
The Formation of Plasma Lipoproteins from Apoprotein in ... HE-02965 and Training Grant 2 TI HE-5273...
9
THE Jonmiar, OF BIOLOGICAL CHE~GTRY Vol. 240, No. 7, July 1965 Printed in U.S.A. The Formation of Plasma Lipoproteins from Apoprotein in Plasma* PAUL S. RoHEIM,t LIBBY MILLER, AND HOWARD A. EDER From the Departments of ikfedicine and Physiology, Albert Einstein College of Medicine, New York, New York 10461 (Received for publication, August 10, 1964) In a previous paper (1) we reported that, when livers from rats fed diets containing 2% cholesterol were perfused with whole blood from normal rats, the concentration of cholesterol and triglyceride in the perfusate increased significantly. These lipids were present almost entirely in the lipoproteins of density (d) < 1.019. In the present investigations the transfer of lipids from the liver into the plasma in the form of lipoproteins has been studied in more detail. We have found that specific plasma proteins participate in the transfer of lipids by combining with lipid in the liver to form lipoproteins. Preliminary reports of this work have been published (24). EXPERIMENTAL PROCEDURE Animals-Male rats of the Sprague-Dawley strain (Holtz- man Laboratories) weighing 400 to 500 g were used. The rats used as blood donors were fed Rockland Farms mouse pellets. Except when specifically noted, the rats used as liver donors were fed the same diet, to which had been added 2 g of cholesterol and 20 g of olive oil per 100 g of diet for periods of from 7 to 17 days. Liver Perfusions-Livers were perfused by the method of Miller et al. (5) and Green and Miller (6) for 3 hours. The blood for perfusion was obtained by cardiac puncture with heparinized syringes from rats under ether anesthesia. The plasma was separated, and the erythroeytes were washed with Ringer’s solution. The perfusate was constituted by mixing the erythrocytes with whole plasma, d > 1.21 protein solution, or Ringer’s solution to give a final hematocrit of approximately 30%. To this perfusate were added 250 mg of glucose and 20 mg of heparin. The total volume of the perfusate varied be- tween 80 and 140 ml. Preparations of d > 1.21 Proteins-The plasma proteins of d > 1.21 were separated from the lipoproteins by ultracentrifuga- tion of rat plasma of cl 1.21 for 60 hours at 15” in a Spinco model L ultracentrifuge at 114,000 x g. The tubes were cut 2 cm from the top, and the top fraction, containing all of the lipoproteins, was removed. The bottom fraction (d > 1.21) was dialyzed against Ringer’s solution containing 0.170 disodium EDTA, at pH 7, for 24 to 48 hours. A final dialysis against Ringer’s solu- tion was carried out. Proteins of d > 1.21, labeled with 14C- amino acids, were prepared in the following manner. -A protein * Supported by United States Public Health Service Research Grant HE-02965 and Training Grant 2 TI HE-5273 and by a grant from the American Heart Association. t A portion of this work was done during the tenure of an Es- tablished Investigatorship of the American Heart Association. hydrolysatel prepared from Chlorella grown in the presence of 14C02 was neutralized to pH 7, and aliquots of this hydrolysate containing between 35 and 80 PC of i4C were injected into the tail veins of the blood donor rats. After 4 to 5 hours the rats were bled, and the d > 1.21 protein was separated as described. Analytical Methods-Plasma lipoproteins were separated from the perfusate by the method of Havel, Eder, and Bragdon (7) into three fractions of d < 1.019, d 1.019 to 1.063, and d 1.063 to 1.21. The lipoprotein fractions were washed by resuspending them in a salt solution of appropriate density and repeating the centrifugation. Proteins were precipitated with trichloroacetic acid at a final concentration of 5%. The d < 1.019 lipoprotein fraction formed a fluffy precipitate of low density which would not sediment upon centrifugation. When ethyl ether was layered over the water phase, there was partial extraction of the lipid, and the precipitate could then be separated by centrifuga- tion. The protein precipitates were washed three times with 5% trichloroacetic acid, and the lipids were extracted with a solution of ethanol-acetone (I: 1) and subsequently with ether, as described by Radding and Steinberg (8). Proteins were dissolved in appropriate amounts of 1 N NaOH, aliquots were taken for protein determination by the method of Lowry et al. (9), and radioactivity was determined in a Packard Tri-Carb liquid scintillation counter by the method of Vaughan, Steinberg, and Logan (lo), as modified by Radding and Steinberg (8). Cholesterol was determined by t,he method of Abel1 et al. (11); triglycerides were measured according to Van Handel’s modifica- tion (12) of the method of Van Handel and Zilversmit (13); and phospholipids were determined by the method of Beveridge and Johnson (14). RESULTS Release of Lipid during Liver Perfusions-Animals fed t.he cholesterol-containing diet had fatty livers (Table I). The cholesterol and triglyceride concentrations were elevated ap- preciably. When such livers were perfused with whole hepa- rinized blood, a gradual release of triglyceride, cholesterol, and phospholipid into the perfusate occurred (Fig. I). Appreciably more triglyceride than cholesterol or phospholipid was released, and more cholesterol than phospholipid was released. When the perfusing solution consisted of washed rat erythrocytes suspended in Krebs-Ringer-bicarbonate solution instead of plasma, con- siderably less lipid was released into the perfusate (Fig. 1). In these experiments the release of lipids ceased after about 2 hours of perfusion with whole blood. That this was not due to changes 1 Obtained from Picker X-Ray, Volk Radiochemical Company, or New England Nuclear Corporation. by guest on June 29, 2018 http://www.jbc.org/ Downloaded from
Transcript of The Formation of Plasma Lipoproteins from Apoprotein in ... HE-02965 and Training Grant 2 TI HE-5273...

THE Jonmiar, OF BIOLOGICAL CHE~GTRY Vol. 240, No. 7, July 1965
Printed in U.S.A.
The Formation of Plasma Lipoproteins from Apoprotein in Plasma*
PAUL S. RoHEIM,t LIBBY MILLER, AND HOWARD A. EDER
From the Departments of ikfedicine and Physiology, Albert Einstein College of Medicine, New York, New York 10461
(Received for publication, August 10, 1964)
In a previous paper (1) we reported that, when livers from rats fed diets containing 2% cholesterol were perfused with whole blood from normal rats, the concentration of cholesterol and triglyceride in the perfusate increased significantly. These lipids were present almost entirely in the lipoproteins of density (d) < 1.019. In the present investigations the transfer of lipids from the liver into the plasma in the form of lipoproteins has been studied in more detail. We have found that specific plasma proteins participate in the transfer of lipids by combining with lipid in the liver to form lipoproteins. Preliminary reports of this work have been published (24).
