
The Chemokine Receptor CXCR3 Is Degraded following Internalization
13
of April 9, 2019. This information is current as Synthesis of Receptor Replenished at the Cell Surface by De Novo Degraded following Internalization and Is The Chemokine Receptor CXCR3 Is Timothy J. Williams and James E. Pease McDonagh, Sarah J. Petit, Namita Saran, Peter C. Clark, Andrea Meiser, Anja Mueller, Emma L. Wise, Ellen M. http://www.jimmunol.org/content/180/10/6713 doi: 10.4049/jimmunol.180.10.6713 2008; 180:6713-6724; ; J Immunol References http://www.jimmunol.org/content/180/10/6713.full#ref-list-1 , 41 of which you can access for free at: cites 68 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2008 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 9, 2019 http://www.jimmunol.org/ Downloaded from by guest on April 9, 2019 http://www.jimmunol.org/ Downloaded from
Transcript of The Chemokine Receptor CXCR3 Is Degraded following Internalization

of April 9, 2019.This information is current as
Synthesis of ReceptorReplenished at the Cell Surface by De NovoDegraded following Internalization and Is The Chemokine Receptor CXCR3 Is
Timothy J. Williams and James E. PeaseMcDonagh, Sarah J. Petit, Namita Saran, Peter C. Clark, Andrea Meiser, Anja Mueller, Emma L. Wise, Ellen M.
http://www.jimmunol.org/content/180/10/6713doi: 10.4049/jimmunol.180.10.6713
2008; 180:6713-6724; ;J Immunol
Referenceshttp://www.jimmunol.org/content/180/10/6713.full#ref-list-1
, 41 of which you can access for free at: cites 68 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Chemokine Receptor CXCR3 Is Degraded followingInternalization and Is Replenished at the Cell Surface byDe Novo Synthesis of Receptor1
Andrea Meiser,2 Anja Mueller,2,3 Emma L. Wise, Ellen M. McDonagh, Sarah J. Petit,Namita Saran, Peter C. Clark, Timothy J. Williams, and James E. Pease4
The chemokine receptor CXCR3 is expressed on the surface of both resting and activated T lymphocytes. We describe in this studythe endocytosis of CXCR3 using T lymphocytes and CXCR3 transfectants. Chemokine-induced CXCR3 down-regulation occurredin a rapid, dose-dependent manner, with CXCL11 the most potent and efficacious ligand. Endocytosis was mediated in part byarrestins, but appeared to occur independently of clathrin and caveolae. In contrast to other chemokine receptors, which arelargely recycled to the cell surface within an hour, cell surface replenishment of CXCR3 occurred over several hours and wasdependent upon mRNA transcription, de novo protein synthesis, and transport through the endoplasmic reticulum and Golgi.Confocal microscopy and Western blotting confirmed the fate of endocytosed CXCR3 to be degradation, mediated in part bylysosomes and proteosomes. Site-directed mutagenesis of the CXCR3 C terminus revealed that internalization and degradationwere independent of phosphorylation, ubiquitination, or a conserved LL motif. CXCR3 was found to be efficiently internalized inthe absence of ligand, a process involving a YXXL motif at the extreme of the C terminus. Although freshly isolated T lymphocytesexpressed moderate cell surface levels of CXCR3, they were only responsive to CXCL11 with CXCL9 and CXCL10 only havingsignificant activity on activated T lymphocytes. Thus, the activities of CXCR3 are tightly controlled following mRNA translation.Because CXCR3� cells are themselves a source of IFN-�, which potently induces the expression of CXCR3 ligands, such tightregulation of CXCR3 may serve as a control to avoid the unnecessary amplification of activated T lymphocyte recruitment. TheJournal of Immunology, 2008, 180: 6713–6724.
T he chemokine receptor CXCR3 is expressed on a widevariety of cells including activated T lymphocytes, NKcells, malignant B lymphocytes, endothelial cells, and
thymocytes (1–6). Three major CXCR3 ligands, CXCL9,CXCL10, and CXCL11, have been identified, all of which areinduced by IFN-� and are therefore thought to promote Th1 im-mune responses (7–9). Recent studies have shown that the CXCR3ligands exhibit unique temporal and spatial expression patterns,suggesting that they have nonredundant functions in vivo. More-over, the CXCR3 ligands share low sequence homology (around40% amino acid identity) and exhibit differences in their potenciesand efficacies at CXCR3 with CXCL11 being the dominant ligandin several assays (8, 10). CXCR3 and its ligands have been im-plicated as playing an important role in the induction and perpet-uation of several human inflammatory disorders including athero-
sclerosis (11), autoimmune diseases (12), transplant rejection (13,14), and viral infections (15). Consequently, the mechanisms un-derlying the regulation of CXCR3 expression at the cell surface areof considerable interest.
The number of receptors on a cell surface results from a balancebetween the rate of internalization and the rate of replacement(recycling and synthesis of nascent receptor). Following ligandbinding, there are two major routes whereby G protein-coupledreceptors (GPCRs),5 as typified by chemokine receptors, are in-ternalized into cells. The first and most well-defined route involvesthe binding of arrestin to the phosphorylated receptor, which inturn initiates the internalization process by binding to clathrin. Thereceptor-arrestin complex is then sequestered in clathrin-coatedpits. This pathway is often considered a default system for degra-dation and recycling of receptors (16, 17). The second pathwayinvolves invaginations of the cell membrane known as caveolaeand functions independently of clathrin-coated pits (18). Althoughthe rate of internalization of a receptor is an important factor indetermining its level at the cell surface, the rate of recycling andthe rate of synthesis of new receptors are also important. Untilrecently, the mechanisms of the recycling process were poorlyunderstood, and internalized receptors were thought to have sev-eral potential fates. The concept of two different classes of receptor(as distinguished by their recycling) has been introduced recently,in which class A receptors traffic to recycling endosomes and are
Leukocyte Biology Section, National Heart and Lung Institute, Faculty of Medicine,Imperial College London, South Kensington Campus, London SW7 2AZ, UnitedKingdom
Received for publication March 14, 2008. Accepted for publication March 14, 2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by Grants PG/2000055 and FS/05/021 from the BritishHeart Foundation, Grant 174240 from the Arthritis Research Campaign, and ProjectGrant 076036/Z/04/Z from the Wellcome Trust.2 A. Meiser and A. Mueller contributed equally to the study.3 Current address: School of Chemical Sciences and Pharmacy, University of EastAnglia, Norwich NR4 7TJ, U.K.4 Address correspondence and reprint requests to Dr. James E. Pease, LeukocyteBiology Section, Faculty of Medicine, National Heart and Lung Institute, Sir Alex-ander Fleming Building, Imperial College London, South Kensington Campus, Lon-don SW7 2AZ, U.K. E-mail address: [email protected]
5 Abbreviations used in this paper: GPCR, G protein-coupled receptor; HA, hemag-glutinin; LAMP, lysosome-associated membrane protein; MEF, mouse embryonicfibroblast; ER, endoplasmic reticulum; WT, wild type.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

rapidly returned to the cell surface (16). In contrast, class B re-ceptors are dephosphorylated in endosomes followed by slow re-cycling back to the plasma membrane. Sequentially, the receptorspass through late endosomes and the Golgi and finally are trans-ported back to the cell surface. Another potential fate is that ofdegradation, which may be perceived to down-regulate receptorexpression. To date, protein synthesis has not been shown to playa role in GPCR replenishment (19–21).
In this study we show that CXCR3 is internalized both consti-tutively and following incubation with CXCL11, resulting in deg-radation of the receptor. We also show that in the absence of de-tectable recycling, cell surface replenishment of CXCR3 isdependent upon de novo protein synthesis.
Materials and MethodsMaterials
All chemicals unless otherwise stated were purchased from Sigma-Aldrich.Chemokines were purchased from PeproTech. Filipin, sucrose, nystatin,monensin, and nocodazole were purchased from Sigma-Aldrich or Calbio-chem. Brefeldin A, actinomycin D, and bafilomycin A1 were obtainedfrom Tocris. Cycloheximide was from ICN Biomedicals. The mouse anti-human CXCR3 mAb (clone 49801.111) and the mouse isotype-matchedcontrol IgG1 (MOPC 21 clone) were obtained from Sigma-Aldrich. Theanti-HA.11 Ab was from Covance, and the anti-�-tubulin Ab was fromAbcam. Secondary Abs were obtained from DakoCytomation. Plasmidsencoding dominant negative mutants of �-arrestin 1 and �-arrestin 2 weregifts of Dr. M. Caron (Duke University Medical Center, Durham, NC).
Plasmids encoding the fusion proteins GFP-DIII and GFP-DIII�2 were giftof Dr. A. Benmarah (Institute Cochin, Paris, France).
Cell culture and transient transfection
The murine pre-B cell line L1.2 was maintained as previously described inRPMI 1640 supplemented medium (22). L1.2 cells stably transfected withpCDNA3 containing the CXCR3A cDNA hemagglutinin (HA)-tagged atthe N terminus (10) were cultured in the same medium with the addition of1 mg/ml geneticin (G418) to maintain selection. Mutant CXCR3 constructswere generated by site-directed mutagenesis using the QuikChange mu-tagenesis kit (Stratagene) with the pCDNA3 HA-CXCR3A plasmid as tem-plate. Transient transfection of L1.2 cells with plasmids was conducted byelectroporation as previously described (23). Cells were cultured for 24 hin medium supplemented with 10 mM sodium butyrate before use to en-hance cell surface receptor expression. Mouse embryonic fibroblasts(MEFs) derived from both wild-type (WT) mice and mice deficient in�-arrestins 1 and 2 were a gift of Dr. R. Lefkowitz (Duke UniversityMedical Center, Durham NC) and were maintained as previously described(24). Transfection of MEFs was by electroporation as previously described(25). Cells were cultured overnight in medium supplemented with 10 mMsodium butyrate before use to enhance cell surface receptor expression.
