
The Effect of Dietary Nucleotide Supplementation on Growth ...
81
The Effect of Dietary Nucleotide Supplementation on Growth and Feed Efficiency of Rainbow Trout (Oncorhynchus mykiss) Fed Fish Meal-Free and Animal Protein-Free Diets By Bing Liu A Thesis presented to The University of Guelph In partial fulfilment of requirements for the degree of Master of Science in Animal Biosciences Guelph, Ontario, Canada © Bing Liu, May, 2016
Transcript of The Effect of Dietary Nucleotide Supplementation on Growth ...

The Effect of Dietary Nucleotide Supplementation on Growth and Feed Efficiency of Rainbow Trout (Oncorhynchus mykiss) Fed Fish Meal-Free
and Animal Protein-Free Diets
By
Bing Liu
A Thesis
presented to The University of Guelph
In partial fulfilment of requirements
for the degree of Master of Science
in Animal Biosciences
Guelph, Ontario, Canada
© Bing Liu, May, 2016

i
ABSTRACT THE EFFECT OF DIETARY NUCLEOTIDE SUPPLEMENTATION ON GROWTH AND
FEED EFFICIENCY OF RAINBOW TROUT (ONCORHYNCHUS MYKISS) FED FISH MEAL-FREE AND ANIMAL PROTEIN-FREE DIETS
Bing Liu Advisor: University of Guelph, 2016 D. P. Bureau Effects of dietary nucleotide supplementation on growth and feed efficiency in rainbow
trout (initial weight = 25 g/fish) were conducted in a 70-day growth trial. One control diet and
two basal diets (25%, 5% and 0% fish meal, respectively) were formulated using high levels of
highly digestible plant protein ingredients to meet all the known nutrient requirements of
rainbow trout. Two basal diets were supplemented with four increasing levels (0, 8, 16 and 24
ppm, respectively) of nucleotides using a commercial supplement (Laltide®, Lallemand Inc.
Qc, Canada). Nucleotide supplementation levels significantly (p<0.05) affected final body
weight and feed efficiency in rainbow trout fed with the 5% fish meal diet. The results of this
study suggest that dietary nucleotide supplementation may be beneficial to young rainbow
trout fed with low fish meal and high levels of plant protein ingredients. The essentiality of
nucleotides to young rainbow trout deserves to be further examined.

ii
Acknowledgement
I would first like to express my sincere heartfelt thanks to my advisor Dr. Dominique
P. Bureau, thank you for providing me this valuable opportunity to study in your lab which
has been my most precious memory in my life. Thank you for your insightful suggestion
and constant encouragement on my work during my time in your lab which guided me
towards making improvements not only as a researcher but also as a person. You have
inspired me with your brilliant ideas, passion and wealth of knowledge for your research.
Thank you to my advisory committee, Dr. Niel Karrow and Dr. Ming Fan for your
encouragement and great ideas that you have provided me at the beginning of my master
study, and thank you to Dr. Fan for providing me a great opportunity to attend to your
class.
I would like to thank Owen Skipper-Horton and Guillaume Pfeuti for their help and
advice during my experiment. I couldn’t deal with those physical work and system
problems without your help. Thank you to Christopher David Powell, for his help in data
analysis. Your code and explanation really helped me understand the practical application
of statistics. I would also like to thank Chunfang Wang and Mingfei Zhao for their tireless
help during my experiment. I would like to thank all of my lab mates, the lab is a warm and
family-like place for me. I would especially like to thank a former lab mate of mine,
Patricio Saez Albornoz, for his kind help and explanation throughout the preparation of my
experiment. Also, I would like to thank Laura Schnablegger, from learning and curriculum
support team at the university McLaughlin Library, for her commons and suggestion on my
writing.

iii
I would like to thank the contributor for the funding of this project, Lallemand Inc.
Thank you to Dr. Mathieu Castex for his valuable discussion with Dr. Bureau at the
beginning of my experiment.
Finally, I would like to thank my family, friends and all the people I love, thank you
for your supporting, understanding and encouraging in the last two years, thank you for
taking accompany with me online during my homesick and confused periods, and thank
you for all the wishes that you sent to me. I would especially thank my mom for giving me
her endless love and teaching me how to be a brave and optimistic person. All of you are
always the main props of mine.

iv
Table of Contents
CHAPTER 1- GENERAL INTRODUCTION ................................................................... 1
Objective ..................................................................................................................................... 3
CHAPTER 2 - LITERATURE REVIEW ........................................................................... 4
2.1 Nucleotides: Structure ........................................................................................................... 4 2.2 The Functions of NT ............................................................................................................... 6 2.3 Biosynthesis of purine NT ...................................................................................................... 8 2.4 Pathways of pyrimidine NT biosynthesis .............................................................................. 12 2.5 Nucleotides in Nutrition ....................................................................................................... 14 2.6 Effects of dietary NT ............................................................................................................ 18 2.6.1 The effect of dietary NT on gastrointestinal tract ................................................................... 19 2.6.2 The effect of dietary NT on growth performance ................................................................... 20 2.6.3 The effect of dietary NT on immune function ......................................................................... 21 2.6.4 The Effect of Dietary NT on Liver Function .............................................................................. 24 2.7 Evolution of Aquaculture and Aquaculture Feed Formulation .............................................. 24 2.8 Conclusion and perspectives ................................................................................................ 27
CHAPTER 3- THE EFFECT OF DIETARY NUCLEOTIDES SUPPLEMENTATION ON GROWTH AND FEED EFFICIENCY OF RAINBOW TROUT (ONCORHYNCHUS MYKISS) FED FISH MEAL-FREE AND ANIMAL PROTEIN-FREE DIETS ................................................................................................... 29 Abstract .................................................................................................................................... 29 3.2 Methods .............................................................................................................................. 32 3.2.1 Experimental Diet Formulation and Preparation .................................................................... 32 3.2.2 Fish, Feeding and Husbandry .................................................................................................. 36 3.2.3 Chemical Analysis .................................................................................................................... 37 3.2.4 Calculations and Statistical Analysis ........................................................................................ 38 3.3 Results ................................................................................................................................. 40 3.3.1 Growth Performance ............................................................................................................... 40 3.3.2 Carcass Composition ............................................................................................................... 43 3.3.3 Retained Nutrients and Retention Efficiencies ....................................................................... 44 3.4 Discussion ............................................................................................................................ 48 3.5 Conclusion ........................................................................................................................... 50
CHAPTER 4 GENERNAL DISCUSSION ....................................................................... 52
References ............................................................................................................................ 58

v
List of Tables
Table 2.1. Types of reactions in the purine biosynthesis de novo pathway ((Henderson &
Paterson, 1973) ..................................................................................................................... 10
Table 2.2 Purine and pyrimidine base content of some aquafeed ingredients (%) (Devresse,
2000) ..................................................................................................................................... 15
Table 2.3 Summary of studies examining the effects of dietary NT in fish ......................... 17
Table 3.1 Ingredient composition of the experimental diets ................................................. 33
Table 3.2 Nucleotide supplementation added level .............................................................. 34
Table 3.3 Total potentially available nucleotides (TPAN) in the Laltide® .......................... 34
Table 3.4 Proximate composition of experimental diets. ..................................................... 35
Table 3.5 Performance of rainbow trout (initial weight = 25 g/fish) in response to being fed
increasing NT content with different fish meal inclusion levels diets for 10 weeks. ........... 45
Table 3.6 Comparison of some parameters of rainbow trout in response to being fed diet 4
with Control diet ................................................................................................................... 45
Table 3.7 Proximate composition of whole carcass of rainbow trout in response to being fed
increasing NT content with different fish meal inclusion levels diets for 10 weeks,
expressed on a wet weight basis. .......................................................................................... 46
Table 3.8 Retained nitrogen (RN), recovered energy (RE), nitrogen retention efficiency
(NRE), and energy retention efficiency (ERE) of rainbow trout in response to being fed
increasing NT content with different fish meal inclusion levels diets for 10 weeks. ........... 47

vi
List of Figure
Figure 2.1. Nucleotide structure. .......................................................................................... 5
Figure 2.2. The de novo synthesis of purines (Henderson & Paterson,1973; King, 2016) ..... 9
Figure 2.3. Catabolism and the salvage pathway of purine NT (Henderson &
Paterson,1973; King, 2016) ................................................................................................. 10
Figure 2.4. The de novo pathway of pyrimidine biosynthesis (Henderson & Paterson,1973;
King, 2016) .......................................................................................................................... 13
Figure 2.4 Digestion and absorption of dietary nucleotides (Hess & Greenberg, 2012). .... 15
Figure 3.1 Live body weight (g/fish) of rainbow trout in response to being fed
experimental diets containing increasing NT levels with different fish meal inclusion levels.
............................................................................................................................................. 41
Figure 3.2 Final body weight (g/fish) of rainbow trout in response to being fed
experimental diets containing increasing NT levels with different fish meal inclusion levels.
............................................................................................................................................. 41
Figure 3.3 Thermal-‐unit growth coefficient (TGC) of rainbow trout in response to being fed
experimental diets containing increasing NT levels with different fish meal inclusion levels.
............................................................................................................................................. 42

1
CHAPTER 1- GENERAL INTRODUCTION
Nucleotides are important metabolites that are involved in almost all cellular processes and
play major roles in structural, metabolic, energetic and regulatory functions (Rudolph 1994). The
physiological functions of these compounds include encoding and deciphering genetic
information, mediating energy metabolism and cell signaling as well as serving as components of
co-enzymes, allosteric effectors and cellular agonists (Carver and Walker, 1995).
Studies have indicated that dietary nucleotide supplementation had positive effect on
various physiological functions in many species, especially during early life stages (Gil, 2002). It
has been hypothesized that during periods of rapid growth or high metabolism, some tissues such
as immune cells and gastrointestinal cells may have high demands for nucleotides. There are
three ways that an animal can obtain the NT: de novo synthesis, the salvage pathway, and
through the diet (Quan et al., 1990). The de novo synthesis and salvage pathways provide
insufficient amounts of nucleotides for the needs of these tissues, which are only partially able to
produce nucleotides or unable to produce them at all (Holen & Jonssona, 2004; Peng et al.,
2013).
Research with various animals has shown that dietary nucleotides can enhance the
proliferation of epithelial cells of the gastrointestinal tract (Sanderson & He, 1994), improve
immune responses (Carver 1994), reduce hepatic lipid accumulation (Carver 1994) and
beneficially modify intestinal microflora (Uauy 1990). Evidence also suggests that the dietary
supply of nucleotides may optimize the function of rapidly dividing tissues (Carver 1994).
Experimental evidence suggests substantial advantages of dietary nucleotides in different
aquatic species, including turbot (Low et al., 2003; Peng et al., 2013), red drum (Li et al 2007a;
Cheng et al., 2011), Coho salmon (Burrells et al., 2001a), Atlantic salmon (Burrells et al.,

2
2001b), rainbow trout (Adamek et al., 1996, Burrells et al., 2001a; Tahmasebi-Kohyani et al.,
2011; Mohebbi et al., 2013), common carp (Sakai et al., 2001), grouper (Lin et al., 2009), tilapia
(Ramadan et al., 1991; Ramadan et al., 1994), hybrid striped bass (Li et al, 2004), puffer fish
(Kiyohara et al., 1975), American lobster (Mackie, 1973), Pacific white shrimp (Li et al., 2007b)
and black tiger shrimp (Huu et al., 2012).
Feed for most aquaculture species were in the past formulated with high levels of fish
meal. However, growth of the demand for aquaculture feeds and the limited supply and high
price of fish meal has led aquaculture feed manufacturers to progressively decrease the level of
fish meal in feeds and increase their reliance on more economical plant protein ingredients.
Studies have shown that good growth and feed efficiency can be achieved with feeds based
primarily on plant ingredients. However, performance of fish generally drops significantly with
complete replacement of fish meal by plant ingredients, especially in young, fast-growing,
animals, even if all known nutrient requirements appear to have been met. Fish meal is an
excellent source of digestible essential amino acids, vitamins, minerals as well as of a wide
variety of compounds, including nucleotides (Zinn et al., 2009; Watanabe et al., 1997). In some
markets, processed animal proteins (PAPs) have filled in void left by decreasing fish meal levels
in aquaculture feeds. However, feed safety regulations and consumer concerns and demands are
preventing the use of PAPs in some markets. Most PAPs are also excellent sources of amino
acids, minerals and various compounds, including nucleotides. Conversely, plant protein
ingredients are significantly poorer sources of nucleotides. In this context, the level of
nucleotides in aquaculture feeds has likely decreased significantly and may, in part, explain the
lower performance of fish fed diets with very high levels of plant protein ingredients. It can be

3
thus hypothesized that nucleotides could potentially be beneficial essential nutrients for young
fast growing fish.
This thesis examines the effect of dietary nucleotide supplementation on growth
performance of young rainbow trout fed plant-based diets with no or only very low levels of fish
meal.
Objective
The objective of this thesis was to investigate the effects of dietary nucleotides
(nucleosides) on growth and the efficiency of feed, protein and energy utilization in rainbow
trout fed plant-based diets containing very low levels of fish meal or no fish meal.

4
CHAPTER 2 - LITERATURE REVIEW
2.1 Nucleotides: Structure
Nucleotides (NT) are low molecular weight molecules consisting of a nitrogenous base, a
pentose sugar, and one to three phosphate groups (Cosgrove, 1998) (Figure 2.1). The
nucleobases are classified as either purines including adenine (A), and guanine (G), or
pyrimidines including cytosine (C), thymine (T), and uracil (U). Pentose sugars include ribose
and deoxyribose. A purine or pyrimidine base attaches a pentose sugar to constitute a nucleoside
(NS), while a phosphate ester of NS is a NT (Carver &Walker, 1995; Li & Gatlin, 2006). NT are
the building blocks for DNA and RNA synthesis.

5
Figure 2.1. Nucleotide structure.

6
2.2 The Functions of NT
NT can either participate in reactions or synthesize other molecules. Purine and pyrimidine
NT generally have five types of functions. These functions of NT are as: 1. structure units; 2.
energy storage; 3. parts of coenzymes; 4. intermediates in some biosynthetic pathways; 5. part of
intracellular signalling molecules (Henderson & Paterson, 1973).
From the genetic perspective, the key roles of NT are the activated precursors of nucleic
acids. These precursors are connected to each other with a phosphodiester bond between the 3’
carbon of one and the 5’ carbon of the next. The pentose sugars contain five atoms of carbon,
which can be numbered from 1’ through 5’, and are attached to a nitrogenous base at the 1’
carbon position. A phosphate group is attached with the 3’ carbon of the one and 5’carbon of the
next, where the phosphodiester bond is formed. Thus, nucleic acids, are formed when NT are
joined as phosphodiester bonds. As a result, an RNA or DNA strand will have a 3’-hydroxyl end
and a 5’-phosphate or triphosphate end (Guttman, 2013).
NT, nucleoside diphosphates and triphosphates, are the main energy carriers in cells
(Guttman, 2013). Adenosine triphosphate (ATP) takes part in many endergonic metabolic
reactions of animals and plants as an energy source, and experiences change in formation,
consumption and recycling (Wischke et al, 2014). In most reactions, ATP transfers one or two
terminal phosphate group(s) or phosphoryl group(s) to some other molecule, leaving behind a
nucleoside diphosphate (ADP) or a nucleoside monophosphate (AMP) (Guttman, 2013). Some
reactions are shown below:

7
NT can also be components of co-enzymes. For example, adenosine along with cysteine
and pantothenate are the precursors of coenzyme A (CoA). Two other important coenzymes,
nicotinamide adenine dinucleotide (NAD) and flavin adenine dinucleotide (FAD), are also
synthesized by adenine NT (Cosgrove, 1998; Pesnot et al, 2011; Deluca & Kaplan, 1958).
NT derivatives are integral participants in many biochemical processes in addition to
intercellular base units (Cosgrove, 1998). For example, UDP-glucose participates in the
biosynthesis of glucose (Cosgrove, 1998). During glycogenesis, UDP-glucose is formed from
glucose-1-phosphate with the help of the enzyme UDP-glucose phosphorylase; and the UTP is
converted into PPi. Phosphoglucomutase and UDP-glucose pyrophosphorylase drive the
biosynthesis of nucleotide sugar precursors. These precursors are involved in the biosynthesis of
polysaccharides (Ji et al., 2015).
The last key role of NT is that they can act as signaling molecules. For example, in the
autocrine purinergic signaling systems, pannexin 1 channels can release ATP from cells to drive
the autocrine activation of P2 receptors (Chekeni et al., 2010; Junger, 2011; Idzko et al., 2014).

