
The Hairless Phenotype of the Hirosaki Hairless Rat Is Due to the ...
24
1 The Hairless Phenotype of the Hirosaki Hairless Rat Is Due to the Deletion of an 80-kb Genomic DNA Containing Five Basic Keratin Genes* Naoki Nanashima 1, 2 , Miki Akita 1, 3 , Toshiyuki Yamada 1 , Takeshi Shimizu 1 , Hajime Nakano 3 , Yang Fan 1 , and Shigeki Tsuchida 1, 4 From the 1 Department of Biochemistry and Genome Biology, Hirosaki University Graduate School of Medicine, Hirosaki 036-8562, Japan, the 2 Department of Biomedical Sciences, Hirosaki University Graduate School of Health Sciences, Hirosaki 036-8564, Japan, and the 3 Department of Dermatology, Hirosaki University Graduate School of Medicine, Hirosaki 036-8562, Japan Running title: Hairless Rat Due to Deletion of Basic Hair Keratin Genes 4 Address correspondence to: Shigeki Tsuchida, Department of Biochemistry and Genome Biology, Hirosaki University Graduate School of Medicine, 5 Zaifu-cho, Hirosaki 036-8562, Japan, Tel. +81-172-39-5018; Fax. +81-172-39-5205; E-mail: [email protected] Most models of hereditary hypotrichosis are due to alterations in growth factors and transcription factors, and the examples of causative mutations in hair keratin genes are limited. Hirosaki hairless rat (HHR) is a mutant strain spontaneously derived from Sprague-Dawley rats (SDR). In this study, the locus of the responsible gene was examined by linkage analysis and mapped on the chromosome 7q36. Since many basic keratin genes are clustered on 7q36, their expression was examined. Reverse transcription-polymerase chain reaction (RT-PCR) and genomic PCR indicated that the Kb21 (Krt81), 23 (Krt83), and 26 (Krt86) genes encoding basic hair keratins were not expressed, and were deleted. Furthermore, 80-kb genomic DNA ranging from the exon 9 of the Kb25 (Krt85) to the exon 9 of the Krt2-25 was deleted. The breakpoints of these genes were within a 95-bp portion shared by the two genes, suggesting the deletion due to non-allelic homologous recombination. Proteins identified as the Kb21, Kb23, and Krt2-25 in SDR hairs by mass spectrometry were not detected in HHR. Instead, the product of a fusion gene became dominant in HHR. Since fusion occurred between the exons of the two genes with the same sequences, the product was identical with the wild-type Kb25 protein. By immunohistochemistry Kb21 was not detected in HHR hair follicles. Kb25 was expressed in the cortex in HHR while in the medulla in SDR. This study clearly illustrates the importance of hair keratin genes in hair growth. The large keratin multigene family comprises cytokeratins, which are differentially expressed in the various types of epithelia, and hair keratins, expressed in hard keratinized structures such as hairs, nails and claws. These keratins can be divided into the acidic type I and the basic-to-neutral type II members, which form the 10-nm intermediate filament network through the obligatory association of equimolar amounts of type I and type II keratins (1). Previous studies on the hair keratins of several mammals reveal the presence of 9 type I and 6 type II members (1, 2). Hair keratins are collectively designated “H” for hair, “b” for the basic members (Hb), and “a” for the acidic members (Ha) (3). In the case of rats, genes http://www.jbc.org/cgi/doi/10.1074/jbc.M802539200 The latest version is at JBC Papers in Press. Published on April 16, 2008 as Manuscript M802539200 Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on April 11, 2018 http://www.jbc.org/ Downloaded from
-
Upload
nguyentuong -
Category
Documents
-
view
219 -
download
0
Transcript of The Hairless Phenotype of the Hirosaki Hairless Rat Is Due to the ...

1
The Hairless Phenotype of the Hirosaki Hairless Rat Is Due to the Deletion of an 80-kb Genomic DNA Containing Five Basic Keratin Genes*
Naoki Nanashima1, 2, Miki Akita1, 3, Toshiyuki Yamada1, Takeshi Shimizu1, Hajime Nakano3, Yang Fan1, and Shigeki Tsuchida1, 4
From the 1Department of Biochemistry and Genome Biology, Hirosaki University Graduate School of Medicine, Hirosaki 036-8562, Japan, the 2Department of Biomedical Sciences, Hirosaki University
Graduate School of Health Sciences, Hirosaki 036-8564, Japan, and the 3Department of Dermatology, Hirosaki University Graduate School of Medicine, Hirosaki 036-8562, Japan
Running title: Hairless Rat Due to Deletion of Basic Hair Keratin Genes 4Address correspondence to: Shigeki Tsuchida, Department of Biochemistry and Genome Biology, Hirosaki University Graduate School of Medicine, 5 Zaifu-cho, Hirosaki 036-8562, Japan, Tel. +81-172-39-5018; Fax. +81-172-39-5205; E-mail: [email protected]
Most models of hereditary hypotrichosis are due to alterations in growth factors and transcription factors, and the examples of causative mutations in hair keratin genes are limited. Hirosaki hairless rat (HHR) is a mutant strain spontaneously derived from Sprague-Dawley rats (SDR). In this study, the locus of the responsible gene was examined by linkage analysis and mapped on the chromosome 7q36. Since many basic keratin genes are clustered on 7q36, their expression was examined. Reverse transcription-polymerase chain reaction (RT-PCR) and genomic PCR indicated that the Kb21 (Krt81), 23 (Krt83), and 26 (Krt86) genes encoding basic hair keratins were not expressed, and were deleted. Furthermore, 80-kb genomic DNA ranging from the exon 9 of the Kb25 (Krt85) to the exon 9 of the
Krt2-25 was deleted. The breakpoints of these genes were within a 95-bp portion shared by the two genes, suggesting the deletion due to non-allelic homologous recombination. Proteins identified as the Kb21, Kb23, and Krt2-25 in SDR hairs by mass spectrometry were not detected in HHR. Instead, the product of a fusion gene became dominant in HHR. Since fusion occurred between the exons of the two genes with the same sequences, the product was identical with the wild-type Kb25 protein. By immunohistochemistry Kb21 was not detected in HHR hair follicles. Kb25 was expressed in the cortex in HHR while in the medulla in SDR. This study clearly illustrates the importance of hair keratin genes in hair growth.
The large keratin multigene family comprises
cytokeratins, which are differentially expressed in the various types of epithelia, and hair keratins, expressed in hard keratinized structures such as hairs, nails and claws. These keratins can be divided into the acidic type I and the basic-to-neutral type II members, which form the 10-nm intermediate filament network
through the obligatory association of equimolar amounts of type I and type II keratins (1). Previous studies on the hair keratins of several mammals reveal the presence of 9 type I and 6 type II members (1, 2). Hair keratins are collectively designated “H” for hair, “b” for the basic members (Hb), and “a” for the acidic members (Ha) (3). In the case of rats, genes
http://www.jbc.org/cgi/doi/10.1074/jbc.M802539200The latest version is at JBC Papers in Press. Published on April 16, 2008 as Manuscript M802539200
Copyright 2008 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
encoding the basic members are designated Kb21-26. In rats, the type I keratin genes are clustered on the chromosome 10q31 and the type II gene cluster on 7q36 (4). The latter cluster includes Kb21-26 on an ~200-kb DNA domain.