For the generation of activated T lymphocytes, PBMC were isolatedfrom blood sampled from healthy donors, according to the Royal BromptonHospital Ethics Committee approved protocol as previously described (26).Lymphocytes were separated from monocytes by allowing the latter toadhere to a tissue culture flask for 2 h at 37°C and were activated by culturein the presence of 100 IU/ml IL-2 and 2 mg/ml Con A for 7–10 days.Purified T lymphocytes were isolated from whole blood using the Rosette-Sep Human T cell Enrichment Cocktail kit (StemCell Technologies),which typically gave a population �95% pure. Nucleofection of purified T
FIGURE 1. Internalization of CXCR3 by PBMCsand L1.2 transfectants. A and B, Dose-dependent natureand the respective kinetics of ligand-induced CXCR3internalization in PBMCs as determined by flow cytom-etry using a specific anti-CXCR3 mAb. The ligandsCXCL11 (f), CXCL10 (F) and CXCL9 (Œ) are shown.C and D, The levels of CXCR3 internalization inPBMCs and L1.2 transfectants, respectively, followingincubation with 50 nM CXCL11 after pretreatment inthe presence or absence of 0.4 M sucrose, 50 �M mo-nensin, 5 �g/ml filipin, and 50 �g/ml nystatin. Cell sur-face CXCR3 levels were measured as described and theuntreated control (�) is shown. ���, p � 0.001 com-pared with CXCL11 treatment alone. E, The effect offilipin pretreatment on the specific binding of 125I-CXCL11 to L1.2 CXCR3 transfectants. Data representthe mean � SEM of at least three different experiments.
6714 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

lymphocytes with plasmids encoding GFP-DIII and GFP-DIII�2 wasachieved by using an Amaxa nucleofector, according to the manufacturer’sinstructions, using program U-014, which typically gave around 60% cellviability as deduced by staining with the dye ToPro3 (Invitrogen).
Internalization assay and flow cytometry analysis
Internalization assays were essentially conducted as previously describedby Sauty et al. (27). Activated T lymphocytes or L1.2-CXCR3 cells wereincubated with serum-free medium for 1 h at 37°C and then resuspended inmedium without serum at 5 � 106 cells/ml. Cells were then incubated withchemokines (50 nM) for various times at 37°C, and washed in ice-cold PBScontaining 1% FCS and 1% NaN3 before flow cytometry analysis. Cellsurface-expressed CXCR3 was detected using an anti-CXCR3 Ab andFITC conjugated anti-mouse IgG. Samples were quantified on a FACS-Calibur, and data processed with CellQuest software (version 3.1; BD Bio-sciences) with dead cells excluded from analysis. The relative CXCR3surface expression was calculated as a percentage using the following:100 � (mean channel of fluorescence (stimulated) � mean channel offluorescence (negative control))/(mean channel of fluorescence (me-dium) � mean channel of fluorescence (negative control)). Pilot experi-ments staining CXCR3 transfectants on ice in either the presence or ab-sence of 50 nM CXCL11 confirmed that binding of ligand by CXCR3 didnot significantly reduce detection by the primary Ab (data not shown).Where inhibitors were used, cells were incubated for 1 h at 37°C withfilipin (5 �g/ml), nystatin (50 �g/ml), monensin (50 �M), sucrose (0.4 M),or cycloheximide (10 �g/ml) before assays of receptor down-regulationwere performed.
Recovery of cell surface receptor levels
Receptor down-regulation was initiated as described. After 30 min incu-bation with chemokines, the cells were washed three times in mediumwithout FCS and resuspended in medium without FCS and incubated at37°C. To remove CXCL11 from endogenous glycosaminoglycans, acti-vated T lymphocytes were washed once in prewarmed 0.5 M NaCl/RPMI1640 as previously described (27) then twice in RPMI 1640 and incubatedat 37°C. Samples were taken at different time points and cells were washedin PBS buffer containing 1% FCS and 1% NaN3. Cells were stained withAbs as described. Where inhibitors were used, brefeldin A (5 �M), acti-
nomycin D (5 �M), bafilomycin A1 (100 nM), or cycloheximide (10 �g/ml) were added to the cells during the recovery phase, following inductionof CXCR3 down-regulation by ligand.
Confocal analysis
The H9 human T cell lymphoma line was washed in RPMI 1640 andresuspended at a concentration of 5 � 106 cells/ml in serum-free RPMI1640 and were incubated at 37°C with 50 nM CXCL11. Samples wereremoved either before or at the indicated times following the addition ofCXCL11. Internalization buffer (1% FCS, 1% NaN3 in PBS) was added tothe samples removed, and tubes were incubated on ice until all time pointswere collected. Each time point was divided into either isotype control orAb-staining tubes. Cells were washed twice in cold PBS before fixation in4% paraformaldehyde for 20 min on ice. Cells were then washed in PBSand permeabilized in 0.5% saponin buffer containing anti-human lyso-some-associated membrane protein (LAMP)-1 Ab (1/20 dilution; BDPharmingen) or an equal concentration of mouse IgG1 isotype control.After incubation with the primary Ab, cells were washed in saponinbuffer before incubation with goat anti-mouse IgG Alexa Fluor 568(1/100 dilution in 0.5% saponin buffer; Invitrogen). Cells were washedand then preblocked with mouse IgG before being incubated withmouse anti-human CXCR3 FITC (1/10 dilution in 0.5% saponin; R&DSystems) or isotype control. Cells were then resuspended in 4% para-formaldehyde and were spun onto poly-L-lysine-coated glass coverslipsin 24-well tissue culture plates at 1200 rpm for 5 min. The supernatantwas removed and the coverslips washed twice in PBS and once indeionized water before being removed from the wells, allowed to airdry, and mounted onto slides in VectorShield Hardset fluorescencemounting medium (Vector Laboratories). Analysis was conducted byconfocal microscopy using a Leica TCS NT confocal microscope witha �40 oil objective. Image analysis was conducted using Leica LCSLite software version 2.61 and the images manipulated for presentationusing Adobe Photoshop version 6.0.
SDS-PAGE and Western blot analysis
L1.2 transfectants expressing WT CXCR3, and CXCR3-AAA, CXCR3-K324R, CXCR3-�4, and CXCR3-�34 constructs were washed in RPMI
FIGURE 2. Neither clathrin nor arrestinsare required for the CXCL11-induced inter-nalization of CXCR3. A–D, CXCL11-in-duced internalization of CXCR3 in purifiedT lymphocytes as deduced by flow cytom-etry following nucleofection in the presenceor absence (mock) of a plasmid containing adominant negative GFP-tagged DIII con-struct that inhibits clathrin-coated pit assem-bly. Nucleofection was conducted 24 h be-fore the induction of CXCR3 internalizationwith 50 nM CXCL11. The percentage ofcells is shown in each quadrant and the ex-periment shown is representative of threeseparate experiments. E, The relative inter-nalization of CXCR3 in WT MEF cells (�)and �-arrestin 2- or �-arrestin 3-deficientMEF cells (f), transiently transfected withCXCR3, following incubation with 50 nMCXCL11 or buffer alone. Data represent themean � SEM of four different experiments.F, The relative internalization of CXCR3 inresponse to 50 nM CXCL11 or buffer alonein a stable L1.2 transfectant cell line follow-ing transfection 24 h previously with eitherplasmid pcDNA3 (mock) or pcDNA3 con-taining the V53D and V54D dominant neg-ative mutants of �-arrestin 1 and �-arrestin2. Data represent the mean � SEM of threedifferent experiments ��, p � 0.01 and ���,p � 0.001 compared with buffer-treatedcontrols.
6715The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
1640 and resuspended at 5 � 106 cells/ml in serum-free RPMI 1640 con-taining 10 �g/ml cycloheximide. Where indicated, 40 �M MG132 or 200�M chloroquine was also added. The cells were preincubated for 30 minat 37°C before the addition of 50 nM CXCL11. Samples were taken at theindicated time points, the cells washed in ice-cold PBS and resuspended inlysis buffer containing 1% N-dodecyl-� D-maltoside, 10% glycerol, 1/1000protease inhibitor cocktail in PBS (28). Equal quantities of cell lysates wereseparated on 4–12% SDS-PAGE gels and were electrophoretically trans-ferred onto a nitrocellulose membrane, which was subsequently blockedwith 5% milk in 0.01 M PBS with 0.05% Tween 20. The blots were in-dependently probed with either anti-HA (1/1000 dilution; Covance) or anti-�-tubulin (1/10,000 dilution; Abcam) as a loading control. Followingwashing and probing with a secondary HRP-conjugated polyclonal goatanti-mouse Ig (1/1000 dilution), blots were developed by ECL (GEHealthcare).
