
The Cathepsin D protease programmed Fas/APO-1 TNF-a · TheEMBOJournal vol.15 no.15 pp.3861-3870,...
10
The EMBO Journal vol.15 no.15 pp.3861-3870, 1996 Cathepsin D protease mediates programmed cell death induced by interferon-y, Fas/APO-1 and TNF-a Louis P.Deiss1, Hamutal Galinka, Hanna Berissi, Ofer Cohen and Adi Kimchi2 Department of Molecular Genetics and Virology, The Weizmann Institute of Science, Rehovot 76100, Israel 'Present address: The George Williams Hooper Foundation, Department of Biophysics and Biochemistry, University of California, San Francisco, CA 94143-0552, USA 2Corresponding author L.P.Deiss and H.Galinka contributed equally to this work A functional approach of gene cloning was applied to HeLa cells in an attempt to isolate positive mediators of programmed cell death. The approach was based on random inactivation of genes by transfections with antisense cDNA expression libraries, followed by the selection of cells that survived in the presence of the external apoptotic stimulus. An antisense cDNA fragment identical to human cathepsin D aspartic protease was rescued by this positive selection. The high cathepsin D antisense RNA levels protected the HeLa cells from interferon-y- and Fas/APO-1-induced death. Pepstatin A, an inhibitor of cathepsin D, sup- pressed cell death in these systems and interfered with the TNF-a-induced programmed cell death of U937 cells as well. During cell death, expression of cathepsin D was elevated and processing of the protein was affected, which resulted in high steady-state levels of an intermediate, proteolytically active, single chain form of this protease. Overexpression of cathepsin D by ectopic expression induced cell death in the absence of any external stimulus. Altogether, these results suggest that this well-known endoprotease plays an active role in cytokine-induced programmed cell death, thus adding cathepsin D to the growing list of proteases that function as positive mediators of apoptosis. Keywords: cathepsin D/cell death/Fas/interferon-y/TNF Introduction Recent evidence has emphasized the indispensable role of programmed cell death (PCD) in the development and maintenance of cellular homeostasis within multicellular organisms. This evolutionarily conserved process removes cells that have either finished their functional role, have the potential to be deleterious to the organism or are redundant surplus cells (Raff, 1992; Schwartz et al., 1993). The identification of the genes that initiate or execute programmed cell death is a major challenge in this field (Steller, 1995; Wyllie, 1995). Invertebrate models employing powerful genetic tools have provided the most elegant approach in isolating cell death-associated genes. Two major genes that function as positive mediators of cell death, ced-3 and ced-4, were isolated from the nematode Caenorhabditis elegans (Ellis et al., 1991; Yuan and Horvitz, 1992; Yuan et al., 1993). Subsequently, the mammaliam homologue of ced-3 was isolated and identified as the interleukin-l1B converting enzyme (ICE). ICE is a cysteine protease that causes PCD when overexpressed in mammalian cells (Miura et al., 1993). Additional members of the ICE family have been identified, including Ich-lINedd2 (Kumar et al., 1994; Wang et al., 1994), CPP32/Yama (Femandes et al., 1994, Tewari et al., 1995), Tx/Ich-2 (Faucheu et al., 1995) and Mch-2 (Femandes et al., 1995). These cysteine proteases were shown to have a central role in PCD by virtue of the findings that ectopic expression of these genes leads to cell death and blocking the activity of these proteases by specific inhibitors will suppress the apoptotic process (Tewari and Dixit, 1995). Drosophila melanogaster is a second important invertebrate model that has employed a powerful genetic screen in isolating cell death genes. Two novel genes, reaper and hid, that appear to be required for cell death during embryogenesis were rescued (White et al., 1994; Grether et al., 1995). In mammalian systems apoptotic genes, such as p53 and c-myc, were isolated and studied for many years as regulators of other cellular processes; only later was their potential function as positive mediators of cell death