8
Also, the extracellular release of NT is associated with inflammatory disease conditions
(Eltzching et al., 2006; Faigle et al., 2008; Lazarowski, 2012).
2.3 Biosynthesis of purine NT
There are two major pathways for synthesis of purine NT: the de novo pathway and the
salvage pathway. These two pathways for NT synthesis are mostly controlled by the liver
(Carver & Walker, 1995; Cosgrove, 1998; Grimble & Westwood, 2000; Fustin et al, 2012).
Biosynthesis of purine NT by the de novo pathway occurs within the cytosol of mammalian cells
from some precursors and small molecules, such as 5-phosphoribosyl-1-pyrophosphate (PRPP),
THF derivatives, CO2, glutamine, glycine, aspartate and formate (Cosgrove, 1998; Zrenner et al.,
2006; Li & Gatlin, 2006). Purine bases and purine nucleosides participate in the salvage
pathways for the synthesis of NT. The de novo pathway is more complicated and consumes more
energy than the salvage pathway (Cosgrove, 1998; Li & Gatlin, 2006).
Synthesis of inosine monophosphate (IMP) de novo starts with PRPP. Enzymes, ATP,
glutamine, glycine, THF derivatives and aspartate are utilized during the biosynthesis of IMP.
IMP can be converted into either AMP or GMP; the conversion of IMP to AMP needs to be
catalyzed by GTP, while ATP catalyzes the conversion of IMP to GMP. Thus, IMP represents a
branch point for purine biosynthesis (Henderson & Paterson, 1973; Zrenner et al., 2006). The 10
enzymatic reactions of the purine biosynthesis de novo pathway are summarized in Figure 2.2
and Table 2.1.
In the purine biosynthesis de novo pathway, the major rate-limiting steps happen at the
activation of ribose-5-phosphate and the first step of the pathway. Ribose-5-phosphate, along
with the PRPP synthetase and ATP are required to generate the activated sugar, PRPP
(Henderson & Paterson, 1973). This activation is feedback-inhibited by purine-5’-nucleotides,

9
especially AMP and GMP (King, 2016). Also, glutamine, glycine and aspartate can be rate-
limiting factors for purine synthesis in some cells (Henderson & Paterson, 1973). Additionally,
the biosynthesis of purine de novo is controlled by the conversion of IMP to AMP and GMP;
ATP and GTP inhibit some enzymes during the biosynthesis of purines de novo (Zrenner et al.,
2006).
Figure 2.2. The de novo synthesis of purines (Henderson & Paterson,1973; King, 2016) Abbreviations in figure 2.2: PRPP: 5-phosphoribosyl-1-pyrophosphate; PRA: 5-phosphoribosylamine; PPi: pyrophosphate; GAR: glycinamide ribonucleotide; 10F-THF:10-formyl tetrahydrofolate; FGAR: formylglycinamide ribonucleotide; FGAM: formylglycinamidine ribonucleotide; AIR: 5-aminoimidazoleribonucleotide; CAIR: 4-carboxy aminoimidazole ribonucleotide; SAICAR: N-succinyl-5-aminoimidazole-4-carboxamide ribonucleotide; AICAR: 5-aminoimidazole-4-carboxamide ribonucleotide; FAICAR: 5-formaminoimidazole-4-carboxamide ribonucleotide; IMP: inosine monophosphate.

10
The catabolism and the salvage pathway of purine NT is summarized in Figure 2.3. Free
bases and nucleosides are from the degradation/catabolism of NT, and then are recycled by the
salvage reaction (Zrenner et al., 2006). Catabolism of the purine nucleotides in animal and other
higher primates leads to the final production of uric acid, which is insoluble and is excreted as
the main source of nitrogen waste (Zrenner et al., 2006).
Figure 2.3. Catabolism and the salvage pathway of purine NT (Henderson & Paterson,1973; King, 2016) Abbrevations in figure 2.3: APRT: adenine phosphoribosyl transferase; HGPRT: hypoxanthineguanine phosphoribosyl transferase; ADA: adenosine deaminase; PNP: purine nucleotide phosphorylases Table 2.1. Types of reactions in the purine biosynthesis de novo pathway ((Henderson & Paterson, 1973) Number Reaction Bond formed or
broken
Reversible ATP
required

11
1 Phosphoribosyl pyrophosphate
amidotransferase
Carbon-nitrogen No No
2 Phosphoribosyl glycineamide
synthetase
Carbon-nitrogen Yes Yes
3 Phosphoribosyl glycineamide
formyltransferase
Carbon-nitrogen No No
4 Phosphoribosyl
formylglycineamidine synthetase
Carbon-nitrogen No Yea
5 Phosphoribosyl aminoimidazole
synthetase
Carbon-nitrogen No Yes
6 Phosphoribosyl aminoimidazole
carboxylase
Carbon-nitrogen Yes No
7 Phosphoribosyl aminoimidazole
succinocarboxamide synthetase
Carbon-nitrogen Yes Yes
8 Adenylosuccinate lyase Carbon-nitrogen Yes No
9 Phosphoribosyl aminoimidazole
carboxamide formyltransferase
Carbon-nitrogen Yes No
10 IMP cyclohydrolase Carbon-nitrogen Yes No

12
2.4 Pathways of pyrimidine NT biosynthesis
The biosynthesis of pyrimidine NT can also occur either by the de novo pathway or by the
salvage pathway (Carver & Walker, 1995; Cosgrove, 1998; Grimble & Westwood, 2000; Fustin
et al, 2012). However, the synthesis of pyrimidine NT is easier than that of purine NT because
the pyrimidine base is simpler. Pyrimidine NT synthesis differs in two significant aspects from
that of purine NT. Firstly, the ring structure is not assembled on PRPP, but built as a free base.
Secondly, there is no branch in the pyrimidine synthesis pathway. The salvage pathway of
pyrimidine NT has not been included as an illustration because it is less significant than that of
purine bases due to the solubility of the byproducts of pyrimidine catabolism.
Within the de novo pathway, pyrimidine NT are synthesized from glutamine, aspartate, and
CO2 to form UMP in the cytosol and the mitochondria of mammalian cells (Li & Gatlin, 2006).
There are six steps in this pathway as diagrammed in the Figure 2.4.

13
Figure 2.4. The de novo pathway of pyrimidine biosynthesis (Henderson & Paterson,1973; King, 2016) Abbreviations in figure 2.4: CAD: CPS-II activity; CA: carbamoylaspartate; DHO: dihydroorotate; OMP: orotidine 5 -monophosphate; UMP: uridine-5-monophosphate
Carbamoyl phosphate (CP) joins the pyrimidine biosynthetic pathway with the help of
aspartate transcarbamoylase (ATCase) and aspartate to form CA; CP is a precursor for arginine
biosynthesis as well (Zrenner et al., 2006). Then, the enzyme dihydroorotase (DHOase) catalyzes
the conversion of CA to produce DHO, which is oxidized by dihydroorotate dehydrogenase
(DHODH) to form orotate. Orotate is condensed with PRPP to form OMP (Zrenner et al., 2006),
which is decarboxylated by orotidylate decarboxylase (ODCase) to generate UMP.

14
2.5 Nucleotides in Nutrition
NT are present as free NT and nucleic acids in various natural foods, for example, organ
meats, seafood and dried legumes contain purines and RNA (Clifford & Story, 1976).
Unfortunately, there are few publications describing the NT content of different foods, but any
biomass of plant, microbial or animal is generally made up of cellular material and should thus
contain nucleotides. High concentrations of NT are found in high cell density foods and
metabolically active tissues. As a result, animal based foods contain more NT than plant based
foods.
In aquaculture feed ingredients, certain animal by-products contain high NT
concentrations, such as poultry by-product (360-950 ppm) (Nates, 2013). In contrast, oils,
oilseeds and grains contain low levels of nitrogenous bases. Levels have been observed to be
well below 5 mg/g (Devresse, 2000). Purine and pyrimidine base content of some aquaculture
feed ingredients is shown in Table 2.2.

15
Table 2.2 Purine and pyrimidine base content of some aquafeed ingredients (%) (Devresse, 2000) Ingredient Adenine Guanine Cytosine Thymine Uracil Total
bases Complete fish meal 0.2 0.9 0.1 0.1 0.1 1.4 Fish solubles 0.2 2.3 0.1 0.1 0.1 2.8 Yeast 0.3 0.3 0.2 0.0 0.2 0.9 Yeast extract 0.7 0.6 0.1 0.0 0.7 2.3 Single cell protein 0.2 0.9 0.1 0.4 0.6 2.1
Figure 2.4 Digestion and absorption of dietary nucleotides (Hess & Greenberg, 2012).

16
In the diet, nucleotides are mostly consumed along with protein bound as nucleoproteins
(Carver, 1995). Digestion takes place in the small intestine as protease and nuclease enzymes
break the nucleoproteins and then nucleotides are degraded into nucleosides; Furthermore,
nucleosides can be degraded by nucleosidases to purines and pyrimidines (Figure 2.4),
nucleoside if the preferred form being absorbed into the cells of the gut (Bronk & Hastewell,
1987; Sanderson & He, 1994; Li & Gatlin, 2006; Hess & Greenberg, 2012). Rumsey et al.
(1992) reported that rainbow trout fed increasing levels of yeast nucleic acid extracts presented
significant increasing in their growth and nitrogen retention, but research on digestibility and
bioavailability of nucleic acids in natural aquaculture feed sources is limited.
In aquatic animals, dietary NT have long been implicated as feed attractants in both
vertebrate and invertebrate species (Carr and Thompson, 1983; Carr et al., 1984). For example,
early research showed that the puffer fish have chemoreceptors on their lips that can respond to
AMP, IMP, UMP and ADP through electrophysiological changes (Kiyohara et al.,1975). Kubitza
et al. (1997) reported that dietary IMP (2.8 g/kg) inclusion enhanced feed intake of largemouth
bass by 46%, compared to non-supplemented soybean meal-based diet. Also, inosine and IMP
have been found to be potent gustatory feeding stimulants and chemo-attractive substance for
turbot and American lobster (Mackie, 1973; Mackie and Adron, 1978).
Research into potential growth and health benefits of dietary NT in aquaculture species did
not begin until Burells et al. (2011a, b) reported that dietary supplementation of NT enhanced
resistance of salmonids to viral, bacterial and parasitic infections as well as improved efficacy of
vaccination and osmoregulation capacity of the animals. A summary of studies that focused on
the effect of dietary NT in a variety of fish species is provided in Table 2.3.
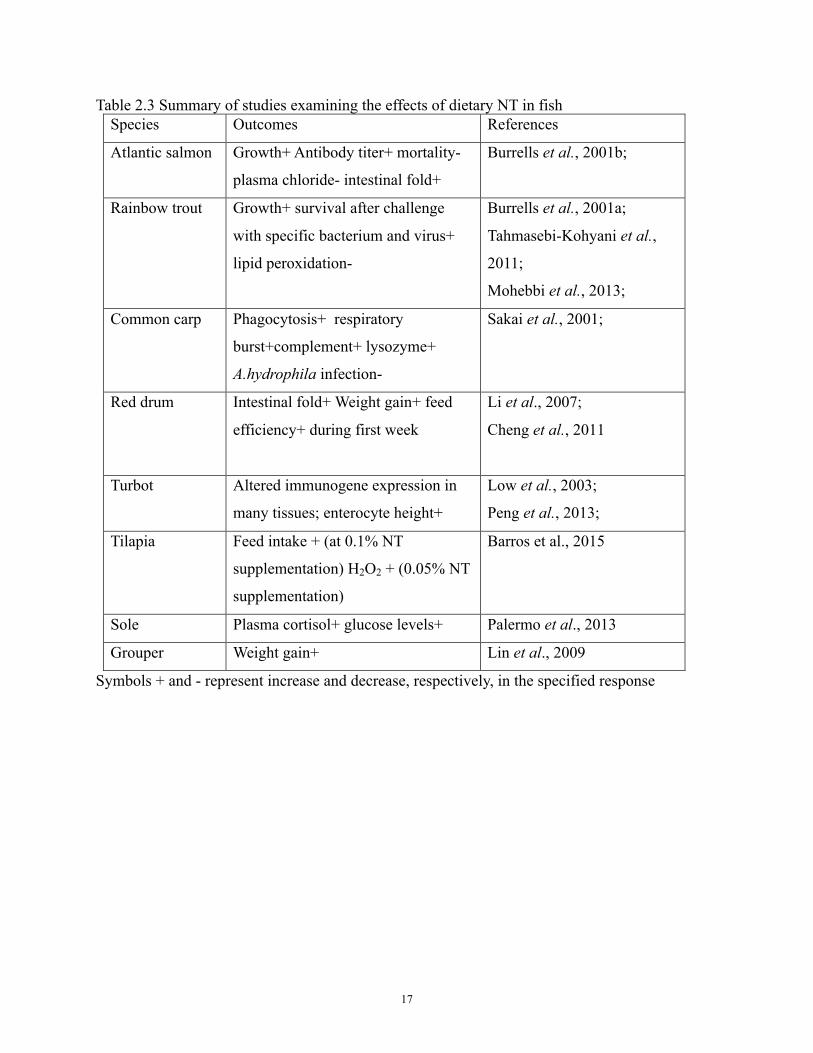
17
Table 2.3 Summary of studies examining the effects of dietary NT in fish Species Outcomes References
Atlantic salmon Growth+ Antibody titer+ mortality-
plasma chloride- intestinal fold+
Burrells et al., 2001b;
Rainbow trout Growth+ survival after challenge
with specific bacterium and virus+
lipid peroxidation-
Burrells et al., 2001a;
Tahmasebi-Kohyani et al.,
2011;
Mohebbi et al., 2013;
Common carp Phagocytosis+ respiratory
burst+complement+ lysozyme+
A.hydrophila infection-
Sakai et al., 2001;
Red drum Intestinal fold+ Weight gain+ feed
efficiency+ during first week
Li et al., 2007;
Cheng et al., 2011
Turbot Altered immunogene expression in
many tissues; enterocyte height+
Low et al., 2003;
Peng et al., 2013;
Tilapia Feed intake + (at 0.1% NT
supplementation) H2O2 + (0.05% NT
supplementation)
Barros et al., 2015
Sole Plasma cortisol+ glucose levels+ Palermo et al., 2013
Grouper Weight gain+ Lin et al., 2009
Symbols + and - represent increase and decrease, respectively, in the specified response

