The influence of excitotoxic basal ganglia lesions on motor - Brain
17
Brain (2000), 123, 1442–1458 The influence of excitotoxic basal ganglia lesions on motor performance in the common marmoset A. Lisa Kendall, F. David, G. Rayment, Eduardo M. Torres, Lucy E. Annett and Stephan B. Dunnett Department of Experimental Psychology and Cambridge Correspondence to: A. Lisa Kendall, Department of Centre for Brain Repair, University of Cambridge, Experimental Psychology, University of Cambridge, Cambridge, UK Downing Street, Cambridge CB2 3EB, UK E-mail: alk1001@cus.cam.ac.uk Summary Huntington’s disease is a genetically inherited neurodegenerative disorder for which currently there is no effective treatment or cure. In order to gauge the potential therapeutic benefits of neuroprotective or restorative treatments, it is necessary to create an animal model that is associated with readily measurable and long-lasting functional impairments. The undifferentiated neostriatum and limited behavioural repertoire of rodents have led to the extension of our investigations into the common marmoset. We have used quinolinic acid to create unilateral excitotoxic lesions of the caudate nucleus or the putamen in this small non-human primate. Keywords: striatum; Huntington’s disease; excitotoxin; primate Abbreviations: DARPP-32 dopamine- and adenosine-3:5-monophosphate-regulated phosphoprotein; GFAP glial fibrillary acidic protein; TBS Tris-buffered saline Introduction The basal ganglia are a group of subcortical nuclei that are involved in the parallel processing of descending cortical information that is relayed back to the cortex via the thalamus (Alexander et al., 1986; Alexander and Crutcher, 1990a; Wichmann and DeLong, 1996). Human and animal studies have demonstrated an involvement of the basal ganglia in motor sequencing, response selection and attentional processes (Alexander, 1994; Graybiel, 1998; Jueptner and Weiller, 1998; Lawrence et al., 1998). We owe much of our understanding of the role of the basal ganglia to the knowledge gained from various neurodegenerative disorders such as Parkinson’s disease and Huntington’s disease. The dopamine depletion associated with the loss of nigrostriatal neurons in Parkinson’s disease produces a clinical syndrome that includes tremor, bradykinesia and impairments in gait and movement initiation (for a review, see Lang and Lozano, 1998). Similarly, striatal pathology in Huntington’s disease is associated with a complex disorder where patients exhibit motor, cognitive and psychiatric disturbances (Vonsattel et al., 1985; de la Monte et al., 1988; Sharp and Ross, 1996; Harper, 1996). © Oxford University Press 2000 Following rigorous investigation of each monkey on a battery of behavioural tests, we found that the unilateral putamen lesion was associated with a contralateral motor impairment that persisted for at least 9 months and withstood repeated testing. However, the unilateral caudate nucleus lesion did not appear to be associated with any detectable motor deficit. The stability and the reproducibility of the unilateral putamen lesion in the marmoset provide a suitable tool for the investigation of potential treatments for neurodegenerative disorders that attack this region of the brain. Huntington’s disease is a genetically inherited disorder for which currently there is no effective treatment or cure (Huntington’s Disease Collaborative Research Group, 1993; Aronin et al., 1999; Reddy et al., 1999). One feature of Huntington’s disease that has received much attention is that, early in the course of the disease, there is a loss of the striatal medium spiny GABAergic projection neurons (Vonsattel and DiFiglia, 1998) yet the NADPH-diaphorase/neuropeptide Y/ somatostatin aspiny interneurons and the large cholinergic interneurons remain relatively preserved (Ferrante et al., 1987; Beal et al., 1988). In parallel, it was found that intra- striatal injections of the endogenous NMDA (N-methyl-D- aspartate) agonist quinolinic acid produced lesions that had a necrotic core but around that core was a transition zone where the neuropeptide Y/somatostatin and the cholinergic neurons were selectively spared (Beal et al., 1986, 1989). This, and the fact that there is a loss of putaminal NMDA receptors early in the course of the disease, led to the suggestion that the cells actually may succumb by way of an excitotoxic mechanism (Kowall et al., 1987; Young et al.,
Transcript of The influence of excitotoxic basal ganglia lesions on motor - Brain

Brain (2000), 123, 1442–1458
The influence of excitotoxic basal ganglia lesionson motor performance in the common marmosetA. Lisa Kendall, F. David, G. Rayment, Eduardo M. Torres, Lucy E. Annett and Stephan B. Dunnett
Department of Experimental Psychology and Cambridge Correspondence to: A. Lisa Kendall, Department ofCentre for Brain Repair, University of Cambridge, Experimental Psychology, University of Cambridge,Cambridge, UK Downing Street, Cambridge CB2 3EB, UK
E-mail: [email protected]
SummaryHuntington’s disease is a genetically inheritedneurodegenerative disorder for which currently there isno effective treatment or cure. In order to gauge thepotential therapeutic benefits of neuroprotective orrestorative treatments, it is necessary to create an animalmodel that is associated with readily measurable andlong-lasting functional impairments. The undifferentiatedneostriatum and limited behavioural repertoire of rodentshave led to the extension of our investigations into thecommon marmoset. We have used quinolinic acid tocreate unilateral excitotoxic lesions of the caudate nucleusor the putamen in this small non-human primate.
Keywords: striatum; Huntington’s disease; excitotoxin; primate
Abbreviations: DARPP-32 � dopamine- and adenosine-3�:5�-monophosphate-regulated phosphoprotein; GFAP � glialfibrillary acidic protein; TBS � Tris-buffered saline
IntroductionThe basal ganglia are a group of subcortical nuclei that areinvolved in the parallel processing of descending corticalinformation that is relayed back to the cortex via the thalamus(Alexander et al., 1986; Alexander and Crutcher, 1990a;Wichmann and DeLong, 1996). Human and animal studieshave demonstrated an involvement of the basal ganglia inmotor sequencing, response selection and attentionalprocesses (Alexander, 1994; Graybiel, 1998; Jueptner andWeiller, 1998; Lawrence et al., 1998). We owe much of ourunderstanding of the role of the basal ganglia to the knowledgegained from various neurodegenerative disorders such asParkinson’s disease and Huntington’s disease. The dopaminedepletion associated with the loss of nigrostriatal neurons inParkinson’s disease produces a clinical syndrome that includestremor, bradykinesia and impairments in gait and movementinitiation (for a review, see Lang and Lozano, 1998). Similarly,striatal pathology in Huntington’s disease is associated witha complex disorder where patients exhibit motor, cognitiveand psychiatric disturbances (Vonsattel et al., 1985; de laMonte et al., 1988; Sharp and Ross, 1996; Harper, 1996).
© Oxford University Press 2000
Following rigorous investigation of each monkey on abattery of behavioural tests, we found that the unilateralputamen lesion was associated with a contralateral motorimpairment that persisted for at least 9 months andwithstood repeated testing. However, the unilateralcaudate nucleus lesion did not appear to be associatedwith any detectable motor deficit. The stability and thereproducibility of the unilateral putamen lesion in themarmoset provide a suitable tool for the investigation ofpotential treatments for neurodegenerative disorders thatattack this region of the brain.
Huntington’s disease is a genetically inherited disorder forwhich currently there is no effective treatment or cure(Huntington’s Disease Collaborative Research Group, 1993;Aronin et al., 1999; Reddy et al., 1999). One feature ofHuntington’s disease that has received much attention is that,early in the course of the disease, there is a loss of the striatalmedium spiny GABAergic projection neurons (Vonsattel andDiFiglia, 1998) yet the NADPH-diaphorase/neuropeptide Y/somatostatin aspiny interneurons and the large cholinergicinterneurons remain relatively preserved (Ferrante et al.,1987; Beal et al., 1988). In parallel, it was found that intra-striatal injections of the endogenous NMDA (N-methyl-D-aspartate) agonist quinolinic acid produced lesions that hada necrotic core but around that core was a transition zonewhere the neuropeptide Y/somatostatin and the cholinergicneurons were selectively spared (Beal et al., 1986, 1989).This, and the fact that there is a loss of putaminal NMDAreceptors early in the course of the disease, led to thesuggestion that the cells actually may succumb by way ofan excitotoxic mechanism (Kowall et al., 1987; Young et al.,

