
The Conserved Chimeric Transcript UPGRADE2 Is Associated with ...
20
The Conserved Chimeric Transcript UPGRADE2 Is Associated with Unreduced Pollen Formation and Is Exclusively Found in Apomictic Boechera Species 1[C][W][OPEN] Martin Mau, José M. Corral, Heiko Vogel, Michael Melzer, Jörg Fuchs, Markus Kuhlmann, Nico de Storme, Danny Geelen, and Timothy F. Sharbel* Leibniz Institute of Plant Genetics and Crop Plant Research, D–06466 Gatersleben, Germany (M.Ma., J.M.C., M.Me., J.F., M.K., T.F.S.); Genomics Research Group, Department of Entomology, Max Planck Institute for Chemical Ecology, D–07745 Jena, Germany (H.V.); and Plant Production, Faculty of Bioscience Engineering, Ghent University, 9000 Ghent, Belgium (N.d.S., D.G.) ORCID ID: 0000-0002-3450-2733 (T.F.S.). In apomictic Boechera spp., meiotic diplospory leads to the circumvention of meiosis and the suppression of recombination to produce unreduced male and female gametes (i.e. apomeiosis). Here, we have established an early flower developmental staging system and have performed microarray-based comparative gene expression analyses of the pollen mother cell stage in seven diploid sexual and seven diploid apomictic genotypes to identify candidate factors for unreduced pollen formation. We identified a transcript unique to apomictic Boechera spp. called UPGRADE2 (BspUPG2), which is highly up-regulated in their pollen mother cells. BspUPG2 is highly conserved among apomictic Boechera spp. genotypes but has no homolog in sexual Boechera spp. or in any other taxa. BspUPG2 undergoes posttranscriptional processing but lacks a prominent open reading frame. Together with the potential of stably forming microRNA-like secondary structures, we hypothesize that BspUPG2 functions as a long regulatory noncoding messenger RNA-like RNA. BspUPG2 has apparently arisen through a three-step process initiated by ancestral gene duplication of the original BspUPG1 locus, followed by sequential insertions of segmentally duplicated gene fragments, with final exonization of its sequence structure. Its genesis reflects the hybridization history that characterizes the genus Boechera. Haploid gamete formation is an important step dur- ing the diplohaplontic plant life cycle, which marks the switch from sporophyte to gametophyte. During female and male sporogenesis, the diploid megaspore mother cell and pollen mother cell (PMC), respectively, un- dergo one reductional and equational mitotic division to form four haploid daughter cells. During subsequent male spore development (microgametogenesis), each of the four microspores undergoes two mitotic divisions to form a tricellular pollen grain containing two mature reduced gametes (generative nuclei) and one vegetative nucleus. Despite a relatively simple structure, the male gametophyte is regulated by complex gamete-specific molecular programs that are characterized by (1) en- richment of transcripts responsible for DNA repair, cell cycle, and chromosome organization, (2) underrep- resentation of the RNA-processing machinery (Pina et al., 2005; Borges et al., 2008), and (3) developmental stage-speci fic transcript sets whose activities vary temporally from the uninuclear microspore toward the mature pollen grain (Mascarenhas, 1989; Honys and Twell, 2004). Loss-of-function studies of male and female meiosis have revealed cell cycle defects (Yang et al., 2003), ef- fects on homologous chromosome recombination (Lu et al., 2008), and defects in meiotic cell fate (Sundaresan et al., 1995; Yang et al., 1999), all of which lead to re- duced meiocyte fertility or death. Other mutations lead to unreduced (2C) gametes (Brownfield and Köhler, 2011). Apomeiosis is a naturally occurring form of sensu stricto unreduced egg formation, which is the first step of gametophytic apomixis or asexual (clonal) seed formation in plants (Nogler, 1984; Grimanelli et al., 2001a). During apomixis, the apomeiotically derived egg cell undergoes parthenogenetic (i.e. without fer- tilization) development, while endosperm develops with (pseudogamous) or without (autonomous) fertili- zation (Nogler, 1984; Grimanelli et al., 2001a). Apomixis bears the potential to fix hybrid genotypes over gener- ations and, therefore, is of high interest to agronomy (Grimanelli et al., 2001a; Grossniklaus, 2001). 1 This work was supported by the Leibniz Institute of Plant Ge- netics and Crop Plant Research and by the European Union COST action Harnessing Plant Reproduction for Crop Improvement (grant no. FA0903). * Address correspondence to [email protected]. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy de- scribed in the Instructions for Authors (www.plantphysiol.org) is: Timothy F. Sharbel ([email protected]). [C] Some figures in this article are displayed in color online but in black and white in the print edition. [W] The online version of this article contains Web-only data. [OPEN] Articles can be viewed online without a subscription. www.plantphysiol.org/cgi/doi/10.1104/pp.113.222448 1640 Plant Physiology Ò , December 2013, Vol. 163, pp. 1640–1659, www.plantphysiol.org Ó 2013 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.org on January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
Transcript of The Conserved Chimeric Transcript UPGRADE2 Is Associated with ...

The Conserved Chimeric Transcript UPGRADE2 IsAssociated with Unreduced Pollen Formation and IsExclusively Found in Apomictic Boechera Species1[C][W][OPEN]
Martin Mau, José M. Corral, Heiko Vogel, Michael Melzer, Jörg Fuchs, Markus Kuhlmann,Nico de Storme, Danny Geelen, and Timothy F. Sharbel*
Leibniz Institute of Plant Genetics and Crop Plant Research, D–06466 Gatersleben, Germany (M.Ma., J.M.C.,M.Me., J.F., M.K., T.F.S.); Genomics Research Group, Department of Entomology, Max Planck Institute forChemical Ecology, D–07745 Jena, Germany (H.V.); and Plant Production, Faculty of Bioscience Engineering,Ghent University, 9000 Ghent, Belgium (N.d.S., D.G.)
ORCID ID: 0000-0002-3450-2733 (T.F.S.).
In apomictic Boechera spp., meiotic diplospory leads to the circumvention of meiosis and the suppression of recombination toproduce unreduced male and female gametes (i.e. apomeiosis). Here, we have established an early flower developmental stagingsystem and have performed microarray-based comparative gene expression analyses of the pollen mother cell stage in sevendiploid sexual and seven diploid apomictic genotypes to identify candidate factors for unreduced pollen formation. Weidentified a transcript unique to apomictic Boechera spp. called UPGRADE2 (BspUPG2), which is highly up-regulated in theirpollen mother cells. BspUPG2 is highly conserved among apomictic Boechera spp. genotypes but has no homolog in sexualBoechera spp. or in any other taxa. BspUPG2 undergoes posttranscriptional processing but lacks a prominent open reading frame.Together with the potential of stably forming microRNA-like secondary structures, we hypothesize that BspUPG2 functions as along regulatory noncoding messenger RNA-like RNA. BspUPG2 has apparently arisen through a three-step process initiated byancestral gene duplication of the original BspUPG1 locus, followed by sequential insertions of segmentally duplicated genefragments, with final exonization of its sequence structure. Its genesis reflects the hybridization history that characterizes thegenus Boechera.
Haploid gamete formation is an important step dur-ing the diplohaplontic plant life cycle, which marks theswitch from sporophyte to gametophyte. During femaleand male sporogenesis, the diploid megaspore mothercell and pollen mother cell (PMC), respectively, un-dergo one reductional and equational mitotic divisionto form four haploid daughter cells. During subsequentmale spore development (microgametogenesis), each ofthe four microspores undergoes two mitotic divisions toform a tricellular pollen grain containing two maturereduced gametes (generative nuclei) and one vegetativenucleus.
Despite a relatively simple structure, the malegametophyte is regulated by complex gamete-specific
molecular programs that are characterized by (1) en-richment of transcripts responsible for DNA repair, cellcycle, and chromosome organization, (2) underrep-resentation of the RNA-processing machinery (Pinaet al., 2005; Borges et al., 2008), and (3) developmentalstage-specific transcript sets whose activities varytemporally from the uninuclear microspore toward themature pollen grain (Mascarenhas, 1989; Honys andTwell, 2004).
Loss-of-function studies of male and female meiosishave revealed cell cycle defects (Yang et al., 2003), ef-fects on homologous chromosome recombination (Luet al., 2008), and defects in meiotic cell fate (Sundaresanet al., 1995; Yang et al., 1999), all of which lead to re-duced meiocyte fertility or death. Other mutations leadto unreduced (2C) gametes (Brownfield and Köhler,2011).
Apomeiosis is a naturally occurring form of sensustricto unreduced egg formation, which is the first stepof gametophytic apomixis or asexual (clonal) seedformation in plants (Nogler, 1984; Grimanelli et al.,2001a). During apomixis, the apomeiotically derivedegg cell undergoes parthenogenetic (i.e. without fer-tilization) development, while endosperm developswith (pseudogamous) or without (autonomous) fertili-zation (Nogler, 1984; Grimanelli et al., 2001a). Apomixisbears the potential to fix hybrid genotypes over gener-ations and, therefore, is of high interest to agronomy(Grimanelli et al., 2001a; Grossniklaus, 2001).
1 This work was supported by the Leibniz Institute of Plant Ge-netics and Crop Plant Research and by the European Union COSTaction Harnessing Plant Reproduction for Crop Improvement (grantno. FA0903).
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Timothy F. Sharbel ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OPEN] Articles can be viewed online without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.113.222448
1640 Plant Physiology�, December 2013, Vol. 163, pp. 1640–1659, www.plantphysiol.org � 2013 American Society of Plant Biologists. All Rights Reserved. www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from
Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Apomixis research can be divided into studies ofde novo induction of apomixis-like processes in sexualplants and naturally occurring apomixis (Savidanet al., 2001). Mutant screening and attempts to intro-gress apomixis into sexual taxa have been hindered byits molecular genetic complexity (d’Erfurth et al., 2008;Hörandl and Temsch, 2009; Marimuthu et al., 2011).Comparative mapping strategies in natural apomicts,with an emphasis on aposporic species, have identifiedcandidate factors, although proof of function in cropplants is still pending (Guerin et al., 2000; Albertiniet al., 2004; Matzk et al., 2005; Schallau et al., 2010).Besides theories of apomixis inheritance through asingle dominant locus (Mogie, 1988) or via a complexof physically linked coadapted genes (Van Dijk et al.,1999), the hybridization-derived floral asynchrony(HFA) theory proposes the deregulation of sexual ge-netic pathways via the asynchronous expression ofduplicated gene sets as a mechanism for apomixis in-duction (Carman, 1997). Reproductive deregulationwas recently supported by heterochronic expressionpatterns during ovule development in sexual andapomictic Boechera spp. (Sharbel et al., 2010), pointingto a network of epigenetic and posttranscriptionalregulation during germline specification (Twell, 2011).The approximately 110 species that constitute the
North American genus Boechera (Brassicaceae) exist asdiploid sexual or diploid and allopolyploid apomicticforms (Böcher, 1951; Al-Shehbaz and Windham, 2010),the latter of which are highly correlated with inter-specific hybridization (Sharbel et al., 2009, 2010; Becket al., 2012). Diploid apomixis is rare and facilitatescomparisons between sex and apomixis without theadded complexity of differing ploidy (Sharbel et al.,2009, 2010). Apomictic Boechera taxa are characterizedby Taraxacum-type pseudogamous diplospory (Rollins,1941; Böcher, 1951). Besides the formation of unre-duced female gametes, apomictic Boechera spp. alsoproduce unreduced pollen, as demonstrated by thefact that diploids and triploids produce seeds almostexclusively with hexaploid (6C = [4Cmaternal] + [2Cpaternal])and nonaploid (9C = [6Cmaternal] + [3Cpaternal]) endosperm(Voigt et al., 2007; Aliyu et al., 2010; Voigt-Zielinskiet al., 2012). Hence, strong selection pressure to main-tain a balanced two maternal-to-one paternal genomeratio (i.e. endosperm balance number; Johnston et al.,1980) during endosperm formation apparently charac-terizes apomictic Boechera spp. (Johnston et al., 1980;Voigt et al., 2007; Aliyu et al., 2010). While we have thusfar focused our transcriptome comparisons on sexualand apomictic ovules (Sharbel et al., 2009, 2010), strongselection for genetically balanced endosperm duringapomictic seed formation has led us to study unreducedpollen formation.Hence, we have undertaken a microarray-based
transcriptome comparison of a single stage of anther-head development differentiating reduced and unre-duced pollen formation between diploid sexual anddiploid apomictic Boechera spp. Here, we describe theisolation and characterization of a differentially
expressed candidate gene, BspUPG2 (for Boecheraspecies UPGRADE2 [unreduced pollen grain de-velopment]), which is associated with unreducedpollen formation and is potentially part of a homology-dependent gene-silencing mechanism. BspUPG2 hasapparently arisen through gene duplication and sub-sequent insertions of genic and nongenic fragments inthe hybrid apomictic genome. Its putative molecularfunction in apomictic genomes of Boechera spp. isdiscussed.
RESULTS
Apomictic Boechera spp. Produce (First DivisionRestitution-Type) Unreduced Gametes
One major mechanism that leads to unreducedpollen formation is the meiotic nuclear restitution,which comprises a failure during the first or secondmeiotic division. Meiotic chromosome behavior (Rosset al., 1996) was examined to determine the exactmechanism by which unreduced pollen is formed infacultative and obligate apomictic Boechera spp. andshowed that pollen formation differed between apo-micts and sexuals (Fig. 1; Supplemental Table S1). Thesexual Boechera stricta ES 612.1 showed expected ho-mologous chromosome pairing, with the juncture ofnonsister chromatids leading to seven bivalents (Fig. 1A).The subsequent two nuclear divisions lead to atetrad with four haploid nuclei (Fig. 1, B–G). In con-trast, the aneuploid obligate apomictic Boechera poly-antha (ES 776.2; 2n = 2x = 15) and the euploid Boecheralignifera (ES 753; 2n = 2x = 14) exhibited complete orpartial chromosomal asynapsis, resulting in univalentsat metaphase I that do not (or only partially) segregateduring meiosis I (Fig. 1, I, J, P, and Q). As a result,metaphase II plates were frequently fused (Fig. 1, Kand R), in contrast to their perpendicular orientation inthe sexual reference accession (Fig. 1D), leading to thegeneration of dyads with balanced and unreducedchromosome numbers (Fig. 1, L, M, S, and T). Lowlevels of chromosomal synapsis leading to tetrad for-mation were also observed in all examined diploidobligate apomicts (Fig. 1, N and U). Compared withobligate apomicts, the facultative apomictic Boecheradivaricarpa ES 514 exhibited higher levels of nuclei withbivalents (Fig. 1, W and Y), which proceeded throughboth meiotic cell divisions (Fig. 1A9, C9, and E9), gen-erating tetrads with four haploid cells (Fig. 1G9).Nonetheless, nuclei with univalents were frequentlyobserved (Fig. 1, V, X, and Z) whose sister chromatidswere equally separate at metaphase II (Fig. 1B9) todevelop balanced dyads (Fig. 1, D9 and F9). Meioticchromosome counts of all apomicts strongly supportfirst division restitution without crossover. Univalentsremain together during meiosis I and disjoin in meiosisII, where sister chromatids are equationally separatedto opposite poles, leading to two balanced diploidchromosome sets in dyads.