Chemotaxis
Chemotaxis assays using either CXCR3 transfectants or purified T lym-phocytes were performed essentially as previously described (10, 29) usingChemoTX plates with a 5-�m pore size, purchased from NeuroProbe. ForT lymphocyte migration, enumeration was conducted using a hemocytom-eter and cell migration to buffer alone was subtracted from the resultingdata, with individual results expressed as a percentage of the total cellsapplied to the filter. For L1.2 transfectant chemotaxis, the same apparatus
was used, although at the end of the assay, cells were transferred from thelower chamber to a white 96-well microtiter plate using a funnel plate(NeuroProbe), and cells were detected with CellTiter Glo (Promega). Lu-minescence was measured using a TopCount microplate scintillation andluminescence counter (PerkinElmer). Data described are expressed as achemotactic index, relative to migration observed to medium alone.
Ligand binding assays125I-CXCL11 and 125I-CXCL10 were purchased from PerkinElmer LifeSciences. Ligand binding was performed as previously described usingcentrifugation through oil to separate bound chemokine from free chemo-kine (26). Data are presented following the subtraction of nonspecific bind-ing, taken as the counts obtained when the labeled chemokine was dis-placed by a 1000-fold excess of homologous cold chemokine.
Data analysis
Data were analyzed using Prism 4.0 (GraphPad Software) by ANOVA withBonferroni’s Multiple comparisons test.
ResultsDown-regulation of cell surface CXCR3 following incubationwith ligand
We initially used activated PBMCs to investigate the process ofCXCR3 down-regulation following incubation of the cells with thethree natural ligands described to date for CXCR3, namelyCXCL9/Mig, CXCL10/IP-10, and CXCL11/I-TAC. ActivatedPBMCs, cultured for 7–10 days with Con A and IL-2, readilyexpressed CXCR3 on their cell surface as detected by flow cytom-etry using a specific mAb. Incubation of PBMCs with all three
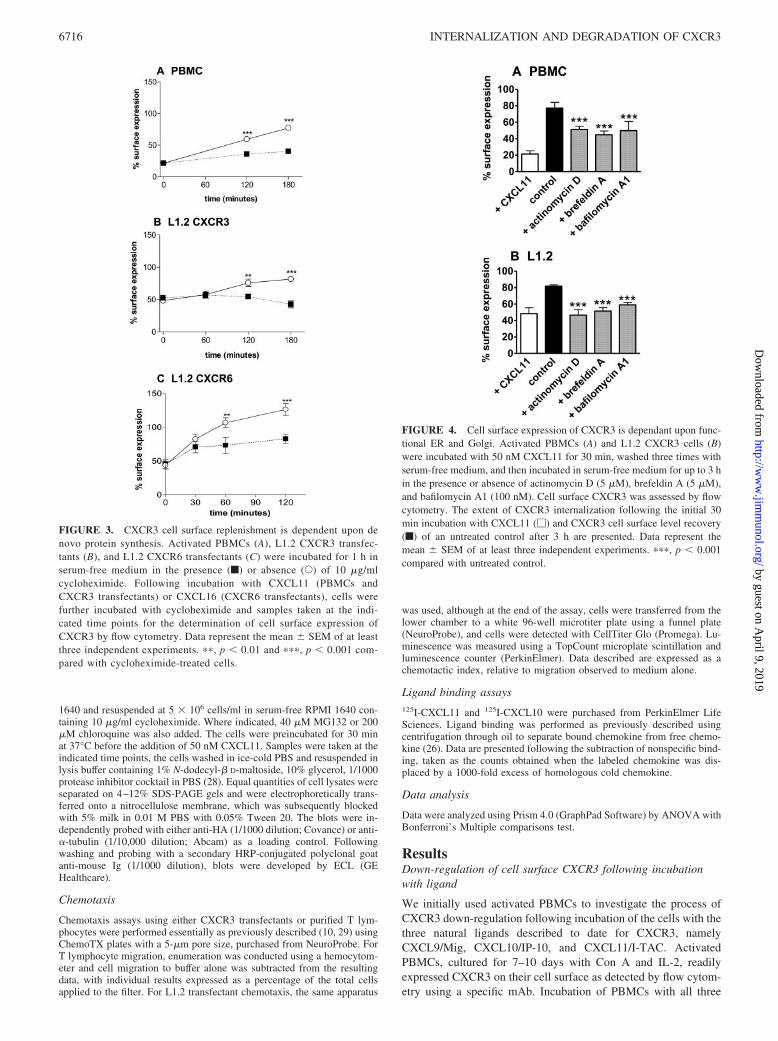
FIGURE 3. CXCR3 cell surface replenishment is dependent upon denovo protein synthesis. Activated PBMCs (A), L1.2 CXCR3 transfec-tants (B), and L1.2 CXCR6 transfectants (C) were incubated for 1 h inserum-free medium in the presence (f) or absence (E) of 10 �g/mlcycloheximide. Following incubation with CXCL11 (PBMCs andCXCR3 transfectants) or CXCL16 (CXCR6 transfectants), cells werefurther incubated with cycloheximide and samples taken at the indi-cated time points for the determination of cell surface expression ofCXCR3 by flow cytometry. Data represent the mean � SEM of at leastthree independent experiments. ��, p � 0.01 and ���, p � 0.001 com-pared with cycloheximide-treated cells.
FIGURE 4. Cell surface expression of CXCR3 is dependant upon func-tional ER and Golgi. Activated PBMCs (A) and L1.2 CXCR3 cells (B)were incubated with 50 nM CXCL11 for 30 min, washed three times withserum-free medium, and then incubated in serum-free medium for up to 3 hin the presence or absence of actinomycin D (5 �M), brefeldin A (5 �M),and bafilomycin A1 (100 nM). Cell surface CXCR3 was assessed by flowcytometry. The extent of CXCR3 internalization following the initial 30min incubation with CXCL11 (�) and CXCR3 cell surface level recovery(f) of an untreated control after 3 h are presented. Data represent themean � SEM of at least three independent experiments. ���, p � 0.001compared with untreated control.
6716 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

CXCR3 ligands induced a dose-dependent loss of CXCR3 fromthe cell surface (Fig. 1A) as deduced by staining with the sameCXCR3-specific mAb.
Notably, CXCL11 was the most efficacious ligand, with a 50 nMconcentration of CXCL11 reducing cell surface staining to �20%of their starting levels. We subsequently examined the kinetics ofthis response, using 50 nM concentrations of each ligand (Fig.1B). Loss of cell surface CXCR3 occurred rapidly, with optimaldown-regulation observed by 30 min in agreement with a pre-vious study (27). Similar data was also obtained using a previ-ously described L1.2 cell line stably expressing CXCR3 (datanot shown) with CXCL11 again the most efficacious of the li-gands, although the maximum level of receptor down-regula-tion observed was reduced to �50% of starting levels as wehave previously observed with L1.2 cells expressing the relatedreceptor CCR3 (30). This reduction likely reflects less effica-cious coupling of the human receptor to murine intracellularmachinery in the transfectants. In all subsequent experiments,we therefore incubated cells with 50 nM CXCL11 to achieveoptimal CXCR3 down-regulation.
Pathways of CXCR3 internalization
Two major pathways are known by which chemokine receptors areinternalized; either via clathrin-coated vesicles following the clath-rin-mediated endocytic pathway or via caveolae. Hypertonic su-crose medium has been shown to block the assembly of clathrin-
coated pits (31), whereas internalization via caveolae can beinhibited with filipin or nystatin (32). Monensin is an inhibitor ofvesicle acidification, a process essential for the sorting events oc-curring during endocytosis of GPCRs such as the �2-adrenergicreceptor (33). We assessed the activity of these inhibitors onCXCR3 down-regulation in PBMCs and L1.2 CXCR3 transfec-tants (Fig. 1, C and D). Neither filipin nor nystatin had any inhib-itory effect on CXCR3 down-regulation in either cell type, sug-gesting that caveolae are not involved in the endocytosis ofCXCR3. Although sucrose had little effect on ligand-inducedCXCR3 down-regulation in PBMCs (Fig. 1C), it was observed tosignificantly reduce the levels of CXCR3 internalization in L1.2CXCR3 cells following treatment with CXCL11 (Fig. 1D). In L1.2transfectants, monensin treatment significantly reduced CXCL11-induced internalization of the receptor, suggesting that vesicularacidification is necessary for the sorting events occurring follow-ing CXCR3 endocytosis. In PBMCs, monensin had a modest in-hibitory effect on CXCR3 endocytosis that did not reach statisticalsignificance. Because it has been previously demonstrated thatcholesterol and lipid rafts are required for the maintenance of che-mokine receptor conformation (34, 35), we also sought to examinethe effects of filipin and nystatin on ligand binding. Although nys-tatin treatment altered the density of CXCR3 transfectants makingthem unable be centrifuged through oil in our ligand binding assay(data not shown) treatment of cells with filipin was observed tohave little effect on CXCL11 binding (Fig. 1E).
FIGURE 5. The effects of C-ter-minal mutation upon CXCR3 func-tion. A, The amino acid identity of theC-terminal CXCR3 mutants ana-lyzed. B, The relative expression lev-els of the constructs following thetransient transfection of L1.2 cells,compared with cell surface stainingobserved with the WT CXCR3 con-struct. �, p � 0.001. C, The relativechemotactic responses to CXCL11 ofthe same CXCR3 mutants. D, depictsthe internalization of the CXCR3 mu-tants in response to increasing con-centrations of CXCL11. Comparisonswere made with untreated transfec-tants in each case.�, p � 0.001 and���, p � 0.01 compared with con-trols. E, Constitutive internalizationin the absence of ligand of both WTCXCR3 and the �4 construct over a6-h period following treatment withcycloheximide to inhibit de novo syn-thesis of receptor. All data representthe mean � SEM of at least threeexperiments.
