
THE JOURNAL OF TURKISH PHYTOPATHOLOGY 20x28.pdfUzunköprü (Edirne) Çeltik Alanlarında Yoğun...
60
THE JOURNAL OF TURKISH PHYTOPATHOLOGY SCIENTIFIC REVIEW BOARD The Editor-in-Chief and Editorial Boards of The Journal of Turkish Phytopathology would like to extend their sincere appreciation to all those who have worked as referees for the Journal. Thank you for sharing your time, effort and professional expertise. Prof. Dr. Gülay TURHAN Prof. Dr. Semih ERKAN Prof. Dr. Ersin ONOĞUR Prof. Dr. Nafiz DELEN Prof. Dr. Filiz ERTUNÇ Prof. Dr. Figen YILDIZ Prof. Dr. Saadettin BALOĞLU Prof. Dr. Abuzer SAĞIR Prof. Dr. Savaş KORKMAZ Prof. Dr. Murat H.SİPAHİOĞLU Prof. Dr. Yeşim AYSAN Prof. Dr. Semra DEMİR Prof. Dr. Emin ONAN Assoc. Prof. Dr. Fikret DEMİRCİ Prof. Nuh BOYRAZ Assoc. Prof. Mona GAZEL Prof. Dr. Yusuf YANAR Assoc. Prof. Havva İLBAĞI Prof. Dr. Mehmet E. GÜLDÜR Assoc. Prof. Dr. Ömer ERİNCİK Prof. Dr. F. Sara DOLAR Assoc. Prof. Dr. Dr. İlhan KAYA Prof. Dr. Berna TUNALI Assoc. Prof. Dr. Dr. Hülya ÖZGENEN Assoc. Prof. Dr. Himmet TEZCAN Asist. Prof. Dr. Kadir İLHAN Assoc. Prof. Dr. Mustafa GÜMÜŞ Asist. Prof. Dr. Mustafa KÜSEK Asist. Prof. Dr. Sibel DERVİŞ Dr. Üftade GÜNER Asist. Prof. Dr. Kubilay K. BAŞTAŞ Dr. Behçet Kemal ÇAĞLAR Asist. Prof. Dr. İsmail Can PAYLAN Dr. Mehmet DEMİRCİ Dr. Aydan KAYA Dr. Suat KAYA All rights of articles published in this journal are reserved by The Turkish Phytopathological Society. Any use of the material, including reproduction in whole or in part requires permission in writing from The Turkish Phytopathological Society. Meta Basım Matbaacılık Hizmetleri 87 Sok. No. 4 / A Bornova (0.232) 343 64 54 [email protected] İzmir, 2014 Basım Tarihi: 28.05.2014 ISSN 0378 - 8024 http://www.fitopatoloji.org.tr
Transcript of THE JOURNAL OF TURKISH PHYTOPATHOLOGY 20x28.pdfUzunköprü (Edirne) Çeltik Alanlarında Yoğun...

THE JOURNAL OF TURKISH
PHYTOPATHOLOGY SCIENTIFIC REVIEW BOARD The Editor-in-Chief and Editorial Boards of The Journal of Turkish Phytopathology would like to extend their sincere appreciation to all those who have worked as referees for the Journal. Thank you for sharing your time, effort and professional expertise. Prof. Dr. Gülay TURHAN Prof. Dr. Semih ERKAN Prof. Dr. Ersin ONOĞUR Prof. Dr. Nafiz DELEN Prof. Dr. Filiz ERTUNÇ Prof. Dr. Figen YILDIZ Prof. Dr. Saadettin BALOĞLU Prof. Dr. Abuzer SAĞIR Prof. Dr. Savaş KORKMAZ Prof. Dr. Murat H.SİPAHİOĞLU Prof. Dr. Yeşim AYSAN Prof. Dr. Semra DEMİR Prof. Dr. Emin ONAN Assoc. Prof. Dr. Fikret DEMİRCİ Prof. Nuh BOYRAZ Assoc. Prof. Mona GAZEL Prof. Dr. Yusuf YANAR Assoc. Prof. Havva İLBAĞI Prof. Dr. Mehmet E. GÜLDÜR Assoc. Prof. Dr. Ömer ERİNCİK Prof. Dr. F. Sara DOLAR Assoc. Prof. Dr. Dr. İlhan KAYA Prof. Dr. Berna TUNALI Assoc. Prof. Dr. Dr. Hülya ÖZGENEN Assoc. Prof. Dr. Himmet TEZCAN Asist. Prof. Dr. Kadir İLHAN Assoc. Prof. Dr. Mustafa GÜMÜŞ Asist. Prof. Dr. Mustafa KÜSEK Asist. Prof. Dr. Sibel DERVİŞ Dr. Üftade GÜNER Asist. Prof. Dr. Kubilay K. BAŞTAŞ Dr. Behçet Kemal ÇAĞLAR Asist. Prof. Dr. İsmail Can PAYLAN Dr. Mehmet DEMİRCİ Dr. Aydan KAYA Dr. Suat KAYA
All rights of articles published in this journal are reserved by The Turkish Phytopathological Society. Any use of the material, including reproduction in whole or in part requires permission in writing from The Turkish Phytopathological Society.
Meta Basım Matbaacılık Hizmetleri
87 Sok. No. 4 / A Bornova (0.232) 343 64 54 [email protected]
İzmir, 2014
Basım Tarihi: 28.05.2014
ISSN 0378 - 8024 http://www.fitopatoloji.org.tr


THE JOURNAL OF TURKISH PHYTOPATHOLOGY
TURKISH PHYTOPATHOLOGICAL SOCIETY
VOL. 41 2013, December NO. 1-3
CONTENTS
Determination of Weeds in Rice Region of Edirne - Uzunköprü and Researches on Chemical Control of Those Weeds Uzunköprü (Edirne) Çeltik Alanlarında Yoğun Olarak Bulunan Yabancı Otların Belirlenmesi ve Kimyasal Savaşımı Üzerinde Araştırmalar K. UZUN, H. DEMİRKAN ......................................................................................................................... 1
Studies on Esca and Petri Diseases in Grapevine Nurseries and Vineyards in Aegean Region Ege Bölgesi Asma Fidanlıkları ve Bağlarında Kav ve Petri Hastalıkları Üzerinde Araştırmalar D. POYRAZ, E. ONOĞUR ....................................................................................................................... 13
Optimization of a Specific Real-Time PCR Assay for Fusarium avenaceum Using Species-specific Couples of Primer Fusarium avenaceum için tür-spesifik primerler kullanılarak real time PCR yönteminin optimizasyonu Ş. E. ARICI, P. KARLOVSKY .................................................................................................................. 29
Identification and Pathogenicitiy of Botryosphaeriaceae Species Causing Wood Canker on Grapevines Asmada Kol ve Sürgün Kurumalarina Neden Olan Botryosphaeriaceae Türlerinin Teşhisi ve Patojenisiteleri D. S. AKGÜL, N. GÜNGÖR SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT ............................ 35
Combination of Irradiation and Sodium Carbonate to Control Postharvest Penicillium Decay of Apples Elmalarda Hasat Sonu Penicillium Çürüklüğünün Engellenmesi için Işınlama ve Sodyum Karbonat Kombine Uygulaması C. TEMUR, O. TİRYAKİ........................................................................................................................... 47


J. Turk. Phytopath., Vol. 42 No. 1-3, 1-12, 2013 ISSN 0378 - 8024
1
Determination of Weeds in Rice Region of Edirne - Uzunköprü and Researches on Chemical Control of Those Weeds
Kayahan UZUN * Hasan DEMİRKAN **
* 153 Sok. No: 66/7 Bornova /İZMİR ** E.Ü.Ziraat Fakültesi, Bitki Koruma Bölümğ, 35100 Bornova/İZMİR, [email protected] Accepted for publication January15, 2014
ABSTRACT
In our cuntry, rice agriculture is commonly applied in center of Edirne, Uzunköprü, Meriç, İpsala, Keşan and Enez regions which meet 25% of sowing area and % 28 of rice production in our country. The weeds which causes some problems are Echinochloa crus-galli, E. oryzoides, Cyperus difformis, Diplachne fusca, Polygonum lapatifolium, Paspalum paspaloides, Lindernia dubia and Ammania coccinea., Diplachne fusca has entered to our country in recent years grass and began to spread very quickly. In 2008, this study was carried out because of the difficulty of conflict with D. fusca and to compete with rice. Experiments were made in two different phenologic development period of D.fusca in Uzunköprü. Assay was established as 4-character and four iteration by Randomized Complate Blocks type design. Before grass tillering period of D. fusca; the effectiveness of cyhalafop – buthyl with 75 ml/da dose was 79% at day 18 and 82% at day 33, and 100% at day 18 and 33 with 150 ml/da dose. The effectiveness of fenoxaprop-p-ethyl + izoxafen effective herbicide at 80 ml/da dose was determined as 95% and 98% at day 18 and 33, respectively. In tillering period of Diplachne fusca; when cyhalafop - buthyl was applied at 100 ml/da dose, it was found to be effective as 71% at day 26, 73% at day 45 and at 150 ml/da dose, 100% at day 26 and 45 days and for Fenoxaprop-p-ethyl + izoxafen at 100 ml/da dose, it was found as 97% at day 26 and 98% at day 45.
Keywords: Rice, Diplachne fusca, herbicide, Edirne
GİRİŞ
Çeltik (Oryza sativa L.) tarımı, M.Ö. 3000 yıllarında Güney Hindistan’dan Çin’e, M.Ö. 1000 yıllarında Java’ya doğru yayılmış, Avrupa’ya Büyük İskender’in Asya seferleri sonunda M.Ö. 300 yıllarında tanıtılmıştır. Türkiye’ye yaklaşık 500 yıl önce Güney’den girdiği sanılmaktadır (Kün, 1985). Dünya’da üretilen çeltiğin işlendikten sonra yaklaşık %90’ını insan beslenmesinde, %3.5’u tohumluk, %1.5’i endüstriyel alanda tüketilmekte ve %4.5’u ise kayıp olmaktadır. Dünyada üretilen çeltiğin %91’lik kısmı Asya ülkeleri tarafından, geriye kalan %6.9’luk kısmı da diğer ülkeler tarafından tüketilmektedir (Finassi, 1979). Batı Marmara (Trakya) Bölgesi Türkiye toplam çeltik ekilişinin %59,32’sini, çeltik üretiminin %57’sini karşılamaktadır. (Anonymous, 2007).
Besin değeri açısından ele alındığında pirinç buğdaya göre daha önemli bir yer tutmaktadır. Çünkü 100 g buğday 350 kalori ve %1.5 yağ içerirken pirinç 360 kalori ve %1.7 yağ içermektedir (Anonymous, 1969). Artan kalori ihtiyacının sağlanmasında pirincin önemli bir yer tutacağı anlaşılmaktadır. Türkiye’de Marmara Bölgesi’ndeki toplam 74.500 ha çeltik ekilişinin 47.000 hektarı Trakya (Avrupa)’da, 27.500 hektarı Marmara (Anadolu)’da yapılmaktadır (Demircan, 2006). Daha kuzeyde kaldığı için Trakya’da çeltik ekimi Marmara’ya göre 15-20 gün daha geç yapılmaktadır.
Çeltik, sürekli su dolu tavalarda yetiştirildiğinden suda yetişen yabancı otlardan darıcan (Echinochloa crus-galli (L.) P.B., E. oryzoides (Ard.) Fritsch), kızotu (Cyperus difformis L.), başta olmak üzere barajotu (Diplachne

DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
2
fusca (L.) P. Beauv), söğütotu (Ammania coccinea Rottb.) ve dipotu (Lindernia dubia (L.) Pennell) çıkıştan itibaren kültür bitkisi ile topraktaki bitki besin elementleri için, rekabet ederek çeltiğin gelişmesine engellemekte, verim azalışına neden olmakta (Işık ve Mennan 2001), hasat ve harman maliyetini arttırmakta, tohumları ürüne karışarak pazar değerini düşürmektedir. Özellikle barajotu, yakın tarihte Türkiye’de görülmüş olmasına rağmen tohumları su ve rüzgar ile kolayca yayıldığından kısa zamanda Trakya’daki çeltik alanlarını kaplamıştır.
Bu çalışmanın amacı, üretim koşulları ve şartları alışagelmiş kültür bitkilerinden daha zahmetli olan ve ülkemizde büyük alanlarda ve miktarlarda üretilen çeltik bitkisinin en büyük problemlerinden biri olan yabancı ot sorununa karşı Uzunköprü ilçesinde yoğun olan yabancı otların belirlenmesi ve bu yabancı otlar içerisinde de bulaşıklığı yakın zamanda (Sürek, 2002) tespit edilmiş olan barajotu’nun kimyasal mücadelesine yönelik araştırma yapmak olmuştur.
Çeltik Üretim Alanları
Dünya’da pirinç ekim alanları giderek genişlemekte ve üretimde de buna paralel bir artış yaşanmaktadır. Pirinç ekim alanlarının toplamı, 1948–1952 arası dönemde ortalama olarak 102,5 milyon ha kadar iken bu değer 2004 yılında 151,3 milyon ha’a ulaşmıştır. Pirinç üretimi ise aynı süre zarfında yaklaşık olarak 165 milyon tondan 605.7 milyon tona çıkmıştır. Pirinç üretildiği yerde tüketilmekte olan bir tahıldır. Bu yüzden dünyada yetiştirilen toplam çeltiğin % 90’ından fazlası Asya ülkelerindedir (Çizelge 1). Fakat dış satımda ise, %1.5’la en yüksek değere ABD sahiptir (Doğanay, 2007).
Çizelge 1. Dünyada pirinç yetiştiren ülkeler (Bulut, 2006).
Ülkeler Ekiliş (ha) Üretim (ton) Verim kg/da
Dünya 146.029.456 579.476.000 396.8
Hindistan 40.000.000 123.000.000 307.5
Çin 28.360.000 177.589.000 626.2
Endonezya 11.500.000 48.654.000 423.0
Bangladeş 10.900.000 39.000.000 357.8
Tayland 9.920.000 27.000.000 272.1
Vietnam 7.539.000 31.319.000 415.4
Myanmar 6.200.000 21.200.000 341.9
Filipinler 4.034.000 12.684.800 314.4
Brezilya 3.174.840 10.489.400 330.3
Japonya 1.700.000 11.264.000 662.5
A.B.D. 1.297.840 9.616.400 740.9
Türkiye 85.000 400.000 470.5
Çin eskiden beri dünya pirinç üretiminde ön sıralarda yer almaktadır. Ülkenin 2006 yılında 28.4 milyon ha ekim alanından üretimi 177.6 milyon tonu bulmuştur. Günümüzde de pirinç üretiminde 1. sırada yer almaktadır. Hindistan ise, son yıllarda 100 milyon ton pirinç üretimiyle 2. sırayı almaktadır. Birim alandan alınan verim dünya ortalamasından düşüktür. Fakat son yıllarda tarımda yaşanan gelişmelere bağlı olarak gelecekte Hindistan birinci sırayı alacaktır. Diğer önemli üreticiler Muson Asyası ülkelerinden olan Endonezya, Bangladeş, Vietnam, Tayland, Myanmar, Flipinler, Japonya vs. diğer ülkelerdir (Çizelge 1) (Özşahin, 2008’e atfen Bulut 2006).
Türkiye'de de kaliteli tohumun yaygınlaşmasıyla birlikte çeltik üretiminde dünya ortalamasının üzerinde verim elde edilmeye başlanmıştır. Daha kaliteli ve verimli çeltik tohumu üzerinde devam eden bilimsel çalışmalar meyvesini vermeye başlamıştır. Yaklaşık 10 yıl öncesinde Türkiye'de yaygın olarak kullanılan çeltik tohumundan dekar başına yaklaşık 500 kg verim elde edilirken, bu rakam şu anda 750 ile 850 kg arasında değişim göster-mektedir.
Ülkemizde çeltik tohumu üzerindeki bilimsel çalışmalar Edirne'de bulunan Trakya Tarımsal Araştırma Enstitüsü tarafından yapılmaktadır (Beşer, 2009).

K. UZUN, H. DEMİRKAN
3
Çeltikte Yabancı Otun Önemi
Çeltikte sorun olan yabancı otlardan Alisma plantago-aquatica (kurbağa kaşığı), Equisetum arvense (at kuyruğu), Butomus umbellatus (su menekşesi) hariç diğerleri tek çenekli yabancı otlardır. E. crus-galli (darıcan) ve Panicum spp. (yabani darı) sadece tohumlarıyla, diğerleri hem tohum, hem de rizom ve stolonlarıyla ürerler. Kışı toprakta rizom veya tohum halinde veya ürün içinde tohum olarak geçirirler. Havaların ısınmasıyla gelişmeye başlar, Haziran ve Temmuz sonuna kadar çeltik tavalarını istila ederler (Akkoyunlu, 2005).
Çeltikte sorun oluşturan yabancı otlarının yayılışları, sulama, sel suları ve hasat sonrası hayvan otlamalarıyla olduğu gibi bulaşık tohum ve yanmamış çiftlik gübrelerinin kullanılması, toprak aletleri, harman makineleri ve rüzgârla olabilmektedir (Akkoyunlu, 2005).
Çeltik tarlalarındaki yabancı otların zararı; besin maddesi, su sarfiyatı, ışıklanma ve kaplama alanı işgal bakımından rekabet temeline dayanır. Yabancı otların gelişme yetenekleri kültür bitkilerine nazaran yüksek olduğundan, genellikle çeltikler bu yarışmaya dayanamaz seyrelirler, gelişmeleri cılız ve bodur kalır, kardeşlenme az olur, alınan ürün istenilen düzey ve nitelikte olmaz.
Çeltik tarımına yeni açılmış alanlar dışında, yabancı ot mücadelesi yapılmaksızın çeltik ziraatını düşünmek mümkün değildir. Yabancı otların çeltiklerde meydana getireceği zarar, tavalardaki ot türlerine, yoğunluklarına ve çevre koşullarına bağlı olarak değişmeler gösterebilir. ABD Arkansas'ta yapılan bir araştırma sonucuna göre; kuruya mibzerle ekilen Lebonnet çeşidinde, 215-270 arasında değişen bitki sıklığında, metrekaredeki 11, 22, 54 ve 108 D. fusca bitkileri, sıra ile %9, 18, 20 ve 36 verim azalmasına neden olmuştur. Metrekaredeki bir adet D. fusca bitkisi dekarda verimi 2.1 kg azaltmıştır. Metrekarede 54-108 arasında değişen D. fusca bitkisi pirinç randımanını ve 108 bitki ise çimlenme oranını düşürmüştür (Smith, 1983).
Yabancı otlar çeltik bitkisi ile rekabeti yanında yabancı ot tohumlarının ürüne karışarak ürünün pazar değerini düşürmektedir. Tohumluk olarak kullanılan çeltik içindeki yabancı ot tohumları temiz tarlalara bulaşarak geniş alanlara yayılmalarına neden olmaktadır. Diplachne fusca ABD, İspanya ve İtalya gibi ılıman iklim böl-gelerinde bulunan çeltik tarlalarında ve Asya'nın bazı bölgelerinde bulunabilmektedir. Bu yabancı ot İtalya'ya İspanya'dan 1990'lı yılların başında, Amerikan Thaibonnet çeşidinin tohumu ile gelmiştir (Damar 2006’ya atfen Romani ve Tabacchi, 2000).
Yabancı ot ile bulaşık çeltik tarlalarında hasat ve harman maliyeti artmakta ve ertesi yıla yabancı ot tohumları ile daha yoğun olarak bulaşık bir tarla kalmasına neden olmaktadır.
Çeltikte dar yapraklı yabancı otlar olan E. crus-galli ve Diplachne fusca benzer kök yapısı, benzer besinleri tüketmesi ve çeltikle birlikte çimlenerek benzer gelişme devresi nedeniyle geniş yapraklı yabancı otlara göre çeltikle daha şiddetli olarak rekabet etmektedir.
Çeltikte yabancı ot rekabetini ortadan kaldırmak amacı ile elle yolma veya ilaçlı mücadele yapılmaktadır.
Yapılan çalışmalarda çeltikte en yoğun olarak bulunan yabancı otlar arasında E. crus-galli ve C. difformis sayılabilmektedir (Chang, 1970).
E. crus-galli’nin bulunuş oranına göre çeltikte neden olduğu ürün kayıpları çok değişmektedir. E. crus-galli bulunmadığında çeltikte verim 580 kg/da, m2’de 10 adet darıcan bulunduğunda ise verim 350 kg/da’a düşmekte ve ürün kaybı %40 oranında olmaktadır. Buna karşın m2’de 50 darıcan bulunduğunda çeltik verimi 200 kg/da olmakta, verim kaybı %66’ya yükselmektedir (Smith et al., 1977).
MATERYAL VE METOD
Materyal
Çalışmanın esas materyalini bölgede çeltik alanlarındaki yabancı otlar ve bunlardan D. fusca’a karşı kullanılan bazı herbisitler oluşturmaktadır. Herbisit denemesi Edirne İli Uzunköprü İlçesi Balabanköy’de yapılmıştır.

DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
4
Çalışmada çeltikte baraj otunun ilaçlı mücadelesi için, Türkiye’de ruhsatlı iki herbisit denemiştir. Bu ilaçların etkili maddeleri ve formülasyon şekli aşağıda verilmiştir (Çizelge 2).
Çizelge 2. Çeltikte barajotuna karşı denemeye alınan ilaçlar
Etkili Madde Adı ve Oranı Formülasyon Şekli
Fenoxaprop-p-ethyl + 75 g/l
isoxadifen-ethyl
EC
Cyhalafop-butyl, 200 g/l EC
İlaçlama aleti: Denemede, sabit basınçta çalışan Matabi marka elektrikli sırt pompası kullanılmıştır. Uygulamalar, 2 bar basınçta, 4 adet yelpaze püskürtme yapan meme (Lurmark marka, No: 01 F 110) ve 2 m iş genişliği olan bum takılı, elektrikli, sabit basınçlı sırt pompası ile yapılmıştır.
Metod
Sürvey çalışması ve yabancı ot türlerinin belirlenmesi
Sürvey çalışması, Edirne ili Uzunköprü ilçesinde tesadüfi olarak seçilen 20 çeltik tarlasında, yabancı otlar keşif sürveyi yapılarak belirlenmiştir (Karaca et al, 1970). Belirlenmiş olan tarlalara girilip, tarlanın köşegenleri yönünde yürünerek var olan yabancı otlar tür düzeyinde belirlenenerek kaydedilmiştir. Yabancı otların teşhisi Davis (1985)’e göre yapılmıştır.
Herbisit denemeleri
Herbisit denemeleri, Osmancık çeltik çeşidi ekilmiş bir tarlada yapılmıştır. Dekara 20 kg çeltik tohumu kullanılmıştır.
Deneme 2008 yılında, bir yıl önce de çeltik ekilmiş tarlada yapılmıştır. Deneme yapılacak tarla kışa girmeden, 2007 yılı Kasım ayında pulluk ile sürülmüş ve 2008 yılı Nisan ayında tiller denilen toprak işleme aleti ile toprak karıştırılmıştır. Çeltik ekiminden önce tırmık çekip 20 kg/da (N15-P25-K15) taban gübresi verilmiştir. Çeltik tavalarındaki setler sağlamlaştırılıp, suyun giriş, çıkış yerleri (peçeler) belirlenmiş, aşınmaya karşı naylon örtülerle sağlamlaştırılmıştır. Tırmık çekme ve gübreleme işlemlerinden önce çeltik ekimi yapılmıştır.
Sulama, sanayi atıklarından dolayı su kirliliği olan Ergene nehrinden motopompla yapılmıştır.
Deneme alanında çeltik ekimi 25.05.2008 tarihinde yapılmıştır. Çeltik tohumları bir gün önceden ıslatılarak burunlanma devresine geldiğinde 8-10 cm su derinliğindeki tavalara serpme olarak ekilmiştir.
Baraj otu’nun iki ayrı fenolojik gelişme döneminde herbisitlerin etkisini belirlemek amacı ile birbirlerine su ve ilaçlı su sızdırmayacak şekilde 1 m genişliğinde set ile ayrılmış iki tarlada, 7 gün ara ile iki deneme kurulmuştur.
I. Deneme: Barajotu’nun kardeşlenme dönemi öncesi,
II. Deneme: Barajotu’nun kardeşlenme devresinde kurulmuştur.
Denemelerde Barajotu’na karşı kullanılan ilaçlar ve dozları Çizelge 3’de verilmektedir.
Denemeler, tesadüf blokları denemeler desenine göre 4 (3 ilaç + şahit) karakterli ve 4 tekerrürlü olarak kurulmuş ve parseller 2 m x 17 m = 34 m2 olarak alınmıştır. Parseller arasında 0.5 m, bloklar arasında 1 m emniyet şeridi bırakılmıştır.

K. UZUN, H. DEMİRKAN
5
Çizelge 3 Barajotu’na karşı kullanılan ilaçlar ve dozları
Barajotu’nun kardeşlenme devresi öncesi uygulama denemesi
Etkili Madde Adı ve Oranı Form. Şekli Kullanılan Doz
Fenoxaprop-p-ethyl + 75 g/l isoxadifen-ethyl EC 80 ml/da
Cyhalafop-butyl, 200 g/l EC 75 ml/da
Cyhalafop-butyl, 200 g/l EC 150 ml/da
Kontrol
Barajotu’nun kardeşlenme devresinde uygulama denemesi
Fenoxaprop-p-ethyl + 75 g/l isoxadifen-ethyl EC 100 ml/da
Cyhalafop-butyl, 200 g/l EC 100 ml/da
Cyhalafop-butyl, 200 g/l EC 150 ml/da
Kontrol
Herbisitlerin Uygulama Zamanları
Barajotu’nun kardeşlenme devresi öncesi uygulama denemesi
Herbisit uygulaması, çeltik ekiminden 18 gün sonra Barajotu’nun 3–4 yapraklı devresinde, 12.06.2008 tarihinde yapılmıştır. Uygulamadan 3 gün önce tavaların suyu kesilmiş, ancak ilaçlama tarla tamamen kurumadan, balçık halde, çizmenin 4-8 cm çamura battığı tavalarda yapılmıştır. İlaçlama sırasında çeltiğin 1-2 yapraklı devrede olduğu belirlenmiştir.
Uygulamadan 24 saat sonra denemenin yapıldığı tarla 8-10 cm derinliğinde su ile doldurulmuştur. Uygulamadan 7 gün (12.06.2008), 18 gün (30.06.2008), 33 gün (15.07.2008) ve 52 gün (03.08.2008) sonra yapılan değerlendirmelerde sonuçlar kaydedilmiş ve COSTAT istatistiki analiz programında Duncan testi ile herbisitlerin etkinlilikleri değerlendirilmiştir.
Barajotu’nun kardeşlenme devresinde uygulama denemesi:
İlaç uygulaması, çeltik ekiminden 25 gün sonra, barajotu’nun 2-3 kardeşli olduğu devrede, 19.06.2008 tarihinde kurulmuştur. Uygulamadan 4 gün önce tavaların suyu kesilmiş, ancak tarla tamamen kurumadan, balçık halde, çizmenin 3-5 cm çamura battığı tavalarda yapılmıştır. İlaçlama sırasında çeltiğin 4-5 yapraklı olduğu devrede belirlenmiştir.
Uygulamadan 11 gün (30.07.2008), 26 gün (15.07.2008), 45 gün (03.08.2008) ve 52 gün (10.08.2008) sonra yapılan değerlendirmelerde sonuçlar kaydedilmiş ve istatistiki analiz programı ile herbisitlerin etkinlilikleri değerlendirilmiştir.
Uygulamadan 24 saat sonra denemenin yapıldığı tarla 10-12 cm derinliğinde su ile doldurulmuştur.
İlaçlamadan önce kalibrasyon yapılarak 4 tekerrüre gidecek su miktarı belirlenmiştir. Uygulamada 31,25 lt/da su kullanılmıştır.
Baraj otunun kardeşlenme devresi öncesi uygulama denemesinde, uygulama anında hava açık, güneşli ve uygulama anındaki hava sıcaklığın 26-28oC, orantılı nemin %55-65, baraj otunun kardeşlenme devresi uygulama denemesinde hava açık ve uygulama anındaki hava sıcaklığın 33oC, orantılı nemin %43 olduğu gölgedeki elektronik meteorolojik alet ile belirlenmiştir.
İlaçların yabancı otlara etkisi, 1-9 EPPO skalasına göre %0 -100 etki değerleri esas alınarak görsel olarak değerlendirilmiştir (Bora ve Karaca, 1970). Değerlendirmede barajotu esas alınarak kontrole göre barajotunda meydana gelen azalmalar, kaplama örtüsü, boyda kısalma yüzdesi saptanmıştır. Skalaya göre; %90 ve üstündeki değerler yeterli, altındaki değerler ise yetersiz etkili olarak kabul edilmiştir.
Elde edilen verilere COSTAT istatistik analiz programında Duncan testi uygulanmıştır.
DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
6
BULGULAR
Sürveyde belirlenen yabancı ot türleri ve rastlanma sıklıkları
Tesadüfen seçilen 20 tarlada yapılan sürveylerde çeltik tarlalarında belirlenen 8 yabancı ot türünün bilimsel adı, familyası ve Türkçe adları Çizelge 4’de, bu yabancı otların rastlanma sıklığı ise Çizelge 5’de verilmiştir.
Çizelge 4. Sürvey çalışmasında belirlenen yabancı otlar
Yabancı ot türleri Familya Türkçe adı
Diplachne fusca (L.) Poaceae Baraj otu
Echinochloa crus-galli (L.) P.B. Poaceae Darıcan, cinek otu
Cyperus difformis L. Cyperaceae Kızotu
Echinochloa oryzoides (Ard.) Fritsch Poaceae Çeltiksi darıcan
Ammania coccinea Rottb. Lythraceae Güvercin ayağı
Lindernia dubia (L.) Pennell Scrophulariaceae Dip otu
Scirpus maritimus L. Cyperaceae Sivri saz
Paspalum paspalodes (Michx.) Schrib. Poaceae Su ayrığı
Çizelge 5 incelendiğinde, yapılan keşif sürveylerinde ele alınan 20 tarlanın tümünde D. fusca, E. crus-galli ve C. difformis’e rastlanmıştır. 8 tarlada E. oryzoides, 7 tarlada P. paspalodes, 4 tarlada A. baccifera, 3 tarlada L. dubia, 2 tarlada ise S. maritimus’ a rastlanmıştır.
Çizelge 5. Keşif sürveyinde belirlenen yabancı otların tarla bazında bulunma oranları
Yabancı ot türleri Bulunduğu tarla sayısı Rastlanma sıklığı %
D. fusca 20 100
E. crus-galli 20 100
C. difformis 20 100
E. oryzoides 8 40
P. paspalodes 7 35
A. baccifera 4 20
L. dubia 3 15
S. maritimus 2 10
Barajotu’na Karşı Yapılan Herbisit Denemesi Sonuçları
Edirne İli Uzunköprü İlçesinde, barajotunun iki farklı fenolojik dönemlerinde kurulmuştur. Bu denemelerden ilki barajotunun kardeşlenme dönemi öncesinde ve ikincisi de barajotunun kardeşlenme devresinde kurulmuştur.
Herbisit uygulamalarından sonra 4 defa değerlendirme yapılmış, her parselde ilaçların baraj otuna etkileri değerlendirilmiş ve sonuçlar Çizelge 6 ve Çizelge 7’te verilmiştir.
Barajotu’nun karşı kardeşlenme öncesi döneminde herbisit uygulama sonuçları
Barajotu’nun kardeşlenme öncesi dönemde uygulama denemesinde herbisit uygulaması 12.06.2008 tarihinde yapılmıştır. Denemede fenoxaprop-p-ethyl + izoxadifen ilacı 80 ml/da dozda, uygulamadan 18 ve 33 gün sonra yapılan değerlendirmelerde sırasıyla % 95 ve %98 etkili bulunmuştur.
Cyhalafop-buthyl 75 ml/da, uygulamadan 18 ve 33 gün sonra yapılan değerlendirmelerde sırasıyla %79 ve %82 yetersiz etkili bulunmuştur. Cyhalafop-buthyl 150 ml/da yapılan değerlendirmelerde sırasıyla %100 ve %100 (Çizelge 6) yeterli etkili bulunmuştur.

K. UZUN, H. DEMİRKAN
7
Çizelge 6. Barajotu’nun kardeşlenme öncesi döneminde yapılan uygulama sonuçları.
Herbisitler Tekerrür 18. gün
% etki
33. gün
% etki
1 95 97
2 97 99
3 93 98
4 95 98
Fenoxaprop-p-ethyl + isoxadifen-ethyl
80 ml/da
Ort. 95 a 98 a
1 77 84
2 81 80
3 78 83
4 80 81
Cyhalafop-butyl
75 ml/da
Ort. 79 b 82 b
1 100 100
2 100 100
3 100 100
4 100 100
Cyhalafop-butyl
150 ml/da
Ort. 100 a 100 a
Barajotu’nun kardeşlenme dönemi öncesinde herbisit uygulama denemesinde Şekil 1’de görüldüğü gibi kontrol parseldeki baraj otu yoğunluğuna karşın fenoxaprop-p-ethyl uygulanan parseldeki baraj otuna % 98’lik etki gözlemlenmiştir.
Şekil 1. Fenoxaprop-p-ethyl uygulanmış ve kontrol parsellerinin görünümü.
Barajotu’nun kardeşlenme döneminde herbisit uygulama sonuçları
Barajotu’nun kardeşlenme döneminde uygulama denemesinde herbisit uygulaması 19.06.2008 tarihinde yapılmıştır. Denemede fenoxaprop-p-ethyl + izoxadifen ilacı 80 ml/da dozda, uygulamadan 26 ve 45 gün sonra yapılan değerlendirmelerde sırasıyla %97 ve %98 yeterli etkili bulunmuştur.
Cyhalafop-buthyl 100 ml/da, uygulamadan 26 ve 45 gün sonra yapılan değerlendirmelerde sırasıyla %71 ve %73 yetersiz bulunmuştur.

DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
8
Cyhalafop-buthyl 150 ml/da, uygulamadan 26 ve 45 gün sonra yapılan değerlendirmelerde sırasıyla %100 ve %100 oranında etkili bulunmuştur. Sonuçlar Çizelge 7’de gösterilmektedir.
Çizelge 7. Barajotu’nun kardeşlenme devresinde yapılan uygulama sonuçları.
Herbisitler Tekerrür 26. gün % etki 45. gün % etki
1 97 97
2 98 99
3 96 98
4 97 98
Fenoxaprop-p-ethyl isoxadifen-ethyl
100 ml/da
Ort. 97 a 98 a
1 73 70
2 70 74
3 72 76
4 69 72
Cyhalafop-butyl
100 ml/da
Ort. 71 b 73 b
1 100 100
2 100 100
3 100 100
4 100 100
Cyhalafop-butyl
150 ml/da
Ort. 100 a 100 a
Baraj otu’nun kardeşlenme devresi sonrası uygulama alanında kontrol parseldeki baraj otu yoğunluğuna karşın; fenoxaprop-p-ethyl uygulanan parselde %97’lik etki ve cyhalafop-buthyl 150 ml/da uygulanan parselde de %100’lük etki Şekil 2’de görülmektedir. İlaçlı parsellerdeki etkilenmiş baraj otları sonrası açığa çıkan çeltik de daha net görülmektedir.
Şekil 2. Barajotunun kardeşlenme devresi sonrası uygulama alanındaki cyhalafop buthyl, kontrol ve fenoxaprop-p-ethyl parsellerinin karşılaştırmalı görünümü.
Fitotoksite gözleminde de deneme süresince yapılan tüm değerlendirmeler sırasında, herbisitlerin çeltiğe herhangi bir fitotoksik etkisinin olmadığı görülmüştür.

K. UZUN, H. DEMİRKAN
9
TARTIŞMA VE SONUÇ
Uzunköprü ilçesinde tesadüfen seçilen 20 tarlada yapılan keşif sürveyinde çeltik tarlalarından; 20 tarlada %100 rastlanma sıklığıyla Diplachne fusca, Echinochloa crus-galli ve Cyperus rotundus’a rastlanmıştır. 20 tarladan; sekizinde %40’lık rastlanma sıklığıyla Echinochloa oryzoides, yedi tarlada %35’lik rastlanma sıklığıyla Paspalum paspalodes’e, dört tarlada %20’lik rastlanma sıklığıyla Ammania baccifera’ya, üç tarlada %15’lik rastlanma sıklığıyla Lindernia dubia’ya ve iki tarlada %10’luk rastlanma sıklığıyla Scirpus maritimus’a rastlanmıştır. Özdemir (1992) yaptığı bir çalışmada, Marmara Bölgesi çeltik ekiliş alanlarında Echinochloa crus-galli (darıcan), Cyperus spp. (topalak), Scirpus spp. (sandalye sazı) ve Alisma plantago aquatica (kurbağa kaşığı)’yı deneme alanının dominant yabancı ot türleri olarak saptamıştır. Güneydoğu Anadolu Bölgesi’nde yapılan bir başka çalışmada 14 tür yabancı ot belirlenmiş, bunlardan Echinochloa spp., Cyperus difformis ve Cyperus fuscus’un biyolojisi, ekolojisi incelenmiştir (Uzun, 1983). Yapılan diğer çalışmalarda bu yabancı otlar önemli olarak bulunmuşlardır (Işık ve ark., 2000, Damar, 2006)
Çalışma alanında yaygın ve yoğun olarak bulunan barajotu kontrolü için denemeye alınan ilaçların dozlarına ve barajotunun fenolojik gelişme dönemlerine göre farklı sonuçlar elde edilmiştir.
Barajotu’nun kardeşlenme dönemi öncesinde yapılan uygulamada; cyhalafop – buthyl 75 ml/da dozun uygulandığı parsellerde, uygulamadan 18. ve 33. gün sonra yapılan değerlendirmelerde barajotuna sırasıyla ortalama %79 ve %82 oranlarında etkili bulunmuştur. 1–9 EPPO skalasına göre ilaçların yabancı otlara etkide kabul edilebilirlik sınırı olan %90’ın altında kalmış olduğundan 75 ml/da dozun barajotunun mücadelesinde önerilemeyeceği kanısına varılmıştır.
Cyhalafop – buthyl 150 ml/da dozun uygulandığı parsellerde, uygulamadan 18. ve 33. gün sonra yapılan değerlendirmelerde barajotuna sırasıyla ortalama %100 ve %100 oranlarında etkili bulunmuştur. 1–9 EPPO skalasına göre ilaçların yabancı otlara etkide kabul edilebilirlik sınırı olan %90’ın üzerinde ve tam etkili olduğundan 150 ml/da dozun barajotu’nun mücadelesinde önerilebileceği kanısına varılmıştır.
Fenoxaprop-p-ethyl + isoxadifen-ethyl 80 ml/da dozun uygulandığı parsellerde, uygulamadan 18. ve 33. gün sonra yapılan değerlendirmelerde barajotu’na sırasıyla ortalama %95 ve %98 etki ile yabancı otlara etkide kabul edilebilirlik sınırı olan %90’ın üzerinde etkili bulunmuş ve barajotu’nun mücadelesinde önerilebileceği kanısına varılmıştır.
Barajotu’nun kardeşlenme döneminde yapılan uygulamada; cyhalafop–buthyl 100 ml/da dozun uygulandığı parsellerde uygulamadan 26 ve 45 gün sonra yapılan değerlendirmelerde barajotu’na sırasıyla ortalama % 71 ve % 73 oranlarında etkili bulunmuştur. 1–9 EPPO skalasına göre ilaçların yabancı otlara etkide kabul edilebilirlik sınırı olan % 90’ın altında kalmış olduğundan 100 ml/da dozun barajotu’nun mücadelesinde önerilemeyeceği kanısına varılmıştır.
Cyhalafop–buthyl 150 ml/da denenen tüm dozları %100 etkili ve Fenoxaprop-p-ethyl + isoxadifen-ethyl 100 ml/da %97 ve %98 oranlarında etkili bulunmuştur.
Sera ve tarla koşullarında yapılan çalışmalarla, D. fusca’yı, cyhalofob-butyl (Gast, 1999, Sürek, 2002), clefoxydim (Fenley et al., 1999), thiobencarb (Smith, 1988) ve clomazone (Pegg et al., 1999) gibi herbisitlerin de kontrol ettiği ortaya konmuştur. Arkansas' da yapılan başka bir çalışmada, D. fusca’nın propanilin yalnız başına ve pendimethalin veya thiobencarb ile birlikte uygulanmasıyla kontrol edilebileceği sonucunu elde edilmiştir. Bunun yanında karışımsız veya karışım halinde propanil, thiobencarb veya pendimethalin ile uygulanan fenoxapropun da bu otu kontrol ettiği saptanmıştır (Khodayari et al.,(1989).
Sonuç olarak; Trakya bölgesinde, tuzlu topraklara ve zor koşullara uyum sağlayabilen; birkaç yılda tohumla ve sulama suyuyla hızla yayılarak yörede ciddi problem oluşturan barajotuna karşı çalışmalarımızda denediğimiz herbisitler, son yıllarda kullanılmaya başlanmıştır. Barajotu (D. fusca)’nun kardeşlenme dönemi öncesi uygulanan fenoxaprop-p-ethyl + isoxadifen-ethyl etkili maddeli ilacın dekara ilaç kullanımı (80 ml/da) ile diğer kullanılan herbisit dozlarına göre daha az olması ve baraj otunun çeltik bitkisi ile rekabete girmeden 18 günde baskı altına alınması ile etki süresinin kısa olmasından dolayı uygun bulunmuştur.

DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
10
Baraj otunun kardeşlenme döneminde uygulanan fenoxaprop-p-ethyl + izoxadifen-ethyl 100 ml/da dozun, baraj otunun kardeşlenme dönemi uygulamasındakine oranla fazla olmasından, bu sebeple maliyetin yüksek ve çeltiğin baraj otu ile rekabet süresinin 26 gün ile daha fazla olması sebebiyle herbisitlerin baraj otunun kardeşlenme dönemi öncesinde uygulanması daha uygun bulunmuştur.
Uzunköprü (Edirne) Çeltik Alanlarinda Yoğun Olarak Bulunan Yabancı Otların Belirlenmesi ve Kimyasal Savaşımı Üzerinde Araştırmalar
ÖZET
Ülkemizde çeltik tarımı yaygın olarak Edirne merkez, Uzunköprü, Meriç, İpsala, Keşan ve Enez ilçelerinde yapılmakta, bu bölge ülkemiz ekim alanının %25'ini, üretimin ise %28'ini karşılamaktadır. Çeltik alanlarında sorun oluşturan yabancı otlar; Echinochloa crus-galli (darıcan), E. oryzoides , Cyperus difformis (kızotu), Diplachne fusca (baraj otu), Polygonum lapatifolium, Paspalum paspaloides, Lindernia dubia ve Ammania coccinea (söğüt otu)’dır. Barajotu ülkemize son yıllarda giriş yapmış ve çok hızlı bir biçimde yayılmaya başlamıştır. Güçlü saçak kök yapısı ile çeltikle rekabet etmesi ve savaşımının zor olması nedeniyle bu çalışma 2008 yılında yapılmıştır. Deneme Uzunköprü’de, barajotunun iki farklı fenolojik gelişme döneminde (kardeşlenme öncesi ve kardeşlenme devresi) yapılmıştır. Denemeler, tesadüf blokları deneme desenine göre 4 karakterli ve dört tekerrürlü olarak kurulmuştur. Baraj otu’nun kardeşlenme öncesi döneminde; cyhalafop – buthyl 75 ml/da dozda 18. günde % 79, 33. günde % 82 etkili, 150 ml/da dozda ise 18. ve 33. günlerde %100 etkili bulunmuştur. Fenoxaprop-p-ethyl + izoxafen etkili maddeli herbisitin 80 ml/da dozunda 18. günde %95.33. günde ise %98 etkili olduğu belirlenmiştir. Barajotu’nun kardeşlenme devresinde cyhalafop – buthyl 100 ml/da dozda uygulandığında 26. günde %71, 45. günde %73 etkili, 150 ml/da dozda ise 26 ve 45. günlerde %100 etkili, Fenoxaprop-p-ethyl + izoxafen 100 ml/da dozda 26. günde %97.45. günde ise % 98 etkili bulunmuştur.
Anahtar kelimeler: Çeltik, Diplachne fusca, baraj otu, herbisit, Edirne
TEŞEKKÜR
Bu çalışma E.Ü.Bilimsel Araştırma Projeleri Komisyonu tarafından desteklenmiştir (Proje no: 2008 ZRF 011)
LITERATURE CITED
Akkoyunlu, Ş., 2005, Çeltik Tarlarında Yabancı Otlar, T.C. Samsun Valiliği, Tarım İl Müdürlüğü, No. B/24, Samsun, 2 s.
Anonymous., 1969. Türkiye’nin Tarımsal Üretim Projeksyonu (1968-200). T.C. Tarım Bakanlığı, Ankara, 1967
Anonymous, 2007, Tarım ve Köyişleri Bakanlığı, TR 2 Batı Marmara Bölgesi Tarım Master Planı, T. C. Tarım ve Köyişleri Bakanlığı Strateji Geliştirme Başkanlığı Yayınları, 125, Ankara 128 sf.
Beşer, N. 2009,“Trakya Tarımsal Araştırma Enstitüsü Müdürü Dr.Necmi Beşer: Türkiye, dünyada çeltikte yüksek verim alan ilk üç ülke arasında”, http://www.tarimmerkezi.com/haber_detay.php?hid=26000 (Erişim: 20 Ekim 2009)
Bora,T., İ.Karaca, 1970. Kültür Bitkilerinde Hastalığın ve Zararın Ölçülmesi. Ege Üniversitesi Ziraat Fak. Yardımcı Ders Kitabı, Yayın No.167 Bornova, 41s.
Bulut, İ., 2006, Genel Tarım Bilgileri ve Tarımın Coğrafi Esasları, (Ziraat Coğrafyası), Gündüz Eğitim ve Yayıncılık, Ankara.
Chang, T.T. 1970. Rice. In Genetic Resources in Plants – Their Exploration and Conservation (eds O.H. Frankel and E. Bennett), IBP Handbook, no. 11 pp. 267-72. Blackwell, Oxford and Edinburgh.