Hairs are produced in hair follicles. The hair follicle includes not only the cortex and medulla but also the inner root sheath, companion layer, and outer root sheath (5). Recent studies have revealed that some specific cytokeratins are expressed in the inner root sheath (6) and other components (4, 7). Numerous growth factors and cytokines such as WNT (8, 9) and transforming growth factor (10, 11) are involved in hair follicle formation, and mutations of these genes cause various types of hypotrichosis. Furthermore, loss-of-function mutation of the transcription factor Foxn1 is responsible for the nude phenotype (12). Deletion or overexpression of a member (Hoxc 13) of the highly conserved Hox multigene family, result in defective hair formation (13). However, causative mutations in keratin genes for hair abnormalities are very limited. Although cytokeratin genes are involved in several human and mouse skin diseases (14), mutations of two basic hair keratin genes are noted to lead to the inherited hair disorder monilethrix (15). Deletion or mutation of genes encoding specific basic cytokeratin forms and the acidic cytokeratin K17 (Krt17), result in hair loss in mice (7, 16).
The Hirosaki hairless rat (HHR) is a mutant strain spontaneously derived from the Sprague-Dawley rat (SDR), and its inheritance is autosomal recessive (17). HHRs are nearly bare with some sparse and short hairs and are also distinguishable from normal rats by characteristic bent whiskers (17). In addition to hair loss, female HHRs show involution of the
mammary gland at early stage of lactation, and most newborn rats die within a week because of less feeding (18). Besides HHR, several hereditary atrichotic or hypotrichotic rat models have been reported. These include Charles River “hairless” (19), naked (20), Rowett nude (21), fuzzy (22), hairless (23), shorn (24) and bald rats (25). With the exception of the nude mutation (26), responsible genes remain to be clarified.
In the present study, we investigated genes responsible for HHR by linkage analysis and identified the hhr locus on the chromosome 7q36. An 80-kb genomic DNA ranging from the exon 9 of Kb25 to the exon 9 of Krt2-25 was deleted in HHR. Furthermore, breakpoints of these genes were within a 95-bp portion shared by the two genes, suggesting the deletion to be due to non-allelic homologous recombination. This is the first report on hypotrichosis due to the deletion of hair keratin genes and the expression of a keratin fusion gene.
EXPERIMENTAL PROCEDURES Rats––HHRs were maintained by
brother-sister mating of hhr/+ heterozygous females and hhr/hhr homozygous males. HHRs and SDRs were housed in plastic cages in air-conditioned rooms with a 12 h light/dark cycle in the Institute for Animal Experiments of Hirosaki University Graduate School of Medicine and had free access to water and food. For linkage analysis, pigmented BN/CrlCrlj females (+/+, C/C, hi/hi) obtained from Charles River Japan Inc. (Tokyo, Japan), were crossed with a single HHR (hhr/hhr, c/c, h/h) male. BN rats are wild for the hhr (+) and homozygous for the dominant Color (C) and dominant irish hooded (hi) alleles. The F1 hybrid females from the initial cross were then backcrossed with
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
hhr/hhr males to produce the N2 progeny. A total of 88 N2 progeny were produced in this study. This study was carried out in accordance with the Guidelines for Animal Experimentation, Hirosaki University.
Genetic Linkage Analysis––Genomic DNA was extracted from liver specimens using the DNeasy® (Qiagen Inc., Valencia, CA). Eighteen microsatellite markers that are polymorphic between HHRs and BNs were used for the primary genome-wide linkage analysis (Table 1). The Polymerase chain reaction (PCR) was carried out in 20 µl of a premix Taq® (TaKaRa, Kyoto, Japan) containing 40 ng of the genomic DNA and 0.5 µM of each of oligonucleotide primer. After initial denaturation for 3 min at 94ºC, 35 cycles of amplification, each consisting of denaturation for 30 s at 94ºC, annealing for 30 s at 55 - 60ºC and extension for 45 s at 72ºC, were carried out with a Thermal Cycler i-Cycler (BIO-RAD, Hercules, CA). The PCR products were separated by 4% NuSive® agarose gel (FMC BioProducts, Rockland, ME) in TBE buffer with 0.1 µg/ml ethidium bromide.
Reverse Transcription (RT)-PCR––Total RNAs were extracted from the skin of HHRs and SDRs using the RNeasy mini kit (Qiagen Inc.). RT-PCR was performed with an Access Quick® RT-PCR System (Promega) by using 0.5 µg RNA and primer pairs. PCR amplification consisted of 30 s at 94ºC, 30 s at 55 to 60ºC and 30 s―2 min at 72ºC for 21 to 30 cycles. The primers used were as follows (5’→3’): VDR, F-ACCCTTGGGCTCTACTCACC and R-TCCTCTCCGTTCAGATCCAG; Wnt10b, F-GGCTGTAATCACGACATGGA and R-GGAGCCATGATTAACCGAAA; Sp1, F-AGGCCTCCAGACCATTAACC and R-TGGGTATTGGCCCATATGTC; Hoxc8,
F-CGGCAGCAAGCACAAAGA and R-GCGGCTCTCTGCTCACTTT; Hoxc13, F-CACGCCTTCTCTCTCCCTCT and R-CATAAATAGTCGGGGCAGGA; Krt7, F-GCCTGAGGTCAGGGAGTT and R-CTTCCACCACATCCTGCAT; Kb21, F-CTGTGCGTGTCAGGTTCCAG and R-GCCAGGGATGACTCAGGATG; Kb23, F-TGAAGAAGAAGTTGCACTCC and R-CCCCTTGCCAGCAGGTAG; Kb24, F-GGCTTGGGTGGTTTTGGTAG and R-ATTCCTCCAATTCTGTAGCC; Kb25, F-ATTGACAGGCTTCAGCAGTC and R-ACTCGTTCTCTGCCGTAGCC; Kb26, F-GATCTCTGTGTCTCCGGTAC and R-TTGCAGCTTTGTGGCCTCCT; K6irs1, F-TCTTCCCCTGTCAGCATCTC and R-TCAGACTGGAGCCCTTTGTT; β-actin, F-AGCCATGTACGTAGCCATCC and R-CTCTCAGCTGTGGTGGTGAA. The RT-PCR products were subjected to electrophoresis in a 2% agarose gel, and visualized with ethidium bromide.
Location of the Deletion Breakpoints––PCR was performed on genomic DNA derived from the livers of HHRs and SDRs. Amplification was cycled 35, each consisting of 30 s at 94ºC, 30 s at 60ºC and 2 min at 72ºC. Primers used for amplifying intron/exon of the individual basic keratin genes were designed according to the Primer 3 (http://primer3.sourceforge.net/). The primers shown in Fig. 3A and B were as follows (5’→3’): Krt7, F-ACTGACGTGGGTATGGCTTT and R-AACCTAGGCTCTGGGACCTC; Krt2-25, F-TTCTCCCTCTCCCACAACAG (corresponding to the intron 1 region) and R-CTTATGGTTGGGGGTTAGCA (intron 2 region); RGD1305207,
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
F-GGACTCTACCAGGTGCTCCA and R-CTCTGCCTTCCCAGCTCTAA; Kb21, F-GGATTCTCCCTGCTCTCTGA and R-ATTCATGAGTCGCCTTTTGG; Kb26, F-CTTCCCAGAGCCCCTCTACT and R-TTCATGTCCTGGGCTTTGAT; Kb23, F-CTGAAGGCACCCTTAGAACG and R-AGGTTGGTCCTCTCCCAGAT; Kb25, F-CCAGTCTGTTGGGGAGACA (corresponding to the intron 1 region) and R-CTGTATCCTGCTTGGGGTGT (intron 1 region); (a), F-AAGTTCCAAAGGGCTTCCAC and R-AATTCCCTTTCTCCCCTCCT; (b), F-GAGCCAACAGTAGGATGAGCA and R-TTTCAGCATTTCGCTTTGC; (c), F-GCTGTTCAAGAAGCCTTTGC and R-GGGATGAGTGGAGAAGGTCA; (d), F-TTCACGAACGTCTGTCTCCA and R-ACCCCAGTGTAAGGGCTGTA; R2, GACTGGATAAGCCCCTTGTG. The sequences of the F1, F2 and R1 were presented in Fig. 3D. The PCR products using the primers F2 and R1 were sequenced with a Big Dye Terminator Kit (PE Biosystem, Foster City, CA) and an ABI DNA sequencer Model 310 (PE Biosystems).