EXPERIMENTAL PROCEDURE
Animals-Male rats of the Sprague-Dawley strain (Holtz- man Laboratories) weighing 400 to 500 g were used. The rats used as blood donors were fed Rockland Farms mouse pellets. Except when specifically noted, the rats used as liver donors were fed the same diet, to which had been added 2 g of cholesterol and 20 g of olive oil per 100 g of diet for periods of from 7 to 17 days.
Liver Perfusions-Livers were perfused by the method of Miller et al. (5) and Green and Miller (6) for 3 hours. The blood for perfusion was obtained by cardiac puncture with heparinized syringes from rats under ether anesthesia. The plasma was separated, and the erythroeytes were washed with Ringer’s solution. The perfusate was constituted by mixing the erythrocytes with whole plasma, d > 1.21 protein solution, or Ringer’s solution to give a final hematocrit of approximately 30%. To this perfusate were added 250 mg of glucose and 20 mg of heparin. The total volume of the perfusate varied be- tween 80 and 140 ml.
Preparations of d > 1.21 Proteins-The plasma proteins of d > 1.21 were separated from the lipoproteins by ultracentrifuga- tion of rat plasma of cl 1.21 for 60 hours at 15” in a Spinco model L ultracentrifuge at 114,000 x g. The tubes were cut 2 cm from the top, and the top fraction, containing all of the lipoproteins, was removed. The bottom fraction (d > 1.21) was dialyzed against Ringer’s solution containing 0.170 disodium EDTA, at pH 7, for 24 to 48 hours. A final dialysis against Ringer’s solu- tion was carried out. Proteins of d > 1.21, labeled with 14C-
amino acids, were prepared in the following manner. -A protein
* Supported by United States Public Health Service Research Grant HE-02965 and Training Grant 2 TI HE-5273 and by a grant from the American Heart Association.
t A portion of this work was done during the tenure of an Es- tablished Investigatorship of the American Heart Association.
hydrolysatel prepared from Chlorella grown in the presence of 14C02 was neutralized to pH 7, and aliquots of this hydrolysate containing between 35 and 80 PC of i4C were injected into the tail veins of the blood donor rats. After 4 to 5 hours the rats were bled, and the d > 1.21 protein was separated as described.
Analytical Methods-Plasma lipoproteins were separated from the perfusate by the method of Havel, Eder, and Bragdon (7) into three fractions of d < 1.019, d 1.019 to 1.063, and d 1.063 to 1.21. The lipoprotein fractions were washed by resuspending them in a salt solution of appropriate density and repeating the centrifugation. Proteins were precipitated with trichloroacetic acid at a final concentration of 5%. The d < 1.019 lipoprotein fraction formed a fluffy precipitate of low density which would not sediment upon centrifugation. When ethyl ether was layered over the water phase, there was partial extraction of the lipid, and the precipitate could then be separated by centrifuga- tion. The protein precipitates were washed three times with 5% trichloroacetic acid, and the lipids were extracted with a solution of ethanol-acetone (I: 1) and subsequently with ether, as described by Radding and Steinberg (8). Proteins were dissolved in appropriate amounts of 1 N NaOH, aliquots were taken for protein determination by the method of Lowry et al. (9), and radioactivity was determined in a Packard Tri-Carb liquid scintillation counter by the method of Vaughan, Steinberg, and Logan (lo), as modified by Radding and Steinberg (8). Cholesterol was determined by t,he method of Abel1 et al. (11); triglycerides were measured according to Van Handel’s modifica- tion (12) of the method of Van Handel and Zilversmit (13); and phospholipids were determined by the method of Beveridge and Johnson (14).
RESULTS
Release of Lipid during Liver Perfusions-Animals fed t.he cholesterol-containing diet had fatty livers (Table I). The cholesterol and triglyceride concentrations were elevated ap- preciably. When such livers were perfused with whole hepa- rinized blood, a gradual release of triglyceride, cholesterol, and phospholipid into the perfusate occurred (Fig. I). Appreciably more triglyceride than cholesterol or phospholipid was released, and more cholesterol than phospholipid was released. When the perfusing solution consisted of washed rat erythrocytes suspended in Krebs-Ringer-bicarbonate solution instead of plasma, con- siderably less lipid was released into the perfusate (Fig. 1). In these experiments the release of lipids ceased after about 2 hours of perfusion with whole blood. That this was not due to changes
1 Obtained from Picker X-Ray, Volk Radiochemical Company, or New England Nuclear Corporation.
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from
July 1965 P. S. Rot&m, L. Miller, and H. A. Eder
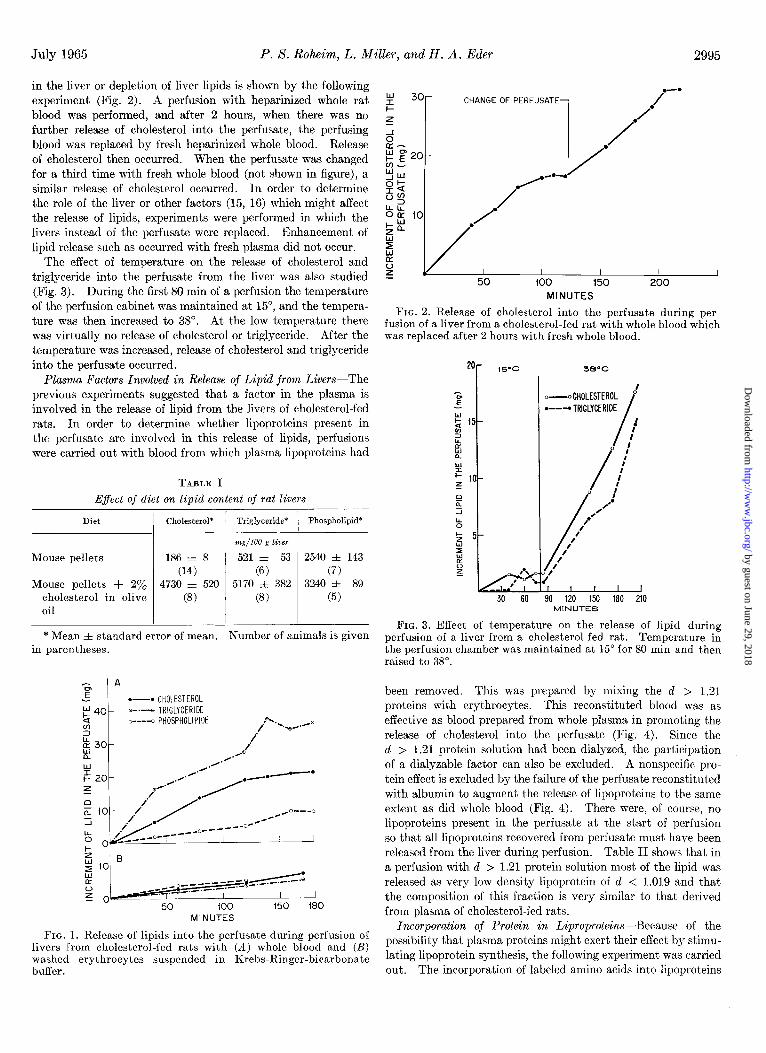
in the liver or depletion of liver lipids is shown by the following experiment (Fig. 2). A perfusion with heparinized whole rat blood was performed, and after 2 hours, when there was no further release of cholesterol into the perfusate, the perfusing blood was replaced by fresh heparinized whole blood. Release of cholesterol then occurred. When the perfusate was changed for a third time with fresh whole blood (not shown in figure), a similar release of cholesterol occurred. In order to determine the role of the liver or other factors (15, 16) which might affect the release of lipids, experiments were performed in which the livers instead of the perfusate were replaced. Enhancement of lipid release such as occurred with fresh plasma did not occur.