K. UZUN, H. DEMİRKAN
11
Damar, İ., 2006, Edirne İli Çeltik Alanlarında Bulunan Yabancı Ot Türleri ve Yoğunluklarının Belirlenmesi. Edirne, Yüksek Lisans Tezi.
Davis, P.H., 1985, Flora of Turkey, 9, Edinburgh Universty Pres, 724 p.
Demircan, A. K., 2006, Gönen Rehberi, Referans Yayınları, İstanbul.
Doğanay, H., 2007, Ekonomik Coğrafya 3 (Ziraat Coğrafyası), Aktif Yayınları, Erzurum.
Fenley, C, M. Landes, B. Sİevemİc'h, U. Missiitz, and U. Schöfl, 1999. BA5625 H- a new post-emergence herbicide for the control of grasse weeds in rice İn 1999 Bringhton crop protection conference: weeds. Proceeding of an international conference, Brightoni UK ,. 15-18 .November 1999.
Finansi, A. 1979. Rice and Food for Development. Fiat Trattori Edution, Torino, İtaly.P.
Gast R.E., 1999. Early watergrass (Echinocba oryzoides) and bearded sprangletop (Diplachne fusca) control with cyhalofob-buthyl in California water seeded rice. In 2nd temperate rice conference. Sacremento, California, USA.
Işık, D., H. Mennan ve O. Ecevit, 2000. Samsun İli Çeltik Ekim Alanlarında Görülen Yabancı Ot Türlerinin Belirlenmesi. Ondokuz Mayıs Üniversitesi, Ziraat Fakültesi Dergisi, 15 (3): 99-104.
Işık, D., H. Mennan, 2001. Çeltikte Darıcan (Echinochloa crus-galli (L.) P. Beauv), Kurbağa Kaşığı (Alisma plantago aquatica L.) ve Sandalye Sazının (Scirpus mucranatus Pollich) Rekabet Yeteneklerinin Araştırılması. Türkiye Herboloji Dergisi, 4 (2),47-57.
Khodayari, K., P. Nastasi, and RJJr., Smith, 1989. Fenoxaprop im grass control in dry seeded rice (Oryza sativa). Weed Technoi, 3:131-135.
Kün, E., 1985, Sıcak İklim Tahılları, A.Ü. Ziraat Fakültesi yayın No: 953, Ankara
Özdemir, C., 1992, Marmara Bölgesinde Çeltikte Sorun Olan Yabancı Otlara Karşı İlaç Denemesi, Zir. Müd. Araş. Yıl., s.219
Özşahin, E., 2008, Gönen Ovasında Pirinç Tarımı, Fırat Üniversitesi Sosyal Bilimler Dergisi Cilt: 1, Sayı: 2, Sayfa: 49-70, Elazığ.
Pegg, I.R., P.V. Grassick, and M.C, Taylor, 1999. Clomazone- a new herbicide for grosse control in water-seeded rice. İn 2nd temperate rice conference. Sacremento, California, USA.
Smith, R. J. Jr., W. T. Flinchum, and D. E. Seaman. 1977. Weed control in U. S. Rice production. U. S. Dep. Agric. Handb. 497. U. S. Gov. Printing Office, Washington, D. C. 78 p.
Smith, R.J.Jr, 1983. Weed control in water-and dry -seeded rice (Oryza sativa). Weed Techn. 2:142-250.
Smith, R.J.Jr, 1988. Assessment of allelopathic potential of barnyard grass (Echinochloa crus-galli) on rice (Oryza sativa L.) cultivars Crop Protection, Volume 20, Issue 10, December 2001, Pages 921-928
Sürek, H., 2002, Çeltik Tarımı. Hasad Yayıncılık Ltd. Şti. İstanbul
Uzun, A., 1983, Güneydoğu Anadolu Bölgesindeki Çeltik Alanlarındaki Bazı Darıcan (Echinochloa spp.) ve Bazı Tek Yıllık Topalak (Cyperus spp.) Türleri Üzerinde Araştırmalar, Doktora çalışması, Diyarbakır, s. 97

DETERMINATION OF WEEDS IN RICE REGION OF EDIRNE - UZUNKÖPRÜ AND RESEARCHES ON CHEMICAL CONTROL OF THOSE WEEDS
12

J. Turk. Phytopath., Vol. 42 No. 1-3, 13-27, 2013 ISSN 0378 - 8024
13
Studies on Esca and Petri Diseases in Grapevine Nurseries and Vineyards in Aegean Region
Dilek POYRAZ * Ersin ONOĞUR **
* Bornova Zirai Mücadele Araştırma İstasyonu, İzmir/ TÜRKİYE, [email protected] ** Ege Üniversitesi Ziraat Fakültesi Bitki Koruma Bölümü, İzmir/ TÜRKİYE Accepted for publication March 5, 2014
ABSTRACT
This study was carried out between the years of 2009-2012 in order to determine fungal agents that cause Esca and Petri diseases in grapevine nurseries and in vineyards of Manisa, Denizli and Izmir provinces of Aegean Region. It was planned to investigate the role and share of pathogens in complex disease symptoms.
The trunk, cane and leaf samples were taken from the grapevine nurseries, young and older vineyards. The isolations from nurseries revealed the presence of Phaeoacremonium aleophilum (Pm) and Phaeomoniella chlamydospora (Pcl) as causal agents of Petri disease, based on their morphological characteristics. The samples from young vineyards were infected primarily with Pm, Pcl and secondly with Fomitiporia mediterrenea (Fom), known as Esca pathogen, whereas the isolations from older vineyards indicated primarily the presence of Fom followed with Pm and Pcl.
In conclusion, the presence of the pathogens Pmand Pcl causing Petri disease in the samples taken from grapevine nurseries was revealed and it was also found that these pathogens were dominant in the samples taken from young vineyards accompanied with Fom, causing Esca symptoms. The vines in the older (10-25 years old) vineyards were however infected mainly with Fom and additionally with Pmand Pcl.
Key words: Grapevine, Petri disease, Esca, Phaeoacremoniumaleophilum, Phaeomoniella chlamydospora, Fomitiporia mediterrenea.
GİRİŞ
Türkiye, bağcılık için dünyanın en elverişli iklim koşullarına sahip ülkelerinden birisidir. Asmanın (Vitis vinifera L.) gen merkezi olmasının yanı sıra çok eski tarihlere dayanan bir bağcılık kültürüne sahip olan Anadolu bağcılığının kökeni M.Ö. 2300 yıllarına dayanmaktadır (Çelik ve ark., 1998).
Türkiye İstatistik Kurumunun 2010 verilerine göre ülkemiz, dünya ülkeleri arasında, bağ alanı yönünden 4.sırada (477.786 ha), üzüm üretimi yönünden ise 6. (4.255.000 ton) sıradadır. Ayrıca Türkiye dünyada en büyük çekirdeksiz kuru üzüm üreticisi ve ihracatçısı konumunda olup, dünya çekirdeksiz kuru üzüm ihracatının %40-45’ini gerçekleştirmektedir. Özetle bağcılık ülke ekonomisi için önemli bir gelir kaynağıdır.
Türkiye’de üzüm yetiştiriciliği bölgelere göre değerlendirildiğinde, Ege Bölgesi’nin gerek alan (1.392.082 da) ve gerekse üretim (1.952.356 ton) açısından ilk sırada yer almaktadır. Bu bölgede Manisa ilinin hem alan (715.895 da) hem de üzüm üretimi (1.372.571 ton) yönünden ilk sırada yer aldığı görülmektedir (Anonim, 2010).
Türkiye’de üzümün yetiştirme sürecinde ortaya çıkan sorunların başında hastalık ve zararlılar gelmektedir. Hastalıkları, önemleri ve yaygınlıklarına göre Külleme (Erysiphe necator Schwein.), Mildiyö (Plasmopara viticola Berl. & De Toni.), Ölükol (Phomopsis viticolaSacc.), Kurşuni Küf (Botrytis cinerea Pers.), Kav (Stereum

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
14
hirsutumPers., PhellinusigniariusQuél., Phaeoacremonium aleophilum W. Gams, Crous, M.J. Wingf. & Mugnai., Phaeomoniella chlamydospora Crous & W. Gams), Antraknoz (Elsinoe ampelina Shear.) ve Eutypa (Eutypa lata Tul. & C. Tul) şeklinde sıralamak mümkündür (Anonim, 2008).
İzmir ve Manisa Ticaret Borsası, Ege İhracatçılar Birliği ve Manisa Bağcılık Araştırma İstasyonu temsilcilerinden oluşan bir komisyonu 18.07-04.08.2011 tarihleri arasında Manisa, İzmir ve Denizli il ve ilçelerini gezmiştir. Hazırladıkları “2011-2012 Ege Bölgesi Çekirdeksiz Kuru Üzüm Rekolte Tahmin Raporu” nda tüm bölge bağlarında Kav Hastalığının yayılma eğiliminde olduğu, ciddi sorunlara yol açtığı görüşüne yer verilmiş, ancak bu raporda yalnızca “Kav” hastalığından söz edilmiş, “Petri” hastalığının varlığından söz edilmemiştir.
Bornova Zirai Mücadele Araştırma İstasyonu Müdürlüğüne, 2007 yılından günümüze (2012) kadar olan süre içerisinde Ege Bölgesi’ndeki il ve ilçelerden gelen şikâyetler, örnekler ve arazi kontrolleri değerlendirilmiştir. Yaşlı bağlarda Kav hastalığı yanında yeni tesis edilmiş genç bağlarda da benzer hastalık belirtilerinin var olduğu,10 yaşın üzerindeki bağlarda sorun olduğu bilinen “Kav Hastalığı”nın genç bağlarda da şikâyet konusu olduğu saptanmıştır. Bu şikâyet ve bulguların gittikçe artan bir şekilde dile getirilmesi sonucu bu iki hastalık üzerinde ayırt edici, kapsamlı bir araştırma yapma gereksinimi doğmuştur.
“Esca” olarak da bilinen “Kav Hastalığı” Türkiye’de ilk defa P. Viala tarafından 1926 yılında İzmir (Smyrna) bağlarında saptanmış ve özellikle yaşlı bağlarda sorun olan, birkaç fungal etmenin birlikte neden olduğu bir hastalık olarak kabul edilmiştir. İlk kez İyriboz (1942), Kav hastalığının Ege Bölgesindeki varlığını ve hastalığa Stereum hirsitum ve Phellinus (Fomes) igniarius fungal etmenlerinin neden olduğunu bildirmiştir. Daha sonra Üzümeri (1947), yaşlı bağların zarar gören odun dokusunda Stereum necator, S. hirsitum, Polyporus igniarius ve P. versicolor adlı fungal etmenleri saptamıştır.
Ege Bölgesi bağlarında Kav hastalığı ile ilgili en son çalışma Erkan ve Larignon (1998) tarafından gerçekleştirilmiş, Manisa ve İzmir bağlarından Kav hastalığının tipik belirtilerini taşıyan örneklerin farklı kısımlarından yapılan izolasyonlar sonucunda Stereum hirsitum, Phelllinus sp., Phaeoacremonium aleophilumve Phaeoacremonium chlamydosporum saptanmıştır. Son iki fungusun Türkiye’deki varlığı ilk olarak bu araştırma ile ortaya konmuş, ancak bu etmenler Kav hastalığı ile ilişkilendirilmiş, Petri hastalığına yol açabildiklerine değinilmemiştir (Erkan 2000). Bu araştırmanın sonucu olarak, Zirai Mücadele Teknik Talimatlarında Kav Hastalığı’na neden olan etmenler arasında Phaeoacremonium aleophilumve Phaeoacremonium chlamydosporum da yer almıştır.
İtalyan bitki patoloğu Lionello Petri tarafından 1912 yılında, İtalya’da yapılan çalışmada,yapraklarında Kav belirtilerini taşıyan asmaların odun dokusundan yapılan izolasyonlar sonucunda Phaeoacremonium ve Phaeomoniella türleri izole edilmiş ve bu etmenlerin yol açtığı belirti tablosu, Kav’dan ayrı bir hastalık olarak, araştırıcının adına izafeten, “Petri Hastalığı” olarak tarif edilmiştir (Penn 2001).
Son yıllarda yurt dışında yapılan birçok çalışmada, Phaeoacremonium ve Phaeomoniella türleri Kav hastalığının tipik yaprak belirtilerini taşıyan veya gösteren genç asmaların odun dokusundan izole edilmiş ve bu etmenlerin yaşlı omcalardaki Kav hastalığının etiolojisi içinde yer alıp almadıkları araştırılmıştır. Fidanlıklardan alınan örnekler üzerinde sürdürülen çalışmalarda, Phaeoacremonium ve Phaeomoniella türlerinin varlığı moleküler tanı yöntemleriyle ortaya konmuş ve sonuçta bu etmenlerin Kav’dan farklı olarak, Petri hastalığına yol açtıkları ortaya konmuştur. Bu çalışmalarda, Petri hastalığının fidanlıkta ve bölgeler arasında yayılmasında bulaşık anaç ve çeliklerin ve dolayısıyla fidanların büyük rol oynadığı ileri sürülmüştür (Scheck et al. 1998, Stamp 2001, Retief et al. 2006, Aroca et al. 2006, Gimenez-Jaime et al. 2006).
Türkiye’de yapılan çalışmalar da incelendiğinde, 1947-1998 yılları arasında ve 1998 yılından günümüze kadar olan süreçte Petri hastalığından söz edilen yayının bulunmadığı dikkati çekmiştir.
Bu araştırmada, Ege Bölgesi, Manisa, Denizli ve İzmir illerindeki özel ve resmi fidanlıklarda, üreticiye ait genç ve yaşlı bağlarda Petri ve Kav hastalıklarının varlığı, bu hastalığa yol açan etmenlerin tanısı yapılmış ve Petri hastalığının kaynağı ve etmenlerin dokuyu işgal süreçlerine ilişkin bulgular sunulmaktadır (Poyraz 2012).

D. POYRAZ, E. ONOĞUR
15
Bu çalışma, T.C. Gıda Tarım ve Hayvancılık Bakanlığı Tarımsal Araştırmalar ve Politikalar Genel Müdürlüğü tarafından TAGEM-BS-09/04-1/02-12 no’lu proje olarak desteklenmiş ve Bornova Zirai Mücadele Araştırma İstasyonu’nda E.Ü. Fen Bilimleri Enstitüsü’ne sunulan doktora tez projesi kapsamında yürütülmüştür.
MATERYAL VE METOT
MATERYAL
Ege Bölgesinde bağ yetiştiriciliğinin yoğun olarak yapıldığı Manisa, Denizli ve İzmir illeri çalışma alanı olarak seçilmiş ve bu illerdeki asma fidanlıklardan alınan örnekler yanında genç ve yaşlı asmalardan alınan örnekler ve bu örneklerden elde edilen fungal çalışmanın ana materyalini oluşturmuştur.
METOT
Fidanlıklardan materyal temini
İl Gıda, Tarım ve Hayvancılık Müdürlükleri’ne kayıtlı olmak koşuluyla; Çizelge 1’de belirtildiği gibi 4-6.05.2009 tarihlerinde Manisa ilinden 10 ve Denizli ilinden 2 fidanlıktan olmak üzere toplam 12 fidanlıktan 10’ar tane aşılı tüplü asma fidanı alınmıştır. Bu örnekler bölgede en çok fidan yetiştiriciliğinin yapıldığı Yuvarlak Çekirdeksiz, Alphonse Lavallée, Superior Seedless, Merlot ve Cabernet Sauvignon’dan oluşmuştur. Örnekler laboratuvara getirildiğinde enine ve boyuna kesitleri alınmış, kesitler fotoğraf makinası ile görüntülenmiş ve örnekler izolasyon çalışmalarında kullanmak üzere +4oC’de saklanmıştır. Kayıtlı fidanlık bulunamadığından İzmir ilinden asma fidanı örneği alınamamıştır.
Çizelge 1. İllere göre asma fidanı örneklerinin alındığı fidanlık sayısı ve kodları ile bu fidanlıklardan alınan asma çeşitleri ve kodları.
İl Fidanlık sayısı Fidanlık kodları Asma çeşitleri
Manisa 10 A-B- C- D- E-F- G- H- I- J
Denizli 2 K- L
1: Yuvarlak Çekirdeksiz
2: Alphonse Lavallée
3: Superior Seedless
4: Merlot
5: CabernetSauvignon
Genç ve yaşlı bağların asmalarından materyal temini
Ege Bölgesi’nde en çok yetiştiriciliği yapılan Yuvarlak Çekirdeksiz üzüm çeşidiyle kurulmuş, 10 yaşından küçük (genç) ve 10-25 yaşında (yaşlı), büyüklüğü 10 da’dan küçük olmayan bağların hastalığın tipik yaprak belirtilerini gösteren asmalarından gövde, en az 2 yaşlı sürgün ve yaprak örnekleri alınmıştır. Örnekleme yapılan ilçeler, örnek sayıları ve örnekleme tarihlerine Çizelge 2’de yer verilmiştir. Fidan örneklerinde olduğu gibi, bu örneklerde laboratuara getirilmiş ve bunlara aynı işlemler uygulanmıştır.
Çizelge 2. İl ve İlçelere göre genç ve yaşlı bağlardan alınan örnek sayıları ve örnekleme tarihleri.
Örnek sayısı Toplam Örnekleme tarihleri İl İlçe
Genç bağ Yaşlı bağ Manisa Alaşehir
Salihli Saruhanlı Merkez Turgutlu Sarıgöl
2 2 2 1 1 2
2 2 2 1 1 2
20
05-06-07. 08.2009
Denizli Buldan Çal Sarayköy
2 2 2
2 2 2
12 14.08.2009
İzmir Menemen Kemalpaşa
3 2
3 2
10 30-31.07.2009
Toplam 21 21 42

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
16
İzolasyon çalışmaları ve tanı
Laboratuvara getirilen fidan ve asmaların ana gövde ve en az 2 yaşındaki sürgün örneklerinden enine ve boyuna kesitler alınmış ve bu kesitlerden belirti görülen kısımları da kapsayacak şekilde yaklaşık 5 mm büyüklüğünde küçük parçalar çıkarılmıştır. Bu parçalar önce %70’lik etil alkol içerisinde 30 sn, sonrasında %3’lük kalsiyum hipoklorit içerisinde 15 sn bekletilmiş ve arkasından steril kurutma kağıtları üzerine alınmışlardır. İzolasyon streptomisin sülfat ilaveli (100mg/l) malt extrakt agar (MEA), potatodextrose agar (PDA) ve rose bengal agar (RBA) içeren petrilerde gerçekleştirilmiş, petriler 20-25oC’de karanlıkta 14 gün inkubasyona bırakılmışlardır. Gelişen kültürler tüplere aktarılarak daha sonraki çalışmalarda kullanmak üzere +4oC’de saklanmışlardır (Erkan ve Larignon 1998).
İzolasyon çalışmaları sonucu elde edilen fungal etmenlerin morfolojik ve mikroskobik özelliklerine göre tanıları Kaliforniya Üniversitesi’nden Dr. Akif Eskalen ve Dr. Jose Ramon Urbez-Torres tarafından gönderilmiş referans izolatlar (Pa/A2:Phaeoacremonium aleophilum, Pc/Pc45: Phaeomoniella chlamydospora, Fm: Fomitiporia mediterrenea) yardımıyla ve literatürle karşılaştırılarak yapılmıştır.
SONUÇLAR VE TARTIŞMA
İzole edilen etmenlerin morfolojik özellikleri
Asma fidanları, genç ve yaşlı bağların asmalarından alınan örneklerden yapılan izolasyonlar sonucu elde edilen etmenlerin makroskobik ve mikroskobik özelliklerine göre tanılarıyla ilgili bulgular aşağıda verilmiştir.
Phaeoacremonium aleophilum(Pm):
Koloniler ortama yapışık, grimsi kahverengi (devetüyü) renkte, gelişme yavaştır (Şekil 1-A). Hifler; bölmeli ve dallı, açık kahverengi renkte, 1,5-4 µm genişliğinde. Konidiler; elips dikdörtgen şeklinde, kenarları oval ve 3-5,5 x 1,5-2 µm boyutlarında, renksiz (Şekil 1-B), klamidospor yoktur. Pm’un koloni gelişimi ve mikroskobik özellikleri literatürle aynı olmakla birlikte konidilerin boyutları Mostert et al. (2006b) ve Fischer and Kassemeyer (2003) tarafından 3-5,5 x 1,5-2 µm olarak bildirilmiştir.
Şekil 1. Phaeoacremonium aleophilum’un koloni gelişimi (MEA)ve mikroskobikgörüntüsü (100x büyütme).
Phaeomoniella chlamydospora (Pcl)
Koloniler ortama yapışık, yüzeysel ve seyrek, koyu zeytin yeşili renkli, gelişme yavaştır (Şekil 2-A). Hifler; bölmeli ve dallı, yeşil renkli, 2-5 µm genişliğindedir. Konidiler; elips-dikdörtgen şeklinde, kenarları oval ve 2,5-5 x 1,5-2 µm boyutlarında, renksiz olup (Şekil 2-B), klamidospor nadiren oluşur. Pcl’nın koloni gelişimi ve mikroskobik özellikleri literatürle aynı olmakla birlikte konidilerin boyutları Crous and Gams (2000) ve Fischer and Kassemeyer (2003) tarafından 1,5-5 x 1-2 µm olarak bildirilmiştir.

