
THE EARLY MIOCENE OF WESTERN VENEZUELA,...
18
A QUANTITATIVE PALYNOLOGICAL RECORD FROM THE EARLY MIOCENE OF WESTERN VENEZUELA, WITH EMPHASIS ON MANGROVES Palynology, 25 (2001): 109–126 © 2001 by AASP Foundation ISSN 0191-6122 VALENTÍ RULL PDVSA Exploration and Production Caracas Venezuela Postal address: PA1394 PO Box 02-5304 Miami, FL 33102-5304 U.S.A. e-mail: [email protected] Abstract The quantitative reconstruction presented in this paper documents paleoecological trends of the northern Maracaibo basin (western Venezuela), during the Early Miocene, through pollen analysis of drill core samples from the La Rosa (shallow marine) and Lagunillas (coastal plain) formations. Palynological assemblages were grouped statistically into three assemblages representing man- groves, herbaceous back-mangrove swamps, and inland palm/ fern swamps. Mangroves were the local vegetation during the deposition of La Rosa Formation and, after a regressive event, were replaced by herbaceous back-mangroves during the sedi- mentation of Lagunillas Formation. The palynological assem- blage representing palm/fern swamps dominates the entire se- quence and is interpreted as a large background signal reflecting transport by rivers. Mangrove communities had few species, and represented a transitional phase in the mangrove community evolution, after the terminal Eocene biotic crisis. INTRODUCTION This paper presents a palynological study of Early Mio- cene La Rosa and Lagunillas formations from the NE of the Maracaibo basin in Venezuela, in order to reconstruct the paleosuccession of coastal plant communities, with empha- sis on mangroves. It is a part of a broader project which is aimed at reconstructing coastal and mangrove communities and their paleosuccession in the Tertiary of the Maracaibo basin using quantitative pollen analysis and statistical paleo- ecological methods to increase the degree of objectivity in both the composition and the interpretation of palynological taphocenoses. Paleocommunities should be reconstructed from suitable taphocenoses, and not with qualitative or semi- quantitative data. In palynology, this is achieved through representative counts including all the palynomorph types present in samples (Rull, 1987), and comparison with mod- ern analogs (Rull, 1998a). Furthermore, empirical associa- tions based solely in the botanical affinities of the fossil taxa and the taxonomic composition of present communities (a common procedure) implicitly assume that communities have not changed through time, which is totally untrue, especially for mangroves (Graham, 1995; Rull, 1998b). To properly record past associations, more objective methods, such as statistical associations, should be applied, before looking at botanical affinities (Rull, 1998c). So far, several Paleocene and Eocene mangroves communities have been reconstructed with these methods (Rull, 1992; 1997b; 1998b; 1998c; 1999; 2000). However, post-Eocene mangroves, although well-known taxonomically (Lorente, 1986; Muller et al., 1987), have not yet been studied with these methods. From a palynostratigraphic point of view, the Early Mio- cene in Venezuela is characterized by two zones (Lorente, 1986): Verrutricolporites and Psiladiporites, which are equi- valent to the northern South American pollen zones Verrutricolporites rotundiporus–Echidiporites barbeitoensis, and Echitricolporites maristellae–Psiladiporites minimus, re- spectively (Muller et al., 1987). The zone of Verrutricolporites is an interval zone, having its base in the first occurrence of Verrutricolporites rotundiporus and its top below the first occurrence of Psiladiporites minimus or Echitricolporites maristellae. In addition, two other taxa: Bombacacidites zuatensis and Psilatricolporites pachydermatus have their first occurrence at the base of this zone. According to Lorente (1986), pollen assemblages for this zone are dominated in the entire basin by Zonocostites ramonae, the other important components being Psilatricolporites crassus, Bombacacidites baumfalki, Retitricolporites irregularis, Retitricolporites hispidus, Retitricolporites simplex, Jandufouria seamrogiformis, Verrucatosporites usmensis, Proxapertites operculatus, Monoporites annulatus, Mauritiidites franciscoi, Striatricolpites catatumbus, Psilatricolporites maculosus, and
Transcript of THE EARLY MIOCENE OF WESTERN VENEZUELA,...

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 109A QUANTITATIVE PALYNOLOGICAL RECORD FROMTHE EARLY MIOCENE OF WESTERN VENEZUELA,WITH EMPHASIS ON MANGROVES
Palynology, 25 (2001): 109–126© 2001 by AASP Foundation ISSN 0191-6122
VALENTÍ RULLPDVSA Exploration and ProductionCaracasVenezuelaPostal address: PA1394PO Box 02-5304Miami, FL 33102-5304U.S.A.e-mail: [email protected]
Abstract
The quantitative reconstruction presented in this paper documentspaleoecological trends of the northern Maracaibo basin (westernVenezuela), during the Early Miocene, through pollen analysis ofdrill core samples from the La Rosa (shallow marine) and Lagunillas(coastal plain) formations. Palynological assemblages weregrouped statistically into three assemblages representing man-groves, herbaceous back-mangrove swamps, and inland palm/fern swamps. Mangroves were the local vegetation during thedeposition of La Rosa Formation and, after a regressive event,were replaced by herbaceous back-mangroves during the sedi-mentation of Lagunillas Formation. The palynological assem-blage representing palm/fern swamps dominates the entire se-quence and is interpreted as a large background signal reflectingtransport by rivers. Mangrove communities had few species, andrepresented a transitional phase in the mangrove communityevolution, after the terminal Eocene biotic crisis.
INTRODUCTION
This paper presents a palynological study of Early Mio-cene La Rosa and Lagunillas formations from the NE of theMaracaibo basin in Venezuela, in order to reconstruct thepaleosuccession of coastal plant communities, with empha-sis on mangroves. It is a part of a broader project which isaimed at reconstructing coastal and mangrove communitiesand their paleosuccession in the Tertiary of the Maracaibobasin using quantitative pollen analysis and statistical paleo-ecological methods to increase the degree of objectivity inboth the composition and the interpretation of palynologicaltaphocenoses. Paleocommunities should be reconstructedfrom suitable taphocenoses, and not with qualitative or semi-quantitative data. In palynology, this is achieved throughrepresentative counts including all the palynomorph typespresent in samples (Rull, 1987), and comparison with mod-ern analogs (Rull, 1998a). Furthermore, empirical associa-
tions based solely in the botanical affinities of the fossil taxaand the taxonomic composition of present communities (acommon procedure) implicitly assume that communitieshave not changed through time, which is totally untrue,especially for mangroves (Graham, 1995; Rull, 1998b). Toproperly record past associations, more objective methods,such as statistical associations, should be applied, beforelooking at botanical affinities (Rull, 1998c). So far, severalPaleocene and Eocene mangroves communities have beenreconstructed with these methods (Rull, 1992; 1997b; 1998b;1998c; 1999; 2000). However, post-Eocene mangroves,although well-known taxonomically (Lorente, 1986; Mulleret al., 1987), have not yet been studied with these methods.
From a palynostratigraphic point of view, the Early Mio-cene in Venezuela is characterized by two zones (Lorente,1986): Verrutricolporites and Psiladiporites, which are equi-valent to the northern South American pollen zonesVerrutricolporites rotundiporus–Echidiporites barbeitoensis,and Echitricolporites maristellae–Psiladiporites minimus, re-spectively (Muller et al., 1987). The zone of Verrutricolporitesis an interval zone, having its base in the first occurrence ofVerrutricolporites rotundiporus and its top below the firstoccurrence of Psiladiporites minimus or Echitricolporitesmaristellae. In addition, two other taxa: Bombacaciditeszuatensis and Psilatricolporites pachydermatus have theirfirst occurrence at the base of this zone. According to Lorente(1986), pollen assemblages for this zone are dominated in theentire basin by Zonocostites ramonae, the other importantcomponents being Psilatricolporites crassus, Bombacaciditesbaumfalki, Retitricolporites irregularis, Retitricolporiteshispidus, Retitricolporites simplex, Jandufouriaseamrogiformis, Verrucatosporites usmensis, Proxapertitesoperculatus, Monoporites annulatus, Mauritiidites franciscoi,Striatricolpites catatumbus, Psilatricolporites maculosus, and

110 PALYNOLOGY, VOLUME 25 — 2001
Podocarpidites sp. The inferred sedimentary environment iscoastal to shallow marine. The interval zone of Psiladiporitesis defined by the first occurrence of Psiladiporites minimus, atthe base. In western Venezuela, the first occurrence ofEchitricolporites maristellae is also characteristic, and thezone is called Psiladiporites–Echitricolporites (Lorente, 1986).The top is defined by the first occurrence of Crassoretitriletesvanraadshooveni. In the Maracaibo basin, this zone can besubdivided into two well-differentiated intervals. The lowerinterval is dominated by Monoporites annulatus,Echitricolporites maristellae, Malvacipollis spinulosa,Psilatricolporites operculatus, Crototricolpites annemariae,Bombacacidites baculatus, Retimonocolpites longicolpatus,Verrucatosporites usmensis, Magnastriatites grandiosus,Psilamonocolpites medius, Zonocostites ramonae,Mauritiidites franciscoi, and Retitricolporites guianensis, andwas deposited under alluvial to lower coastal plain condi-tions, with some geographical variations. The upper intervalis similar but in it Zonocostites ramonae decreases in abun-dance, and disappears in some areas. The sedimentary envi-ronments were alluvial to coastal plains, with more marineconditions in the south (Lorente, 1986).