Plant Physiol. Vol. 163, 2013 1641
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Approximately 28,000 meiocytes from single anther-heads of six apomictic genotypes and one sexual ref-erence genotype were counted to quantify meioticproduct frequencies. The sexual reference genotypeproduced the expected high frequency of tetrads (88%),whereas the tetrad frequency varied among all testedapomicts (0%–87%) between individuals of one geno-type and even between different flower buds from thesame individual (Fig. 2A; Supplemental Table S2). Lowlevels of triads and tetrads were observed for apomicticgenotypes ES 805.2 (0.55% and 0.35%, respectively) andES 776.2 (0.00% and 0.00%, respectively), both of whichshowed high levels of monad and dyad formation.However, surprisingly high levels of triads and tetradswere found for other individuals of both obligate (300.9,24.98% and 57.98%; ES 753, 32.78% and 62.16%) andfacultative (ES 514, 11.22% and 87.00%; ES 524.2,15.67% and 64.53%) apomicts (Fig. 2A). The formationof high levels of balanced dyads confirmed that unre-duced pollen in apomictic Boechera spp. is produced bya defect in chromosome segregation during meiosis I.However, unequal sister chromatid segregation duringmeiosis II, resulting in the formation of extra nuclei, waslow (in total, 0.19% meiocytes with micronuclei and0.02% polyads) and mainly detected in the apomict ES753 (0.88% meiocytes with micronuclei; SupplementalFig. S1).
Considering the highly variable pattern of meiocytefrequencies between individuals and between flowersof single specific apomictic genotypes (Fig. 2A), onlyplants producing exclusively unreduced pollen werechosen for comparative gene expression analyses.Plant selection, therefore, was based upon measure-ments of the nuclear DNA content of pollen and seednuclei for all individuals (Supplemental Fig. S1). In-terestingly, flow cytometric seed screen data of alltested obligate apomictic individuals suggest success-ful fertilization with only unreduced pollen, althoughalso some individuals produce high levels of reducedpollen (embryo:endosperm ratio of 2C [1Cmaternal +1Cpaternal]:6C [4Cmaternal + 2Cpaternal]; Fig. 2A; SupplementalFig. S1; Supplemental Table S3).
Staging of Meiosis in Boechera spp.
To optimize for comparative gene expression analy-ses between reduced (sex) and unreduced (apomeiosis)pollen, the morphological development of antherheadswas quantified through staging (Sharbel et al., 2010).Analyses of early flower development from about 780flower buds in six diploid Boechera spp. genotypes(Figs. 2, B and C, and 3; Table I; Supplemental TableS3) showed a linear relationship between bud length
Figure 1. Meiotic chromosome behavior of male meiocytes. Chromosome spreads of a diploid obligate sexual (A–G), ananeuploid (H–N) and a euploid (O–U) obligate apomictic, and a high facultative apomictic (V–G9) Boechera spp. genotype aredisplayed. Arrowheads show synapsed homologs in sexual (A) and nonsynapsed homologs in apomictic (H and O) Boecheraspp. In addition, homologs with close juxtaposition were occasionally observed in apomictic Boechera spp. (asterisk). Bars = 5 mm.
1642 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

and flower organ size (e.g. for stages S4 to S12antherheads, r2apo = 0.87, F = 1273.50, P, 0.001; r2sex = 0.95,F = 4237.23, P , 0.001). Minor variations amonggenotypes, but no overall differences between sex andapomixis, were detected [i.e. stages predict similarorgan size classes; general linear model for regressionline slopes (b) of stages S4 to S12 antherheads: H0, theeffective null hypothesis which assumes that thereis no relation between two phenomena, bapo 5 bsex;r2 5 0.95, F(44, 8) 5 0.68, P 5 0.804; SupplementalFig. S2].
We focused on four major histodifferentiation stepsof male gametophyte development that are character-ized by their premeiotic, meiotic, and postmeiotic ap-pearance (Fig. 1; Table I). Despite some marker-specificvariations in correlation strength, flower bud lengthpredicts the gametophytic stage of anthers with rela-tively high accuracy (n = 455; sporocyte, 100%, PMC,100%, meiosis, 52.27%, microspores, 58.24%; Fig. 2B).Antherhead length predicts the gametophytic stage ofanther development, except for meiotic and micro-spore stages in sexuals, which overlapped in anther-head size class (one-way ANOVA with Tukey’shonestly significant difference [HSD] post hoc test, be-tween P , 1022 and P , 1029; Fig. 2C). The correlationbetween antherhead length and gametophyte stageled to the selection of 400- 6 30-mm antherheads, astage that is significantly enriched for PMCs, beingclose to meiosis in both sexual and apomictic geno-types, for the collection of total RNA (P , 0.001,Fisher’s exact test; Supplemental Table S4; the deci-sion to take the PMC stage is detailed in “Materialsand Methods”).
Identification of the Apo-Specific UPGRADE Transcript
In search for candidate transcripts that are differ-entially expressed between the sexual and the apo-mictic microsporogenesis programs, we performedpairwise comparisons of RNA expression levels inmicrodissected antherheads from seven sexual andseven apomictic genotypes using a custom Agilent
Figure 2. Meiocyte constitution at the tetrad stage and correlation offlower bud size and antherhead length with different gametophyte
stages. A, Squashes of single antherheads from several individuals pergenotype demonstrate meiocyte constitution at the tetrad stage. Romannumbers denote groups of individuals per genotype primarily (greaterthan 50%) producing reduced gametes (I) versus groups of individualsper genotype primarily (greater than 50%) producing unreducedgametes (II). B, Relationship between flower bud stages S3 to S12 andproportion of anthers at a particular developmental stage of gameto-phytes in Boechera spp. C, Horizontal bars above the box plotsdemonstrate significant comparisons between gametophytic stages perantherhead length within each genotype (*P , 0.05, **P , 0.01,***P , 0.001, n.s. = not significant) as conducted for a 95% binomialproportion confidence interval with a one-way ANOVA includingTukey’s HSD post hoc test. Despite observations of antherhead-relatedgametophyte stages for flower bud stages S3 to S12, the correlationanalysis focuses only on S8 to S12. Me, Meiosis; Msp, microspore;Sp, sporocyte.
Plant Physiol. Vol. 163, 2013 1643
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

microarray (detailed in “Materials and Methods”). Usinga 2-fold or greater change threshold and correctedP # 0.05, while allowing one outlier sample, five micro-array probes were identified to be highly up-regulated inall apomicts, except in genotype ES 753 (B. lignifera),exhibiting absolute fold change levels between 48.74 and849.31 compared with sexuals (Supplemental Fig. S3;Supplemental Table S5).
Homologous complementary DNAs (cDNAs) corre-sponding to the five candidate array probes were iden-tified from a flower-specific cDNA library (SupplementalTable S5; Sharbel et al., 2009). A BLASTN (Altschulet al., 1997) search of these cDNAs against the wholeGenBank nucleotide collection (http://www.ncbi.nlm.nih.gov/genbank/) revealed hits for only two ofthe candidate probes; probe Sharb1199059 (GenBank/EMBL/DNA Data Bank of Japan [DDBJ] accession no.ERS317552) was homologous to an Arabidopsis (Arabi-dopsis thaliana) S-locus lectin protein kinase involvedin pollen recognition, and probe Sharb0501554(GenBank/EMBL/DDBJ accession no. ERS317556)matched two S-LOCUS RECEPTOR KINASE genes(S-12 and S-15 type) encoding a Brassica oleracea
S-locus receptor kinase involved in pollen self-recognitionspecificity (Supplemental Table S5).
Real-time quantitative reverse transcription (qRT)-PCR confirmed that four of the five microarray probeshad expression profiles corresponding to the micro-array data (Supplemental Fig. S4; SupplementalTables S6 and S7). The single probe that could notbe qRT-PCR validated (Sharb1199059; lectin homolog)was furthermore characterized by a negative bacterialartificial chromosome (BAC) screen result (SupplementalTable S8) and was not considered for further analysis.With the exception of the apomictic outlier sample(ES 753), the four microarray probes were highly up-regulated in apomictic antherheads. In contrast tosexuals, in which the relative mRNA levels were closeto the detection limit, ubiquitous expression of all probesequences was detected in apomictic somatic andreproductive tissues. Moreover, for microarray probesSharb0931225, Sharb0501554, and Sharb0690829, ex-pression was substantially higher in antherheads com-pared with the levels in leaf tissue (one-way ANOVAwith Tukey’s HSD post hoc test, P = 0.022, P = 0.008,and P = 0.027, respectively; Supplemental Fig. S4;Supplemental Table S7).
RACE was performed using gene-specific primersfrom microarray probe Sharb0690829 (SupplementalTable S5) to obtain full-length cDNA including the 39and 59 cDNA ends. Although total RNA of anther-heads and whole flower tissue from two sexuals andeight apomicts was used, 59 and 39 RACE generatedsimilar DNA fragments exclusively from apomicts,except for the apomictic outlier ES 753 (SupplementalFig. S5).
All 39 and 59 RACE fragments from five apomictswere cloned and sequenced and led to the identificationof a single polyadenylated full-length transcript, BspUPG(GenBank/EMBL/DDBJ accession no. HF930769; Fig. 4)with length of about 2,648 nucleotides (nt; for B. divaricarpaES 524), excluding the poly(A) sequence. The microarrayprobes Sharb0931225, Sharb0501554, and Sharb0690829were found in the most 39 exon of the BspUPG transcript(Fig. 4). A comparison of different BspUPG full-lengthcDNA variants with genomic DNA by isolation of 39RACE and the various 59 RACE fragments in differentBoechera spp. genotypes (Supplemental Fig. S5, whitearrows) showed that the gene has an overall length of3,156 nt (for B. divaricarpa ES 524) and contains twoputative alternative splicing sites, 61 bp (intron 1) and303 bp (intron 2), in addition to a 144-bp intron com-mon to all apomicts (intron 3; Fig. 4; SupplementalFig. S5). The U2-dependent classified introns (59-GT/39-AG splicing site; Simpson and Filipowicz, 1996) arelocated toward the 59 end of the transcript.
UPGRADE Is a Chimeric Gene Encoding a Transcriptwithout Translational Capacity
BspUPG fragments are homologous to sequences inArabidopsis located on chromosomes 1, 4, and 5,
Figure 3. Light microscopic analysis of Boechera spp. antherheaddevelopment and corresponding gametophyte stages during pollenformation. A, Flower buds and appropriate antherheads of differentdevelopmental stages S3 to S12 in a sexual genotype. B to E, Lightmicroscopy images of semithin sections of resin-embedded anther-heads after histological staining displaying the development of thesporocyte into a mature microspore, used for cytological validation ofmajor gametophyte stages. C, Connective; E, endodermis; En, endo-thecium; MC, meiotic cell; Me, meiosis; Msp, microspores; P, parietalcells; Sp, sporocyte; T, tapetum; V, vascular region. [See online articlefor color version of this figure.]
1644 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

which encode an elongation factor TU/EF1-A protein(EFTU/EF1-A; AT4G02930), an RNA recognitionmotif-containing protein (RNAR; AT5G19960), and theHRD3 protein (for 3-hydroxy-3-methyl glutaryl-CoAreductase degradation), which is homologous to compo-nents of the yeast HRD1 complex (AT1G18260), respec-tively (Fig. 4). From these candidates, AtHRD3, whichplays a central role in endoplasmic reticulum-associatedprotein degradation, exhibits the highest similarityto BspUPG, with fragments of 145 nt (83% identity;P , 5E-31), 118 nt (87% identity; P , 6E-30), and 47 nt(89% identity; P , 0.0002).The transcript carries a poly(A) tail with some var-
iation in length and initiation position. In search forregulatory motifs on BspUPG, a putative near-upstream element (AATAAA) of a polyadenylationsignal, which is common in plants (Hunt, 1994), wasidentified 31 nt upstream of the polyadenylation site.Furthermore, a single CpG island was detected at the59 end of the transcript locus (+140 to 312 nt, C + G .60%; Fig. 4; Supplemental Fig. S6C). Sequences that arerich in the CpG/CpNpG patterns are predominantlynonmethylated and tend to be associated with genesthat are frequently switched on (Deaton and Bird,2011). A screen for putative regulatory features in theupstream sequences (between +47,809 and +48,921nt on Assembly 2, PlantPAN database; Chang et al.,2008; http://plantpan.mbc.nctu.edu.tw/) detected asingle tandem repeat (2726 to 2985 nt, 82-nt con-sensus size, 3.2 copies; Fig. 4) and multiple tran-scription factor-binding sites (Supplemental Table S9).
Coding potential calculation of BspUPG (for ES 524;Kong et al., 2007), excluding the constitutive intron 3,revealed a single short open reading frame (ORF) foreach strand. The sense strand gave an ORF with 196 nt(66 amino acids, +895 to +1,090 nt, coding potentialscore = 20.0298), and the reverse complement strandrevealed a 202-nt ORF (68 amino acids, +1,958 to+2,159 nt, coding potential score = 21.180). The lack ofan ORF length typical for a protein encoding mRNAsuggested a long non-protein-coding mRNA-like RNA(lncRNA) product of BspUPG (ORF typically morethan 100 amino acids; Kondo et al., 2007).