Sequencing of Kb21, Kb23 and Krt2-25 cDNAs in SDR––These full-length cDNAs were prepared by RT-PCR employing the Omniscript® Reverse Transcription Kit (Qiagen) and the primers described below. PCR was performed essentially according to manufacturer’s instructions. Kb21, F-ATGACCTGTGGATCAGGATT and R-GAAGCCAGTATTCACTTC; Kb23, F-ATGACCTGTGGCTTTCAAACTG and R-GCAGCAAGCCCCTTGC; Krt2-25, F-ATGTCCTGCTTCTCCTCCC and R-AGCAAAGCGCACAGAGC. The PCR products were sequenced as described above.
Extraction of Rat Hair Proteins––Hair proteins were extracted from hairs of SDRs and HHRs, as described by Winter et al. (15).
SDS-Polyacrylamide Gel Electrophoresis (PAGE) and Western Blotting––SDS-PAGE was carried out by the method of Laemmli (27) on a 10% (w/v) polyacrylamide gels. Gels were stained with Coomassie Brilliant Blue R250. Western blotting was performed, according to the method of Towbin et al. (28). To purify basic hair keratins, SDR 51-kDa proteins and HHR 56-kDa protein resolved by SDS-PAGE, the gel portions containing the respective proteins were cut with a razor, homogenized in 1% SDS, 20 mM Tris-HCl (pH 8.0), and then rotated overnight. The supernatant fractions after centrifugation at 15,000×g for 10 min were dialyzed against distilled water and then lyophilized.
Production of Anti-Kb25 and Anti-Kb21 Antibodies––To produce anti-Kb25 antibody, the HHR 56-kDa protein thus purified was dialyzed against phosphate-buffered saline. The protein (200 µg protein/1.5 ml) was emulsified with an equal volume of Freund’s complete adjuvant (Iatron Laboratories, Tokyo, Japan), and then injected subcutaneously 4 times into an albino rabbit at 1-week intervals. The rabbit was bled on the seventh day after the last injection, and antiserum was purified by ammonium sulfate fractionation. Specificity of the antibody was examined by Western blotting. To produce anti-Kb21 antibody, the C-terminal peptide of Kb21 was synthesized by a peptide synthesizer (model 432A, Applied Biosystems, Foster City, CA), based on the amino acid sequence of the rat Kb21 (473SAVSCGRKC481) that was specific to the keratin (4). The peptide was coupled to keyhole limpet hemocyanine with
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
m-maleimidobenzoyl N-hydroxysuccinimide ester (29). This peptide-hemocyanine complex was injected into a rabbit to produce anti-Kb21 antibody, as described above.
Liquid Chromatography/Electrospray-Ionization Mass Spectrometry (LC/ESI-MS)––After reduced with dithiothreitol and alkylated with iodoacetamide, proteins were dissolved in 100 mM NH4HCO3 containing 50 ng/µl of trypsin and incubated at 37ºC for 16 h. The samples were applied to a Nano Frontier LC column, C18 (75 µm id x 150 mm, Hitachi-Hitec, Tokyo, Japan) and eluted by a gradient flow of acetonitrile and distilled water containing 0.1% formic acid (flow rate 200 nl/min). The peptide fragments eluted were analyzed using an online coupled linear trap electrospray-ionization mass spectrometry (Nano Frontier L, Hitachi-Hitec) at a heated capillary temperature of 140ºC, and voltage of 1.0 kV. Peptide sequence analysis was performed using BioLynx software (Micromass). The sequence information was submitted to the MASCOT programs (http://www.matrixscience.com/), and the MASCOT score of more than 34 were considered to be the corresponding proteins.
Immunohistochemistry––Skin tissues from the anterior dorsal regions of rats were fixed in 10% formaldehyde and embedded in paraffin. Two-week-old rats were used to obtain hairs in an anagen phase. Immunohistochemical staining for Kb21 or Kb25 was performed by the avidin-biotin-peroxidase complex (ABC) method (30) with the respective antibodies. The biotinylated anti-rabbit antibody and Vectastain ABC kit were obtained from Vector Laboratories (Burlingame, CA). The specific binding was visualized with a 3,3’-diaminobenzidine
tetrahydrochloride solution. Sections were then lightly counterstained with hematoxylin for microscopic examination. The specimens were examined and photographed using a microscope (COOLSCOPE, Nikon, Tokyo, Japan) interfaced with a computer.
RESULTS Mapping of the hhr––To map the hhr locus,
the backcross N2 progeny were produced from the (BN × HHR) F1 mated with HHR. Linkage analysis was performed by typing DNA samples from randomly selected 12-52 hairless N2 animals for 2 visible coat colors (the color on the chromosome 1 and the hooded on the chromosome 14) and 18 microsatellite markers on other chromosomes. Linkage of the hhr was first detected with the D7Rat105 on Chr 7 (Table 1). DNA samples of the 88 progeny were further typed for additional Chr 7 markers. One hairless rat exhibited the heterozygosity at the D7Rat173 gene but all hairless rats showed the homozygosity at the D7Rat3 and D7Rat105 loci without exception (Fig. 1A), indicating that the hhr was localized to the region between the D7Rat173 and the telomere on 7q36. Hair growth-related genes and the basic keratin gene cluster are known to be localized on 7q36 (Fig. 1B). RT-PCR was performed to examine whether expression of these genes is altered in HHR. The Kb21, 23, and 26 genes were not expressed in HHR, but other basic keratin genes were expressed (Fig. 2). Expression of the Wnt10b, Hoxc13 and other genes related to hair growth was retained in HHR.
Deletion of Multiple Basic Keratin Genes in HHR––Since the loci of the Kb21, 23 and 26 genes are localized adjacently, the loss of their expression raised the possibility of the deletion
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
of the genes. To examine this, PCR was preformed for neighboring basic keratin genes (from Krt7 to Kb25, Fig. 3A) in HHR and SDR. The Krt7 and Kb25 genes were amplified, but neither the Krt2-25, RGD1305207, Kb21, Kb26 nor Kb23 were amplified in HHR (Fig. 3A), indicating the deletion of a region containing Krt2-25 to Kb23 gene loci. PCR was repeated to define a deletion range precisely. A DNA portion over 850-bp downstream from the stop codon of the Krt2-25 (broken line (a) in Fig. 3B) and that around the exon 6 of the Kb25 ((d) in Fig. 3B) were amplified in HHR (panels a and d in Fig. 3B), but a region near the exon 6 of the Krt2-25 ((b) in Fig. 3B) or a portion over 850-bp downstream from the stop codon of the Kb25 ((c) in Fig. 3B) were not. Furthermore, PCR products were not amplified with the F1 and R1 primers in HHR, but amplified with the F2 and R2 primers (Fig. 3C). These results suggested that the F1 or R1 portions of the Kb25 were not present in HHR. A single PCR product of about 1.3-kb with the F2 and R2 primers in HHR was much shorter than about 80-kb theoretically expected from the SDR genomic sequence. PCR with the F2 and R1 primers was performed to define the junction portion in HHR, and the DNA sequence of the product was compared with that of SDR (Fig. 3C and D). The HHR product showed the sequence of the Kb25 in the upstream (boxes 1-4 in Fig. 3D and E) but that of the Krt2-25 in the downstream (box 5), while the SDR product exhibited only the Kb25 sequence in all boxes. These results indicated that breakpoints were within a 95-bp portion shared by both the exon 9 of the Kb25 and the exon 9 of the Krt2-25.
