The Effect of Glycerol on the Lactate Metabolism in the Perfused Rat Liver
7
Acta pharmacol. et toxicol. 1975, 36, 441-447. From the Institute of Physiology, University of Aarhus, Universitetsparken, DK-8000 Aarhus C, Denmark The Effect of Glycerol on the Lactate Metabolism in the Perfused Rat Liver BY Christian OIsen (Received November 19, 1974; Accepted December 10, 1974) Abstract: The effect of glycerol on the lactate metabolism was studied on the perfused rat liver. Addition of glycerol to the perfusion medium caused a signif- icant output of lactate and uptake of pyruvate. The lactate/pyruvate- and p- hydroxybutyratdacetoacetate ratios rose indicating an effect of glycerol on the cytoplasmic and mitochondria1 redox potentials which became more reduced. The output of lactate by the liver may be a result of the reduction in the redox level induced by the oxidation of glycerol. Key-words: Glycerol - lactate - pyruvate - p-hydroxybutyrate - acetoacetate - redox potentials - liver. The concentration of lactate in the blood has been shown to increase and the clearance of lactate produced during muscular work to decrease when the concentration of glycerol is raised in man (OLSEN & PETERSEN 1974). Elevated concentrations of lactate in the blood after injection of glycerol to the rat have previously been described (WILLIAMSON et al. 1969). A major fraction of the glycerol (LARSEN 1963) and the lactate (BERRY 1967) metabolism takes place in the liver which may be the target organ for this effect of glycerol. Lactate enters the metabolism via pyruvate: Lactate + NAD' + Pyruvate + NADH + H' Elevated concentrations of lactate in the liver must thus be the result of increased pyruvate concentration, increased NADH/NAD+ ratio or decreased intracellular pH. The initial steps in the gluconeogenesis from glycerol are:
-
Upload
christian-olsen -
Category
Documents
-
view
212 -
download
0
Transcript of The Effect of Glycerol on the Lactate Metabolism in the Perfused Rat Liver

Acta pharmacol. et toxicol. 1975, 36, 441-447.
From the Institute of Physiology, University of Aarhus, Universitetsparken, DK-8000 Aarhus C, Denmark
The Effect of Glycerol on the Lactate Metabolism in the Perfused Rat Liver
BY
Christian OIsen (Received November 19, 1974; Accepted December 10, 1974)
Abstract: The effect of glycerol on the lactate metabolism was studied on the perfused rat liver. Addition of glycerol to the perfusion medium caused a signif- icant output of lactate and uptake of pyruvate. The lactate/pyruvate- and p- hydroxybutyratdacetoacetate ratios rose indicating an effect of glycerol on the cytoplasmic and mitochondria1 redox potentials which became more reduced. The output of lactate by the liver may be a result of the reduction in the redox level induced by the oxidation of glycerol.
Key-words: Glycerol - lactate - pyruvate - p-hydroxybutyrate - acetoacetate - redox potentials - liver.
The concentration of lactate in the blood has been shown to increase and the clearance of lactate produced during muscular work to decrease when the concentration of glycerol is raised in man (OLSEN & PETERSEN 1974). Elevated concentrations of lactate in the blood after injection of glycerol to the rat have previously been described (WILLIAMSON et al. 1969).
A major fraction of the glycerol (LARSEN 1963) and the lactate (BERRY 1967) metabolism takes place in the liver which may be the target organ for this effect of glycerol.
Lactate enters the metabolism via pyruvate:
Lactate + NAD' + Pyruvate + NADH + H'
Elevated concentrations of lactate in the liver must thus be the result of increased pyruvate concentration, increased NADH/NAD+ ratio or decreased intracellular pH.