The effect of temperature on the release of cholesterol and triglyceride into the perfusate from the liver was also studied (Fig. 3). During the first 80 min of a perfusion the temperature of the perfusion cabinet was maintained at 15”, and the tempera- ture was then increased to 38”. At the low temperature there was virtually no release of cholesterol or triglyceride. After the temperature was increased, release of cholesterol and triglyceride into the perfusate occurred.
Plasma Factors Involved in Release of Lipid from Livers-The previous experiments suggested that a factor in the plasma is involved in the release of lipid from the livers of cholesterol-fed rats. In order to determine whether lipoproteins present in the perfusate are involved in this release af lipids, perfusions were carried out with blood from which plasma lipoproteins had
TABLE I E$ect of diet on lipid content of rat livers
Diet Cholesterol* Triglyceride*
mg/lOO g liver
Mouse pellets 186 f 8 ’ 521 f 53 (14) (6)
Mouse pellets + 2% 4730 f 520 5170 f 382 cholesterol in olive (8) (8)
- I - Phospholipid”
2540 f 143 (7)
3240 zk 89 (5)
I
* Mean f standard error of mean. Number of animals is given in parentheses.
.-.CHOLESTEAOL F 40 e-.--e TRIGLYCERIDE
4 o----o PHOSPHOLIPIDE .A.,,
/ -eti ./a
MINUTES
FIG. 1. Release of lipids into the perfusate during perfusion of livers from cholesterol-fed rats with (A) whole blood and (B) washed erythrocytes suspended in Krebs-Ringer-bicarbonate buffer.
MINUTES
FIG. 2. Release of cholesterol into the perfusate during per- fusion of a liver from a cholesterol-fed rat with whole blood which was replaced after 2 hours with fresh whole blood.
zo- 15-c 380c
l ---.TRlGLYCERlDE if 15- 2
L
30 60 90 120 150 160 210 MINUTES
FIG. 3. Effect of temperature on the release of lipid during perfusion of a liver from a cholesterol-fed rat. Temperature in the perfusion chamber was maint.ained at 15” for 80 min and then raised to 38”.
been removed. This was prepared by mixing the cl > I.21 proteins with erythrocytes. This reconstituted blood was as effective as blood prepared from whole plasma in promoting the release of cholesterol into the perfusate (Fig. 4). Since the cl > 1.21 protein solution had been dialyzed, the participation of a dialyzable factor can also be excluded. A nonspecific pro- tein effect is excluded by the failure of the perfusate reconstituted with albumin to augment the release of lipoproteins to the same extent as did whole blood (Fig. 4). There were, of course, no lipoproteins present in the perfusate at the start of perfusion so that all lipoproteins recovered from perfusate must have been released from the liver during perfusion. Table II shows that in a perfusion with cl > 1.21 protein solution most of the lipid was released as very low density lipoprotein of d < 1.019 and that the composition of this fraction is very similar to that derived from plasma of cholesterol-fed rats.
Incorporation of Protein in Liproproteins-Because of the possibility that plasma proteins might exert their effect by stimu- lating lipoprotein synthesis, the following experiment was carried out, The incorporation of labeled amino acids into lipoproteins
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

2996 Apoprotein of Plasma Lipoproteins Vol. 240, No. 7
- x---WHOLE BLOOD .---•dti.21 PROTElNStERYTHROCYTES .-.
z m-*ALBUMIN+ERYTHROCYTES -
I 40 80 120 160
MINUTES
FIG. 4. Release of cholesterol into the perfusate during per- fusions of livers of cholesterol-fed animals with various plasma proteins.
TABLE II
Release of lipids and proteins into lipoprotein fractions of perfusate
Livers from cholesterol-fed rats were perfused with blood re- constituted with erythrocytes and d > 1.21 proteins. The values represent the total amount released into various lipoproteins of the perfusate. The recovery of cholesterol in the fractions was 89%. Numbers in parentheses are the amounts present in lipo- protein fraction prepared from 10 ml of pooled plasma from choles- terol-fed rats.
Lipoprotein fraction Cholesterol
mg
d < 1.019 6.34 (7.9)
d 1.019-1.063 0.33 (2.10)
d 1.063-1.21 1.69 (2.55)
Triglyceride Phospholipid Protein
w mg WY
13.0 2.37 1.56 (11.2) (5.30) (1.60) 0.004 0.01 0.03
(0.21) (1.26) (0.74) 0.64 0.01 0.47
(0.37) (2.98) (3.44)
TABLE III
Incorporation of “C-labeled amino acids in perfusions of livers from cholesterol-fed rats
During each perfusion 25.9 PC of W-labeled Chlorella hydroly- sate (New England Nuclear Corporation) at pH 7 were added; half of this was added directly to the perfusate at the start of the per- fusion, and the rest was infused into the perfusate at a constant rate. In Experiment 374, the perfusate was washed red blood cells suspended in Ringer’s solution; in Experiment 376, it was washed red blood cells suspended in d > 1.21 protein solution.
Lipoprotein fraction 1 Specific activity / Total incorporation
Experiment 374 d < 1.019.. d 1.019-1.063. d 1.063-1.21.... d > 1.21.
Experiment 376 d < l.OlQ..... d 1.019-1.063. d 1.063-1.21.... d > 1.21.