Loss of Basic Hair Keratins Kb21 and Kb23 Expression in HHR Hairs––To examine whether
hair protein profiles are different between HHR and SDR, proteins extracted from hairs of 5 months-old rats were subjected to SDS-PAGE. Protein staining with Coomassie brilliant blue revealed two major bands at 51-kDa (basic hair keratin, Hb) and 40-kDa (acidic hair keratin, Ha) in SDR, while in HHR the 51-kDa band was markedly decreased and instead a 56-kDa band (Fig. 4A) became dominant. The SDR 51-kDa band was broad in electrophoretic mobility. The size of acidic keratins was not different between HHR and SDR. To examine immunochemical relationship between the HHR 56-kDa and SDR 51-kDa proteins, Western blotting was performed with anti-Kb25 and anti-Kb21 antibodies. Anti-Kb25 antibody reacted not only with the HHR 56-kDa protein but also with the SDR 56-kDa protein that was very faint on protein staining (Fig. 4B). This antibody also reacted weakly with the HHR 51-kDa and SDR 51-kDa proteins. On the other hand, anti-Kb21 antibody reacted only with SDR 51-kDa proteins but not with HHR 56-kDa protein (Fig. 4C). These results indicated that the HHR 56-kDa protein was immunologically related to but not identical with the SDR 51-kDa proteins.
To identify the basic hair keratins in the SDR 51-kDa proteins, the sample recovered from SDS gels was digested with trypsin and their fragments were subjected to LC/ESI-MS analysis. This revealed the peptide fragments characteristic of Kb21 (ENSRNOT 00000012845), Kb23 (ENSRNOT 00000011222) and Krt2-25 (AB 354637) (underlined in Fig. 5). As the Kb21, Kb23, and Krt2-25 cDNAs, the respective two to four different forms have been reported (4, 31). To confirm the data of the amino acid sequences by LC/ESI-MS and also to identify the keratin
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
forms expressed in SDR skin, cDNAs encoding these keratins were prepared by RT-PCR as described in the Experimental Procedures and sequenced. This demonstrated that the ENSRNOT 00000012845 form and ENSRNOT 00000011222 form were expressed as the Kb21 (481 aa) and Kb23 (479 aa), respectively. Although four forms (XP_001064518 (497 aa), ENSRNOT00000031257 (493 aa), XP_001060591 (491 aa) and XP_001060532 (440 aa)) have been reported as the rat Krt2-25 cDNA, this analysis also revealed that a new form with 495 aa was expressed in SDR. The nucleotide sequence of this form (accession number AB354637) as well as our data of Kb21 (AB355638) and Kb23 cDNAs (AB355639) is available in the DDBJ/EMBL/GenBank databases.
Expression of Kb25 as the Fusion Gene Product in HHR Hair––The HHR 56-kDa protein was also treated with trypsin and subjected to LC/ESI-MS analysis. The peptides exhibited the sequences of Kb25 reported as the ENSRNOP00000011644 (507 aa form), a splice variant of NP_001008811 (4) (472 aa form) and these were different only in the C-terminal side encoded by the exons 8 and 9 (Fig. 6). In this region the sequence of Kb25 (the 507 aa form) is very similar to the Krt2-25 (AB354637) except at the 479th residue, the base at the box 4 in Fig. 3D. Thus, the HHR 56-kDa protein was suggested as the product of a fusion gene of Kb25 and Krt2-25, but identical with the Kb25 itself in amino acid sequence. To demonstrate this, the fusion gene cDNA was prepared by RT-PCR employing the primers for the exon 5 of the Kb25 and for the 3’ downstream of the Krt2-25 (R1 primer in Fig. 3D). Since the R1 primer was common for the exon 9 of the Kb25,
the SDR Kb25 cDNA was also prepared with the same primer set. Sequencing of the SDR Kb25 cDNA indicated the sequence of the ENSRNOP 00000011644 (Fig. 7A), and it differed from the Krt2-25 cDNA at 3 bases in the exon 8 (shaded boxes in Fig. 7A) and 7 bases in the exon 9 (shaded one base corresponding to the box 4 in Fig. 3D and shaded 6 bases in the 3’ untranslated region).
Characterization of HHR Hair Follicles by Immunohistochemistry––To examine the expression of Kb25 and Kb21 in HHR hair follicles, immunohistochemistry was performed. With anti-Kb21 antibody the hair cortex of SDR showed positive reaction but HHR hair follicles were negative (Fig. 8), confirming the deletion of basic keratin genes, including the Kb21 in HHR. Staining with anti-Kb25 antibody revealed positive reaction in the cortex (black arrows in Fig. 8) and companion layer (open arrowheads) of HHR. In SDR, the medulla (red arrows) and companion layer were stained but the hair cortex was not. Thus, Kb25 expression in the cortex and medulla was opposite between HHR and SDR.
DISCUSSION
The present study has revealed that genes responsible for HHR are located on the chromosome 7q36 (Fig. 1), and an 80-kb genomic DNA ranging from the downstream of the Kb25 locus to the Krt2-25 is deleted (Fig. 7B). This region includes the Krt2-25, RGD130527, Kb21, Kb26 and Kb23 genes. In accordance with the deletion of these genes, the Kb21, Kb23 and Krt2-25 proteins (51-kDa proteins) were not expressed in HHR (Figs. 2 and 4). In place of the 51-kDa proteins, the 56-kDa protein that was very minor in SDR was
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
dominant in HHR. The nucleotide sequence of the Krt2-25 gene is highly homologous to that of the Kb25, and the breakpoints of these genes are within a 95-bp portion shared by both the terminal region of the exon 9 of the two genes (Figs. 3 and 7), suggesting that the deletion is due to non-allelic homologous recombination. Fusion of the Kb25 gene to the Krt2-25 in HHR raises the possible production of a fusion protein. The sequence of the HHR fusion gene cDNA was identical with that of the SDR Kb25 cDNA in most parts but with the Krt2-25 cDNA in the 3’ untranslated region. In fact, mass spectrometry analysis revealed that the sequence of the HHR 56-kDa protein was identical with that of the wild-type Kb25. These findings suggest that the fusion of the two genes occurs between the exons with the same nucleotide sequence without any alterations in codon frame.
A unique basic cytokeratin, K6hf (Krt75), is reported to be expressed in the medulla and companion layer of the mouse and human (32, 33). The present study revealed the Kb25 to be expressed in the same regions as K6hf. Although the human equivalent of the rat Krt2-25 is considered as a pseudogene (4, 31, 34), it is transcribed and translated in SDRs. Thus, different basic keratins are expressed in the particular regions of hair follicles between SDRs and humans or mice. This study also suggested that different Kb21, Kb23, Kb25 and Krt2-25 forms due to alternative splicing are expressed between Norway rats and SDRs (4). Thus, differences in splicing of the individual keratin transcripts may be involved in the multiplicity of keratins in some strains of the rat.
