QUT Seminar 08.08.2014 36 / 41
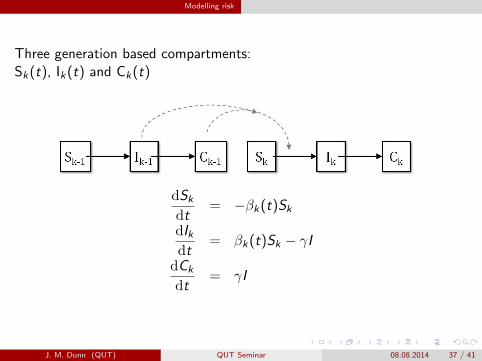
Modelling risk
Three generation based compartments:Sk(t), Ik(t) and Ck(t)
dSkdt
= −βk(t)Sk
dIkdt
= βk(t)Sk − γI
dCk
dt= γI
J. M. Dunn (QUT) QUT Seminar 08.08.2014 37 / 41

Modelling risk
Force of Infection
The force of infection is related to the unfed nymphs from the previousyear k − 1
βk(t) =1
DNνk Z̄N(t)qN .
with the proportion of infected unfed nymphs, νk , in year k is given by
νk =
∫ 365
0aL(t)p̄
Ik−1
Nk−1dt
J. M. Dunn (QUT) QUT Seminar 08.08.2014 38 / 41
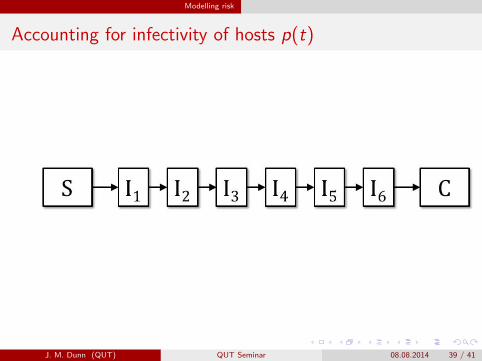
Modelling risk
Accounting for infectivity of hosts p(t)
J. M. Dunn (QUT) QUT Seminar 08.08.2014 39 / 41

Modelling risk
dSkdt
= −βk(t)Sk + b(t)Nk − (µ+Nk
K)Sk
dIk,1dt
= βk(t)Sk − (µ+Nk
K)Ik,1 − γI1
dIk,2dt
= γI1 − (µ+Nk
K)Ik,2 − γI2
dIk,3dt
= γI2 − (µ+Nk
K)Ik,3 − γI3
dIk,4dt
= γI3 − (µ+Nk
K)Ik,4 − γI4
dIk,5dt
= γI4 − (µ+Nk
K)Ik,5 − γI5
dIk,6dt
= γI5 − (µ+Nk
K)Ik,6 − γI6
dCk
dt= γI6 − (µ+
Nk
K)C
J. M. Dunn (QUT) QUT Seminar 08.08.2014 40 / 41
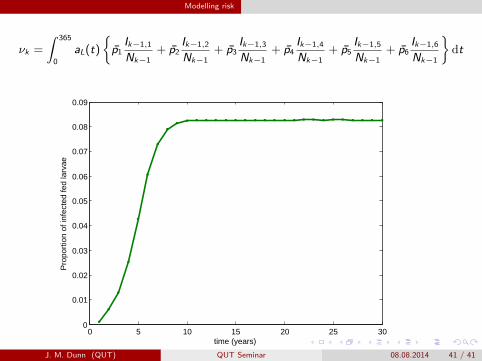
Modelling risk
νk =
∫ 365
0
aL(t)
{p̄1
Ik−1,1
Nk−1+ p̄2
Ik−1,2
Nk−1+ p̄3
Ik−1,3
Nk−1+ p̄4
Ik−1,4
Nk−1+ p̄5
Ik−1,5
Nk−1+ p̄6
Ik−1,6
Nk−1
}dt
0 5 10 15 20 25 300
0.01
0.02
0.03
0.04
0.05
0.06
0.07
0.08
0.09
time (years)
Pro
port
ion
of in
fect
ed fe
d la
rvae
Student Version of MATLAB
J. M. Dunn (QUT) QUT Seminar 08.08.2014 41 / 41


