&%?z/mg x 10-z
234 159 110 17.6
76.9 2.0 9.2 6.2 X lo3
85.4 14.3 37.4
4.1
apm x 10-a
196 1.3
14.5 7.4 x 103
TABLE IV
Incorporation of ‘YXabeled d > 1.21 proteins into plasma lipoproteins
The perfusate consisted of erythrocytes and d > 1.21 plasma protein solution which had been labeled biosynthetically.
Lipoprotein fraction
Experiment 318
Specific activity Total incorporation
hww am
d < 1.019. d 1.019-1.063. d 1.063-1.21............. d > 1.21.
Experiment 324
2530 3950 1270 40
710 340 1570 2.1 x 10”
d < 1.019. 5820 9400 d 1.019-1.063. 2460 90 d 1.063-1.21............ 1460 220 d > 1.21. .., ., 3320 2.5 X 106
was measured in perfusions with erythrocytes reconstituted with Ringer’s solution instead of plasma and in perfusions with eryth- rocytes reconstituted with the d > 1.21 protein fraction (Table III). The presence of protein in the perfusate had no significant effect on the total incorporation of amino acids into the d < 1.019 lipoprotein fraction; however, the specific activity of this fraction was markedly reduced when the proteins were added, since the newly formed lipoproteins derived from incorporation of the amino acids were probably diluted by the lipoproteins formed directly from the d > 1.21 proteins. These experiments have eliminated the possibility that the plasma proteins in some way stimulate lipoprotein synthesis. The possibility that plasma proteins are utilized in lipoprotein synthesis was investi- gated by measuring the incorporation into lipoproteins of d > 1.21 proteins which had been labeled biosynthetically. Livers were perfused with blood containing the labeled d > 1.21 pro- teins. At the end of 3 hours of perfusion, the lipoproteins were separated from the perfusate, and the specific activities and total incorporation were determined (Table IV). There was appreci- able labeling of the lipoproteins, especially the d < 1.019 lipopro- teins, while the d 1.019 to 1.063 and the d 1.063 to 1.21 fractions were labeled to a lesser extent. In both experiments the d < 1.019 lipoproteins had specific activities higher than that of the d > 1.21 proteins added to the perfusate. Total incor- poration was appreciably higher in the d < 1.019 fraction than in the other lipoprotein fractions.
It is, of course, possible that combination in vitro between lipid-free protein and lipoproteins, such as by nonspecific adsorp- tion or by a more specific process such as that described by Scanu and Hughes (17), could account for the presence of labeled protein in the lipoproteins. In addition, it is well known that preparation of lipoproteins completely free of other proteins is extremely difficult. Studies by Avigan, Eder, and Steinberg (18) have shown that, while repeated washing of lipoprotein fractions does reduce contamination of the lipoproteins, it is impossible to obtain lipoprotein fractions entirely free of con- tamination. Labeled d > 1.21 fractions were therefore incu- bated with unlabeled d < 1.21 lipoprotein fractions; the lipo- protein fractions were then separated, and their specific radioactivities were determined. These experiments were per- formed with d < 1.21 fractions obtained from rats fed a normal diet (Table V, Experiment A), and from rats fed diets supple-
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

July 1965 P. S. Roheim, L. Miller, and H. A. Eder 2997
mented with cholesterol and olive oil (Table V, Experiment B). In the latter animals the concentration of cl < 1.019 lipoproteins was markedly increased. It is evident from Table V that radioactivity appeared in the lipoprotein fractions. However, the specific activities of the lipoprotein fractions in both experi- ments were much lower than those in the experiments in which livers were perfused with labeled d > 1.21 proteins of comparable specific activity. In the perfusion experiments the specific activities of the cl < 1.019 lipoproteins were greater than those of the d > 1.21 proteins. Studies were also carried out in which homogenates of liver from cholesterol-fed rats were incubated with labeled d > 1.21 plasma proteins. Despite the presence of as much as 80 mg of cholesterol and 100 mg of triglyceride in the incubation vessel, the specific activities of the recovered lipo- proteins were less than 30% of those of the d > 1.21 proteins. The extent of labeling was not diminished by carrying out the reaction at 4”, whereas the incorporation of proteins into lipo- proteins in the liver perfusion is markedly temperature-depend- ent. It thus appears that the labeling of the lipoproteins that occurs during liver perfusion is accomplished by a process other than that which occurs in vitro.
The fact that the lipoproteins contained radioactivity after perfusion with the labeled plasma proteins suggests that protein functions as a lipoprotein precursor. Such a precursor might function by combining directly with lipid to form lipoproteins, or it could be degraded into its constituent amino acids or pep-
TABLE V
Incubation in vitro of ‘T-labeled d > 1.21 plasma proteins with unlabeled d < 1.21 lipoproteins
X-Labeled d > 1.21 protein fractions were prepared from plasma of normal rats as previously described. The d < 1.21 lipoprotein fraction was prepared from the top 2-cm fractions ob- tained after ultracentrifugation of whole serum at d 1.21 under the same conditions used to obtain the d > 1.21 fraction. Unlabeled d < 1.21 solution was incubated for 4 hours at 37” with ‘Glabeled d > 1.21 protein solution. The three lipoprotein fractions were separated by ultracentrifugation and washed as described. Ex- periment A was carried out with a d < 1.21 fraction prepared from plasma of rats on normal diets. The labeled d > 1.21 protein solution contained 1460 mg of protein with a specific activity of 1760 dpm per mg. The d < 1.21 lipoprotein fraction contained 41 mg of protein. Experiment B was carried out with d < 1.21 lipoprotein fractions prepared from cholesterol-fed rats with a plasma cholesterol concentration of 126 mg/lOO ml and plasma triglyceride content of 118 mg/lOO ml. The d > 1.21 fraction contained 957 mg of protein with a specific activity of 2470 dpm per mg. The d < 1.21 protein contained 46.1 mg of protein. In Experiment A 2.05 mg of protein were recovered in the d < 1.019 fraction; in Experiment B 7.95 mg of protein were recovered in that fraction.
Lipoprotein fraction Specific activity
Experiment A Experiment B
dm/w d < 1.019*. d 1.019-1.063*. d 1.06331.21* _. _. d > 1.21t.. _. _. _,
* Unlabeled. t ‘%-Labeled.