Human HB1, 3, and 6, the orthologs of the rat Kb21, 23, and 26, respectively, are reported to be expressed in the hair cortex (5). We
confirmed Kb21 to be expressed in the cortex of SDR (Fig. 8). In HHR, Kb25, instead of Kb21, was expressed in the cortex but not expressed in the medulla, the site of its expression in SDR. Not only loss of Kb21, 23 and 26 genes but also altered regulation of Kb25 gene is suggested to be responsible for the defective hair follicle formation in HHR. Detailed characterization of HHR hair follicles is now under active study. Although anti-Kb25 antibody cross-reacted with the SDR 51-kDa proteins on Western blotting (Fig. 4B), the hair cortex was not stained on immunohistochemistry (Fig. 8A). Since the latter was carried out with the antibody at a very low concentration, only Kb25 seemed to be stained.
The shorn (shn) and CR hairless rats share similar properties in hair as HHRs. However, life span is normal in HHR but shorn rats die before 14 months of age. The shn gene is mapped between the D7Got143 and D7Rat94 in 7q36 (35), but responsible genes have not been identified. The gene responsible for hereditary hypotrichotic mouse, hague, is mapped on the telomeric region of the chromosome 15, very close to the basic keratin gene cluster of the mouse (36), but is not identified. Thus, these rat and mouse models seem to share mutations in or around the basic keratin gene cluster.
Homologous recombination is the exchange or replacement of a DNA region between the homologous chromosomes or chromatids. It also occurs between similar DNA sequences of non-homologous chromosomes or within a chromosome (37). These rearrangements lead to the loss or gain of genes (38). Since basic hair keratin genes have highly homologous sequences, these loci are possible hot spots for non-allelic homologous recombination. Frequent
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
assignment of genes responsible for alopecia to the vicinity of these loci in the rodent models supports this possibility. However, the multiplicity of basic keratin genes may compensate for the loss of particular types by the expression of the other types. Thus, the deletion of massive DNA domain may be
required for abnormal hair formation. The rodent models of hypotrichosis due to alterations in keratin genes will be useful to explore responsible genes in such patients, because the arrangement of the individual keratin gene loci is conserved between humans and the animal species (31).
ACKNOWLEDGMENTS We thank Dr. Kosuke Kasai, Department of Pathologic Analysis, Hirosaki University Graduate
School of Health Sciences, for advice and support, and Dr. Noriko Kobayashi, Hitachi-Hitec, Tokyo, Japan, for mass spectrometry.
FOOTNOTES *This work was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science and Technology of Japan and from the Food Safety Commission of Japan and by grants from Hirosaki University School of Medicine and the Hirosaki Foundation for Science and International Exchange. 1 To whom correspondence should be addressed: Department of Biochemistry and Genome Biology, Hirosaki University Graduate School of Medicine, 5 Zaifu-cho, Hirosaki 036-8562, Japan. Tel.: +81-172-39-5018; Fax: +81-172-39-5205; E-mail: [email protected] 2 The Abbreviations used are: RT-PCR, reverse transcription-polymerase chain reaction; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; SDR, Sprague-Dawley rat; BN, Brown-Norway; HHR, Hirosaki hairless rat.
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
REFERENCES 1. Langbein, L., Rogers, M. A., Winter, H., Praetzel, S., and Schweizer, J. (2001) J.
Biol. Chem. 276, 35123-35132 2. Langbein, L., Rogers, M. A., Winter, H., Praetzel, S., Beckhaus, U., Rackwitz, H.
R., and Schweizer, J. (1999) J. Biol. Chem. 274, 19874-19884 3. Heid, H. W., Werner, E., and Franke, W. W. (1986) Differentiation 32, 101-119 4. Hesse, M., Zimek, A., Weber, K., and Magin, T. M. (2004) Eur. J. Cell Biol. 83, 19-26
5. Langbein, L., and Schweizer, J. (2005) Int. Rev. Cytol. 243, 1-78 6. Langbein, L., Rogers, M. A., Praetzel, S., Winter, H., and Schweizer, J. (2003) J.
Invest. Dermatol. 120, 512-522 7. Wojcik, S. M., Longley, M. A., and Roop, D. R. (2001) J. Cell Biol. 154, 619-630 8. Kishimoto, J., Burgeson, R. E., and Morgan, B. A. (2000) Genes Dev. 14, 1181-1185 9. Millar, S. E., Willert, K., Salinas, P. C., Roelink, H., Nusse, R., Sussman, D. J., and
Barsh, G. S. (1999) Dev. Biol. 207, 133-149 10. Luetteke, N. C., Qiu, T. H., Peiffer, R. L., Oliver, P., Smithies, O., and Lee, D. C.
(1993) Cell 73, 263-278 11. Mann, G. B., Fowler, K. J., Gabriel, A., Nice, E. C., Williams, R. L., and Dunn, A.
R. (1993) Cell 73, 249-261 12. Schorpp, M., Schlake, T., Kreamalmeyer, D., Allen, P. M., and Boehm, T. (2000)
Dev. Dyn. 218, 537-543 13. Godwin, A. R., and Capecchi, M. R. (1998) Genes Dev. 12, 11-20 14. McLean, W. H., and Lane, E. B. (1995) Curr. Opin. Cell Biol. 7, 118-125 15. Winter, H., Hofmann, I., Langbein, L., Rogers, M. A., and Schweizer, J. (1997) J.
Biol. Chem. 272, 32345-32352 16. Tong, X., and Coulombe, P. A. (2006) Genes Dev. 20, 1353-1364 17. Hanada, K., Chiyoya, S., Suzuki, K., Hashimoto, I., and Hatayama, I. (1988) J.
Dermatol. 15, 257-262 18. Nanashima, N., Asano, J., Hayakari, M., Nakamura, T., Nakano, H., Yamada, T.,
Shimizu, T., Akita, M., Fan, Y., and Tsuchida, S. (2005) J. Biol. Chem. 280, 43010-43016 19. Panteleyev, A. A., and Christiano, A. M. (2001) Comp. Med. 51, 49-55 20. Castle, W., Dempster, E., and Shurrager, H. (1955) J. Hered. 46, 9-14 21. Festing, M. F., May, D., Connors, T. A., Lovell, D., and Sparrow, S. (1978) Nature
274, 365-366 22. Palm, J., and Ferguson, F. G. (1976) J. Hered. 67, 284-288 23. Roberts, E., Quisenberry, J. E., and Tomas, L. C. (1940) J. Invest. Dermatol. 3, 1-29
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
24. Moemeka, A. N., Hildebrandt, A. L., Radaskiewicz, P., and King, T. R. (1998) J.
Hered. 89, 257-260 25. Inazu, M., Kasai, K., and Sakaguchi, T. (1984) Lab. Anim. Sci. 34, 577-583 26. Nehls, M., Pfeifer, D., Schorpp, M., Hedrich, H., and Boehm, T. (1994) Nature 372, 103-107
27. Laemmli, U. K. (1970) Nature 227, 680-685 28. Towbin, H., Staehelin, T., and Gordon, J. (1979) Proc. Natl. Acad. Sci. U S A. 76, 4350-4354 29. Peeters, J. M., Hazendonk, T. G., Beuvery, E. C., and Tesser, G. I. (1989) J.