Basal ganglia lesions in the marmoset 1443
1988). Since that discovery, quinolinic acid has been used toinvestigate the anatomical and functional consequences ofstriatal lesions in both rodents and primates (for examples,see Sanberg et al., 1989; Block et al., 1993; Ferrante et al.,1993; Storey et al., 1994; Brasted et al., 1998). However, inprimate studies, there has been relatively little investigationof the long-term, non-drug-induced behavioural effects ofexcitotoxic striatal lesions. Instead, the main aim has beento concentrate upon the development of primate models thatexhibit the chorea and dyskinesia frequently seen in theearly course of Huntington’s disease (Hantraye et al., 1990;Kanazawa et al., 1990; Burns et al., 1995).
The present study was undertaken to provide a directcomparison of the functional and anatomical consequencesof unilateral quinolinic acid lesions of the caudate nucleusor the putamen in the common marmoset. We wish to developa striatal lesion that is associated with long-lasting and readilymeasurable functional deficits that can be used as a stablebaseline against which to gauge the potential therapeuticbenefits of neural transplantation and neuroprotectivestrategies. The use of unilateral lesions allows each animalto act as its own control, allows explicit comparison betweenthe functional effects of selective caudate versus putamenlesions, and leaves one side of the brain intact to reduce thegeneral debility associated with bilateral damage. We reporton the utility of an extensive battery of motor functiontests, first developed to investigate the effects of unilateralnigrostriatal lesions and middle cerebral artery occlusions inthe marmoset (Annett et al., 1992a, 1994; Marshall andRidley, 1996), to characterize the deficits associated withselective striatal lesions in the marmoset.
MethodsQuinolinic acid lesionsTwelve common marmosets (Callithrix jacchus), seven malesand five females, aged between 2 and 3 years were used inthis study. The animals were housed in a temperature-controlled room (25 � 2°C) that was kept on a 12 h light–12 h dark cycle. The monkeys were fed daily on speciallyprepared sandwiches (containing egg, high protein dietsupplement and powdered primate pellets) and one piece ofapple or banana, plus once weekly forage mixes. All monkeyshad free access to water. All of the experimental procedures,and the animals’ health and welfare, were monitored inaccordance with the UK Animals (Scientific Procedures) Act1986 and associated codes of practice.
Prior to the lesion, each monkey was pre-trained and testedon all of the behavioural measures except for apomorphinerotation (see below). The monkeys were divided into threegroups of four, caudate (C15, C16, C22 and C24), putamen(P12, P14, P20 and P21) and sham, where two of the shammonkeys received sham surgery according to the caudateprotocol (SC18 and SC25) and the other two according tothe putamen protocol (SP17 and SP23). After pre-medication
Table 1 Lesion coordinates
Site AP ML DV
Caudate 1 12.5 3.5, 2.5 12.25, 11.25Caudate 2 11.5 2.0, 3.25 12.0, 12.75Caudate 3 10.5 2.5, 3.5 12.25, 13.0Caudate 4 9.5 2.25, 3.25 12.25, 13.25Caudate 5 8.0 4.0, 2.25 13.5, 12.75Putamen 1 11.5 5.25, 4.5 11.0, 10.25Putamen 2 10.5 5.25, 6.0 10.25, 11.0Putamen 3 9.5 5.7, 5.0 11.2, 9.5Putamen 4 8.5 6.25, 5.5 11.25, 10.5Putamen 5 7.5 6.25, 6.5 10.75, 9.75Putamen 6 6.5 6.5 10.25
AP � anterior–posterior; ML � medial–lateral ; DV � dorsal–ventral.
with 0.05 ml of ketamine (Vetalar®, Parke-Davis Veterinary),stereotaxic surgery was conducted under Saffan® (Pitman-Moore, UK) anaesthesia at a dose of 0.1 ml/100 g. Thecaudate group received 10 injections of 0.15–0.25 µl of0.1 M quinolinic acid (Sigma), in 0.1 M phosphate buffer(pH 7.4), and the final amount of toxin injected was 200nmol. The lesion coordinates were based on the marmosetatlas of Stephan and colleagues (Stephan et al., 1980) andare given in Table 1. The putamen group received 205 nmolquinolic acid delivered in 11 injections of 0.15–0.25 µl of0.1 M quinolic acid (coordinates in Table 1). The sham-operated monkeys were injected with appropriate volumesof 0.1 M phosphate buffer (pH 7.4). All of the injectionswere conducted using a 2 µl Hamilton syringe with a 28gauge dome-tipped needle, and the toxin or vehicle wasinjected at a rate of 0.1 µl per min. Any postoperativeseizures were controlled using 0.05–0.2 ml injections ofdiazepam (Valium, Roche, 5 mg/ml), and the monkeys werekept in a darkened room until the following day when, oncefully recovered, they were returned to the home cage.
Behavioural analysis of unilateral striatallesions in the marmosetThe staircase taskThe staircase apparatus consists of a Perspex© box fitted ontothe home cage door (illustrated in Fig. 1A). A central dividerprevents the animals reaching across the front of the apparatus,and a triangular segment fitted at the back of the box reducesmovements to the sides, providing separate measures for thereaching ability of each hand without the need for restraint(Marshall and Ridley, 1996). The monkeys were trained toretrieve small pieces of bread soaked in glucose syrup, andone reward was placed on each step on the left and/or rightstaircase. The monkeys were allowed a maximum time of300 s to clear the staircase for both bilateral and unilateraltrials. This task required minimal pre-training and wasconducted pre-lesion and on a monthly basis up to 10 monthspost-lesion. Each test session was conducted such that a

1444 A. L. Kendall et al.
Fig. 1 Pictorial representations of each staircase. (A) The normalstaircase (reprinted with permission from Nature Medicine;Kendall et al., 1998) has an opening on each side and a centralbarrier to prevent reaching across the centre. (B) The centralstaircase (reprinted with permission from ExperimentalNeurology; Marshall et al., 1999) has only one central openingthereby forcing the monkey to cross over and use the contralateralhand in ipsilateral body space. Both staircases have a triangularback piece (not shown) which prevents movement to the sideonce inside the apparatus.
bilateral trial was followed by two unilateral trials (one oneach side) and this was repeated twice such that the monkeyperformed nine trials and had to retrieve a total of 60 rewards.All test sessions were videotaped to allow full analysis ofall aspects of the monkeys’ performance.
The central staircase taskThis task involved a slight modification to the apparatus usedin the standard staircase task such that there was only onecentral opening into the front and each stair ascended up thelateral edge of the Perspex© (see Fig. 1B). The single, central,opening means that the monkey had to perform crossoverreaches such that the ipsilateral hand had to reach into thecontralateral half of the monkey’s body space therebyallowing a direct measurement of sensorimotor neglect tocomplement the motor assessment performed in the previoustask. The rewards were the same and the monkey was stillallowed 300 s per trial, but this time each of the six trialsper session was bilateral. This task was conducted with the
same frequency as the staircase task, and all test sessionswere videotaped.
Tube reaching taskThe next test of skilled motor function was a tube reachingtask (Annett et al., 1994). This provides a more stringentassessment of individual hand-reaching ability whilerestricting use of the opposite hand. The opposite hand waswrapped in gauze that was then fixed in place using stickingplaster. The monkeys used their free hand to reach into atube where sugared bread rewards were placed inside a small,16 mm diameter, tube and the monkey was allowed amaximum of 2 min to retrieve the reward. Each monkeyundertook a series of trials at different reaching distances,starting at 1 cm and increasing in 1 cm steps to a maximumof 6 cm. This task was conducted pre-lesion and at 1, 2, 3,6 and 9 months post-lesion.
Sticky label taskThis task is a sensitive measure of sensorimotor neglect aswell as motor coordination and has been used extensively toassess the functional consequences of dopamine depletion inrats and marmosets (Schallert et al., 1982; Annett et al.,1992a, 1994). A cuff of adhesive postal label ~6 cm longand 2 cm wide was wrapped around each foot, the monkeywas returned to the home cage and then allowed a maximumof 10 min to remove each label. The investigator observedfrom in front of the cage and recorded the times of contactand removal as well as the number of bites and scratchestaken to remove each label. This task was performed pre-lesion and at 1, 2, 3, 6 and 9 months post-lesion.
RotationAlthough the exact mechanism which causes rotation isunknown, it has been shown that excitotoxic lesions, whichdestroy intrinsic striatal output neurons, induce an ipsilateralrotation bias following activation by either direct (e.g.apomorphine) or indirect (e.g. amphetamine) dopamineagonists in rats (Schwarcz et al., 1979; Dunnett et al., 1988).Three separate measures of rotational bias were used in thisstudy, and for each assessment the monkey was placed in aspecially designed observation cage (43 � 55 � 44 cminternal dimensions) that had a clear front and top to allowfilming. Prior to the start of all rotation tests, each monkeywas placed in the observation cage on at least six separateoccasions for a total of at least 3 h for habituation to thecage, the video camera, the lighting and the room. Drug-independent rotation was measured by injecting 0.1 ml/300 gof 0.9% saline intramuscularly and then the monkey wasplaced in the observation cage for 60 min after the injection.The effects of dopamine imbalance were measured using twodrugs and there was at least 3 days between the administrationof either of the drugs. The mixed dopamine agonist