183 400 208 340 112 200
1760 2310
TABLE VI Incorporation of i4C-labeled d > 1.21 plasma proteins
into lipoproteins in presence of excess of unlabeled amino acids
Livers were perfused with reconstituted blood consisting of erythrocytes and labeled d > 1.21 plasma proteins. In both ex- periments 3 ml of the amino acid solution were added at the be- ginning of the perfusion and 2.2 ml were infused continuously during the duration of the perfusion. The amino acid solution contained all of the essential and 10 nonessential amino acids in concentrations ranging from 2.5 to 17.8 mM. The proportions were similar to those of the free amino acid content of normal rat plasma (19).
Lipoprotein fraction Sped&c activity Total incorporation --
d@n/?ng &‘m Experiment 332
d < 1.019. d 1.019-1.063. d 1.063-1.21.. _. d > 1.21.
Experiment 335
2,590 7,240 1,520 270
900 790 1,680 2.3 X 106
d < 1.019. 5,320 14,800 d 1.019-1.063. 1,450 440 d 1.063-1.21. _. _. _, 1,340 2,100 d > 1.21. 3,210 4.8 x 106
-
tides, which would then be preferentially incorporated into lipoproteins. To test this latter possibility, perfusions with labeled d > 1.21 proteins were carried out in the presence of an excess of unlabeled amino acids (Table VI). Both specific activity and total incorporation were similar to those found in the experiments carried out without added amino acids (Table IV). Had precursor protein been broken down to amino acids, these amino acids would have been diluted by the amino acids added, and the newly synthesized lipoproteins would have had lower specific activities. The results shown in Table VI suggest that breakdown of the protein to amino acids did not occur.
The question of resynthesis of protein from amino acids re- sulting from the d > 1.21 protein was also studied in animals treated with puromycin, which has been shown to inhibit lipo- protein synthesis by liver slices (20). The administration of puromycin to the liver donor rats markedly inhibited the incor- poration of 14C-labeled amino acids into the lipoproteins and other proteins of plasma (Table VII). In the puromycin- treated rat, 0.06% of the administered amino acids was incor- porated into plasma proteins as compared to 24y, in the un- treated animal. However, when puromycin was used in a perfusion with i4C-labeled d > 1.21 proteins (Table VIII), there was no significant decrease in specific activities of the very low density lipoprotein fraction. The other two fractions showed somewhat higher specific activities after administration of puro- mycin. The total incorporation of the labeled d > 1.21 protein into d < 1.019 lipoproteins was reduced to about 10% of the control value, contrasted to the very much greater inhibition of amino acid incorporation into that fraction. These experiments indicate that the protein precursor does not break down to con- stituent amino acids but rather is incorporated directly into lipoprotein by combination with lipid in the liver.
That the incorporation of d > 1.21 proteins can occur during perfusion of the liver of a rat fed a diet not supplemented with cholesterol or olive oil as well as during the perfusion of livers
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

2998 Apoprotein of Plasma Lipoproteins Vol. 240, 1-o. 7
from cholesterol-fed animals is shown in Table IX. The differ- ences in specific activities between the various lipoprotein frac- tions were not as great as those seen in the experiments in which livers from cholesterol-fed animals were perfused. The specific activities of the d < 1.019 lipoproteins in this experiment were less than that of the d > 1.21 proteins. However, the total incorporation of label into the d < 1.019 lipoprotein fraction was still considerably greater than that into the other lipoprotein fract,ions. It thus is evident that incorporation of d > 1.21 proteins into lipoproteins also occurs with normal livers.
SpeciJicity of Protein Incorporation into Lipoproteins--The
TABLE VII
E$ect of puromycin on lipoprotein synthesis from 14C-labeled amino
acids
Puromycin-HCl (American Cyanamid Company) was dissolved and adjusted to pH 5. The liver donor rats were given intraperi- toneal injections of 18 mg of puromycin 3, 2, and 1 hour prior to death, and 15 min before they were killed they received 9 mg of puromycin. Puromycin (40 mg) was added to the perfusate at the start of the perfusion, and a continuous infusion of 12 mg per hour was maintained during the entire perfusion. Livers were perfused with reconstituted blood consisting of erythrocytes and d > 1.21 plasma protein. During each perfusion a total of 15.6 PC of 1%.labeled Chlorella hydrolysate (Volk Radiochemical Company) at pH 7 were added. At the start of the perfusion 4.5& were added directly to the perfusate, and the rest was con- tinuously infused at a rate of 3.7 ,& per hour throughout the per- fusion.
Lipoprotein fraction Puromycin Sped? activit] i Total incorporation
dial%? -
79,500 14,900 18,900
4,590
+ 350
1,160 660 110
Experiment 339 cl < 1.019. d 1.019-1.063.. _. _. d 1.063-1.21.. cl > 1.21.
Experiment 337
260,000 6,000
12,300 8.0 x 106
d < 1.019.. d 1.019-1.063.. d 1.063-1.21.. _. _. _. d > 1.21..
150 350 990
1.9 x 104
TABLE VIII
Effect of puromycin on lipoprotein synthesis from labeled d > 1.21 proteins
Livers were perfused with reconstituted blood consisting of erythrocytes and ‘%-labeled d > 1.21 plasma protein. Puro- mycin was administered as in Table VII.
Lipoprotein fraction
Experiment 335 cl < 1.019.. cl 1.019-1.063.. d 1.063-1.21.. _. cl>1.21
Experiment 338 d < 1.019. _. d 1.019-1.063.. _. d 1.063-1.21.. _. cl > 1.21..
Puromvcin Specific activity Total incorporation
5,320 14,800 1,450 440 1,340 2,100 3,210 4.8 X 106
4,710 1,950 4,356 1,700 2,700 2,300 3,920 6.9 X 106
TABLE IX
Incorporation of ‘YZabeled d > 1.21 protein into lipoproteins during perfusion of liver from rat fed diet without
added cholesterol
The liver was perfused with reconstituted blood consisting of erythrocytes and ‘Glabeled d > 1.21 plasma protein.
Lipoprotein fraction Specific activity Total incorporation
Experiment 328 d<1.019 .._. d 1.019-1.063. d 1.063-1.21. d > 1.21.
Experiment 423 d < 1.019. d 1.019-1.063. . d 1.063-1.21. d > 1.21.