Immunol. Methods 120, 133-143 30. Hsu, S. M., Raine, L., and Fanger, H. (1981) J. Histochem. Cytochem. 29, 577-580 31. Rogers, M. A., Edler, L., Winter, H., Langbein, L., Beckmann, I., and Schweizer, J.
(2005) J. Invest. Dermatol. 124, 536-544 32. Winter, H., Langbein, L., Praetzel, S., Jacobs, M., Rogers, M. A., Leigh, I. M.,
Tidman, N., and Schweizer, J. (1998) J. Invest. Dermatol. 111, 955-962 33. Wang, Z., Wong, P., Langbein, L., Schweizer, J., and Coulombe, P. A. (2003) J.
Invest. Dermatol. 121, 1276-1282 34. Schweizer, J., Bowden, P. E., Coulombe, P. A., Langbein, L., Lane, E. B., Magin, T. M., Maltais, L., Omary, M. B., Parry, D. A. D., Rogers, M. A., and Wright, M. W. (2006) J. Cell Biol.
174, 169-174 35. Chrissluis, R. R., Stoklasek, T. A., Loman, J. A., Guariglia, A., and King, T. R.
(2002) Mol. Genet. Metab. 76, 335-339 36. Poirier, C., Yoshiki, A., Fujiwara, K., Guenet, J. L., and Kusakabe, M. (2002)
Genetics 162, 831-840 37. Lupski, J. R. (2006) Nat. Genet. 38, 974-976 38. Sharp, A. J., Hansen, S., Selzer, R. R., Cheng, Z., Regan, R., Hurst, J. A., Stewart, H., Price, S. M., Blair, E., Hennekam, R. C., Fitzpatrick, C. A., Segraves, R., Richmond, T. A., Guiver, C., Albertson, D. G., Pinkel, D., Eis, P. S., Schwartz, S., Knight, S. J., and Eichler, E. E. (2006) Nat.
Genet. 38, 1038-1042
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
FIGURE LEGENDS
FIGURE 1. Genetic linkage studies and physical map. (A) Segregation of the alleles of three microsatellite marker loci of 7q36 in 88 N2 progeny obtained from backcrosses between HHRs and the F1 hybrid derived from BN strain. Mutant denotes hypotrichotic phenotype (HHR) while non-mutant does wild phenotype. Open boxes represent the homozygosity of the individual marker alleles for HHR and closed boxes do the heterozygosity. The number of N2 progeny with each haplotype are indicated at the bottom. (B) Physical map of the marker genes, hair growth-related genes, and the basic keratin gene cluster on rat chromosome 7q36.
FIGURE 2. Gene expression analysis of hair growth-related genes localized on 7q36 by RT-PCR. RT-PCR was performed using the primers for the individual genes and mRNA isolated from the skin of HHR and SDR. VDR, vitamin D receptor; Kb21, basic hair keratin 21; Krt7, cytokeratin 7; K6irs1, cytokeratin 6 inner root sheath 1.
FIGURE 3. Deletion of an 80-kb genomic DNA containing basic keratin gene loci in HHR and identification of breakpoints. (A) Map of the seven basic keratin genes (Krt7––Kb25) clustered on 7q36 and their PCR amplification results in HHR and SDR. (B) Scheme for exon structure of Krt2-25 and Kb25 genes and results of genomic PCR. Open boxes represent the deleted exons of Krt2-25 and closed boxes represent remaining ones in HHR. Solid lines show highly homologous regions and broken lines non-homologous ones between Krt2-25 and Kb25. The broken lines (a) and (c) represent the region over 850-bp downstream from the stop codons of Krt2-25 and Kb25, respectively. The (b) and (d) denote the introns upstream from the exon 6 of Krt2-25 and Kb25, respectively. F1 and F2 represent forward primers for Kb25; R1 and R2 represent reverse primers for Krt2-25. R1 also works as the primer for Kb25. (C) Abnormal PCR product in HHR by the deletion of multiple genes. Lane M, DNA molecular marker. The primers, F1, F2, R1 and R2, were the same as in (B). (D) The highly homologous sequences of parts of Kb25 intron 8/exon 9 and Krt2-25 intron 8/exon 9. F1 and F2 represent forward primers for Kb25 as described in (B). R1 does a reverse primer for both Kb25 and Krt2-25. An open box shared by both Kb25 and Krt2-25 represents break and fusion points of the two genes. Shaded boxes (1-5) denote different sequences between Kb25 and Krt2-25 genes. An asterisk represents the stop codon of Kb25 and Krt2-25 genes. (E) Sequence results of the products amplified with F2-R1 in (C). Boxes 1-4 indicated Kb25 sequences in HHR and SDR. The box 5 indicated Krt2-25 sequence in HHR but Kb25 in SDR.
FIGURE 4. SDS-PAGE of hair keratins of HHR and SDR and Western blotting with anti-Kb25 and Kb21 antibodies. (A) Hair proteins of HHR and SDR resolved by SDS-PAGE. Each 20 µg of protein was applied to a lane and stained with Coomassie Brilliant Blue. Hb, basic hair keratin; Ha, acidic hair keratin. (B and C) Western blotting with anti-Kb25 antibody and anti-Kb21 antibody, respectively.
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
FIGURE 5. Demonstration of the Kb21, Kb23, and Krt2-25 forms in the SDR 51-kDa proteins by liquid chromatography/electrospray-ionization mass spectrometry (LC/ESI-MS). The SDR 51-kDa proteins eluted from the SDS gels were digested with trypsin and then subjected to LC/ESI-MS, as described in the Experimental Procedures. The amino acid sequences of the Kb21, Kb23, and Krt2-25 were deduced from the sequences of the respective cDNAs. Kb21, Kb23 and Krt2-25 cDNA forms expressed in SDR were determined by RT-PCR as ENSRNOT00000012845, ENSRNOT00000011222 and AB354637, respectively. The peptides identified as those of Kb21, Kb23, and Krt2-25 by LC/ESI-MS are underlined. Among the peptides with the same sequences, only one peptide is underlined for simplicity. This does not indicate that it is derived from the underlined keratin form.
FIGURE 6. Identification of the HHR 56-kDa protein as the Kb25 form by LC/ESI-MS. The HHR 56-kDa protein eluted from the SDS gels was also digested with trypsin and subjected to LC/ESI-MS. The amino acid sequence of Kb25 was deduced from the cDNA sequence, and the Kb25 cDNA form expressed in HHR and SDR was determined as ENSRNOP00000011644 by RT-PCR (one asterisk). The peptides identified as those of Kb25 by LC/ESI-MS are underlined. The amino acid sequence of another form of Kb25 (NP001008811, two asterisks) is added for reference.