The genomic sequence information of the candidatelocus was augmented by a screen of a B. divaricarpaBAC library. Colony PCR using target-specific primersfrom the chromosome-walking approach (detailed in“Materials and Methods”; Supplemental Table S10) ledto the identification of 12 Boechera spp. BAC clones thathybridized with one or several probes simultaneously(Supplemental Table S8). None of the clones werepositive for probe Sharb1199059, whereas three singlehits for probe Sharb0425060 and nine triple hits forprobes Sharb0931225, Sharb0501554, and Sharb0690829were detected, which confirmed the results from theRACE experiment (Fig. 4). Restriction digests of the 12BACs suggested partial overlap of their DNA inserts.Sanger sequencing was thus performed on BAC clonesA4O22, E7K5, C8B11, and F8G11, each being positivefor three of the five microarray probes (Sharb931225,Sharb501554, and Sharb690829). The C8B11 BAC se-quence contig (Assembly 1; GenBank/EMBL/DDBJ
Table I. Developmental markers of pollen formation corresponding to gametophyte and flower stages
Flower Stage Size RangeArabidopsis
Flower StageaPollen Stageb
Developmental
Marker Identifier
Developmental Marker
and Histological Eventsc
mm
S1 0–0.1 n/ad n/a n/a n/aS2 0.1–0.2 n/a n/a n/a n/aS3 0.2–0.3 7–8 n/a Sp Sporocytes, premeiotic, anther in differentiation stage,
occurrence of 1˚ and 2˚ parietal cells, vascular region initiatedS4 0.3–0.4 9 3 Pattern of anther defined, all four locules presentS5 0.4–0.5 d d Rapid lengthening of all flower organs, especially antherheads
and filamentS6 0.5–0.6S7 0.6–0.7S8 0.7–0.8 PMC PMC enlarged and clearly separated from tapetum, prior to meiosis,
no callose depositionS9 0.8–0.9 4 PMC enters pollen meiosis, crushed middle layer, vacuolated tapetum,
initial callose deposition, binucleate tapetumS10 0.9–1.0 4–6 Me Meiosis, tetrads of microspores, accumulated callose depositionS11 1.0–1.1 10 5–6 Msp Microspores in interphase, early-vacuolated, minorities of tetrads
or binucleate microsporesS12 1.1–1.2 11 6–8 Microspores vacuolated and undergo first mitotic division,
callose degradation, binucleate microspores
aArabidopsis flower developmental stages taken from Smyth et al. (1990). bArabidopsis pollen developmental stages after Regan and Moffatt(1990). cObserved histological events characterized according to Sanders et al. (1999). dn/a, Not applicable.
g
Plant Physiol. Vol. 163, 2013 1645
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

accession no. HF954100), which could not be alignedtogether with the sequence contigs of the other threeBAC clones, reached 57,458 bp in length with 33.5%average GC content, which increased to 40.1% in genicregions. BAC sequence contigs from clones A4O22,F8G11, and E7K5 overlapped and were assembled intothe 58,769-bp Assembly 2 (GenBank/EMBL/DDBJaccession no. HF954101), with 32.9% average GCcontent increasing to 42.1% in genic regions.
Annotation of Assembly 1 identified one transposon-related gene, five protein-encoding genes, and twofragments of protein-encoding genes, all of which(except for TER4) were homologous to genes locatedon Arabidopsis chromosome 1 (Fig. 5A; SupplementalFig. S6, A and B; Supplemental Table S11). In contrast,Assembly 2 contained a higher level of transposon-related genes (three) and gene fragments (nine), whosehomologs were found on all Arabidopsis chromosomesexcept for chromosome 3 (Fig. 5A; Supplemental Fig. S6,A and C; Supplemental Table S11). Comparison of bothassemblies showed that both fragments share severalhighly similar sequences (e.g. for the two protein-encoding genes MtN21 and RRP4, for a fragmentof the protein-encoding gene MBOAT, and for the 39end of BspUPG). Allowing for rearrangements, Assem-bly 1 and Assembly 2 aligned along orthologous regionscovering 27.15 kb of Assembly 1 and 46.20 kb of Assem-bly 2. Synteny gaps in Assembly 2 contained transposon-related genes (e.g. TER1 and TER2) and insertionscomposed of short protein-encoding gene fragmentsflanked by inverted repeat sequences (e.g. IR7; Fig. 5A;Supplemental Fig. S6C; Supplemental Table S12),suggesting that parts of Assembly 2 arose via partialduplications of Assembly 1. Hence, considering thepartial presence of the candidate gene at both loci, wesubsequently labeled the original locus in Assembly 1BspUPG1 and the duplicated variant on Assembly 2BspUPG2 (Fig. 5A; Supplemental Fig. S6).
Both apomictic and sexual gene variants were alignedwith Assembly 1 and Assembly 2 (i.e. BAC DNA se-quences), allowing for rearrangements. The 39 end of allgene variants from sexuals and apomicts, here namedlocally collinear block (LCB) 1 (LCB1 in violet; see defi-nition in “Materials and Methods”), shares high simi-larity with both assemblies. The 59 ends of apomicticgene copies (LCB2 in brown) were only identified onAssembly 2 (Fig. 5, A and B; Supplemental Fig. S6).
Unlike apomictic gene variants (now BspUPG2; see no-menclature above), the 59 ends (LCB3–LCB5) of variantsfrom sexuals (now BspUPG1) share high sequence simi-larity with Assembly 1 but also exhibit a mosaic distri-bution of their LCBs relative to the duplicated locus ofAssembly 2 (Supplemental Figs. S6 and S7).
Transcriptional Activity of BspUPG2 Is Restrictedto Apomictic Boechera spp.
The transcriptional functionality of BspUPG1 andBspUPG2 was inferred by mapping sexual and apo-mictic cDNAs (Sharbel et al., 2009) independently ontoboth and revealed that only the apo-specific LCB2 istranscribed in both sexuals and apomicts (see sexual/apomictic cDNA reads; Fig. 5C). In contrast, apomicticcDNA maps to LCB1 and LCB2, including the readswith homology to the candidate microarray 60-meroligonucleotide probes (Sharb931225, Sharb501554,and Sharb690829; Fig. 5C). No cDNA maps to any ofthe remaining LCBs. PCR on genomic DNA usingspecific primers for both loci demonstrates the pres-ence of BspUPG1 in sexuals and apomicts but absenceusing cDNA as a PCR template. In contrast, genomiccopies and transcripts of BspUPG2 were solely identi-fied in apomicts (Fig. 5, D and E).
BspUPG1 is flanked by four genes centered in thecentromeric region on the lower arm of Arabidopsischromosome 1 (Fig. 5A; Supplemental Fig. S8;Supplemental Table S11; approximately 0.57-Mb dis-tance from T3P8-sp6; Hosouchi et al., 2002). Interest-ingly, two genetic markers (Bst006701 and BstES0032)from the synthetic F2 linkage map of sexual B. stricta(Schranz et al., 2007), both of which are homologous tothe Arabidopsis locus identifiers AT1G51310 andAT1G43245, flank BspUPG1. Both markers span theinterval of genomic block C1, which localizes BspUPG1on the BstLG1 linkage group of B. stricta, for whichvery low levels of recombination were detected(Schranz et al., 2007). The putative location of the du-plicated BspUPG2 is unknown.
BspUPG2 Has High Secondary Structure Folding Potential
Apart from other lncRNA types in plants, somelncRNAs act as precursors for small non-protein-coding RNAs (npcRNAs), such as microRNAs
Figure 4. Schematic representation ofputative splicing forms of BspUPG2.All four splice forms could be extractedby end-to-end sequencing in five apo-mictic Boechera spp. Exons are repre-sented by gray boxes, and numbersindicate intron-exon boundary posi-tions. BLAST search results from theGenBank nucleotide collection and theTAIR10 gene database are given below.
1646 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
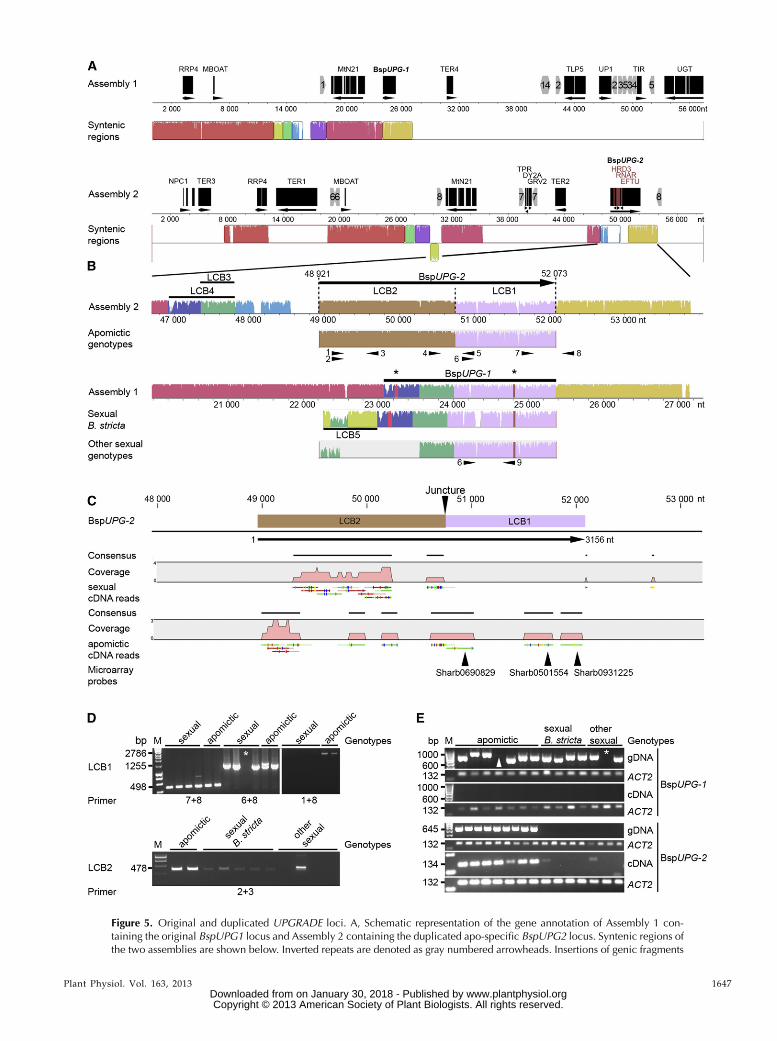
Figure 5. Original and duplicated UPGRADE loci. A, Schematic representation of the gene annotation of Assembly 1 con-taining the original BspUPG1 locus and Assembly 2 containing the duplicated apo-specific BspUPG2 locus. Syntenic regions ofthe two assemblies are shown below. Inverted repeats are denoted as gray numbered arrowheads. Insertions of genic fragments
Plant Physiol. Vol. 163, 2013 1647 www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from
Copyright © 2013 American Society of Plant Biologists. All rights reserved.

(miRNAs) or endogenous trans-acting small interfer-ing RNAs, and may regulate translation in cis or trans(Hirsch et al., 2006; Pikaard et al., 2008). A search formiRNA-binding sites of known Arabidopsis (www.mirbase.org; default parameter, one mismatch) andBoechera spp. miRNAs (Amiteye et al., 2011, 2013)revealed no mature miRNAs matching the candidatetranscript. Considering this, in addition to the obviouslack of a long ORF on BspUPG2, a bioinformatics ap-proach was used to examine whether thermodynamicallystabile npcRNA structural elements could be predictedfrom its genomic sequence (detailed in SupplementalProtocol S1).
As shown in Figure 6A, the BspUPG forward andreverse strands contain eight regions (i.e. overlappingwindows are combined to determine the total length ofa candidate region; Supplemental Table S13) charac-terized by significant Z-score values, suggesting thatparts of BspUPG are able to form stable secondarystructures (Fig. 6A; Supplemental Fig. S9). These can-didate regions span only exons, comprising together25% and 27% of the total length of each strand ofBspUPG, whereby npcRNAs 1/6, 2/7, and 4/8 coversimilar regions on both strands of the genomic se-quence. The predicted secondary structures were clas-sified based upon their (1) minimal folding free energy,(2) adjusted minimal folding free energy, (3) minimalfolding free energy index, and (4) A + U content (Seffensand Digby, 1999; Bonnet et al., 2004; Zhang et al., 2006).Interestingly, all detected secondary structures metmost of the criteria for miRNA precursors (Fig. 6B;Supplemental Fig. S9; Supplemental Table S14). Com-parable with known primary miRNAs (pri-miRNAs) inBoechera spp. (Amiteye et al., 2011, 2013) and other plantspecies (Zhang et al., 2006), npcRNAs 1 to 8 demon-strate a lower minimal folding free energy (rangingfrom 218.27 to 273.72 kcal mol21), have an elevated
A + U level (ranging from 56.70% to 70.00%), and, withthe exception of npcRNA 5 (0.70), have a minimalfolding free energy index greater than 1.07, which ismuch higher compared with tRNA (0.64), ribosomalRNA (0.59), or random mRNA (0.65).
A BLASTN search of the GenBank nucleotide andThe Arabidopsis Information Resource 10 (TAIR10)genes databases showed that three npcRNAs are sig-nificantly similar to known protein-coding genes (i.e.potential regulatory targets). Two of these potentialtargets, npcRNAs 5 and 8, encode for proteins withribosomal functions. The 70-nt npcRNA 5 (position+1,090 nt on the forward strand and +1,160 nt on thereverse strand of BspUPG) covers 75% of the third exonof the EFTU/EF-1A protein homolog in Arabidopsis,which binds tRNAs in a GTP-dependent reaction tothe acceptor site of ribosomes (AT1G18260; 92.86%similarity; P = 7.00E-24; Fig. 6, B and C; SupplementalTable S14). A fragment of npcRNA 8 maps to a ho-molog in Arabidopsis lyrata and Arabidopsis that isassociated with a U3 small nucleolar RNA involvedin the processing of pre-ribosomal RNA (AT3G06530;P = 0.008). The third, npcRNA 3, maps to exon 25of the myosin XI B protein-coding gene (AT1G04160;P = 0.01), which belongs to a class of myosins poten-tially involved in the process of spindle/phragmoplastalignment during cytokinesis (Hepler et al., 2002) andwhose mutation led to abnormal pollen developmentin rice (Oryza sativa), suggesting a crucial role of myosinXI B in pollen formation (Jiang et al., 2007).
BspUPG2 Is Highly Conserved in Apomictic Boechera spp.