18
Traditionally, NT have been deemed to be non-essential nutrients because the synthesis of
NT through the salvage or the de novo pathways is adequate to meet the metabolic needs of
animals (Carver & Walker, 1995; Li and Gatlin III, 2006). The absence of NT from human and
animal diet does not cause a classic clinical deficiency syndrome. However, evidence is
mounting that endogenous supply, either de novo or salvage synthesis, may be inadequate for
normal function of certain tissues under certain conditions (Carver & Walker, 1995; Van Buren &
Rudolph, 1997). Dietary NT may be highly beneficial and may be considered semi- and/or
conditionally essential nutrients, particularly during periods of rapid cell replication (Carver &
Walker, 1995; Van Buren & Rudolph, 1997; Li and Gatlin 2006; Welker et al. 2011).
Dietary NT may help in provision of physiologically required levels of NT owing to
limited synthetic capacity of certain tissues (e.g. intestinal mucosa, haematopoietic cells,
lymphocytes and the brain), the high cost for de novo synthesis, immune-endocrine interactions
and modulation of gene expression patterns (Saikai et al. 2001; Low et al. 2003). Various studies
have suggested that dietary NT deficiency may lead damage into important tissues such as liver,
heart and intestine, and may also affect the functionality of immune cells (Grimble and
Westwood, 2000).
2.6 Effects of dietary NT
Studies indicated that growth and feed utilization of juvenile Atlantic salmon (initial body
weight: 35 g and 43 g), juvenile red drum (10 g), juvenile grouper (6 g and 10 g), post-larvae
Pacific white shrimp (1 g) and juvenile black tiger shrimp (6 g and 6 g) benefitted from dietary
NT supplementation (Burell et al. 2001b; Li et al. 2007a, b; Lin et al. 2009; Huu et al., 2012).
Experimental evidence from several studies suggest that dietary NT can exert positive effects on

19
the gastrointestinal tract, growth performance, immune response and liver function of different
species (Li & Gatlin, 2006; Sauer et al, 2011; Peng et al., 2013).
In the majority of studies, commercial NT products have been supplemented in fish
diets. These products are derived from yeast and contain other components such as
polysaccharides and trace elements (Lin et al., 2009), which are also known to provide
immunostimulation in fish (Sakai, 1999). Only a handful of studies have evaluated the effects of
purified nucleotide mixtures added to diets, such as those on red drum (Li et al., 2007a) and
Pacific white shrimp (Li et al., 2007b).
2.6.1 The effect of dietary NT on gastrointestinal tract
Dietary NT play an important role in the development of proper functions in the
gastrointestinal (GI) tract of different species. For example, the intestine of Atlantic salmon fed a
diet with a commercial NT supplement (“Optimun” 0.03% of the total feed) had a significantly
greater mean fold height of the proximal, mid and distal intestine than that of fish fed the control
diet, which may have led to a greater gut surface area (Burrells et al., 2001b). Similar results
were found in turbot (Peng et al., 2013) and red drum (Cheng et al., 2011) where researchers
observed that the intestinal morphology, including distal intestine fold height, enterocyte height
and microvillus height of fish fed a NT-supplemented diet (0.1% nucleotide premix (Sigma
Aldrich) in turbot; 0.5% nucleotide product (Canadian Bio-Systems Inc.) in red drum) was
significantly improved compared to those of fish fed control diets. In studies with rats and
piglets, dietary NT can also influence intestinal morphology including higher mitosis and lower
apoptosis rates (Sauer et al., 2011).

20
Dietary NT also has a potential role in influencing the gastro-intestinal flora. Sauer et al.,
(2010) observed that dietary NT had the ability to modulate the composition of the
gastrointestinal microbiota, after weaning in piglets exposed to stressors (Andrés-Elias et al.,
2007; Moore et al., 2011). The authors suggested that dietary NT could have the potential to be
used as alternatives to antimicrobial growth promoters in piglets.
2.6.2 The effect of dietary NT on growth performance
Dietary NT have effects on growth performance for some species; notably in the early
growth period, such as in Atlantic salmon (Burrells et al., 2001a,b), tilapia (Ramadan et al.,
1994), rainbow trout (Tahmasebi-Kohyani et al., 2011; Mohebbi et al., 2013), grouper (Lin et al.,
2009), Pacific white shrimp (Li et al., 2007b), weanling pigs (Li et al., 2015) and rats (Xu et al.,
2013).
During early growth stages, de novo synthesis might be insufficient to support rapid cell
replication, so dietary NT could reduce the high demand for NT synthesis (Li & Gatlin, 2006).
This positive effect on growth performance might also be related to the amount of feed
consumed by animals since NT may have feed attractant properties. For example, IMP
supplementation (2.8%) has increased feed intake of largemouth bass (Micropterus salmoides)
significantly (Kubitza et al., 1997). However, some experiments showed that the positive effects
of dietary NT were not related to the feed attractant properties of these compounds since similar
quantities of the diet consumed across treatments (Carver, 1994).

21
2.6.3 The effect of dietary NT on immune function
The maintenance of normal immune function may require dietary NT (Carver et al., 1990).
Studies with terrestrial animals have indicated that NT can influence phagocyte activity (Gil,
2002), interleukin-2 production (Van et al., 1985) and natural killer cell activity (Carver et al.,
1990). Also, dietary NT can also affect lymphocyte activity and immunoglobulin production
(Leonardi et al., 2003; Li & Gatlin, 2006).
Dietary NT have a positive effect on resisting infection, and are conducive to the
immunoglobulin response in early life (Gil, 2002). Enhancing resistance to infection is the
evidence that NT can affect immune function. For example, groups of mice fed NT-
supplemented diets showed less mortality after being challenged with the infection of
Staphylococcus aureus and Candida albicans compared to the groups of mice fed non-NT diets
(Kulkarni et al., 1986a,b; Carver, 1994). Also, the number of Aeromonas hydrophila was
reported to be significantly lower in liver, kidney and blood after an intraperitoneal injection
challenge in the fish fed a NT supplemented diet (Sakai et al., 2001). This may be because of the
increased phagocytic activity of murine peritoneal macrophages (Kulkarni et al., 1986a), the
enhanced T-cell dependent antibody production (Jyonouchi, 1994), the increased interleukin-2
(IL-2) production (Carver, 1994) and the intensive bone marrow cell and peripheral neutrophil
number (Matsumoto et al., 1995). The exogenous supply of a nucleoside-nucleotide mixture
might be necessary to enhance the proliferation of some host defense cells, such as bone marrow
cells and peripheral blood neutrophils, after induction of sepsis (Matsumoto et al., 1995).
Li et al. (2005) reported that dietary inclusion of 0.2% NT (Optimun) did not exert any
significant impact on growth, stress tolerance (assessed by plasma cortisol following
confinement) and in situ challenge by co-habitation with A. ocellatum in juvenile red drum. Li et

22
al. (2007a) also investigated the possible effect of purified mixtures of NT in juvenile red drum,
and a transient growth enhancing effect was found in the first week, and then it disappeared.
However, dietary supplementation of 0.5 or 1% NT (Ascogen) was effective in enhancing
immune function as assessed by superoxide anion production of head kidney macrophages and
intestine fold and microvillus height in the same fish species (Cheng et al. 2011).
Burrells et al. (2001a) observed that dietary NT (0.03%, Optimun) inclusion improved
survival rates of rainbow trout challenged with V. anguillarum and with the infectious salmon
anaemia virus. Dietary NT (Optimun) reduced plasma cortisol release, and increased number of
B lymphocytes and resistance to challenge with infectious pancreatic necrosis virus (Leonardi et
al. 2003). Recently, Tahmasebi-Kohyani et al. (2011, 2012) found that dietary 0.15-0.2% NT
supplementation promoted growth, immune function (ACH50 level, lysozyme activity and IgM
level), handling and crowding stress tolerance (plasma cortisol, glucose and ion concentration)
and resistance (survival) of fingerling rainbow trout to S. iniae.
Although dietary supplement of 0.2% NT (Aquagen) didn’t improve the growth of red-tail
black shark (Epalzeorhynchos bicolor), its inclusion enhanced the resistance (survival) to S. iniae
infection in vaccinated and unvaccinated fish (Russo et al., 2006). Similarly, 0.4% yeast RNA
inclusion in purified diets improved blood innate immune capacity (leucocyte count, respiratory
burst activity, total protein, globulin and A/G ratio) and the relative percent survival after
challenge with Aeromonas hydrophila in rohu (Labeo rohita) juveniles (Choudhury et al. 2005).
Haemato-immunological response and resistance of Catla catla juveniles to A. hydrophila also
increased when 0.8% yeast RNA was added to the purified diets (Jha et al. 2007).
Dietary NT inclusion of (0.2 or 0.5%) exerted positive impact on the innate immunity of
Pacific White Shrimp, Litopenaeus vannamei (Murthy et al. 2009). Huu et al. (2012)

23
investigated the NT supplementation (0.44% and 0.56%) of a semi-purified diet could obtain
optimal growth rate of juvenile black tiger shrimp (Penaeus monodon) at different days. They
also indicated declining requirement for NT with the increase in prawn size in this dose-titration
stud. Lin et al. (2009) also studied the NT requirement of grouper employing a purified diet
supplemented with a purified mixture of NT or single NT, and found that dietary 0.15% of
purified mixture or AMP was effective in promoting growth and immune function of the fish.
Dietary single supplementation of inosine monophosphate was also studies in olive flounder
(Paralichthys olivaceus), and it was effective in improving growth at levels of about 0.1-0.2%.
IMP inclusion can enhance innate immunity (myeloperoxidase and lysozyme activities) and
disease resistance against S. iniae at levels ranging from 0.1-0.4%.
Dietary 0.25% NT (Optimun) inclusion was reported to improve growth performance,
blood innate immune response (blood protein, albumin, albumin/globulin ratio, red blood cells,
white blood cells, lymphocyte content, alkaline phosphatase and lysozyme activity and cortisol
in serum) and stress tolerance of confinement and salinity of Caspian brown trout (Salmo trutta
caspius) (Kenari et al. 2013). In another study, 0.25-0.35% NT supplement increased the
tolerance of low water level stress in Beluga sturgeon Huso huso (Yousefi et al. 2012). However,
Welker et al. (2011) fed channel catfish with graded levels (0, 0.1%, 0.3%, 0.9% and 2.7%) of
purified nucleotide mixture for 8 weeks and observed that addition of nucleotides produced a
dose-dependent reduction in survival of to Edwardsiella ictaluri. It is hypothesized that the high
levels of NT in that study may have led to the decrease in disease resistance, which was not
related to innate immune and adaptive antibody responses.

24
2.6.4 The Effect of Dietary NT on Liver Function
A study examining the effects of dietary NT on hepatic composition and morphology
indicated that the liver weight as a percentage of the body weight was significantly higher in the
animals fed NT-supplemented diets compared to that of the animals fed basal diets (Carver,
1994). Moreover, the total serum bilirubin level and total lipid, lipid phosphorous and cholesterol
levels in the livers were lower, while the glycogen level in the livers was higher in the animals
fed NT-supplemented diets than the animals fed basal diets (Carver, 1994). A nucleotide-
nucleoside mixture diet is recommended to provide a better nutritional supplement to the
cirrhotic rats following partial hepatectomy, because the diet stimulated a hepatic RNA level and
a hepatic fraction protein synthetic rate (Usami et al., 1996).
Research on the role of dietary NT has shown in consistent results in a variety of species;
therefore, further study on these components is warranted.
2.7 Evolution of Aquaculture and Aquaculture Feed Formulation
Fish, crustacean and mollusk production increased from 3.9 percent of total by weight in
1970 to 33 percent in 2005, and about one third of fish consumed in the world is farm raised
(Gatlin III et al., 2007). The growth and the intensification of aquaculture production has
contributed to the higher aquaculture feed production. In 2015, the production of aquaculture
feed is 4% share of the global livestock feed market at 41 million tonnes, where 1.8 million
tonnes of aquaculture feed was produced in North America (Alltech Global Feed Survey, 2015).
Aquaculture feed formulations have traditionally relied on fish meal as a major protein
source. Fish meal is rich in highly digestible amino acids, essential fatty acids, vitamins, minerals

25
and nucleotides (Zinn et al., 2009; Watanabe et al., 1997). The increasing demand for this
ingredient combined with limited supply has led to a significant increase in the price of fish
meal. Aquaculture feed manufacturers have had to increasingly rely on more economical protein
sources of animal and plant origins (Wang et al., 2010; Hu et al., 2013; Zinn et al., 2009).
Studies have shown that good growth and feed efficiency can be achieved with feeds
based primarily on plant ingredients. However, performance of fish generally drops significantly
with complete replacement of fish meal by plant ingredients, especially in young, fast-growing,
animals, even if all known nutrient requirements appear to have been met. Fish meal is an
excellent source of digestible essential amino acids, vitamins, minerals as well as of a wide
variety of compounds, including nucleotides (Zinn et al., 2009; Watanabe et al., 1997). In some
markets, processed animal proteins (PAPs) have filled in the role traditionally played by fish
meal in aquaculture feeds. However, feed safety regulations and consumer concerns and
demands are preventing the use of PAPs in some markets. Most PAPs are also excellent sources
of amino acids, minerals and various compounds, including nucleotides.
Conversely, plant protein ingredients are significantly poorer sources of nucleotides
because they have low cell density compare to animal protein sources, although some plant
ingredients are high in protein content and well balanced amino acids, and have a reasonable
price and stable supply (Storebakken et al., 2000; Mente et al., 2003; Burel et al., 2000a). In this
context, the level of nucleotides in aquaculture feeds has likely decreased significantly and may,
in part, explain the lower performance of fish fed diets with very high levels of plant protein
ingredients. Thus, it is hypothesized that nucleotides are beneficial essential nutrients for young
fast growing fish.

26
From nutritional and environmental perspectives, partially or totally replacing fish meal
with plant protein is reliable and cost effective, even though the presence of some anti-nutritional
factors, such as protease inhibitors, phytates, lectins, saponins, and high fiber content in plant
protein ingredients may limit how much of these ingredients are used in feeds (Spinelli et al.,
1983; Davies et al., 1990; Krogdahl et al., 1994; Storebakken et al., 1998; Hendricks, 2002).
Several studies have examined replacing fish meal with plant protein in a multitude of species,
such as Atlantic salmon (Mente et al., 2003), rainbow trout (Oliva-Teles et al., 1994; Kaushik et
al., 1995; Refstie et al., 2000; Yurkowski et al., 1978; Burel et al., 2000b), Chinook salmon
(Higgs et al., 1982), tilapia (Davies et al., 1990), common carp (Viola et al., 1982), grass carp
(Dabrowski & Kosak, 1979; Tan et al., 2013), turbot (Regost et al., 1999; Fournier et al., 2004),
Japanese flounder (Pham et al., 2007), cobia (Chou et al., 2004), puffer fish (Zhong et al., 2011),
tiger puffer (Lim et al., 2011), African catfish (Toko et al., 2008), red sea bream (Biswas et al.,
2007), gilthead seabream (Robaina et al., 1997; Pereira & Oliva-Teles, 2003), sunshine bass
(Lewis & Kohler, 2008) and Japanese seabass (Men et al., 2014).
Most studies have led to the conclusion that it is possible to produce successful feeds
containing high levels of plant protein ingredients and minimal fish meal levels (Hua and
Bureau, 2012). However, complete replacement of fish meal and other animal ingredients has
proved very difficult, notably in diets for salmonids and other carnivorous fish species, even
when formulating to digestible essential nutrient levels in excess of all known requirements of
these animals. Growth and feed efficiency of rainbow trout fed fish meal and animal protein free
diets are generally significantly less good than that of fish fed diets with some levels of fish meal
and/or animal protein ingredients (Cho et al., 1974; Gomes et al., 1995; Adelizi et al., 1998;
Glencross et al., 2011; Burr et al., 2012). Review of evidence suggest that the lower performance