D. POYRAZ, E. ONOĞUR
17
Şekil 2. Phaeomoniella chlamydospora’nınkoloni gelişimi (MEA)vemikroskobik görüntüsü (100x büyütme).
Fomitiporia mediterrenea(Fom):
Koloniler pamuksu, miselyum havai, turuncu-kahverengi renkte, diğer etmenlere göre daha hızlı gelişmedir (Şekil 3-A).Hifler; bölmeli ve dallı, turuncu-kahverengi renkte, 1,6-4 µm genişliğinde, konidi yoktur (Şekil 3-B). Fom’nin koloni morfolojisi literatürle aynı olmakla birlikte hiflerin genişliği Fischer (2002) ve Fischer and Kassemeyer (2003) tarafından 1,5-5,5 µm olarak bildirilmiştir.
Şekil 3. Fomitioporia mediterrenea’nın koloni gelişimi ve mikroskobik görüntüsü (40x büyütme).
Asma fidanlarının gövdelerindeki hastalık belirtileri ve etmenler
Bölgede en çok fidan üretimi yapılan Yuvarlak Çekirdeksiz, Alphonse Lavaillée, Superior Seedless, Merlot, Cabernet Sauvignon fidanlıklarından alınan örnekler üzerinde yapılan makroskobik gözlemlerde herhangi bir belirti gözlenmemiştir. Ancak bu örneklerden hazırlanan enine ve boyuna kesitlerde tipik belirtiler saptanmış, bu belirtiler Şekil 5’de görüntülenmiş ve tarif edilmişlerdir.
Fidanların enine kesitlerinde ilk anda ksilem dokularında tylosis oluşumuna işaret eden siyah noktacıklar (a) görülmüş ve birkaç dakika içinde bu siyah noktacıklardan boncuk şeklinde koyu amber-siyah renginde sıvı (b) çıkışı olmuştur. Enine kesitte siyah noktacıklara sahip kısımlardan boyuna kesitler alındığında bu noktacıkların ksilem dokusu boyunca çizgisel, siyah nekrozlar (c) şeklinde oluştuğu görülmüştür (Şekil 5).Tarif edilen belirtileri taşıyan kısımlardan yapılan izolasyon sonuçları, Çizelge 3’te verilmiştir.Örnek alınan çeşitler arasında bu belirtiler yönünden herhangi bir farklılık gözlenmemiştir.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
18
Şekil 5. Asma fidanlarının enine ve boyuna kesitlerindeki hastalık belirtileri.
Çizelge 3. Asma fidanlarının gövdelerinden hastalık belirtilerine göre izolasyon sonucunda elde edilen fungal etmenler.
Belirti Tipleri Fungal Etmenler
a b c
Phaeomoniella chlamydospora (Pcl) + + +
Phaeoacremonium aleophilum(Pm) + + +
Fomitiporia mediterrenea (Fom) - - -
İzolasyon sonucunda asma fidanlarından Petri hastalığına neden olan etmenler Pm ve Pcl elde edilmiştir. Asma fidanlarındaki bu belirtiler ve tarafımızdan yapılan belirti tarifleri, literatürle paralellik göstermektedir (Mugnai et al.1999,Surico 2001, Penn 2001, Mostert et al. 2006a, Surico et al. 2006, White 2010). Bu belirtiler Petri hastalığının tipik belirtileridir.
Diğer ülkelerde yapılan pek çok çalışmada da asma fidanlarında Petri hastalığı saptandığı ve hastalığın bölgeler arasında yayılmasında bulaşık aşı kalemi, çelik gibi üretim materyalinin, fidanların büyük rol oynadığı bildirilmektedir (Scheck et al. 1998, Stamp 2001, Retief et al. 2006, Aroca et al. 2006, Gimenez-Jaime et al. 2006). Bu da, Petri hastalığına neden olan etmenlerin üretim materyalinin temin edildiği 1-2 yıllık sürgünlere kadar ulaştığını, ancak Kav hastalığı etmenlerinin ana gövdede kaldığını göstermektedir.
Genç asmaların gövdelerindeki hastalık belirtileri ve etmenler
Yuvarlak Çekirdeksiz üzüm çeşidine ait 10 yaşından genç asmalardan alınan ana gövde örneklerinde hazırlanan enine ve boyuna kesitlerde ortaya çıkan belirtiler Şekil 6’daki gibi görüntülenmiştir.

D. POYRAZ, E. ONOĞUR
19
Bu kesitlerde de, fidanlardaki belirtilerde olduğu gibi, enfekteli ksilem demetlerinde önce siyah renkli noktacıklar görülmüş ve bunlardan birkaç dakika içinde boncuk şeklinde koyu amber-siyah renginde sıvı çıkışı olmuş (a), boyuna kesitlerde ise bu noktacıkların ksilemde çizgisel siyah nekrozlar şeklinde uzandığı görülmüştür (b). Bazı asmalarda ise, öz kısmının etrafındaki ksilem dokusunda beyaz çürüklük şeklinde açık renkli, yumuşak dokulu kavlamış bir kısmın varlığı (c) ve bu kısmın koyu renkli, sert dokulu bir kuşak ile (d) çevrilmiş olduğu belirlenmiştir. Bu belirtilere ek olarak, yine ksilem dokusunda koyu renkli, nekrotik alanlar (e) da görülmüştür (Şekil 7). Bu 5 ayrı belirti tipinden yapılan izolasyon sonuçları Çizelge 4’de verilmiştir.
Çizelge 4. Genç asmaların gövdelerinde belirlenen farklı belirti tiplerinden yapılan izolasyonlar sonucunda elde edilen fungal etmenler.
Belirti Tipleri Fungal Etmenler
a b c d e
Phaeomoniella chlamydospora (Pcl) + + - - -
Phaeoacremonium aleophilum (Pm) + + - + +
Fomitiporia mediterrenea (Fom) - - + + -
Şekil 6. Genç asmaların ana gövdelerinden hazırlanan enine ve boyuna kesitlerindehastalık belirtileri.
Görüldüğü gibi genç asmalarda, Petri hastalığına neden olan etmenlerin (Pcl, Pm) yanı sıra Kav hastalığına neden olan etmenlerden Fom‘da izole edilmiştir. Bu veri Literatürde Kav hastalığına yol açtığı kayıtlı diğer 2 etmen bu örneklerden izole edilmemiştir.
Genç asmalardaki bu belirtiler ve tarafımızdan yapılan belirti tarifleri, literatürle paralellik göstermektedir (Morton, 1999, Larignon, 1999, Surico, 2001, Penn, 2001, Surico et al. 2006, Luque et al. 2009, White, 2010, Andolfi et al., 2011, Kuntzmann, 2012, Lecomte et al. 2012).
Yaşlı asmaların gövdelerindeki hastalık belirtileri ve etmenler
Yuvarlak Çekirdeksiz üzüm çeşidine ait 10-25 yaşlı asmalardan alınan ana gövde örneklerinden hazırlanan enine ve boyuna kesitlerde saptanan belirtiler Şekil 7’de yansıtılmıştır.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
20
Şekil 7.Yaşlı asmalarda ana gövdenin enine ve boyuna kesitlerindeki hastalık belirtileri
Bu örneklerin enine kesitlerinde, öz kısmı ile birlikte ksilem dokusunda yumuşak dokulu, kavlamış, açık renkli, bir beyaz çürüklük (a), bu kısmın çevresinde ise koyu renkli, sert dokulu bir kuşak halkasının varlığı saptanmıştır(b). Bu belirtiler yanında, yine ksilem dokusunda koyu renkli nekrotik alanlar (c) görülmüştür. Tarif edilen bu kısımlardan yapılan izolasyon sonuçları Çizelge 5’te yer almıştır.
Çizelge 5. Yaşlı asmaların gövdelerinde belirlenen farklı belirti tiplerinden yapılan izolasyonlar sonucunda elde edilen fungal etmenler.
Belirti Tipleri Fungal Etmenler
a b c
Phaeomoniella chlamydospora(Pcl) - - +
Phaeoacremonium aleophilum(Pm) - + +
Fomitiporia mediterrenea(Fom) + + -
Görüldüğü gibi, yaşlı asmalardan Petri hastalığına neden olan etmenler (Pcl, Pm) yanında Kav hastalığına neden olan etmenlerden Fom izole edilmiştir. Bu bulgulara göre, Pcl “c” tipi belirti vererek, Pm ise bu belirtiye ek olarak “b” tipi belirtiye yol açarak asmanın ksilem dokusuna yerleşmekte, Fom ise “a” ve “b” tipi belirtilere iştirak ederek ksilem dokusuna ve öz kısmına yerleşmektedir.
Yaşlı asmalardaki bu belirtiler ve tarafımızdan yapılan belirti tarifleri, literatürle paralellik göstermektedir (Karaca, 1965, Onoğur, 1995, Mugnai et al., 1999, Morton, 1999, Larignon, 1999, Surico, 2001, Penn, 2001, White, 2010).

D. POYRAZ, E. ONOĞUR
21
Ege Bölgesi bağlarında Erkan ve Larignon (1998) tarafından yapılan çalışmada, Kav hastalığının yaprak belirtilerini taşıyan yaşlı asmalardan Ste, Ph, Pm ve Pcl etmenleri saptanmış ve son iki etmen yapraktaki belirtilerin benzerliği nedeniyle Kav hastalığı ile ilişkilendirilmiştir. Ancak bu yayında Petri hastalığına değinilmemiştir.
Bu çalışmada ise, genç bağlardan alınan örneklerde ağırlıklı olarak Petri hastalığına neden olan etmenlerden Pm ve Pcl saptanırken, yaşlı bağ örneklerinde ise ağırlıklı olarak Kav hastalığı etmenlerinden Fom saptanmıştır. Erkan and Larignon’un (1998) yaşlı bağlarda saptadıkları Ste ve Ph etmenleri kendi çalışmamızda saptanmamış ve buna göre Ste’un bölgemizdeki Kav tablosunda rolünün olmadığı düşünülmüştür. Ph etmeninin ise bazı çalışmalarda isimlendirmesinin hatalı olduğu, bu etmenin aslında Fom olduğu, bazı yayınlarda ise bunun tam net olarak belirtilmediği görülmektedir (Fischer 2002). Nitekim son yapılan çalışmalarda, Şekil 6, 8 ve 10’da gösterilen belirtileri taşıyan genç ve yaşlı asmalarda Fom, Pm ve Pcl’nin varlığından söz edilirken (Surico 2000, Surico 2009, White, 2010) Ph etmeninin adının geçmediği dikkati çekmektedir.
Genç ve yaşlı bağların asmalarında yaprak ve tane belirtileri
Genç ve yaşlı bağlardan Temmuz-Ağustos aylarında yapılan örnek alımı sırasında yapraklarda tipik belirtiler görülürken, nadiren bazı asmaların üzüm tanelerinde hastalık belirtileri görülmüştür. Bu belirtiler aşağıda verilmiş ve tarif edilmişir.
Bu asmaların yapraklarında, Kav hastalığının bilinen belirtilerine uygun olarak; damar aralarında önce klorotik lekeler belirmekte, daha sonra bu lekeler birleşip, sarımsı-kahverengi veya kızıl- kahverengi bir renk kazanarak nekroze olmakta (Şekil 8) ve bu yapraklar ağır su kaybı sonucu özellikle sıcak yaz günlerinde aniden kurumaktadırlar (Şekil 9).
Şekil 8.Genç ve yaşlı asmaların yapraklarında belirti gelişimi.
Şekil 9.Yaşlı asmalarda yaz aylarında ani kurumalar.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
22
Bu belirtiler, Karaca (1965), Onoğur (1995), Mugnai et al. (1999), Morton (1999), Larignon (1999), Surico et al. (2006) ve White (2010) tarafından tarif edilen yaşlı asmalardaki Petri ve Kav hastalığının yaprak belirtileriile paralellik göstermekte, onlara uymaktadır.
Yaprak belirtileri taşıyan bazı omcaların salkımlarındaki üzüm tanelerinin yüzeyinde, koyu mor, nokta şeklinde lekeler görülmüş ve bu lekelerin zamanla salkımın tüm tanelerini kapladığı kaydedilmiştir (Şekil 10). Bu tanelerin tadına bakıldığında bunların daha tatlı oldukları, ancak tanenin kabuk kısmının buruk bir tada sahip olduğu hissedilmiştir.
Şekil 10. Üzüm tanelerinde hastalık belirtileri.
Üzüm tanelerindeki bu belirtiler de literatürde açıklanan, tarif edilen belirtilere uymaktadır (Karaca 1965, Mugnai et al., 1999, Rooney-Latham et al., 2005, Eskalen et al., 2005, Vasquez, 2012, White, 2010).
Görüldüğü gibi bu iki hastalığın yapraktaki ve üzüm tanelerindeki belirtileri benzemekte ve kanımıza göre bu özellik Petri hastalığının gözden kaçmasına neden olmakta ve ayırım ancak odun dokusunun incelenmesi sonucu yapılabilmektedir. Bu belirti benzerliği ve etmenlerin bir arada olması nedeniyle, araştırıcılar arasında Esca “bir hastalık kompleksi mi?”, yoksa “hastalıkların kompleksi mi?” diye iki teori üzerinde farklı görüşler ortaya konmuştur. İlk başlarda, birkaç fungal etmenin neden olduğu ve diğer faktörlerin de bir arada olabildiği bir hastalık kompleksi olarak düşünülmüş, ancak daha sonraki çalışmalarla en az iki hastalığın kompleksi olduğu kabul edilmiştir. Böylece “Esca” Kav ve Petri hastalığının bir arada olabildiği bir hastalık kompleksi olarak kabul edilmiştir (Larignon and Dubos 1997, Graniti et al., 2000, Surico, 2001, Surico, 2009, White, 2010). Bu nedenle “Esca”, Kav ve Petri hastalıklarının birlikte anıldığı bir hastalık olarak literatürde yer almaktadır.
Petri ve Kav hastalığı etmenlerinin asmaya yerleşme sıraları
Larignon (1999), asma gövdesinin enine kesitini gösteren şemada (Şekil 11), hastalık belirtilerini ve bu belirtilerden elde edilen etmenleri göstermiştir. Buna göre, asma ksilem dokusunda önce siyah noktacıklar şeklinde belirtiler ortaya çıkmakta, sonra bu noktacıkların birleşmesiyle siyah çizgiler belirmekte ve bu belirtilere Pcl etmeninin neden olmaktadır. Daha sonra ksilemden öz dokusuna kadar ilerleyen üçgen şeklinde koyu renkli nekrozlar ortaya çıkmakta, nekrozlar zamanla daire şeklini aldığı ve bu belirtilerden Pcl ile birlikte Pm de izole edilmektedir.Daha sonraki şekillerde ise dokuda koyurenk ile gösterilmiş beyaz bir çürüklüğün meydana geldiği bulgusuna yer verilmekte ve bu belirtiye Fom’un yol açtığı gösterilmektedir.
Bu çalışmada elde ettiğimiz Petri ve Kav hastalığı etmenlerinin (Çizelge 5) ne zaman ve hangi sırayla asmaya yerleştikleri, gerek elde ettiğimiz bulgular ve gerekse Surico (2001)’nun verdiği şemanın tarafımızdan modifiye edilmiş şekli yardımıyla Şekil 12’de açıklanmıştır.

D. POYRAZ, E. ONOĞUR
23
Şekil 11. Asmaların gövdelerinin enine kesitindeki Petri ve Kav hastalığına neden olan etmenlerin oluşturdukları belirti tiplerinin şematik
görüntüsü (Larignon, 1999).
Bu verilere göre;
a. Petri hastalığının kaynağı ana bitkiden alınan bulaşık üretim materyalidir. Sonuçta bağda gelişen fidan ve genç bitki hastalıklıdır. Bu omca ya ölür ya da sadece Petri hastalığını taşıyarak ömrüne devam eder. Ancak bu arada Kav hastalığı da tabloya iştirak edebilir ve omca iki hastalığı birlikte taşır.
b. Ana bitkiden alınan üretim materyali sağlıklıdır, ancak elde edilen fidan Petri hastalığı ile karşılaşır. Bu fidan toprağa dikildiğinde genç bitkide hastalık devam eder. Bu bitki ya ölür ya da sadece Petri hastalığını taşıyarak ömrüne devam eder. Ancak bu arada Kav hastalığı da tabloya iştirak edebilir ve omca iki hastalığı birlikte taşır.
c. Ana bitkiden alınan üretim materyali sağlıklıdır, bundan üretilen fidan da sağlıklıdır, Ancak bağa dikilen fidan bağda Petri hastalığı ile karşılaşır. Bu bitki ya ölür ya da sadece Petri hastalığını taşıyarak ömrüne devam eder. Ancak bu arada Kav hastalığı da tabloya iştirak edebilir ve omca iki hastalığı birlikte taşır.
d. Ana bitkiden alınan üretim materyali sağlıklıdır, bundan üretilen fidan ve bağdaki genç bitki de sağlıklıdır. Bu bitki ya sağlıklı olarak ömrüne devam eder ya da sonraki dönemde sadece Petri veya sadece Kav hastalığı ile karşılaşır ya da her ikisine de yakalanır.
Çizelge 6. Asma fidanı ile genç ve yaşlı asmalarda saptanan fungal etmenler.
Fungal Etmenler Asma Fidanı
Genç Asma
Yaşlı Asma
Üretim Materyali
Phaeomoniella chlamydospora (Pcl) x x x x
Phaeoacremonium oleophilum (Pm) x x x x
Fomitiporia mediterrenea (Fom) - x x -
Fomitiporia punctata (Fop) - - - -
Phellinusigniarius (Ph) - - - -
Stereum hirsitum (Ste) - - - -
Sonuç olarak; asma fidanlarında Petri hastalığı, genç asmalarda ağırlıklı olarak Petri hastalığı görülür, ancak Kav hastalığı da sonradan iştirak edebilir. Yaşlı asmalarda ise ağırlıklı olarak Kav hastalığı görülür ancak Petri hastalığı da sonradan budama yaralarından giriş yaparak tabloya iştirak edebilir. Sonuçta bu asmadaki hastalık toptan ESCA olarak adlandırılır, böylelikle ESCA iki hastalığın bir arada olduğu bir hastalık kompleksi olarak kabul edilir.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
24
Şekil 12. Kav ve Petri hastalığına neden olan etmenlerin asmaya yerleşme sıraları ve zamanları (Surico, 2001).
Çalışmamızda elde ettiğmiz bulgular ışığında önerilerimiz aşağıda özetlenmiştir:
- Ege Bölgesinde daha çok yaşlı asmalarda sorun olduğu bilinen Kav hastalığının genç bağlarda da yaygınlaşma eğiliminde olduğu görüşü aslında Petri hastalığına dayanmaktadır.Genç asmaların yaprak ve üzüm danelerinde Petri hastalığı da Kav’ı andıran belirtilere yol açmakta ve sonuçta omca Kav’a tutulmuş olarak kabul edilmektedir. Gerek yeşil aksam belirtileri ve gerekse odun dokusundaki belirtilerin benzerliği veya kompleks görünümleri Petri hastalığının varlığını gizlemekte, belirtiler Kav hastalığına dayandırılmaktadır.
- Dünyada ve Türkiye’de “Esca” adı Kav hastalığının sinonimi olarak kullanılmaktadır. Bu adın hem Petri ve hem de Kav hastalığına işaret etmesi, literatürde böyle anılması bu hastalıkların mücadelesi açısından yararlı olacaktır.
- Fidanlıkların Petri hastalığı etmenleri ile bulaşık olması bu hastalığın neden yayıldığına da işaret etmektedir. Fidanların bulaşık olması üretim materyalinin de bulaşık olduğunu göstermektedir. Bu nedenle fidanlıklarda temiz üretim materyali kullanılmalı, yeni bağlar temiz fidanlarla kurulmalıdır.
-Türkiye’de Petri hastalığı henüz kayda geçmemiştir. Bu nedenle teknik elemanlara ve üreticilere Petri hastalığı ile ilgili eğitim verilmesi yararlı olacaktır.
-Petri hastalığı etmenleri Pcl ve Pm iç ve dış karantina listelerine dahil edilmelidir.