In the mangrove communities, Zonocostites ramonaewas abundant and geographically widespread in westernVenezuela, during the Early Miocene. This form-speciescorresponds to the fossil pollen of Rhizophora, the domi-nant genus in present-day Neotropical mangrove commu-nities (Chapman, 1976; Tomlinson, 1986). The fossil recordshows that this pollen appeared in the Late Eocene in lowquantities, and has been dominant in lower coastal man-grove communities from the Miocene to the present. Othermangrove-forming trees progressively appeared in theMiocene (Avicennia), Pliocene (Laguncularia) and Qua-ternary (Conocarpus) (Graham, 1995). The Eocene man-grove communities were radically different in composi-tion. Their main elements are nowadays extinct in theNeotropics (Nypa, Brevitricolpites variabilis) or reducedto a relict area (Pelliciera). The only extant component isthe fern Acrostichum aureum, represented in the fossilrecord by Deltoidospora adriennis. The evolutionary changefrom Eocene to Miocene–Recent mangroves is assumed tohave been not gradual, and related to the terminal Eocenebiotic crisis, probably due to climatic causes (Rull, 1998b).
MATERIAL AND METHODS
Study Site and Lithology
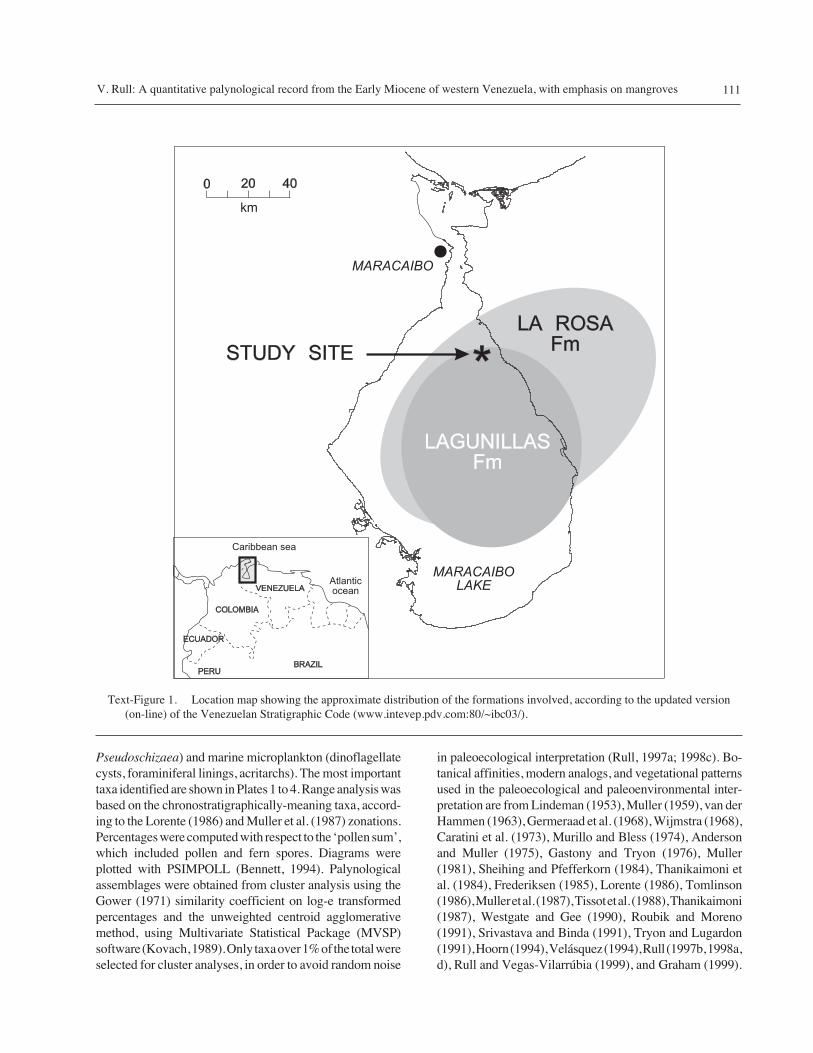
This study was carried out on core samples from well TJlocated at the NE part of the Maracaibo basin, in westernVenezuela (Text-Figure 1). The geological formations
involved are La Rosa and lower Lagunillas. The La RosaFormation unconformably overlies the Eocene Misoa For-mation. It is approximately 200 m thick in the type locality,but can be as thick as 1000 m in the eastern part of the basin.It consists of greenish fossiliferous clayey shales withintercalated sandstone layers, and has been subdivided intofour units: the basal Santa Bárbara Member (unconsoli-dated clayey sandstones), the La Rosa shales (with thetypical lithology described for the formation), the Interme-diate Sand (with a mixture of the main La Rosa lithologies)and the La Rosa sands (fine-grained sandstones, and shaleswith mollusks and foraminifers). The fossil content indi-cates an Early Miocene age, and initial deposition in aneritic environment. The Santa Bárbara Member repre-sents the initial transgression on the eroded Eocene, laterculminating in a shallow sea covering most of the Maracaibobasin (represented by La Rosa shales). During the subse-quent regression, the ‘intermediate sand’ and the ‘La Rosasand’ (informal names) were deposited in estuarine andbeach, and ultimately deltaic environments, transitional tothe overlying Lagunillas Formation (González de Juana etal., 1980).
The Lagunillas Formation is 300 to ~1000 m thick,consisting of unconsolidated sandstones, with intermingledclays, shales, and lignites. In the eastern part of the basin,it has been subdivided into three members: Lower Lagunillas(fine-grained sandstones with intercalated shales), Laguna(gray fossiliferous shales and gray sandstones), andBachaquero (thick clayey sandstones with clays and shales).The Lagunillas Formation ranges from Lower to MiddleMiocene in age, and represents the continuation of theregressive sequence initiated in the La Rosa Formation,although an intermediate increase of marine conditions arerepresented by the Laguna Member. The Lower LagunillasMember was deposited in a deltaic environment, prograd-ing from the S–SE, and the Bachaquero Member wasdeposited in fluvial and deltaic environments (González deJuana et al., 1980). In the present study, only the LowerLagunillas member has been examined.
Laboratory and Analytical Methods
Palynological analyses were carried out on all the shalyintervals of the selected cores (31 samples). Samples weredigested in HF and HCl, and centrifuged with ZnBr2, beforemounting in glycerin-jelly. In order to have statistically-representative proportions, counts were done on randomtransects until the saturation of diversity (Rull, 1987). Finalaverage counts were 424 for sporomorphs (pollen and fernspores) and 497 for the total, which include fungal spores,freshwater algal remains (Pediastrum, Botryococcus,
V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 111
Pseudoschizaea) and marine microplankton (dinoflagellatecysts, foraminiferal linings, acritarchs). The most importanttaxa identified are shown in Plates 1 to 4. Range analysis wasbased on the chronostratigraphically-meaning taxa, accord-ing to the Lorente (1986) and Muller et al. (1987) zonations.Percentages were computed with respect to the ‘pollen sum’,which included pollen and fern spores. Diagrams wereplotted with PSIMPOLL (Bennett, 1994). Palynologicalassemblages were obtained from cluster analysis using theGower (1971) similarity coefficient on log-e transformedpercentages and the unweighted centroid agglomerativemethod, using Multivariate Statistical Package (MVSP)software (Kovach, 1989). Only taxa over 1% of the total wereselected for cluster analyses, in order to avoid random noise
in paleoecological interpretation (Rull, 1997a; 1998c). Bo-tanical affinities, modern analogs, and vegetational patternsused in the paleoecological and paleoenvironmental inter-pretation are from Lindeman (1953), Muller (1959), van derHammen (1963), Germeraad et al. (1968), Wijmstra (1968),Caratini et al. (1973), Murillo and Bless (1974), Andersonand Muller (1975), Gastony and Tryon (1976), Muller(1981), Sheihing and Pfefferkorn (1984), Thanikaimoni etal. (1984), Frederiksen (1985), Lorente (1986), Tomlinson(1986), Muller et al. (1987), Tissot et al. (1988), Thanikaimoni(1987), Westgate and Gee (1990), Roubik and Moreno(1991), Srivastava and Binda (1991), Tryon and Lugardon(1991), Hoorn (1994), Velásquez (1994), Rull (1997b, 1998a,d), Rull and Vegas-Vilarrúbia (1999), and Graham (1999).