FIGURE 7. Identification of the HHR 56-kDa protein as the product of Kb25-Krt2-25 fusion gene. (A) Nucleotide sequences of RT-PCR products from a HHR Kb25-Krt2-25 fusion gene, SDR Kb25 and SDR Krt2-25 genes. RT-PCR was carried out employing primers for the exon 5 of Kb25 and for the 3’ downstream of Krt2-25, as described in the Experimental Procedures. The partial sequences of the regions corresponding to the exons 7––9 of the genes are presented. The sequences of the HHR fusion gene cDNA and SDR Krt2-25 cDNA are shown by hyphens when identical with those of SDR Kb25 cDNA. Shaded 3 bases in the exon 8 and one base in the exon 9 (corresponding to the box 4 in Fig. 3D) differ between Kb25 and Krt2-25 genes. Shaded 6 bases in the 3’ untranslated region (box 5 in Fig. 3D) are present in SDR Kb25 cDNA but absent in Krt2-25 cDNA and HHR fusion gene cDNA (bold solid lines). Amino acid sequences are also described in a single letter. An asterisk denotes the stop codon. (B), Schematic representation of the deletion of basic keratin genes in an 80-kb region, breakpoints, and the fusion of Kb25 and Krt2-25 genes in HHR. Open boxes represent deleted genes and closed boxes remaining ones in HHR. Grey boxes and grey line represent the exon 9 and the 3’ downstream region of Krt2-25, respectively. An asterisk represents the stop codon of Kb25 and Krt2-25. Broken lines represent 95-bp break and fusion points shared by both the terminal side of the exon 9 of Kb25 (hatched boxes) and Krt2-25 (grey boxes).
FIGURE 8. Immunohistochemical analysis of SDR and HHR hair follicles with anti-Kb25 and anti-Kb21 antibodies. Skin sections of two-week-old SDR (A, C and E) and HHR (B, D and F) were stained with anti-Kb25 antibody (A and B), anti-Kb21 antibody (C and D) and non-immune control
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
γ-globulin (E and F). Black arrows, red arrows, and open arrowheads denote the hair cortex, medulla, and companion layer, respectively. All images were at 400x magnification.
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Table 1 Segregation ratio of marker loci in mutant (hhr /hhr ) N2
backcross progeny
Color (D1)b 23 29 0.48 0.25<P<0.5
D2Rat187 14 10 0.38 0.5<P<0.75
D3Arb3 8 16 2.04 0.1<P<0.25
D4Arb15 11 5 1.56 0.1<P<0.25
D5Rat116 12 12 0.42 0.5<P<0.75
D6Arb5 10 6 0.56 0.25<P<0.5
D7Rat105 24 0 22.0 P<0.005
D8Arb5 8 8 0.06 0.75<P<0.9
D9Mit7 5 11 1.56 0.1<P<0.25
D10Rat103 8 8 0.06 0.75<P<0.9
D11Rat27 9 6 0.27 0.5<P<0.75
D12Rat49 7 5 0.08 0.75<P<0.9
D13Arb11 9 11 0.05 0.75<P<0.9
Hooded (D14)b 15 13 0.04 0.75<P<0.9
D15Rat29 5 9 0.64 0.25<P<0.5
D16Rat31 11 9 0.05 0.75<P<0.9
D17Rat76 10 14 0.38 0.5<P<0.75
D18Mit1 7 11 0.50 0.25<P<0.5
D19Rat67 8 8 0.06 0.75<P<0.9
D20Rat46 9 7 0.06 0.75<P<0.9
aHomozygosity denotes homozygous for the individual marker alleles for HHR.
b Color and Hooded are visible coat color marker loci on chromosomes
1 and 14, respectively.
χ² PMicrosatellite
Homozygositya Heterozygosity
No. of rats
marker
15
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

D7Rat173
D7Rat1054245 1
Mutant Non-mutantA
135
137
139
141
Mb
Rat105
Rat3
Rat173
Kb1, Kb15, Kb20, Krt 7, Kb21, Kb26, Kb23, Kb25, Kb24, Kb22, K6irs
VDR Wnt10b
Sp1 Hoxc8, Hoxc13
B
hhr
D7Rat3
tel
Fig. 1
16
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

VDR
Wnt10b
Sp1
Hoxc8
Hoxc13
β-actin
Kb21
Kb23
Kb26
Kb24
Krt7
K6irs1
HHR HHRSDR SDR
Kb25
Fig. 2
17
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

A
B
M HHR SDR HHR SDR HHR SDR
500bp →
F1 - R1 F2 - R2 F2 - R1C
679
Krt2-25 Kb25
HHR SDR HHR SDR HHR SDR HHR SDR
a b c
da b c
dF1 F2R2 R1
Fig. 3
HHR SDR HHR SDR HHR SDR HHR SDR HHR SDR HHR SDR HHR SDR
Krt7 Krt2-25 RGD1305207 Kb21 Kb23Kb26 Kb25 tel
679 8
500bp
1 2 3 4 5
Krt2-25
HHR
EKb25
SDR
R1
D
Kb25Krt2-25
TGAAAA-GTAGAACCTGTCAG-----CAACTGATCCCTTGAGGAGGAGGCAGGTCCTGTGTGAAAAAGCAGAACCTGTCATGTTAACCACTGACCCCTTGAGGAAAAGGCAGGTCCTGTG
TGTGAAGCCCTCAGAGACTTCTGGTGGGTGGGT CATCA TGTGAAGCCCTCAGGGACTTCTGGTGGGTGGGT CATCT
CAGTGATGGCCCCTGACTCCTGCTCTCCTTGCCAGCCCCGTGCCTCCAGCTTCACCTGTGCAGTGATGGCCCCTGACTCCTGCTCTCCTTGCCAGCCCCGTGCCTCCAGCTTCACCTGTG
GGAGCAGCCGCTCTGTGCGCTTTGCTTAGTGCCAGGACCTCCAAATGAGCCACCTCTCCTGGAGCAGCCGCTCTGTGCGCTTTGCTTAGTGCCAG------CAAATGAGCCACCTCTCCT
CTGGCTTTACTTTAGAAGGTCATTCATTCCCAACCCGTCC CTGGCTTTACTTTAGAAGGTCATTCATTCCCAACCCGTCC
F2
R1
F1
1 2
3 4
5
exon 9exon 9
*
8
18
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from
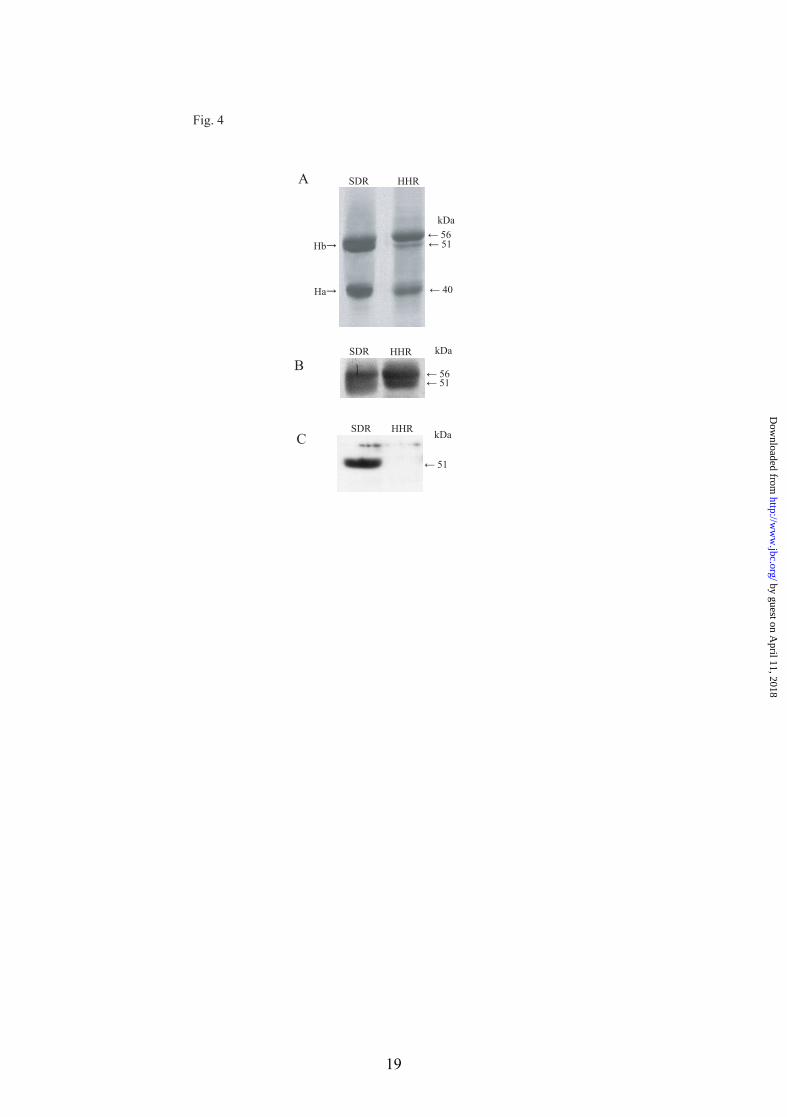
Fig. 4
HHRSDR
Hb→
Ha→
← 56← 51
← 40
kDa
A
B
C
SDR
SDR
HHR
HHR
kDa
← 51← 56
← 51
kDa
19