1950 5050 1350 230 1500 1100 2500 2.8 X lo6
1120 1270 N.S.* N.S.*
1000 190 2350 2.9 x 106
* Not significant.
finding of higher specific activity in the d < 1.019 lipoproteins than in the labeled d > 1.21 proteins used for perfusion suggested that in this mixture there must be a fraction with higher specific activity which could serve as precursor for the lipoproteins. Miller and Bale (21) separated plasma proteins by starch block electrophoresis and found that the a-globulin fraction has specific activities considerably higher than that of the total proteins. We have done a similar study on the d > 1.21 proteins and have shown that the specific activity in the a-globulin fraction was twice as high as that in the mixed proteins. The specific activity of this fraction of the plasma proteins could readily satisfy the requirements of a precursor-product relationship. Evidence of specificity of certain proteins for lipoprotein synthesis is provided by the following experiments. The labeled d > 1.21 plasma proteins were separated into two fractions by precipitation with 55 y0 saturated ammonium sulfate. The fractions were analyzed, and the supernatant consisted almost entirely of albumin while the precipitate contained all of the plasma globulins but also some albumin. In Table X are shown the results of perfusions with these fractions. It is apparent that the precipitate was a more active fraction than the supernatant. With the precipitate the specific activity of the newly formed d < 1.019 lipoproteins was almost as high as that of the added protein, while incorpora- tion into the other lipoprotein fractions was relatively low. With the supernatant the specific activity of the d < 1.019 lipoprotein fraction was considerably lower than that of the added protein. Furthermore, t,otal incorporation was about the same in all lipoprotein fractions. It thus appears that the active precursor of the lipoprotein is in the globulin-containing fraction, and this suggests that there is specificity in protein utilization for lipoprotein synthesis.
Studies of lipoprotein synthesis have also been carried out in intact rats. In Table XI are shown the results of experiments in which 1°Clabeled d > 1.21 proteins were injected into the tail veins of rats that had been fed the cholesterol-containing diet for a period of 5 days. These rats were killed 2 hours after injec- tion, and their plasma lipoproteins were separated. Incorpora- tion of the injected proteins into the lipoprotein fractions oc- curred. In these experiments the specific activity of the d < 1.019 lipoproteins was not consistently greater than those of the
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

July 1965 P. S. Roheim, L. Miller, an.d H. A. Eder
TABLE XII
Incorporation of ~4C-labeled “globulin” and LLalbumin” into plasma lipoproteins of intact rats
Rats were given intravenous injections of a solution of i4C- labeled protein fractions prepared by ammonium sulfate precipi- tation as in Table X. The rats had been fed the diet containing added cholesterol and olive oil for 10 to 16 days. The animals were bled 90 to 135 min after the injection.
other lipoprotein fractions, and furthermore, the specific activi- ties of the lipoprotein fractions were considerably lower than that of the d > 1.21 proteins. These findings can, in part, be explained by the fact that the plasma of the intact rats contains preformed lipoproteins and that the newly formed lipoproteins are diluted by them, whereas in the perfusion with d > 1.21 proteins no lipoproteins are present at the start of the perfusion.
Experiments with fractions prepared by ammonium sulfate precipitation were also carried out in intact rats (Table XII).
TABLE X Incorporation of “C-labeled “albumin” and “globulin”
into lipoproteins
The protein fractions were prepared from “C-labeled cl > 1.21 plasma protein solution with a specific activity of 4250 dpm per mg. Saturated ammonium sulfate was added slowly with con- tinuous stirring to a final concentration of 557, (v/v). The sus- pension was centrifuged at 1900 rpm for 90 min, and the superna- tant p-as removed and dialyzed against 0.85% NaCl containing 0.1% EDTA and then with Ringer’s solution. The precipitate was dissolved in water and dialyzed for 2 days against 0.85y0 NaCl containing 0.1% EDTA and then with Ringer’s solution. In Experiment 371 the perfusate consisted of erythrocytes and 82 ml of protein solution (800 mg). The specific activity of the protein was 2520 dpm per mg. In Experiment 372 the perfusate consisted of erythrocytes and 72 ml of protein solution (1300 mg). The specific activity of the protein was 5260 dpm per mg.
hlount ejected
Specific activity of lipoprotein
fraction
Protein fraction injected
Specific activity of injection
Lipoprotein fraction
Experiment 336. d < 1.019. d 1.019-1.063. _. d 1.063-1.21. _. d > 1.21.
Experiment 340.. d < 1.019. d 1.019-1.063. d 1.063-1.21... _. d > 1.21.
Experiment 341. d < 1.019. d 1.019-1.063.. d 1.063-1.21.. d > 1.21.
Experiment 333. d < 1.019. d 1.019-1.063.. _. d 1.06331.21.. d > 1.21.
Experiment 336. cl < 1.019. cl 1.019-1.063. d 1.063-1.21.. d > 1.21.
w 37
80
150
140
230
@mlw Supernatant
Supernatant
Supernatant
Precipitate
Precipitate
3400
2130
690
2600
3600
0 0 0
60
0 0 0
120
0 0 0
50
260 120
40 280
200 60 40
280
- Specific
activity Total incorpo
ration
dPm
230 60 390 50 600 100
1130 1.2 x 106
4400 4750 1020 160 1390 140 5080 6.3 X 10”
Protein fraction injected Lipoprotein fraction
Experiment 371. d < 1.019.. d 1.019-1.063.. d 1.063-1.21.. d > 1.21..
Experiment 372. d < 1.019. d 1.019-1.063.. d 1.063-1.21. _. d > 1.21..
Supernatant
Precipitate
There was no incorporation of supernatant proteins into the lipoproteins while there was appreciable incorporation of proteins of the precipitate. In Experiment 336, the total amount of supernatant protein injected was small in relation to the amount of the precipitate protein. However, in Experiments 340 and 341 the amounts of the supernatant fraction injected were closer to the amount, of precipitate fraction injected in Experiments 333 and 336. Nevertheless, when these larger amounts of pro- tein were injected, no incorporation of label into lipoprotein occurred, as compared to the appreciable incorporation of the proteins of the precipitate fractions. These experiments in- dicate that formation of lipoproteins from specific precursor proteins occurs in the intact animal as well as in the isolated perfused liver.
DISCUSSION
Since the first isolation of lipoproteins, attempts have been made to free the lipoproteins of lipid in order to obtain an apo- protein capable of recombining with lipid. Macheboeuf (22), using a lipoprotein prepared from horse serum, probably a high density cu-lipoprotein, showed that it was possible to extract some of the lipid but that additions of these lipids did not result in recombination into the original lipoprotein. Oncley, Gurd, and Melin (23) extracted the major portion of the lipid from
TABLE XI
Incorporation of 14C-labeled d > 1.21 plasma proteins into plasma lipoproteins of intact rats
IGLabeled d > 1.21 plasma protein solution (4 ml; 325 mg) with a specific activity of 8150 dpm per mg was injected into the tail vein of rats which had been fed the diet containing added cholesterol and olive oil for 5 days. The animals were bled 2 hours after the injection.
Specific activity Lipoprotein fraction
Experiment 362-5 d < 1.019.. d 1.019-1.063. d 1.0633121. d > 1.21..