Within-individual (i.e. allelic) polymorphisms werenot identified for BspUPG2 among nine tested apo-micts. Furthermore, both the original locus (BspUPG1)
Figure 5. (Continued.)are marked in red letters. RRP4, Exosome complex component RRP4; MBOAT, membrane-bound O-acyl transferase-likeprotein; MtN21, nodulin MtN21/EamA-like transporter protein; BspUPG1, original locus of candidate geneUPGRADE; TER1-4,transposable element-related protein; TLP5, Tubby-like F-box protein5; UP1, uncharacterized protein; TIR, Toll/interleukin1receptor-nucleotide binding site class of disease resistance protein; UGT, sterol 3b-glucosyltransferase; NPC1, Niemann-PickC1 protein; TPR, tetratricopeptide repeat domain-containing protein; DY2A, dynamin2A; GRV2, DNAJ heat-shock N-terminaldomain-containing protein; HRD3, HRD3-like protein; BspUPG2; duplicated locus of UPGRADE. B, Mapping of genomicsequences of BspUPG1 in apomictic (e.g. B. divaricarpa, ES 514) and sexual (e.g. B. stricta, ES 612.1) genotypes onto both BACclone assemblies identified by rearrangements of LCBs. Black asterisks denote indels for the sex-specific identity of LCB1 andLCB4. The LCB5 position on Assembly 1 and Assembly 2 is displayed in Supplemental Figure S5. Black arrowheads denoteprimers 1 (CON234X2L), 2 (CON234X14L), 3 (CON234X10R), 4 (PC1pol1L), 5 (PC1pol1R), 6 (GSP4), 7 (TSP33R),8 (CON234X5R), and 9 (Indel9minus). C, CLC Genomics Workbench (version 4.5.1) output file showing different distributionsof separate mapped cDNA reads from sexual and apomictic genotypes onto BspUPG2. Both sense-oriented (green) and an-tisense-oriented (red) cDNA reads are displayed below gray regions that demonstrate cDNA coverage (in pink). Colored verticalbars on cDNA reads show single-nucleotide polymorphisms in comparison with genomic DNA of BspUPG2. D, PCR withindependent primer pairs, one located on LCB1 and one on the cDNA mapping positions of LCB2 in sexual and apomicticgenomes illustrate that LCB1 is highly conserved in both sexuals and apomicts and that LCB2 is also present in sexual genotypesbut in a different position in the genome. Primer locations are shown as numbered black arrows in B. The white asterisk denotesa missing band for genotype 105.18, which lacks the priming site for primer 6. E, PCR with primers (4 and 5) combining LCB1and LCB2 shows the apo-specific presence of BspUPG2, whereas BspUPG1 is present in both sexual and apomictic genomes(primers 6 and 9). In contrast to BspUPG1, which is not transcribed in sexuals and apomicts (primers 6 and 9), BspUPG2 issolely transcribed from apomicts (primers CON234B2L and CON234B2R; Supplemental Table S6). The white arrowhead marks a faintband for genotype ES 753, and the white asterisk marks a missing band for genotype 105.18, which lacks the priming site for primer 6.
1648 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Figure 6. Secondary structure prediction for BspUPG2. A, The Z-score for the sliding window with length between 50 and300 nt (step size = 10) is plotted versus the position on BspUPG2. Any window producing a significant Z-score during thescanning process was considered a candidate region for a structural npcRNA. If multiple, overlapping windows of severallengths produced significant Z-scores, the region encompassed by all the overlapping windows defined the candidate region
Plant Physiol. Vol. 163, 2013 1649 www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from
Copyright © 2013 American Society of Plant Biologists. All rights reserved.

and its duplicated variant (BspUPG2) are highly con-served at the 39 end between all tested genotypes,which decreases markedly toward the 59 end (Fig. 7A;Supplemental Table S15). Sequence conservation ishigh between all apomicts (i.e. BspUPG2), includingthe transcriptional outlier ES 753, with the highestnucleotide divergence between genotypes ES 753 andES 514 (99.33% 6 0.13% similarity), while BspUPG2 inall other apomicts shares complete sequence identity(Fig. 7A). In contrast, sexual sequences can be split intotwo subgroups based on low sequence conservation atthe 59 end of BspUPG1 (39.95% 6 0.02% similarity),one representing B. stricta genotypes (99.69% 6 0.05%similarity between B. stricta genotypes) and the secondrepresenting the remaining sexual genotypes (98.87% 60.30% similarity between non-B. stricta genotypes;Fig. 7A; Supplemental Table S15).
Among all insertions/deletions (indels) that weredetected between BspUPG1 and BspUPG2 (less than 50nucleotides; Albers et al., 2011; Supplemental TableS16), a single 27-nt sequence was exclusively present inBspUPG1 but not in BspUPG2 (Fig. 7A, indel 9). In-terestingly, Chellappan et al. (2010) identified a new27-nt small RNA species that is associated with AGO4to regulate gene expression at the transcriptional levelby directing DNA methylation to some of their targetloci in trans. Hence, indel 9 was tested as a small RNA-binding site by northern-blot screening of small RNAsfrom sexual and apomictic pooled flower buds using spe-cific sense and antisense probes (detailed in SupplementalProtocol S2; Supplemental Fig. S10). Expression of thehighly conserved plant miRNA167, which was usedas a positive control, was observed in all flower tissues,whereas no signal was detected for any indel 9-specificprobe in both apomictic and sexual flower tissues(Supplemental Fig. S10).
Together, the lack of intraindividual allelic variationand the absence of BspUPG2 at any other locus in bothsexuals and apomicts (Fig. 5) point to a homozygousor hemizygous state for BspUPG2 in apomicts.
BspUPG2 Arose via Genome Rearrangements
Two lines of evidence were used to validate theproposed fusion of the 59 (LCB1) and 39 (LCB2) endsof the transcriptionally active BspUPG2. A BLASTNsearch between BspUPG2 and the complete genomicsequence-read archive of a sexual B. stricta identified agap at the juncture between all reads mapping to LCB1and LCB2, while all reads per LCB overlapped, sug-gesting their separate genomic origins in the sexual
genotype (position +1,820 nt; Fig. 7C). We confirmedthis result by amplifying a 478-bp fragment in sexualgenotypes using primers corresponding to a fragmentof the BspUPG2-specific LCB2. Additionally, this frag-ment is mapped in sexual and apomictic Boechera spp.by cDNA belonging to the Boechera spp. homolog ofAtHRD3 (BspHRD3; Figs. 4 and 7B).
BspHRD3 was used to test whether the 59 end ofBspUPG2 derived directly from parental genes or fromputative duplicated variants. Although BspHRD3cDNA is conserved with Arabidopsis HRD3 (93.0%),variation (i.e. insertions) between the transcript andthe detected genomic copy, which are not caused byRNA splicing, was identified (Fig. 7B). This wouldpoint to two different genomic variants of BspHRD3:(1) one from which the transcript could be detected butnot its genomic copy and that lacks the insertions, and(2) a second variant from which the genomic copycould be sequenced but not the transcript and thatincludes the inserts (Fig. 7B). Interestingly, comparingthe genomic sequence of BspHRD3 with BspUPG2, a550-bp fragment, identified it as being homologous toBspUPG2, including these insertions (93.6% similarity;Fig. 7B, blue boxes denote insertions), thus pointing tothe integration of a fragment of one of the two genomicvariants of BspHRD3 into BspUPG2.
As gene duplication is strongly correlated with in-sertions and pseudogenization (Ohno, 1970; Kaessmann,2010), we tested for duplication by mapping a setof sequence tags from an array-based comparativegenome hybridization experiment in sexual and apo-mictic Boechera spp. (Aliyu et al., 2013) againstBspUPG2 (Fig. 7C). In total, 79 array probes from thecomparative genome hybridization experiment map-ped to BspUPG2. Whereas most probes mapping ontoBspUPG2 show no copy number variations (CNVs) inapomicts (96%, with the remaining 4% showing de-pletion), 68% of the tags were depleted in at least onesexual genotype. Approximately half of the depletedsequence tags in sexual genotypes were situated at the39 end of BspUPG2, representing the start of LCB1. Theremaining depleted sequence tags were distributedtoward the 59 end of BspUPG2, between +0 and +1,081nt. To summarize, both LCB1 and the extreme 59 endof BspUPG2 are present in fewer copies in sexualcompared with apomictic genomes, whereas the mid-dle part of BspUPG2 shows no variation in copynumber in either reproductive mode. The duplicationof LCB1 in apomictic genomes is explained by its pres-ence in both assemblies. The duplication of the 59 end inapomictic genomes is evidenced by the presence of
Figure 6. (Continued.)(represented as boxes). Numbers in the boxes are putative npcRNAs having the most negative Z-scores (Supplemental Fig. S8;Supplemental Tables S13 and S14). B, Proposed minimum free energy structures of putative npcRNAs with significant BLASTNhits in the GenBank and TAIR10 genes databases. The structures above are colored by base-pairing probabilities from 0%(violet; see color scale) to 100% (red). For unpaired regions, the color denotes the probability of being unpaired. C, Overlap ofnpcRNA 5 with exon 3 of EFTU/EF-1A (AT4G02930). Red letters indicate dissimilarities between sequences.
1650 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

duplicated source gene insertions covering almost ex-actly the region represented by depleted sequence tagsin sexuals (HRD3, +199 to +731 nt; RNAR, +732 to+948 nt; EFTU, +1,041 to +1,191 nt; Figs. 5 and 7C).Since transposable elements (TEs) are considered to
drive genome rearrangements, including duplications,after interspecific hybridization, Assembly 1 andAssembly 2 were screened for Viridiplantae transpos-able elements. In total, 100 and 113 TEs mapped ontoAssembly 1 and Assembly 2, respectively, many ofwhich are simple repeats with low sequence complex-ity. Excluding all simple repeats left 68 and 73 TEs thatmapped onto Assembly 1 and Assembly 2, respectively,with about half of them identified in Arabidopsis
(27 and 35, respectively; Supplemental Fig. S11). The majorrepeat families are copia-like (18 and 16) and gypsy-like(8 and 9) long terminal repeat (LTR) retrotransposons,followed by DNA transposons En-Spm (9 and 12),MuDR (11 and 7), hAT (7 and 7), and Helitrons (6 and 9).Proportions of the TE superfamilies do not vary sig-nificantly between Assembly 1 and Assembly 2 but dowhen compared with their distribution across the wholeArabidopsis genome. Copia-like LTR retrotransposonsrepresent the largest proportion of Arabidopsis TEs(57.70%) and those on the Boechera spp. BAC clone as-semblies (26.47% and 21.92%, respectively). Nonetheless,copia-like LTR retrotransposons are more prominentacross the whole Arabidopsis genome compared with
Figure 7. Genesis of BspUPG2 via duplication from the original locus BspUPG1 and sequential insertion of genic fragments.A and B, Structural overview of full-length BspUPG2 in apomictic and sexual Boechera spp. (Table I), adapted and modifiedfrom the progressive Mauve algorithm, exhibits LCBs between the candidate genes in various genotypes (A) and betweenBspUPG2 and the source genes from which BspUPG2 hosts fragments (B). Identical color displays highly similar sequences;black zigzag lines denote truncated sequences due technical reasons. gDNA, Genomic DNA. C, Distribution of 454 whole-genome sequencing (WGS) reads mapping across sliding windows (bins) of BspUPG2 versus CNV in 10 sexual versus 10apomictic genotypes (Aliyu et al., 2013; for details, see “Materials and Methods”; note that “genic inserts” refers to positions+199 to +1,191 nt in B and that whole-genome sequencing reads did not overlap between LCB1 and LCB2). D, Schematicrepresentation of parental and chimeric BspUPG genes. Colored boxes illustrate different LCBs. +, Transcription activity; 2, notranscription activity.
Plant Physiol. Vol. 163, 2013 1651
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

both Boechera spp. assemblies [one-tailed Fisher’s exacttest; LTR/Copia: P(Assembly1) = 1.83E-07, P(Assembly2) =5.23E-10], in contrast to DNA transposons of the En-Spm and hAT class and non-LTR retrotransposons ofthe LINE/L1 class, which are more prominent on bothBoechera spp. assemblies [one-tailed Fisher’s exact test;DNA/En-Spm: P(Assembly1) = 4.21E-10, P(Assembly2) =3.83E-14; DNA/hAT: P(Assembly1) = 2.00E-06, P(Assembly2) = 3.24E-06; LINE/L1: P(Assembly1) =0.001, P(Assembly2) = 0.008; Supplemental Fig. S11;Supplemental Tables S17 and S18].
Together, these results (Figs. 5 and 7) indicate aduplication of BspUPG1 fragments from the originallocus on Assembly 1 to form the basis of BspUPG2,which subsequently underwent sequential insertionsof genome fragments derived from at least two un-linked genomic regions. Thereby, LCB1 was insertedbetween LCB2 and LCB4. The newly formed locus wasthen the insertion target for the duplicated variant of atleast one functional gene (e.g. BspHRD3), with subse-quent exonization leading to a gain of BspUPG2 tran-scriptional activity (Fig. 7D).
DISCUSSION
High Quantitative Variation for Unreduced PollenFormation in Apomictic Boechera spp.
High variability in pollen morphology and unre-duced pollen formation, in addition to tolerance todeviations from the sexual endosperm balance num-ber, have been described for some Boechera taxa(Böcher, 1951, 1954; Voigt et al., 2007; Aliyu et al., 2010;Voigt-Zielinski et al., 2012). Despite this variability,castration experiments (Böcher, 1951) and extensiveflow cytometric analyses of seeds (Aliyu et al., 2010)strongly support selection pressure for the mainte-nance of unreduced pollen development to fulfill en-dosperm balance requirements in diploid apomicts.
Here, we have performed detailed quantitative analy-ses of anther growth and microsporogenesis to identifythe optimal developmental stage for comparative ex-pression profiling of reduced (sexual) and unreduced(apomictic) pollen formation in 14 genotypes. A strongcorrelation between microsporogenesis and anthergrowth was evident at the premeiotic and postmeioticstages, whereas it was difficult to identify antherheadlengths corresponding to the meiotic stage (Fig. 2B).Considering the spatial and temporal variability of ex-pression profiles in reproductive tissues (Mascarenhas,1989; Honys and Twell, 2004), the identification of aspecific stage of antherhead development and lengthcharacterized by PMCs at the onset of meiosis enabledtargeted expression profiling of the meiotic stage, whichdifferentiates reduced (sexual) and unreduced (apomei-otic) pollen formation (Supplemental Table S4). Variablelevels of apomeiosis frequencies have been reported forBoechera spp. (Aliyu et al., 2010), and here, both obligateand highly facultative apomicts were analyzed.No evidence for sex- or apomixis-specific flower
morphological variation was found, although meiosisand the appearance of microspores are develop-mentally uncoupled in apomicts relative to sexuals(Fig. 2A; Supplemental Fig. S1). This latter observationsupports the HFA theory, which describes the tem-poral shift (heterochrony) between both reproduc-tive modes (Carman, 1997), which has also beenidentified on the transcriptomic level (Sharbel et al.,2010).
The meiotic chromosome behavior of apomicts pro-ducing unreduced pollen (the same used for expressionprofiling) was primarily asynaptic (Fig. 1), as has pre-viously been shown (asyndetic; Böcher, 1951). Althoughultimately producing higher levels of dyads comparedwith sexuals, the same apomicts demonstrated varia-bility in terms of monad, dyad, triad, and tetrad for-mation (Fig. 2A), mirroring reported genotype-specificvariability for meiotic chromosome synapsis potentialin both diploid and polyploid Boechera spp. (Böcher,1951; Naumova, 2001; Kantama et al., 2007). Interest-ingly, variation between individuals of the same clonallineage for dyad and tetrad formation was also ap-parent in the majority of tested apomictic genotypes(Fig. 2A); hence, phenotypic variability for pollen for-mation exists despite genetically clonal reproduction. Inthis light, the observed sexual allopolyploid progenyfrom both diploid and triploid obligate apomicts (e.g.ES 753, B. divaricarpa; Schranz et al., 2005) could beexplained by fertilization with reduced self-pollen ratherthan with pollen from another plant (Aliyu et al., 2010),since the genus Boechera is a highly selfing system (Roy,1995). On a broader scale, our data indicate that fixationof the genotype via apomixis does not necessarily lead tophenotypic stability of male meiocytes.