Basal ganglia lesions in the marmoset 1445
apomorphine (apomorphine-HCl, Sigma) was injected at adose of 0.5 mg/kg intramuscularly and the monkey wasobserved for 60 min. The indirect agonist amphetamine(D-amphetamine, Sigma) was also injected at a dose of 0.5mg/kg intramuscularly but this time the monkey was firstreturned to the home cage for 30 min and then placed in theobservation cage for a 30 min filming period. For all threerotational measures, the total number of 360° turns recordedon the videotape was counted and the net rotation score wascalculated (ipsilateral–contralateral). Saline and amphetaminemeasures were conducted pre-lesion and all three rotationmeasures were conducted at 1, 2, 3, 6 and 9 months post-lesion. Apomorphine rotation was not measured pre-lesionas this dose is particularly high and high levels ofapomorphine can be associated with distressing behavioursin the intact marmoset (Ridley et al., 1980). However, inpilot studies, it was the only dose found to result in consistentlevels of ipsilateral rotation.
Anatomy of unilateral striatal lesions in themarmosetFollowing completion of the behavioural assessments, themarmosets were terminally anaesthetized with a lethal doseof barbiturate (Expiral) and transcardially perfused with apre-wash solution of 0.1 M phosphate buffer followed by4% paraformaldehyde which was prepared in 0.1 M phosphatebuffer at pH 7.3. After fixation, the brains were removed andthen post-fixed in paraformaldehyde for 24 h. The brainswere cryopreserved in a 25% sucrose solution until theysank, and then were sectioned on a freezing microtome at athickness of 60 µm. One in six sections were stained usinga Nissl stain (1% cresyl fast violet) for the assessment ofcell viability and general morphology. A second series ofsections (1 : 6) were stained using a modified thiocholinesilver precipitation method (Koelle, 1955) to visualize theactivity of acetylcholinesterase (AChE; for protocol seeFricker et al., 1997a). Immunohistochemical staining wasconducted on free-floating sections. The sections (1 : 12series) were quenched with 10% hydrogen peroxide/10%methanol in distilled water, washed three times in Tris-buffered saline (TBS), pH 7.4, and then placed in TBS,containing 3% normal goat serum/0.2% Triton-X 100, for1 h before three further washes in TBS. Antibodies werediluted in TBS containing 1% normal goat serum/0.2%Triton-X 100.
Sections were then incubated for 60 h at 4°C in primaryantibodies to dopamine- and adenosine-3�:5�-monophos-phate-regulated phosphoprotein (DARPP-32, 1 : 20 000, akind gift from Drs P. Greengard and H. Hemmings), tyrosinehydroxylase (1 : 3000, Institute Jacques Boy, France) andglial fibrillary acidic protein (GFAP, 1 : 2000, DAKO). Thiswas followed by three further washes with TBS andincubation in either goat anti-mouse or goat anti-rabbitbiotinylated secondary antibodies (DAKO, UK) for 2 h. After
further washing in TBS, the sections were incubated instreptavidin–biotin complex (Vectastain ABC kit, VectorLaboratories, Burlingame, Calif., USA) in TBS for 1.5 h.Sections were washed in TBS followed by Tris non-salinebuffer, pH 7.4, and then developed in a 1 : 20diaminobenzidine solution (Sigma) with 0.002% hydrogenperoxide. Sections were mounted on gelatin-coated slides,dehydrated in alcohols, cleared in xylene and coverslippedusing DPX mountant.
The areas of remaining tissue on the lesioned side and theareas of intact caudate nucleus and putamen on the non-lesioned side were measured on DARPP-32-stained sectionsusing a Seescan image analysis system (Seescan ImageAnalysis, Cambridge). A line was traced around the outeredge of the caudate or the putamen using 1 : 6 sections fromthe most rostral extent of each region to the post-commissurallevel of 7.5 as defined on the atlas of Stephan and colleagues(Stephan et al., 1980). A threshold function was used tostandardize the signal obtained from the DARPP-32-positiveareas. The area (in mm2) was then used to estimate caudateor putamen volume as the sum of cross-sectional areasmultiplied by the distance interval between measured sections.
Statistical analysesFull analysis was conducted using two or three factor analysesof variance with the three groups as the between-subjectfactor and the test session time points and ipsilateral versuscontralateral as within-subject factors. All analyses wereperformed using Genstat 5 software. Where appropriate,post hoc analyses were performed by Newman–Keuls (andassociated t-tests) to correct for multiple comparisons.
ResultsBehaviourIn the immediate post-lesion phase, the putamen-lesionedmonkeys exhibited contralateral limb dystonia whichpersisted for 24–48 h after the surgery. After that time, noneof the lesioned monkeys exhibited any noticeable limbweakness and all monkeys maintained a constant, healthybody weight. There was no evidence of spontaneousdyskinesias as reported in lesion studies with larger primates(Brownell et al., 1994; Storey et al., 1994; Burns et al.,1995). One of the sham-lesioned monkeys (caudate protocol)had to be killed at 8 months post-surgery because of anunrelated illness and, consequently, there are behaviouraldata for only three sham monkeys for months 8–10.
The most marked motor impairments detected on eachbehavioural task were in the putamen-lesioned group. Incontrast, the caudate-lesioned monkeys appeared to exhibitlittle or no sensorimotor deficits. The deficits shown by theputamen-lesioned monkeys were immediately apparent onthe first post-lesion assessment at 1 month and, despite someminor practice effects, particularly on the staircases, the

1446 A. L. Kendall et al.
Fig. 2. Data measurements taken from the staircase task. In each graph, the open symbols or bars represent the ipsilateral side and theclosed symbols and bars represent the contralateral side. In (A), the mean (� SEM) latency to clear each side of the staircase, measuredover all trials, is shown for each of the three groups. When trials were not completed, the maximum latency of 300 s was ascribed tothat trial. There was a significant effect of the putamen lesion such that the time taken to clear the contralateral side was greater thanthat taken to clear the ipsilateral side and also greater than that taken to clear each side in both of the other groups. This effect of theputamen lesion was stable for the entire post-lesion assessment period. (B) The mean (� SEM) number of reaches recorded for eachtrial where a reach was counted each time the monkey formed a complete grasp with its hand when the arm was inside the staircase.The lateralized motor impairment seen in the putamen group produced excessive clumsiness such that the contralateral hand graspedthe space in front of the reward many times before actually making contact with the reward, and this led to a significantly larger numberof reaches on that side. There was no significant difference between the number of reaches on the ipsilateral or the contralateral side foreach of the other two groups. (C) The mean (� SEM) number of fast clears per trial where a fast clear was defined as a reward removedin 1 s or less. There was a significant tests by groups interaction on this measure [F(4,8) � 7.17, P � 0.01] which was related to thefact that a practice effect of increased numbers of fast clears each month was recorded on both sides, for all groups, except thecontralateral side of the putamen group.
deficits were relatively stable up to 9 and 10 months post-lesion. The putamen-lesioned monkeys did not exhibit anynoticeable spontaneous motor deficits apart from occasionalfitting that sometimes was triggered by handling or excitementabout testing.
The staircase taskOn the staircase task, the putamen-lesioned monkeysexhibited significant difficulties in using their contralateralhand and arm to pick up and remove the small rewards fromthe contralateral side. As shown in Fig. 2A, the latency toclear the contralateral side was significantly increased incomparison with the ipsilateral side and also in comparisonwith the other two groups [tests � sides � groups interaction
F(20,87) � 2.15, P � 0.01]. There was no significantdifference between the groups for the latency to clear theipsilateral side, which contrasts with the paw-reaching taskused in rats where striatal lesions are associated withsignificant reaching impairments on both sides (Fricker et al.,1997b). Detailed analysis of the videotapes revealed a numberof interesting features about each of the lesion groups’performances. The putamen lesion was associated with asignificant clumsiness such that the contralateral hand grabbedrepeatedly to try and pick up the reward and frequentlydropped the reward before finally removing it from thestaircase (referred to as ‘reaches’). The reach numbers areshown in Fig. 2B, and the putamen-lesioned group made, onaverage, more than twice the number of reaches of thecaudate and sham groups when clearing the contralateral side