Text-Figure 1. Location map showing the approximate distribution of the formations involved, according to the updated version(on-line) of the Venezuelan Stratigraphic Code (www.intevep.pdv.com:80/~ibc03/).
112 PALYNOLOGY, VOLUME 25 — 2001
RESULTS AND INTERPRETATION
Palynostratigraphy
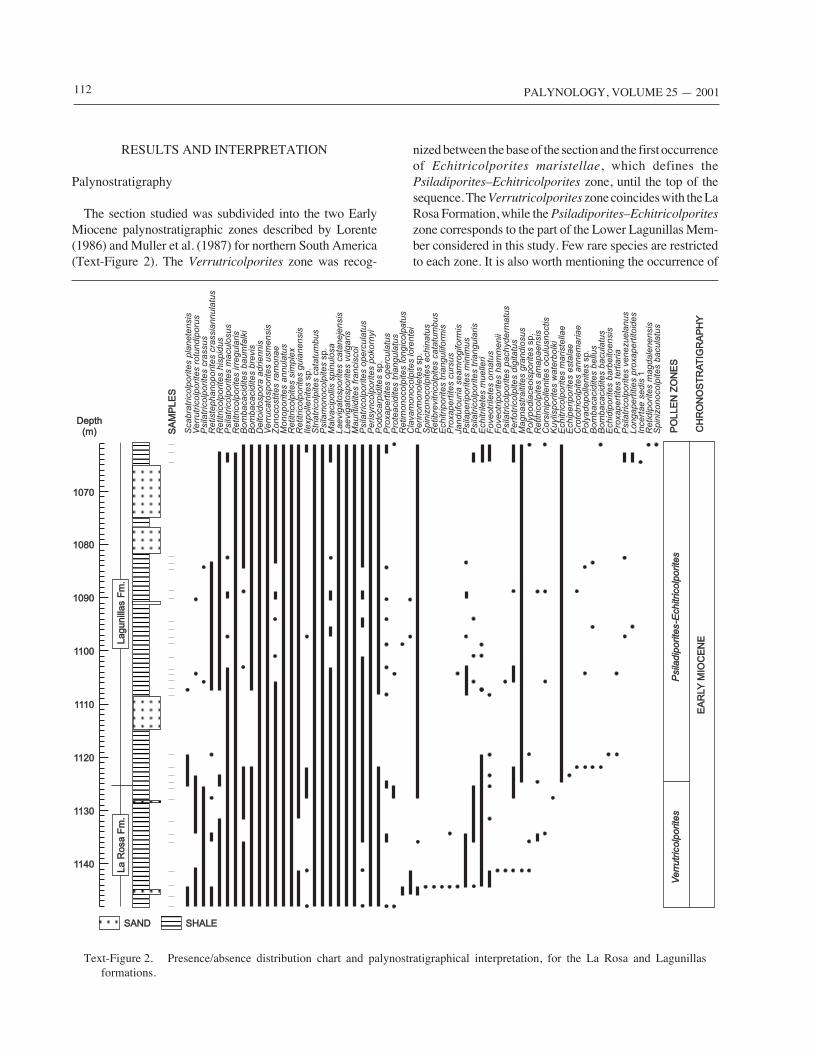
The section studied was subdivided into the two EarlyMiocene palynostratigraphic zones described by Lorente(1986) and Muller et al. (1987) for northern South America(Text-Figure 2). The Verrutricolporites zone was recog-
nized between the base of the section and the first occurrenceof Echitricolporites maristellae, which defines thePsiladiporites–Echitricolporites zone, until the top of thesequence. The Verrutricolporites zone coincides with the LaRosa Formation, while the Psiladiporites–Echitricolporiteszone corresponds to the part of the Lower Lagunillas Mem-ber considered in this study. Few rare species are restrictedto each zone. It is also worth mentioning the occurrence of
Text-Figure 2. Presence/absence distribution chart and palynostratigraphical interpretation, for the La Rosa and Lagunillasformations.
Sca
bra
tric
olp
orite
s p
lan
ete
nsis
Ve
rru
tric
olp
orite
s r
otu
nd
ipo
rus
Psila
tric
olp
orite
s c
rassu
sR
etiste
ph
an
op
orite
s c
rassia
nn
ula
tus
Re
titr
ico
lpo
rite
s h
isp
idu
sP
sila
tric
olp
orite
s m
acu
losu
sR
etitr
ico
lpo
rite
s irr
eg
ula
ris
Bo
mb
aca
cid
ite
s b
au
mfa
lki
Bo
mb
aca
cid
ite
s b
revis
De
lto
ido
sp
ora
ad
rie
nn
isV
err
uca
tosp
orite
s u
sm
en
sis
Zo
no
co
stite
s r
am
on
ae
Mo
no
po
rite
s a
nn
ula
tus
Re
titr
ico
lpite
s s
imp
lex
Re
titr
ico
lpo
rite
s g
uia
ne
nsis
Ile
xp
olle
nite
s s
p.
Str
iatr
ico
lpite
s c
ata
tum
bu
sP
sila
mo
no
co
lpite
s s
p.
Ma
lva
cip
olli
s s
pin
ulo
sa
La
evig
ato
sp
orite
s c
ata
ne
jen
sis
La
evig
ato
sp
orite
s v
ulg
aris
Ma
uritiid
ite
s fra
ncis
co
iP
sila
tric
olp
orite
s o
pe
rcu
latu
sP
erisyn
co
lpo
rite
s p
oko
rnyi
Po
do
ca
rpid
ite
s s
p.
Pro
xa
pe
rtite
s o
pe
rcu
latu
sP
rote
acid
ite
s t
ria
ng
ula
tus
Re
tim
on
oco
lpite
s lo
ng
ico
lpa
tus
Cla
va
mo
no
co
lpite
s lo
ren
tei
Pe
rin
om
on
ole
tes s
p.
Sp
iniz
on
oco
lpite
s e
ch
ina
tus
Re
tib
revitrico
lpite
s c
ata
tum
bu
sE
ch
itrip
orite
s t
ria
ng
ulif
orm
isP
roxa
pe
rtite
s c
urs
us
Ja
nd
ufo
uria
se
am
rog
ifo
rmis
Psila
pe
rip
orite
s m
inim
us
Psila
tric
olp
orite
s t
ria
ng
ula
ris
Ech
itrile
tes m
ue
lleri
Fo
ve
otr
ilete
s o
rna
tus
Fo
ve
otr
ipo
rite
s h
am
me
nii
Psila
tric
olp
orite
s p
ach
yd
erm
atu
sP
erf
otr
ico
lpite
s d
igita
tus
Ma
gn
astr
aitite
s g
ran
dio
su
sP
oly
po
dia
ce
ois
po
rite
s s
p.
Re
titr
ico
lpite
s a
ma
pa
en
sis
Co
rsin
ipo
llen
ite
s o
cu
lusn
octis
Ku
ylis
po
rite
s w
ate
rbo
lki
Ech
itrico
lpo
rite
s m
ariste
llae
Ech
ipe
rip
orite
s e
sta
lae
Cro
totr
ico
lpite
s a
nn
em
aria
eP
oly
ad
op
olle
nite
s s
p.