While these data are consistent with previous work(Kantama et al., 2007; Voigt et al., 2007), the lack of cor-relation with apomeiosis expression (e.g. egg cell forma-tion as measured by flow cytometric seed screen;Supplemental Fig. S1; Supplemental Table S1) suggestsseparate mechanisms leading to unreduced male versusfemale gametes in Boechera spp. and supports inde-pendent genetic control for at least some of the de-velopmental steps required to form apomictic seeds(Van Dijk et al., 1999; Noyes and Rieseberg, 2000;Matzk et al., 2005).
The analyses of seed production, anther develop-ment, meiosis, and pollen formation have shown that(1) unreduced pollen formation occurs via first divi-sion restitution in apomictic Boechera spp., (2) thismechanism is not fully penetrant, and (3) selectionpressure for a balanced endosperm in apomictic Boe-chera spp. leads to the almost exclusive contribution ofunreduced pollen to endosperm formation.
BspUPG2, Unreduced Pollen, and Stable Apomixisin Boechera spp.
The detailed phenotypic analyses and selection of aspecific antherhead stage that differentiated reduced
1652 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.
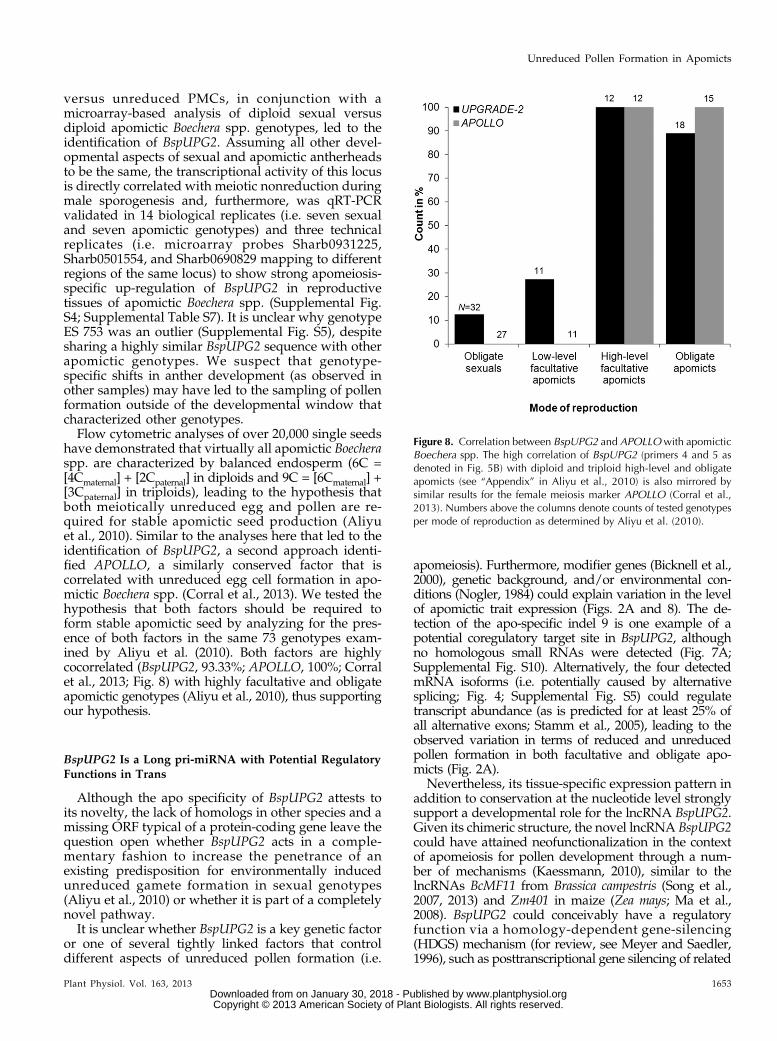
versus unreduced PMCs, in conjunction with amicroarray-based analysis of diploid sexual versusdiploid apomictic Boechera spp. genotypes, led to theidentification of BspUPG2. Assuming all other devel-opmental aspects of sexual and apomictic antherheadsto be the same, the transcriptional activity of this locusis directly correlated with meiotic nonreduction duringmale sporogenesis and, furthermore, was qRT-PCRvalidated in 14 biological replicates (i.e. seven sexualand seven apomictic genotypes) and three technicalreplicates (i.e. microarray probes Sharb0931225,Sharb0501554, and Sharb0690829 mapping to differentregions of the same locus) to show strong apomeiosis-specific up-regulation of BspUPG2 in reproductivetissues of apomictic Boechera spp. (Supplemental Fig.S4; Supplemental Table S7). It is unclear why genotypeES 753 was an outlier (Supplemental Fig. S5), despitesharing a highly similar BspUPG2 sequence with otherapomictic genotypes. We suspect that genotype-specific shifts in anther development (as observed inother samples) may have led to the sampling of pollenformation outside of the developmental window thatcharacterized other genotypes.Flow cytometric analyses of over 20,000 single seeds
have demonstrated that virtually all apomictic Boecheraspp. are characterized by balanced endosperm (6C =[4Cmaternal] + [2Cpaternal] in diploids and 9C = [6Cmaternal] +[3Cpaternal] in triploids), leading to the hypothesis thatboth meiotically unreduced egg and pollen are re-quired for stable apomictic seed production (Aliyuet al., 2010). Similar to the analyses here that led to theidentification of BspUPG2, a second approach identi-fied APOLLO, a similarly conserved factor that iscorrelated with unreduced egg cell formation in apo-mictic Boechera spp. (Corral et al., 2013). We tested thehypothesis that both factors should be required toform stable apomictic seed by analyzing for the pres-ence of both factors in the same 73 genotypes exam-ined by Aliyu et al. (2010). Both factors are highlycocorrelated (BspUPG2, 93.33%; APOLLO, 100%; Corralet al., 2013; Fig. 8) with highly facultative and obligateapomictic genotypes (Aliyu et al., 2010), thus supportingour hypothesis.
BspUPG2 Is a Long pri-miRNA with Potential RegulatoryFunctions in Trans
Although the apo specificity of BspUPG2 attests toits novelty, the lack of homologs in other species and amissing ORF typical of a protein-coding gene leave thequestion open whether BspUPG2 acts in a comple-mentary fashion to increase the penetrance of anexisting predisposition for environmentally inducedunreduced gamete formation in sexual genotypes(Aliyu et al., 2010) or whether it is part of a completelynovel pathway.It is unclear whether BspUPG2 is a key genetic factor
or one of several tightly linked factors that controldifferent aspects of unreduced pollen formation (i.e.
apomeiosis). Furthermore, modifier genes (Bicknell et al.,2000), genetic background, and/or environmental con-ditions (Nogler, 1984) could explain variation in the levelof apomictic trait expression (Figs. 2A and 8). The de-tection of the apo-specific indel 9 is one example of apotential coregulatory target site in BspUPG2, althoughno homologous small RNAs were detected (Fig. 7A;Supplemental Fig. S10). Alternatively, the four detectedmRNA isoforms (i.e. potentially caused by alternativesplicing; Fig. 4; Supplemental Fig. S5) could regulatetranscript abundance (as is predicted for at least 25% ofall alternative exons; Stamm et al., 2005), leading to theobserved variation in terms of reduced and unreducedpollen formation in both facultative and obligate apo-micts (Fig. 2A).
Nevertheless, its tissue-specific expression pattern inaddition to conservation at the nucleotide level stronglysupport a developmental role for the lncRNA BspUPG2.Given its chimeric structure, the novel lncRNA BspUPG2could have attained neofunctionalization in the contextof apomeiosis for pollen development through a num-ber of mechanisms (Kaessmann, 2010), similar to thelncRNAs BcMF11 from Brassica campestris (Song et al.,2007, 2013) and Zm401 in maize (Zea mays; Ma et al.,2008). BspUPG2 could conceivably have a regulatoryfunction via a homology-dependent gene-silencing(HDGS) mechanism (for review, see Meyer and Saedler,1996), such as posttranscriptional gene silencing of related
Figure 8. Correlation between BspUPG2 and APOLLOwith apomicticBoechera spp. The high correlation of BspUPG2 (primers 4 and 5 asdenoted in Fig. 5B) with diploid and triploid high-level and obligateapomicts (see “Appendix” in Aliyu et al., 2010) is also mirrored bysimilar results for the female meiosis marker APOLLO (Corral et al.,2013). Numbers above the columns denote counts of tested genotypesper mode of reproduction as determined by Aliyu et al. (2010).
Plant Physiol. Vol. 163, 2013 1653
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

genes in trans (Eamens et al., 2008), which has been foundfor chimeric Helitron and Pack-MULE RNAs in maize(Jiang et al., 2004; Morgante et al., 2005), or via the mod-ulation of DNAmethylation patterns, such as reported forlncRNA-like loci that are associated with polycombcomponents and histone modifications (De Lucia andDean, 2011).
Consistent with this are the sequence fragments ho-mologous to three different functional parental genesacross the 59 end of BspUPG2, which putatively couldserve as targets for HDGS (BspHRD3, RNAR, and EFTU/EF-1A; Figs. 4, 5A, and 7B). Thereby, BspUPG2 couldbelong to a novel class of miRNA-containing lncRNAsthat serve as a matrix (i.e. as long pri-miRNA) for severalunidentified precursor miRNAs (premiRNAs; Altuviaet al., 2005) or endogenous trans-acting short interferingRNAs. Such regulatory elements are processed from theexcised intronic region of a spliced transcript, or from thefull-length unspliced transcript, and have been detectedin vegetative developmental pathways (Peragine et al.,2004; Vazquez et al., 2004; Hirsch et al., 2006) as well asin generative tissues (e.g. in mature pollen; Grant-Downtonet al., 2009b).
Computational analysis of RNA folding detectedseveral sections of BspUPG2 that were predicted toform with high probability nonrandom and stablesecondary structures (Fig. 6, A and B; SupplementalFig. S8; Supplemental Table S13) and that fulfill mostcriteria for premiRNAs (e.g. elevated A + U content;Supplemental Table S14). Interestingly, these potentialpremiRNAs were not detected in previous screens(Amiteye et al., 2011, 2013), which may reflect (1) itstissue-specific and short-term up-regulation at theonset of male meiosis or (2) limitations of homology-based analyses considering that BspUPG2 is unique toapomictic Boechera spp. The detection of a stable sec-ondary structure in a region that is homologous to aknown protein-coding gene with translation elonga-tion activities during polypeptide synthesis at theribosome and activities in signal transduction (i.e.npcRNA 5 similar to GTP-binding EFTU/EF-1A fam-ily protein; E = 7.00E-24; GO:0003746, AT4G02930)could be a first indication for a HDGS function ofBspUPG2. For example, a homologous factor to EFTU/EF-1A has been implicated with meiosis progression inXenopus laevis (Bellé et al., 1990; Peters et al., 1995),while one of the other two sequence fragments of the59 end of BspUPG2 is also homologous to a gene withnucleotide-binding function (RNAR; GO:0003723).
High Sequence Conservation and Hemizygous orHomozygous Status Imply a Selective Advantageof BspUPG2
Importantly, BspUPG2 is a chimera of nongenic (e.g.LCB1) and genic (e.g. HDR3, RNAR, EFTU) regions inwhich the genic regions were derived from partial du-plicated variants of their parental genes (e.g. BspHRD3,homologous to AT1G18260; i.e. a pseudogene derived
through duplication; Ohno, 1972; Fig. 7, C and D). Al-though the long-term survival of chimeric genes is de-scribed to be rare (Bennetzen, 2005), BspUPG2 exhibitedan unexpectedly high degree of conservation in DNAsequence (Fig. 7A; Supplemental Table S15) and ex-pression (Supplemental Fig. S4) between apomicticBoechera spp. representing different taxa and geographicorigins. In contrast, higher sequence variation wasidentified for BspUPG1 in sexuals (e.g. 59 end; Fig. 7A;Supplemental Table S15). Thus, BspUPG2 appears to beunder selective maintenance (Casillas et al., 2007),which would be consistent with its assumed role inunreduced pollen formation for balanced endosperm inapomicts (Aliyu et al., 2010).
The hemizygous or homozygous status of BspUPG2reflects dominant inheritance as a proposed character-istic for an apomeiosis-controlling locus (Grossniklauset al., 2001; Table I), as has been found, for example, inPennisetum squamulatum (Ozias-Akins et al., 1998).Hemizygosity would reflect an extended period of lowto no recombination in its genomic location and wouldbe consistent with the distribution of TEs at both theoriginal and duplicated loci (Supplemental Fig. S11).The similar abundance of TEs from gypsy-type andEn-Spm superfamilies, which insert preferentially intogene-poor regions (i.e. heterochromatic regions; Fiston-Lavier et al., 2012) and which are enriched in both as-semblies compared with their abundance within thetotal number of Arabidopsis TEs (Supplemental Fig. S11),points to a colocalization of the original and duplicatedloci of BspUPG in the same pericentromeric region(Supplemental Fig. S9) or to regions with similar char-acteristics (i.e. interstitial heterochromatic regions com-parable to hk4S in Arabidopsis; Fransz et al., 2000).
We hypothesize that the genesis of BspUPG2 is as-sociated with the recurrent interspecific hybridizationthat is highly correlated with the origins of apomicticBoechera spp. (Schranz et al., 2005; Beck et al., 2012).Assuming colocalization of BspUPG2 with BspUPG1and the formation of highly heterochromatic regions inapomicts as protected zones (e.g. Het and Del chro-mosomes; Kantama et al., 2007), the genome-wideeffects of hybridization (Comai, 2000) could haveprovided the mechanism through which BspUPG2originated and gained regulatory function.
To summarize, despite variability for unreducedpollen formation in apomictic Boechera spp., a singlenovel transcription unit (BspUPG2) is consistently up-regulated in apomictic flower tissues at the PMC stage,and its chimeric sequence structure might reflect theinterspecific hybridization history of this genus. Wehypothesize that BspUPG2 arose via sequential (seg-mental) duplication-insertion events involving at leastfive loci, from which three originated from transcribedgenes (Fig. 7D). In this regard, it is interesting that onlythe apomixis-specific duplicated locus (BspUPG2)shows transcriptional activity, while the original locus(BspUPG1), which is present in both sexuals and apo-micts, does not (Fig. 5E). Whereas many studies havefocused on mutation accumulation and deregulation
1654 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

with respect to the origins of apomixis elements(Tucker et al., 2003; d’Erfurth et al., 2008), the emer-gence of novel genes in apomicts has not been appreci-ated, although various identified apomixis-associated locisuggest species-specific apomixis factors (Grossniklauset al., 2001) for which gains in function are hypothesized(Vielle-Calzada et al., 1996).