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kb21 1 MTCGSGFCGR AFSCASACGP RPGRCCITAA PYRGISCYRG LSGGFGSRSV CGAFRSGSCG
Kb23 1 MTCGFGTG NFG------- ---------- ---------- ---------- ----------
Krt2-25 1 MSCFS SRLGASCGV- ---------- ---------- ---------- -T-------- ----------
61 RSFGYRSGGV CGPSPPCITT VSVNESLLTP LNLEIDPNAQ CVKHEEKEQI KCLNSRFAAF
59 ---------- ---------- ---------- ---------- ---------- ----------
66 ---------- ---------- ---------- ---------- ---------- ----------
121 IDKVRFLEQQ NKLLETKWQF FQNRKCCESN VEPLFEGYIE TLRREAECVE ADSGRLAAEL
119 ---------- ---------- ---------- ---------- ---------- ----------
126 ---------- ---------- Y--------- L--------- -------Y-- ----------
181 NHAQEAMEGY KKRYEEEVSL RATAENEFVA LKKDVDCAYL RKSDLEANTE ALTQEIDFLR
179 ---------- --------A- ---------- ---------- --------A- ----------
186 --V------- --------A- ---------- ---------- --------A- ----------
241 RLYEEETRIL HAHISDTSVI VKMDNSRDLN MDCVIAEIKA QYDDIASRSR AEAESWYRTK
239 ---------- ---------- ---------- ---------- ---------- ----------
246 --------L- ---------- ---------- ----V----- ---------- ----------
301 CEEIKATVIR HGETLRRTRE EINELNRIIQ RLTAEIENAK CQNTKLEAAV TQSEQQGEAA
299 ---------- ---------- ---------- ---------- ---------- ----------
304 ---M------ ---------- -M-------- ---------- --RA---T-- AEA-------
361 LTDARCKLAE LEAALQKAKQ DMACLLKEYQ EVMNSKLGLD IEIATYRHLL EGEEQRLCEG
359 ---------- ---------- ---------- ---------- -------R-- ----------
364 -----G--T- ---------- ---------- ---------- -------R-- ----H-----
421 VGAVNVCVSS SRGGVVCGDL CVSGSRPVTG SACSAPCSGN LAVNTGLCAP CGSAVSCGRK
419 ---------- ---------- ---------- -V-------- V--S------ ---GPCHQGR
424 --S------- -----T--G- TYGTTPGRQI ASGPSVTGGS ISVMAPDSCS PCQPRASSFT
481 C
479 C
484 CGSSRSVRFA
Fig. 5
20
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Kb25* 1 MSCRSYRISP GCGVTRNFSS CSAVAPKTGN RCCISAAPFR GVSCYRGLTG FSSRSLCNPS Kb25** ---------- ---------- ---------- ---------- ---------- ----------
61 PCGPRMAVGG FRSGSCGRSF GYRAGGVCGP SPPCITTVSV NESLLTPLNL EIDPNAQCVK ---------- ---------- ---------- ---------- ---------- ----------
121 HEEKEQIKCL NSKFAAFIDK VRFLEQQNKL LETKWQFYQN QRCCESNLEP LFGGYIETLR ---------- ---------- ---------- ---------- ---------- ----------
181 REAECVEADG GRLAAELNHV QEAMEGYKKR YEEEVALRAT AENEFVVLKK DVDCAYLRKS ---------- ---------- ---------- ---------- ---------- ----------
241 DLEANVEALV EESSFLKRLY EEEVRVLQAH ISDTSVIVKM DNSRDLNMDC VVAEIKAQYD ---------- ---------- ---------- ---------- ---------- ----------
301 DVASRSRAEA ESWYRTKCEE MKATVIRHGE TLRRTREEMN ELNRIIQRLT AEIENAKCQR ---------- ---------- ---------- ---------- ---------- ---------- 361 AKLETAVAEA EQQGEAALTD ARGKLAELEA ALQKAKQDMA CLLKEYQEVM NSKLGLDIEI ---------- ---------- ---------- ---------- ---------- ----------
421 ATYRRLLEGE EHRLCEGVGS VNVCVSSSRG GVTCGGLTYG TTPGRQIASG PSVTGGSITV ---------- ---CQQLPWR SHMWGPHLWH YPRAPDCLWT LCHRGQHHSD GP
481 MAPDSCSPCQ PRASSFTCGS SRSVRFA
Fig. 6
21
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Krt2-25Kb25
80 kb
95 bp100 bpbreak point
Kb24Krt7
Krt7 Krt2-25 RGD1305207 Kb21 Kb26 telKb24
SDR
HHR
Kb23
exon 9
Kb25
B
exon 9*
Aexon 8 exon 9
SDR Krt2-25
exon 7 SDR Kb25
HHR Kb25-Krt2-25 fusion gene
SDR Kb25 protein HHR Kb25-Krt2-25 fusion protein
SDR Krt2-25 protein
SDR Kb25 HHR Kb25-Krt2-25 fusion gene
SDR Krt2-25
Nucleotide sequence
Nucleotide sequence
Amino acid sequence
Amino acid sequence
GAGCACAGGCTATGCGAGGGTGTGGGCTCTGTGAATGTCTGTGTCAGCAGCTCCCGTGGA-----------------------------------------------------------------------G--T--A------------------------------------------ E H R L C E G V G S V N V C V S S S R G - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - - -
GGAGTCACA ATCACAGTGATGGCCCCTGA TTTGCTTAGTGCCAGGACCTCCAA --------- -------------------- --------------- ------------ G ---T----------------V ---------------T --- G V T I T V M A P D F A * - - - - - - - - - - - - - - - -V M - S - - - - - - - -
SDR Kb25 protein HHR Kb25-Krt2-25 fusion protein
SDR Krt2-25 protein
Fig. 7
22
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

SDR HHR
Fig. 8
A B
C D
FE
23
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Yang Fan and Shigeki TsuchidaNaoki Nanashima, Miki Akita, Toshiyuki Yamada, Takeshi Shimizu, Hajime Nakano,
genomic DNA containing five basic keratin genesThe hairless phenotype of the hirosaki hairless rat is due to the deletion of an 80-kb
published online April 16, 2008J. Biol. Chem.
10.1074/jbc.M802539200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