27
is unlikely due to differences in digestible amino acids, energy or essential nutrient contents of
the diets or due to the effect of anti-nutritional factors. The results of studies appear to indicate
that fish meal may have contributed some dietary components that may be beneficial to these
fish. Growth performance and feed efficiency of the fish remain nonetheless relative good and
the fish did not present any overt signs of deficiency or nutritional pathologies. This may indicate
that the dietary components contributed by fish meal may be nutrients or metabolites that the
animal is able to synthesize but unable to synthesize sufficient quantities of in order to enable
higher growth and higher efficiency of feed and nutrient utilization.
Since studies have indicated that dietary nucleotide supplementation had positive effect on
various physiological functions in many species, especially during early life stages (Gil, 2002).
Dietary NT supplementation have been observed to have positive effect on fish growth
performance, immune responses and GI tract of different fish species (Burrells et al., 2001b;
Sakai et al., 2001; Li & Gatlin, 2006; Cheng et al., 2011; Sauer et al, 2011; Tahmasebi-Kohyani
et al., 2011; Mohebbi et al., 2013; Peng et al., 2013). It has been hypothesized to be due to high
metabolic demands for nucleotides. Fish meal is rich in nucleotides while plant protein
ingredients are poor sources of these compounds. It has therefore been suggested that NT
supplementation of diet containing high levels of plant protein ingredients and low levels of fish
meal or animal protein ingredients may be an effective strategy to improve performance achieved
with such diets (Watanabe et al., 1997; Zhang et al., 2012).
2.8 Conclusion and perspectives
NT are ubiquitous intracellular compounds with important roles in cellular function and
metabolism (Cosgrove, 1998). They have been traditionally thought of as non-essential nutrients,

28
but recent research has suggested that they are semi-essential nutrients for different animal
species. An abundance of data from nutritional studies with fish species indicates that dietary NT
supplementation can influence aquatic animals’ biology, mainly in terms of growth performance,
GI tract morphology and functionality, resistance to diseases, and efficiency of nutrient
utilization.
Feeds containing high levels of fish meal probably contain sufficient quantity of NT to
meet the requirement of the animals (Huu et al., 2012). The increase in the demand and cost of
fish meal is leading feed manufacturers to reduce fish meal levels in feeds to a minimal level and
to increasingly rely on plant protein ingredients. However, feed formulated with very high levels
of plant protein ingredients and no fish meal generally perform relatively poorly. Plant protein
ingredients are less good sources of certain nutrients than fish meal and contain certain anti-
nutritional factors. Plant protein ingredients are also much less good sources of NT than fish
meal. Dietary NT supplementation has also been shown to improve growth, intestinal
morphology and functionality and immune function of fish and other animals. It would be of
interest to examine the effect of dietary NT supplementation of diets formulated to contain high
levels of plant protein ingredients and low fish meal and other animal protein ingredients.

29
CHAPTER 3- THE EFFECT OF DIETARY NUCLEOTIDES SUPPLEMENTATION ON GROWTH AND FEED EFFICIENCY OF RAINBOW TROUT (ONCORHYNCHUS MYKISS) FED FISH MEAL-FREE AND ANIMAL PROTEIN-FREE DIETS
Abstract
The effect of dietary nucleotide supplementation on growth and feed efficiency of rainbow
trout (initial weight = 25.3±0.7 g/fish) was examined in a 70-day growth trial. One control diet
was formulated to contain 25% fish meal and no supplemental nucleotides. Two basal diets were
formulated using high levels of highly digestible plant protein ingredients and to meet all the
known nutrient requirements of rainbow trout. One of the basal diet contained 5% fish meal
while the other contained 0% fish meal. These two diets were supplemented with four increasing
levels (0, 8, 16 and 24 ppm, respectively) of nucleotides (measured as total potentially available
nucleotides, TPAN) using a commercial supplement (Laltide®, Lallemand Inc. Qc, Canada).
Differences (p<0.05) in final body weight, thermal-unit growth coefficient (TGC), feed
efficiency, retained nitrogen (RN) and recovered energy (RE) were observed among diets with
different fish meal inclusion levels. Trout fed the fish meal-free diet performed less well
(p<0.05) than trout fed the fish meal-containing diets. Nucleotide supplementation levels
significantly (p<0.05) affected final body weight and feed efficiency in rainbow trout fed with
the 5% fish meal diet. The growth rate of fish fed the 5% fish meal diet with 16 ppm of dietary
supplementation of nucleotides was not different from that of the fish fed the control diet with
25% fish meal. The feed efficiency of fish fed the control diet with 25% fish meal was better
than of fish fed all the other diets. The results of this study suggest that dietary nucleotide
supplementation may be beneficial to young rainbow trout fed with low fish meal and high levels
of plant protein ingredients. The essentiality of nucleotides to young rainbow trout deserves to be
further examined.

30
3.1 Introduction
Nucleotides (NT), the phosphate esters of nucleosides (NSs), are the building blocks for
DNA and RNA, which provide energy for normal cellular bioprocesses, thus, are essential to
growth performance and development of humans and animals (Van Buren et al., 1994). An
animal can obtain NT in three ways, the de novo pathway, the salvage pathway and through the
diet. Dietary NT have been characterized as non-essential nutrients, but this position has been
over-turned by recent research, indicating that dietary NT are semi-essential nutrients. Since the
de novo synthesis pathway may be limiting under conditions like infection, rapid growth and
development, dietary NT is essential when the de novo pathway and the salvage pathway are not
able to synthesize sufficient amount of NT to meet the needs of all tissues. The results from a
number of studies suggest that dietary NT supplementation has the effects on growth, intestinal
morphology and functionality and immune function of different fish species. (Ramadan et al.,
1994; Adamek et al., 1996; Sakai et al., 2001; Burrells et al., 2001a, b; Li & Gatlin, 2006; Sauer
et al, 2011; Cheng et al., 2011; Tahmasebi-Kohyani et al., 2011; Mohebbi et al., 2013; Peng et
al., 2013).
Fish can obtain NT from their diets, notably diets that contain significant levels of fish
meal. Fish meal is a good source of nutrients, such as amino acids, minerals and NT. An
increasing demand of fish meal due to the growth of the aquaculture feed industry in recent years
has led to it becoming a very expensive commodity. The replacement of fish meal by other more
cost-effective ingredients has been the focus of much research in aquaculture nutrition. Studies
have indicated that feeds can be formulated with high levels of plant protein sources provided
attention is paid to meeting the nutritional requirements of the animals and that diets do not
contain excessive levels of anti-nutritional factors (Watanabe et al., 1997; Hua and Bureau,

31
2012).
However, feed formulated with very high levels of plant protein ingredients and no fish
meal generally perform relatively poorly. Plant protein ingredients are less good sources of
certain nutrients and other less well characterized components than are found in abundant
quantity in fish meal. Plant protein ingredients are notably much less good sources of NT than
fish meal. The performance of fish generally drops significantly with complete replacement of
fish meal by plant ingredients, especially in young, fast-growing animals, even if all known
nutrient requirements appear to have been met. In this situation, the level of NT in aquaculture
feeds has likely decreased significantly and may partly explain the lower performance of fish fed
diets with high levels of plant protein ingredients. Thus, it is hypothesized that nucleotides are
beneficial essential nutrients for young fast growing fish.
Currently, there are gaps in existing knowledge about supplementation of nucleotides in
diets of fish and the optimal dose of NT supplementation in rainbow trout. Therefore, it has been
suggested that NT supplementation of diet containing high levels of plant protein ingredients and
low levels of fish meal or animal protein ingredients may be an effective strategy to improve
performance, achieved with such diets (Watanabe et al., 1997; Zhang et al., 2012).Thus, it would
be of interest to examine the effect of dietary NT supplementation of diets formulated to contain
high levels of plant protein ingredients and low fish meal and other animal protein ingredients on
growth and efficiency of feed and nutrient utilization in rainbow trout.

32
3.2 Methods
3.2.1 Experimental Diet Formulation and Preparation
In total, nine (9) diets were prepared in this experiment (Table 3.1). All the experimental
diets were formulated to contain 38% digestible protein and 18 MJ/kg digestible energy and
exceed all the known nutritional requirements for rainbow trout recommended by NRC (2011). A
diet containing 25% fish meal, with no NT supplementation was included in the study as positive
control. Two basal diets were formulated to contain 0% and 5% of fish meal and using a
combination of several highly digestible plant protein ingredients. Eight (8) experimental diets
were produced by supplementing two basal diets with four (4) graded levels of a commercial NT
supplement (Laltide®, Lallemand Inc. Quebec, Canada) (Table 3.2) to produce four levels of NT,
(0, 8, 16 and 24 ppm of total potentially available nucleotides (TPAN) as presented as Table 3.3).
All the dry ingredients of each diet were first mixed as one batch in a Hobart mixer for 10
min (Hobart Ltd, Don Mills, ON, Canada). Fish oil and canola oil were subsequently added
slowly after mixing of dry ingredients and the resulting mash was mixed for an additional 10
min. The nine (9) resulting mashes were stored at 4°C until pelleted. Diets were steam-pelleted
using a laboratory-scale team pellet mill using a 3 mm diameter dye and a pellet length of 3 mm
(California Pellet Mill, San Francisco, CA, USA), and dried under forced air at 36°C for 24 h
using a drying oven (Precision & Scientific Co., USA). Pelleted diets were sieved to remove fine
and broken pellets, and stored at 4°C until used. The fines and broken pellets were collected and
stored at 4°C as well. Samples of the dried diets were analyzed as described in Section 3.2.3 and
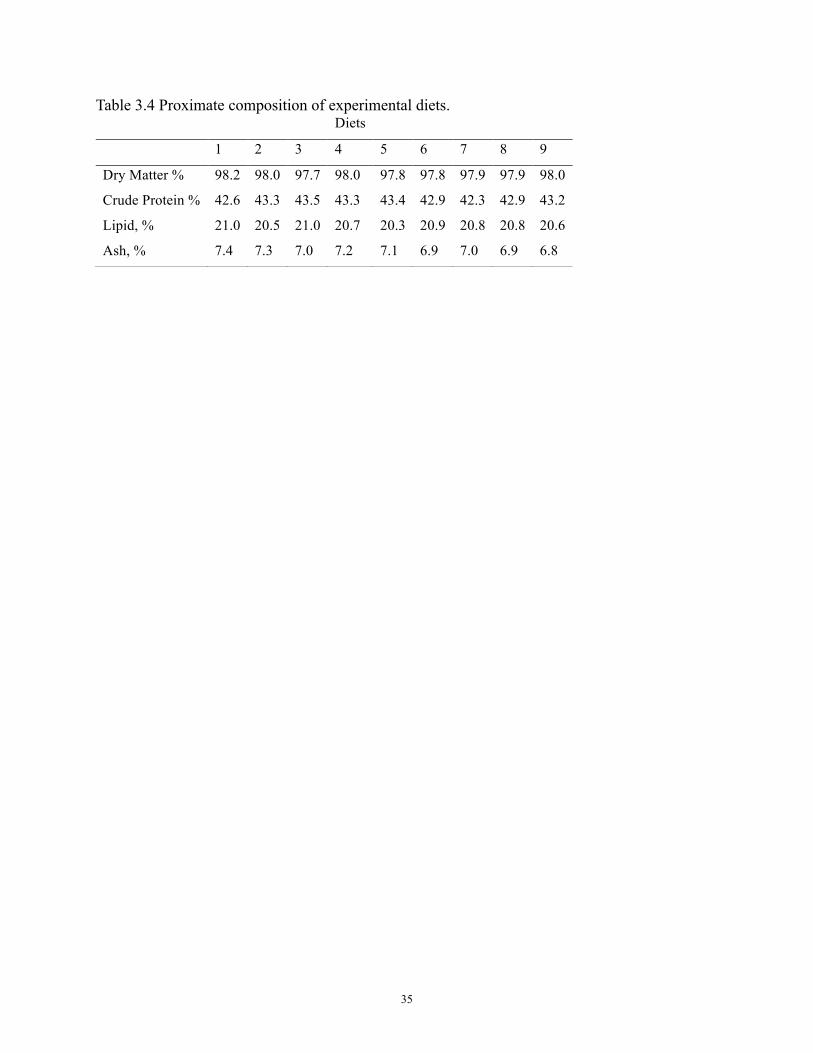
the proximate composition of the diets is reported in Table 3.4.

33
Table 3.1 Ingredient composition of the experimental diets Diets Ingredients (g/kg) 1 2 3 4 5 6 7 8 9 Fish meal, herring, 68% CP 250 50 50 50 50 0 0 0 0 Rapeseed protein conc, 62% CP 20 108 108 108 108 150 150 150 150 Rapeseed meal, 44% CP 140 140 140 140 140 145 145 145 145 Wheat gluten, 80% CP 50 50 50 50 50 50 50 50 50 High oil soy protein conc, 60% 20 150 150 150 150 160 160 160 160 Corn gluten meal, 60% CP 60 60 60 60 60 60 60 60 60 Hipro sunflower meal, 46% CP 150 150 150 150 150 150 150 150 150 Wheat middlings, 17% CP 113 68.5 68.5 68.5 68.5 56.5 56.5 56.5 56.5 Fish oil 80 90 90 90 90 90 90 90 90 Canola oil 80 80 80 80 80 80 80 80 80 Vitamin premixa 5 5 5 5 5 5 5 5 5 Vitamin E premixb 1 1 1 1 1 1 1 1 1 Bio-Lys®(51% lys) 10 20 20 20 20 20 20 20 20 DL-Met (99%) 1 1.5 1.5 1.5 1.5 2 2 2 2 L-Histidine 0 1 1 1 1 1.5 1.5 1.5 1.5 Choline chloride 60% 3 3 3 3 3 3 3 3 3 Mineral premixc 2 2 2 2 2 2 2 2 2 Dicalcium phosphate 10 15 15 15 15 15 15 15 15 Sodium chloride 2 2 2 2 2 2 2 2 2 Rovimix Stay-C 1 1 1 1 1 1 1 1 1 Carophyll® pink 1 1 1 1 1 1 1 1 1 Laltide (g/kg) 0 0 0.05 0.1 0.15 0 0.05 0.1 0.15 aProvides per kg of diet: Retinyl acetate (vit. A), 75mg; Cholescalciferol (vit. D), 60mg; dl-a-tocopherol-acetate (vit. E), 300mg; Menadione Na-bisulfate (vit. K), 1.5mg; Cyanocobalamine (vit. B12), 30mg; Ascorbic acid monophosphate, 300mg; Biotin, 210mg; choline chloride (chloride, 50%), 15mg; D-calcium pantothenate, 32.6mg; pyrodpxone-HCL (vit. B6), 7.5mg; Riboflavin (vit. B2), 9mg; Thiamin-HCL (vit. B1), 1.5mg; Caro-Pink (Astaxanthin), 500mg. bProvides per Kg of diet: Vitamin E, 0.375g; Wheat middling, 1g. cProvides per kg: sodium chloride (NaCl, 39% Na, 61% Cl), 3077mg; potassium iodide (KI, 24% K, 76%I), 10.5mg; ferrous sulphate (FeSO4-H20, 20% Fe),65mg; manganese sulphate (MnSO4, 36% Mn), 88.9mg; zinc sulphate (ZnSO4-H2O, 40% Zn), 150mg; copper sulphate (CuSO4-H20, 25% Cu), 28mg; yttrium oxide, 100mg.