Bo
mb
aca
cid
ite
s b
ellu
sB
om
ba
ca
cid
ite
s b
acu
latu
sE
ch
idip
orite
s b
arb
eito
en
sis
Pro
xa
pe
rtite
s t
ert
iaria
Psila
tric
olp
orite
s v
en
ezu
ela
nu
sL
on
ga
pe
rtitie
s p
roxa
pe
rtito
ide
sIn
ce
rta
e s
ed
is 1
Re
tid
ipo
rite
s m
ag
da
len
en
sis
Sp
iniz
on
oco
lpite
s b
acu
latu
s
Verr
utr
icolp
orite
sV
err
utr
icolp
orite
sP
sila
dip
orite
s-E
ch
itricolp
orite
sP
sila
dip
orite
s-E
ch
itricolp
orite
s
EA
RLY
MIO
CE
NE
EA
RLY
MIO
CE
NE
PO
LLE
N Z
ON
ES
PO
LLE
N Z
ON
ES
CH
RO
NO
ST
RA
TIG
RA
PH
YC
HR
ON
OS
TR
AT
IGR
AP
HY
10701070
Depth(m)
Depth(m)
10801080
10901090
11001100
11101110
11201120
11301130
11401140
SANDSAND SHALESHALE
La R
osa
Fm
.L
a R
osa
Fm
.
SA
MP
LE
SS
AM
PLE
S
La
gu
nill
as F
m.
La
gunill
as F
m.

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 113
several Paleogene taxa, such as Spinizonocolpites echinatus,Spinizonocolpites baculatus, Echitriporites trianguliformis,Proxapertites cursus, Foveotriporites hammenii, andRetidiporites magdalenensis. They usually show single oc-currences in isolated samples, and can be assumed to bereworked from older upstream paleo-outcrops.
Paleoecology and Paleoenvironments
Fern and allied spores are generally dominant throughoutthe whole section (Text-Figure 3). Pollen and fungal sporesshow lower values, and aquatic elements (freshwater andmarine together) are only important in the La Rosa Forma-
Text-Figure 3. Percentage diagram for the major groups of palynomorphs.
418418
countscountsPollen
Pollen
Spo
res
Spo
res
Fungi
Fungi
Aqu
atics
Aqu
atics
0 20 400 20 40 0 20 40 600 20 40 60 0 10 0 10 0 20 %0 20 %
413413514514
449449540540411411
370370
547547
527527
521521529529
431431
444444
372372
565565
409409
471471
396396
458458
512512
542542
623623
503503
468468
484484
648648
502502
429429
607607
452452
661661
Ve
rru
tric
olp
orite
sV
err
utr
ico
lpo
rite
sP
sila
dip
orite
s-E
ch
itrico
lpo
rite
sP
sila
dip
orite
s-E
ch
itrico
lpo
rite
s
EA
RLY
MIO
CE
NE
EA
RLY
MIO
CE
NE
10701070
Depth(m)
Depth(m)
10801080
10901090
11001100
11101110
11201120
11301130
11401140
SANDSAND
SHALESHALE
La
Ro
sa
Fm
.L
a R
osa
Fm
.
SA
MP
LE
SS
AM
PLE
S
La
gu
nill
as F
m.
La
gun
illa
s F
m.
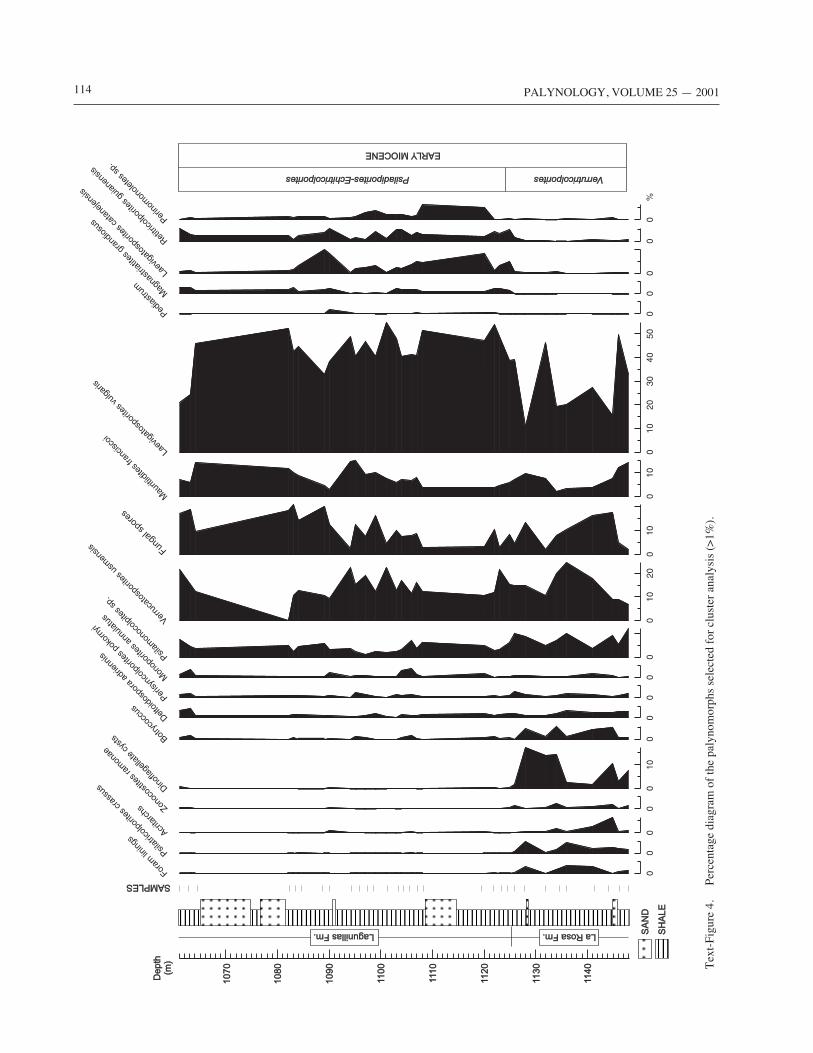
114 PALYNOLOGY, VOLUME 25 — 2001
Tex
t-Fi
gure
4.
Perc
enta
ge d
iagr
am o
f th
e pa
lyno
mor
phs
sele
cted
for
clu
ster
ana
lysi
s (>
1%).
Verrutricolporites VerrutricolporitesPsiladiporites-Echitricolporites Psiladiporites-Echitricolporites
EARLY MIOCENE EARLY MIOCENE
10
70
10
70
De
pth
(m)
De
pth
(m)
10
80
10
80
10
90
10
90
11
00
11
00
111
0111
0
11
20
11
20
11
30
11
30
11
40
11
40
SA
ND
SA
ND
SH
AL
ES
HA
LE
La Rosa Fm. La Rosa Fm.
SAMPLES SAMPLES
Lagunillas Fm. Lagunillas Fm..
.
Funga
l spo
res

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 115
tion (Verrutricolporites zone). In the percentage diagramfor selected taxa (>1%), a clearer differentiation can beseen than using only presence/absence data (Text-Figure4). Dominant and less abundant elements show little varia-tions across the section, although Laevigatosporites vul-garis is slightly more abundant in the Lagunillas Formation(Psiladiporites–Echitricolporites zone). Other elementswith intermediate abundance, however, are clearly distinc-tive. Characteristic of the La Rosa Formation are themangrove elements, Psilatricolporites crassus andZonocostites ramonae, the marine representatives (foram-iniferal linings and acritarchs) and the freshwater algaBotryococcus, indicating sedimentation in coastal and shal-low marine environments, close to mangrove vegetation.Botryococcus, although more typical of freshwater envi-ronments, can also occur in slightly brackish water, due toits tolerance to salinity (Rull, 1997b). The main elementsthat characterize the Lower Lagunillas Member of theLagunillas Formation are fern spores (Magnastriatitesgrandiosus, Laevigatosporites catanejensis, Perino-monoletes sp.), together with Retitricolporites guianensis(of unknown affinity), and the freshwater alga Pediastrum.This is consistent with the presence of alluvial and coastalplain environments flooded by freshwater.