D. POYRAZ, E. ONOĞUR
25
Ege Bölgesi Asma Fidanlıkları ve Bağlarında Kav ve Petri Hastalıkları Üzerinde Araştırmalar
ÖZET
Bu çalışma, Ege Bölgesi Manisa, Denizli ve İzmir illerindeki fidanlıklar ile üretici bağlarında Kav ve Petri hastalıklarına yol açan fungal etmenleri saptamak ve karmaşık belirti tablosunu aydınlatmak amacıyla planlanmış ve 2009-2012 yılları arasında yürütülmüştür.
Çalışma, asma fidanlıklarından, genç ve yaşlı bağlardan alınan gövde, dal ve yaprak örneklerine dayalı olarak yürütülmüştür. Etmenlerin morfolojik özelliklerine ayırd edilmesi için uygun geliştirme ortamları kullanılmış ve tanıda A.B.D.’den temin edilen referans izolatlardan yararlanılmıştır.Araştırma sonunda, fidanlıklarda Petri hastalığına neden olan Phaeoacremonium aleophilum (Pm) ve Phaeomoniella chlamydospora (Pcl) etmenlerinin varlığı belirlenmiş, genç asmalara sahip bağlarda ağırlıklı olarak yine Petri hastalığının etmenleri saptanırken, bu etmenlere ek olarak Kav hastalığı etmeni Fom’nın da belirti tablosuna iştirak ettiği görülmüştür. Yaşlı bağlarda (10-25 yıllık)ise asmaların ağırlıklı olarak Kav (Fomitiporia mediterrenea (Fom)) ve yanında Petri hastalığı (Pmve Pcl) ile bulaşık olduğu saptanmıştır.
Anahtar kelimeler: Asma, Petri hastalığı, Kav hastalığı,Phaeoacremonium aleophilum, Phaeomoniella chlamydospora, Fomitiporia mediterrenea.
LITERATURE CITED
Anonim. 2008. Zirai Mücadele Teknik Talimatları. Tarımsal Araştırmalar ve Politikalar Genel Müd., Ankara.
Anonim. 2010. Türkiye İstatistik Kurumu.http://www.tuik.gov.tr/ (Erişim tarihi: 03.02.2012).
Andolfi, A., Mugnai, L., Luque, J., Surico, G., Cimmino, A. and Evidente, A. 2011. Phytotoxins Produced by Fungi Associated with Grapevine Trunk Diseases.Toxins, 3(12): 1569-1605.
Aroca, A., Garcia-Figueres, F., Bracamonte, L. and Raposo, R. 2006. A Survey of Trunk Disease Pathogens within Rootstocks of Grapevines in Spain. Europen Journal of Plant Pathology, 115:195-202.
Crous, P.W. and Gams, W. 2000. Phaeomoniella chlamydospora gen. et comb. nov., a causal organism of Petri grapevine decline and esca, Phytopathologia Mediterranea 39: 112–118.
Çelik H., Ağaoğlu, Y.S., Fidan, Y., Marasalı, B. ve Söylemezoğlu, G. 1998. Genel Bağcılık, Sunfidan A.Ş. Mesleki Kitaplar Serisi:1.
Erkan, M. and Larıgnon, P. 1998. Fungi assosiated with esca disease in grapevines in the Aegen Region. Turkey, Journal Turkısh Phytopathology, 27 (2-3): 137-143.
Erkan, M. 2000. A general approach for esca disease in the vineyards of Turkey. Phytopathologia Mediterranea, 39: 35-37.
Eskalen, A., Rooney-Latham, S. and Gubler, W.D. 2005. Occurrence of Togninia fraxinopennsylvanica on Esca Diseased Grapevines (Vitis vinifera) and Declining Ash Trees (Fraxinus latifolia) in California. Plant Disease, 89(5): 528.
Fischer, M. 2002. A new wood-decaying basidiomycete species associated with esca of grapevine: Fomitiporia mediterranea (Hymenochaetales). Mycological Progress 1(3): 315-324.
Fischer, M. and Kassemeyer, H. M.. 2003. Fungi associated with esca disease of grapevine in Germany. Vitis 42 (3): 109-116.
Giménez-Jaime, A., Aroca, A., Raposo, R., García-Jiménez, J. and Armengol, J. 2006. Occurrence of fungal pathogens associated with grapevine nurseries and the decline of young vines in Spain.Journal of Phytopathology154: 598-602.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
26
Graniti, A., Surico, G. and Mugnai, L. 2000. Esca of Grapevine: A Disease Complex or A Complex of Diseases. Phytopathologia Mediterranea, 39: 16-20.
İyriboz, N. 1942. Bağ hastalıkları (2.Basım). Ziraat Vekâleti Neşriyatı, sayı:323–2, İzmir, 232s.
Karaca, L. 1965. Sistematik Bitki Hastahklan (Phycomycetes, Basidiomycetes) Cilt II. Ege Üniv. Matbaası. Yayın No: 107, İzmir, 180 s.
Kuntzmann, P., Villaume, S., Larignon, P. and Bertsch, C. 2012.Esca, BDA and Eutypiosis: Foliar Symptoms, Trunk Lesions and Fungi Observed in Diseased Vinestocks inTwo Vineyards in Alsace.Vitis, 49 (2):71-76.
Larignon, P. and Dubos, B. 1997. Fungi Associated with Esca Disease in Grapevine, European Journal of Plant Pathology. 103: 147-157.
Larignon, P. 1999. Esca Disease from a European Perspective, Black Goo Symptoms and Occurrence of Grape Declines, International Ampelography Society: Fort Valley, Virginia, USA, 43-55p.
Lecomte, P., Darrieutort, G., Liminana, J.-M., and Comont, G. 2012.New Insights into Esca of Grapevine: The Development of Foliar Symptoms and Their Association with Xylem Discoloration.July 2012, 96 (7): 924-934.
Luque, J., Martos, S., Aroca, A., Raposo, R. and Garcia-Figueres, F. 2009.Symptoms and Fungı Associated with Declining Mature Grapevine Plants in Northeast Spaın. Journal of Plant Pathology, 91 (2): 381-390.
Morton, L. 1999. Black Goo Symptoms and Occurrence of Grape Declines. International Ampelography Society: Fort Valley, Virginia, USA, 132p.
Mostert, L., Hallen, F., Fourıe, P. and Crous, P.W. 2006a. A Review of Phaeoacremonium species involved in Petri Disease and Esca of Grapevines. Phytopathology Mediterranean 45: 12-29.
Mostert, L., Groenewald, J.Z., Gams, W. and Summerbell , R. and Crous, P.W. 2006b. Taxonomy and pathology of Togninia (Diaporthales) and its Phaeoacremonium anamorphs. Studies in Mycology 54: 1-115.
Mugnai, L., Graniti, A. and Surico, G. 1999. Esca (Black measles) and Brown Wood Streaking: Two Old and Elusive Diseases of Grapevines. Plant Disease 83 (5): 404-418.
Onoğur, E. 1995. Bağ Hastalıkları, Alaşehir Meslek Yüksek Okulu Yayınları, Yayın No: 1., Ege Univ. Basımevi, Bornova-İzmir, 97 s.
Penn, C. 2001. From Mystery Disease to Discovery of Pathogens.http://winebusiness.com/html/MonthlyArticle (Erişim Tarihi: 24.10.2008).
Poyraz, D. 2012. Ege Bölgesindeki Bağlarda Petri ve Kav Hastalığına Neden Olan Fungal Etmenlerin Moleküler Yöntemlerle Saptanması ve Mücadelesi Üzerine Araştırmalar. Doktora Tezi. Ege Üniversitesi. Fen Bilimleri Enstitüsü. Bitki Koruma Ana Bilim Dalı. Bornova-İZMİR.
Retief, E., Mcleod, A. and Fourie, P.H. 2006. Potential İnoculum Sources of Phaeomoniella chlamydospora in South African Grapevine Nurseries. European Journal of Plant Pathology, 115: 331-339.
Rooney-Latham, S., Eskalen, A., and Gubler, W. D. 2005. Ascospore release of Togninia minima, cause of esca and grapevine decline in California.http://plantpathology.ucdavis.edu/ (Erişim Tarihi: 24.10.2008).
Scheck, H.J., Vasquez, S.J. and Gubler, W.D. 1998. First Report of Three Phaeoacremonium spp. Causing Young Grapevine Decline in California. Plant Disease, 82: 590.
Stamp, J.A. 2001. The Contribution of İmperfections in Nursery Stock to The Decline of Young vine in California. Phytopathology Mediterranean, 40: 369-375.
Surico, G. 2000. The grapevine and wine production through the ages. Phytopathologia Mediterranea 39: 3-10.
Surico, G. 2001. Towards Commonly Agreed Answers to Some Basic Questions on Esca. Phytopathologia Mediterranea, 40: 487-490.

D. POYRAZ, E. ONOĞUR
27
Surico, G., Mugnai, L. and Marchi, G., 2006, Older and more recent observations on esca: a critical overview, Phytopathologia Mediterranea, 45: 68-86.
Surico, G. 2009. Towards a redefinition of the diseases within the esca complex of grapevine. Phytopathologia Meditterranea, 48: 5-10.
Üzümeri, M.E. 1947. Bağ Hastalıkları, Tarım bakanlığı Neşriyat Müdürlüğü, Sayı:636, Ankara, 245s.
Vasquez S. 2012. Grapevine Measles,http://www.extension.org/ (Erişim Tarihi: 01.08.2012).
Viala, P. 1926. Recherches sur les maladies de la vigne, Esca, Annales des Epiphyties Fasc. 1 et 2: 1-108.
White, C. L. 2010. The Characterization of The Basıdıomycetes and other Fungı Associeted with Esca of Grapevines in South Africa, Thesis presented in partial fulfillment of the requirements for the degree of Master of Science at Stellenbosch University, 157 p.

STUDIES ON ESCA AND PETRI DISEASES IN GRAPEVINE NURSERIES AND VINEYARDS IN AEGEAN REGION
28

J. Turk. Phytopath., Vol. 42 No. 1-3, 29-34, 2013 ISSN 0378 - 8024
29
Optimization of a Specific Real-Time PCR Assay for Fusarium avenaceum Using Species-specific Couples of Primer
Şerife Evrim ARICI * Peter KARLOVSKY **
* Department of Plant Protection, Faculty of Agriculture, Süleyman Demirel University, 32260, Isparta, Turkey
** Molecular Phytopathology and Mycotoxin Research, Georg August University Göttingen, Grisebachstrasse 6, 37077 Göttingen, Germany
Accepted for publication March 26, 2014
ABSTRACT
Fusarium head blight (FHB) is an important cereal disease and may result in the accumulation of toxins in grains. Although F. graminearum is globally the most prevalent FHB causing species, F. avenaceum is found in contaminated samples. Real-time PCR (qPCR) is the the standard analytical method for species-specific, quantitative estimation of fungal biomass in the tissue of host organisms. qPCR is useful for quantifying fungal colonization of crops while distinguishing among species. Recently, species-specific PCR primers have been developed for most Fusarium species that cause head blight. In some cases, the species-specific primers synthesized with other Fusarium species and can give the wrong results. In this study, two species-specific primer pairs for F. avenaceum were tested by using with different fungi DNA. JIA primer pairs were amplified only F. avenaceum, whereas MGA primer pairs were amplified with F. equiseti and F. tricinctum. Results indicate that primer pair JIA is effective in determination of F. avenaceum.
Key words: Fusarium avenaceum, real-time PCR, wheat, species-specific primer
INTRODUCTION
Fusarium head blight (FHB), also known as scab or ear blight, is one of the most serious diseases in wheat growing areas (Parry et al. 1995; Goswami and Kistler 2004). FHB pathogens result in severe yield losses and a decline in cereal quality. Furthermore, infections by these pathogens lead to contamination of grain and straw by a wide array of mycotoxins. These fungal metabolites pose serious threats to human and animal health (Bennett and Klich 2003). FHB may be caused by 17 dierent Fusarium species, the most common of which are Fusarium graminearum, F. culmorum, F. avenaceum, F. poae, and F. nivale (Parry et al., 1995; Goswami and Kistler 2004). The predominant Fusarium head blight species besides F. graminearum are F. culmorum F. avenaceum and F. Poae, F. avenaceum is widespread throughout the cool temperature cereal growing areas and produces moniliformin, enniatins and fusarin (Desjardin, 2006; Yli-Mattila et al., 2006). Traditional diagnostic methods for detection and identification of F. avenaceum are based on micro- and macro morphological features developing on a medium. This process is time consuming and requires extensive training. In addition, it can often be difficult to distinguish between species having similar morphological characteristics. Recently, polymerase chain reaction (PCR) and Real-time quantitative-PCR are sensitive and rapid methods that can be used for the detection and screening of fungal microorganisms. Real-time PCR (qPCR) is the standard analytical method for species-specific, quantitative estimation of fungal biomass in the tissue of host organisms. qPCR is useful for quantifying fungal colonization of crops while distinguishing among species. Real-time quantitative-PCR is a sensitive and rapid method that can be used for the detection and screening of F. avenaceum. More recently, real-time quantitative PCR assays with

OPTIMIZATION OF A SPECIFIC REAL-TIME PCR ASSAY FOR FUSARIUM AVENACEUM USING SPECIES-SPECIFIC COUPLES OF PRIMER
30
fluorogenic sequence-specific probes for F. avenaceum have been developed (Bluhm et al., 2004; Reischer et al., 2004; Waalwijk et al., 2004; Klemsdal et al., 2006; Yli-Mattila et al., 2006). Real-time PCR provides a means for detection and quantification of DNA targets by monitoring PCR product accumulation during the thermal cycling. The two main chemistries employed by real time-PCR are SYBR Green I and dual-labelled fluorescent probes, or TaqMan probes. Real-time PCR enables simpler and more rapid analysis of data, and has higher sensitivity and a wider dynamic range compared with conventional PCR (McKillip and Drake, 2004). Species-specific primers have been used for detection and screening of F. avenaceum (Schilling et al., 1996; Turner et al., 1998; Doohan et al., 1998; Waalwijk et al., 2003, 2004) and primers should be designed only for F. avenaceum. In some cases, the species-specific primers synthesized with other Fusarium species and can give the wrong results. The purpose of present study was to comparison of the different species-specific PCR primers for the identification of F. avenaceum.
MATERIALS AND METHODS
Fungal Isolates
The fungal strains used in this study are listed in Table 1. Fusarium spp. isolates for DNA isolation were grown in 100 ml of mungbean media with shaking (200 rpm) at 25 °C for 4 to 5 days. The mycelium was harvested by filtration and then freeze-dried. The other fungal isolates were grown in broth potato dextrose media.
DNA Extraction
A variant of the cetyltrimethyl ammonium bromide (CTAB) method described by Brandfass and Karlovsky, (2006) previously was used, and the quality and quantity of DNA were estimated by electrophoresis in 0.8% (w/v) agarose gel. The gel was stained with ethidium bromide (2 mg/lt) and documented with a digital imaging system (Vilber Lourmat, Marne la Vallee, France). The densitometry was performed using Multi-Analyst (Bio-Rad, Hercules, CA, USA). The concentration of fungal DNA was calculated by comparing a dilution series with defined amounts of DNA of lambda phage (methylated, from Escherichia coli host strain W3110).
Two couples of primer were used for F. avenaceum. One of them is JIAF (GCT AAT TCT TAA CTT ACT AGG GGC C) and JIAR (CTG TAA TAG GTT ATT TAC ATG GGC G) designed by Turner et al. 1998. The other one is MGB-F (CCATCGCCGTGGCTTTC) and MGB-R (CAAGCCCACAGACACG TTGT) designed by Waalwijk et al. 2004.
Real Time-PCR
The specificity of both PCR assays was determined with DNA extracted from pure cultures of 39 fungal isolates (35 Fusarium species and 4 isolates of other fungal species, Table 1). Real-time PCR was carried out in a total of 4 μl consisting of 10X buffer (Bioline, Germany), 2 mM MgCl2, 150 µM dNTP (Bioline, Germany), 0.3 μM concentration of each primer, 0.025 u Taq DNA poymerase, (BIOTaq, Bioline, Germany), 0.1× SYBR Green I (Invitrogen, Karlsruhe, Germany), and 1 μl template DNA. PCR reactions were performed in duplicate on all samples. Genomic DNA from pure cultures was diluted 1:100 before PCR.
PCR was performed on CFX384 Real-Time PCR Detection Systems (Bio-Rad, CA, USA). The amplification for F. avenaceum consisted of an initial denaturation at 94 °C for 2min, followed by 35 cycles of 30 s denaturation at 94 °C, 20 s annealing at 60 °C, and 30 s elongation at 72 °C. The final elongation step was performed for 5 min at 72 °C (Dastjerdi et al., unpublished).

Ş.E. ARICI, P KARLOVSKY
31
Table 1. Fungal strains used in this study
Species Strain Source
F. subglutinans Fsub2202 A
F. subglutinans Fsub2215 A
F. subglutinans Fsub2210 A
F. concolar Fconc2 A
F. crookwellense FCKW3 A
F. crookwellense FCKW4 A
F. culmorum FC15 E
F. culmorum FC2 D
F. culmorum FC22 E
F. culmorum FCH69 A
F. graminearum DSM6212 A
F. graminearum DSM62722 A
F. graminearum DSM67638 A
F. oxyporium FO125 A
F. oxyporium Foxy436 A
F. oxyporium Foxy119 A
F. oxyporium Foxy6 A
F. proliferatum Fpro2 G
F. proliferatum Fpro5 G
F. proliferatum Fpro10 G
F. sacchari Fsac1 A
F. sacchari Fsac2 A
F. verticillioides FRC8114 F
F. verticillioides Fv A
F. poea F. poe B
F. poea F. poe A
F. equiseti F. equiseti B
F. equiseti F. equiseti B
F. tricinctum F. trincinctum B
F. tricinctum F. trincinctum B
Septoria nodorum SN1 A
Michrodochium nivale GN7 E
Michrodochium nivale GN35 E
Michrodochium nivale GN36 E
F. avenaceum Fa14 A
F. avenaceum Fa16 C
F. avenaceum Fa17 C
F. avenaceum Fa18 A
F. avenaceum Fa5-2 A
A: Culture Collection at the Göttingen University, the Division of Plant Pathology and Crop Protection, Germany; B: Marcel Tillmann, Göttingen University, the Division of Plant Pathology and Crop Protection, Germany; C: Berna Tunali, University of Ondokuz Mayis , Agricultural Faculty, Department of Plant Protection, Samsun, Turkey; D: H. Nirenberg (BBA Berlin, Germany) via E. Möller; E: T. Miedaner, State Plant Breeding Institute, University of Hohenheim, Stuttgart, Germany, via E. Möller; F: A. Desjardins, USA, Mexico, via E. Möller; G: A. Szecsi, Budapest, Hungary via E. Möller
RESULTS AND DISCUSSION
We investigated the species-specific PCR primers for F. avenaceum against 3 strains of F. subglutinans, one strain of F. concolar, two strains of F. crookwellense, four strains of F. culmorum, three strains of F. graminearum, four strains of F. oxysporum, three strains of F. proliferatum, two strains F. sacchari, two strain of F. verticillioides, two strains of F. poe, two strains of F. equiseti two strains of F. tricinctum, five strains of

OPTIMIZATION OF A SPECIFIC REAL-TIME PCR ASSAY FOR FUSARIUM AVENACEUM USING SPECIES-SPECIFIC COUPLES OF PRIMER
32
F. avenaceum, one strain of Septoria nodorum and three strains of Michrodochium nivale. Two couples of primer were evaluated for the specific real-time PCR assay for F. avenaceum. JIA and MGB which are specific for F. avenaceum were used to test Fusarium isolates. A standardized PCR protocol was established according to optimal conditions of all two primers (2mM MgCl2, 200 μM dNTP, annealing temperature: 60 °C for 20 sec) (Dastjerdi et al., unpublished). Preliminary PCR assays, including melt curve analysis, were carried out with a BioRad iCycler (BioRad, Hercules, California, USA).
Primer couple of JIA specially amplified target DNA (F. avenaceum). All isolates of 14 non-target fungal species tested negatively. Samples of non-target fungal DNA generated no amplification products or unspecific products (Fig 1).
Fig 1. Melting curve analysis of PCR products obtained with primer JIA for F. avenaceum
Primer couple of MGB specially amplified target DNA (F. avenaceum). All isolates of 12 non-target fungal DNA generated no amplification products or unspecific products, but primer couple of MGB amplified F. equiseti and F. tricinctum (non-target DNA) with same melting temperature (Fig 2).
Fig 2. Melting curve analysis of PCR products obtained with primer MGA for F. Avenaceum
F. avenaceum
Unspecific product
no amplification product
F. tricintum
F. equisetiF. avenaceum

Ş.E. ARICI, P KARLOVSKY
33
Diagnostics for plant pathogens are important to identify fungal contaminants in crops and are invaluable for quarantine purposes. To confirm the morphological identification of species, PCR is performed by testing isolates with species-specific primers. Waalwijk et al (2002) tested seven primer pairs for their ability to identify isolates of Fusarium avenaceum, F. culmorum, F. graminearum, F. poae, F. proliferatum and M. nivale var. majus and M. nivale var. nivale. Each primer pair only generated a PCR product with the corresponding Fusarium species and all PCR fragments had different molecular sizes. Turner et al, (1998) tested the specificity of primers using primer pairs JIA for F. avenaceum against 2 strains of F. graminearum, two strains of F. culmorum, one strain of F. poae, one strain of F. sporotrichioides, thirty strains of F. avenaceum, one strain of F. chlamydosporum, one strain of F. crookwellense, one strain of F. flocciferum, one strain of F. heterosporum, four strains of F. sambucinum, one strain of F. solani, one strain of F. sporotrichoides, thirteen strains of F. tricinctum, and one or strains other fungal species. The primer was found to be specific for F. avenaceum. Yli-Mattila, (2002) tested primer pairs JIA for Fusarium avenaceum against Fusarium avenaceum/F. arthrosporioides/F. tricinctum species. It was reported that JIA primer pairs was specific for F. avenaceum, but F. avenaceum was most similar to F. tricinctum isolates. In conclusion, primer pairs JIA, performed under real-time PCR conditions, generated positive signals only for the target species (F. avenaceum), while primer pairs MGB amplified non target DNA (F. equiseti, F. tricinctum). Primer pairs JIA can therefore be regarded as species-specific in real-time mode for F. avenaceum, respectively.
ÖZET
Fusarium avenaceum için tür-spesifik primerler kullanılarak real time PCR yönteminin optimizasyonu
Fusarium başak yanıklığı hastalığı tahıllarda çok önemli bir hastalık olup danelerde toksin oluşturmaktadır. Başak yanıklığı hastalığına dünyada en yaygın F. graminearum neden olmasına rağmen F. avenaceum’da hastalıklı örneklerde bulunmaktadır. Gerçek zamanlı PCR (qPCR) konukçu organizmaların dokusundaki fungal biyokütlenin, kantitatif olarak belirlenmesi için kullanılan standart analitik bir yöntemdir. qPCR türler arasındaki farklılığı belirlemede kullanıldığı gibi, kültür bitkileri içerisinde fungal kolonizasyon miktarının belirlenmesinde kullanılmaktadır. Son yıllarda başak yanıklığı hastalığına neden olan bir çok Fusarium türü için türe özgü primerler geliştirilmiştir. Bu çalışmada farklı funguslar ve F. avenaceum kullanılarak F. avenaceum'a özel iki primer (JIA ve MGA) test edilmiştir. JIA primer çifti sadece F. avenaceum'a bağlanırken, MGA primer çifti F.equiseti ve F.tricinctum ile bağlanarak çoğalmıştır. Sonuç olarak JIA primeri F. avenaceum'un tespitinde daha etkili bulunmuştur.
Anahtar kelimeler: Fusarium avenaceum, gerçek zamanlı PCR, buğday, tür-spesifik primer
LITERATURE CITED
Bennett, J.W., Klich, M. 2003. Mycotoxins. Clinical Microbiology Reviews 16:497–516.
Bluhm, B.H., Cousin, M.A., and Woloshuk, C.P. 2004. Multiplex real-time PCR detection of fumonisin-producing and trichothecene-producing groups of Fusarium species. Journal of Food Protection 67: 536–543.
Brandfass, C., and Karlovsky, P. 2006. Simultaneous detection of Fusarium culmorum and F. graminearum in plant material by duplex PCR with melting curve analysis. BMC Microbiology, 1-10 doi:10.1186/1471-2180-6-4.
Desjardins, A.E., Hohn, T.M., McCormick, S.P. 1993. Trichothecene biosynthesis in Fusariumspecies – chemistry, genetics, and significance. Microbiol Rev. 57:595–604.
Doohan, F.M., Parry, D.W., Jenkinson, P., and Nicholson, P. 1998. The use of species-specific PCR-based assays to analyse Fusarium ear blight of wheat. Plant Pathology 47:197–205.
Goswami, R.S., Kistler, H.C. 2004. Heading for disaster: Fusarium graminearum on cereal crops. Molecular Plant Pathology 5:515–525.