34
Table 3.2 Nucleotide supplementation added level
Diet Diet Type + Nucleotide Supplementation
(expressed as ppm TPAN)
1 Control (25% fish meal)
2 Basal diet (5% fish meal) + 0 ppm
3 Basal diet (5% fish meal) + 8 ppm
4 Basal diet (5% fish meal) + 16 ppm
5 Basal diet (5% fish meal) + 24 ppm
6 Basal diet (0% fish meal) + 0 ppm
7 Basal diet (0% fish meal) + 8 ppm
8 Basal diet (0% fish meal) + 16 ppm
9 Basal diet (0% fish meal) + 24 ppm
Table 3.3 Total potentially available nucleotides (TPAN) in the Laltide®
LALTIDE® inclusion Actual TPAN added
g/kg ppm
0 0
0.05 8
0.1 16
0.15 24
35
Table 3.4 Proximate composition of experimental diets. Diets
1 2 3 4 5 6 7 8 9
Dry Matter % 98.2 98.0 97.7 98.0 97.8 97.8 97.9 97.9 98.0
Crude Protein % 42.6 43.3 43.5 43.3 43.4 42.9 42.3 42.9 43.2
Lipid, % 21.0 20.5 21.0 20.7 20.3 20.9 20.8 20.8 20.6
Ash, % 7.4 7.3 7.0 7.2 7.1 6.9 7.0 6.9 6.8

36
3.2.2 Fish, Feeding and Husbandry
Rainbow trout fingerlings were obtained from Alma Aquaculture Research Station (Elora,
Ontario, Canada) and transferred to the Fish Nutrition Research Laboratory at the University of
Guelph before the start of the experiment. The juvenile rainbow trout, average initial weight of
25.3±0.7g, were stocked into 27 fiberglass tanks (60L) with a stocking density of 12 fish per
tank. Water was supplied through a partial recirculation system (+/- 30% make up water per
pass) at a flow rate of ~3L/min per tank. Before the start of the experiment, biomass in each tank
was equalized to be within 3% of the mean. Water temperature was maintained at 15.0 °C. Fish
was kept under a 12 h light: 12 h dark photoperiod regime. The nine (9) experimental diets were
each allocated to three replicates groups (tanks) using a complete randomized design.
Fish were acclimated to the experimental conditions for two weeks prior to the start of
the experiment. During the acclimation period, fish was fed a commercial trout feed (Profishent,
Martin Mills Inc., Elmira, ON, Canada) once daily. During the 10-week-trial (70 days), fish was
hand-fed to near satiation three times daily on weekdays, 9:00, 13:00 and 16:00 and once daily
on weekends. Feed intake was recorded weekly and bulk weight of the fish in each tank were
recorded every 28 days. Experimental tanks were cleaned twice a week on each Monday and
Thursday according to the Standard Operating Procedures of the Fish Nutrition Research
Laboratory. All animals were treated in compliance with the guidelines of the Canadian Council
on Animal Care and the University of Guelph Animal Care Committee.

37
3.2.3 Chemical Analysis
At the beginning of the trial, an initial pooled sample of ten fish was taken for the
determination of the initial carcass composition. At the end of the trial, a sample of five fish per
tank was taken for the determination of the final carcass composition. Fish was humanly killed
by an overdose of tricaine methane sulfonate (MS-222) followed by a sharp blow to the head.
Live weights were recorded and 10 (initial) and/or 5 (final) fish per tank were pooled into plastic
autoclave bags and stored at -20°C until processing. Samples for carcass composition
determination were thawed overnight prior to being cooked in an autoclave (LabTech LAC-
5100S Autoclave). After autoclaving, samples were thoroughly ground into homogeneous slurry
using a food processor (Cuisinart DLC-X PLUS), freeze-dried (FTS Systems Dura Dry MP
freeze drier), and ground into a fine powder using a food processor and kept at -20 °C until
further analysis.
Another sample of three fish per tank was taken for determination of individual body
weight and length. Viscera and liver from each fish were dissected, weighed, and snap frozen in
liquid nitrogen and stored at -80°C.
Diets, ingredients, initial and final carcass samples were analysed in duplicate. Dry
matter was determined with a drying oven at 100°C for 12 h (Fisher Scientific Isotemp oven,
Markhan, ON, Canada). Ash was determined using a muffle furnace at 550°C for 4 hours (Fisher
Scientific Isotemp muffle furnace, Markham, ON, Canada), crude protein (% N × 6.25) was
determined by LECO (LECO Corp., St. Joseph, MI, USA) and lipid analysis was performed with
an Ankom XT20 fat analyser (Ankom Technology, Macedon, NY, USA) using petroleum ether.
Total carbohydrate content was determined by the following equation, total carbohydrate
content= 100 – (crude protein + lipids + ash), and gross energy (GE) was determined by the

38
following equation, GE=[(CP*23.4 kJ/g)+(Lipid*39.2 kJ/g)+(DM-Ash-CP-Lipid
kJ/g)*17.2]/100 (Cho, 1982).
3.2.4 Calculations and Statistical Analysis
The effect of dietary nucleotide level in the diet, level of fish meal and possible
interaction between the two, on final body weight, thermal-unit growth coefficients (TGC), feed
intake (FI), feed efficiency (FE), retained nitrogen (RN), recovered energy (RE), nitrogen
retention efficiency (NRE), energy retention efficiency (ERE) and proximate carcass
composition were investigated using SAS general linear model in addition to use linear and
quadratic contrasts (PROC GLM, SAS Version 9.2, SAS Institute, Cary, NC, USA). Growth
performance and proximate composition data were analyzed using a complete randomized
design with ANOVA with fish meal level and NT level as sources of variation. In all these
analyses the tank was the experimental unit and the significance level was P<0.05.
Growth performance parameters were calculated as follows:
a) Weight gain (WG, g/fish)
WG= FBW – IBW
Where: FBW= final body weight (g/fish); IBW= initial body weight (g/fish).
b) Thermal unit growth coefficient (TGC)
TGC= 100×[(FBW1/3−IBW1/3)×(sum T×D)-1]
where: FBW= final body weight (g/fish); IBW= initial body weight (g/fish); sum T×D= sum

39
Celsius degrees×days.
c) Feed efficiency (FE, gain:feed)
FE= live body weight gain/ feed intake (DM basis)
Where: live body weight gain= (FBW/final number of fish)−(IBW/initial number of fish);
feed intake= total dry feed/number of fish.
d) Retained nitrogen (RN, g/fish)
RN= (FBW×N contentfinal)−(IBW×N contentinitial)
Where: N contentfinal= nitrogen content (%) of the final carcass sample;
N contentinitial= nitrogen content (%) of the initial carcass sample
e) Recovered energy (RE, kJ/fish)
RE= (FBW×GE contentfinal)−(IBW×GE contentinitial)
Where: GEfinal= gross energy (kJ/g) content of the final carcass sample;
GEinitial= gross energy (kJ/g) content of the initial carcass sample
f) Nitrogen retention efficiency (NRE) and energy retention efficiency (ERE)
NRE (% IN)= [[(FBW×N contentfinal)−(IBW×N contentinitial)]/IN]×100
ERE (% IE)= [[(FBW×GE contentfinal)−(IBW×GE contentinitial)]/IE]×100
where: N contentfinal= nitrogen content (%) of the final carcass sample; N contentinitial= nitrogen
content (%) of the initial carcass sample; GEfinal= gross energy (kJ/g) content of the final carcass
sample; GEinitial= gross energy (kJ/g) content of the initial carcass sample; IN=ingested nitrogen
(g/fish); IE= ingested energy (kJ/fish).

40
g) Viscerosomatic index (VSI, %)
VSI =100×Individual viscera weight/Individual FBW
Where: Individual viscera weight = weight of viscera (g) at the end of experimental period
h) Hepatosomatic index (HIS, %)
HIS =100×Individual liver weight /Individual FBW
Where: Individual liver weight = weight of liver at the end of experimental period.
3.3 Results
3.3.1 Growth Performance
During the 70-day trial, no mortality was observed. Figure 3.1 presents the live body
weight of the fish fed the nine experimental diets over the 70-day trial. The results of final body
weight, TGC and feed efficiency, of fish fed diets containing increasing levels of NT with
different levels of fish meal inclusion, are presented in Figure 3.2, 3.3 and 3.4, respectively.

41
Figure 3.1 Live body weight (g/fish) of rainbow trout in response to being fed experimental diets containing increasing NT levels with different fish meal inclusion levels.
Figure 3.2 Final body weight (g/fish) of rainbow trout in response to being fed experimental diets containing increasing NT levels with different fish meal inclusion levels.
1 Dietary NT supplementation level.
0.0
20.0
40.0
60.0
80.0
100.0
120.0
140.0
160.0
180.0
200.0
Diet 1 Diet 2 Diet 3 Diet 4 Diet 5 Diet 6 Diet 7 Diet 8 Diet 9Day 0 Day 28 Day 56 Day 70
Live
body weight (g/fish)
1

42
Figure 3.3 Thermal-unit growth coefficient (TGC) of rainbow trout in response to being fed experimental diets containing increasing NT levels with different fish meal inclusion levels.
1 Dietary NT supplementation level. Figure 3.4 Feed efficiency of rainbow trout in response to being fed experimental diets containing increasing NT levels with different fish meal inclusion levels.
1 Dietary NT supplementation level. Lines regressed using a 2nd order polynomial function.
1
1

43
The growth performance parameters of rainbow trout fed different diets are presented in
Table 3.5. Increases in final body weight (p<0.001), TGC (p<0.001) and feed efficiency (p<0.01)
of rainbow trout were found in response to increasing fish meal level in diets, but no significant
effect on VSI and HIS score occurred when comparing different fish meal levels. However,
dietary NT supplementation level affected feed intake (p<0.05) and final body weight (p<0.05)
significantly.
NT supplementation has shown effects on feed intake (p<0.05) and final body weight
(p<0.05) of rainbow trout were fed with 5% fish meal diets in significant linear responses, no
such difference was found in fish fed with 0% fish meal diets (Figure 3.2; Figure 3.3). In Table
3.6, feed intake, final body weight and TGC of rainbow trout showed no statistical difference
between fish fed with Diet 4 and fish with control diet. However, feed efficiency of fish fed Diet
4 was significantly lower (p<0.01) than that of fish fed control diet.
Only a significant quadratic increase in response to increasing dietary NT levels in VSI
(p<0.05) was observed in rainbow trout fed with 0% fish meal diets. Nonetheless, no statistical
interaction effect between fish meal levels and NT levels on growth performance was detected in
this 70-day trial.
3.3.2 Carcass Composition
The results from proximate analysis of pooled carcass samples are presented in Table 3.7.
Fish meal and NT supplementation did not significantly influence whole body composition with
the exception of ash content of fish fed with 0% fish meal which increased in a significantly
decreased (p<0.05) in a linear fashion. However, significant interactions were observed between

44
fish meal level and NT level on moisture (p<0.05), lipid (p<0.01), ash (p<0.05) and gross energy
(p<0.05) which indicates influence of NT supplementation on whole body composition as a
function of fish meal level.
3.3.3 Retained Nutrients and Retention Efficiencies
Results for retained nitrogen (RN), recovered energy (RE), nitrogen retention efficiency
(NRE) and energy retention efficiency (ERE) are presented in Table 3.8. Fish meal level had a
significant positive effect on RN (p<0.05) and RE (p<0.05), but no effect on NRE and ERE. NT
supplementation did not significantly affect RN, RE, NRE and ERE. However, NRE of fish fed
0% fish meal inclusion diets responded to NT supplementation significantly (p<0.05) in a
quadratic fashion. A significant interaction response on ERE (p<0.05) was noted between fish
meal levels and NT supplementation.

45
Table 3.5 Performance of rainbow trout (initial weight = 25 g/fish) in response to being fed increasing NT content with different fish meal inclusion levels diets for 10 weeks.
Inclusion level of
NT
Feed Intake
Final Weight
TGC1 Feed efficiency2
gain:FI
VSI3 HIS4
Ppm g/fish g/fish % gain:feed % %
Control5 - 147 194 0.274 1.17 13.6 1.11 5 % FM Diet 2 - 133 163 0.247 1.04 15.4 1.20 Diet 3 8 137 171 0.257 1.09 15.0 1.15 Diet 4 16 152 187 0.270 1.09 16.0 1.18 Diet 5 24 142 176 0.259 1.06 15.6 1.16 S.E.M 2.8 3.4 0.003 0.01 0.2 0.03 Significance6 Linear * * N.S N.S N.S N.S Quadratic N.S N.S N.S * N.S N.S 0 % FM Diet 6 - 133 162 0.246 1.03 16.5 1.22 Diet 7 8 122 151 0.233 1.04 15.0 1.09 Diet 8 16 134 165 0.249 1.04 15.5 1.24 Diet 9 24 127 156 0.236 1.02 16.7 1.26 S.E.M 2.4 2.6 0.003 0.01 0.371 0.03 Significance Linear N.S N.S N.S N.S N.S N.S Quadratic N.S N.S N.S N.S * N.S Among FM Levels *** ** *** ** N.S N.S Among NT Levels * * N.S N.S N.S N.S FM Levels*NT Levels
N.S N.S N.S N.S N.S N.S
Control vs. Average7
* *** *** *** ** N.S
1TGC=thermal unit growth coefficient, %TGC= 100 x (FBW1/3-IBW1/3)/Σ(Temp (◦C) x Number of days). 2Feed efficiency= (final body weight - initial body weight) : feed intake as dry matter basis. 3Viscerosmatic index = 100 x viscera weight (g)/ whole body weight (g). 4Hepatosomatic index=100 x liver weight (g)/ whole body weight (g). 5Containing 25% fish meal 6Significance indicated by *, **, *** at the p<0.05, 0.01 and 0.001 level; N.S = not statistically significant (p≤0.05). 7Average of Diet 2, 3, 4, 5, 6, 7, 8 and 9. Table 3.6 Comparison of some parameters1 of rainbow trout in response to being fed diet 4 with Control diet
Feed intake g/fish
Final body weight g/fish
TGC %
Feed efficiency %
Diet 4 vs. Control diet N.S N.S N.S 0.007 1parameters with significant effects after comparison, on the contrary, other parameters are not significantly affected by the NT supplementation. Significance indicated by *at the p<0.05level; N.S = not statistically significant (p≤0.05).

46
Table 3.7 Proximate composition of whole carcass of rainbow trout in response to being fed increasing NT content with different fish meal inclusion levels diets for 10 weeks, expressed on a wet weight basis.
Inclusion level of NT (ppm)
Moisture %
Crude Protein
%
Lipid %
Ash %
GE1 kJ/g
Control2 - 66.8 15.2 14.9 2.4 9.4 5 % FM Diet 2 - 66.7 15.3 15.0 2.5 9.3 Diet 3 8 65.3 15.5 16.2 2.7 9.9 Diet 4 16 67.6 14.7 15.2 2.2 9.1 Diet 5 24 66.2 15.8 15.0 2.5 9.6 S.E.M 0.4 0.2 0.2 0.1 0.1 Significance3 Linear N.S N.S N.S N.S N.S Quadratic N.S N.S N.S N.S N.S 0 % FM Diet 6 - 65.4 15.0 16.2 2.6 9.9 Diet 7 8 66.0 16.0 14.7 2.6 9.6 Diet 8 16 65.3 15.6 15.7 2.6 9.9 Diet 9 24 66.9 15.3 14.9 2.2 9.5 S.E.M 0.2 0.2 0.2 0.1 0.1 Significance Linear N.S N.S N.S * N.S Quadratic N.S N.S N.S N.S N.S Among FM Levels N.S N.S N.S N.S N.S Among NT Levels N.S N.S N.S N.S N.S FM Levels*NT Levels
* N.S ** * *
Control vs. Average4
N.S N.S N.S N.S N.S
1 Gross energy (kJ/g)
2 Containing 25% fish meal 3Significance indicated by *, **, *** at the p<0.05, 0.01 and 0.001 level; N.S = not statistically significant (p≤0.05). 4Average of Diet 2, 3, 4, 5, 6, 7, 8 and 9.