6717The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
CXCR3 can be internalized independently of clathrin and�-arrestin 1 and �-arrestin 2
Because sucrose and monensin were without effect on CXCR3internalization in PBMCs we sought to confirm our findings byusing an alternative strategy to inhibit clathrin. T lymphocyteswere purified from blood and underwent nucleofection either inbuffer alone (mock nucleofection) or in buffer containing plasmidencoding a GFP-tagged construct, DIII (DIII transfection). Thisconstruct inhibits clathrin-coated pit assembly and therefore clath-rin-dependent internalization (36, 37). Twenty-four hours afternucleofection, cells were harvested and incubated at 37°C either inthe presence or absence of CXCL11 before staining for CXCR3expression. A significant percentage of mock-nucleofected T lym-phocytes were shown to express CXCR3 (Fig. 2A, top left quad-rant), which was seen to be reduced following CXCR3 treat-ment (Fig. 2C, top left quadrant). Nucleofection of Tlymphocytes led to the identification of two populations ofCXCR3-positive cells, a major population not expressing theDIII-GFP fusion protein (Fig. 2B, top left quadrant) and a mi-nor population expressing the DIII-GFP fusion protein (Fig. 2B,top right quadrant). CXCL11 treatment was seen to signifi-cantly reduce the number of cells within both populations (Fig.2D, top right and left quadrants). A similar lack of effect uponCXCL11-induced CXCR3 endocytosis was also seen followingnucleofection of T lymphocytes with the control protein GFP-DIII�2, which corresponds to the GFP-DIII construct lackingall AP-2 binding sites (data not shown). Collectively this sug-gests that ligand-driven endocytosis of CXCR3 in T lympho-cytes occurs independently of clathrin.
The clathrin-dependent pathway for endocytosis of GPCRs typ-ically involves the binding of arrestins to the intracellular face ofthe phosphorylated receptor. To examine whether CXCR3 inter-nalization is dependent upon arrestins, we transiently transfectedMEFs obtained from both WT and �-arrestin 1- and �-arrestin
2-deficient mice. Internalization was then induced by incubationwith CXCL11 and CXCR3 down-regulation assessed as before byflow cytometry. CXCR3 down-regulation in WT MEFs was sim-ilar to that seen in L1.2 transfectants, with CXCL11 reducing cellsurface levels to around 50% of their starting levels (Fig. 2E). InMEFs from �-arrestin 1- and �-arrestin 2-deficient mice, CXCR3down-regulation in response to ligand was observed, but at a re-duced level, with only a 20% reduction of CXCR3 cell surfacelevels in response to CXCL11, suggestive of an incomplete re-quirement for � arrestin in the down-modulation process. Simi-larly, transfection of L1.2 CXCR3 transfectants with either emptyplasmid or plasmids encoding the V53D and V54D dominant formof �-arrestin 1 and �-arrestin 2 were without effect upon CXCR3down-regulation induce by CXCL11 (Fig. 2F). Collectively, thedata suggest the existence of a �-arrestin-independent pathway forthe endocytosis of CXCR3.
Inhibition of CXCR3 cell surface replenishment
After the induction of down-regulation by ligand, the recovery ofcell surface CXCR3 levels was relatively slow in both PBMCs andL1.2 transfectants (Fig. 3, A and B), with only �70–80% recoveryof the original CXCR3 cell surface levels observed a full 3 h afterincubation with CXCL11. This was in contrast to another Th1-expressed chemokine receptor CXCR6, which showed 100% re-covery of cell surface levels within 1 h of ligand-induced down-regulation (Fig. 3C) and is typical of receptor recycling to the cellsurface as described for other chemokine receptors (19, 38, 39).The slow recovery of cell surface CXCR3 levels suggested to usthat upon ligand-induced internalization, CXCR3 is either slowlyrecycled, as is the case for class B GPCRs such as the vasopressintype 2 receptor (40) or alternatively, is degraded. In the case ofdegradation, cell surface replenishment would therefore require denovo synthesis of receptor.
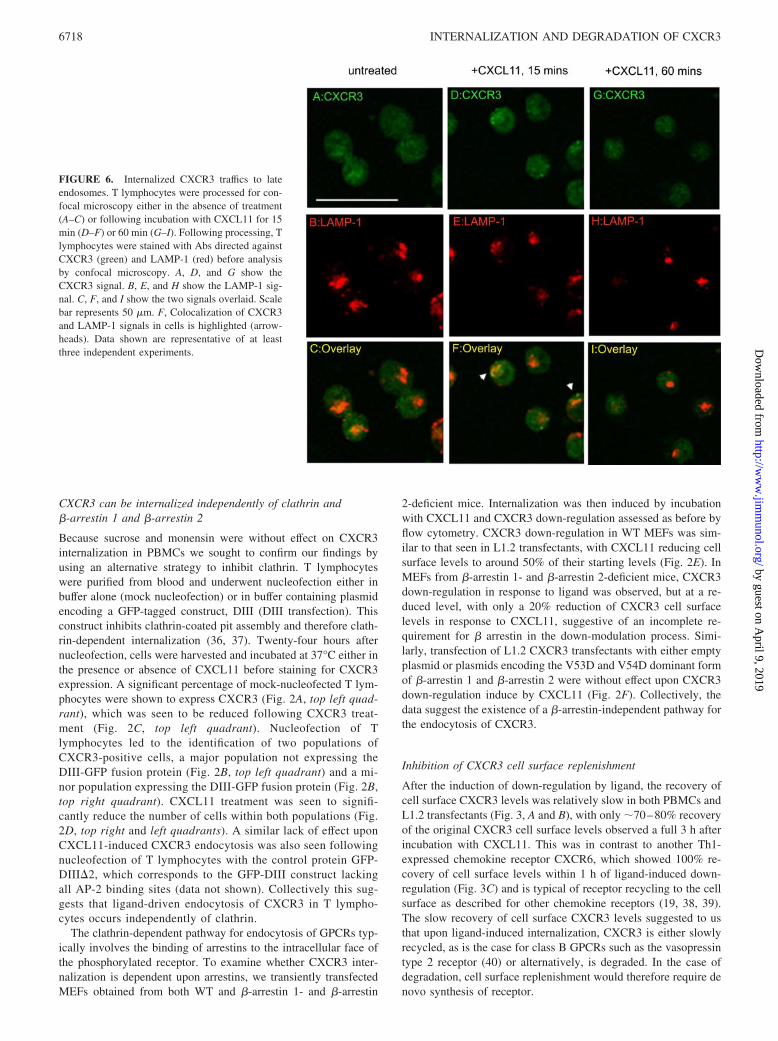
FIGURE 6. Internalized CXCR3 traffics to lateendosomes. T lymphocytes were processed for con-focal microscopy either in the absence of treatment(A–C) or following incubation with CXCL11 for 15min (D–F) or 60 min (G–I). Following processing, Tlymphocytes were stained with Abs directed againstCXCR3 (green) and LAMP-1 (red) before analysisby confocal microscopy. A, D, and G show theCXCR3 signal. B, E, and H show the LAMP-1 sig-nal. C, F, and I show the two signals overlaid. Scalebar represents 50 �m. F, Colocalization of CXCR3and LAMP-1 signals in cells is highlighted (arrow-heads). Data shown are representative of at leastthree independent experiments.
6718 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

To examine this latter hypothesis, we preincubated the cells for1 h with cycloheximide, induced CXCR3 internalization withCXCL11, and let the cells recover in the presence of cyclohexi-mide. Treatment with cycloheximide ablated the recovery ofCXCR3 in both PBMCs and L1.2 transfectants, whereas recoveryof cell surface CXCR6 levels to 80% of the starting levels wasobserved at the 2-h point. This latter value was approximately halfof the level of staining seen at the same time point with CXCR6transfectants that had not been treated with cycloheximide, sug-gesting that both recycling and de novo synthesis cooperate inmaintaining CXCR6 cell surface levels in the transfectant systemused (Fig. 3C).
Thus, in contrast to CXCR6 and other chemokine receptors de-scribed in the literature (19, 20, 41), cell surface replenishment ofCXCR3 is dependent upon de novo protein synthesis. If this pos-tulate is true, then CXCR3 cell surface replenishment should alsobe dependent upon mRNA transcription and efficient transportthrough the endoplasmic reticulum (ER) and Golgi. We thereforepreincubated PBMCs or L1.2 transfectants for 1 h in the pres-ence or absence of actinomycin D (an inhibitor of transcrip-tion), or brefeldin A and bafilomycin A1, which have beenshown to inhibit function of the ER and Golgi apparatus, re-spectively, and therefore inhibit transport of receptors throughthese compartments (41, 42). Internalization was induced with50 nM CXCL11 and the expression of CXCR3 was monitoredat 3 h post-internalization. In PBMCs, cell surface replenish-ment of CXCR3 was also significantly inhibited, although notreduced to basal levels (Fig. 4A). In L1.2 transfectants, cell
surface CXCR3 levels remained at baseline following incuba-tion with CXCL11 in the presence of actinomycin D, brefeldinA, or bafilomycin A1 (Fig. 4B). Collectively, the data suggestthat the observed recovery of CXCR3 at the cell surface isdependent upon newly synthesized receptor trafficking throughfunctional Golgi apparatus in the cell, in contrast to chemokinereceptors such as CCR4, CCR5, and CXCR6, which appear tobe replenished by a recycling mechanism (19, 20).