The initial steps in the gluconeogenesis from glycerol are:

442 CHRISTIAN OLSEN
Glycerol + ATP + L-3-glycerophosphate + ADP L-3-glycerophosphate + NAD' *Dihydroxyacetone phosphate + NADH.
which lead to reduction of NAD' to NADH. Moreover, as glycerol can be converted to pyruvate, it is likely that one or more of the above factors determining the lactate concentration are influenced by glycerol metabolism. The present experiments were undertaken to study the effect of glycerol on the isolated liver with regard to the factors governing the lactate metabolism. A preliminary account of this work has been presented (OLSEN 1974).
Materials and methods
Male Wistar rats fed ad libitum and weighing about 180 g were used. The perfusion technique and medium were as described by OLSEN & KRARUP (1974) except for the haematocrit and the concentration of pyruvate which were about 30 % and 0.5 mM respectively. The medium was equilibrated with a gas mixture containing 75 % Nz, 20 % 0 2 , and 5 % C q , and recirculated for the first 40 min. after the operation. From that time on the perfusion was continued without recirculation. The flow was recorded continuously and related samples of the affluent and effluent medium were taken. The first samples served as controls. Glycerol was then added to a concentration of about 10 mM in the medium. The glycerol used was redistilled (Merck 4094) and free of ethanol. 4 perfusions were performed as described above but without the addition of glycerol. The concentrations of lactate, pyruvate, 0-hydroxybutyrate and acetoace- tate were determined by an enzymatic, fluorimetric micromethod (OLSEN 1971). Oxygen saturation was determined spectrophotometrically (SIGGAARD-ANDERSEN et a/ . 1962). Blood pH was measured at 38" with a microelectrode (Radiometer, Copenhagen). The uptake or output of oxygen, lactate and pyruvate by the liver were calculated by multiplying the arterio-venous differences by the blood flow. P-values have been ob- tained from t-tests based on paired comparisons except for the ratios on which Wil- coxon's signed rank test was used (SOKAL & ROHLF 1969).
Results
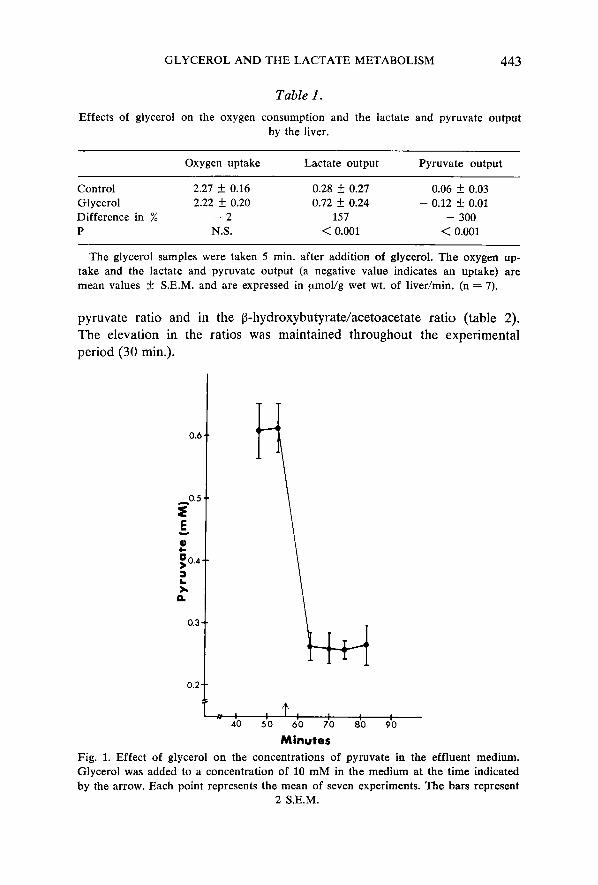
From table 1 it appears that the lactate output by the liver was insigni- ficant in the control period but increased significantly after addition of glycerol to the medium, whereas the oxygen consumption remained un- changed. The addition of glycerol reversed the small output of pyruvate to a highly significant uptake as a result of the markedly diminished con- centration of pyruvate in the effluent medium (fig. 1). The output of glucose from the liver was enhanced whereas the produc-
tion of ketone bodies was reduced (not shown). There was a slight but con- sistent increase in the pH of the effluent medium from a mean value of 7.38 to 7.41. Glycerol caused a prompt and significant rise in the lactatel
GLYCEROL AND THE LACTATE METABOLISM 443
0.6.-
0.5
5̂ E - 0 t
0.4 -- 3 h P
L
0.3 --
Table I. Effects of glycerol on the oxygen consumption and the lactate and pyruvate output
by the liver.