OPTIMIZATION OF A SPECIFIC REAL-TIME PCR ASSAY FOR FUSARIUM AVENACEUM USING SPECIES-SPECIFIC COUPLES OF PRIMER
34
Klemsdal, S.S., Yli-Mattila, T., Paavanen-Huhtala, S., and Elen, O. 2006. Sensitive and quantitative real-time detection of trichothecene producing Fusarium spp. in cereal samples. European Journal of Plant Pathology (submitted).
McKillip, J.L., and Drake M.A. 2004. Real-time nucleic acid-based detection methods for pathogenic bacteria in food J. Food Prot., 67 (2004), pp. 823–832.
Nielsen, L.K., Jensen, J.D., Nielsen, G.C., Jensen, J.E., Spliid, N.H., Thomsen, I.K., Justesen, A.F., Collinge D.B., and Jorgensen, L.N., 2011. Fusarium head blight of cereals in Denmark: Species Complex and Related Mycotoxins, Population Biology, 101(8):960-969.
Parry, D.W., Jenkinson, P., McLeod, L. 1995. Fusarium ear blight (scab) in small grain cereals - a review. Plant Pathology 44:207–238.
Reischer, G.H., Lemmens, M., Farnleitner, A., Adler, A., and Mach, R.L. 2004. Quantification of Fusarium graminearum in infected wheat by species-specific real-time PCR applying a TaqMan Probe. J. Microbiol. Methods 59, 141–146.
Schilling, A.G., Möller, E.M., and Geiger, H.H., 1996. Polymerase chain reaction-based assays for species-specific detection of Fusarium culmorum, F. graminearum and F. avenaceum. Phytopathology 86: 515-522.
Turner, A.S., Lees, A.K., Rezanoor, H.N., and Nicholson, P. 1998. Refinement of PCR-detection of Fusarium avenaceum and evidence from DNA marker studies for phenetic relatedness to Fusarium tricinctum. Plant Pathology 47:278–288.
Waalwijk, C., Kastelein, P., deVries, I., Kereny, Z., van der Lee, T., Hesselink, T. , Kohl, J.,and Kema, G. 2003.Major changes in Fusarium spp. in wheat in the Netherlands Eur. J. Plant Pathol., 109: 743–754.
Waalwijk, C., van der Heide, R., De vries, I., Van der Lee, T., Schoen, C., Costrel-de Corainville, G., Hauser-Hahn, I., Kastelein, P., Kohl, J., Lonnet, P., Demarquet, T., and Kema, G.H.J., 2004. Quantitative detection of Fusarium species in wheat using TaqMan. Eur. J. Plant Pathol. 110, 481–494.
Yli-Mattila, T., Paavanen-Huhtala, S., Parikka, P., Konstantinova, P., Gagkaeva, T., Eskola, M., Jestoi, M., and Rizzo, A. 2002. Occurrence of Fusarium fungi and their toxins in Finnish cereals in 1998 and 2000. J Appl Gen 43A:207–214.
Yli-Mattila, T., Paavanen-Huhtala, S., Parikka P., Konstantinova, P., and Gagkaeva, T. Y. 2004. Molecular and morphological diversity of Fusarium species in Finland and north western Russia. Eur. J. Plant Pathol.110:573-585.
Yli-Mattila, T., Paavanen-Huhtala, S., Parikka, P., Jestoi, M. Klemsdal, S., and Rizzo A.2006. Genetic variation, metabolites and mycotoxins of Fusarium avenaceum and related species. Mycotoxin Res., 22: 79–86.

J. Turk. Phytopath., Vol. 42 No. 1-3, 35-45, 2013 ISSN 0378 - 8024
35
Identification and Pathogenicitiy of Botryosphaeriaceae Species Causing Wood Canker on Grapevines
D. Soner AKGÜL* Nurdan GÜNGÖR SAVAŞ* Turcan TEKER*
Bilgehan KARAHAN* Bilge KEYKUBAT**
* Manisa Viticulture Research Station, PK:12, 45125, Yunus Emre, Manisa, Turkey
** İzmir Commodity Exchange, GaziBulvarı, No:2, 35250, Konak / İzmir, Turkey Accepted for publication April 21, 2014
ABSTRACT
Wood canker and dieback diseases, caused by Botryosphaeriaceae fungi, are among the most important fungal trunk diseases of grapevines. In recent years, the symptoms of local dead arm, shoot dieback and V-shaped discolorations in woody tissues have dramatically increased in vines of Turkey. During the 2012 growing season, symptomatic wood samples (from Ankara, Çorum, Izmir, and Manisa cities) were taken and standard mycological isolations were done to determine the fungal agents of disease. A high proportion (69.4%) of Botryosphaeriaceae fungi was isolated from these samples. In the first stage of the isolates’identification, fungal DNA was extracted and amplified in Real-Time thermocycler by using the genus spesific primers (BOT100F-BOT472R). In this way, the members of Botryosphaeriaceae species were distinguished from other similar species. In molecular identification of the isolates, ITS 1 and ITS 2 ribosomal DNA fragments were sequenced and the gene sequences were compared with those deposited in NCBI Gene Bank database.It has been determined that 4 different species Botryosphaeria dothidea (Anamorph; Fusicoccum aesculi), Diplodia seriata (An; B. obtusa) Lasiodiplodia theobromae (An; B. rhodina) and Neofusicoccum parvum (An; B. parva) were found in the sequenced 15 isolates. Colony morphology on PDA media andpicniospore shapes on woody tissues were examined in morphological / microscopic identification. To fulfill Koch’s postulates, pathogenicity tests for all isolates were conducted on 1-year-old potted vines. Four months after inoculation, the lesion lengths on woody tissues were evaluated. Pathogenicity tests revealed that Neofusicoccum parvum species produced larger lesions on woody tissues when compared to control and other species.
Key words: Botryosphaeriaceae, Grapevine, Trunk Diseases.
INTRODUCTION
Fungal trunk diseases have become a growing threat to vineyards in Turkey and throughout the world. Trunk diseases of grapevine are caused by fungal pathogens that invade through pruning wounds located on the woody parts of the plant. Botryosphaeriaceae canker (Bot canker) is one of the most common fungal trunk diseases in grape growing areas of the world. These diseases reduce yields and increase production costs. Eutypa dieback and Bot canker were estimated to cause over 260 million dollars yield loss in California US, and up to 20% yield loss in Bordeaux Region, France (Siebert, 2001; Wilcox, 2011).To date, 21 species in the Botryosphaeriaceae family have been reported in 17 grape producing countries (Urbez-Torres, 2011) and four species of them found in Turkey (Akgül et al. 2014).
The disease was characterized by the presence of wedge or pie-shaped perennial cankers in spurs, cordons or trunks (Figure 1a). Cankers start to develop primarily from pruning wounds, cracks, as well as big cuts. In the initial stages of cankers, browning streaks appear internal woody tissues, by the time they prolong to other tissues. Due to high enzymatic activity and toxin production, affected tissues die and local or entire vine drying can occur

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
36
eventually (Figure 1b). These kind of wood symptoms can also be seen in Eutypalata, Diaporthe and Phomopsis infectionson vines (Urbez-Torres, et al., 2013).
a b
Figure 1.Wedge-shaped discolorations on wood (a), local drying in a mature vine (b).
The Botryosphaeriaceae fungi produce a variety of toxic metabolites having an important role on their virulence and pathogenesis. The production of toxins and some exopolysaccharides (EPS) can induce phytotoxic effects on Botryosphaeriaceae-infected grapevines. Withering of the leaves, interveinal necrotizations and discoloured spots were shown when grapevine leaves were immersed in toxin suspensions (Evidenteet al. 2010). Environmental conditions are other factors affecting the virulence of Botryosphaeriaceae fungi. Some species are more virulent than the others in higher temperature, so these species predominate in that specific region (Rolshausenet al. 2008). The virulence varies between species and isolates of the same species in different countries (Urbez-Torres and Gubler, 2009).On the other hand, these differences may affect the control strategies against Bot species in that region. To date, no study has been initiated about the etiology, epidemiology and control of Bot-canker pathogens on grapevine in Turkey. Doing accurate identification and fulfilling pathogenicity tests are essential steps for any research in plant pathology.
In this study, identification of Botryosphaeriaceae species by classic and molecular biology methods and pathogenicity tests on grapevines were introduced.
MATERIALS AND METHODS
Pathogen Isolation and Colony Discrimination
Symptomatic grapevine wood samples, having typical wedge-shaped discoloration, were surface disinfested with 95% ethanol and flame sterilized and the discolored outer bark was cut away. The internal tissues (0.5 cm2) were excised, then they were plated onto Potato Dextrose Agar (PDA: Merck) amended with streptomycin-sulphate (150 g/L) (Sigma Aldrich). After 4 days of incubation (24°C temp, in the dark), fast growing colonies were subcultured and the pure cultures were incubated for 20-25 days in 24°C temp.- under 16 h light/8 h darkfor visual and microscopic examination.
Molecular Identification
DNA Extraction and PCR Tests
Fungal DNA was extracted by following the extraction protocol of Cenis (1992). Mycelial mats (approximately 50 mg) were directly taken from the fresh cultures of the isolates with a sterile surgical blade and

D. S. AKGÜL, N. G. SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT
37
they were crushed in micro-centrifuge tubes (1.5 ml) with a sterile plastic pestle by adding 550 µl DNA extraction buffer (200 mM Tris-HCl (pH:8.5), 250 mMNaCl, 25 mM EDTA and 2% Sodium Dodecyl Sulphate). After homogenization, 150 µl of 3M Sodium Acetate (NaOAc) was added into tubes and they were placed at -20°C for 15 min. The homogenates were centrifuged at 14.000 rpm for 10 min and the supernatant (200 µl) was transferred to the new tubes. Equal volume of isopropanol (2-propanol) was added and mixed gently about five times, and the tubes were placed at 0°C for 10 min. After that, DNA pellet was precipitated by centrifugation at 14.000 rpm for 10 min and supernatant was discarded. DNA was washed with 1 ml of 70% ethanol for 1-2 sec and the pellet was dried for 10 min. under hood. Finally, DNA was suspended with 75 µl of TE (1M Tris-HCl, pH:8 and 0.5M EDTA) buffer and stored at -20°C.The concentration and purity of extracted DNA were measured with nano-drop device (Thermo Scientific MultiScan Go) by dropping 2 µl sample.
The genus specific primers (BOT 100F – BOT 472R) were used to discriminate the Botryosphaeriaceae isolates from the other fungi in PCR tests (Table 1). 10 µl Roche FastStart Essential DNA Green Master Mix (2X conc.), 7.4 µl of RNAse free water, 0.3 µl of forward and reverse primers (20 mM conc.), 2 of µl template DNA were included in thermocycler tubes. DNA samples were amplified with a real-time thermocycler (Roche Light Cycler® Nano) according to the amplification protocol (initial denaturation 95°C for 10 min and followed by 35 cycles at 95°C 10s., 58°C 10 s., 72°C 15 s. and 72°C 10 min final extension). PCR products were seperated 1.5% agarose gel electrophoresis and stained with ethidium-bromide (EtBr) to visualize amplicons on gel documentation unit (DNR Bio Imaging Systems). To check the specificity of genus specific primer pair, DNA samples of Alternaria alternata, Fomitiporia mediterranea and one isolate of Botryosphaeria dothidea (from olive) fungi were included in PCR experiments as control and nuclease free water (instead of template) was used for the negative control.
In addition to classic PCR tests, Botryosphaeriaceae isolates were identified with both direction sequence analysis. Ribosomal DNA fragments (ITS1, 5.8S ITS2 rDNA loci) were amplified with ITS4 / ITS5 universal primers (Table 1) by using real-time thermocycler amplification protocol (initial denaturation 95°C for 10 min and followed by 35 cycles at 95°C 10s., 51°C 10 s., 72°C 15 s. and 72°C 10 min final extension).
After that, PCR products were sequenced by Macrogen Co, South Korea, and the sequences were compared with those deposited in NCBI Gene Bank database. The ITS sequences were also submitted to the NCBI GenBank and accession numbers were obtained (Table 2).
Table 1. Primer sets used to amplify two loci analyzed in this study.
Primer Amplicon Annealing
Pairs PrimerSequences
Size (bp) Temp. (°C) Reference
ITS 4 5 ‘ TCC TCC GCT TAT TGA TAT GC 3’
ITS 5 5’ GGA AGT AAA AGT CGT AAC AAG G 3’ 580 51 (White et al., 1990)
BOT 100 F 5’ AAA CTC CAG TCA GTR AAC 3’
BOT 472 R 5’ TCC GAG GTC AMC CTT GAG 3’ 372 58 (Ridgway et al., 2011)
Morphological and Microscopic Identification
The Botryosphaeriaceae-like fungi were grown on PDA medium in the incubator conditions (24°C constant temperature, under fluorescent lamps, 16h light/8h dark) for 20-25 days and colonies were allowed to make their typical morphology. After incubation, colony color at the bottom of petri plates, hyphal morphology and aerial mycelium constitution and picniospore morphology were examined under light microscope, if they were produced on PDA. In addition, all the isolates were inoculated onto uninfected canes to induce picnidia and picniospore formation. Twenty cm mature canes were washed under tap water and the surfaces were cleaned with 70% ethanol. These canes were wounded by removing bark with 5-mm cork borer and fresh mycelial plugs were inoculated into the holes and covered with parafilm. After inoculation, the bottom ends of canes were placed in tap water (in plastic beakers) and they were incubated in climate-room (24°C temp, 80% relative humidity). Three weeks later, picnidia and picniospores were examined under light microscope and conidia dimensions were measured.
Pathogenicity Tests
Pathogenicity tests were conducted under greenhouse conditions on 1-year-old own rooted grapevine (Vitis vinifera cv. Sultana Seedless) plants. These plants were inoculated (as described previously) with mycelial plugs of

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
38
all isolates and they were grown in greenhouse for 12-16 weeks. After this, outer bark of stem was carefully scraped and the plants were examined for the extent of wood discoloration and recovery of fungal isolates. Lesion lengths were recorded and data were analyzed by performing Duncan’s multiple range test (<0.05). To complete Koch’s postulates, the isolates were re-isolated from the inoculation sites. Five plants were used for each isolate and 75 plants were in total.
RESULTS
The Botryosphaeriaceae and the others (wood colonizing saprophytic fungi: Alternaria spp., Arthrinium sp. Chaetomium sp. Fusarium equiseti and Nectria sp.) were isolated from the symptomatic wood samples with 69.4% and 30.1% isolation rates respectively.
PCR Tests and Sequence Analysis Results
Sequence and NCBI-BLASTn analysis of the PCR products (amplified by universal primer pair, ITS4/ITS5) revealed that four Botryosphaeriaceae species (Botryosphaeria dothidea (Anamorph; Fusicoccum aesculi), Diplodia seriata (An; B.obtusa) Lasiodiplodia theobromae (An; B. rhodina) and Neofusicoccum parvum (An; B. parva)) were obtained from the symptomatic wood samples. The DNA sequences of these species showed 98-100% similarity with those previously deposited sequences (submitted from other countries) in NCBI Gene Bank database. The species names, location, their accession number records, nucleotid sizes, maximum identity percentages and BLASTn comparisons (with the other isolates) were presented in Table 2.
Table 2. List of Botryosphaeriaceae fungi isolated from the grapevines with their corresponding GenBank accession numbers and data of BLASTn results obtained from GenBank.
GenBank Query BLASTn Maximum Species Isolate Location
Accession No Lenght (bp) Accession No Identity (%)
Botryosphaeria dothidea MBAi25AG Manisa KF182329 451 FJ790847 99
MBAi48CL Saruhanlı KJ596525 558 JQ411394 99
MBAi135AG Manisa KJ596531 450 KF876691 99
Diplodia seriata MBAi23AG Manisa KF182328 583 KC020170 98
MBAi30AG Manisa KF988138 581 KF182332 99
MBAi32AG Manisa KF182332 581 KF988138 99
MBAİ183AG Salihli KJ596528 597 KC020170 99
MBAi185AG İzmir KJ596530 594 KC020170 99
Lasiodiplodia theobromae MBAi28AG Manisa KF182331 641 JQ245975 99
MBAi39CL Ankara KJ596523 554 JQ245975 99
MBAi184AG Alaşehir KJ596529 548 FJ904841 99
Neofusicoccum parvum MBAi27AG Manisa KF182330 581 JN135282 99
MBAi34AG Çorum KF798191 583 KC706919 99
MBAi44CL Salihli KJ596524 579 JN135282 99
MBAi105AG Turgutlu KJ596526 585 KF182330 100
MBAİ131AG Turgutlu KJ596527 581 JN135282 99
On the other hand, the genus-specific primer pair used in this study (BOT 100F / BOT 472R) amplified DNA of the 15 grapevine and 1 olive isolates. The amplified products’ size (for Bot isolates) was 372 bp in agarose gel electrophoresis but DNA samples of Fomitiporia mediterranea and Alternaria alternata could not be amplified (Figure 2). Discrimination of Botryosphaeriaceae fungi from Alternaria or the other wood colonizers was facilitated by using these primers, so these fungi were understood to be the members of Botryosphaeriaceae.
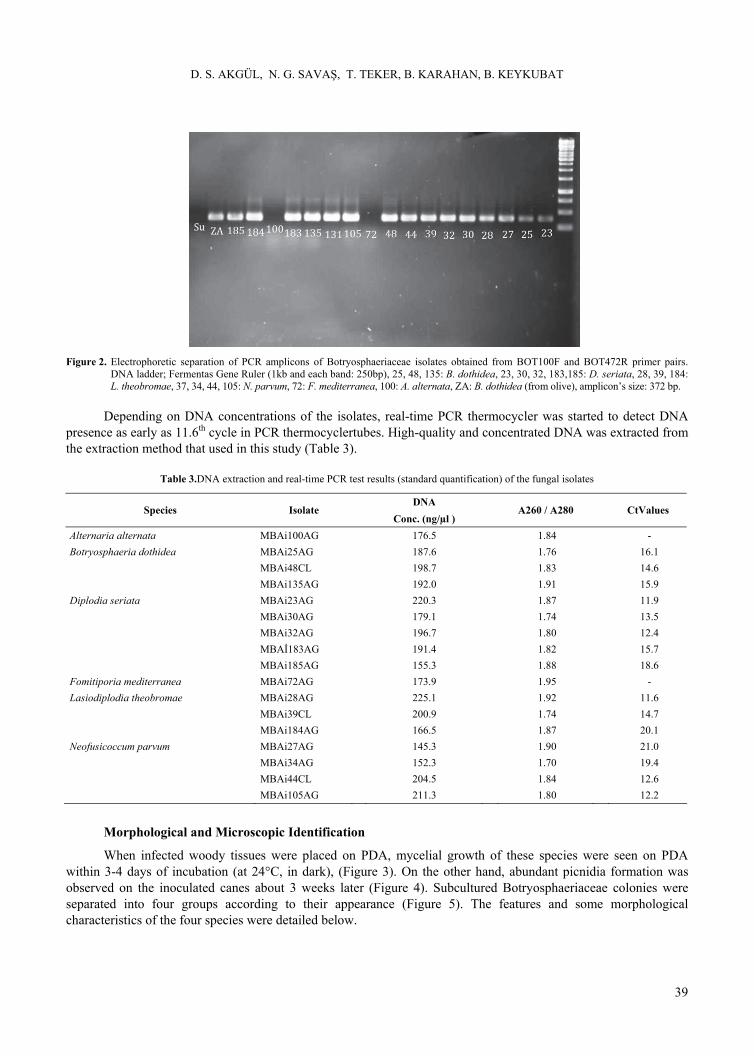
D. S. AKGÜL, N. G. SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT
39
Figure 2. Electrophoretic separation of PCR amplicons of Botryosphaeriaceae isolates obtained from BOT100F and BOT472R primer pairs.
DNA ladder; Fermentas Gene Ruler (1kb and each band: 250bp), 25, 48, 135: B. dothidea, 23, 30, 32, 183,185: D. seriata, 28, 39, 184: L. theobromae, 37, 34, 44, 105: N. parvum, 72: F. mediterranea, 100: A. alternata, ZA: B. dothidea (from olive), amplicon’s size: 372 bp.
Depending on DNA concentrations of the isolates, real-time PCR thermocycler was started to detect DNA presence as early as 11.6th cycle in PCR thermocyclertubes. High-quality and concentrated DNA was extracted from the extraction method that used in this study (Table 3).
Table 3.DNA extraction and real-time PCR test results (standard quantification) of the fungal isolates
DNA Species Isolate
Conc. (ng/µl ) A260 / A280
CtValues
Alternaria alternata MBAi100AG 176.5 1.84 -
Botryosphaeria dothidea MBAi25AG 187.6 1.76 16.1
MBAi48CL 198.7 1.83 14.6
MBAi135AG 192.0 1.91 15.9
Diplodia seriata MBAi23AG 220.3 1.87 11.9
MBAi30AG 179.1 1.74 13.5
MBAi32AG 196.7 1.80 12.4
MBAİ183AG 191.4 1.82 15.7
MBAi185AG 155.3 1.88 18.6
Fomitiporia mediterranea MBAi72AG 173.9 1.95 -
Lasiodiplodia theobromae MBAi28AG 225.1 1.92 11.6
MBAi39CL 200.9 1.74 14.7
MBAi184AG 166.5 1.87 20.1
Neofusicoccum parvum MBAi27AG 145.3 1.90 21.0
MBAi34AG 152.3 1.70 19.4
MBAi44CL 204.5 1.84 12.6
MBAi105AG 211.3 1.80 12.2
Morphological and Microscopic Identification
When infected woody tissues were placed on PDA, mycelial growth of these species were seen on PDA within 3-4 days of incubation (at 24°C, in dark), (Figure 3). On the other hand, abundant picnidia formation was observed on the inoculated canes about 3 weeks later (Figure 4). Subcultured Botryosphaeriaceae colonies were separated into four groups according to their appearance (Figure 5). The features and some morphological characteristics of the four species were detailed below.

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
40
Figure 3. Mycelial growth of a Botryosphaeriaceae Fungus
(Neofusicoccum parvum) around infected wood samples after 3-4 days incubation (at 24°C, in dark).
Figure 4. Picnidia formation on grapevine canes after 20-22 days of incubation.
Figure 5. Colony morphology of Diplodia seriata (a), Botryosphaeria dothidea (b), Neofusicoccum parvum (c) and Lasiodiplodia theobromae
(d) on PDA, after 20-25 days incubation.
Botryosphaeria dothidea (Moug.Ex Fr.) Ces.& De Not.
Colonies developing on PDA were initially colourless, turning dark olive from the centre with the dark colouration and it spread the entire colony with time. Upper view of the original (not subculted for many times) colony was generally white and it had a typical pattern (Figure 5b). The conidia were hyaline, smooth and aseptate so they were looked like a bowling lobut (Figure 6a-b).The conidia sizes were around 22.1 – 24.7 x 4.9 – 5.2 µm.