Cluster analysis provided 3 groups, EM1–3, which wereinterpreted according to the known botanical affinities oftheir components and the available modern analog studies(see Table 1 for a summary). Group EM1 is also recognizedin the percentage diagram, and is typical of the La Rosa
Formation (Text-Figures 5 and 6). From a vegetationalpoint of view, it is considered to be a mangrove assemblagedeposited near the mangrove belt or in adjacent shallowbrackish water. The lower Lagunillas Member is character-ized by group EM2 (Text-Figure 6), which is composed ofelements typical of vegetation flooded by fresh or low-salinity water (Magnastriatites grandiosus, Deltoidosporaadriennis, Laevigatosporites catanejensis, Pediastrum). Italso includes other elements with a wide range of environ-mental conditions, which also present in coastal swampsand marshes (Monoporites annulatus, Perisyncolporitespokornyi), and a form-species with unknown botanicalaffinity and environmental requirements (Retitricolporitesguianensis). The majority of these components are fernsor herbs, hence, they probably represent herbaceousswamps and marshes behind the mangrove belt. GroupEM3 is composed of palms (Mauritiidites franciscoi,Psilamonocolpites sp.), ferns (Verrucatosporites usmensis,Laevigatosporites vulgaris), and fungal spores, represent-ing another type of coastal swamps (palm/fern swamps)common in the Neotropics. The palm Mauritiiditesfranciscoi typically develops almost monotypical stands(‘morichales’) in the freshwater swamps of upper coastalplains and savannas, beyond the limit of tidal influence.Therefore, this assemblage is thought to represent the moreinland vegetation recorded in this study.
The succession of these assemblages through timeshows a dominant ‘background’ pollen signal from palm/fern–back-mangrove swamps (EM3) with little changes,
Text-Figure 5. Cluster analysis on the percentages of selected palynomorphs (>1%).
-0.2 0 0.2 0.4 0.6 0.8 1
Gower General Similarity Coefficient
Mauritiidites franciscoiPsilamonocolpites sp.Laevigatosporites vulgarisVerrucatosporites usmensisFungal sporesMonoporites annulatusPerisyncolporites pokornyiDeltoidospora adriennisPerinomonoletes sp.Retitricolporites guianensisLaevigatosporites catanejensisMagnastriatites grandiosusPediastrumPsilatricolporites crassusForam liningsZonocostites ramonaeAcritarchsBotryococcusDinoflagellate cysts
EM1
EM2
EM3

116 PALYNOLOGY, VOLUME 25 — 2001
and the replacement of mangroves (EM1) by herbaceousback-mangrove swamps (EM2), in the La Rosa–Lagunillas transition. Indeed, during deposition of theLa Rosa Formation (Verructricolporites zone), pollensedimentation was almost exclusively composed of palm/
fern swamp sporomorphs and pollen locally depositedunder or in front of the mangrove fringe, with a negli-gible contribution from herbaceous back-mangroveswamps. During deposition of the Lagunillas Formation(Psiladiporites–Echitricolporites zone), although sporo-
1, 2 Deltoidospora adriennis (Potonié & Gelletich 1933)Frederiksen 1983. 3572' (106.4–17.6), 3673' (76.3–25.0).
3, 4 Foveotriletes ornatus Regali et al., 1974. 3481' (84.1–1.5).
5, 6 Magnastriatites grandiosus (Kedves & Solé de Porta)Dueñas 1980. 1963. 3551' (85.0–22.3); 3686' (108.8–2.6); bar = 16 µm for these two specimens.
PLATE 1
7, 8 Laevigatosporites vulgaris Potonié 1934. 3727' (89.9–3.6), 3623' (81.3–17.3).
9, 10 Laevigatosporites catanejensis Muller et al., 1987.3713' (108.3–8.2), 3673' (94.3–18.8).
11, 12 Verrucatosporites usmensis (van der Hammen 1956)Germeraad et al., 1968. 3767' (75.7–5.0).
13, 14 Perinomonoletes sp. Krutzsch 1967. 3673' (110.5–7.5),3760' (112.2–19.9).
Text-Figure 6. Percentage diagram showing the quantitative assemblages established, and the sporomorphs under 1%, withsignificant correlation.
EM
1EM
2EM
3Oth
ers
Ech
itrile
tes m
uelle
ri
Ech
itrile
tes m
uelle
ri
Ret
itricolpo
rites
hispidu
s
Ret
itricolpo
rites
hispidu
s
Ret
itricolpo
rites
irre
gularis
Ret
itricolpo
rites
irre
gularis
Psilatri
colpor
ites m
aculos
us
Psilatri
colpor
ites m
aculos
us
Pod
ocar
pidite
s sp
Pod
ocar
pidite
s sp
.
Ret
itricolpite
s sim
plex
Ret
itricolpite
s sim
plex
Psilatri
colpor
ites op
ercu
latu
s
Psilatri
colpor
ites op
ercu
latu
s
Ech
itricolpo
rites
mar
iste
llae
Ech
itricolpo
rites
mar
iste
llae
Ve
rru
tric
olp
orite
sV
err
utr
ico
lpo
rite
sP
sila
dip
orite
s-E
ch
itrico
lpo
rite
sP
sila
dip
orite
s-E
ch
itrico
lpo
rite
s
EA
RLY
MIO
CE
NE
EA
RLY
MIO
CE
NE
0 50 100 % 0 2 %0 2 %
10701070
Depth(m)
Depth(m)
10801080
10901090
11001100
11101110
11201120
11301130
11401140
SANDSAND
SHALESHALE
La
Ro
sa
Fm
.L
a R
osa
Fm
.
SA
MP
LE
SS
AM
PL
ES
La
gu
nill
as F
m.
La
gu
nill
as F
m.

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 117Plate 1
8 µm

118 PALYNOLOGY, VOLUME 25 — 2001
1 Polyadopollenites sp. Pflug & Thompson 1953. 3681'(109.5–24.0)
2, 3 Podocarpidites sp. Cookson 1947 ex Couper 1953.3492' (104.2–5.4), ; 3760' (102.1–1.4),
4, 5 Psilamonocolpites spp. van der Hammen & García deMutis 1965. 3673' (79.0–14.9), 3755' (104.6–8.4).
6 Mauritiidites franciscoi (van der Hammen 1956) vanHoeken-Klinkenberg 1964. 3481' (99.7–7.5).
7, 8 Echidiporites barbeitoensis Muller et al., 1987. 3673'(11.2–11.2).
9 Verrutricolporites rotundiporus van der Hammen &Wijmstra 1964. 3745' (91.5–3.0).
PLATE 2
10, 11 Psilatricolporites maculosus Regali et al 1974. 3551'(72.8–11.5), 3673' (93.5–20.2).
12, 13, Zonocostites ramonae Germeraad et al., 1968. 3486'14 (91.6–23.0), 3755' (98.1–3.7), 3589' (92.5–18.0).15 Psilatricolporites venezuelanus Lorente 1986. 3551'
(98.7–21.0)16, 17 Psilatricolporites operculatus van der Hammen &
Wijmstra 1964. 3767' (89.7–6.3), 3673' (94.3–3.6).18 Psilatricolporites crassus van der Hammen & Wijmstra
1964. 3700' (105.1–22.5).19 Perfotricolpites digitatus González 1967. 3572' (98.5–
3.9).
TABLE 1. Pearson product-moment coefficients between the palynomorph assemblages and the taxa below 1%. Significance level(α): * 0.05, ** 0.01, *** 0.001.
EM1 EM2 EM3Bombacacidites baumfalki 0.1096 -0.1441 -0.1388Bombacacidites brevis 0.1923 -0.5312** 0.0998Clavamonocolpites lorentei 0.1286 0.0905 -0.2168Corsinipollenites oculus-noctis 0.3186 -0.0051 -0.3785*Echitricolporites maristellae -0.4762** 0.7853*** -0.0028Foveotriletes ornatus 0.0665 -0.0098 -0.1311Ilexpollenites sp. 0.1859 -0.1786 -0.1932Malvacipollis spinulosa -0.0652 0.2543 -0.1381Perfotricolpites digitatus -0.1354 0.3478 -0.1322Podocarpidites sp. 0.4691** -0.3523 -0.4291*Proteacidites triangulatus 0.1287 -0.0779 -0.1857Psilaperiporites minimus 0.1206 0.0718 -0.1970Proxapertites operculatus 0.0059 0.0012 -0.1475Psilatricolporites maculosus 0.6771*** -0.2457 -0.7173***Psilatricolporites operculatus -0.3077 0.6683*** -0.2226Psilatricolporites venezuelanus -0.1764 0.1041 0.0977Psilatricolporites triangularis 0.2878 -0.3527 -0.2090Retistephanoporites crassiannulatus 0.3406 -0.3669* -0.1147Retitricolpites simplex 0.3580* -0.0305 -0.3996*Retitricolporites hispidus 0.3617* -0.3138 -0.2963Retitricolporites irregularis 0.5083** -0.3890* -0.4308*Scabratricolporites planetensis 0.1273 0.0576 -0.2704Striatricolpites catatumbus 0.1373 -0.1027 -0.2323Echitriletes muelleri 0.5745*** -0.1980 -0.5522**Polypodiaceoisporites sp. 0.1249 -0.0403 -0.2356
morphs from palm/fern swamps continued to be domi-nant, sporomorphs from herbaceous swamps increased,and the pollen from mangroves was substantially re-duced to very few grains, probably transported landwardby wind. Therefore, the replacement of mangrove as-
semblage EM1 (coastal) by assemblage EM2 (continen-tal) across the boundary between the La Rosa and theLagunillas formations (Text-Figure 6) can be inter-preted as a regressive trend, from shallow shelf to back-mangrove environments of deposition.