CONCLUSION
The identification of the novel apo-specific BspUPG2supports the HFA theory, which proposes that apo-meiosis, and in a broader perspective apomixis, originatesfrom hybrid-specific “genome collisions” and associatedinduction of gene duplication and TE activation (Carman,1997). BspUPG2 could have a potential application instabilizing the endosperm balance in the course of theimplementation of apomixis into crop plants. HowBspUPG2 has undergone neofunctionalization to developa trans-regulatory function remains to be clarified.
MATERIALS AND METHODS
Plant Material and Cultivation Conditions
Several sets of the same diploids, 10 apomictic and 12 sexual Boechera spp.,were used for all experiments (Supplemental Table S1; note the recent taxo-nomic information; Koch et al., 2010). Ten seeds per genotype were culturedon moist filter paper in sealed petri dishes and vernalized at 4°C in the darkfor 2 weeks until germination. Seedlings were transplanted to plastic pots(11 3 11 3 13 cm) containing autoclaved substrate and grown in a phytotronwithout insecticides and herbicides under long-day conditions (16 h of lightand 8 h of dark, 21°C).
Measurement of Relative Nuclear DNA Content andReproductive Mode
Relative nuclear DNA content (referred to as ploidy) in leaf, seed (Matzket al., 2000), and pollen (De Storme et al., 2013) was quantified using leaf tissuefrom a diploid sexual Boechera stricta (ES 558.2; Supplemental Table S1) as anexternal control. Leaf and seed material (20 seeds per individual plant) werechopped with a razor blade in a drop of Galbraith’s buffer containing 4 mg mL21
49,6-diamino-phenylindole (Galbraith et al., 1983). Tissue-specific ploidymeasurements were performed on a FACSAria II (BD Biosciences) equippedwith a 375-nm near-UV laser. Data were measured using the FACSDivaSoftware (version 6.1; BD Biosciences) and analyzed using WinMDI version2.9 (Scripps Research Institute; http://facs.scripps.edu/software.html); “C”refers to DNA content of a haploid anaphase cell, and “x” is the basic chro-mosome number.
Microscopy Analysis of Gametophyte Stages
Pollen developmental stages were defined (Fig. 3; Regan and Moffatt, 1990)from each of 12 flower bud size stages (S1–S12) that differed in 100-mm lengthincrements (Smyth et al., 1990; Sanders et al., 1999). For dissection of flowerbuds at stage S3 and antherheads at flower developmental stages S8 to S12using a Zeiss Discovery V20 (Carl Zeiss), stereomicroscope sterile glass nee-dles bent to an angle of approximately 100° to 120° and tip size of 50 to 100 mmusing a Narishige PC-10 puller (Narishige Group) were used. Sample prepa-ration of size-staged buds and antherheads for histology and light microscopywas carried out according to Grasser et al. (2009).
The decision to take the PMC stage for comparative gene expression pro-filing is based on two criteria: (1) apomeiosis candidate genes should affectearly meiotic stages (Grimanelli et al., 2001b), and (2) in previous analyses, thespike of gene expression change between sexual and apomictic ovules was
detected in the microspore mother cell (Sharbel et al., 2010), a stage that cor-responds to the PMC in antherheads.
Cytochemical Analysis of Fixated Boechera spp. Anthers
Cytological observations of meiosis and PMCs were made from sexuals andapomicts at flower bud stages S9 to S10. Meiotic chromosome spreads ofanthers were prepared according to Ross et al. (1996) with minor changes andobserved with a Zeiss Axioplan 2 imaging microscope (Carl Zeiss). Photo-graphs were taken with an AxioCam HRc Rev. 2 camera under 100-foldmagnification using a 49 DAPI BP reflector block. Meiocytes at the tetrad stagewere examined by separate squashes of all six antherheads per flower bud,from one sexual and six apomictic genotypes, to determine the number ofmonads, dyads, triads, and tetrads in each antherhead. Anthers were squashedaccording to Peterson et al. (2010), stained (Alexander’s stain; Alexander, 1969),and examined with a Zeiss Axioplan 2 imaging microscope (Carl Zeiss). Allavailable meiocytes per anther were counted, and statistical analyses of meiocytebehavior and anther size correlations of gametophyte stages were evaluatedwith SPSS version 11.5 (LEAD Technologies).
RNA Isolation, Microarray Hybridization, and qRT-PCR ofAntherhead Tissue
Microdissections and total RNA extractions were prepared using tools and adissection area cleaned with ethanol, treated with RNAseZap (Ambion), andwashed with diethyl phosphorocyanidate-treated distilled water. Approxi-mately 30 antherheads of each genotype, corresponding to the PMC stage, werelive microdissected from fresh whole flower buds using a Zeiss Discovery V20stereomicroscope (Carl Zeiss) using sterile glass needles (see above) and col-lected in 500 mL of RNAlater (Qiagen). Eighteen microliters of RNase-freeDNase I (Qiagen)-digested total RNA extracts (RNeasy Micro kit; Qiagen)was eluted through RNeasy MinElute Spin columns (Qiagen) and quantifiedon a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies). TheRNA quality of all samples was assessed with the RNA 6000 Nano LabChipKit II (Agilent Technologies) on the Agilent 2100 Bioanalyzer. Purified totalRNA was stored at 280°C.
To ensure optimal copy RNA yield after labeling, 200 ng of total RNA withabsorbance readings of A260/A280 . 1.8 and A260/A230 . 1.8 (recommended byAgilent) was used for the labeling procedure. Approximately 1.5 mg of thegenerated Cy3-labeled copy RNA per sample (One-color Quick Amp Labelingkit; Agilent Technologies) was hybridized for 18 h at 65°C to the customBoechera spp. whole flower-105k-Agilent microarrays, which were scanned at5-mm double-pass resolution with an Agilent G2565BA Microarray Scanner.The One-color RNA Spike-in kit (Agilent Technologies) was used to ensureoptimal microarray processing. Microarray hybridization quality was assessedwith Feature Extraction 10.1 software (Agilent Technologies), whereas quan-tile normalization with baseline-to-median transformation and gene expres-sion analysis were performed with GeneSpring GX 10 software (AgilentTechnologies). Differentially expressed microarray probes were consideredvalidated with P # 0.05 as assessed by an unpaired Student’s t test with amean difference of 2-fold or greater. P values were corrected for the family-wise error rate as a control for false positives using the Bonferroni method.
Real-time PCR, using primers that were designed on the cDNA readshomologous to the candidate microarray probes Sharb0690829 (GenBank/EMBL/DDBJ accession no. ERS317557), Sharb0501554 (GenBank/EMBL/DDBJ accession no. ERS317556), and Sharb0931225 (GenBank/EMBL/DDBJaccession no. ERS317555; Primer3 version 0.4.0; http://frodo.wi.mit.edu/primer3/), was performed according to Sharbel et al. (2010). Seven biologicaland four technical replicates were run for each probe and tissue on a 384-wellplate together with two endogenous control genes tested on Boechera spp.anther material (ACTIN2 and ELONGATION FACTOR a1; Pellino et al., 2011),negative template, and reverse transcriptase controls. PCR efficiencies andnormalized cycle threshold values of each set of four technical replicates wereprocessed with the Real-time PCR Miner version 2.0 software (Zhao andFernald, 2005). Relative quantification and normalized cycle threshold valuesof the amplified targets were calculated separately with reference to the ex-pression levels of each of the two housekeeping genes employing the ΔΔCtmethod (Pfaffl, 2001) using a calibrator sample (ES 910.2; Supplemental Table S7).The corresponding mean relative expression ratio for each genotype was calcu-lated with SPSS version 11.5 (LEAD Technologies), and significant differencesbetween samples were evaluated using a one-way ANOVA (a = 0.05) withTukey’s HSD post hoc test for differences between multiple pairs of means.
Plant Physiol. Vol. 163, 2013 1655
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

RACE
The SMARTer RACEmethod (Clontech) was employed to obtain 59 end andpoly(A) site information from BspUPG2 and BspHRD3, an Arabidopsis (Arabi-dopsis thaliana) homolog of HRD3 in Boechera spp. Primers (Primer3 version0.4.0) for BspUPG2 RACE were derived from microarray probe homologouscDNA read ERS317557 (59 end primer GSP3, 59-TCTTCGCCATCGTTCATGTT-TACTTCCG-39; 39 end primer GSP4, 59-TCATCATGTCTTCTTCGCCATCGTTCA-39),and those for BspHRD3 RACE were derived from the LCB2 of BspUPG2(59 end primer GSP11, 59-TAATGCCCACTGGGGTCGTCATTGT-39; 39 endprimer CON234X14_L, 59-ACTGGAATTGGGTACTTGTATGTCA-39). PCR wasperformed with the Advantage 2 PCR Kit (Clontech), and PCR fragments werecloned (pCR4-TOPO TA; Life Technologies) and Sanger sequenced (see above).
Computational Analysis of RNA-Folding Probabilitiesof BspUPG
Structural RNAs are usually characterized by an unusual thermodynamicstability and a conserved secondary structure. The minimum folding energy asa measure of thermodynamic stability for a sequence (i.e. negative valuesindicate that a sequence is more stable) was calculated using the RNAfold andRNAz version 1.0 software forWindows of the Vienna RNA package (Hofackeret al., 1994; http://www.tbi.univie.ac.at/ivo/RNA). The presence of statis-tically significant secondary structures of BspUPG2 was monitored usingZ-score values as described by Crespi et al. (1994) and Bonnet et al. (2004) usingsimilar thresholds to Kavanaugh and Dietrich (2009). All significant secondarystructures of BspUPG2 were classified according to Zhang et al. (2006). The“npcRNA [number]” names were chosen to follow the naming convention estab-lished by previous investigators (Hirsch et al., 2006).
BAC Probe Preparation and Screening of a Boechera spp.BAC Library
Candidate 60-mer microarray probes were mapped against Boechera spp.454 FLX cDNA libraries (Sharbel et al., 2009) using CLC Genomics Workbenchversion 4.5.1 (CLC Bio; standard parameters). Flanking regions to the mappedmicroarray probes were used to design specific primers (Primer3 version 0.4.0)for chromosome walking in sexuals and apomicts using the DNA WalkingSpeedUp Premix Kit I from Seegene (Supplemental Table S10). Resultantproducts were transformed into Escherichia coli TOP10 cells using the TOPOTA Cloning Kit for Sequencing (Invitrogen), and individual clones were am-plified using the TempliPhi DNA Sequencing Template Amplification Kit(Reagin et al., 2003) and Sanger sequenced on an ABI 3730 XL sequencingsystem. Sequence analysis and assembly were carried out with Lasergene 8(DNAStar). The E. coli TOP10 subcloned, PCR-amplified, and finally gel-purified DNA walking products (NucleoSpin Extract II kit; Macherey-Nagel)were hybridized against a gridded BAC library (48 3 384 spotted wells onto a22- 3 22-cm filter membrane; binary vector pCLD04541; Bancroft, 1997). In-dividually radiolabeled and pooled probes were hybridized as a group usingthe overgo hybridization method (Ross et al., 1999).
Vector inserts of 500 ng of pure BAC clone DNA extracts were restrictiondigested to completion with HindIII, BglII, and BamHI (Amersham PharmaciaBiotech) at 37°C (HindIII and BglII) and 30°C (BamHI) for 8 h and examined ona 1% agarose gel. Based on their partial overlapping restriction patterns, DNAwas isolated from four BAC clones (A4O22, E7K5, C8B11, and F8G11) usingNucleobond Xtra Midi Kits (Macherey-Nagel). BAC DNA was randomlysheared (Hydroshear; Digilab) and size fractionated by agarose gel electro-phoresis in approximately 1-kb and approximately 4- to 5-kb size classes.These fragments were end repaired, blunt-end ligated into pUC19 (LifeTechnologies), transformed into E. coli ELECTROMAX DH5a-E electro-competent cells (Invitrogen), and sequenced on an ABI 3730 XL automaticDNA sequencer (PE Applied Biosystems). Vector clipping, quality trimming,and sequence assembly using stringent conditions (e.g. 95% sequence identitycutoff, 25-bp overlap) were done using Lasergene 8 (DNASTAR) and Staden(http://staden.sourceforge.net/). Assembly of the complete BAC clone se-quences was performed using Seqman and the Gap4 algorithm implementedin Staden. Remaining gaps in the contiguous BAC sequences were manuallyinspected and closed with primer walking or PCR products crossing the gapsfrom adjacent contigs. The resulting sequences were assembled using Laser-gene 8 (DNASTAR) set to an overlap minimum of 20 bp with 95% identity. AllBAC assemblies were annotated using BLASTN and BLASTX searches againstthe nonredundant GenBank nucleotide and protein databases, respectively(http://www.ncbi.nlm.nih.gov/genbank/).
Processing of DNA Sequences and Sequence Analysis
The allelic constitution of BspUPG2 in nine each sexual and apomicticgenotypes was determined by sequencing proofreading polymerase-amplified(Phusion high-fidelity polymerase; Thermo Scientific), gel-purified (Nucleo-Spin Extract II kit; Macherey-Nagel), and multiply cloned (CloneJet PCR cloning kit;Fermentas) PCR fragments using specific primers (59 end primer in apomicticgenotypes, CON234X5L, 59-TCCGACCTAAATCCTACCAAACTGA-39; in sexualB. stricta, CON234X11L, 59-CAAAAATAAAAGATTTGATGTAGATTGC-39, and inother sexual genotypes, FLTsexX2L, 59-GAAGAAAGAGCTACGGCGTGAT-39;39 end primer CON234X5R, 59-TGCTCAATTTTGAACATCTTATTTGC-39). Laser-gene 8 (DNASTAR) was used for assembly and similarity analysis of Phred 20quality-trimmed sequences. Coding potential was calculated from all six frames ofall BspUPG2 splicing forms using the Coding Potential Calculator software (Konget al., 2007).
Pairwise ClustalW comparisons in the presence of rearrangements wereperformed with Mauve (version 2.3.1; progressiveMauve, default parameters;Darling et al., 2004) to detect LCBs (conserved sequence segments, which areinternally free from genome rearrangements) in BspUPG1 and BspUPG2 be-tween different genotypes.
Local BLASTN search of the complete genomic sequence read archive(SRP007750; http://www.ncbi.nlm.nih.gov/sra/; 454 GS FLX) and sequenceextraction were conducted with CLC Genomics Workbench (costs = match 1,mismatch 3, existence 5, extension 2; E value = 10; word size = 11, filtercomplexity = yes). For mapping of both 454 whole-genome sequencing andCNV sequence tags (Aliyu et al., 2013), the genomic sequence of BspUPG2 wasdivided into 18 200-bp bins with 20-bp overlap. All existing CNVs for eachsingle sequence tag are represented, and relative CNV frequencies are con-sidered independently for sexual and apomictic genotypes.