D. S. AKGÜL, N. G. SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT
41
Diplodia seriata De Not.
On PDA culture, colonies appear greyish-black with dense aerial mycelium on top but it becomes black at the bottom of petri plates (Figure 5a). Growth rate is very fast, so fungus covers petri plates within 4-5 days on PDA in optimal conditions. Conidia are initially hyaline, becoming dark brown and moderately thick-walled and they are generally aseptate but rarely one-septate (Figure 6 c-d).The conidia sizes were around 22.6 – 26.5 x 11.2 – 14.3 µm.
Lasiodiplodia theobromae (Pat.) Griff. & Maubl.
The colonies were grey-brown to black with dense fluffy aerial mycelia on the media (PDA), while the underside becomes black with time. The fungus produced dark brown picnidia and picniospore on PDA after 3-4 weeks of incubation (Figure 5 c). Conidia were sub-ovoid to ellipsoid, thick walled and their color was initially hyaline but turned dark brown with age (Figure 6 e-f). The most characteristic feature of conidia was that they had longitudinal strations and one-septate (when they were mature). The conidia sizes were about 23.7 – 27.1 x 12.7 – 14.1 µm.
Neofusicoccum parvum (Pennycook&Samuels) Crous, Slippers& A.J.L. Phillips
Original colony morphology of this species was white with fluffy aerial mycelium, turning pale olivaceous gray from the middle of colony after 3–4 days (Figure 5c). The aerial mycelium resembled a cloudy seeming and touched to the petri dish’s lid within 8-10 days. The bottom color of colony was olivaceous grey in the first week but it turned blackish color in time. Conidia shape was generally ellipsoidal with round apices (Figure 6 g-h). Their sizes were around 14.5 – 19.2 x 6.5 – 8.0 µm.
Pathogenicity Test Results
All of the Botrysphaeriaceae isolates used in pathogenicity tests were able to colonize woody tissues of the test plants. After 15-16 weeks, blackish-brown discolorations have been observed on woody tissues of young vines and the isolates could be re-isolated (by 95.6%) from necrotic tissues successfully.Lesions extended both upward and downward from the point of inoculation.Depending on virulence, each of the isolates produced moderate or severe lesions on infected plants. Mean lesion lengths were 17.2 mm for B. dothidea, 13.9 mm for D. seriata, 13.1 mm for L. theobromae and 83.5 mm for N. parvum, but it was only 6.3 mm for the control (Table 4). N. parvum was the most virulent one among these species and lesion length mean of this species was longer than that of other species statistically. Although slight wood discoloration was seen just around inoculation point of the control plants, none of Botryosphaeriaceae isolate could be isolated from these tissues.
Table 4. Mean lesion lengths produced by Botryosphaeriaceae species on Sultana Seedless grapevine plants.
Lesion Species Name
Lengths (mm)
Botryosphaeria dothidea 17.2 b*
Diplodia seriata 13.9 b
Lasiodiplodia theobromae 13.1 b
Neofusicoccum parvum 83.5 c
Sterile agarinoculation (Control) 6.3 a
* Mean values within a column are significantly different based on Duncan (0.05) multiple range test

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
42
40 X objective magnification 100 X objective magnification
Figure 6. Conidia shapes of Botryosphaeria dothidea (a-b), Diplodia seriata (c-d), Lasiodiplodia theobromae (e-f) and Neofusicoccum parvum (g-h)

D. S. AKGÜL, N. G. SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT
43
DISCUSSION
In this study, four Botryosphaeriaceae species were identified with classic and molecular biology methods. In classic identification, colony growth patterns, colony color, hyphal morphology and picniospore shapes were documented and these characteristics were also confirmed with the studies of Crous et al. (2006); Urbez-Torres et al. (2006); Amponsah, 2010; Pitt et al. (2010). Classic identification of Botryosphaeriaceae fungi relied on morphological features in the past. However many evaluations by several researchers of these fungi have revealed that these characteristics were very variable and often overlapped between species. Morphological characteristics can also be affected in the isolates that collected from different locations (Amponsah, 2010). Conclusively, it has been advised by many authors that DNA based molecular methods should be used for accurate identification of these fungi (Burruano et al., 2008).
The genus specific primer pairs (BOT100F/BOT472R) discriminated Botryosphaeriaceae fungi from the others successfully. Ridgway et al. (2011) found that these fungi could also be detected even in soil by using these primers.Moreover these primers may also be suitable for fast detection of these pathogens in grapevine propagation systems and in certification of young grafted plants. In addition, it may be useful for the first molecular discrimination before sequence analysis. Although the isolates were isolated from different locations, maximum identity (up to 100%) has been observed between two Neofusicoccum parvum isolates, MBAi27AG and MBAi105AG in sequence analysis. It can be concluded from this similarity that one of the isolates may have been transmitted from one location to another by vegetative propagation materials.
Among the identified isolates, two different virulence groups occurred in pathogenicity tests. N. parvum was in virulent and B. dothidea, D. seriata and L. theobromae were in moderately virulent group, so N. parvum produced longer lesions, when compared to B. dothidea, D. seriata and L. theobromae on woody tissues. Former studies indicated that there were three different groups in pathogenicities of Botryosphaeriaceae species. Neofusicoccum spp. and Lasiodiplodia spp. were described as highly virulent, B. dothidea and Diplodia spp. as moderately virulent and Dothiorella spp. and Spencermartinsia viticola as slightly virulent (van Niekerket al., (2004); Luque et al., (2009); Urbez-Torres and Gubler, (2009)). Our findings corroborate these studies in some extent, although we described our L. theobromae isolates as moderately virulent in our pathogenicity tests. Difference in virulence was attributed to toxin production of these species during pathogen growth in plant tissues. Martos et. al., (2008) found that Botryosphaeriaceae fungi were capable of producing hydrophilic high-molecular weight compounds with phytotoxic properties, which were thought to be exopolysaccharides. Their study showed that maximum toxin production was assessed from the Neofusicoccum spp. inoculated grapevines in pathogenicity tests. These findings may explain in some extent our pathogenicity results.
Botryosphaeriaceae canker is an important disease in Aegean Region Vineyards. According to the unpublished survey results, the incidence of disease has been estimated to reach 21.0% in some Sultana Seedless vineyards in Manisa and İzmir production areas (Akgül et al.). Identification is the first stage of disease management in all culture plants, and accurate identification is very important to launch correct disease control strategy.
ACKNOWLEDGEMENTS
This research was supported by İzmir Commodity Exchange and Manisa Viticulture Research Station, under the project name “Research on vine decline and bud burst disorders caused by fungal pathogens in Sultana Seedless vineyards of Manisa and İzmir Cities”. We thank Işınsu Kestelli, İlhan Zincircioğlu, Mustafa Yağcıoğlu, Dr. Selçuk Karabat and Akay Ünal for personal contribution.
ÖZET
Asmada Kol ve Sürgün Kurumalarina Neden Olan Botryosphaeriaceae Türlerinin Teşhisi ve Patojenisiteleri
Botryosphaeriaceae funguslarının neden olduğu kol, sürgün kurumaları (veya kangren) ve geriye ölüm hastalıkları, asmanın en önemli fungal gövde hastalıkları arasında yer almaktadır. Son yıllarda Türkiye’deki

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
44
asmalarda lokal kol kurumaları, sürgünlerde geriye ölüm ve odunsu dokularda “V” şeklindeki renk değişimi belirtileri önemli oranda artmıştır. 2012 yılı üretim sezonunda, Ankara, Çorum, Izmir ve Manisa’dan bu belirtileri taşıyan 15 odunsu doku örneği alınmış ve hastalığa neden olan fungal etmenleri saptamak için standart mikolojik izolasyonlar yapılmıştır. Bu örneklerden yüksek oranda (%69,4) Botryosphaeriaceae üyesi funguslar izole edilmiştir. İzolatların teşhisindeki ilk aşamada fungal DNA ekstrakte edilmiş ve Real-Time thermocycler’da cinse özgü primer çifti ile (BOT100F-BOT472R) çoğaltılmışlardır. Bu sayede Botryosphaeriaceae üyesi türler diğer benzer türlerden ayırt edilmişlerdir. İzolatların tür düzeyindeki moleküler tanısında ITS 1 ve ITS 2 ribosomal DNA bölgeleri diziletilmiş ve gen dizileri NCBI Gen Bankası’ndaki mevcut dizilerle karşılaştırılmıştır. Dizi analizi yapılan 15 izolat içerisinde 4 farklı türün; Botryosphaeria dothidea (Anamorf; Fusicoccum aesculi), Diplodia seriata (An; B. obtusa) Lasiodiplodia theobromae (An; B. rhodina) ve Neofusicoccum parvum (An; B. parva) bulunduğu tespit edilmiştir. Morfolojik ve mikroskobik tanıda türlerin PDA ortamı üzerindeki koloni morfolojileri ve odunsu dokulardaki pikniospor şekilleri incelenmiştir. Koch pastülatları için tüm izolatların patojenisitesi 1 yaşındaki tüplü asma fidanları üzerinde yapılmıştır. İnokulasyondan 4 ay sonra, odun dokulardaki lezyonların uzunluğu kontrole göre değerlendirilmiştir. Patojenisite testleri, Neofusicoccum parvum türünün kontrol ve diğer türlerle kıyaslandığında dokular üzerinde daha büyük lezyonlar meydana getirdiğini ortaya koymuştur.
Anahtar kelimeler: Botryosphaeriaceae, Asma, Gövde Hastalıkları
LITERATURE CITED
Akgül, D.S., Savaş, N.G., and Eskalen, A. 2014. First report of wood canker caused by Botryosphaeria dothidea , Diplodia seriata, Neofusicoccum parvum, and Lasiodiplodia theobromae on grapevine in Turkey. Plant Disease: 98(4): 568p.
Amponsah, N. T. 2010. Epidemiology of Botryosphaeriaceous species associated with grapevines in New Zealand. Lincoln University, PhD thesis, 251 p.
Burruano S., V. Mondello, G. Conigliaro, A. Alfonso, A. Spagnoloand L. Mugnai, 2008. Grapevinedecline in Italycaused by Lasiodiplodia theobromae. Phytopathologia Mediterranea, 47: 132–136.
Cenis, J. L. 1992. Rapid extraction of fungal DNA for PCR amplification. Nucleic Acid Research, 20 (9): 2380p.
Crous P.W., B. Slippers, M.J. Wingfield, J. Rheeder, W.F.O. Marasas, A.J.L. Phillips, A. Alves, T. Burguess, P. Barberand J.Z. Groenewald, 2006. Phylogeneticlineages in theBotryosphaeriaceae. Studies in Mycology, 55: 235–253.
Evidente A., B. Punzo, A. Andolfi, A. Cimmino, D. Melckand J. Luque, 2010. Lipophilic phytotoxins producedby Neofusicoccum parvum, a grapevine canker agent. Phytopathologia Mediterranea, 49: 74–79.
Luque J., S. Martos, A. Aroca, R. Raposoand F. Garcia- Fihueres, 2009. Symptoms and fungi associated with declining mature grapevine plants in northeast Spain. Journal of Plant Pathology,91: 381–390.
Martos S., A. Andolfi, J. Luque, L. Mugnai, G. Suricoand A. Evidente, 2008. Production of phytotoxic metabolites by five species of Botryosphaeriaceae causing decline on grapevines, with special interest in the species Neofusicoccum luteum and N. parvum. European Journal of Plant Pathology, 121: 451–461.
Pitt, W. M., Huang, R., Steel, C. C., Savocchia, S. 2010. Identification, distribution and current taxonomy of Botryosphaeriaceae species associated with grapevine decline in New South Wales and South Australia. Australian Journal of Grapeand Wine Research, 16: 258–271.
Ridgway, H.J., Amponsah, N.T., Brown, D.S., Baskarathevan, J., Jones, E.E., and Jaspers, M.V. 2011. Detection of Botryosphaerious species in environmental samples using a multi-species primer pair. PlantPathology, 60: 1118–1127.
Rolshausen P.E., L.C. Greve, J.M. Labavitch, N.E. Mahoney, R.J. Molyneux and W.D. Gubler, 2008. Pathogenesis of Eutypalata in grapevine: Identification of virulence factors and biochemical characterization of cordon dieback. Phytopathology, 98: 222–229.

D. S. AKGÜL, N. G. SAVAŞ, T. TEKER, B. KARAHAN, B. KEYKUBAT
45
Siebert, J. B. 2001. Eutypa: The economic toll on vineyards. Wines Vines Apr: 50-56.
vanNiekerk J.M., P.W. Crous, J.Z. Groenewald, P.H. Fourieand F. Halleen, 2004. DNA phylogeny, morphology and pathogenicity of Botryosphaeria species on grapevines. Mycologia,96: 781–798.
Wilcox W.F., 2011. Disease Management. In: 2011 New York and Pennsylvania Pest Management Guidelines for Grapes (T.H. Weigleand A.J. Muza, ed.). Cornell University Cooperative Extension, Geneva, NY, USA and Pennsylvania State University, College of Agricultural Sciences, State College, PA, USA. Online publication: http://ipmguidelines.org/Grapes/
White, White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Snisky JJ, White TJ, eds. PCR protocols: a guide to methods and applications. San Diego: Academic Press. p 315–322.
Úrbez-Torres, J. R., Leavitt, G. M., Voegel, T. M., andGubler, W. D. 2006. Identification and distribution of Botryosphaeria spp. associated with grapevine cankers in California. Plant Disease, 90:1490-1503.
Úrbez-Torres J.R. and W.D. Gubler, 2009. Pathogenicity of Botryosphaeriaceae species isolated from grapevine cankers in California. Plant Disease,93: 584–592.
Urbez-Torres, J. R. 2011. The status of Botryosphaeriaceae species infecting grapevines. Phytopathologia Mediterranea, 50 (Supplement), S5−S45.
Úrbez-Torres, J. R.,Peduto, F., Smith, R. J., andGubler, W. D. 2013. Phomopsis dieback: A grapevine trunk disease caused by Phomopsis viticola in California. Plant Disease, 97:1571-1579.

IDENTIFICATION AND PATHOGENICITIY OF BOTRYOSPHAERIACEAE SPECIES CAUSING WOOD CANKER ON GRAPEVINES
46

J. Turk. Phytopath., Vol. 42 No. 1-3, 47-56, 2013 ISSN 0378 - 8024
47
Combination of Irradiation and Sodium Carbonate to Control Postharvest Penicillium Decay of Apples
Cemile TEMUR* Osman TİRYAKİ **
* Department of Plant Protection, Faculty of Seyrani Agriculture, Erciyes University, Kayseri, 38039, Turkey
** Department of Plant Protection, Faculty of Agriculture, Çanakkale Onsekiz Mart Universitesi, Terzioğlu Campus, Çanakkale, 17020, Turkey.
Accepted for publication May 2, 2014
ABSTRACT
Penicillium expansum is an important pathogen causing considerable postharvest losses in apples. Due to some restrictions applied for chemicals, there is a need for new approaches and combined treatments for inhibition of fungi. In present study, O2 isolate of P. expansum, isolated from apples in cold storages of Kayseri, was used. Inoculated apples were irradiated with 3.0, 3.5 and 4.0 kGy and stored at 3-4ºC for 40 days. While the lesion diameter of untreated apples at 40th day was 20.28 mm, the apples treated with 3.5 kGy had a diameter of 7.85 mm. Differences in lesion diameters were found to be significant (p<0.05). In second part of study, inoculated apples were treated with sodium carbonate (SC,%3)+2.5 kGy, SC+3.0 kGy and SC+3.5 kGy combinations and stored at 3-4ºC for 40 days. At the 40th day, lesion diameters were measured as 49.00 mm and 19.18 mm for control samples and SC+3.0 kGy treatment, respectively. The area under the disease progress curve (AUDPC) was evaluated to calculate individual and total AUDPC. Long-term impacts of treatments on P.expansum were also evaluated by performing re-isolation from untreated and SC+3.0 kGy treatments. There were no significant differences between control and treated ones with regard to cultural characteristics and pathogenicity of fungus.
Key words: Postharvest disease; Penicillium expansum; irradiation; combined treatment
INTRODUCTION
Postharvest losses are estimated to be 30-40% in Turkey and sometimes it may reach to 50% (Karabulut and Baykal, 2004; Anonymous, 2013). The major postharvest pathogen of apples in Turkey is Penicillium expansum Link. Today, customers demand residue-free food products in one hand; several fungi develop resistance against common fungicides on the other hand. Alternative control strategies are under investigation due to health concerns with regard to chemicals. However, such alternatives have not been able to provide a control as effective as chemical agents have. Combinations of several alternatives increase their effectiveness and decrease negative effect of application by exposure to lower doses when compared to individual treatments (Beraha et al., 1960; Ragsdale and Sisler 1994; Tiryaki et al., 1994; Conway et al., 2004). Temur and Tiryaki (2013a) reported advantages and disadvantages of irradiation and combined treatments of sodium carbonate (SC), cold storage, chemicals, heat treatment, modified atmosphere packaging and bio-control agent in details.
The purpose of all food production techniques is to minimize decay and extend shelf life. Previous studies have demonstrated that irradiation, particularly gamma ray, produced from 60Co, is used for controlling postharvest decays and extending the shelf-life of fresh fruit (Mostafavi et al., 2011; Duvenhage et al., 2012).Gamma irradiation not only decreases postharvest decays and but also decreases the chemical needs for postharvest treatments (Geweely and Nawar 2006; Cia et al., 2007). The penetrating power of gamma rays is more than fungicides. They can uniformly penetrate deep into fruit and easily treat every kind of fruits regardless of the shape and size (Jarrett, 1982). Irradiation increases major phenolic compounds of fruit peels. Such an increase is related to phenylalanine

COMBINATION OF IRRADIATION AND SODIUM CARBONATE TO CONTROL POSTHARVEST PENICILLIUM DECAY OF APPLES
48
ammonia-lyase enzyme activity. Ionizing radiation can induce fruit resistance against pathogens and stimulate the biosynthesis of constitutive phenolic. Researchers also concluded that the direct effects of irradiation on fungal structures growing in the rind were more important for disease reduction than a possible indirect effect on fruit defense mechanisms. This assumption is further supported by considerable inhibition of fungi sporulation through irradiation (Palou et al., 2007). DNA fragmentation is the basic mechanism of irradiation to kill microbes. Complex organisms are more sensitive to irradiation than simple ones (Shea et al., 2000).
Lethal radiation doses required for pathogens in the host are higher than in the culture media (Beraha et al., 1960). Dose rate is also important for inhibition of fungal development. Beraha (1964) investigated the effect of dose rate and reported higher effects of high dose rates than low rates. Botrytis cinerea infection was inhibited with 1.25-1.50 kGy of irradiation with an implementation rate of 250 Gymin-1.Whereas the infection was not inhibited with 2kGy at 25Gy min-1.Darras et al., (2010) emphasized the importance of order of application. The irradiation after inoculation with B. cinerea was found to be more effective than before inoculation. The time elapsed between inoculation and irradiation is also significant point (Kuhn et al., 1968). An irradiation dose of 2kGy after 24 h of inoculation yielded 10% decrease in lesions over peaches. Researchers observed 60, 80 and 90% increase in lesion incidence by postponing irradiations 36, 48 and 60 h after inoculation, respectively. Low irradiation doses stimulated fungal development for both in vitro and in vivo. After 40 days of irradiation, lesion diameters were 36.21 mm and 34.75 mm respectively for 1 kGy and control treatment in Ankara pears inoculated with Penicillium expansum (Tiryaki and Maden 1991). Similarly, 1kGy stimulated aflatoxin incidence, whereas 3-4 kGy inhibited fungal and mycotoxin development (Kabak and Var 2005).
SC solutions have been used to control postharvest fungi on fruits since 1928 (Conway et al., 2004). It is an inexpensive readily available method with minimal risk of injury over the fruits (Palou et al., 2002; Zamani et al., 2009). Inhibitory activity of SC mostly depends on the presence of salt residues within the infected sites and the interactions between residual salt andpeel constituents. SC primarly act as fungistatic and not provide persistent protection if fruit are infected after treatment (Smilanick et al., 1999; Usall et al., 2008).
Palou et al., (2007) investigated the effects of X-ray and SC treatments on postharvest Penicillium decay in mandarins. SC treatment with 0.875 kGywas found to be more effective for the inhibition of fungus. P. digitatum lesion diameter was significantly reduced only by the highest dose of irradiation as 0.875 kGy with storage at 20C. In a recent study carried out with P. digitatum and P. italicum in mandarins revealed that low pressure-fruit rinsing is considered as an efficient method to eliminate adverse effects of SC treatments on quality parameters of mandarins. On the other hand, Smilanick et al., (1999) reported significant loss of SC effectiveness against green mold by rinsing. Palou et al., (2007) therefore observed higher SC effectiveness on un-rinsed Clementine mandarins.
While heat treatment is very effective in eradicating decay if infection occurs prior to heating, it has little protective effect if infection occurs after heating (Klein et al., 1997). Synergistic effects of heat and SC solutions to control Penicillium decay on citrus fruit were also observed in previous research (Smilanick et al., 1999; Palou et al., 2001). Heating SC solution until 50oC was considered as an effective treatment to control Penicillum decay in mandarins (Palou et al., 2002). Similar findings were also reported by Conway et al., (2004), suggesting the use of heated SC solution (38oC) against the P.expansum.
The safety of foods irradiated up to 10 kilogray (kGy, 1kGy=100 krad) is guaranteed by joint FAO/IAEA/WHO Expert Committee for Food Irradiation (JECFI) (Anonymous, 1987; WHO, 1999).The limit of 10 kG yis accepted in Turkish Food Codex in 1999 (Anonymous, 1999). Radioactivity is impossible since any chain reactions occur to make foods radioactive. Irradiation disappears when the energy source is removed (Brennand, 1995;Farkas, 2006).Unless the specified optimum doses are not exceeded for certain food stuff, ionizing radiation has minimal or almost zero effect on nutritional characteristics, taste and quality (Diehl and Josephson, 1994).
The aim of this study was to investigate the effects of combined treatment of SC and gamma irradiation on the growth of Penicillium expansum in artificially inoculated Golden Delicious apples. The most virulent P.expansum O2 isolate from cold storages of Kayseri Province was used to evaluate the effects of individual and combined treatments.