The role of the CXCR3 C terminus in internalization
Because the C terminus of several GPCRs has been implicated inthe internalization process, we sought to examine the role of thismotif in the internalization of CXCR3. Site-directed mutagenesisof the CXCR3 cDNA was performed to generate four mutant con-structs (Fig. 5A). The first of these mutated a triple LLL motif toAAA, thereby losing two potential LL motifs previously reportedto be involved in CXCR2 internalization (43). The second muta-tion targeted the sole intracellular lysine residue, K324. Ubiquiti-nation of internalized GPCRs has been shown to target them fordegradation, a process whereby the 74 aa ubiquitin is covalentlyattached to intracellular lysine residues. The two remaining mu-tations introduced premature stop codons within the cDNA,truncating the receptor by either 4 aa (�4 construct) or 34 aa(�34 construct). These mutations removed a YXXL motif at theextreme C terminus and the entire repertoire of C-terminalserine and threonine residues, respectively. The latter constructallowed us to examine the requirement for phosphorylation ofCXCR3 in the internalization process. All four mutants were
FIGURE 7. CXCR3 is degraded following constitu-tive or CXCL11-induced internalization. L1.2 cellstransiently transfected with plasmids encoding CXCR3were preincubated for 30 min at 37°C with 10 �g/mlcycloheximide and an aliquot was reserved (0 h timepoint). Incubation of the remaining cells was then al-lowed to proceed in the presence or absence of 50 nMCXCL11 for 3 h. Cell lysates were generated and ana-lyzed by Western blotting using anti-HA mAb (top).Blots were subsequently stripped and reprobed with ananti-�-tubulin (aT) Ab as a loading control (bottom). A,Both constitutive (untreated) and CXCL11-induceddegradation of WT CXCR3 over the 3-h time period asdeduced by a loss of immunoreactivity. B, The effects ofpreincubation of either 40 �M MG132 or 200 �M chlo-roquine on CXCL11-induced degradation. C, TheCXCL11-induced degradation of WT CXCR3 and C-terminal CXCR3 mutants. Data shown are from one ex-periment representative of three different experiments.
6719The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

transiently expressed in L1.2 cells, and cell surface expressionwas monitored by flow cytometry. All four mutants trafficked tothe cell membrane, although the �34 mutant was expressed atlevels significantly below those of WT CXCR3 (Fig. 5B). Con-versely, the �4 mutant was consistently expressed at greaterlevels than found in WT CXCR3, although this did not reachsignificance. All four constructs were able to mediate chemo-taxis of cells in response to CXCL11, with the typical bell-shaped responses optima around the 3 nM concentration (Fig.5C). Likewise, internalization of CXCR3 in response toCXCL11 was unimpaired by mutation, with the 50 and 100 nMconcentrations of ligand inducing significant internalizationcompared with untreated cells (Fig. 5D). Because the �4 con-struct appeared to be expressed at higher levels than WTCXCR3 (Fig. 5B), we postulated that CXCR3 might be inter-nalized constitutively, i.e., in the absence of ligand, and the lossof the four most C-terminal residues might inhibit this process.We subsequently examined the expression of both WT CXCR3and the �4 construct over a 6-h period, following pretreatmentwith cycloheximide to inhibit de novo synthesis. WT CXCR3was seen to be quite rapidly lost from the cell surface in theabsence of ligand, with approximately half of the original cellsurface levels of CXCR3 observed after 4 h of incubation. Incomparison, internalization of the �4 construct was less effi-cacious, with the remaining cell surface levels of mutant recep-tor at the 6-h time point significantly greater than receptor lev-els of WT CXCR3.
Endocytosed CXCR3 passes through late endosomes orlysosomes before its degradation
We subsequently turned our attention to the fate of CXCR3 fol-lowing its internalization, using confocal microscopy to examineintracellular localization of the receptor in permeablized T lym-phocytes. A predominantly granular intracellular staining patternfor CXCR3 was evident in untreated cells (Fig. 6A), identical withthat previously described by Gasser and colleagues (44). Likewise,a similar pattern was seen for staining with the late endosomemarker LAMP-1 (Fig. 6B) with little colocalization of signals seen(Fig. 6C). Treatment with CXCL11 for 15 min resulted in clus-tering of LAMP-1� vesicles with apparent colocalization ofCXCR3 with LAMP-1 in some but not all cells (Fig. 6, E and F).This result may reflect either a rapid loss of CXCR3 immunore-activity following trafficking to lysosomes or the fact that this path-way is not the sole route of CXCR3 degradation. Little, if any,colocalization of CXCR3 with LAMP-1 staining was observed incells 60 min following treatment with CXCL11, suggesting thatdegradation of CXCR3 may be complete by this point (Fig. 6,G–I).
To further examine CXCR3 degradation, we used Western blot-ting methodologies. HA-tagged CXCR3 was expressed transientlyin L1.2 cells and following pretreatment with cycloheximide toinhibit de novo synthesis, the cells were incubated for varying timeperiods in the presence or absence of CXCL11. Cell lysates werethen examined by SDS-PAGE, followed by Western blotting. As
FIGURE 8. CXCR3 function in T lymphocytesis strictly controlled. A–F, A series of experimentsto compare CXCR3 expression and function infreshly isolated T lymphocytes (day 0) and T lym-phocytes cultured for 10 days in medium supple-mented with IL-2 and Con A (day 10). A, The rel-ative expression levels of CXCR3 on T lymphocytesas deduced by flow cytometry using an anti-CXCR3Ab. ��, p � 0.01. B–D, The relative chemotacticresponses of T lymphocytes to increasing concen-trations of CXCL9, CXCL10, and CXCL11. E, Therelative induction of CXCR3 internalization onfreshly isolated T lymphocytes as deduced by flowcytometry, following incubation with 50 nM ofCXCL9, CXCL10, or CXCL11. �, p � 0.05 com-pared with controls. F, The relative levels of specificbinding of 0.1 nM 125I-CXCL10 and 125I-CXCL11by T lymphocytes. All data represent the mean �SEM of at least three experiments.
6720 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

can be seen in Fig. 7A, CXCR3 appears as a band of �50 kDa.Following 3 h of incubation at 37°C, either in buffer alone orsupplemented with CXCL11, the band representing CXCR3 wasseen to reduce considerably in intensity, suggestive of a degrada-tive fate. Additional pretreatment of CXCR3 transfectants witheither the proteosome inhibitor MG132 or the lysosomal inhibitorchloroquine, before treatment with CXCL11, was observed to in-hibit the degradative process (Fig. 7B). We subsequently examinedthe panel of four C-terminal CXCR3 mutants to examine the ef-fects of mutation upon degradation. Compared with untreatedcells, obvious degradation of each construct was observed, sug-gesting that none of the C-terminal motifs we examined are criticalfor targeting CXCR3 for degradation (Fig. 7C). Thus it appearsthat CXCR3 is readily degraded in the presence or absence ofligand, by pathways involving both the proteosomes and lyso-somes, and that ubiquitination of CXCR3 is not a fundamental partof this process.
Previous reports have detailed the strict control of CXCR3mRNA expression in freshly isolated PBMCs. Although a signif-icant proportion of freshly isolated PBMCs express CXCR3 at thecell surface, mRNA transcripts remain undetectable and the cellsare unresponsive to CXCL9 and CXCL10 in assays of chemotaxisand calcium flux (1, 2). This unresponsive phenotype is reversedfollowing culture of PBMCs for several days in a medium con-taining IL-2 and a mitogen such as PHA, and correlates withmRNA induction and increased levels of CXCR3 at the cell sur-face. We revisited these data using freshly isolated T lymphocytesused either immediately after isolation or following culture for 10days in medium supplemented with IL-2 and Con A. As previouslydescribed for PBMCs, freshly isolated T lymphocytes expressedmodest levels of cell surface CXCR3, which were significantlyup-regulated following 10 days of culture (Fig. 8A). Modest num-bers of freshly isolated cells were seen to migrate in response toincreasing concentrations of CXCL9 and CXCL10, responseswhich were significantly enhanced following 10 days of culture(Fig. 8, B and C). In contrast, chemotactic responses of bothfreshly isolated and cultured T lymphocytes to CXCL11 were ro-bust, notably at the optimal concentration of 10 nM (Fig. 8D). Thegreater efficacy of CXCL11 on freshly isolated cells was also ev-ident when internalization assays were performed, with CXCL11but neither CXCL9 nor CXCL10 inducing significant internaliza-tion of CXCR3 (Fig. 8E). Radioligand binding assays competing125I-CXCL10 and 125I-CXCL11 with homologous unlabelled li-gand (Fig. 8F) suggested a greater number of binding sites forCXCL11 on freshly isolated T lymphocytes than were evidentfor CXCL10, despite both ligands reporting to have similarnanomolar affinities at CXCR3 (8, 10, 45). Culture for 10 daysin the presence of IL-2 and Con A resulted in a trend for anincrease in the number of binding sites for both ligands, al-though this increase was not statistically significant. Thus, de-spite freshly isolated cells expressing CXCR3 at the cell sur-face, the responses to CXCR3 ligands appear to be muted, withonly CXCL11 inducing significant biological function.
DiscussionAlthough there is growing information regarding the mecha-nisms of GPCR internalization and recycling, no data concern-ing the fate of internalized CXCR3 have been published to date.Once internalized, a GPCR can experience one of two fates,namely dissociation of ligand and recycling of functional re-ceptor back to the plasma membrane or degradation. These fatesare not mutually exclusive, as CXCR4 has been shown to un-dergo both processes following engagement with ligand (46,
47). In this study, we provide several lines of evidence to sug-gest that the fate of endocytosed CXCR3 is predominantly oneof degradation, with de novo synthesis of CXCR3 required forthe recovery of CXCR3 cell surface levels. Indeed, little if anyCXCR3 was seen to reappear at the cell surface when transcrip-tion, translation, or the trafficking of nascent proteins throughthe golgi-ER was perturbed with appropriate pharmacologicalinhibitors.