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 119Plate 2
8 µm

120 PALYNOLOGY, VOLUME 25 — 2001
Minor taxa
Elements under 1% (called here minor taxa) were alsoinvestigated for paleoenvironmental and paleoecologicalpurposes by computing their correlations with the threeassemblages found (Rull, 1991). For this analysis, thereworked sporomorphs and the very sparse taxa withonly single occurrences in a few samples, such asBombacacidites baculatus, Bombacacidites bellus,Crototricolpites annemariae, Echidiporites barbeitoensis,Jandufouria seamrogiformis, Kuylisporites waterbolki,Polyadopollenites sp., Proxapertites tertiaria, Psila-tricolporites pachydermatus, Retibrevitricolpitescatatumbus, Retimonocolpites longicolpatus, Retitri-colpites amapaensis were excluded. Four minor taxa havehighly significant correlations (α < 0.01) with the man-grove assemblage (Table 1, Text-Figure 6): Podocarpidites,Psilatricolporites maculosus, Retitricolporites irregularis,and Echitriletes muelleri. The first taxon is the pollen of thegymnosperm tree genus Podocarpus, living in highlandrain forests of northern South America; its pollen is widelydispersed by wind. The form-species Retitricolporitesirregularis corresponds to Amanoa (Euphorbiaceae), a treefrom upper delta environments (Germeraad et al., 1968;Lorente, 1986). Little ecological information is availablefor Psilatricolporites maculosus and Echitriletes muelleri,that have been related to the families Sapotaceae andSelaginellaceae respectively, with wide ranges of occur-rence (Van der Hammen, 1963; Lorente, 1986; Tissot et al.,1988). Two other taxa (Retitricolpites simplex andRetitricolporites hispidus) show lower but still significantcorrelations (α < 0.05). They are similar to the pollen ofinland forest trees of the families Anacardiaceae andFlacourtiaceae, respectively (Lorente, 1986; and observa-tions of the author from herbarium material). Rull (1998c)included Retitricolpites simplex in an Eocene assemblagerepresenting inland forest communities.
Two taxa show highly significant correlations with theback-mangrove herb swamps component: Echitricolporitesmaristellae and Psilatricolporites operculatus. The firstone is a marker species having its first occurrence at the
base of Psiladiporites–Echitricolporites zone; therefore,the significant correlation could be due in part to evolution-ary and not environmental factors. However, when only therange of occurrence of the species is considered, the corre-lation is still significant (r = 0.6772, α < 0.001), indicatinga real statistical association. The botanical affinity ofEchitricolporites maristellae is not well established, al-though it has been related to the Malvaceae or Bombacaceae(Lorente, 1986). The most probable botanical affinity ofProxapertites operculatus is with the palm Astrocaryum,from inland and back-mangrove communities (van derHammen, 1963; Lorente, 1986). None of the taxa consid-ered have a positive significant relationship with the assem-blage EM3. However, the negative correlations of someelements are worth mentioning. Almost all the taxa withsignificant positive correlations with the mangrove assem-blage are negatively associated to the assemblage EM3,except for Retitricolporites hispidus (Table 1).
DISCUSSION
The ‘background’ palynological assemblage from palm/fern swamps is considered to be transported from inlandcommunities, probably upper delta environments, consid-ering that 1) it originated in these environments, 2) themajor transport agents for palynomorphs in coastal envi-ronments are water currents flowing towards the sea, and 3)the La Rosa and Lagunillas Formations were deposited inmore distal environments (mangroves and back-mangroves,respectively). The large amounts of this background signal,however, are surprising. In modern analog studies fromnorthern South America, such quantities are never attainedin shelf environments (Rull, 1998a). This can be explainedby an increased water-transport capacity, which involves avery humid climate, the existence of a major fluvial system,or both. Current paleogeographic reconstructions supportthese points. According to Díaz de Gamero (1996), duringthe Early Miocene, the proto-Orinoco fluvial system drainedmost of the western part of northern South America and itsdelta was situated in the northeast Maracaibo basin.
1, 2 Perfotricolpites digitatus González 1967. 3572' (98.5–3.9), 3690' (79.0–23.8).
3, 4, 5 Bombacacidites brevis Dueñas 1979. 3767' (93.3–10),3713 (106.7–8.9), 3594' (111.2–25.1).
6, 7, 8 Bombacacidites baumfalki Lorente 1986. 3755' (102.8–2.5), 3745' (82.9–21.8).
PLATE 3
9, 10, 11 Retitricolpites simplex González 1967. 3636' (110.1–13.2; 85.6–13.6), 3745' (79.4–14.4).
12, 13, Retitricolporites hispidus van der Hammen & Wijmstra14 1964. 3727' (84.3–3.4; 72.8–13.4).15, 16, Retitricolporites guianensis van der Hammen &17 Wijmstra 1964. 3636' (97.6–9.6; 110.3–6.8), 3673',
(77.8–25.0).

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 121Plate 3
8 µm

122 PALYNOLOGY, VOLUME 25 — 2001
The Early Miocene mangrove communities are repre-sented in this study by the assemblage EM1 of theVerrutricolporites zone (La Rosa Formation), and prob-ably associated minor taxa. The only known mangrove-forming trees present were Rhizophora (Zonocostitesramonae) and Pelliciera (Psilatricolporites crassus), form-ing an intermediate association between typical Eoceneand post-Eocene mangroves, and representing a transi-tional phase after the terminal Eocene impoverishment ofthese communities and their progressive Neogene enrich-ment (Rull, 1998b; Graham, 1995). None of the minor taxastatistically associated with the mangrove assemblage inthe present study has been previously considered a man-grove or mangrove-related element (Graham, 1995). In thecase of Podocarpus, the correlation can be explained bywind transport to coastal environments (Muller, 1959). Apotential source in the southeast could have been theincipient northern Andes, where uplift was initiated shortlybefore, during the late Oligocene (Rull, 1997b). Some ofthe other minor elements also correspond to present mon-tane taxa (Sapotaceae, Alchornea and Anacardiaceae) andhave been found in modern mangrove sediments (Rull andVegas-Vilarrúbia, 1999). Therefore, in the study area,Early Miocene mangroves seem to have been of lowdiversity. However, more work is needed to establish thebotanical affinities of many minor taxa in order to infer abetter paleovegetational history.