47
Table 3.8 Retained nitrogen (RN), recovered energy (RE), nitrogen retention efficiency (NRE), and energy retention efficiency (ERE) of rainbow trout in response to being fed increasing NT content with different fish meal inclusion levels diets for 10 weeks.
Inclusion level of NT
(ppm)
RN g/fish
RE kJ/fish
NRE %IN
ERE %IE
Control1 - 4.0 1645 40.3 49.0 5 % FM Diet 2 - 3.3 1344 36.0 44.2 Diet 3 8 3.6 1512 37.5 48.2 Diet 4 16 3.7 1528 35.3 43.9 Diet 5 24 3.8 1506 38.3 46.8 S.E.M 0.1 39 0.5 0.8 Significance2 Linear N.S N.S N.S N.S Quadratic N.S N.S N.S N.S 0 % FM Diet 6 - 3.2 1431 35.2 46.9 Diet 7 8 3.2 1270 38.9 45.8 Diet 8 16 3.5 1452 37.6 47.4 Diet 9 24 3.1 1286 35.3 44.2 S.E.M 0.1 32 0.7 0.5 Significance Linear N.S N.S N.S N.S Quadratic N.S N.S * N.S Among FM Levels * * N.S N.S Among NT Levels N.S N.S N.S N.S FM Levels*NT Levels N.S N.S N.S * Control vs. Average3 ** ** * *
1 Containing 25% fish meal
2Significance indicated by *, **, *** at the p<0.05, 0.01 and 0.001 level; N.S = not statistically significant (p≤0.05). 3Average of Diet 2, 3, 4, 5, 6, 7, 8 and 9.

48
3.4 Discussion
In this study, a series of experimental diets were formulated to contain essential nutrients
in excess of all the known nutritional requirements for rainbow trout according to NRC (2011)
recommendations. One control diet contained 25% fishmeal was formulated. The other two basal
diets were formulated with two fish meal levels (5% fish meal and 0% fish meal) and each of
them had been supplemented with four increasing levels (0, 8, 16 and 24 ppm, respectively) of
NT using a commercial supplement (Laltide®, Lallemand Inc. Qc, Canada).
In the present study, the level of fish meal in diets decreased from 25% down to 5% and
0%, which negatively affected the growth and feed efficiency of rainbow trout. Plant protein
ingredients in this study have been previously examined and shown to be sources of highly
digestible essential amino acids, contain low levels of anti-nutritional factors and be of high
nutritive value to rainbow trout (unpublished results). Previous results from related studies on the
same ingredients are suggesting that the differences in performance achieved with the diets with
different fish meal and plant protein ingredients levels were not likely due to different in
digestible amino acids, energy or essential nutrient contents of the diets or to the effect of anti-
nutritional factors. Similar results were observed in another fish meal replacement trial, rainbow
trout fed diets containing low (less than 20%) fish meal source exhibited decreased growth
(Gomes et al., 1995; Adelizi et al., 1998; Glencross et al., 2011), although the nutrient contents
in the experimental diets appeared to be similar and meet all known requirements of rainbow
trout (NRC, 2011). However, complete replacement of fish meal and other animal ingredients
has proved to be very difficult in salmonid diets even when formulating to digestible essential
nutrient levels in excess of all known requirements of these animals. For example, Burr et al.

49
(2012) observed that rainbow trout which received 0% fish meal had significantly lower weight
gain, lower TGC and lower feed efficiency than fish which received higher fish meal levels.
In this study, the feed intake in rainbow trout fed with diet 4 was not significantly different
from that of fish fed with control diet. It could be hypothesized that dietary NT may have a
potential effect on palatability of diet since some NTs have been found to have significant
attractant properties for aquatic animals. For example, inosine and AMP were found to be potent
chemo-attractants for lobster (Mackie, 1973) and dietary IMP supplementation (2.8%)
significantly increased feed intake of largemouth bass (Micropterus salmoides) fed soybean-
based diet (Kubitza et al., 1997). However, the fact that NT supplementation was only effective
for feed containing 5% fish meal suggest that enhancement of performance could be related to
different mechanisms.
NT supplementation level significantly affected feed intake, TGC and feed efficiency of
rainbow trout but only for the basal diet containing 5% fish meal. Positive effect of NT
supplementation of growth and efficiency of feed utilization in several other studies with
different species, such as weanling pigs (Andres-Elias et al., 2007; Li et al., 2015), rats (Xu et
al., 2013), Atlantic salmon (Burrells et al., 2001b), rainbow trout (Tahmasebi-Kohyani et al.,
2011; Zhang et al., 2012), juvenile turbot (Peng et al., 2013), Hybrid tilapia (Ramadan et al.,
1991) and black tiger shrimp (Huu et al., 2012).
In the present study, no positive effect of NT supplementation was observed in fish fed
with 0% fish meal diets. This may suggest that some nutrients may have prevented the fish from
responding to NT supplementation. Theoretically, these different responses of fish should have
resulted in an interaction response between fish meal level and NT supplementation level.
However, no statistical interaction response in growth parameter was detected in this study. This

50
is thought to be the variability of feed intake, final body weight and feed efficiency among tanks
fed the same diet was relatively large. Thus, future studies should be designed to be more
statistically powerful by using larger number of replicates per experimental diet, exploring more
NT inclusion levels to better determine whether NT supplementation is truly beneficial or
whether the results are a simple experimental artifact.
In current study, the basal diets were designed to contain a low percentage of fish meal.
Moreover, the levels of dietary NT supplementation in this study were lower than the other
studies referred to above. More work needs to be done with greater range and types of NT
supplements and more replicates in order to assess whether NT supplementation truly exert a
positive effect on salmonid fish.
3.5 Conclusion
The objective of this study was to evaluate the effect of supplemental dietary NT,
provided through a commercial feed additive, on growth, feed efficiency and efficiency of
protein and energy utilization in rainbow trout fed diets formulated to contain low levels of NT.
The combination of high quality plant protein ingredients used in this study appeared to result in
fairly highly nutritive and palatable diets. The results suggest that dietary NT supplementation
may have a beneficial effect on fish fed low fish meal diets. A significant reduction in
performance with reducing fish meal level in this study also indicated that gaps still exist in our
understanding of nutritional requirements of salmonid fish.
Future studies could put more efforts to on examining the effect of NT and other dietary
components on digestion, cell, tissue or organ integrity, multiplication and functionality, the

51
activity of key enzymes and the expression of genes known to be associated with growth
processes. The effect of these compounds on immune function and disease resistance would also
be highly valuable.

52
CHAPTER 4 GENERAL DISCUSSION
The objective of this study was to evaluate the effect of supplemental dietary NT,
provided through a commercial feed additive, on growth, feed efficiency and efficiency of
protein and energy utilization in rainbow trout fed diets formulated to contain low levels of NT.
Traditionally, aquaculture feeds were formulated to contain high levels of fish meal, which is
characterized by an excellent profile of essential amino acid, minerals, omega-3 fatty acids, fat
soluble vitamins, phospholipids, taurine, and numerous other compounds (Watanabe et al., 1997;
Zinn et al., 2009). Due to high cost and limited global supply of fish meal, modern aquaculture
feeds are increasingly formulated to contain low levels of fish meal and high levels of more cost-
effective ingredients, such as plant protein ingredients. Several limitations, such as the presence
of anti-nutritional factors and poorer levels of essential nutrients, are associated with use of plant
ingredients in feeds. However, it is possible to produce successful rainbow trout feeds containing
high levels of plant protein ingredients at minimal fish meal levels (Hua and Bureau, 2012).
Complete replacement of fish meal and other animal ingredients, however, has proved to be very
difficult in salmonid diets even when formulating to digestible essential nutrient levels in excess
of all known requirements of these animals. Growth and feed efficiency of rainbow trout fed fish
meal and animal protein free diets are generally significantly lower than that of fish fed diets
with some levels of fish meal and/or animal protein ingredients (Cho et al., 1974; Gomes et al.,
1995; Adelizi et al., 1998; Glencross et al., 2011; Burr et al., 2012).
In the present study, the level of fish meal in the diet was from 25% down to 5% and 0%,
and this negatively affected the growth and feed efficiency of rainbow trout, which is consistent
with the findings of earlier studies. In the present study, a series of experimental diets was
formulated with plant protein ingredients that have previously been shown to be a source of

53
highly digestible essential amino acids, contain low levels of anti-nutritional factors and be of
high nutritive value to rainbow trout (unpublished results). This makes it unlikely that
differences in performance were due to different in digestible amino acids, energy or essential
nutrient contents of the diets or to the effect of anti-nutritional factors.
The results appear to indicate that fish meal may have contributed some dietary
components that may be beneficial to these fish. Growth performance and feed efficiency of the
fish remain nonetheless relative good and the fish did not present any overt signs of deficiency or
nutritional pathologies. This may indicate that the dietary components contributed by fish meal
may be nutrients or metabolites that the animal is able to synthesize but unable to synthesize
sufficient quantities of in order to enable higher growth and higher efficiency of feed and
nutrient utilization.
Nucleotides are natural components of all living organisms that are involved in a wide
variety of metabolic and cellular processes. Animals are capable of de novo synthesis of
nucleotides but they can also acquire them through the salvage pathway or through the diet
(Quan et al., 1990). It has been observed that de novo synthesis and the salvage pathways
occasionally provide insufficient amounts of NT for the needs of certain tissues.
Studies have indicated that dietary NT supplementation had positive effect on various
physiological functions in many species, especially during early life stages (Gil, 2002). It has
been hypothesized that during periods of rapid growth or high metabolism, some tissues such as
immune cells and gastrointestinal cells may have high demands for NT. Dietary NT
supplementation can have a positive effect on fish growth performance, immune responses and
GI tract of different fish species (Burrells et al., 2001b; Sakai et al., 2001; Li & Gatlin, 2006;

54
Cheng et al., 2011; Sauer et al, 2011; Tahmasebi-Kohyani et al., 2011; Mohebbi et al., 2013;
Peng et al., 2013).
Fish meal is generally recognized as a good source of NTs, whereas plant protein
ingredients are not (Devresse, 2000). In this thesis, the effect of supplementation of diets
containing with 5% or 0% fish meal with graded levels of NT fed to rainbow trout was
examined. Results suggested that dietary NT supplementation had a beneficial effect on rainbow
trout fed diets with 5% fish meal. The feed intake and final body weight of fish increased
significantly in a linear fashion, and feed efficiency increased significantly in a quadratic fashion
with increasing NT supplementation. However, the fish fed the diet with no fish meal did not
show any significant response to NT supplementation. This may indicate that other nutrients may
have become deficient in this diet and prevented the fish from responding to NT
supplementation. In theory, this difference in response of the fish to NT supplementation should
have resulted in a significant interaction between fish meal level and nucleotide supplementation.
However, no such significant statistical interaction response was detected in this study. The p
value for the interaction response on TGC was 0.17 which is close to the established significance
level (p ≤ 0.05). Overall, variability in terms of feed intake, final body weight and feed
efficiency among tanks fed the same diet was relatively large. Additional feeding trials should be
carried out to determine if the results of this study can be replicated. Future studies should be
designed to be more statistically powerful by using larger number of replicates per experimental
diet, exploring broader and more numerous NT inclusion levels to help better determine whether
NT supplementation is truly beneficial or a simple experimental artifact.
The potential positive effect of NT supplementation in low fish meal diet observed in
this study as well as in other studies (Barros et al., 2015) could be related to different

55
mechanisms. Certain nucleotides have been found to have significant attractant properties for
fish diets. Dietary IMP supplementation (2.8%) significantly enhanced feed intake of largemouth
bass (Micropterus salmoides) fed soybean-based diet (Kubitza et al., 1997). Electrophysiological
studies indicated that, AMP, IMP, UMP and ADP were detected by lips chemoreceptors of
puffer fish (Fugu pardalis) (Kiyohara et al., 1975). Free adenine was found to have potent ability
to inhibit feed intake in rainbow trout (Rumsey et al., 1992). These evidences indicate that NTS
and their metabolites can influence palatability of feeds.
In the present study, the feed intake in rainbow trout fed with Diet 4 (5% fish meal, 16
ppm NT supplementation) was no statistically different (P=0.39) from that of the fish fed with
control diet with 25% fish meal, no NT supplementation. However, the feed efficiency of fish
fed Diet 4 was still significantly (p<0.01) lower than those of fish fed the control diet. These
results suggest a potential effect of NT supplementation on palatability of the diet with 5% fish
meal. However, increase feed intake can be the results of increase palatability or other biological
or metabolic effects. NT could increase metabolic efficiency or growth potential and thus
indirectly influence the feed intake of the animals. The use of a pair-feeding protocol in which all
the groups of fish would have received the same amount of feed would have been an effective
way of determining if the NT supplementation exerted its effect through metabolic effects. Truly
determining the effects of NT on palatability of feeds would require an elaborate protocol. This
could be done through a feed preference assay (Suresh et al., 2011).
As reviewed in some level of details in Chapter 2, NT supplementation has been found to
have significant effect on number of models, notably in young fast growing animals like the
young rainbow trout in this study. Nucleotides are building blocks for rapidly dividing tissues

56
and play many important roles in intermediary metabolisms of all living organisms. As
mentioned above.
The NT supplement used here was a commercial yeast-based feed additive containing
approximately 15% total potentially available nucleotides (TPAN) as well as several other
components, such as amino acids, vitamins, cell wall components, etc. Since NT
supplementation also was done concurrently with these other components, a response to one or
several of these other components cannot be ruled out. Limited information is available on the
actual NT composition of this commercial supplement. Analysis of the NT content of the
experimental diet is still in progress and no results can be reported at this time.
Most other studies examining the effects of dietary NT supplementation have been based
on the use of commercial supplements of poorly characterized composition. Very few studies
have used purified single nucleotides (Kubitza et al., 1997; Lin et al., 2009; Huu et al., 2013).
Different individual NTs or combinations of NTs in different ratios may have different effects.
For example, 0.4% GMP and 0.1% AMP and 0.4% GMP and 0.1% IMP improved growth rate
and weight gain in black tiger prawn (P. monodon) (Huu et al., 2013). Supplementation of the
diet with a nucleotide mixture (IMP:AMP:GMP:UMP:CMP) significantly increased weight gain
in grouper (Epinephelus malabaricus) in a 8-week trial but only increased weight gain
significantly in red drum (Sciaenop ocellatus) during the first week of a 4-week trial (Li et al.,
2007a; Lin et al., 2009). Dietary supplementation of purified AMP in grouper resulted in
significantly higher superoxide anion (O2-) production, a measure of activity of phagocytic cells
(Secombes, 1990), compared to that of fish fed with other purified nucleotides (Lin et al., 2009).
The present study produced valuable results that opens up the door to additional studies.
The reduction in performance in diet with low or no fish meal indicates that our understanding of

57
nutritional requirements for salmonid species is still imperfect. Certain nutrients, compounds or
properties of importance to salmonid fish may be contributed by fish meal and these are poorly
characterized. This also suggests that dietary NT may have a beneficial effect and could
potentially be considered semi or conditionally essential nutrients. These observations deserve to
be examined very carefully in future research efforts.
The experimental diets used in this study represent an interesting model for future studies
on the effect of NT and other dietary components. The combination of high quality plant protein
ingredients used in this study appeared to result in fairly highly nutritive and palatable diets. The
5% fish meal and 0% diets could be supplement with various nutrients and chemical compounds
alone or in combination to determine what is apparently “missing” from fish meal-free diets.
The scope of future studies could be expanded to include assessment of the effect of NT
and other dietary components on digestion, cell, tissue or organ integrity, multiplication and
functionality, assessment of the activity of key enzymes and the expression of genes known to be
associated with growth processes. The effect of these compounds on immune function and
disease resistance would also be highly interesting since dietary NT supplementation have been
observed to have a significant effect on immune function and disease resistance in several animal
models. As such, immune function may represent a highly sensitive indicator of the response of
the animals to dietary NT.