Although the related receptor CXCR4 has been shown to un-dergo lysosomal degradation in a ubiquitin-dependent manner (47,48), both the lysosome and the proteosome appear to facilitate thedegradation of CXCR3, as deduced by sensitivity to both MG132,a 26 S proteasome inhibitor (49), and chloroquine, an inhibitor ofintralysosomal catabolism (50). Taken in the context of our con-focal microscopy data, we suggest that internalized CXCR3 trafficsto late endosomes or lysosomes that may communicate in part withthe proteosome for CXCR3 degradation. Cooperation betweenboth the lysosomal and proteosomal pathways has been describedfor the degradation of other receptors including the growth hor-mone receptor (51) and the IL-2R/IL-2R ligand complex (52). Inthe case of the growth hormone receptor, endocytosis occurs in theabsence of receptor ubiquitination but still requires intact protea-somal activity, suggesting that an adaptor protein targets the re-ceptor to the proteosome (53). In the case of CXCR3, ubiquitina-tion of the receptor appears not to be required for eitherinternalization or degradation, as mutation of the sole intracellularlysine residue had little effect upon either process. This may sug-gest the existence of an additional motif within the CXCR3 intra-cellular regions that targets it for degradation by this route. Alter-natively, CXCR3 may be envisaged to interact with an adapterprotein, which itself undergoes ubiquitination, targeting both pro-teins to the proteosome. Such a process has been described for thelectin Siglec-7, which is targeted to the proteosome as a complexwith SOCS-3 (54).
Two main routes have been described for the internalization ofGPCRs following their activation. The best-characterized pathwayuses clathrin-coated pits. In this pathway the phosphorylated re-ceptor is bound by arrestins and located to clathrin-coated pits,where the complex is internalized in vesicles. These vesicles aresubsequently released from the plasma membrane by dynamin andtransported to endosomes, where dephosphorylation of the recep-tor occurs and resensitized receptor is recycled to the plasma mem-brane (55). The clathrin-mediated pathway has been demonstratedfor the internalization of other chemokine receptors of the CXCclass, notably CXCR1 (56), CXCR2 (57), and CXCR4 (38). Asecond pathway of internalization depends on caveolae (58), cho-lesterol rich, highly organized membrane structures that have beenshown to be involved in the internalization of other GPCRs in-cluding the chemokine receptors CCR4 and CCR5 (19, 20). Al-though caveolae have been described in macrophages (59), there isstill some debate as to whether lymphocytes contain caveolae (60,61). Evidence for the use of either pathway of receptor internal-ization is often provided through the overexpression of dominantnegative constructs (e.g., arrestin, dynamin, and clathrin mutants)or the use of pharmacological inhibitors to invoke or preclude theuse of a particular pathway (62). In both human PBMCs and anestablished transfectant system expressing the human orthologof CXCR3 (10), cell surface levels of CXCR3 were rapidlyreduced in a concentration- and time-dependant manner follow-ing exposure to ligand. In CXCR3 transfectants, use of inhib-itors suggested that the pathway mediating ligand-induced en-docytosis did not appear to involve caveolae but involvedclathrin. In contrast, treatment of activated PBMCs with hyper-tonic sucrose did not inhibit the internalization of CXCR3, and
6721The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

the use of an inhibitor of clathrin-coated pit assembly had noeffect upon the down-regulation of CXCR3 in purified T lym-phocytes. Likewise, we found no absolute requirement for ar-restin in the internalization process. In MEFs from mice defi-cient in �-arrestins 1 and 2 (24), internalization of CXCR3 wassignificantly reduced, but not completely abolished, whereas theuse of dominant negative arrestin mutants was without effectupon CXCR3 internalization in our transfectant system. Col-lectively, our results suggest an alternative pathway for the en-docytosis of CXCR3, one that is independent of clathrin orarrestin or a combination. This finding is in agreement with aprevious study in which CXCL11-induced internalization ofCXCR3 in a transfectant system was found to occur in a dy-namin and �-arrestin 2-independent manner (28).
The cellular motifs controlling ligand-driven internalizationand targeting it for subsequent degradation remain elusive. Re-moval of potential phosphorylation sites in the C terminus bytruncation had no effect on CXCL11-induced internalization, aspreviously described for CXCR3 transfectants in both HEK-293and 300-19 cell lines (28, 63). Likewise, mutation of the LLLmotif was without effect on CXCL11-induced internalizationagain in agreement with a study using 300-19 transfectants (28)but in disagreement with a study using HEK-293 transfectantsin which some inhibition of CXCL11-induced internalizationwas observed (63). This likely reflects differences in the intra-cellular machinery to which CXCR3 is coupled in either cellsystem.
Of interest was the finding that CXCR3 is constitutively de-graded in the absence of ligand, a robust process that was me-diated to a significant extent by a canonical YXX� motif at theextreme of the C terminus. Because such motifs have been im-plicated in the sorting of transmembrane proteins to endosomesand lysosomes (64), we hypothesize that the YSGL motif in-teracts with currently unknown intracellular proteins and con-trols the constitutive internalization of CXCR3. Supportive ofour hypothesis, a distal YKKL motif within the C terminus ofthe GPCR PAR1 directs constitutive receptor internalizationthat is clathrin- and dynamin-dependent but independent of ar-restins (65, 66).
Cell surface levels of CXCR3 are tightly regulated by both con-stitutive and ligand-driven degradation and the replenishment ofcell surface CXCR3 appears not to be dependent upon recycling ashas been shown for other chemokine receptors, but upon de novosynthesis of CXCR3 protein and its subsequent transportationthrough the Golgi apparatus. To our knowledge, this is the firstexample of a GPCR in which protein synthesis is essential for thereplenishment of the receptor on the cell surface following stim-ulation with ligand. This strict control is in addition to other mech-anisms of posttranslational regulation of CXCR3 function. Al-though expressing significant amounts of CXCR3 on the cellsurface, freshly isolated T lymphocytes were poorly responsive tothe CXCR3 ligands CXCL9 and CXCL10 as previously described(2), with the exception of the ligand CXCL11. This phenotype wascorrected upon activation of the T lymphocytes by prolonged in-cubation with IL-2 and a mitogen such as Con A, a process thatcorresponds with CXCR3 mRNA induction (2), increased cell sur-face expression of the receptor, and the acquisition of robust func-tional responses to all three ligands. CXCL10 and CXCL11 havepreviously been described as allotopic ligands of CXCR3, withactivated T cells expressing a significant population of CXCR3molecules that can bind 125I-CXCL11 but not 125I-CXCL10 (45).The binding of CXCL10 is thought to be controlled at the level ofG protein coupling because treatment of cell membranes withGTP�S (guanosine 5-O-(3-thiotriphosphate) or pertussis toxin re-
sulted in a total loss of CXCL10 binding. In contrast, CXCL11 canbind to both coupled and uncoupled CXCR3 (45). Supportive ofthis idea, both resting and activated T lymphocytes were observedto bind significantly more 125I-CXCL11 than125I-CXCL10. Re-cently published data from studies using mice deficient in the Gprotein � subunits G�i2 and G�i3 found that although T lympho-cytes from mice lacking G�i2 subunits exhibited no chemotaxis toCXCR3 ligands, T lymphocytes from mice lacking G�i3 displayedsignificant increases in both migration and GTP�S binding andmigration as compared with WT T lymphocytes (67). This sug-gests that in mice, G�i2 subunits are crucial for CXCR3 signaling,and that G�i3 subunits can act as intracellular inhibitors of CXCR3function, thereby modulating CXCR3 responsiveness. Examiningour findings in the light of these data, we can hypothesize thatup-regulation of CXCR3 itself does not necessarily result in re-sponsiveness to CXCL10 and that CXCR3 function in the humanis likely be modulated at the intracellular level by interaction withG proteins.
CXCR3 has previously been reported to be expressed in an in-tracellular compartment within T lymphocytes, which can rapidlybe mobilized to the cell surface by treatment with arachidonicacid (44). This rapid, transient mobilization of receptor hasbeen postulated to enable the cells to respond timely to changesin the microenvironment in vivo. Such a capacity for increasedcell surface expression is likely to be counterbalanced by thedegradative fate of CXCR3 we describe in this study. It is note-worthy that the CXCR3 ligands are all readily induced by IFN-�(7–9) and that the Th1-polarized lymphocytes specifically at-tracted by these chemokines are themselves a source of IFN-�(68). It can be postulated that such fine tuning of CXCR3 ac-tivity by degradation of internalized receptor serves to avoid theunnecessary amplification of T lymphocyte recruitment in vivo,which would have undesirable consequences for the host. Gen-eration of an artificial CXCR3 ligand that promotes the cellulardegradation of CXCR3 in the absence of intracellular signalingmay represent an alternative strategy for the therapeutic mod-ulation of CXCR3 with potential benefit in a wide variety ofdisease processes.
AcknowledgmentsWe are grateful to Dr. Robert Lefkowitz for providing the WT and �-ar-restin-deficient MEF, to Dr. Marc Caron for the provision of plasmidsencoding arrestin mutants, and to Dr. Alexandre Benmerah for providingplasmids encoding the DIII and DIII�2 constructs. We thank ProfessorMark Marsh, University College London, and Dr. Richard Colvin, Mas-sachusetts General Hospital, for helpful discussions.