Compared with modern analog studies, in which themangrove pollen commonly ranges from 20% to more than70% (Muller, 1963; van der Hammen, 1963; Tissot et al.,1988), the mangrove signal found in this study is not strong(Text-Figure 4). The same is true for several sites of theregion (Root et al., 1998). This probably indicates that thesediments were deposited some distance away from anydense mangrove vegetation, although both marinepalynomorphs (Text-Figure 4) and micropaleontologicalanalysis clearly indicated a shallow-water (0–20 m depth)coastal environment of deposition (Echeverría and Ruiz,1999). Therefore, mangroves were probably reduced and/
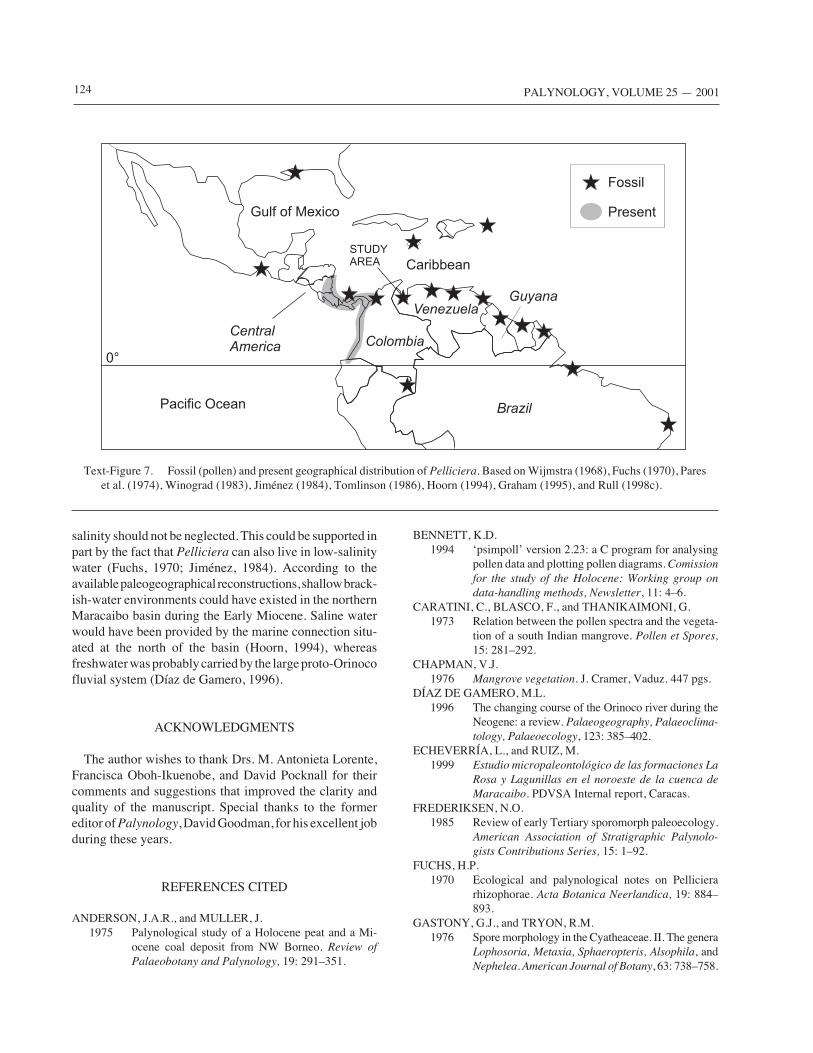
or were not dense in the region, contrasting with other areasof the same basin and eastern Venezuela, where mangrovepollen is abundant in the Early Miocene (Lorente, 1986). A‘dilution’ effect due to the background EM3 assemblage isundoubtedly present, but insufficient to explain such lowvalues. A possible explanation for Pelliciera emerges afterthe consideration of its biogeography. Aside from doubtfulrecords in Europe and Africa (Muller, 1981), Pelliciera isknown to be common in Eocene to Miocene records ofCentral America, northern South America and the Carib-bean–Gulf of Mexico region (Text-Figure 7). A dramaticreduction began in the Miocene (Wijmstra, 1968), leadingto its present relict distribution. It disappeared from thenorthern limits of its range beginning in the Miocene(Graham, 1995), but remained as a scarce element innorthern South America until the Pliocene. Therefore, theEarly Miocene mangrove communities of the Maracaibobasin were probably affected by the initial reduction.
The relative scarcity of Rhizophora, however, remains tobe explained. In the available modern analog studies for theregion that include mangroves, similar numbers of thispollen occur only inland, in freshwater environments, mostprobably due to wind transport (Muller, 1959; ten Broekand Nijssen, 1971). However, inland fluvial paleoenviron-ments for the La Rosa Formation are unlikely because ofthe abundance of marine palynomorphs (Text-Figure 4),and the micropaleontological evidence (see above). Lowpercentages of Rhizophora pollen have been also foundwithin a disturbed mangrove stand surrounded by montaneforests (Rull and Vegas-Vilarrúbia, 1999; Rull et al., 1999).These modern pollen assemblages, however, are very dif-ferent from those found in the present work, because theyare dominated by herbaceous pollen (Gramineae,Cyepraceae, Chenopodiaceae) with negligible percentagesof marine elements. On the other hand, It is known thatRhizophora can survive in low-saline, even in freshwater,where is not as abundant as in truly brackish and marineones (Lindeman, 1953; Van der Hammen, 1963). There-fore, the possibility of shallow marine environments of low
1 Retitricolporites guianensis van der Hammen &Wijmstra 1964. 3623' (86.7–1.9).
2, 3, 4, 5 Retitricolporites irregularis van der Hammen &Wijmstra 1964. 3693' (99.3–24.1), 3686' (78.0–10.8).
6, 7, 8 Striatricolpites catatumbus González 1967. 3600'(110.2–14.5), 3481' (104.1–17.4).
9 Ilexpollenites sp. Thiegart 1937 ex Potonié 1960. 3481'(75.0–19.6).
10, 11, Echitricolporites maristellae Muller et al., 1987. 3628'
PLATE 4
12 (94.7–16.7), 3486' (95.5–21.0).13, 14 Psilaperiporites minimus Regali et al., 1974. 3636'
(105.1–20.8).15 Perisyncolporites sp. Germeraad et al., 1968. 3727'
(93.3–10.0).16 Perisyncolporites pokornyi Germeraad et al., 1968.
3628' (90.4–17.4).17 Malvacipollis spinulosa Frederiksen 1983. 3673' (77.8–
8.4).

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 123Plate 4
8 µm
124 PALYNOLOGY, VOLUME 25 — 2001
salinity should not be neglected. This could be supported inpart by the fact that Pelliciera can also live in low-salinitywater (Fuchs, 1970; Jiménez, 1984). According to theavailable paleogeographical reconstructions, shallow brack-ish-water environments could have existed in the northernMaracaibo basin during the Early Miocene. Saline waterwould have been provided by the marine connection situ-ated at the north of the basin (Hoorn, 1994), whereasfreshwater was probably carried by the large proto-Orinocofluvial system (Díaz de Gamero, 1996).
ACKNOWLEDGMENTS
The author wishes to thank Drs. M. Antonieta Lorente,Francisca Oboh-Ikuenobe, and David Pocknall for theircomments and suggestions that improved the clarity andquality of the manuscript. Special thanks to the formereditor of Palynology, David Goodman, for his excellent jobduring these years.
REFERENCES CITED
ANDERSON, J.A.R., and MULLER, J.1975 Palynological study of a Holocene peat and a Mi-
ocene coal deposit from NW Borneo. Review ofPalaeobotany and Palynology, 19: 291–351.
BENNETT, K.D.1994 ‘psimpoll’ version 2.23: a C program for analysing
pollen data and plotting pollen diagrams. Comissionfor the study of the Holocene: Working group ondata-handling methods, Newsletter, 11: 4–6.
CARATINI, C., BLASCO, F., and THANIKAIMONI, G.1973 Relation between the pollen spectra and the vegeta-
tion of a south Indian mangrove. Pollen et Spores,15: 281–292.
CHAPMAN, V.J.1976 Mangrove vegetation. J. Cramer, Vaduz. 447 pgs.
DÍAZ DE GAMERO, M.L.1996 The changing course of the Orinoco river during the
Neogene: a review. Palaeogeography, Palaeoclima-tology, Palaeoecology, 123: 385–402.
ECHEVERRÍA, L., and RUIZ, M.1999 Estudio micropaleontológico de las formaciones La
Rosa y Lagunillas en el noroeste de la cuenca deMaracaibo. PDVSA Internal report, Caracas.
FREDERIKSEN, N.O.1985 Review of early Tertiary sporomorph paleoecology.
American Association of Stratigraphic Palynolo-gists Contributions Series, 15: 1–92.
FUCHS, H.P.1970 Ecological and palynological notes on Pelliciera
rhizophorae. Acta Botanica Neerlandica, 19: 884–893.
GASTONY, G.J., and TRYON, R.M.1976 Spore morphology in the Cyatheaceae. II. The genera
Lophosoria, Metaxia, Sphaeropteris, Alsophila, andNephelea. American Journal of Botany, 63: 738–758.
Text-Figure 7. Fossil (pollen) and present geographical distribution of Pelliciera. Based on Wijmstra (1968), Fuchs (1970), Pareset al. (1974), Winograd (1983), Jiménez (1984), Tomlinson (1986), Hoorn (1994), Graham (1995), and Rull (1998c).