Basal ganglia lesions in the marmoset 1447
Fig. 3 Data from the central staircase task. Due to the design of this apparatus, the open symbolsrepresent data from the ipsilateral hand (which removed rewards on the contralateral side) and theclosed symbols represent data from the contralateral hand (which removed rewards on the ipsilateralside). The mean (� SEM) latency required to remove rewards by each hand is shown in (A) and therewas a significant trend for the contralateral hand of the putamen-lesioned monkeys to be slower thanthe ipsilateral hand and slower than each hand of the other groups. As for the staircase task, the motorimpairment dominated performance of the putamen-lesioned monkeys such that the mean number offast clears measured on the contralateral hand performance was significantly lower than the ipsilateralhand and than the other groups (B). The lack of sensory neglect is discussed in detail in the Resultssection.
[sides � groups interaction F(2,9) � 10.60, P � 0.01]. Thepractice effect exhibited by each group on this task was mostnoticeable on a measurement, referred to as fast clears, wherethe reward was removed in 1 s or less after contact. Figure2C shows the fast clears for all three groups, and aseach monkey’s performance improved the number increased,except for the contralateral side measurements of the putamen-lesioned group, which remained significantly low at �2 pertrial [tests � groups interaction F(4,8) � 7.17, P � 0.01]. Itis interesting to note that even with this more detailed analysisof the performance of each monkey, no impairments weredetected in any of the measurements for the caudate nucleus-lesioned monkeys.
The central staircase taskA similar pattern of impairments was detected on the centralstaircase but, because the putamen-lesioned monkeys used
their ipsilateral hands to clear many of the lower pieces onthe ‘wrong’ side, the latencies to clear the ipsilateral side(where normally the contralateral hand would be used)were much faster than expected [Fig. 3A; sides � groupsinteraction, F(2,9) � 4.63, P � 0.05]. The clumsiness wasstill very much in evidence on this task, and the number ofreaches taken by the contralateral hand was significantlygreater than those taken by the ipsilateral hand and eachhand of the other two groups [data not shown;tests � sides � groups, F(8,35) � 2.80, P � 0.05]. Similarlythe lower number of fast clears supported the dominance ofthe motor impairment exhibited by the contralateral handin the putamen-lesioned group [Fig. 3B; tests �sides � groups interaction, F(8,35) � 2.40, P � 0.05]. Thevalue of this task is that the monkey must use its ipsilateralhand in the contralateral body space such that any sensoryneglect would affect performance on the contralateral side.If the deficit is predominantly motor, it would be expected

1448 A. L. Kendall et al.
Fig. 4 The maximum distance (mean � SEM) reached into thetube by the contralateral hands of each group. There was asignificant effect of the putamen lesion such that the monkeyscould only reach as far as 3 cm (P � 0.001), and this impairmentremained stable for the entire post-lesion assessment period.
that the time taken to clear rewards by the contralateral handwould be greater than that taken by the ipsilateral hand,regardless of the fact that the monkey is working in ipsilateralbody space, and Fig. 3A shows that this was indeed the case.One way in which neglect might be detected is in the latencyto contact each side for the first time, such that a monkeyexhibiting neglect would contact the ipsilateral side first eventhough it was using its contralateral hand; however, analysisof this first contact also did not reveal any evidence of neglectin any of the groups [data not shown; tests � sides � groups,F(8,35) � 1.29, NS]. Both of the staircases clearlydemonstrated that the putamen lesion is associated with asignificant motor impairment with no evidence to suggestany overt sensory neglect.
Tube reaching taskThe tube reaching task further confirmed the existence of asignificant motor impairment after a unilateral putamen lesionbut not after a caudate lesion. Figure 4 shows the maximumdistance reached by the contralateral hand. The distancereached by the putamen group was 3 cm or less, which wasin contrast to the maximum 6 cm reached by the othertwo groups [tests � groups interaction, F(10,44) � 8.65,P � 0.001].
Sticky label taskIn the sticky label task (Fig. 5), there were no overalldifferences between the three groups or between the timetaken to contact or remove the ipsilateral or the contra-lateral label in the putamen group [contact data:tests � sides � groups, F(10,44) � 1.00, NS]. This test hasbeen used previously to demonstrate sensory neglect in both
Fig. 5 The mean (� SEM) latency to contact each label on eachfoot in the sticky label task. There was no overall significantdifference between the three groups nor was there a significanteffect of the putamen lesion as there was no difference betweenthe times taken to contact, or remove, the labels from theipsilateral and the contralateral feet.
rats and marmosets (Schallert et al., 1982; Annett et al.,1992), and the lack of difference between the two sides inthe putamen group supports the data in the staircase taskssuggesting that this lesion produces little overt sensoryneglect.
RotationThe rotation measurements were extremely variable for allthree groups, and only the apomorphine-induced rotationyielded a consistent post-lesion effect. Figure 6A shows therotation measured for 60 min after a saline injection and,despite some contralateral rotation evident in the caudate-lesioned group, there was no overall significant differencebetween the groups, nor was there a significant effect of thelesion [tests � groups interaction, F(10,44) � 0.7, NS].Figure 6B shows that the contralateral bias exhibited bythe caudate-lesioned monkeys was also evident after theadministration of amphetamine and, as the direction wasmaintained post-lesion with some increase in the overallnumber of turns, this effect was significant [tests � groups,F(10,38) � 2.36, P � 0.05]. However, closer inspection ofindividual monkey performances showed that thiscontralateral bias in both saline and amphetamine rotationwas due to the extremely large number of turns exhibited byonly one monkey in this group. The putamen lesion was not

Basal ganglia lesions in the marmoset 1449
Fig. 6 The mean (� SEM) net rotation scores for (A) the salineand (B) the amphetamine (0.5 mg/kg) rotation tests. There was nosignificant effect of the sham treatment or the putamen lesion oneach test, but there was a significant contralateral bias shown bythe caudate-lesioned group on the amphetamine test (P � 0.05).
associated with any significant degree of rotation after eithersaline or amphetamine administration.
Apomorphine administration produced a profoundipsilateral bias in the putamen lesion (Fig. 7) as well as asmaller bias in both the caudate and the sham group. Therewas a significant difference between the three groups, as thenumber of turns exhibited by the putamen-lesioned monkeysexceeded 200 in comparison with �50 turns seen in theother two groups [groups, F(2,9) � 10.84, P � 0.01]. Thesmall number of turns exhibited by the sham and caudategroups is probably related to the non-specific effects ofapomorphine on the overall activity of the monkeys (Ridleyet al., 1980), but it is interesting that this cancelled thecontralateral bias seen previously in the caudate group. Therewas no overall tests by groups interaction [F(4,8) � 0.42,NS], suggesting that the apomorphine tests were performedfar enough apart to avoid the conditioning and sensitizationeffects seen in other studies (Annett et al., 1992).
Fig. 7 The mean (� SEM) net rotation score followingapomorphine (0.5 mg/kg) administration for each group. Therewas a significant difference between the sham and putamen, andbetween the caudate and putamen groups (P � 0.01) because ofthe large numbers of ipsilateral turns performed by the monkeyswith putamen lesions.
In previous striatal lesion studies with large primates, thesame dose of apomorphine has been used to elicit dyskinesias(Burns et al., 1995). Although there were no overt dyskinesiasseen in the present study, the putamen-lesioned monkeys didexhibit marked contralateral arm and hand dystonia as wellas some oro-facial dyskinesias during the acute drug phase.Neither the sham nor the caudate monkeys exhibited anydetectable dystonia, but there were occasional repetitivebehaviour patterns, such as licking the front panel of thecage, which were probably related to the high dose used inthis study. At the end of the study, lower doses of apomorphinewere used (0.05 and 0.1 mg/kg, 10 months post-lesion); inthe caudate monkeys, the contralateral bias evident in thesaline and amphetamine measurements returned and therewere no stereotypies. The lower doses did still produceipsilateral rotation in the putamen-lesioned monkeys but toa much lower degree (0.05 mg/kg mean net score �15.67 � 6.89; 0.1 mg/kg mean net score � 44.00 � 29.36).
In summary the behavioural analysis of the effects ofunilateral caudate nucleus lesions demonstrated that therewere no significant motor impairments nor was there anyevidence of sensory neglect, but there was a slight tendencyto rotate in a contralateral direction. The functionalconsequences of a unilateral putamen lesion were extremelymarked and persisted for the entire post-lesion assessmentperiod. The monkeys exhibited a significant impairment inthe complex sequencing of motor actions with thecontralateral arm and hand as well as a marked degree ofipsilateral rotation in response to apomorphine, but there wasno evidence of sensory neglect.