DisclosuresThe authors have no financial conflict of interest.
References1. Loetscher, M., B. Gerber, P. Loetscher, S. A. Jones, L. Piali, I. Clark-Lewis,
M. Baggiolini, and B. Moser. 1996. Chemokine receptor specific for IP10 andmig: structure, function, and expression in activated T-lymphocytes. J. Exp. Med.184: 963–969.
2. Loetscher, M., P. Loetscher, N. Brass, E. Meese, and B. Moser. 1998. Lympho-cyte-specific chemokine receptor CXCR3: regulation, chemokine binding andgene localization. Eur. J. Immunol. 28: 3696–3705.
3. Trentin, L., C. Agostini, M. Facco, F. Piazza, A. Perin, M. Siviero, C. Gurrieri,S. Galvan, F. Adami, R. Zambello, and G. Semenzato. 1999. The chemokinereceptor CXCR3 is expressed on malignant B cells and mediates chemotaxis.J. Clin. Invest. 104: 115–121.
4. Qin, S., J. B. Rottman, P. Myers, N. Kassam, M. Weinblatt, M. Loetscher,A. E. Koch, B. Moser, and C. R. Mackay. 1998. The chemokine receptorsCXCR3 and CCR5 mark subsets of T cells associated with certain inflammatoryreactions. J. Clin. Invest. 101: 746–754.
5. Van Der Meer, P., S. H. Goldberg, K. M. Fung, L. R. Sharer,F. Gonzalez-Scarano, and E. Lavi. 2001. Expression pattern of CXCR3, CXCR4,
6722 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

and CCR3 chemokine receptors in the developing human brain. J. Neuropathol.Exp. Neurol. 60: 25–32.
6. Romagnani, P., F. Annunziato, E. Lazzeri, L. Cosmi, C. Beltrame, L. Lasagni,G. Galli, M. Francalanci, R. Manetti, F. Marra, et al. 2001. Interferon-inducibleprotein 10, monokine induced by interferon �, and interferon-inducible T-cell �chemoattractant are produced by thymic epithelial cells and attract T-cell receptor(TCR) ��� CD8� single-positive T cells, TCR��� T cells, and natural killer-type cells in human thymus. Blood 97: 601–607.
7. Farber, J. M. 1990. A macrophage mRNA selectively induced by �-interferonencodes a member of the platelet factor 4 family of cytokines. Proc. Natl. Acad.Sci. USA 87: 5238–5242.
8. Cole, K. E., C. A. Strick, T. J. Paradis, K. T. Ogborne, M. Loetscher,R. P. Gladue, W. Lin, J. G. Boyd, B. Moser, D. E. Wood, et al. 1998. Interferon-inducible T cell � chemoattractant (I-TAC): a novel non-ELR CXC chemokinewith potent activity on activated T cells through selective high affinity binding toCXCR3. J. Exp. Med. 187: 2009–2021.
9. Luster, A. D., J. C. Unkeless, and J. V. Ravetch. 1985. �-Interferon transcrip-tionally regulates an early-response gene containing homology to platelet pro-teins. Nature 315: 672–676.
10. Xanthou, G., T. J. Williams, and J. E. Pease. 2003. Molecular characterization ofthe chemokine receptor CXCR3: evidence for the involvement of distinct extra-cellular domains in a multi-step model of ligand binding and receptor activation.Eur. J. Immunol. 33: 2927–2936.
11. Mach, F., A. Sauty, A. S. Iarossi, G. K. Sukhova, K. Neote, P. Libby, andA. D. Luster. 1999. Differential expression of three T lymphocyte-activatingCXC chemokines by human atheroma-associated cells. J. Clin. Invest. 104:1041–1050.
12. Sorensen, T. L., M. Tani, J. Jensen, V. Pierce, C. Lucchinetti, V. A. Folcik,S. Qin, J. Rottman, F. Sellebjerg, R. M. Strieter, et al. 1999. Expression of spe-cific chemokines and chemokine receptors in the central nervous system of mul-tiple sclerosis patients. J. Clin. Invest. 103: 807–815.
13. Hancock, W. W., B. Lu, W. Gao, V. Csizmadia, K. Faia, J. A. King, S. T. Smiley,M. Ling, N. P. Gerard, and C. Gerard. 2000. Requirement of the chemokinereceptor CXCR3 for acute allograft rejection. J. Exp. Med. 192: 1515–1520.
14. Hancock, W. W., W. Gao, V. Csizmadia, K. L. Faia, N. Shemmeri, andA. D. Luster. 2001. Donor-derived IP-10 initiates development of acute allograftrejection. J. Exp. Med. 193: 975–980.
15. Liu, M. T., B. P. Chen, P. Oertel, M. J. Buchmeier, D. Armstrong,T. A. Hamilton, and T. E. Lane. 2000. The T cell chemoattractant IFN-inducibleprotein 10 is essential in host defense against viral-induced neurologic disease.J. Immunol. 165: 2327–2330.
16. Shenoy, S. K., and R. J. Lefkowitz. 2003. Multifaceted roles of �-arrestins in theregulation of seven-membrane-spanning receptor trafficking and signalling. Bio-chem. J. 375: 503–515.
17. Pelchen-Matthews, A., N. Signoret, P. J. Klasse, A. Fraile-Ramos, and M. Marsh.1999. Chemokine receptor trafficking and viral replication. Immunol. Rev. 168:33–49.
18. Orlandi, P. A., and P. H. Fishman. 1998. Filipin-dependent inhibition of choleratoxin: evidence for toxin internalization and activation through caveolae-like do-mains. J. Cell Biol. 141: 905–915.
19. Mueller, A., E. Kelly, and P. G. Strange. 2002. Pathways for internalization andrecycling of the chemokine receptor CCR5. Blood 99: 785–791.
20. Mariani, M., R. Lang, E. Binda, P. Panina-Bordignon, and D. D’Ambrosio. 2004.Dominance of CCL22 over CCL17 in induction of chemokine receptor CCR4desensitization and internalization on human Th2 cells. Eur. J. Immunol. 34:231–240.
21. Roseberry, A. G., and M. M. Hosey. 1999. Trafficking of M2 muscarinic acetyl-choline receptors. J. Biol. Chem. 274: 33671–33676.
22. Sabroe, I., D. M. Conroy, N. P. Gerard, Y. Li, P. D. Collins, T. W. Post, P. J. Jose,T. J. Williams, C. Gerard, and P. D. Ponath. 1998. Cloning and characterisationof the guinea pig eosinophil eotaxin receptor, CCR3: blockade using a monoclo-nal antibody in vivo. J. Immunol. 161: 6139–6147.
23. Auger, G. A., J. E. Pease, X. Shen, G. Xanthou, and M. D. Barker. 2002.Alanine scanning mutagenesis of CCR3 reveals that the three intracellularloops are essential for functional receptor expression. Eur. J. Immunol. 32:1052–1058.
24. Kohout, T. A., F. S. Lin, S. J. Perry, D. A. Conner, and R. J. Lefkowitz. 2001.�-Arrestin 1 and 2 differentially regulate heptahelical receptor signaling and traf-ficking. Proc. Natl. Acad. Sci. USA 98: 1601–1606.
25. Galliera, E., V. R. Jala, J. O. Trent, R. Bonecchi, P. Signorelli, R. J. Lefkowitz,A. Mantovani, M. Locati, and B. Haribabu. 2004. �-Arrestin-dependent consti-tutive internalization of the human chemokine decoy receptor D6. J. Biol. Chem.279: 25590–25597.
26. Sabroe, I., M. J. Peck, B. Jan Van Keulen, A. Jorritsma, G. Simmons,P. R. Clapham, T. J. Williams, and J. E. Pease. 2000. A small molecule antagonistof the chemokine receptors CCR1 and CCR3: potent inhibition of eosinophilfunction and CCR3-mediated HIV-1 entry. J. Biol. Chem. 275: 25985–25992.
27. Sauty, A., R. A. Colvin, L. Wagner, S. Rochat, F. Spertini, and A. D. Luster.2001. CXCR3 internalization following T cell-endothelial cell contact: preferen-tial role of IFN-inducible T cell � chemoattractant (CXCL11). J. Immunol. 167:7084–7093.
28. Colvin, R. A., G. S. Campanella, J. Sun, and A. D. Luster. 2004. Intracellulardomains of CXCR3 that mediate CXCL9, CXCL10, and CXCL11 function.J. Biol. Chem. 279: 30219–30227.
29. Xanthou, G., C. E. Duchesnes, T. J. Williams, and J. E. Pease. 2003. CCR3functional responses are regulated by both CXCR3 and its ligands CXCL9,CXCL10 and CXCL11. Eur. J. Immunol. 33: 2241–2250.
30. Sabroe, I., A. Jorritsma, V. E. Stubbs, G. Xanthou, L. A. Jopling, P. D. Ponath,T. J. Williams, P. M. Murphy, and J. E. Pease. 2005. The carboxyl terminus ofthe chemokine receptor CCR3 contains distinct domains which regulate chemo-tactic signaling and receptor down-regulation in a ligand-dependent manner. Eur.J. Immunol. 35: 1301–1310.
31. Okamoto, Y., H. Ninomiya, S. Miwa, and T. Masaki. 2000. Cholesterol oxidationswitches the internalization pathway of endothelin receptor type A from caveolaeto clathrin-coated pits in Chinese hamster ovary cells. J. Biol. Chem. 275:6439–6446.