- -
Oxygen uptake Lactate output Pyruvate output
Control 2.27 f 0.16 0.28 f 0.27 0.06 f 0.03 Glycerol 2.22 f 0.20 0.72 f 0.24 - 0.12 f 0.01 Difference in % - 2 157 - 300 P N.S. < 0.001 < 0.001
The glycerol samples were taken 5 min. after addition of glycerol. The oxygen up- take and the lactate and pyruvate output (a negative value indicates an uptake) are mean values f S.E.M. and are expressed in pmol/g wet wt. of liver/min. (n = 7).
pyruvate ratio and in the p-hydroxybutyrate/acetoacetate ratio (table 2). The elevation in the ratios was maintained throughout the experimental period (30 min.).
0.2t
40 5 0 60 $0 sb $0 L, !
Minutes Fig. 1. Effect of glycerol on the concentrations of pyruvate in the effluent medium. Glycerol was added to a concentration of 10 mM in the medium at the time indicated by the arrow. Each point represents the mean of seven experiments. The bars represent
2 S.E.M.

444 CHRISTIAN OLSEN
Table 2. Effects of glycerol on the lactate/pyruvate and fi-hydroxybutyrate/acetoacetate ratios
in the effluent medium.
Lactatdpyruvate 0-hy droxybutyratd
acetoacetate
Control Glycerol Difference in % P
17.1 44.3 159
< 0.01
1.95 2.83
45 < 0.01
The glycerol samples were taken 5 min. after addition of glycerol. The ratios are mean values of the concentration ratios in the effluent medium from the liver. (n = 7).
From the lactate/pyruvate and the p-hydroxybutyrate/acetoacetate ratios one could calculate the NAD+-mediated redox potential in the cytoplasm and mitochondria, respectively (SCHOLZ 1968). Estimated in this way the mitochondrial compartment was the most reduced during the control period as well as after the addition of glycerol (from table 2). It appears that the lactate/pyruvate ratio increased more than the fl-hydroxybutyrate/acetoace- tate ratio, indicating that the redox potential difference between the mito- chondria and the cytoplasm was reduced. The significance of the above ef- fects of glycerol remained the same whether compared to the control per- fusion before addition of glycerol (as in tables 1 and 2) or to the perfusions without addition of glycerol (not included).
Discussion
Addition of glycerol to the medium resulted in a significant output of lactate from the liver. Among the above hypothetical factors responsible for this effect of glycerol a more reduced hepatic redox level seems the most likely as the pyruvate concentration in the effluent medium was significantly diminished, and the pH of the effluent medium was slightly increased (see below).
It has been demonstrated, that the concentrations of pyruvate in the intra- and extracellular compartments of the perfused rat liver are nearly equal (SCHIMASSEK 1963). As the concentration of pyruvate in the effluent medium was reduced, it is not likely that the enhanced lactate output was a result of an elevated concentration of intracellular pyruvate.
A rise in the lactate output may also be caused by a fall in the intracellular pH, but this explanation is unlikely, although not excluded, as the pH in the effluent medium remained unchanged or even slightly increased.

GLYCEROL AND THE LACTATE METABOLISM 445
The mitochondrial energy production is reduced during hypoxia resulting in enhanced glycolysis and lactate production in the cytoplasm. The observed rise in the f3-hydroxybutyrate/acetoacetate ratio (see table 2) indicates a more reduced mitochondrial redox level and might be due to hypoxia (e. g. SCHOLZ 1968). However since the oxygen consumption was unchanged on addition of glycerol (see table 1) and not limited by the oxygen supply, the elevated lactate output does not appear to be due to hypoxia. Moreover the lactate/pyruvate ratio rose relatively more than the p-hydroxybutyrate/ acetoacetate ratio which does not indicate hypoxia, but rather an enhanced production of reducing equivalents in the cytoplasm (OLSEN 1972).