1450 A. L. Kendall et al.
Fig. 8 Each row shows representative sections, at the anterior commissural level, taken from one monkey in each group. The top row(A–D) is taken from a sham-lesioned brain (caudate procedure SC-25), the middle row (E–H) from a caudate-lesioned brain (C15) andthe bottom row (I–L) from a putamen-lesioned brain (P21). Nissl-stained sections are shown in A, E and I; acetylcholinesterase-stainedsections in B, F and J; DARPP-32-stained sections in C, G and K; and GFAP-stained sections in D, H and L. The scale bar (in L)represents 500 µm.
HistologyIn the sham-lesioned monkeys, the needle tracks weredetectable in those sections that were stained with Nissl andGFAP, but there was no overt cell loss (see Fig. 8A and D).Levels of immunoreactivity for DARPP-32 and tyrosinehydroxylase were not altered in any of the sham-operatedbrains (Fig. 8, top row). There was, however, limited damage
to the superficial layers of the overlying cortical region thatmay have been related to subdural scar tissue at the corticalsurface associated with multiple needle penetrations.
The caudate lesions generally spared the most anteriorportion of the head and tail segments, but the body of thecaudate was significantly reduced, with a correspondingcollapse of the nearby brain regions and ventricular
Basal ganglia lesions in the marmoset 1451
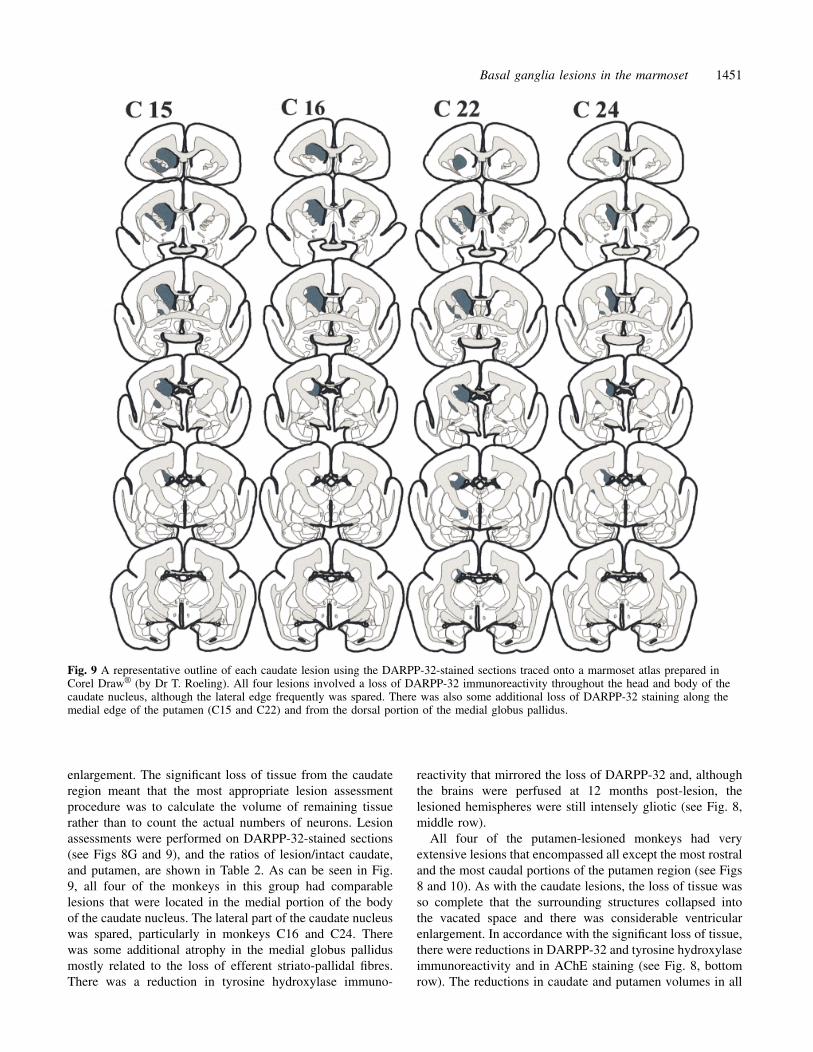
Fig. 9 A representative outline of each caudate lesion using the DARPP-32-stained sections traced onto a marmoset atlas prepared inCorel Draw® (by Dr T. Roeling). All four lesions involved a loss of DARPP-32 immunoreactivity throughout the head and body of thecaudate nucleus, although the lateral edge frequently was spared. There was also some additional loss of DARPP-32 staining along themedial edge of the putamen (C15 and C22) and from the dorsal portion of the medial globus pallidus.
enlargement. The significant loss of tissue from the caudateregion meant that the most appropriate lesion assessmentprocedure was to calculate the volume of remaining tissuerather than to count the actual numbers of neurons. Lesionassessments were performed on DARPP-32-stained sections(see Figs 8G and 9), and the ratios of lesion/intact caudate,and putamen, are shown in Table 2. As can be seen in Fig.9, all four of the monkeys in this group had comparablelesions that were located in the medial portion of the bodyof the caudate nucleus. The lateral part of the caudate nucleuswas spared, particularly in monkeys C16 and C24. Therewas some additional atrophy in the medial globus pallidusmostly related to the loss of efferent striato-pallidal fibres.There was a reduction in tyrosine hydroxylase immuno-
reactivity that mirrored the loss of DARPP-32 and, althoughthe brains were perfused at 12 months post-lesion, thelesioned hemispheres were still intensely gliotic (see Fig. 8,middle row).
All four of the putamen-lesioned monkeys had veryextensive lesions that encompassed all except the most rostraland the most caudal portions of the putamen region (see Figs8 and 10). As with the caudate lesions, the loss of tissue wasso complete that the surrounding structures collapsed intothe vacated space and there was considerable ventricularenlargement. In accordance with the significant loss of tissue,there were reductions in DARPP-32 and tyrosine hydroxylaseimmunoreactivity and in AChE staining (see Fig. 8, bottomrow). The reductions in caudate and putamen volumes in all

1452 A. L. Kendall et al.
Table 2 DARPP-32 volume ratios
Monkey Caudate Mean � SEM Putamen Mean�SEM
SP 17 0.93 0.95 � 0.02 1.11 0.99 � 0.10SC 18 0.92 –SC 23 1.00 0.80SC 25 0.95 1.07
C 16 0.23 0.27 � 0.07 0.94 0.81 � 0.07C 15 0.11 0.68C 22 0.29 0.71C 24 0.43 0.92
P 12 0.99 0.96 � 0.03 0.11 0.20 � 0.04P 14 0.86 0.21P 20 0.98 0.31P 21 1.01 0.16
The volumes of DARPP-32-immunoreactive caudate and putamenon the intact and operated sides were calculated for each monkey.A ratio of lesion/intact was then calculated for each monkey,where S represents shams, C represents caudate-lesioned and Prepresents putamen-lesioned monkeys.
three groups were compared and there was a significantvolume � groups interaction [F(2,8) � 80.76, P � 0.001]related to the loss of DARPP-32 immunoreactivity in therespective areas for the caudate-lesioned and the putamen-lesioned groups. Post hoc analyses confirmed that were wasno significant reduction in DARPP-32 staining in the caudatenucleus of the putamen-lesioned monkeys and there was nosignificant loss of DARPP-32 staining in the putamen ofcaudate-lesioned monkeys. As shown in Fig. 10, the putamenlesions were highly reproducible. Additional atrophy wasseen in the dorsal regions of both segments of the globuspallidus in all four monkeys, which may, in part, be relatedto loss of efferent fibres. The GFAP-stained sections indicatedan intense gliotic reaction around the lesioned area andparticularly in the overlying cortex (see Fig. 8L).
Correlation of the post-mortem anatomy withbehavioural data.The DARPP-32 ratios of lesion/intact caudate and putamenfor all 12 monkeys were correlated with behavioural scoresthat had been calculated for each test. The behavioural scoreswere calculated using means of all of the post-lesion datafor each test, and then either the ratio of contralateral/ipsilateral (as for staircase latencies) or the difference betweencontralateral and ipsilateral (as for net rotation scores) wascalculated. The correlations were performed using Pearson’sr, and a summary of the results is shown in Table 3.
The ratio of lesioned over intact caudate volume did notcorrelate with any of the behavioural measurements, whichis consistent with the lack of functional deficit seen incaudate-lesioned monkeys on all of the tests used in thisstudy. However, there were significant correlations betweenthe putamen ratios and all of the behavioural measurementsexcept for saline and amphetamine rotation and the sticky
label task, where no deficits had been seen in the lesionedgroup. Two of the most significant correlations, betweenputamen ratios and latency to clear the staircase andapomorphine rotation, are illustrated in Fig. 11.
In summary, in the common marmoset, excitotoxic lesionsof either the caudate nucleus or the putamen are highlyreproducible, with a significant loss of neurons and DARPP-32 and tyrosine hydroxylase immunoreactivity in both cases.The putamen lesions are largely complete and also impinge onthe neighbouring globus pallidus. The significant correlationsbetween putamen ratios and the behavioural scores may berelated to this additional damage.
DiscussionFunctional consequences of basal ganglialesionsWe have shown that it is possible to create relatively focallesions of either the caudate nucleus or the putamen inthe common marmoset using multiple small injections ofquinolinic acid. In addition, we demonstrate that putamenlesions are associated with a range of contralateral motorimpairments that appear to remain stable over repeatedassessments and over a long period of time. However, thecaudate nucleus lesion was not associated with any detectablemotor impairment.
In the marmoset, the loss of between 69 and 89% of theputamen (assessed by DARPP-32 staining, see Table 2) wasassociated with significant skilled motor deficits exhibitedduring the reaching and retrieval of rewards by thecontralateral arm. Such deficits are consistent with rodentstudies where impairments in skilled forelimb use have beenshown following excitotoxic striatal lesions (Whishaw et al.,1986; Fricker et al., 1996, 1997b), particularly when thoselesions target the sensorimotor area in the dorsolateral striatum(Fricker et al., 1996). The extent of the reaching impairmentsseen in the putamen-lesioned monkeys was similar to thatfound in unilateral 6-hydroxydopamine-lesioned monkeyswhere reaching into tubes was also significantly impaired(Annett et al., 1994). Annett and colleagues have also foundthat reaching for rewards on the staircase task is impairedafter dopamine depletion but, rather than speed and accuracy,it is the order of reward removal which is most noticeablyaffected (Annett et al., 1992b). The dopamine-depletedmonkeys exhibit a marked contralateral neglect such that onthe staircase task ipsilateral rewards are removed first andon the sticky label task the ipsilateral label is always contactedbefore the contralateral label (Annett et al., 1992a). In thisstudy, the destruction of intrinsic striatal neurons does notappear to be associated with contralateral neglect since thereis no obvious bias on the sticky label task nor is there atendency to prefer the ipsilateral body space in the centralstaircase task. In tasks employed to differentiate the sensoryand motor elements of behavioural responses, it has beenshown that rats with unilateral striatal lesions, or unilateral