BAC sequence assemblies were annotated for transposable elements byscreening the green plant section (Viridiplantae) of the Repbase repetitiveelement database (Jurka, 1998; http://www.girinst.org/server/Maps/AT/index.html) using CENSOR (Kohany et al., 2006). The programs einverted andEMBOSS (Rice et al., 2000) were employed to identify inverted repeats with80% or more matches, and the Pipmaker software was used to identify simplerepeats and CpG islands (Schwartz et al., 2003). LTR analyses of Assembly 1 andAssembly 2 were performed with LTR FINDER software (Xu and Wang, 2007).
Sequence data from this article can be found in the GenBank/EMBL datalibraries under accession numbers ERS317552, ERS317553, ERS317554, ERS317555,ERS317556, ERS317557, ERS317558, HF930769, HF954100, HF954101, HG007837and HG007838.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. Representative meiocytes at the tetrad stage andflow cytometric ploidy confirmation.
Supplemental Figure S2. Correlation of flower organs and flower budlength for sexual and apomictic Boechera spp. genotypes.
Supplemental Figure S3. Constantly differentially regulated microarrayprobes in apomictic compared with sexual Boechera spp. genotypes.
Supplemental Figure S4. Validation of differentially expressed microarrayprobes by qRT-PCR.
Supplemental Figure S5. Rapid amplification of cDNA 39 and 59 ends ofthe candidate gene UPGRADE.
Supplemental Figure S6. Percentage identity plots showing the originalUPGRADE locus and its duplicated variant.
Supplemental Figure S7. Distribution of locally collinear sequence blocksalong the BAC clone Assembly 1 and Assembly 2.
Supplemental Figure S8. Distribution of Arabidopsis homologous locicorresponding to Assembly 1 and Assembly 2 in Boechera spp. alongArabidopsis chromosomes.
Supplemental Figure S9. Thermodynamically stable nonrandom second-ary structures of BspUPG2.
Supplemental Figure S10. Northern-blot analysis for putative small RNAshomologous to indel 9 at BspUPG2.
1656 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Supplemental Figure S11. Distribution of the TE superfamilies along theoriginal and duplicated UPGRADE locus.
Supplemental Table S1. List of Boechera spp. genotypes with populationinformation used for histological examination of microsporogenesis andcandidate gene analyses.
Supplemental Table S2. Analysis of meiocyte constitution at the tetradstage in diploid sexual and high facultative and obligate apomictic Boe-chera spp. genotypes.
Supplemental Table S3. High-level 1C pollen individuals with corre-sponding flow cytometric seed screen data.
Supplemental Table S4. Frequencies of microspores, meiotic cells, andPMCs relative to antherhead length.
Supplemental Table S5. Microarray probes demonstrating significant ab-solute fold change up-regulation in apomictic genotypes.
Supplemental Table S6. qRT-PCR primers used for the validation ofmicroarray candidate probes.
Supplemental Table S7. Relative mRNA expression values for four micro-array probes in somatic and reproductive tissues.
Supplemental Table S8. Overview of BAC clones carrying one to threecandidate microarray probes.
Supplemental Table S9. Transcription factor-binding sites upstream ofBspUPG2.
Supplemental Table S10. Primers used for chromosome walking onmicroarray candidate probes and the source gene BspHRD3.
Supplemental Table S11. Gene annotation of Boechera spp. BAC cloneAssembly 1 and Assembly 2.
Supplemental Table S12. Distribution of inverted repeats on Assembly1 and Assembly 2.
Supplemental Table S13. Detection of candidate regions for stable non-protein-coding RNA secondary structures of BspUPG2.
Supplemental Table S14. Characteristics of thermodynamically stable sec-ondary non-protein-coding RNA structures with highest Z-score fromBspUPG2 in apomictic Boechera spp. genotypes.
Supplemental Table S15. Sequence divergence of BspUPG1 and BspUPG2.
Supplemental Table S16. Indels on BspUPG2 isolates of sexual and apo-mictic genotypes using a ClustalW multiple sequence alignment.
Supplemental Table S17. Green plant (Viridiplantae) homologous repeti-tive DNA element sequence mapping on Assembly 1.
Supplemental Table S18. Green plant (Viridiplantae) homologous repeti-tive DNA element sequence mapping on Assembly 2.
Supplemental Protocol S1. Computational analysis of RNA-folding prob-abilities of BspUPG2.
Supplemental Protocol S2. Small RNA northern blot.
ACKNOWLEDGMENTS
The authors thank Heidi Block, Vera Katzorke, Monika Wiesner and InesWalde for technical support, Ingo Schubert for constructive comments on themanuscript, and Thomas Mitchell-Olds and Eric Schranz for seed material.
Received June 4, 2013; accepted October 9, 2013; published October 15, 2013.
LITERATURE CITED
Albers CA, Lunter G, MacArthur DG, McVean G, Ouwehand WH,Durbin R (2011) Dindel: accurate indel calls from short-read data. Ge-nome Res 21: 961–973
Albertini E, Marconi G, Barcaccia G, Raggi L, Falcinelli M (2004) Isolation ofcandidate genes for apomixis in Poa pratensis L. Plant Mol Biol 56: 879–894
Alexander MP (1969) Differential staining of aborted and nonabortedpollen. Stain Technol 44: 117–122
Aliyu OM, Schranz ME, Sharbel TF (2010) Quantitative variation forapomictic reproduction in the genus Boechera (Brassicaceae). Am J Bot97: 1719–1731
Aliyu OM, Seifert M, Corral JM, Fuchs J, Sharbel TF (2013) Copy numbervariation in transcriptionally active regions of sexual and apomicticBoechera demonstrates independently derived apomictic lineages. PlantCell http://dx.doi.org/10.1105/tpc.113.113860
Al-Shehbaz IA, Windham M (2010) Brassicaceae. In Flora of North AmericaEditorial Committee, eds, Flora of North America North of Mexico, Vol7. Oxford University Press, New York and Oxford, pp 224–746
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W,Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation ofprotein database search programs. Nucleic Acids Res 25: 3389–3402
Altuvia Y, Landgraf P, Lithwick G, Elefant N, Pfeffer S, Aravin A,Brownstein MJ, Tuschl T, Margalit H (2005) Clustering and conser-vation patterns of human microRNAs. Nucleic Acids Res 33: 2697–2706
Amiteye S, Corral JM, Vogel H, Kuhlmann M, Mette MF, Sharbel TF(2013) Novel microRNAs and microsatellite-like small RNAs in sexualand apomictic Boechera species. MicroRNA 2: 46–63
Amiteye S, Corral JM, Vogel H, Sharbel TF (2011) Analysis of conservedmicroRNAs in floral tissues of sexual and apomictic Boechera species.BMC Genomics 12: 500
Bancroft I (1997) A strategy involving the use of high redundancy YACsubclone libraries facilitates the contiguous representation in cosmidand BAC clones of 1.7 Mb of the genome of the plant Arabidopsisthaliana. Weeds World 4: 1–9
Beck JB, Alexander PJ, Allphin L, Al-Shehbaz IA, Rushworth C, BaileyCD, Windham MD (2012) Does hybridization drive the transition toasexuality in diploid Boechera? Evolution 66: 985–995
Bellé R, Cormier P, Poulhe R, Morales J, Huchon D, Mulner-Lorillon O(1990) Protein phosphorylation during meiotic maturation of Xenopusoocytes: cdc2 protein kinase targets. Int J Dev Biol 34: 111–115
Bennetzen JL (2005) Transposable elements, gene creation and genomerearrangement in flowering plants. Curr Opin Genet Dev 15: 621–627
Bicknell RA, Borst NK, Koltunow AM (2000) Monogenic inheritance ofapomixis in two Hieracium species with distinct developmental mech-anisms. Heredity (Edinb) 84: 228–237
Böcher TW (1951) Cytological and embryologal studies in the amphiapo-mictic Arabis holboellii complex. Biologiske Skrifter/Kongelige DanskeVidenskabernes Selskab 6: 1-59
Böcher TW (1954) Experimental taxonomical studies in the Arabis holboelliicomplex. Sven Bot Tidskr 48: 31–44
Bonnet E, Wuyts J, Rouzé P, Van de Peer Y (2004) Evidence that micro-RNA precursors, unlike other non-coding RNAs, have lower folding freeenergies than random sequences. Bioinformatics 20: 2911–2917
Borges F, Gomes G, Gardner R, Moreno N, McCormick S, Feijó JA,Becker JD (2008) Comparative transcriptomics of Arabidopsis spermcells. Plant Physiol 148: 1168–1181
Brownfield L, Köhler C (2011) Unreduced gamete formation in plants:mechanisms and prospects. J Exp Bot 62: 1659–1668
Carman JG (1997) Asynchronous expression of duplicate genes in angio-sperms may cause apomixis, bispory, tetraspory, and polyembryony.Biol J Linn Soc Lond 61: 51–94
Casillas S, Barbadilla A, Bergman CM (2007) Purifying selection maintainshighly conserved noncoding sequences in Drosophila. Mol Biol Evol 24:2222–2234
Chang WC, Lee TY, Huang HD, Huang HY, Pan RL (2008) PlantPAN:Plant Promoter Analysis Navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene group. BMCGenomics 9: 561
Chellappan P, Xia J, Zhou X, Gao S, Zhang X, Coutino G, Vazquez F,Zhang W, Jin H (2010) siRNAs from miRNA sites mediate DNAmethylation of target genes. Nucleic Acids Res 38: 6883–6894
Comai L (2000) Genetic and epigenetic interactions in allopolyploid plants.Plant Mol Biol 43: 387–399
Corral JM, Vogel H, Aliyu OM, Hensel G, Thiel T, Kumlehn J, SharbelTF (2013) A conserved apomixis-specific polymorphism is correlatedwith exclusive DEDDh exonuclease expression in premeiotic ovules ofapomictic Boechera Species. Plant Physiol 163: 1660–1672
Crespi MD, Jurkevitch E, Poiret M, d’Aubenton-Carafa Y, Petrovics G,Kondorosi E, Kondorosi A (1994) enod40, a gene expressed duringnodule organogenesis, codes for a non-translatable RNA involved inplant growth. EMBO J 13: 5099–5112
Plant Physiol. Vol. 163, 2013 1657
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

Darling ACE, Mau B, Blattner FR, Perna NT (2004) Mauve: multiplealignment of conserved genomic sequence with rearrangements. Ge-nome Res 14: 1394–1403
Deaton AM, Bird A (2011) CpG islands and the regulation of transcription.Genes Dev 25: 1010–1022
De Lucia F, Dean C (2011) Long non-coding RNAs and chromatin regu-lation. Curr Opin Plant Biol 14: 168–173
d’Erfurth I, Jolivet S, Froger N, Catrice O, Novatchkova M, Simon M,Jenczewski E, Mercier R (2008) Mutations in AtPS1 (Arabidopsis thalianaparallel spindle 1) lead to the production of diploid pollen grains. PLoSGenet 4: e1000274
De Storme N, Zamariola L, Mau M, Sharbel TF, Geelen D (2013) Volume-based pollen size analysis: an advanced method to assess somatic andgametophytic ploidy in flowering plants. Plant Reprod 26: 65–81
Eamens A, Wang MB, Smith NA, Waterhouse PM (2008) RNA silencing inplants: yesterday, today, and tomorrow. Plant Physiol 147: 456–468
Fiston-Lavier AS, Vejnar CE, Quesneville H (2012) Transposable sequenceevolution is driven by gene context. Cornell University Library, http://arxiv.org/abs/1209.0176
Fransz PF, Armstrong SJ, de Jong JH, Parnell LD, van Drunen C, Dean C,Zabel P, Bisseling T, Jones GH (2000) Integrated cytogenetic map ofchromosome arm 4S of A. thaliana: structural organization of hetero-chromatic knob and centromere region. Cell 100: 367–376
Galbraith DW, Harkins KR, Maddox JM, Ayres NM, Sharma DP,Firoozabady E (1983) Rapid flow cytometric analysis of the cell cycle inintact plant tissues. Science 220: 1049–1051
Grant-Downton R, Le Trionnaire G, Schmid R, Rodriguez-Enriquez J, HafidhS, Mehdi S, Twell D, Dickinson H (2009b) MicroRNA and tasiRNA di-versity in mature pollen of Arabidopsis thaliana. BMC Genomics 10: 643
Grasser M, Kane CM, Merkle T, Melzer M, Emmersen J, Grasser KD(2009) Transcript elongation factor TFIIS is involved in Arabidopsis seeddormancy. J Mol Biol 386: 598–611
Grimanelli D, Leblanc O, Perotti E, Grossniklaus U (2001a) Develop-mental genetics of gametophytic apomixis. Trends Genet 17: 597–604
Grimanelli D, Tohme J, González-De-León D (2001b) Applications ofmolecular genetics in apomixis research. In Y Savidan, JG Carman, TDresselhaus, eds, The Flowering of Apomixis: From Mechanisms toGenetic Engineering. Centro Internacional de Mejoramiento de Maíz yTrigo (CIMMYT), Institut de Recherche pour le Developpernent (IRD),European Commission DG VI (FAIR), Mexico, D.F., pp 83–94
Grossniklaus U (2001a) From sexuality to apomixis: molecular and geneticapproaches. In Y Savidan, JG Carman, T Dresselhaus, eds, The Flow-ering of Apomixis: From Mechanisms to Genetic Engineering. CentroInternacional de Mejoramiento de Maíz y Trigo (CIMMYT), Institut deRecherche pour le Developpernent (IRD), European Commission DG VI(FAIR), Mexico, D.F., pp 168–211
Grossniklaus U, Nogler GA, van Dijk PJ (2001) How to avoid sex: thegenetic control of gametophytic apomixis. Plant Cell 13: 1491–1498
Guerin J, Rossel JB, Robert S, Tsuchiya T, Koltunow A (2000) A DEFI-CIENS homologue is down-regulated during apomictic initiation in ovules ofHieracium. Planta 210: 914–920