58
References
Adamek, Z., Hamackova, J., Kouril, J., Vachta, R., Stibranyiova, I. (1996). Effect of ascogen
probiotics supplementation on farming success in rainbow trout (Oncorhynchus mykiss)
and wels (Silurus glais) under conditions of intensive culture. Krmiva (Zagreb) 38, 11 –
20.
Adelizi, P., Rosati, R., Warner, K., Wu, Y., Muench, T., White, M., & Brown, P. (1998).
Evaluation of fish-meal free diets for rainbow trout, Oncorhynchus mykiss. Aquac
Nutrition Aquaculture Nutrition, 4(4), 255-262.
Global Feed Survey, 2015. Alltech. Kentucky, USA.
Andrés-Elias, N., Pujols, J., Badiola, I., & Torrallardona, D. (2007). Effect of nucleotides and
carob pulp on gut health and performance of weanling piglets. Livestock Science, 108(1-
3), 280-283.
Barros, M. M., Guimarães, I. G., Pezzato, L. E., Orsi, R. D., Junior, A. C., Teixeira, C. P., Fleuri,
L. F., Padovani, C. R. (2015). The effects of dietary nucleotide mixture on growth
performance, haematological and immunological parameters of Nile tilapia. Aquaculture
Research Aquac Res, 46(4), 987-993.
Biswas, A. K., Kaku, H., Ji, S. C., Seoka, M., & Takii, K. (2007). Use of soybean meal and
phytase for partial replacement of fish meal in the diet of red sea bream, Pagrus major.
Aquaculture, 267(1-4), 284-291.
Bronk, J. R., & Hastewell, J. G. (1987). The transport of pyrimidines into tissue rings cut from
rat small intestine. The Journal of Physiology, 382(1), 475-488.

59
Burel, C., Boujard, T., Tulli, F., & Kaushik, S. J. (2000a). Digestibility of extruded peas,
extruded lupin, and rapeseed meal in rainbow trout (Oncorhynchus mykiss) and turbot
(Psetta maxima). Aquaculture, 188(3-4), 285-298.
Burel, C., Boujard, T., Escaffre, A., Kaushik, S. J., Boeuf, G., Mol, K. A., Van der Geyten, S.,
Kühn, E. R. (2000b). Dietary low-glucosinolate rapeseed meal affects thyroid status and
nutrient utilization in rainbow trout (Oncorhynchus mykiss). BJN British Journal of
Nutrition, 83(06), 653.
Burr, G. S., Wolters, W. R., Barrows, F. T., & Hardy, R. W. (2012). Replacing fish meal with
blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus
mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture, 334-
337, 110-116.
Burrells, C., Williams, P., & Forno, P. (2001a). Dietary nucleotides: a novel supplement in fish
feeds 1. effects on resistance to disease in salmonids. Aquaculture, 199(1-2), 159-169.
Burrells, C., Williams, P., Southgate, P., & Wadsworth, S. (2001b). Dietary nucleotides: a novel
supplement in fish feeds 2. effects on vaccination, salt water transfer, growth rates and
physiology of Atlantic salmon (Salmo salar L.). Aquaculture, 199(1-2), 171-184.
Carr, W. E., & Thompson, H. W. (1983). Adenosine 5'-monophosphate, an internal regulatory
agent, is a potent chemoattractant for a marine shrimp. Journal of Comparative
Physiology. A J. Comp. Physiol., 153(1), 47-53.
Carr, W. E., Netherton, J. C., & Milstead, M. L. (1984). Chemoattractants of the shrimp,
Palaemonetes pugio: variability in responsiveness and the stimulatory capacity of
mixtures containing amino acids, quaternary ammonium compounds, purines and other

60
substances. Comparative Biochemistry and Physiology Part A: Physiology, 77(3), 469-
474.
Carver, J.D., Cox, W.I., Barness, L.A. (1990). Dietary nucleotide effects upon murine
naturakiller cell activity and macrophage activation. J Parenter. Enter. Nutr. 14, 18-22.
Carver, J. D. (1994). Dietary nucleotides: cellular immune, intestinal and hepatic system effects.
The Journal of Nutrition, 124, 144S-148S.
Carver, J. D., & Walker, W. A. (1995). The role of nucleotides in human nutrition. The Journal
of Nutritional Biochemistry, 6(2), 58-72.
Chekeni, F., Elliott, M., Sandilos, J., Walk, S., Kinchen, J., Lazarowski, E., Armstrong, A. J.,
Penuela, S., Laird, D. W., Salvesen, G. S., Isakson, B. E., Ravichandran, K. (2010).
Pannexin 1 channels mediate ‘find-me’ signal release and membrane permeability during
apoptosis. Nature, 467(7317), 863.
Cheng, Z., Buentello, A., & Iii, D. M. (2011). Dietary nucleotides influence immune responses
and intestinal morphology of red drum Sciaenops ocellatus. Fish & Shellfish
Immunology, 30(1), 143-147.
Cho, C. Y., Bayley, H. S., & Slinger, S. J. (1974). Partial replacement of herring meal with
soybean meal and other changes in a diet for rainbow trout (Salmo gairdneri). J. Fish.
Res. Bd. Can. Journal of the Fisheries Research Board of Canada, 31(9), 1523-1528.
Chou, R., Her, B., Su, M., Hwang, G., Wu, Y., & Chen, H. (2004). Substituting fish meal with
soybean meal in diets of juvenile cobia Rachycentron canadum. Aquaculture, 229(1-4),
325-333.
Choudhury, A., Pierce, M. E., Nguyen, D., Storace, L., & Confalone, P. N. (2005). Synthesis of
D-D4FC, a biologically active nucleoside via an unprecedented palladium mediated

61
ferrier rearrangement-type glycosidation with an aromatization prone xylo-furanoid
glycal. Tetrahedron Letters, 46(47), 8099-8102.
Cliddord, A., & Story, D. (1976). Levels of purines in foods and their metabolic effects in rats.
Journal of Nutrition, 106(3), 435-442.
Cosgrove, M. (1998). Nucleotides. Nutrition, 14(10), 748-751.
Dabrowski, K., & Kozak, B. (1979). The use of fish meal and soyabean meal as a protein source
in the diet of grass carp fry. Aquaculture, 18(2), 107-114.
Davies, S. J., Mcconnell, S., & Bateson, R. I. (1990). Potential of rapeseed meal as an alternative
protein source in complete diets for tilapia (Oreochromis mossambicus Peters).
Aquaculture, 87(2), 145-154.
Deluca, C., & Kaplan, N. O. (1958). Flavin adenine dinucleotide synthesis in animal tissues.
Biochimica Et Biophysica Acta, 30(1), 6-11.
Devresse, B., 2000. Nucleotides—a key nutrient for shrimp immune system. Feed Mix 8, 20–22.
Eltzschig, H. K., Eckle, T., Mager, A., Kuper, N., Karcher, C., Weissmuller, T., Boengler, K.,
Schulz, R., Robson, S. C., Colgan, S. P. (2006). ATP release from activated neutrophils
occurs via connexin 43 and modulates adenosine-dependent endothelial cell function.
Circulation Research, 99(10), 1100-1108.
Faigle, M., Seessle, J., Zug, S., Kasmi, K. C., & Eltzschig, H. K. (2008). ATP release from
vascular endothelia occurs across Cx43 hemichannels and is attenuated during hypoxia.
PLoS ONE, 3(7).
Fournier, V., Huelvan, C., & Desbruyeres, E. (2004). Incorporation of a mixture of plant
feedstuffs as substitute for fish meal in diets of juvenile turbot (Psetta maxima).
Aquaculture, 236(1-4), 451-465.

62
Fustin, J.M., Masao, D., Yamada, H., Komatsu, R., Shimba, S., Okamura, H. (2012). Rhythmic
nucleotide synthesis in the liver: temporal segregation of metabolites. Cell Reports, 341-
349.
Gatlin III, D. M., Barrows, F. T., Brown, P., Dabrowski, K., Gaylord, T. G., Hardy, R. W.,
Herman, E., Hu, G., Krogdahl, A., Nelson, R., Overturf, K., Rust, M., Sealey, W.,
Skonberg, D., Souza, E. J., Stone, D., Wilson, R., Wurtele, E. (2007). Expanding the
utilization of sustainable plant products in aquafeeds: A review. Aquaculture Research
Aquaculture Res, 38(6), 551-579.
Gil, A. (2002). Modulation of the immune response mediated by dietary nucleotides. European
Journal of Clinical Nutrition, S1-S4.
Glencross, B., Rutherford, N., & Hawkins, W. (2011). A comparison of the growth performance
of rainbow trout (Oncorhynchus mykiss) when fed soybean, narrow-leaf or yellow lupin
meals in extruded diets. Aquaculture Nutrition, 17(2).
Gomes, E. F., Rema, P., & Kaushik, S. J. (1995). Replacement of fish meal by plant proteins in
the diet of rainbow trout (Oncorhynchus mykiss): digestibility and growth performance.
Aquaculture, 130(2-3), 177-186.
Grimble, G.K., Westwood, M.R. (2000). Nucleotides. In: Gershwin, M.E., German, J.B., Keen,
C.L. (Eds.), Nutrition and Immunology: Principles and Practice. Humana Press, Totowa,
NJ, USA, pp. 135-167.
Guttman, B. (2013). Nucleotide. Brenner's Encyclopedia of Genetics, 139.
Henderson, J., & Paterson, A. (1973). Nucleotide metabolism; an introduction. New York:
Academic Press.

63
Hendricks, J. D. (2002). Adventitious toxins. In Halver, J. E., & Hardy, R. W. (Ed.), Fish
Nutrition (pp. 601-649). San Diego: Academic Press.
Hess, J. R., & Greenberg, N. A. (2012). The role of nucleotides in the immune and
gastrointestinal systems: potential clinical applications. Nutrition in Clinical Practice,
27(2), 281-294.
Higgs, D. A., Mcbride, J. R., Markert, J. R., Dosanjh, B. S., Plotnikoff, M., & Clarke, W. (1982).
Evaluation of tower and candle rapeseed (canola) meal and bronowski rapeseed protein
concentrate as protein supplements in practical dry diets for juvenile chinook salmon
(Oncorhynchus tshawytscha). Aquaculture, 29(1-2), 1-31.
Holen, E., & Jonsson, R. (2004). Dietary nucleotides and intestinal cell lines: I. modulation of
growth. Nutrition Research, 24(3), 197-207.
Hu, L., Yun, B., Xue, M., Wang, J., Wu, X., Zheng, Y., & Han, F. (2013). Effects of fish meal
quality and fish meal substitution by animal protein blend on growth performance, flesh
quality and liver histology of Japanese seabass (Lateolabrax japonicus). Aquaculture,
372-375, 52-61.
Hua, K., & Bureau, D. P. (2012). Exploring the possibility of quantifying the effects of plant
protein ingredients in fish feeds using meta-analysis and nutritional model simulation-
based approaches. Aquaculture, 356-357, 284-301.
Huu, H.D., Tabrett, S., Hoffmann, K., Koppel, P., Lucas, J.S., Barnes, A.C. (2012). Dietary
nucleotides are semi-essential nutrients for optimal growth of black tiger shrimp
(Penaeus monodon). Aquaculture 366-367, 115-121.
Idzko, M., Ferrari, D., & Eltzschig, H. K. (2014). Nucleotide signalling during inflammation.
Nature, 509(7500), 310-317.

64
Jha, A. K., Pal, A., Sahu, N., Kumar, S., & Mukherjee, S. (2007). Haemato-immunological
responses to dietary yeast RNA, ω-3 fatty acid and β-carotene in Catla catla juveniles.
Fish & Shellfish Immunology, 23(5), 917-927.
Ji, S., Liu, R., Ren, M., Li, H., & Xu, J. (2015). Enhanced production of polysaccharide through
the overexpression of homologous uridine diphosphate glucose pyrophosphorylase gene
in a submerged culture of lingzhi or reishi medicinal mushroom, ganoderma lucidum
(higher basidiomycetes). International Journal of Medicinal Mushrooms Int J Med
Mushrooms, 17(5), 435-442.
Junger, W. G. (2011). Immune cell regulation by autocrine purinergic signalling. Nat Rev
Immunol Nature Reviews Immunology, 11(3), 201-212.
Jyonouchi, H. (1994). Nucleotide actions on humoral immune response. J. Nutr. 124 ŽSuppl. 1S.,
138S–143S.
Kaushik, S., Cravedi, J., Lalles, J., Sumpter, J., Fauconneau, B., & Laroche, M. (1995). Partial or
total replacement of fish meal by soybean protein on growth, protein utilization, potential
estrogenic or antigenic effects, cholesterolemia and flesh quality in rainbow trout,
Oncorhynchus mykiss. Aquaculture, 133(3-4), 257-274.
Kenari, A. A., Mahmoudi, N., Soltani, M., & Abediankenari, S. (2013). Dietary nucleotide
supplements influence the growth, haemato-immunological parameters and stress
responses in endangered caspian brown trout (Salmo trutta caspius Kessler, 1877).
Aquacult Nutr Aquaculture Nutrition, 19(1), 54-63.
King, M. W., 1996-2016. Nucleotide metabolism: Nucleic acid synthesis.
Kiyohara, S., Hidaka, I., Tamura, T., 1975. Gustatory response in the puffer-II. single fiber
analysis. Bull. Jpn. Soc. Sci. Fish. 41, 383– 391.

65
Krogdahl, Å, Lea, T. B., & Olli, J. J. (1994). Soybean proteinase inhibitors affect intestinal
trypsin activities and amino acid digestibilities in rainbow trout (Oncorhynchus mykiss).
Comparative Biochemistry and Physiology Part A: Physiology, 107(1), 215-219.
Kubitza, F., Lovshin, L.L., Lovell, R.T., 1997. Identification of feed enhancers for largemouth
bass Micropterus salmoides. Aquaculture 148, 191– 200.
Kulkarni, A.D., Fanslow, W.C., Drath, D.B., Rudolph, F.B., Van Buren, C.T. (1986a). Influence
of dietary nucleotide restriction on bacterial sepsis and phagocytic cell function in mice.
Arch. Surg. 121, 169–172.
Kulkarni, A.D., Fanslow, W.C., Rudolph, F.B., Van Buren, C.T. (1986b). Effect of dietary
nucleotides on response to bacterial infections. J. Parenter. Enteral Nutr. 10, 169–171.
Lazarowski, E. R. (2012). Vesicular and conductive mechanisms of nucleotide release.
Purinergic Signalling, 8(3), 359-373.
Leonardi, M., Sandino, A.M., Klempau, A. (2003). Effect of a nucleotide-enriched diet on the
immune system, plasma cortisol levels and resistance to infectious pancreatic necrosis
(IPN) in juvenile rainbow trout (Oncorhynchus mykiss). Bull. Eur. Assoc. Fish Pathol.
23, 52–59.
Lewis, H. A., & Kohler, C. C. (2008). Corn gluten meal partially replaces dietary fish meal
without compromising growth or fatty acid composition of sunshine bass. North
American Journal of Aquaculture, 70(1), 50-60.
Li, P., Lewis, D. H., & Gatlin III, D. M. (2004). Dietary oligonucleotides from yeast RNA
influence immune responses and resistance of hybrid striped bass (Morone chrysops X
Morone saxatilis) to Streptococcus iniae infection. Fish & Shellfish Immunology, 16(5),
561-569.