32. Harder, T., R. Kellner, R. G. Parton, and J. Gruenberg. 1997. Specific release ofmembrane-bound annexin II and cortical cytoskeletal elements by sequestrationof membrane cholesterol. Mol. Biol. Cell. 8: 533–545.
33. Liang, W., P. K. Curran, Q. Hoang, R. T. Moreland, and P. H. Fishman. 2004.Differences in endosomal targeting of human �1- and �2-adrenergic receptorsfollowing clathrin-mediated endocytosis. J. Cell Sci. 117: 723–734.
34. Nguyen, D. H., and D. Taub. 2002. CXCR4 function requires membrane cho-lesterol: implications for HIV infection. J. Immunol. 168: 4121–4126.
35. Nguyen, D. H., and D. Taub. 2002. Cholesterol is essential for macrophage in-flammatory protein 1 � binding and conformational integrity of CC chemokinereceptor 5. Blood 99: 4298–4306.
36. Benmerah, A., M. Bayrou, N. Cerf-Bensussan, and A. Dautry-Varsat. 1999. In-hibition of clathrin-coated pit assembly by an Eps15 mutant. J. Cell Sci. 112:1303–1311.
37. Benmerah, A., C. Lamaze, B. Begue, S. L. Schmid, A. Dautry-Varsat, andN. Cerf-Bensussan. 1998. AP-2/Eps15 interaction is required for receptor-medi-ated endocytosis. J. Cell Biol. 140: 1055–1062.
38. Signoret, N., A. Pelchen-Matthews, M. Mack, A. E. Proudfoot, and M. Marsh.2000. Endocytosis and recycling of the HIV coreceptor CCR5. J. Cell Biol. 151:1281–1294.
39. Amara, A., S. L. Gall, O. Schwartz, J. Salamero, M. Montes, P. Loetscher,M. Baggiolini, J. L. Virelizier, and F. Arenzana-Seisdedos. 1997. HIV coreceptordownregulation as antiviral principle: SDF-1�-dependent internalization of thechemokine receptor CXCR4 contributes to inhibition of HIV replication. J. Exp.Med. 186: 139–146.
40. Innamorati, G., C. Le Gouill, M. Balamotis, and M. Birnbaumer. 2001. The longand the short cycle: alternative intracellular routes for trafficking of G-protein-coupled receptors. J. Biol. Chem. 276: 13096–13103.
41. Signoret, N., T. Christophe, M. Oppermann, and M. Marsh. 2004. pH-indepen-dent endocytic cycling of the chemokine receptor CCR5. Traffic 5: 529–543.
42. Klausner, R. D., J. G. Donaldson, and J. Lippincott-Schwartz. 1992. Brefeldin A:insights into the control of membrane traffic and organelle structure. J. Cell Biol.116: 1071–1080.
43. Fan, G. H., W. Yang, X. J. Wang, Q. Qian, and A. Richmond. 2001. Identificationof a motif in the carboxyl terminus of CXCR2 that is involved in adaptin 2binding and receptor internalization. Biochemistry 40: 791–800.
44. Gasser, O., T. A. Schmid, G. Zenhaeusern, and C. Hess. 2006. Cyclooxygenaseregulates cell surface expression of CXCR3/1-storing granules in human CD4�
T cells. J. Immunol. 177: 8806–8812.45. Cox, M. A., C. H. Jenh, W. Gonsiorek, J. Fine, S. K. Narula, P. J. Zavodny, and
R. W. Hipkin. 2001. Human interferon-inducible 10-kDa protein and human in-terferon-inducible T cell � chemoattractant are allotopic ligands for humanCXCR3: differential binding to receptor states. Mol. Pharmacol. 59: 707–715.
46. Signoret, N., J. Oldridge, A. Pelchen-Matthews, P. J. Klasse, T. Tran, L. F. Brass,M. M. Rosenkilde, T. W. Schwartz, W. Holmes, W. Dallas, et al. 1997. Phorbolesters and SDF-1 induce rapid endocytosis and down modulation of the chemo-kine receptor CXCR4. J. Cell Biol. 139: 651–664.
47. Marchese, A., and J. L. Benovic. 2001. Agonist-promoted ubiquitination of the Gprotein-coupled receptor CXCR4 mediates lysosomal sorting. J. Biol. Chem. 276:45509–45512.
48. Marchese, A., C. Raiborg, F. Santini, J. H. Keen, H. Stenmark, and J. L. Benovic.2003. The E3 ubiquitin ligase AIP4 mediates ubiquitination and sorting of the Gprotein-coupled receptor CXCR4. Dev. Cell 5: 709–722.
49. Lee, D. H., and A. L. Goldberg. 1998. Proteasome inhibitors: valuable new toolsfor cell biologists. Trends Cell Biol. 8: 397–403.
50. de Duve, C., T. de Barsy, B. Poole, A. Trouet, P. Tulkens, and F. Van Hoof. 1974.Commentary: lysosomotropic agents. Biochem. Pharmacol. 23: 2495–2531.
51. van Kerkhof, P., and G. J. Strous. 2001. The ubiquitin-proteasome pathway reg-ulates lysosomal degradation of the growth hormone receptor and its ligand.Biochem. Soc. Trans. 29: 488–493.
52. Yu, A., and T. R. Malek. 2001. The proteasome regulates receptor-mediatedendocytosis of interleukin-2. J. Biol. Chem. 276: 381–385.
53. Strous, G. J., and P. van Kerkhof. 2002. The ubiquitin-proteasome pathway andthe regulation of growth hormone receptor availability. Mol. Cell Endocrinol.197: 143–151.
54. Orr, S. J., N. M. Morgan, R. J. Buick, C. R. Boyd, J. Elliott, J. F. Burrows,C. A. Jefferies, P. R. Crocker, and J. A. Johnston. 2007. SOCS3 targets Siglec 7for proteasomal degradation and blocks Siglec 7-mediated responses. J. Biol.Chem. 282: 3418–3422.
55. Conner, S. D., and S. L. Schmid. 2003. Regulated portals of entry into the cell.Nature 422: 37–44.
56. Barlic, J., M. H. Khandaker, E. Mahon, J. Andrews, M. E. DeVries,G. B. Mitchell, R. Rahimpour, C. M. Tan, S. S. Ferguson, and D. J. Kelvin. 1999.�-arrestins regulate interleukin-8-induced CXCR1 internalization. J. Biol. Chem.274: 16287–16294.
57. Yang, W., D. Wang, and A. Richmond. 1999. Role of clathrin-mediated endo-cytosis in CXCR2 sequestration, resensitization, and signal transduction. J. Biol.Chem. 274: 11328–11333.
6723The Journal of Immunology
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

58. Anderson, R. G. 1998. The caveolae membrane system. Annu. Rev. Biochem. 67:199–225.
59. Kiss, A. L., and H. J. Geuze. 1997. Caveolae can be alternative endocytoticstructures in elicited macrophages. Eur. J. Cell Biol. 73: 19–27.
60. Fra, A. M., E. Williamson, K. Simons, and R. G. Parton. 1994. Detergent-insol-uble glycolipid microdomains in lymphocytes in the absence of caveolae. J. Biol.Chem. 269: 30745–30748.
61. Fra, A. M., E. Williamson, K. Simons, and R. G. Parton. 1995. De novo forma-tion of caveolae in lymphocytes by expression of VIP21-caveolin. Proc. Natl.Acad. Sci. USA 92: 8655–8659.
62. Marchese, A., C. Chen, Y. M. Kim, and J. L. Benovic. 2003. The ins and outsof G protein-coupled receptor trafficking. Trends Biochem. Sci. 28: 369 –376.
63. Dagan-Berger, M., R. Feniger-Barish, S. Avniel, H. Wald, E. Galun,V. Grabovsky, R. Alon, A. Nagler, A. Ben-Baruch, and A. Peled. 2006. Role ofCXCR3 carboxyl terminus and third intracellular loop in receptor-mediated mi-gration, adhesion and internalization in response to CXCL11. Blood 107:3821–3831.
64. Bonifacino, J. S., and L. M. Traub. 2003. Signals for sorting of transmem-brane proteins to endosomes and lysosomes. Annu. Rev. Biochem. 72:395– 447.
65. Paing, M. M., B. R. Temple, and J. Trejo. 2004. A tyrosine-based sorting signalregulates intracellular trafficking of protease-activated receptor-1: multiple reg-ulatory mechanisms for agonist-induced G protein-coupled receptor internaliza-tion. J. Biol. Chem. 279: 21938–21947.
66. Paing, M. M., C. A. Johnston, D. P. Siderovski, and J. Trejo. 2006. Clathrinadaptor AP2 regulates thrombin receptor constitutive internalization and endo-thelial cell resensitization. Mol. Cell Biol. 26: 3231–3242.
67. Thompson, B. D., Y. Jin, K. H. Wu, R. A. Colvin, A. D. Luster, L. Birnbaumer,and M. X. Wu. 2007. Inhibition of G�i2 activation by G�i3 in CXCR3-mediatedsignaling. J. Biol. Chem. 282: 9547–9555.
68. Mosmann, T. R., and R. L. Coffman. 1989. Th1 and Th2 cells: Different patternsof lymphokine secretion lead to different functional properties. Annu. Rev. Im-munol. 7: 145–173.
6724 INTERNALIZATION AND DEGRADATION OF CXCR3
by guest on April 9, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from


