Hepler PK, Valster A, Molchan T, Vos JW (2002) Roles for kinesin andmyosin during cytokinesis. Philos Trans R Soc Lond B Biol Sci 357: 761–766
Hirsch J, Lefort V, Vankersschaver M, Boualem A, Lucas A, Thermes C,d’Aubenton-Carafa Y, Crespi M (2006) Characterization of 43 non-protein-coding mRNA genes in Arabidopsis, including the MIR162a-derived transcripts. Plant Physiol 140: 1192–1204
Hofacker IL, Fontana W, Stadler PF, Bonhoeffer SL, Tacker M, Schuster P(1994) Fast folding and comparison of RNA secondary structures. ChemicalMonthly 125: 167–188
Honys D, Twell D (2004) Transcriptome analysis of haploid male game-tophyte development in Arabidopsis. Genome Biol 5: R85
Hörandl E, Temsch EM (2009) Introgression of apomixis into sexual speciesis inhibited by mentor effects and ploidy barriers in the Ranunculusauricomus complex. Ann Bot (Lond) 104: 81–89
Hosouchi T, Kumekawa N, Tsuruoka H, Kotani H (2002) Physical map-based sizes of the centromeric regions of Arabidopsis thaliana chromo-somes 1, 2, and 3. DNA Res 9: 117–121
Hunt AG (1994) Messenger RNA 39 end formation in plants. Annu RevPlant Physiol Plant Mol Biol 45: 47–60
Jiang N, Bao Z, Zhang X, Eddy SR Sr, Wessler SR (2004) Pack-MULE trans-posable elements mediate gene evolution in plants. Nature 431: 569–573
Jiang SY, Cai M, Ramachandran S (2007) ORYZA SATIVA MYOSIN XI Bcontrols pollen development by photoperiod-sensitive protein localiza-tions. Dev Biol 304: 579–592
Johnston SA, den Nijs TPM, Peloquin SJ, Hanneman RE Jr (1980) Thesignificance of genic balance to endosperm development in interspecificcrosses. Theor Appl Genet 57: 5–9
Jurka J (1998) Repeats in genomic DNA: mining and meaning. Curr OpinStruct Biol 8: 333–337
Kaessmann H (2010) Origins, evolution, and phenotypic impact of newgenes. Genome Res 20: 1313–1326
Kantama L, Sharbel TF, Schranz ME, Mitchell-Olds T, de Vries S, de JongH (2007) Diploid apomicts of the Boechera holboellii complex displaylarge-scale chromosome substitutions and aberrant chromosomes. ProcNatl Acad Sci USA 104: 14026–14031
Kavanaugh LA, Dietrich FS (2009) Non-coding RNA prediction and veri-fication in Saccharomyces cerevisiae. PLoS Genet 5: e1000321
Koch MA, Karl R, Kiefer C, Al-Shehbaz IA (2010) Colonizing the Amer-ican continent: systematics of the genusArabis in North America (Brassicaceae).Am J Bot 97: 1040–1057
Kohany O, Gentles AJ, Hankus L, Jurka J (2006) Annotation, submissionand screening of repetitive elements in Repbase: RepbaseSubmitter andCensor. BMC Bioinformatics 7: 474
Kondo T, Hashimoto Y, Kato K, Inagaki S, Hayashi S, Kageyama Y (2007)Small peptide regulators of actin-based cell morphogenesis encoded bya polycistronic mRNA. Nat Cell Biol 9: 660–665
Kong L, Zhang Y, Ye ZQ, Liu XQ, Zhao SQ, Wei L, Gao G (2007) CPC:assess the protein-coding potential of transcripts using sequence fea-tures and support vector machine. Nucleic Acids Res 36: 345–349
Lu X, Liu X, An L, Zhang W, Sun J, Pei H, Meng H, Fan Y, Zhang C (2008)The Arabidopsis MutS homolog AtMSH5 is required for normal meiosis.Cell Res 18: 589–599
Ma J, Yan B, Qu Y, Qin F, Yang Y, Hao X, Yu J, Zhao Q, Zhu D, Ao G(2008) Zm401, a short-open reading-frame mRNA or noncoding RNA, isessential for tapetum and microspore development and can regulate thefloret formation in maize. J Cell Biochem 105: 136–146
Marimuthu MPA, Jolivet S, Ravi M, Pereira L, Davda JN, Cromer L,Wang L, Nogué F, Chan SWL, Siddiqi I, et al (2011) Synthetic clonalreproduction through seeds. Science 331: 876
Mascarenhas JP (1989) The male gametophyte of flowering plants. PlantCell 1: 657–664
Matzk F, Meister A, Schubert I (2000) An efficient screen for reproductivepathways using mature seeds of monocots and dicots. Plant J 21: 97–108
Matzk F, Prodanovic S, Bäumlein H, Schubert I (2005) The inheritance ofapomixis in Poa pratensis confirms a five locus model with differences ingene expressivity and penetrance. Plant Cell 17: 13–24
Meyer P, Saedler H (1996) Homology-dependent gene silencing in plants.Annu Rev Plant Physiol Plant Mol Biol 47: 23–48
Mogie M (1988) A model for the evolution and control of generative apo-mixis. Biol J Linn Soc Lond 35: 127–153
Morgante M, Brunner S, Pea G, Fengler K, Zuccolo A, Rafalski A (2005)Gene duplication and exon shuffling by helitron-like transposons gen-erate intraspecies diversity in maize. Nat Genet 37: 997–1002
Naumova TN (2001) Reproductive development in apomictic populationsof Arabis holboellii (Brassicaceae). Sex Plant Reprod 14: 195–200
Nogler GA (1984) Gametophytic apomixis. In BM Johri, ed, Embryology ofAngiosperms. Springer-Verlag, Berlin, pp 475–518
Noyes RD, Rieseberg LH (2000) Two independent loci control agamo-spermy (apomixis) in the triploid flowering plant Erigeron annuus. Ge-netics 155: 379–390
Ohno S (1970) Evolution by Gene Duplication. Springer-Verlag, BerlinOhno S (1972) So much “junk” DNA in our genome. Brookhaven Symp Biol
23: 366–370Ozias-Akins P, Roche D, Hanna WW (1998) Tight clustering and hemi-
zygosity of apomixis-linked molecular markers in Pennisetum squamu-latum implies genetic control of apospory by a divergent locus that mayhave no allelic form in sexual genotypes. Proc Natl Acad Sci USA 95:5127–5132
Pellino M, Sharbel TF, Mau M, Amiteye S, Corral JM (2011) Selection ofreference genes for quantitative real-time PCR expression studies ofmicrodissected reproductive tissues in apomictic and sexual Boechera.BMC Res Notes 4: 303
Peragine A, Yoshikawa M, Wu G, Albrecht HL, Poethig RS (2004) SGS3and SGS2/SDE1/RDR6 are required for juvenile development and the
1658 Plant Physiol. Vol. 163, 2013
Mau et al.
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.

production of trans-acting siRNAs in Arabidopsis. Genes Dev 18: 2368–2379
Peters HI, Chang YWE, Traugh JA (1995) Phosphorylation of elongationfactor 1 (EF-1) by protein kinase C stimulates GDP/GTP-exchange ac-tivity. Eur J Biochem 234: 550–556
Peterson R, Slovin JP, Chen C (2010) A simplified method for differentialstaining of aborted and non-aborted pollen grains. International Journalof Plant Biology 1: 66–69
Pfaffl MW (2001) A new mathematical model for relative quantification inreal-time RT-PCR. Nucleic Acids Res 29: 2002–2007
Pikaard CS, Haag JR, Ream T, Wierzbicki AT (2008) Roles of RNA polymeraseIV in gene silencing. Trends Plant Sci 13: 390–397
Pina C, Pinto F, Feijo JA, Becker JD (2005) Gene family analysis of the Arabi-dopsis pollen transcriptome reveals biological implications for cell growth,division control, and gene expression regulation. Plant Physiol 138: 744–756
Reagin MJ, Giesler TL, Merla AL, Resetar-Gerke JM, Kapolka KM,Mamone JA (2003) TempliPhi: a sequencing template preparation pro-cedure that eliminates overnight cultures and DNA purification. J Bio-mol Tech 14: 143–148
Regan SM, Moffatt BA (1990) Cytochemical analysis of pollen develop-ment in wild-type Arabidopsis and a male-sterile mutant. Plant Cell 2:877–889
Rice P, Longden I, Bleasby A (2000) EMBOSS: the European MolecularBiology Open Software Suite. Trends Genet 16: 276–277
Rollins RC (1941) Monographic study of Arabis in western North America.Rhodora 43: 289–483
Ross KJ, Fransz P, Jones GH (1996) A light microscopic atlas of meiosis inArabidopsis thaliana. Chromosome Res 4: 507–516
Ross M, LaBrie T, McPherson S, Stanton VPS (1999) Screening large-insertlibraries by hybridization. In Current Protocols in Human Genetics. JohnWiley & Sons, New York, unit 5.6
Roy BA (1995) The breeding systems of six species of Arabis (Brassicaceae).Am J Bot 82: 869–877
Sanders PM, Bui AQ, Weterings K, McIntire KN, Hsu YC, Lee PY, TruongMT, Beals TP, Goldberg RB (1999) Anther developmental defects inArabidopsis thaliana male-sterile mutants. Sex Plant Reprod 11: 297–322
Savidan Y (2001) Transfer of apomixis through wide crosses. In Y Savidan,JG Carman, T Dresselhaus, eds, The Flowering of Apomixis: FromMechanisms to Genetic Engineering. Centro Internacional de Mejor-amiento de Maíz y Trigo (CIMMYT), Institut de Recherche pour le De-veloppernent (IRD), European Commission DG VI (FAIR), Mexico, D.F.,pp 153–167
Schallau A, Arzenton F, Johnston AJ, Hähnel U, Koszegi D, Blattner FR,Altschmied L, Haberer G, Barcaccia G, Bäumlein H (2010) Identifica-tion and genetic analysis of the APOSPORY locus in Hypericum per-foratum L. Plant J 62: 773–784
Schranz ME, Dobeš C, Koch MA, Mitchell-Olds T (2005) Sexual repro-duction, hybridization, apomixis, and polyploidization in the genusBoechera (Brassicaceae). Am J Bot 92: 1797–1810
Schranz ME, Windsor AJ, Song BH, Lawton-Rauh A, Mitchell-Olds T(2007) Comparative genetic mapping in Boechera stricta, a close relativeof Arabidopsis. Plant Physiol 144: 286–298
Schwartz S, Elnitski L, Li M, Weirauch M, Riemer C, Smit A, Green ED,Hardison RC, Miller W (2003) MultiPipMaker and supporting tools:alignments and analysis of multiple genomic DNA sequences. NucleicAcids Res 31: 3518–3524
Seffens W, Digby D (1999) mRNAs have greater negative folding freeenergies than shuffled or codon choice randomized sequences. NucleicAcids Res 27: 1578–1584
Sharbel TF, Voigt ML, Corral JM, Galla G, Kumlehn J, Klukas C,Schreiber F, Vogel H, Rotter B (2010) Apomictic and sexual ovules ofBoechera display heterochronic global gene expression patterns. PlantCell 22: 655–671
Sharbel TF, Voigt ML, Corral JM, Thiel T, Varshney A, Kumlehn J, VogelH, Rotter B (2009) Molecular signatures of apomictic and sexual ovulesin the Boechera holboellii complex. Plant J 58: 870–882
Simpson GG, Filipowicz W (1996) Splicing of precursors to mRNA inhigher plants: mechanism, regulation and sub-nuclear organisation ofthe spliceosomal machinery. Plant Mol Biol 32: 1–41
Smyth DR, Bowman JL, Meyerowitz EM (1990) Early flower developmentin Arabidopsis. Plant Cell 2: 755–767
Song JH, Cao JS, Wang CG (2013) BcMF11, a novel non-coding RNA genefrom Brassica campestris, is required for pollen development and malefertility. Plant Cell Rep32: 21–30
Song JH, Cao JS, Yu XL, Xiang X (2007) BcMF11, a putative pollen-specificnon-coding RNA from Brassica campestris ssp. chinensis. J Plant Physiol164: 1097–1100
Stamm S, Ben-Ari S, Rafalska I, Tang Y, Zhang Z, Toiber D, Thanaraj TA,Soreq H (2005) Function of alternative splicing. Gene 344: 1–20
Sundaresan V, Springer P, Volpe T, Haward S, Jones JDG, Dean C, Ma H,Martienssen R (1995) Patterns of gene action in plant development re-vealed by enhancer trap and gene trap transposable elements. GenesDev 9: 1797–1810
Tucker MR, Araujo AC, Paech NA, Hecht V, Schmidt ED, Rossell JB, DeVries SC, Koltunow AM (2003) Sexual and apomictic reproduction inHieracium subgenus pilosella are closely interrelated developmentalpathways. Plant Cell 15: 1524–1537
Twell D (2011) Male gametogenesis and germline specification in floweringplants. Sex Plant Reprod 24: 149–160
Van Dijk PJ, Tas IC, Falque M, Bakx-Schotman T (1999) Crosses betweensexual and apomictic dandelions (Taraxacum). II. The breakdown ofapomixis. Heredity (Edinb) 83: 715–721
Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, MalloryAC, Hilbert JL, Bartel DP, Crété P (2004) Endogenous trans-actingsiRNAs regulate the accumulation of Arabidopsis mRNAs. Mol Cell 16:69–79
Vielle-Calzada JP, Crane CF, Stelly DM (1996) Apomixis: the asexualrevolution. Science 274: 1322–1323
Voigt ML, Melzer M, Rutten T, Mitchell-Olds T, Sharbel TF (2007)Gametogenesis in the apomictic Boechera holboellii complex: the maleperspective. In E Hörandl, U Grossniklaus, PJ van Dijk, TF Sharbel, eds,Apomixis: Evolution, Mechanisms and Perspectives. A.R.G. Gantner Verlag,Ruggell, pp 235–257
Voigt-Zielinski ML, Piwczy�nski M, Sharbel TF (2012) Differential effectsof polyploidy and diploidy on fitness of apomictic Boechera. Sex PlantReprod 25: 97–109
Xu Z, Wang H (2007) LTR_FINDER: an efficient tool for the prediction offull-length LTR retrotransposons. Nucleic Acids Res 35: 265–268
Yang WC, Ye D, Xu J, Sundaresan V (1999) The SPOROCYTELESS gene ofArabidopsis is required for initiation of sporogenesis and encodes anovel nuclear protein. Genes Dev 13: 2108–2117
Yang X, Makaroff CA, Ma H (2003) The Arabidopsis MALE MEIOCYTEDEATH1 gene encodes a PHD-finger protein that is required for malemeiosis. Plant Cell 15: 1281–1295
Zhang BH, Pan XP, Cox SB, Cobb GP, Anderson TA (2006) Evidence thatmiRNAs are different from other RNAs. Cell Mol Life Sci 63: 246–254
Zhao S, Fernald RD (2005) Comprehensive algorithm for quantitative real-time polymerase chain reaction. J Comput Biol 12: 1047–1064
Plant Physiol. Vol. 163, 2013 1659
Unreduced Pollen Formation in Apomicts
www.plantphysiol.orgon January 30, 2018 - Published by Downloaded from Copyright © 2013 American Society of Plant Biologists. All rights reserved.


