Glycerol is a precursor of glucose as well as of pyruvate. Although the concentration of pyruvate was decreased after addition of glycerol a two- fold increase in the pyruvate + lactate pool was observed (table 1).
The oxidation of glycerol to dihydroxyacetone phosphate leads to reduc- tion of equivalent amounts of NAD+ to NADH in the hepatic cytosol. The described effects of glycerol are probably the results of this rise in the N A D W A D ' ratio. Pyruvate from the glycerol metabolism and pyruvate transported to the liver by the blood is thus trapped as lactate.
This view is supported by comparing the effects of glycerol with the re- ported effects of ethanol, as the major part of the ethanol oxidation also takes place in the liver (LUNDSGAARD 1938), causing a reduction of NAD+ to NADH in the hepatic cytoplasm (FORSANDER et al. 1965). Addition of ethanol to the medium of the perfused rat liver results in qualitatively identical effects as the above effects of glycerol on the lactate metabolism, the concentration of pyruvate, the ratios lactate/pyruvate and 0-hydroxybu- tyrate/acetoacetate (KREBS et al. 1969b) and the fall in the redox potential difference between mitochondria and cytoplasm (OLSEN 1972).
The rise in the lactate/pyruvate ratio, reflecting a rise in the cytoplasmic NADH/NAD+ ratio, seems to be due to the enhanced production of reducing equivalents from the oxidation of glycerol and ethanol, respectively. The changes in the intracellular concentrations of lactate and pyruvate cause corresponding changes in the concentrations of the effluent medium and consequently in the arteriovenous deficits.
The reoxidation of NADH takes place mainly in the respiratory chain in the mitochondria, and hence the reducing equivalents produced in the cyto- plasm must pass the mitochondrial membrane. The rise in the p-hydroxy- butyrate/acetoacetate ratio indicates an increased mitochondrial NADH/ NAD' ratio (WILLIAMSON et al. 1967). This suggests an increased supply of reducing equivalents to the mitochondrial compartment, as the oxygen con- sumption and thus the activity of the respiratory chain remained constant. Although the mitochondrial redox level became more reduced when glycerol or ethanol were oxidized, this change was far more pronounced in the cyto-

446 CHRISTIAN OLSEN
plasmic redox level by which the redox potential difference across the mito- chondrial membrane is diminished. This may facilitate the uphill transport of reducing equivalents from cytoplasm to mitochondria.
The production of carbon dioxide by the liver decreases when ethanol is oxidized (LUNDSGAARD 1938). This is probably due to a reduction in the citric acid cycle activity by the elevated mitochondrial NADWNAD' ratio (FORSANDER et al. 1965). The glycerol oxidation results in a similar rise in the mitochondrial NADWNAD' ratio, and reduced hepatic production of carbon dioxide is consequently to be expected when glycerol is metabolized. The present results do not include carbon dioxide measurements but never- theless indicate reduced carbon dioxide production after addition of glycerol to the perfusion medium, since pH in the effluent medium was constant or slightly increased, while the sum of the organic acids measured (lactate, pyruvate, p-hydroxybutyrate and acetoacetate) was increased.
In conclusion, the present experiments demonstrate a significant effect of glycerol on the hepatic lactate metabolism, presumably due to increased pro- duction of reducing equivalents. In experiments on cats (KRARUP et al. 1974) the hepatic uptake of lactate was reversed to an output by glycerol even at glycerol concentrations below saturation of the glycerol metabolizing system. The elevated lactate concentration in the blood, at rest, and the reduced clearance of lactate after exercise when elevated glycerol concentrations were induced in man (OLSEN & PETERSEN 1974) may thus - at least partly - be ascribed to the effect of glycerol on the hepatic metabolism. The analogy between the effects of glycerol and ethanol is also true in this situation, since ethanol intake exerts the same effects in man, at rest and after exercise (KREBS et al. 1969a).