Basal ganglia lesions in the marmoset 1453
Fig. 10 The putamen lesions, assessed by loss of DARPP-32 immunoreactivity. The Corel Draw® atlas was used to provide templatesover which lesions have been drawn. The putamen lesions were particularly extensive and encompassed virtually all of the pre- and post-commissural putamen region in all four monkeys. In addition, there was also a loss of DARPP-32 staining in both segments of theglobus pallidus.
dopamine depletion, are not impaired in detectingcontralateral stimuli but rather in making motor responses tothe contralateral side (Carli et al., 1989; Brown and Robbins,1989a, b; Brasted et al., 1999). It has also been suggestedthat the contralateral neglect seen in unilateral 6-hydroxy-dopamine-lesioned marmosets is related more to problemswith initiating motor responses, confounded by a profoundspontaneous rotation, than to sensory detection on thecontralateral side (Annett et al., 1992). The tests employedin this study cannot prove the lack of a sensory element tothe impairments seen in the putamen-lesioned monkeys, butthe lack of neglect phenomena and the extent of the motorproblems suggest that most, if not all, of the deficits aremotor in origin.
Drug-induced rotation following administration of director indirect dopamine agonists is a measure that is commonlyused in the analysis of basal ganglia lesions (Ungerstedt andArbuthnott, 1970; Schwarcz et al., 1979; Pycock, 1980;Dunnett et al., 1988). We found that unilateral putamenlesions were associated with an ipsilateral rotation followingapomorphine administration, but there was no consistentresponse following the administration of amphetamine. Thiscontrasts with the consistent ipsilateral rotation toamphetamine and contralateral rotation to apomorphine seenin marmosets after nigrostriatal bundle lesions (Annett et al.,1992). In fact, a similar discrepancy is seen in rats, in whichnigrostriatal lesions also yield consistent ipsilateral rotationunder amphetamine and contralateral rotation under

1454 A. L. Kendall et al.
Table 3 Correlations between DARPP-32 ratios andbehaviour
Task Caudate P Putamen P
Staircase latency 0.269 NS –0.691 �0.01Staircase fast clears 0.462 NS –0.918 �0.01Central stair latency 0.479 NS –0.655 �0.02Central stair fast clears 0.395 NS –0.707 �0.01Tube reaching 0.409 NS –0.781 �0.01Amphetamine rotation 0.190 NS –0.058 NSApomorphine rotation 0.323 NS –0.762 �0.01
Behavioural scores for each test were calculated (see text fordetails) and then correlated (using Pearson’s r) with the ratio oflesion/intact DARPP-32 immunoreactivity for each region. Whenperforming the correlations, the caudate ratios or the putamenratios for all of the monkeys in all three groups were used.
Fig. 11 The correlation between the latency to clear the staircase(top) and the net apomorphine rotation score (bottom). DARPP-32-stained sections were analysed using a Seescan image analysissystem, and the ratio of lesioned/intact caudate and putamenvolume was calculated for each monkey. These ratios were thencorrelated with scores from each of the behavioural tests(Pearson’s r correlation—see Results section). In both cases, theputamen ratios correlate significantly (P � 0.01) with thebehaviour but the caudate ratios do not. For simplicity, the graphsillustrate the putamen ratios from the sham and putamen lesiongroups and the caudate ratios from the sham and the caudatelesion groups, but all correlation statistics were performed usingall of the data.
apomorphine (Ungerstedt, 1971a, b), whereas excitotoxiclesions of the striatum sometimes induce ipsilateral rotationin response to both drugs (Schwarcz et al., 1979; Dunnettet al., 1988) and sometimes produce opposite turning orcontralateral responses in particular to apomorphine (Normanet al., 1992). Attempts to resolve this conflict in rats havesuggested that, whereas the rate and direction of turning inanimals with nigrostriatal lesions are related to the extentof denervation and development of postsynaptic dopaminereceptor supersensitivity (Ungerstedt, 1971a; Hefti et al.,1980), the direction of turning in rats with neostriatal lesionsis more related to the topography of the lesion. In particular,the most consistent contralateral rotation is obtained withlateral striatal lesions, whereas ipsilateral turning is associatedmore often with anterior and medial lesions (Norman et al.,1992; Fricker et al., 1996). Surprisingly, animals withrelatively large lesions may show little overall rotation, whichis most plausibly attributed to invasion of areas influencingboth ipsilateral and contralateral turning bias. Thus, in thepresent study, and in contrast to marmosets with nigrostriatallesions, it is the putamen site, equivalent to lateral neostriatumin rats, that produces rotation after excitotoxic lesion inmarmosets and this is more consistent for apomorphine thanamphetamine. Nevertheless, although the phenomenon ofrotation may provide a simple quantitative measure ofdysfunction, the cellular mechanisms underlying such rotationremain ambiguous.
Comparison with previous primate striatallesion studiesIt is difficult to make comparisons with previous striatallesion studies in primates since the majority have lesionedboth the caudate and the putamen in an effort to match theneuropathological damage of Huntington’s disease (Hantrayeet al., 1990; DeLong, 1990; Ferrante et al., 1993; Brouilletand Hantraye, 1995). The main aim of primate investigationsin the past has been to elicit chorea and dyskinesias similarto those seen in Huntington’s disease patients. Theadministration of dopamine agonists such as L-dopa orapomorphine to monkeys that have received kainic acid,ibotenic acid or quinolinic acid lesions of the caudate–putamen leads to the onset of a range of dyskinetic anddystonic posturing (Hantraye et al., 1990; Kanazawa et al.,1990; Brownell et al., 1994; Storey et al., 1994; Burnset al., 1995). Spontaneous dyskinesias are only seen in theimmediate post-lesion phase following excitotoxic lesionsand only if the lesion includes the putamen region (Brownellet al., 1994; Burns et al., 1995). If the lesion encompassed�60% of the caudate–putamen or if a ventral striatal lesionwas added to the existing caudate–putamen lesion, then drug-induced dyskinesias were not seen (Hantraye et al., 1990;Kanazawa et al., 1990). This requirement for some remainingportion of the striatum implies that it is a combination of theconsequence of removing part of the caudate–putamen and