Pacific Ocean
0°
Caribbean
Brazil
GuyanaVenezuela
ColombiaCentralAmerica
STUDYAREA
Gulf of Mexico
Fossil
Present

V. Rull: A quantitative palynological record from the Early Miocene of western Venezuela, with emphasis on mangroves 125
GERMERAAD, J.H., HOPPING, C.A., and MULLER, J.1968 Palynology of Tertiary sediments from tropical ar-
eas. Review of Palaeobotany and Palynology, 6:189–348.
GONZÁLEZ DE JUANA, C., ITURRALDE DE AROZENA,J.M., and PICARD, X.
1980 Geología de Venezuela y de sus cuencas petrolíferas.FONINVES, Caracas. 1031 pgs.
GOWER, J.C.1971 A general coefficient of similarity and some of its
properties. Biometrics, 27: 857–874.GRAHAM, A.
1995 Diversification of the Gulf/Caribbean mangrove com-munities through Cenozoic time. Biotropica, 27: 20–27.
GRAHAM, A.1999 Studies in Neotropical paleobotany. XIII. An Oligo-
Miocene palynoflora from Simojovel (Chiapas,Mexico). American Journal of Botany, 86(1): 17–31.
HOORN, C.1994 Miocene palynostratigraphy and paleoenvironments
of northwestern Amazonia. Unpublished Ph.D. dis-sertation, University of Amsterdam. 156 pgs.
JIMÉNEZ, J.A.1984 A hypothesis to explain the reduced distribution of
the mangrove Pelliciera rhizophorae Tr. & Pl.Biotropica, 16: 304–308.
KOVACH, W.L.1989 Comparisons of multivariate analytical techniques
for the use in pre Quaternary plant palaeoecology.Review of Palaeobotany and Palynology, 60: 255–282.
LINDEMAN, J.C.1953 The vegetation of Suriname. Van Eedenfonds, Am-
sterdam. 135 pgs.LORENTE, M.A.
1986 Palynology and palynofacies of the upper Tertiary inVenezuela. Dissertationes Botanicae, 99: 1–222.
MULLER, J.1959 Palynology of recent Orinoco delta and shelf sedi-
ments. Micropaleontology, 5: 1–23.1981 Fossil pollen records of extant angiosperms. The
Botanical Review, 47(1): 1–141.MULLER, J., DI GIACOMO, E., and VAN ERVE, A.W.
1987 A palynological zonation for the Cretaceous, Ter-tiary, and Quaternary of northern South America.American Association of Stratigraphic Palynolo-gists Contributions Series, 19: 7–76.
MURILLO, M.T., and BLESS, M.J.M.1974 Spores of recent Colombian Pteridophyta. I. Trilete
spores. Review of Palaeobotany and Palynology, 18:223–269.
PARES, M., UESUGUI, N., and SANTOS, A.1974 Palinologia dos sedimentos meso-cenozoicos do
Brasil (I). Boletin Técnico Petrobras, 17: 177–191.ROOT, S.A., CURRY, R.P., and RODGERS, B.K.
1998 Biostratigraphic and environmental analyses of theLagunillas and La Rosa formations in the LL-03
field, Lago Maracaibo, Venezuela. Global BiostratServices, Dallas. 69 pgs.
ROUBIK, D.W., and MORENO, J.E.1991 Pollen and spores of Barro Colorado Island. Mis-
souri Botanical Garden Monographs in SystematicBotany, 36. 268 pgs.
RULL, V.1987 A note on pollen counting in palaeoecology. Pollen
et Spores, 29: 471–480.1991 Palaeoecological significance of chrysophycean
stomatocysts: a statistical approach. Hydrobiologia,220: 161–165.
1992 Paleoecología y análisis secuencial de una seccióndeltaica terciaria de la cuenca de Maracaibo. Boletínde la Sociedad Venezolana de Geólogos, 46: 16–26.
1997a Sequence analysis of Western Venezuelan Creta-ceous to Eocene sediments using palynology. Chrono-palaeoenvironmental and palaeovegetational ap-proaches. Palynology, 21: 79–90.
1997b Oligo-Miocene palynology of the Rio Chama se-quence (Western Venezuela), with comments onfossil algae as paleoenvironmental indicators. Pa-lynology, 21: 213–229.
1998a Middle Eocene mangroves and vegetation changesin the Maracaibo basin. Palaios, 13: 287–296.
1998b Evolución de los manglares neotropicales: La crisisdel Eoceno. Interciencia, 23: 355–362.
1998c Modern and Quaternary palynological studies in theCaribbean and Atlantic coasts of Northern SouthAmerica: A paleoecologically-oriented review.Boletín de la Sociedad Venezolana de Geólogos, 23:5–24.
1998d Biogeographical and evolutionary considerations ofMauritia (Arecaceae), based on palynological evi-dence. Review of Palaeobotany and Palynology,100: 109–122.
1999 Palaeofloristic and palaeovegetational changesacross the Paleocene–Eocene boundary in northernSouth America. Review of Palaeobotany and Pa-leoecology, 107: 83–95.
2000 Ecostratigraphic study of Paleocene and EarlyEocene palynological cyclicity in northern SouthAmerica. Palaios, 15: 14–24.
RULL, V., and VEGAS-VILARRÚBIA, T.1999 Surface palynology of a small coastal basin from
Venezuela and its potential paleoecological applica-tions. Micropaleontology, 45: 365–393.
RULL, V., VEGAS- VILARRÚBIA, T., and ESPINOZA DEPERNÍA, N.
1999 Palynological record of an Early–Mid Holocenemangrove in Eastern Venezuela. Implications forsea-level rise and disturbance history Journal ofCoastal Research, 15: 496–504.
SHEIHING, M.H., and PFEFFERKORN, H.W.1984 The taphonomy of land plants in the Orinoco delta: a
model for the incorporation of plant parts in clasticsediments of Late Carboniferous age of Euamerica.Review of Palaeobotany and Palynology, 41: 205–240.

126 PALYNOLOGY, VOLUME 25 — 2001
SRIVASTAVA, S.K., and BINDA, P.L.1991 Depositional history of the early Eocene Shumaysi
Formation, Saudi Arabia. Palynology, 15: 47–61.TEN BROEK, A.N.Ch., and NIJSSEN, L.
1971 Palynological investigation of recent sediments fromthe Rio Baudo area, Colombia, Pacific coast. Shellinternal report, Rijswijk. 14 pgs.
THANIKAIMONI, G.1987 Mangrove palynology. Intitut français de Pondichéry,
travaux de la section scientifique et technique, 24: 1–100.
THANIKAIMONI, G., CARATINI, C., VENKATACHALA,B.S., RAMANUJAM, C.G.K., and KAR, R.K.
1984 Selected Tertiary angiosperm pollen from India andtheir relationship with African Tertiary pollens. Intitutfrançais de Pondichéry, travaux de la sectionscientifique et technique, 19: 1–92.
TISSOT, C., DJUWANSAH, M.R., and MARIUS, C.1988 Evolution de la mangrove en Guyane au cours de
l’Holocene. Etude palynologique. Institut françaisde Pondichéry, travaux de la section scientifique ettechnique, 25: 125–137.
TOMLINSON, P.B.1986 Mangrove Botany. Cambridge Univ. Press, Cam-
bridge. 413 pgs.TRYON, A.F., and LUGARDON, B.
1991 Spores of the Pteridophyta. Springer-Verlag, NewYork. 648 pgs.
VAN DER HAMMEN, T.1963 A palynological study on the Quaternary of British
Guiana. Leidse Geologische Mededelingen, 29: 125–180.
VELÁSQUEZ, J.1994 Plantas acuáticas vasculares de Venezuela.
Uiversidad Central de Venezuela, Caracas. 992 pgs.WESTGATE, J.W., and GEE, C.T.
1990 Paleoecology of a middle Eocene mangrove biota(vertebrates, plants, and invertebrates) from south-west Texas. Palaeogeography, Palaeoclimatology,Palaeoecology, 78: 163–177.
WIJMSTRA, T.A.1968 The identity of Psilatricolporites and Pelliciera.
Acta Botanica Neerlandica, 17: 114–116.WIJMSTRA, T.A.
1971 The palynology of the Guiana coastal basin. Drukkerijde Kempenaer, Oegstgeest. 62 pgs.
WINOGRAD, M.1983 Observaciones sobre el hallazgo de Pelliciera
rhizophorae (Theaceae) en el Caribe Colombiano.Biotropica, 15: 297–298.


