further revealed (Yonish-Rouach et al., 1991; Evan et al., 1992). This stemmed, in part, from the lack of functional strategies for the rescue of mammalian cell death-causing genes. A significant breakthrough was provided by the finding that PCD can be initiated by activation of various cell surface receptors. Among the external triggers of PCD, a group of cytokines play a crucial role, including diffusible cytokines [e.g. TNF-oc, interferon-y (IFN-y) and TGF-,], as well as membrane bound proteins (e.g. the ligand to the Fas/APO-1 receptor) (Laster et al., 1988; Trauth et al., 1989; Itoh et al., 1991; Lin and Chou, 1992; Novelli et al., 1994; Suda and Nagata, 1994; Deiss et al., 1995). These findings suggested that exposure of cell cultures to a specific cytokine may provide a well- controlled in vitro system for the isolation of genes that mediate PCD. Yet, it is only recently that the great potential that resides in this direction started to be exploited. One of the approaches employed the yeast two-hybrid selection system for the rescue of proteins that directly interact with a functional domain (the death domain) that constitutes the intracellular region of TNF receptor 1 and the Fas/ APO- 1 receptor. This led to the isolation of a few novel death domain-containing proteins that may function as critical mediators of PCD, including FADD/MORT- 1, RIP and TRADD (Boldin et al., 1995; Chinnalyan et al., 1995; Heu et al., 1995; Stanger et al., 1995). A second approach employed in our laboratory consisted of a functional selection of death-causing genes from cell cultures that ( Oxford University Press 3861
Transcript of The Cathepsin D protease programmed Fas/APO-1 TNF-a · TheEMBOJournal vol.15 no.15 pp.3861-3870,...

The EMBO Journal vol.15 no.15 pp.3861-3870, 1996
Cathepsin D protease mediates programmed celldeath induced by interferon-y, Fas/APO-1 and TNF-a
Louis P.Deiss1, Hamutal Galinka,Hanna Berissi, Ofer Cohen and Adi Kimchi2Department of Molecular Genetics and Virology, The WeizmannInstitute of Science, Rehovot 76100, Israel
'Present address: The George Williams Hooper Foundation,Department of Biophysics and Biochemistry, University of California,San Francisco, CA 94143-0552, USA
2Corresponding author
L.P.Deiss and H.Galinka contributed equally to this work
A functional approach of gene cloning was applied toHeLa cells in an attempt to isolate positive mediatorsof programmed cell death. The approach was basedon random inactivation of genes by transfections withantisense cDNA expression libraries, followed by theselection of cells that survived in the presence ofthe external apoptotic stimulus. An antisense cDNAfragment identical to human cathepsin D asparticprotease was rescued by this positive selection. Thehigh cathepsin D antisense RNA levels protected theHeLa cells from interferon-y- and Fas/APO-1-induceddeath. Pepstatin A, an inhibitor of cathepsin D, sup-pressed cell death in these systems and interfered withthe TNF-a-induced programmed cell death of U937cells as well. During cell death, expression of cathepsinD was elevated and processing of the protein wasaffected, which resulted in high steady-state levels ofan intermediate, proteolytically active, single chainform of this protease. Overexpression of cathepsin Dby ectopic expression induced cell death in the absenceof any external stimulus. Altogether, these resultssuggest that this well-known endoprotease plays anactive role in cytokine-induced programmed cell death,thus adding cathepsin D to the growing list of proteasesthat function as positive mediators of apoptosis.Keywords: cathepsin D/cell death/Fas/interferon-y/TNF
IntroductionRecent evidence has emphasized the indispensable role ofprogrammed cell death (PCD) in the development andmaintenance of cellular homeostasis within multicellularorganisms. This evolutionarily conserved process removescells that have either finished their functional role, havethe potential to be deleterious to the organism or areredundant surplus cells (Raff, 1992; Schwartz et al., 1993).The identification of the genes that initiate or executeprogrammed cell death is a major challenge in this field(Steller, 1995; Wyllie, 1995).