Basal ganglia lesions in the marmoset 1455
the action of the remaining portion that is involved in theinitiation of dyskinesias. Although we did not witness a truedyskinetic profile in our putamen-lesioned monkeys, we didsee elements of dystonia and some oro-facial movementsduring the acute drug phase following the administration ofapomorphine. It is possible that the dose of apomorphineused in this study was too low since the same dose in ibotenicacid-lesioned baboons did not produce hyperkinesia ordyskinesia and most primate studies have used a dose of 1or 2 mg/kg (Hantraye et al., 1990; Brownell et al., 1994;Storey et al., 1994). We chose the lower dose becausemarmosets are extremely sensitive to the action ofapomorphine and, in pilot studies, we found that higher dosesproduced movements more akin to stereotypies (Ridley et al.,1980). It is of interest that the administration of apomorphinein kainic acid-lesioned macaque monkeys did not elicitdyskinesias, whereas L-dopa and met-amphetamineadministration did produce responses (Kanazawa et al., 1990).Alternatively, it may be that the extent of the putamen lesionwas simply too great for the true appearance of dyskinesias,which is consistent with a previous study where, in rhesusmonkeys, the same dose of apomorphine produceddyskinesias following a posterior putamen lesion but thesubsequent addition of an anterior putamen lesion abolishedthe response to apomorphine (Burns et al., 1995). Since itwas not the main aim of the study to create another modelof drug-induced dyskinesias, the use of a low dose ofapomorphine that elicited a consistent ipsilateral rotationseems justified.
Functional significance of the differencebetween the caudate and the putamen lesionedmonkeysAnatomical tracing studies have demonstrated that motor,premotor and somatosensory cortical areas send corticostriatalprojections primarily to the putamen region in primates,whereas the head and body of the caudate nucleus mostlyreceive efferent input from associational cortical areas suchas prefrontal and cingulate cortex (Kemp and Powell, 1970;Kunzle, 1975, 1977, 1978; Selemon and Goldman-Rakic,1985; Takada et al., 1998). This separation of corticostriatalprojections is by no means complete but forms the basis oftheories of basal ganglia function that rely upon the parallelprocessing of information along functionally segregatedcortico-basal ganglia–thalamic loops (Alexander andCrutcher, 1990a; Alexander et al., 1990). Physiologicalstudies further confirm the complex regional topographynoted in anatomical studies such that there is a clear distinctionbetween sensorimotor responses found in the putamen andthe more complex behavioural responses seen in caudatenucleus neurons. Microstimulation of neurons in the putamencan elicit movement of specific body parts depending on thearea that is stimulated, but that response is abolished after afocal injection of ibotenic acid (Alexander and DeLong,
1985). In addition, passive movement of limbs is nearlyalways associated with neuronal responses in the putamenand those neurons may display a sensitivity for the directionand/or preparation of movement (Alexander and DeLong,1985; Crutcher and Alexander, 1990; Alexander and Crutcher,1990b). The motor circuit, described by Alexander andcolleagues, combines input from primary motor cortex withthat from supplementary and premotor cortex along withsomatosensory cortex to the putamen, which in turn sendstopographic projections to both segments of the globuspallidus and the substantia nigra reticulata. Projections fromthe globus pallidus and substantia nigra to the thalamusmaintain the topography, and the projections from thethalamus to premotor and motor cortex complete the loop. Itseems reasonable to assume that removal of a criticalcomponent of the motor circuit, namely the putamen, willdisrupt motor performance. The disruption seen in this studyappears to be related more to the execution of motor actsrather than to initiation, although more specific tests may beable to tease out the exact nature of the motor problems.
The caudate nucleus is not considered to be part of themotor loop, rather, contemporary evidence suggests that thecaudate nucleus is implicated more in cognitive processessuch as attention and response selection in specificenvironmental contexts that are assigned to the two prefrontalcortico-striatal loops (Alexander et al., 1990). Followingexcitotoxic lesions of the caudate nucleus, we did not observeany significant motor impairment and, since the lesions wereunilateral, no cognitive tasks were performed.
Application of unilateral putamen lesions in thecommon marmosetThe primary aim of our study was to develop a lesion modelin the common marmoset that can be used for evaluatingtransplantation strategies for striatal repair. The criterion forsuch a model is that it is associated with a range of quantifiablemotor impairments and that it is stable over repeated testingand over a long period of time. A unilateral quinolinic acidlesion of the putamen region in the marmoset meets theserequirements and can therefore be applied in the studyof potential restorative and neuroprotective strategies fordiseases, such as Huntington’s, that attack the striatum. Inparticular, following extensive studies in rodents, we haverequired a primate model to evaluate the effects oftransplantation in the differentiated basal ganglia (withseparate caudate and putamen nuclei) which also applies inman. The marmoset is ideally suited for such investigationsbecause of its small size, ease of handling, relatively lowcost and the fact that it breeds readily in captivity, whichis essential for the supply of foetal material in neuraltransplantation studies.
In this study, we did not seek to replicate the pathologyor the symptoms of Huntington’s disease but rather to developa consistently reproducible lesion with motor impairments

1456 A. L. Kendall et al.
which have been used to validate our battery of behaviouraltests. As an application of this unilateral putamen lesionmodel in the marmoset, we recently have demonstrated partialamelioration of the motor deficits following implantation offoetal striatal tissue allografts (Kendall et al., 1998).
AcknowledgementsThe authors would like to thank Drs H. Hemmings and P.Greengard for the generous donation of the DARPP-32antibody. In addition, thanks are due to Ms H. Cox fortechnical assistance in the preparation of the histologicalmaterial. This work was supported by an MRC programmegrant.
ReferencesAlexander GE. Basal ganglia–thalamocortical circuits: their role incontrol of movements. J Clin Neurophysiol 1994; 11: 420–31.
Alexander GE, Crutcher MD. Functional architecture of basalganglia circuits: neural substrates of parallel processing. [Review].Trends Neurosci 1990a; 13: 266–71.
Alexander GE, Crutcher MD. Preparation for movement: neuralrepresentations of intended direction in three motor areas of themonkey. J Neurophysiol 1990b; 64: 133–50.
Alexander GE, DeLong MR. Microstimulation of the primateneostriatum. I. Physiological properties of striatal microexcitablezones. J Neurophysiol 1985; 53: 1401–6.
Alexander GE, DeLong MR, Strick PL. Parallel organization offunctionally segregated circuits linking basal ganglia and cortex.[Review]. Annu Rev Neurosci 1986; 9: 357–81.
Alexander GE, Crutcher MD, DeLong MR. Basal ganglia–thalamocortical circuits: parallel substrates for motor, oculomotor,‘prefrontal’ and ‘limbic’ functions. [Review]. Prog Brain Res 1990;85: 119–46.
Annett LE, Rogers DC, Hernandez TD, Dunnett SB. Behaviouralanalysis of unilateral monoamine depletion in the marmoset. Brain1992a; 115: 825–56.
Annett LE, Torres EM, Clark DJ, Dunnett SB. A spatial componentin the neglect produced by unilateral 6-OHDA lesions in marmosets[abstract]. J Psychopharmacol: BAPS and EBPS meeting, 1992bAug 2–7; 189.
Annett LE, Martel FL, Rogers DC, Ridley RM, Baker HF, DunnettSB. Behavioral assessment of the effects of embryonic nigral graftsin marmosets with unilateral 6-OHDA lesions of the nigrostriatalpathway. Exp Neurol 1994; 125: 228–46.
Aronin N, Kim M, Laforet G, DiFiglia M. Are there multiplepathways in the pathogenesis of Huntington’s disease? [Review].Philos Trans R Soc Lond B Biol Sci 1999; 354: 995–1003.
Beal MF, Kowall NW, Ellison DW, Mazurek MF, Swartz KJ, MartinJB. Replication of the neurochemical characteristics of Huntington’sdisease by quinolinic acid. Nature 1986; 321: 168–71.
Beal MF, Ellison DW, Mazurek MF, Swartz KJ, Malloy JR, BirdED, et al. A detailed examination of substance P in pathologicallygraded cases of Huntington’s disease. J Neurol Sci 1988; 84: 51–61.
Beal MF, Kowall NW, Swartz KJ, Ferrante RJ, Martin JB.Differential sparing of somatostatin–neuropeptide Y and cholinergicneurons following striatal excitotoxin lesions. Synapse 1989; 3:38–47.
Block F, Kunkel M, Schwarz M. Quinolinic acid lesion of thestriatum induces impairment in spatial learning and motorperformance in rats. Neurosci Lett 1993; 149: 126–8.
Brasted PJ, Dobrossy MD, Robbins TW, Dunnett SB. Striatal lesionsproduce distinctive impairments in reaction time performance intwo different operant chambers. Brain Res Bull 1998; 46: 487–93.
Brasted PJ, Robbins TW, Dunnett SB. Distinct roles for striatalsubregions in mediating response processing revealed by focalexcitotoxic lesions. Behav Neurosci 1999; 113: 253–64.
Brouillet E, Hantraye P. Effects of chronic MPTP and 3-nitropropionic acid in nonhuman primates. [Review]. Curr OpinNeurol 1995; 8: 469–73.
Brown VJ, Robbins TW. Deficits in response space followingunilateral striatal dopamine depletion in the rat. J Neurosci 1989a;9: 983–9.
Brown VJ, Robbins TW. Elementary processes of response selectionmediated by distinct regions of the striatum. J Neurosci 1989b; 9:3760–5.
Brownell AL, Hantraye P, Wullner U, Hamberg L, Shoup T, ElmalehDR, et al. PET- and MRI-based assessment of glucose utilization,dopamine receptor binding, and hemodynamic changes after lesionsto the caudate–putamen in primates. Exp Neurol 1994; 125: 41–51.
Burns LH, Pakzaban P, Deacon TW, Brownell AL, Tatter SB,Jenkins BG, et al. Selective putaminal excitotoxic lesions in non-human primates model the movement disorder of Huntingtondisease. Neuroscience 1995; 64: 1007–17.
Carli M, Jones GH, Robbins TW. Effects of unilateral dorsal andventral striatal dopamine depletion on visual neglect in the rat: aneural and behavioural analysis. Neuroscience 1989; 29: 309–27.
Crutcher MD, Alexander GE. Movement-related neuronal activityselectively coding either direction or muscle pattern in three motorareas of the monkey. J Neurophysiol 1990; 64: 151–63.
de la Monte SM, Vonsattel JP, Richardson EP Jr. Morphometricdemonstration of atrophic changes in the cerebral cortex, whitematter, and neostriatum in Huntington’s disease. J Neuropathol ExpNeurol 1988; 47: 516–25.
DeLong MR. Primate models of movement disorders of basalganglia origin. [Review]. Trends Neurosci 1990; 13: 281–5.
Dunnett SB, Isacson O, Sirinathsinghji DJ, Clarke DJ, BjorklundA. Striatal grafts in rats with unilateral neostriatal lesions. III.Recovery from dopamine-dependent motor asymmetry and deficitsin skilled paw reaching. Neuroscience 1988; 24: 813–20.
Ferrante RJ, Kowall NW, Beal MF, Martin JB, Bird ED, RichardsonEP. Morphologic and histochemical characteristics of a spared subsetof striatal neurons in Huntington’s disease. J Neuropathol ExpNeurol 1987; 46: 12–27.