66
Li, P., Burr, G. S., Goff, J., Whiteman, K. W., Davis, K. B., Vega, R. R., Neill, W. H., Gatlin, D.
M. (2005). A preliminary study on the effects of dietary supplementation of brewers
yeast and nucleotides, singularly or in combination, on juvenile red drum (Sciaenops
ocellatus). Aquaculture Research Aquaculture Res, 36(11), 1120-1127.
Li, P., Gatlin III, D.M. (2006). Nucleotide nutrition in fish: Current knowledge and future
applications. Aquaculture 251:141-152.
Li, P., Gatlin, D. M., & Neill, W. H. (2007a). Dietary supplementation of a purified nucleotide
mixture transiently enhanced growth and feed utilization of juvenile red drum, Sciaenops
ocellatus. J World Aquaculture Soc Journal of the World Aquaculture Society, 38(2),
281-286.
Li, P., Lawrence, A.L., Gastille, F.L., Gatlin, D.M. (2007b). Preliminary evaluation of a purified
nucleotide mixture as a dietary supplement for Pacific white shrimp Litopenaeus
vannamei (Boone). Aquac Res 38, 887-890.
Li, H., Zhao, P., Lei, Y., Li, T., & Kim, I. (2015). Response to an Escherichia coli K88 oral
challenge and productivity of weanling pigs receiving a dietary nucleotides supplement.
Journal of Animal Science and Biotechnology J Animal Sci Biotechnol, 6(1).
Lim, S., Kim, S., Ko, G., Song, J., Oh, D., Kim, J., Kim, J., Lee, K. (2011). Fish meal
replacement by soybean meal in diets for tiger puffer, Takifugu rubripes. Aquaculture,
313(1-4), 165-170.
Lin, Y., Wang, H., & Shiau, S. (2009). Dietary nucleotide supplementation enhances growth and
immune responses of grouper, Epinephelus malabaricus. Aquaculture Nutrition, 15(2),
117-122.

67
Low, C., Wadsworth, S., Burrells, C., & Secombes, C. (2003). Expression of immune genes in
turbot (Scophthalmus maximus) fed a nucleotide-supplemented diet. Aquaculture, 221(1-
4), 23-40.
Mackie, A.M., 1973. The chemical basis of food detection in the lobster Homarus gammarus.
Mar. Biol. 21, 103– 108.
Mackie, A.M., Adron, J.W., 1978. Identification of inosine and inosine 5V-monophosphate as
the gustatory feeding stimulants for the turbot, Scophthalmus maximus. Comp. Biochem.
Physiol. 60A, 79–83.
Matsumoto, Y., Adje, A.A., Yamauchi, K., Kise, M., Nakasone, Y., Shinegawa, Y., Yokoyama,
H., Yamamoto, S. (1995). Mixture of nucleosides and nucleotides increases bone marrow
cell and peripheral neutrophil number in mice infected with methicillin-resistant
Staphylococcus aureus. Biochemical and molecular roles of nutrients. J. Nutr. 125, 815–
822.
Mawson, R., Heaney, R. K., Piskula, M., & Kozlowska, H. (1993). Rapeseed meal-
glucosinolates and their antinutritional effects part 1. rapeseed production and chemistry
of glucosinolates. Food / Nahrung Nahrung, 37(2), 131-140.
Men, K., Ai, Q., Mai, K., Xu, W., Zhang, Y., & Zhou, H. (2014). Effects of dietary corn gluten
meal on growth, digestion and protein metabolism in relation to IGF-I gene expression of
Japanese seabass, Lateolabrax japonicus. Aquaculture, 428-429, 303-309.
Mente, E., Deguara, S., Santos, M. B., & Houlihan, D. (2003). White muscle free amino acid
concentrations following feeding a maize gluten dietary protein in Atlantic salmon
(Salmo salar L.). Aquaculture, 225(1-4), 133-147.

68
Mohebbi, A., Nematollahi, A., Gholamhoseini, A., Tahmasebi-Kohyani, A., & Keyvanshokooh,
S. (2013). Effects of dietary nucleotides on the antioxidant status and serum lipids of
rainbow trout (Oncorhynchus mykiss). Aquacult Nutr Aquaculture Nutrition, 19(4), 506-
514.
Moore, K., Mullan, B., Pluske, J., Kim, J., & D'souza, D. (2011). The use of nucleotides,
vitamins and functional amino acids to enhance the structure of the small intestine and
circulating measures of immune function in the post-weaned piglet. Animal Feed Science
and Technology, 165(3-4), 184-190.
Murthy, H. S., Li, P., Lawrence, A. L., & Gatlin, D. M. (2009). Dietary β-glucan and nucleotide
effects on growth, survival and immune responses of Pacific white shrimp, Litopenaeus
vannamei. Journal of Applied Aquaculture, 21(3), 160-168.
Nates, S. F. (2013). The role of renderers in aquaculture feeds. Latin American Rendering
Association.
NRC (National Research Council). 2011. Nutrient requirements of fish and shrimp. The National
Academy Press, Washington, DC.
Oliva-Teles, A., Gouveia, A., Gomes, E., & Rema, P. (1994). The effect of different processing
treatments on soybean meal utilization by rainbow trout, Oncorhynchus mykiss.
Aquaculture, 124(1-4), 343-349.
Peng, M., Xu, W., Ai, Q., Mai, K., Liufu, Z., Zhang, K. (2013). Effects of nucleotide
supplementation on growth, immune responses and intestinal morphology in juvenile
turbot fed diets with graded levels of soybeam meal (Scophtalmus maximus L.).
Aquaculture 392-395, 51-58.

69
Pereira, T. G., & Oliva-Teles, A. (2003). Evaluation of corn gluten meal as a protein source in
diets for gilthead sea bream (Sparus aurata L.) juveniles. Aquaculture Research Aquac
Research, 34(13), 1111-1117.
Pesnot, T., Kempter, J., Schemies, J., Pergolizzi, G., Uciechowska, U., Rumpf, T., Sippl, W.,
Jung, M., Wagner, G. K. (2011). Two-step synthesis of novel, bioactive derivatives of the
ubiquitous cofactor nicotinamide adenine dinucleotide (NAD). J. Med. Chem. Journal of
Medicinal Chemistry, 54(10), 3492-3499.
Pham, M. A., Lee, K., Lim, S., & Park, K. (2007). Evaluation of cottonseed and soybean meal
as partial replacement for fish meal in diets for juvenile Japanese flounder Paralichthys
olivaceus. Fisheries Science Fisheries Sci, 73(4), 760-769.
Quan, R., Barness, L.A., Uauy, R. (1990). Do infants need nucleotide supplemented formula for
optimal nutrition. Journal of Pediatric Gastroenterology and Nutrition 11, 429-434.
Ramadan, A., Atef, M., & Afifi, N. (1991). Effect of the biogenic performance enhancer
(Ascogen “S”) on growth rate of tilapia fish. Acta Veterinaria Scandinavica, 304-306.
Ramadan, A., Afifi, N. A., Moustafa, M., & Samy, A. (1994). The effect of ascogen on the
immune response of tilapia fish to Aeromonas hydrophila vaccine. Fish & Shellfish
Immunology, 4(3), 159-165.
Refstie, S., Korsøen, Ø J., Storebakken, T., Baeverfjord, G., Lein, I., & Roem, A. J. (2000).
Differing nutritional responses to dietary soybean meal in rainbow trout (Oncorhynchus
mykiss) and Atlantic salmon (Salmo salar). Aquaculture, 190(1-2), 49-63.
Regost, C., Arzel, J., & Kaushik, S. (1999). Partial or total replacement of fish meal by corn
gluten meal in diet for turbot (Psetta maxima). Aquaculture, 180(1-2), 99-117.

70
Robaina, L., Moyano, F., Izquierdo, M., Socorro, J., Vergara, J., & Montero, D. (1997). Corn
gluten and meat and bone meals as protein sources in diets for gilthead seabream (Sparus
aurata): nutritional and histological implications. Aquaculture, 157(3-4), 347-359.
Rudolph, F. (1994). The biochemistry and physiology of nucleotides. J. Nutr. 124(1), 124s-127s.
Rumsey, G.L., Winfree, R.A., Hughes, S.G., 1992. Nutritional value of dietary nucleic acids and
purine bases to rainbow trout (Oncorhynchus mykiss). Aquaculture 108, 97–110.
Russo, R., Mitchell, H., & Yanong, R. P. (2006). Characterization of Streptococcus iniae isolated
from ornamental cyprinid fishes and development of challenge models. Aquaculture,
256(1-4), 105-110.
Sanderson, I., & He, Y. (1994). Nucleotide uptake and metabolism by intestinal epithelial cells.
The Journal of Nutrition, 124(1), 131-137.
Sakai, M., 1999. Current research status of fish immunostimulants. Aquaculture 172, 63– 92.
Sakai, M., Taniguchi, K., Mamoto, K., Ogawa, H., Tabata, M. (2001). Immunostimulant effects
of nucleotide isolated from yeast RNA on carp, Cyprinus carpio L.. Journal of Fish
Diseases, 433-438.
Sauer, N., Bauer, E., Vahjen, W., Zentek, J., & Mosenthin, R. (2010). Nucleotides modify
growth of selected intestinal bacteria in vitro. Livestock Science, 133(1-3), 161-163.
Sauer, N., Mosenthin, R., & Bauer, E. (2011). The role of dietary nucleotides in single-
stomached animals. Nutr. Res. Rev. Nutrition Research Reviews, 24(01), 46-59.
Secombes C.J. (1990) Isolation of salmonid macrophages and analysis of their killing activity.
In: Techniques in Fish Immunology, Vol. 1 (ed. by J.S. Stolen, T.C. Fletcher, D.P.
Anderson, & B.S. Roberson), pp. 137–163. SOS Publications, Fair Haven, NJ, USA.

71
Spinelli, J., Houle, C. R., & Wekell, J. C. (1983). The effect of phytates on the growth of
rainbow trout (Salmo gairdneri) fed purified diets containing varying quantities of
calcium and magnesium. Aquaculture, 30(1-4), 71-83.
Storebakken, T., Shearer, K., & Roem, A. (1998). Availability of protein, phosphorus and other
elements in fish meal, soy-protein concentrate and phytase-treated soy-protein-
concentrate-based diets to Atlantic salmon, Salmo salar. Aquaculture, 161(1-4), 365-379.
Storebakken, T., Refstie, S., & Ruyter, B. (2000). Soy products as fat and protein sources in fish
diets for intensive aquaculture. Soy in Animal Nutrition, 127-170.
Suresh, A. V., Vasagam, K. K., & Nates, S. (2011). Attractability and palatability of protein
ingredients of aquatic and terrestrial animal origin, and their practical value for blue
shrimp, Litopenaeus stylirostris fed diets formulated with high levels of poultry
byproduct meal. Aquaculture, 319(1-2), 132-140.
Tahmasebi-Kohyani, A., Keyvanshokooh, S., Nematollahi, A., Mahmoudi, N., & Pasha-Zanoosi,
H. (2011). Dietary administration of nucleotides to enhance growth, humoral immune
responses, and disease resistance of the rainbow trout (Oncorhynchus mykiss) fingerlings.
Fish & Shellfish Immunology, 30(1), 189-193.
Tahmasebi-Kohyani, A., Keyvanshokooh, S., Nematollahi, A., Mahmoudi, N., & Pasha-Zanoosi,
H. (2011). Effects of dietary nucleotides supplementation on rainbow trout
(Oncorhynchus mykiss) performance and acute stress response. Fish Physiol Biochem
Fish Physiology and Biochemistry, 38(2), 431-440.
Tan, Q., Liu, Q., Chen, X., Wang, M., & Wu, Z. (2013). Growth performance, biochemical
indices and hepatopancreatic function of grass carp, Ctenopharyngodon idellus, would be
impaired by dietary rapeseed meal. Aquaculture, 414-415, 119-126.

72
Toko, I. I., Fiogbe, E. D., & Kestemont, P. (2008). Mineral status of African catfish (Clarias
gariepinus) fed diets containing graded levels of soybean or cottonseed meals.
Aquaculture, 275(1-4), 298-305.
Uauy, R., Stringel, G., Thomas, R., Quan, R., 1990. Effect of dietary nucleotides on growth and
maturation of the developing gut in the rat. J. Pediatr. Gastroenterol Nutr. 10, 497–503.
Usami, M., Iso, A., Kasahara, H., Kotani, G., Haji, S., Kanamaru, T., & Saitoh, Y. (1996). Effect
of a parenteral nucleoside-nucleotide mixture on hepatic metabolism in partially
hepatectomized cirrhotic rats. Nutrition, 12(6), 436-439.
Van Buren, C. T., Kulkarni, A. D., Fanslow, W. C., & Rudolph, F. B. (1985). Dietary
nucleotides, a requirement for helper/inducer T lymphocytes. Transplantation, 40(6),
694-697.
Van Buren, C. T., & Rudolph, F. (1997). Dietary nucleotides: A conditional requirement.
Nutrition, 470-472.
Viola, S., Mokady, S., Rappaport, U., & Arieli, Y. (1982). Partial and complete replacement of
fish meal by soybean meal in feeds for intensive culture of carp. Aquaculture, 26(3-4),
223-236.
Wang, Y., Kong, L., Li, C., & Bureau, D. (2010). The potential of land animal protein
ingredients to replace fish meal in diets for cuneate drum, Nibea miichthioides , is
affected by dietary protein level. Aquaculture Nutrition, 16(1), 37-43.
Watanabe, T., Verakunpiriya, V., Watanabe, K., Kiron, V., & Satoh, S. (1997). Feeding of
rainbow trout with non-fish meal diets. Fisheries Science, 63(2), 258-266.
Windsor, M. (1971). Fish meal. Aberdeen: Torry Research Station.

73
Welker, T. L., Lim, C., Yildirim-Aksoy, M., & Klesius, P. H. (2011). Effects of dietary
supplementation of a purified nucleotide mixture on immune function and disease and
stress resistance in channel catfish, Ictalurus punctatus. Aquaculture Research, 42(12),
1878-1889.
Wischke, C., Weigel, J., Bulavina, L., & Lendlein, A. (2014). Sustained release carrier for
adenosine triphosphate as signaling molecule. Journal of Controlled Release, 195, 86-91.
Xu, M., Liang, R., Guo, Q., Wang, S., Zhao, M., Zhang, Z. (2013). Dietary nucleotides extend
the life span in sprague-dawley rats. The Journal of Nutrition, Health & Aging, 223-229.
Yousefi M, Abtahi B, Kenari A (2011) A hematological, serum biochemical parameters, and
physiological responses to acute stress of beluga sturgeon (Huso huso, Linnaeus 1785)
juveniles fed dietary nucleotide. Comp Clin Pathol.
Yurkowski, M., Bailey, J. K., Evans, R. E., Tabachek, J. L., Ayles, G. B., & Eales, J. (1978).
Acceptability of rapeseed proteins in diets of rainbow trout (Salmo gairdneri). J. Fish.
Res. Bd. Can. Journal of the Fisheries Research Board of Canada, 35(7), 951-962.
Zhang, Y., Øverland, M., Shearer, K. D., Sørensen, M., Mydland, L. T., & Storebakken, T.
(2012). Optimizing plant protein combinations in fish meal-free diets for rainbow trout
(Oncorhynchus mykiss) by a mixture model. Aquaculture, 360-361, 25-36.
Zhong, G., Hua, X., Yuan, K., & Zhou, H. (2011). Effect of CGM on growth performance and
digestibility in puffer (Takifugu fasciatus). Aquaculture International Aquacult Int, 19(3),
395-403.
Zinn, K. E., Hernot, D. C., Fastinger, N. D., Karr-Lilienthal, L. K., Bechtel, P. J., Swanson, K.
S., & Fahey, G. C. (2009). Fish protein substrates can substitute effectively for poultry
by-product meal when incorporated in high-quality senior dog diets. Journal of Animal

74
Physiology and Animal Nutrition, 93(4), 447-455.
Zrenner, R., Stitt, M., Sonnewald, U., & Boldt, R. (2006). Pyrimidine and purine biosynthesis
and degradation in plants. Annual Review of Plant Biology Annu. Rev. Plant Biol., 57(1),
805-836.


