Experiment 362-6 d < 1.019.. d 1.019~1.063. d 1.063-1.21 d > 1.21. _. _.
360 290 220
1090
380 480 190
1110
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

3000 Apoprotein of Plasma Lipoproteins Vol. 240, No. 7
@-lipoproteins and obtained a soluble protein residue, but they did not study its recombination with lipids. Avigan (24) found that treatment of low density lipoproteins with ether at 4” removed almost all of the cholesterol and a smaller portion of the phospholipid, leaving a soluble protein with its electrophoretic mobility unaltered.
More recently Scanu, Lewis, and Bumpus (25) extracted high density lipoproteins with ethanol and ether and obtained a protein residue containing only 0.6% lipid. Scanu and Hughes (17) found that when this protein was labeled and mixed with serum it interacted or exchanged with high density lipoproteins, resulting in the appearance of radioactivity in the high density lipoproteins. In the absence of high density lipoproteins the protein would interact with low density lipoproteins, chylomi- crons, or fat emulsions with the appearance of radioactivity in low or very low density particles. This reaction was independent of temperature and the presence of various enzyme inhibitors. The formation of lipoproteins by perfusion of livers with plasma proteins appears to be a very different phenomenon. This process favors the formation of very low density lipoproteins despite the presence of appreciable amounts of high density lipoproteins, and it is markedly temperature-dependent. The incubation experiments shown in Table V suggest that very low density lipoproteins do adsorb plasma proteins, but not to an extent sufficient to explain the high specific activities of the d < 1.019 lipoprotein fractions produced during perfusion of livers with labeled d > 1.21 proteins.
Our studies point to the existence of a native protein which has the capacity to combine with lipid in the liver and become a lipoprotein. The fact that the specific activities of the d < 1.019 proteins are higher than that of the mixed d > 1.21 proteins (Table IV) and the fact that the crude globulin preparation is incorporated into the lipoproteins to a far greater extent than the albumin preparation (Table X) suggest that a specific frac- tion of the plasma proteins may serve as the precursor of the lipoproteins. By analogy with other complex proteins we may consider the protein portion of the lipoprotein to be its apopro- tein. It seems likely that the plasma protein fraction which serves as precursor of the lipoproteins would contain one or more of these apoproteins.
The identification of the newly formed lipoproteins has not yet been made. The chemical data in Table II show that the composition of the d < 1.019 lipoproteins released during per- fusion is very similar to the composition of those present in the plasma of cholesterol-fed rats. A similar comparison with the other lipoprotein fractions is not meaningful because of the very small amounts of material released. Preliminary immuno- chemical studies of the precursor protein fraction have shown that it reacts with antibodies to the plasma lipoproteins of normal rats (26). However, immunochemical studies of the d < 1.019 lipoproteins released during perfusion have not been made.
In the present experiments the incorporation was far greater into the d < 1.019 lipoproteins than into the other fractions. This may be in part due to the fact that the system used (the fatty liver from cholesterol-fed rats) is one which favors the release of the very low density lipoproteins. However, even when livers from rats fed normal diets were perfused, there was greater incorporation into the very low density lipoproteins (d < 1.019). In the intact animal there was also greater incor- poration into d < 1.019 lipoproteins, but the differences were
not so striking. It must be emphasized that the incorporation of the protein into the other lipoprotein fractions occurred in all situations, but to a much lesser extent than occurred with the very low density lipoproteins.
Until a pure apoprotein is isolated and characterized, it is impossible to know whether a single apoprotein serves as pre- cursor for all lipoprotein classes or whether there are apoproteins for each lipoprotein class. Although data on the rat are not available, differences in amino acid composition in the protein portions of the various lipoproteins have been found in other species (27, 28). Such data suggest that at least two and per- haps more apoproteins might be necessary for the formation of the various types of lipoproteins. However, Shore and Shore (29) have shown that after lipolysis of human very low density lipoproteins by post-heparin plasma the proteins of this class ap- pear in lipoprotein classes of higher density, resulting in lipopro- tein classes that are heterogenous with respect to protein. Such heterogeneity of the various lipoprotein classes may well occur normally, and under these circumstances an apoprotein, while incorporated predominantly into a single lipoprotein class, could be found in other fractions. Immunochemical studies have shown considerable cross-reactivity between various low density lipoprotein classes (30, 31).
The observation that lipoproteins can be formed from plasma apoprotein provides information as to the nature of lipoprotein formation. The lack of effect of added amino acids (Table VI) and the relatively slight effect of puromycin (Table VIII) sug- gest that the protein combines directly with the lipid rather than being broken down into amino acids which are t’hen reincor- porated into the lipoproteins. It thus appears that in lipopro- tein synthesis lipid is not att,ached to amino acids during peptide formation but is attached to the protein after its synthesis has been completed.
In order to determine the mechanism of lipoprotein formation, it is necessary to know the nature of the bond between lipid and protein in the lipoproteins. Because much of the lipid of the lipoproteins exchanges rapidly between the various lipoprotein fractions (32) and because much of the lipid is readily extractable at neutral pH, it has been generally believed that the lipid is attached to the protein by relat’ively weak hydrophobic and electrostatic bonds. However, since not all of the lipid classes are readily exchangeable, e.g. cholesterol esters (I), and since not all of the lipid of the lipoproteins can be readily extracted, other types of bonds must be considered. Recently Fisher and Gurin (33) have found that small amounts of fatty acids could not be extracted from the protein moiety of the lipoproteins and have suggested that these fatty acids are linked to the protein by covalent bonds. However, the specificity of lipid composi- tion could be determined by a specific conformation of t,he apo- protein without the formation of covalent bonds. Finally, there could be several types of binding: a small part of the lipid could be joined to the protein by covalent bonds while the major por- tion is bound by hydrophobic bonds which are dependent in part for their specificity on the conformation of the protein.
The biological role of apoprotein in plasma remains to be deter- mined. Its existence in plasma suggests that apoproteins may function in the transport of lipids from the liver to the plasma, especially when the lipid content of the liver is increased. The fact that amino acid incorporation into lipoproteins is not in- creased when livers from cholesterol-fed animals are perfused (34) suggests that the liver does not have the capacity to increase
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

July 1965 P. X. Roheim, L. Miller, and H. A. Eder 3001
the rate of synthesis of proteins to meet an increased lipid load. Under these circumstances the apoprotein could be utilized for lipoprotein formation. The data in Fig. 2 indicate t.hat the amount of apoprotein present in the plasma is limited relative to the amount of lipid present in a fatty liver so that the plasma apoprot.ein might function only for a brief time, such as after a fat feeding, unless its rate of production were to be increased. Nevertheless, the plasma apoprotein could play an important role in maintaining a fairly constant rate of transfer of lipid from the liver to the plasma.