Basal ganglia lesions in the marmoset 1457
Ferrante RJ, Kowall NW, Cipolloni PB, Storey E, Beal MF.Excitotoxin lesions in primates as a model for Huntington’s disease:histopathologic and neurochemical characterization. Exp Neurol1993; 119: 46–71.
Fricker RA, Annett LE, Torres EM, Dunnett SB. The placement ofa striatal ibotenic acid lesion affects skilled forelimb use and thedirection of drug-induced rotation. Brain Res Bull 1996; 41: 409–16.
Fricker RA, Torres EM, Dunnett SB. The effects of donor stage onthe survival and function of embryonic striatal grafts in the adultrat brain. I. Morphological characteristics. Neuroscience 1997a; 79:695–710.
Fricker RA, Torres EM, Hume SP, Myers R, Opacka-Juffrey J,Ashworth S, et al. The effects of donor stage on the survival andfunction of embryonic striatal grafts in the adult rat brain. II.Correlation between positron emission tomography and reachingbehaviour. Neuroscience 1997b; 79: 711–21.
Graybiel AM. The basal ganglia and chunking of action repertoires.[Review]. Neurobiol Learn Mem 1998; 70: 119–36.
Hantraye P, Riche D, Maziere M, Isacson O. A primate model ofHuntington’s disease: behavioral and anatomical studies of unilateralexcitotoxic lesions of the caudate–putamen in the baboon. ExpNeurol 1990; 108: 91–104.
Harper PS. Huntington’s disease. 2nd edn. London: W.B.Saunders; 1996.
Hefti F, Melamed E, Sahakian BJ, Wurtman RJ. Circling behaviorin rats with partial, unilateral nigro-striatal lesions: effect ofamphetamine, apomorphine, and DOPA. Pharmacol Biochem Behav1980; 12: 185–8.
Huntington’s Disease Collaborative Research Group. A novel genecontaining a trinucleotide repeat that is expanded and unstable onHuntington’s disease chromosomes. Cell 1993; 72: 971–83.
Jueptner M, Weiller C. A review of differences between basalganglia and cerebellar control of movements as revealed byfunctional imaging studies. [Review]. Brain 1998; 121: 1437–49.
Kanazawa I, Kimura M, Murata M, Tanaka Y, Cho F. Choreicmovements in the macaque monkey induced by kainic acid lesionsof the striatum combined with L-dopa. Pharmacological, biochemicaland physiological studies on neural mechanisms. Brain 1990; 113:509–35.
Kemp JM, Powell TP. The cortico-striate projection in the monkey.Brain 1970; 93: 525–46.
Kendall AL, Rayment FD, Torres EM, Baker HF, Ridley RM,Dunnett SB. Functional integration of striatal allografts in a primatemodel of Huntington’s disease. Nature Med 1998; 4: 727–9.
Koelle GB. The histochemical identification of acetylcholinesterasein cholinergic, adrenergic and sensory organs. J Pharmacol ExpTher 1955; 114: 167–84.
Kowall NW, Ferrante RJ, Martin JB. Patterns of cell loss inHuntington’s disease. Trends Neurosci 1987; 10: 24–9.
Kunzle H. Bilateral projections from precentral motor cortex to theputamen and other parts of the basal ganglia. An autoradiographicstudy in Macaca fascicularis. Brain Res 1975; 88: 195–209.
Kunzle H. Projections from the primary somatosensory cortex tobasal ganglia and thalamus in the monkey. Exp Brain Res 1977;30: 481–92.
Kunzle H. An autoradiographic analysis of the efferent connectionsfrom premotor and adjacent prefrontal regions (areas 6 and 9) inMacaca fascicularis. Brain Behav Evol 1978; 15: 185–234.
Lang AE, Lozano AM. Parkinson’s disease. [Review]. N Engl JMed 1998; 339: 1044–53.
Lawrence AD, Sahakian BJ, Robbins TW. Cognitive functions andcorticostriatal circuits: insights from Huntington’s disease. TrendsCogn Sci 1998; 2: 379–88.
Marshall JW, Ridley RM. Assessment of functional impairmentfollowing permanent middle cerebral artery occlusion in a non-human primate species. Neurodegeneration 1996; 5: 275–86.
Marshall JW, Cross AJ, Ridley RM. Functional benefit fromclomethiazole treatment after focal cerebral ischemia in a nonhumanprimate species. Exp Neurol 1999; 156: 121–9.
Norman AB, Norgren RB, Wyatt LM, Hildebrand JP, SanbergPR. The direction of apomorphine-induced rotation behavior isdependent on the location of excitotoxin lesions in the rat basalganglia. Brain Res 1992; 569: 169–72.
Pycock CJ. Turning behaviour in animals. [Review]. Neuroscience1980; 5: 461–514.
Reddy PH, Williams M, Tagle DA. Recent advances in understandingthe pathogenesis of Huntington’s disease. [Review]. Trends Neurosci1999; 22: 248–55.
Ridley RM, Baker HF, Crow TJ. Behavioural effects ofamphetamines and related stimulants: the importance of speciesdifferences as demonstrated by a study in the marmoset. In: CaldwellJ, editor. Amphetamines and related stimulants: chemical, biological,clinical and sociological aspects. Boca Raton (FL): CRC Press;1980. p. 97–116.
Sanberg PR, Calderon SF, Giordano M, Tew JM, Norman AB.The quinolinic acid model of Huntington’s disease: locomotorabnormalities. Exp Neurol 1989; 105: 45–53.
Schallert T, Upchurch M, Lobaugh N, Farrar SB, Spirduso WW,Gilliam P, et al. Tactile extinction: distinguishing betweensensorimotor and motor asymmetries in rats with unilateralnigrostriatal damage. Pharmacol Biochem Behav 1982; 16: 455–62.
Schwarcz R, Fuxe K, Agnati LF, Hokfelt T, Coyle JT. Rotationalbehaviour in rats with unilateral striatal kainic acid lesions: abehavioural model for studies on intact dopamine receptors. BrainRes 1979; 170: 485–95.
Selemon LD, Goldman-Rakic PS. Longitudinal topography andinterdigitation of corticostriatal projections in the rhesus monkey. JNeurosci 1985; 5: 776–94.
Sharp AH, Ross CA. Neurobiology of Huntington’s disease.Neurobiol Dis 1996; 3: 3–15.
Stephan H, Baron G, Schwerdtfeger WK. The brain of the commonmarmoset (Callithrix jacchus): a stereotaxic atlas. [Review]. Berlin:Springer-Verlag; 1980.

1458 A. L. Kendall et al.
Storey E, Cipolloni PB, Ferrante RJ, Kowall NW, Beal FM.Movement disorder following excitotoxin lesions in primates.Neuroreport 1994; 5: 1259–61.
Takada M, Tokuno H, Nambu A, Inase M. Corticostriatal projectionsfrom the somatic motor areas of the frontal cortex in the macaquemonkey: segregation versus overlap of input zones from the primarymotor cortex, the supplementary motor area, and the premotorcortex. Exp Brain Res 1998; 120: 114–28.
Ungerstedt U. Postsynaptic supersensitivity after 6-hydroxy-dopamine induced degeneration of the nigro-striatal dopaminesystem. Acta Physiol Scand Suppl 1971a; 367: 69–93.
Ungerstedt U. Striatal dopamine release after amphetamine or nervedegeneration revealed by rotational behaviour. Acta Physiol ScandSuppl 1971b; 367: 49–68.
Ungerstedt U, Arbuthnott GW. Quantitative recording of rotationalbehavior in rats after 6-hydroxy-dopamine lesions of the nigrostriataldopamine system. Brain Res 1970; 24: 485–93.
Vonsattel JP, DiFiglia M. Huntington disease. [Review]. JNeuropathol Exp Neurol 1998; 57: 369–84.
Vonsattel JP, Myers RH, Stevens TJ, Ferrante RJ, Bird ED,Richardson EP Jr. Neuropathological classification of Huntington’sdisease. J Neuropathol Exp Neurol 1985; 44: 559–77.
Whishaw IQ, O’Connor WT, Dunnett SB. The contributions ofmotor cortex, nigrostriatal dopamine and caudate–putamen to skilledforelimb use in the rat. Brain 1986; 109: 805–43.
Wichmann T, DeLong MR. Functional and pathophysiologicalmodels of the basal ganglia. [Review]. Curr Opin Neurobiol 1996;6: 751–8.
Young AB, Greenamyre JT, Hollingsworth Z, Albin R, D’AmatoC, Shoulson I, et al. NMDA receptor losses in putamen frompatients with Huntington’s disease. Science 1988; 241: 981–3.
Received November 26, 1999. Revised February 9, 2000.Accepted February 10, 2000