C. TEMUR, O. TİRYAKİ
49
MATERIALS and METHODS
Pathogens
Penicillium expansum was isolated from decayed Golden Delicious apples in cold storages of Kayseri. Ten P. expansum isolates were collected, namely, EL8 ve EL5 (Elbuz Apple Marketing Co.), EM3 ve EM10 (Eminsu Cold Storage Co.), G2 ve G9 (Gülüm Agriculture Cold Storage Co.), D1 ve D8 (Demir Medical Cold Storage Co.), O2 ve O3 (Special Provincial Administration Cold Storage Co). Pathogenicity test was performed with the 10 P.expansum isolates in our previous work (Temur and Tiryaki, 2013b). High virulent isolate of P. expansum was O2 isolate. This isolate was identified and cultured on potato dextrose agar (PDA Merck, KGaA Damstadt, Germany) in Petri dishes at 23±1 ºC for 7–10 days. A high-density conidial suspension (106conidiamL-1) was prepared in Tween 80 (0.05%, w v-1) in sterile water, passed through two layers of cheesecloth, measured with a hemacytometer, and diluted with sterile water to achieve the desired inoculum density (Palou et al., 2007).
Inoculation of apples with P.expansum
The “Golden Delicious” apples were surface disinfected with 0.5% NaOCl solution. Dried apples were wounded at two sites (the opposite sides of fruit tip with 50 mm distance) with a dissecting needle (1.5 mm diameter x 2.0 mm deep). On each fruit, both wounds were inoculated with P. expansum by immersing needle into a suspension of 106 conida mL-1 (Karabulut and Baykal 2004).
Irradiation treatments
Under in vitro conditions (culture media), effective gamma irradiation dose for P. expancum was found to be 3.0 kGy in a previous study (Temur and Tiryaki 2013b). Hence, 0.0 kGy (control) 3.0 kGy, 3.5 kGy, and 4.0 kGy were used in in vivo studies. Each treatment was applied to 8 replicates with 1 apples each.
The apples were inoculated with P. expansum by using the procedure described above, and placed polyethylene bags and stapled. They kept until the irradiation in a sample storage box at 3-4ºC. Irradiation was carried out by using 60Co gamma source located at the Sarayköy Nuclear Research and Training Center, Ankara, Turkey, with the 1026.2 Ci radioactivity and 0.668 kGyh-1 dose rate. Irradiation was performed by placing apples in 1 L irradiation chamber of 60Co gamma source. Irradiation application was illustrated in Fig. 1. After the irradiation, treated and untreated apples were incubated at 20ºC for 24 h, then stored at 3-4ºC for 46 days. Lesion diameters were measured again in 3-day intervals.
Figure 1. Irradiation of apples in 60Co gamma source.

COMBINATION OF IRRADIATION AND SODIUM CARBONATE TO CONTROL POSTHARVEST PENICILLIUM DECAY OF APPLES
50
Combined treatment of irradiation and sodium carbonate
About 2 hours after inoculation, apples were placed in stainless steel grid baskets and submerged into 22.5 L stainless steel buckets with aqueous 3.0% (w v-1) SC solutions for 150 s (Merck KGaA 64271 Darsmstadt/ Germany) at 40ºC (Palou et al., 2007). Temperature of the solutions was not allowed to change ± 0.5ºC during the treatments. Inoculated and SC-treated apples were placed in polyethylene bags and stapled. Each treatment was applied to 9 apples. SC-treated and control apples were incubated at 23±1ºC for 24 h and stored at 3-4ºC for 39 days. Lesion diameters were measured in 3-day intervals.
Irradiation dose of 2.5, 3.0 and 3.5 kGy were combined with SC (3%) to control P. expansum decay. Inoculated and SC-treated apples irradiated in accordance with the methods described above. Treated and untreated apples were incubated at 23±1ºC for 24 h, and then stored at 3-4ºC for 46 days. Lesion diameter of fruits was measured in 3-day intervals. In this part of experiment, the total number of treatment, including control samples (with neither SC nor irradiation treatment) are four. Similarly, each treatment was applied to 8 replicates with 1 apples each.
Long-term effects of combined treatment on pathogenicity and change on cultural characteristics of
P. expansum
P. expansum re-isolated from combine treated (3.0 kGy+3.0% SC) fruits and the fungus were cultured on PDA. For the pathogenecity test, apples were inoculated with re-isolated P. expansum suspension. Then, incubated at 23±1ºC for 24 h. Control samples were also passed through same methods. Each treatment was applied to 5 replicates with 1 apples each. Inoculation was performed as described above. After inoculation, one lot of fruit (5 replicates) was placed in storage at 3-4ºC and another similarly treated lot was stored at 23±1ºC. Later on, the diameter of lesions was measured in 3-day intervals (Tiryaki et al., 1994).
For observing of the sporulation of fungus, 5 mm agar disk was removed from re-isolated cultures and transferred to fresh PDA and incubated at 23±1ºC for 24 h. Each treatment was applied to 10 replicates and each was placed into separate petri dishes. The experiment was divided into two parts; 5 replicates were stored at 23±1ºC and another 5 replicates were stored at 3-4ºC. The diameters of colonies were measured in 3-day intervals.
Evaluations
SPPS software (SPSS Inc., Chicago, IL, USA) was used for statistical analysis. Duncan’s multiple range test was used to compare the means of treatments. The lesion diameters were used to calculate the area under the disease progress curve (AUDPC) by using Equation 1 (de Capteville et al., 2002).
AUDPC = ∑[(yi+ yi+1)/2 × (ti+1 – ti)] (1)
Where, yi is the diameter of a lesion at time ti, in days, yi+1 is the diameter of the lesion at time ti+1 and (ti+1-ti) is the number of days between two evaluations.
The total AUDPC assessment was performed by the procedure specified by Palau et al., (2007).
RESULTS
Result of irradiation treatments
The lesion sizes on irradiated apples with gamma rays at 3.0, 3.5 and 4.0 kGy are presented in Fig.2. Irradiation dose of 3.5 kGy significantly reduced lesion diameters at 37 and 40 days of cold storage. These effects were also found to be significant at p< 0.05 level. At 40th day, while the lesion diameter of untreated apples was 20.28 mm, the apples treated with 3.5 kGy had a diameter of 7.85 mm. Therefore, the effective dose was assumed as

C. TEMUR, O. TİRYAKİ
51
3.5 kGy in this part of experiment. In other words, gamma-ray dose of 3.5 kGy considerably reduced the size of decay lesion on apples.
Figure 2. Lesion sizes on apple artificially inoculated with the and irradiated with gamma rays and incubated at 3-4◦C for 37 and 40 days. Bars with same letter within same day were not significantly different at p< 0.05.
Disease control ability of combined treatments
A few researchers reported that combinations of several applications increased effectiveness of single treatments (Beraha et al., 1960; Tiryaki et al., 1994; Conway et al., 2004). In present study, the combined effects of SC and irradiation treatments were also evaluated on Golden Delicious apples. Inoculated and SC-treated apples were irradiated at three gamma-ray doses of 2.5, 3.0 and 3.5 kGy. Lesion diameter of apples, subjected to SC+3kGy treatment, at 37 and 40 days of measurement are shown in Fig.3. At 40th day, the diameter of lesions caused by P. expansum was significantly reduced from 49.00 mm (control apples: non-irradiated and non-SC-treated) to 19.18 mm (SC+3 kGy treatment). In addition, the effect of SC +3.0 kGy combination was also photographed in Fig.4 at 40th day of observation. Therefore, the SC+3.0 kGy treatment was determined as the optimum treatment for P. expansum decay control.
Figure 3. Lesion size on apple inoculated with the P. expansum and combined treated with SC+2.50 kGy, SC+3.0 kGy and SC+3.5 kGy and
incubated at 3-4◦Cfor 37and 40days. The bars with same letter within same day were not significantly different at p< 0.05.
In present work, complying with the previous ones, while the effective gamma radiation dose for controlling P. expansum decay in apple was found to be 3.5 kGy; SC (3%)+3.0 kGy (by exposure lower radiation dose) combination was considered as optimum treatment to contol P. expansum decay on apples. The efficiency of combined treatments was significantly superior to that of single treatment. Another issue to be highlighted here in is the larger lesion size in SC + 3.5 kGy than in SC + 3.0 kGy (Fig.4) since high irradiation doses may increase sensitivity of fruits to the fungal infection.

COMBINATION OF IRRADIATION AND SODIUM CARBONATE TO CONTROL POSTHARVEST PENICILLIUM DECAY OF APPLES
52
Figure 4. Lession development over apples inoculated with P. expansum andtreated with the combination of irradiation and SC, after 40 days storage at 3-4ºC.
Long-term effects of combined treatment on cultural characteristics and pathogenicity of
P. expansum
P. expansum was re-isolated from SC+3.0 kGy treated apples, which was found to be the optimum treatment in in-vivo studies, and development and sporulation of pathogen were evaluated. Colony diameters of P. expansum on PDA in Petri dishes were measured at both 3-4ºC and 23±1ºC incubation. In both incubation conditions, there were no significant differences between control and treated ones. Microscobic evaluations also did not show any differences on development of PDA, hypha and structure of spore.
For the pathogenecity test, apples were inoculated with re-isolated P. expansum from untreated and SC+3.0 kGy treated apples and incubated at 23±1ºC. The lesion diameter of apples for a storage period of 16 days was measured. Any difference was not observed between controls and treated ones. Lesion diameters on apples stored at 3-4ºC were also measured for a storage period of 54 days. There were no differences between control and SC + irradiation treatment with regard to lesion size.
This part of study revealed that the effect of irradiation+SC treatment on pathogen was not persistent. These findings support the idea assuming food irradiation is a physical application and disappears when the energy source is removed (Urbain, 1986; Farkas, 2006).
AUDPC evaluations
The area under the disease progress curve was calculated for each individual treatment and observation day (de Capdeville et al., 2002).The effect of irradiation on P. expansum development was evaluated with AUDPC

C. TEMUR, O. TİRYAKİ
53
calculated by using Equation 1 (Fig. 5). The figure indicates the lowest AUDPC value for 3.5 kGy treatment in all 3 observations.
Figure 5. AUDPC evaluation of irradiation treatment on P. expansum development.
Similarly, the effect of combined treatment (SC+irradiation) on P. expansum development was evaluated with AUDPC (Fig. 6). As it can be seen in the figure, the lowest AUDPC value was found at SC+3.0 kGy treatment in all 3 observations.
Figure 6. AUDPC evaluation of SC+ irradiation treatment on P. expansum development.
For the evaluation of AUDPC, Palou et al., (2007) used the total AUDPC values. Total AUDPC was calculated with the data obtained from irradiation treatment and irradiation+SC treatment parts of experiments (Fig.7). The data were subjected to Duncan’s multiple range test of SPSS software. Fig.7 shows reduced rate of AUDPC in 3.5 kGy irradiation treatment and SC+3.0 kGy treatment. But the differences between the 3.0 and 3.5 kGy dose was not significant in irradiation+SC treatment.

COMBINATION OF IRRADIATION AND SODIUM CARBONATE TO CONTROL POSTHARVEST PENICILLIUM DECAY OF APPLES
54
Figure 7. Total AUDPC values of irradiation and SC+irradiation treatment on P. expansum development. The bars with same letter within same treatment were not significantly different at p< 0.05.
DISCUSSION
Several advantages of combined treatments, including irradiation and SC, to control post harvest fungi are reported in literature. Therefore, an attempt was made in present study to evaluate the effects of combined gamma radiation and SC on growth of P.expansum in apples.
Although not included in objectives of present study, it was observed that ripening levels of apples was also a dominant factor in controlling postharvest decays. The more ripened the fruits are, the more they sensitive to pathogen. Lesion diameter at control apples on 37th day and 40th day reached up to 19.40 mm and 20.28 mm, respectively in initial experiment (Fig. 2). But the decay development was over the control apples at later experiment, 45.81 mm and 49.00 mm on 37th day and 40th day of observations, respectively (Fig. 3). Such differences were mainly due to different ripening levels of apples used in both experiments. There was one month between the two set of experiments. Therefore, irradiation and irradiation+SC combination were evaluated as separate experimental designs. That is why, not only gamma doses of 3.5 kGy and 3.0 kGy, but also 2.5 kGy was used considering varying effects of irradiation based on fruit ripening levels.
In present study, it can be finally concluded that SC treatment with 3.0 kGy irradiation was the optimum implementation to control postharvest decay of P.expansum in apples. However, any treatment to decrease postharvest losses should be focused not only on inhibition of pathogen, but also on quality of fruit. For instance, if any treatment inhibits pathogen, this treatment should not have adverse effect of organoleptic properties of treated fruit, and vice versa. Therefore these findings should also be supported by further studies on the evaluation for organoleptic properties of treated apples (Prakash et al., 2000).
ÖZET
Elmalarda Hasat Sonu Penicillium Çürüklüğünün Engellenmesi için Işınlama ve Sodyum Karbonat Kombine Uygulaması
P. expansum elmalarda hasat sonu kayıplara neden olan en önemli fungal patojendir. Fungusun engellenmesi için, kimyasalların kısıtlamasından dolayı yeni yaklaşımlara ve kombine uygulamalara ihtiyaç duyulmaktadır. Bu çalışmada Kayseri’deki soğuk hava depolarından elmalardan izole edilen O2 kodlu P. expansum izolatı kullanılmıştır. İnokule edilmiş elmalar 3.0, 3.5 ve 4.0 kGy dozlarda ışınlanmış ve 3-4ºC’de 40 gün muhafaza edilmiştir. 40. günde kontrol örneklerinde 20.28 mm, 3.5 kGy dozda ise 7.85 mm çürüme çapı gözlemlenmiştir. Lezyon çapları arasındaki farklılık önemli bulunmuştur (p<0.05). Çalışmanın ikinci bölümünde inokule edilmiş elmalara sodyum karbonat (SC)+2.5 kGy, SC+3.0 kGy ve SC+3.5 kGy uygulamaları yapılmış ve 3-4ºC’de 40 gün muhafaza edilmiştir. 40. günde hiçbir işlem görmemiş kontrol örneklerinde 49.00 mm lezyon çapı görülürken, SC+3.0 kGy uygulamasında bu değer 19.18 mm olmuştur. Hastalık gelişimini gösteren eğrinin altındaki alan (AUDPC) değerlendirmesi yapılarak, uygulamaların bireysel ve toplam AUDPC değerleri hesaplanmıştır. P. expansum üzerine uygulamaların uzun süreli etkileri, kontrol ve SC+3.0 kGy uygulamalarından re-izolasyon yapılarak değerlendirilmiştir. Fungusun kültürel özellikleri ve patojenisitesinde, uygulama yapılmış ve kontrol örnekleri arasında önemli bir farklılık bulunmamıştır.
Anahtar Kelimeler:Hasat sonu hastalıkları; Penicillium expansum; Işınlama; Kombine uygulamalar
ACKNOWLEDGEMENTS
This study presents is the partial results of M.Sc. project supported by Scientific Research Projects Commission of Erciyes University (Project No: FBY_11_3555).

C. TEMUR, O. TİRYAKİ
55
LITERATURE CITED
Anonymous, 1987. Int. Atomic Energy Agency, Food Irradiation. Newsletter11(2):6.
Anonymous, 1999. Gıda Işınlama Yönetmeliği. Resmi Gazete 06.11.1999, 23868.
Anonymous, 2013. Hasat sonrası kayıplar.http://www.bahcesel.com/forumsel/taze-meyve-ve-sebzelerin-muhafazasi/ 20385-taze-meyve-ve-sebzelerde-hasat-sonrasi/ (accessed 28.03.13).
Beraha, L., Ramsey,G.B., Smith M.A.,Wright,W.R.,1960. Gamma radiation dose response some decay pathogens. Phytopathology 50:474-476.
Beraha, L.,1964. Influence of gamma radiation dose rate on decay of citrus, pears, peaches on Penicillium italicum and Botrytis cinerea in vitro. Phytopathology 54(7): 755-759.
Brennand, C.P., 1995. Food Irradiation. Radiation Information Network’s. Idaho State University, http://www.physics.isu.edu/radinf/food.htm (accessed: 28.03.13).
Cia, P., Pascholati, S.F., Benato, E.A., Camili, E.C., Santos, C.A., 2007. Effects of gamma and UV-C irradiation on the postharvest control of papaya anthracnose. Postharvest Biology and Technology. 43:366–373.
Conway, W.S., Leverentz, B., Janisiewicz, W.F., Blodgett, A.B, Saftner,R.A., Camp, M.J., 2004. Integrating heat treatment, biocontrol and sodium bicarbonate to reduce postharvest decay of apple caused by Colletotrichum acutatumand P. expansum. Postharvest Biology and Technology. 34: 11–20.
Darras, A.I., Joyceb,D.C.,Terrya, L.A., 2010. Postharvest UV-C irradiation on cut Freesia hybrida L. inflorescences suppresses petal specking caused by Botrytis cinerea, Postharvest Biology and Technology.55: 186–188.
de Capdeville, G., Wilson, C.L., Beer,S.V., Aist, J.R., 2002. Alternative diseasecontrol agents induce resistance to blue mold in harvested ‘Red Delicious’apple fruit. Phytopathology 92: 900–908.
Diehl, J.F., Josephson, E.S., 1994. Assessment of wholesomeness of irradiated food: A Review. Acta Aliment.2(23): 195–214.
Duvenhage, A.J., Jooste, M., Johnson, S.A., 2012. Irradiation as a postharvest quarantine treatment for a new pluot cultivar. Scientia Horticulturae 147:64–70.
Farkas, J., 2006.Irradiation for better foods.Trends in Food Science andTechnology17:148-152.
Geweely, N.S.I., Nawar, L.S., 2006. Sensitivity to gamma irradiation of post-harvest pathogens of pear. Int. J. Agric. Biol. 8(6):710-716.
Harris, B., 1998. The battle to minimise losses due to mycotoxins. World Poultry Magazine on Production Prosessing and Marketing. 14(4): 52-54.
Jarrett, R.D., 1982. Isotope Radiation Sources. In: Josephson, E.S., Peterson, M.S.(Eds) Preservation of Food by Ionizing Radiation. CRC Press, Boca rato´n pp. 137–163.
Kabak, B., Var, I., 2005.Effect of irradiation on mould growth and mycotoxin control. Gıda30(1): 197-201.
Karabulut, O.A., Baykal, N., 2004. Integrated control of postharvest diseases of peaches with a yeast antagonist, hot water and modified atmosphere packaging. Crop Protection.23: 431–435.
Klein, J.D., Conway, W.S., Whitaker, B.D., Sams, C.E., 1997. Botrytiscinereadecay in apples is inhibited by posthar vestheat and calcium treatments. J. Am. Soc. Hort. Sci. 122, 91–94.
Kuhn, G.D, Merkley, M.S., Dennison, R.A., 1968.Irradiation inactivation of brown rot infections on peaches. Food Technology. 22(7): 91-92.
Lurie, S., Droby, S., Chalupowicz, L., Chalutz, E., 1995.Efficacy of Candida oleophilastrain182 in preventing Penicilliumexpansuminfection of nectarine fruits. Phytoparasitica. 23: 231–234.
Margosan, D.A., Smilanick,J.L., Henson, D.J., 1997. Combination of hot water andethanol to control postharvest decay of peaches and nectarines. Plant Disease. 81: 1405–1409.

COMBINATION OF IRRADIATION AND SODIUM CARBONATE TO CONTROL POSTHARVEST PENICILLIUM DECAY OF APPLES
56
Mostafavi, H.A., Mirmajlessi, S.M., Fathollahi, H., Vaheh Minassyan, V., Mirjalili, S.M., 2011. Evaluation of gamma irradiation effect and Pseudomonas flourescens against Penicillium expansum. African Journal of Biotechnology.10(54): 11290-11293.
Palou, L., Smilanick, J.L, Usall, .J., Vinas, I., 2001. Control of postharvest blue and green molds of orange by hot water, sodium carbonate, and sodium bicarbonate. Plant Disease. 85(4): 371-376.
Palou, L., Usall, J., Munoz, J., Smilanick, J.L., Vinas, I., 2002. Hot water, sodium carbonate, and sodium bicarbonate for the control of postharvest green and blue molds of clementine mandarins. Postharvest Biology and Technology. 24: 93–96.
Palou, L., Marcilla, A., Rojas-Argudo, C., Alonso, M., Jacas,J.A., Angel del Rio, M., 2007. Effects of X-ray irradiation and sodium carbonate treatments on postharvest Penicilliumdecay and quality attributes of clementine mandarins.Postharvest Biology and Technology. 46: 252–261.
Prakash, A., Inthajak, P., Huibregtse, H., Caporaso, F., Foley, D.M., 2000. Effect of low dose gamma irradiation and conventional treatments on shelf life and quality characteristics of diced celery. J. Food Sci. 65: 1070-1075.
Ragsdale, N.N., Sisler, H.D., 1994. Social and political implications of managing plant diseases with decreased availability of fungicides in the United States. Annu. Rev. Phytopathology. 32: 545–557.
Shea, K.M., M.D., M.P.H., the Committee on Environmental Health, 2000. Technical Report: Irradiation of Food. American Academy of Pediatrics. Pediatrics106(6): 1505-1510.
Smilanick, J.L., Margosan, D.A., Mlikota, F., Usall, J., Michael,I.F., 1999. Control of citrus green mold by carbonate and bicarbonate salts and the influence of commercial postharvest practices on their efficacy. Plant Disease. 83: 139–145.
Temur, C.,Tiryaki, O., 2013a. Irradiation alone or comined with other alternative treatments to control postharvest diseases. African Journal of Agricultural Research8(5): 421-434.
Temur, C., Tiryaki, O., 2013b). Effect of gamma irradiation on Penicillium expansum isolated from “Golden Delicious” apples. The Journal of Turkish Phytopathol. 40(1-3):1-11.
Tiryaki, O., Maden, S., 1991. Penicillium expansum, Botrytis cinerea ve Rhizopus nigricans ile enfekteli Ankara armutlarında gamma radyasyonunu ile standart depolama koşullarında çürümenin engellenmesi. VI. Türkiye Fitopatoloji Kongresi, 7-11 Ekim, İzmir.
Tiryaki, O., Aydın,G., Gürer, M., 1994. Post-harvest disease control of apple, quince, onion and peach, with radiation treatment, The Journal of Turkish Phytopathol. 23 (3): 143-152.
Urbain, W. M., 1986. Food Irradiation. Academic Press Inc., Orlando, pp.1-351.
Usall, J., Smilanic, J., Palou, L., Denis-Arrue, N., Teixidó, N., Torres,R., Viñas, I., 2008. Preventive and curative activity of combined treatmentsof sodium carbonates and Pantoea agglomerans CPA-2 tocontrol postharvest green mold of citrus fruit.Postharvest Biology andTechnology. 50: 1–7.
WHO (World Health Organization), 1999. High-dose irradiation: wholesomeness of food irradiated with doses above 10 kGy. Report of a Joint FAO/IAEA/WHO Study Group, Technical Report Series N_890. Geneva, Switzerland: WHO.
Zamani, M., Sharifi Tehrani, A., Ahmadzadeh, M., Hosseininaveh,V., Mostofy, Y., 2009. Control of Penicillium digitatum on orange fruit combining Pantoea agglomerans with hot sodium bicarbonate dipping. Journal of Plant Pathology 91(2): 437-442.


