Recent studies have indicated that the apoprotein can be made in the liver (26). Therefore, what is found in the plasma may merely be a fraction of apoprot’ein which has not combined with lipid to form lipoproteins. The apoprotein could also be derived by the removal of lipids during the metabolism of plasma lipo- proteins. Thus, during fat absorption by the intestine there is synthesis of lipoprotein protein by the intestinal wall (35). It is possible that, when the lipid of these newly formed chylomicrons is taken up by tissues, the protein remains in the circulation. Since the release of lipid from intestinal mucosa is in many ways similar to that in the liver, the apoprotein might also play a role in the formation of chylomicrons.
On the basis of these studies it appears likely that the release of lipid from the liver into the plasma is a stepwise process in which lipids present in the liver combine with a completely formed protein to form the lipoproteins. The lipoprotein is then released from the liver into the plasma. The rate of release of lipid from the liver will then be limited by (a) the amount of lipid present in the liver, (b) the availability of apoprotein which may come from the plasma or directly from synthesis in the liver, (c) the rate of combination between lipid and protein, and (d) the rate of release of lipoprotein from the liver. Investigations of the mechanisms of the transport of lipid from the liver require consideration of each of these steps.
SUMMARy
Rat plasma contains an apoprotein which combines with lipid in the liver and is released into the plasma as lipoproteins. The highest rate of incorporation is into the density (d) < 1.019 lipoprotein fraction although there is some incorporation into the other lipoprotein fractions. Ammonium sulfate fractiona- tion of the plasma proteins has shown that the apoprotein is in the globulin fraction.
2. 3. 4.
5.
tj:
8.
9.
10.
11.
12. 13.
14.
15. 16. 17. 18.
LUNDSGaaRD, E., Biochim. et Biophys. Acta, 4, 322 (1950). BRAUER, R. W., Phwsiol. Revs., 43. 115 (1963). SCANU, A., AND HUG&S, W. L.; J. &al. ‘Chek.: 236,2876 (1960). AVIGAN, J., EDER, H. A., STEINBERG, D., Proc. Xoc. Exptl.
BioZ. Med., 96,429 (1957). 19.
20.
HENDERSON, L. M., SCHURR, P. E., AND ELVEHJEM, C. A., J. BioZ. Chem., 177,815 (1949).
ROBINSON, D. S., AND SEAKINS, A., Biochim. et Biophys. Acta, 62, 163 (1962).
21. MILLER, L. L., AND BALE, W. F., J. Exptl. Med., 99,125 (1954). 22. MACHEBOEUF, M. M., Bull. sac. chim. biol., 11, 485 (1929). 23. ONCLEY, J. L:, GURD; F. R. N., AND MEL&, M., J. hm. khem.
Sac., 72,458 (1950). 24. AvIG.QN, J., J. Biol. Chem., 226, 957 (1957). 25. SC.QNU, A., LEWIS, L. A., BND BUMPUS, F. M., Arch. Biochem.
Biophys., 74,390 (1958). 26.
27. 28. 29.
EDER, H. A., ROHEIM, P. S., AND SWITZER, S., Trans. Assoc. Am. Physicians, 77, 259 (1964).
SHORE, B., Arch. Biochem. Biophys., 71, 1 (1957). RODBELL. M.. Science. 127, 701 (1958). SHORE, B., AND SHORE, V., J.‘ Athkroscler. Research, 2, 104
(1962). 30. LEVINE, L., KAUFFMAN, D. L., AND BROWN, R. K., J. Exptl.
Med., 102, 105 (1955). AL.~DJEIM, F., LIEBERMAN, M., AND GOFMAN, J. W., J. Exptl.
Med., 106, 49 (1957). 31.
32. EDER, H. A., Am. J. Med., 23,269 (1957). 33. FISHER, W. R., AND GURIN, S., Science, 143, 362 (1964).
ROHEIM, P. S., AND EDER, H. A., Circulation, 24, 1101 (1961). ROHEIM, P. S., AND EDER, H. A., Physiologist, 5, 206 (1962). EDER, H. A., ROHEIM, P. S., GIDEZ, L. I., AND SWITZER, S., in
H. C. MENG (Editor), Proceedings of an International Sympo- sium on Lipid Transport, Nashville, 1963, Charles C Thomas, Publisher, Springfield, Ill., 1964, p. 202.
MILLER, L. L., BLY, C. G., WATSON, M. L., AND BALE, W. F., J. Exptl. Med., 94, 431 (1951).
GREEN, M., AND MILLER, L. L., J. Biol. Chem., 235,3202 (1960). HAVEL, R. J., EDER, H. A., AND BRAGDON, J. H., J. Clin.
Invest., 34, 1345 (1955). RADDING, C. M., AND STEINBERG, D., J. Clin. Invest., 39, 1560
(1960). LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL,
R. J., J. Biol. Chem., 193, 265 (1951). VAUGHAN, M., STEINBERG, D., AND LOGAN, J., Science, 126,
446 (1957). ABELL, L. L., LEVY, B. B., BRODIE, B. B., AND KENDALL, F. E.,
J. Biol. Chem., 195, 357 (1952). VAN HANDEL, E., CZin. Chem., 7, 249 (1961). VAN HaNDEL, E., AND ZILVERSMIT, D. B., J. Lab. Clin. Med.,
50, 152 (1957). BEVERIDGE, J. M. R., AND JOHNSON, S. E., Can. J. Research,
Sect. E, 27, 159 (1949).
REFERENCES 34. HAFT, D. E., ROHEIM, P. S., WHITE, A., AND EDER, H. A., J.
CZin. Invest., 41, 842 (1962). 1. ROHEIM, P. S., HAFT, D. E.. GIDEZ, L. I., WHITE, A., AND 35. ROHEIM, P. S., GIDEZ, L. I., AND TINDEL, S., Federation Proc.,
EDER, H. A., J. Clin. Invest., 42, 1277 (1963). 21, 291 (1962).
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from

Paul S. Roheim, Libby Miller and Howard A. EderThe Formation of Plasma Lipoproteins from Apoprotein in Plasma
1965, 240:2994-3001.J. Biol. Chem.
http://www.jbc.org/content/240/7/2994.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/240/7/2994.citation.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on June 29, 2018http://w
ww
.jbc.org/D
ownloaded from


