A c k n o w l e d g e m e n t s
ledged. The excellent technical assistance from Lissi Hansen is gratefully acknow-
R E F E R E N C E S
Berry, M. N.: The liver and lactic acidosis. Proc. R . Soc. Med. 1967, 60, 52-54. Forsander, 0. A., N. Raiha, M. Salaspuro & M. Maenpaa: Influence of ethanol on the
liver metabolism of fed and starved rats. Biochem. J . 1965, 94, 259-265. Krarup, N., J. A. Larsen & C. Olsen: The kinetics of splanchnic elimination of ethanol
and glycerol in cats and related metabolic effects. Acta physiol. scand. 1974, 91,
Krebs, H. A., D. J. C. Cunningham, M. Stubbs & D. J. A. Jenkins: Effect of ethanol on
Krebs, H. A., R. A. Freedland, R. Hems & M. Stubbs: Inhibition of hepatic gluconeo-
Larsen, J. A.: Elimination of glycerol as a measure of the hepatic blood flow in the
1 OA-1 1 A.
postexercise lactacidemia. Israel. J . Med. Sci. 1969a, 5, 959-962.
genesis by ethanol. Biochem. J . 1969b, 112, 117-124.
cat. Acta physiol. scand. 1963, 57, 224-234.

GLYCEROL AND THE LACTATE METABOLISM 447
Lundsgaard, E.: Alcohol oxidation as a function of the liver. Compt. Rend. Trav. Lab. Carlsberg Ser. Chim. 1938, 22, 333-337.
Olsen, C.: An enzymatic fluorimetric micromethod for the determination of aceto- acetate, 8-hydroxybutyrate, pyruvate and lactate. Clin. Chem. Acta 1971, 33,
Olsen,C.: Effect of ethanol and hypoxia on the redox potential difference across the mitochondria1 membrane. Digestion 1972, 5, 224.
Olsen, C.: Lactate metabolism by the perfused rat liver after addition of glycerol. Acta physiol. scand. 1974, 91, 23A-24A.
Olsen, C. & N. Krarup: Effects of phenformin and phenformin-ethanol synergism on the perfused rat liver. Acta pharmacol. et toxicol. 1974, 34, 232-240.
Olsen, C. & E. S. Petersen: The effect of glycerol on the postexercise lactate clearance. Acta physiol. scand. 1974, 91, 76-82.
Schimassek, H.: Der Einfluss der Leber auf den extracellularen Redox-Quotienten LactaUPyruvat. Biochem. Z. 1963, 336, 468473.
Scholz, R.: Untersuchungen zur Redoxkompartmentierung bei der hamoglobinfrei per- fundierten Rattenleber. In: Stofjwechsel der isoliert perfundierten Leber. Ed.: W. Staib & R. Scholz. Springer Verlag, Berlin-Heidelberg, 1968, pp. 24-27.
Siggaard-Andersen, K., K. JBrgensen & N. Naeraa: Spectrophotometric determination of oxygen saturation in capillary blood. Scand. J . Clin. Lab. Invest. 1962, 14,
Sokal, R. R. & F. J. Rohlf: The principles and practice o f statistics in biological research. Biometry. San Francisco, W. H. Freeman and Company. 1969.
Williamson, D. H., P. Lund & H. A. Krebs: The redox state of free nicotinamide- adenine dinucleotide in the cytoplasm and mitochondria of rat liver. Biochem. J.
Williamson, D. H., D. Veloso, E. V. Ellington & H. A. Krebs: Changes in the concen- trations of hepatic metabolites on administration of dihydroxyacetone or glycerol to starved rats and their relationship to the control of ketogenesis. Biochem. J .
293-300.
298-302.
1967, 103, 514-527.
1969, 114, 575-584.