Invertebrate models employing powerful genetic toolshave provided the most elegant approach in isolating celldeath-associated genes. Two major genes that function as
positive mediators of cell death, ced-3 and ced-4, wereisolated from the nematode Caenorhabditis elegans (Elliset al., 1991; Yuan and Horvitz, 1992; Yuan et al., 1993).Subsequently, the mammaliam homologue of ced-3 wasisolated and identified as the interleukin-l1B convertingenzyme (ICE). ICE is a cysteine protease that causes PCDwhen overexpressed in mammalian cells (Miura et al.,1993). Additional members of the ICE family have beenidentified, including Ich-lINedd2 (Kumar et al., 1994;Wang et al., 1994), CPP32/Yama (Femandes et al., 1994,Tewari et al., 1995), Tx/Ich-2 (Faucheu et al., 1995) andMch-2 (Femandes et al., 1995). These cysteine proteaseswere shown to have a central role in PCD by virtue ofthe findings that ectopic expression of these genes leadsto cell death and blocking the activity of these proteasesby specific inhibitors will suppress the apoptotic process(Tewari and Dixit, 1995). Drosophila melanogaster is asecond important invertebrate model that has employed apowerful genetic screen in isolating cell death genes. Twonovel genes, reaper and hid, that appear to be requiredfor cell death during embryogenesis were rescued (Whiteet al., 1994; Grether et al., 1995).
In mammalian systems apoptotic genes, such as p53and c-myc, were isolated and studied for many years asregulators of other cellular processes; only later was theirpotential function as positive mediators of cell deathfurther revealed (Yonish-Rouach et al., 1991; Evan et al.,1992). This stemmed, in part, from the lack of functionalstrategies for the rescue of mammalian cell death-causinggenes. A significant breakthrough was provided by thefinding that PCD can be initiated by activation of variouscell surface receptors. Among the external triggers ofPCD, a group of cytokines play a crucial role, includingdiffusible cytokines [e.g. TNF-oc, interferon-y (IFN-y) andTGF-,], as well as membrane bound proteins (e.g. theligand to the Fas/APO-1 receptor) (Laster et al., 1988;Trauth et al., 1989; Itoh et al., 1991; Lin and Chou, 1992;Novelli et al., 1994; Suda and Nagata, 1994; Deiss et al.,1995). These findings suggested that exposure of cellcultures to a specific cytokine may provide a well-controlled in vitro system for the isolation of genes thatmediate PCD. Yet, it is only recently that the great potentialthat resides in this direction started to be exploited. Oneof the approaches employed the yeast two-hybrid selectionsystem for the rescue of proteins that directly interact witha functional domain (the death domain) that constitutesthe intracellular region of TNF receptor 1 and the Fas/APO- 1 receptor. This led to the isolation of a few noveldeath domain-containing proteins that may function ascritical mediators of PCD, including FADD/MORT- 1, RIPand TRADD (Boldin et al., 1995; Chinnalyan et al., 1995;Heu et al., 1995; Stanger et al., 1995). A second approachemployed in our laboratory consisted of a functionalselection of death-causing genes from cell cultures that
( Oxford University Press 3861

L.P.Deiss et al.
were continously exposed to the killing effects of acytokine, as detailed below.Our functional strategy was based on random inactiva-
tion of genes via the introduction of antisense cDNAexpression libraries prepared from cytokine-treated cells.The antisense cDNA library was expressed from an EBV-based episomal vector (named pTKO-1). The genes ofinterest were selected and cloned by virtue of the definedphenotypic change, i.e. a reduced susceptibility to cyto-kine-induced cell death, which resulted from inactivationof these genes (Deiss et al., 1991; Kimchi, 1992). Thestrategy was specifically adapted for the rescue of genesthat mediate the IFN-y-induced PCD in HeLa cells (Deisset al., 1995). Cells that survived in the continuous presenceof IFN-y after transfection with the antisense cDNA librarywere selected and the episomal vectors they carriedwere rescued. Each recombinant vector was independentlyassayed in a second round of transfection. The plasmidsthat provided some growth advantage under the negativepressure of IFN-y were then subjected to further analysis.An interferon-responsive element was introduced into thepTKO-1 vector, in order to impose high expression levelsof antisense RNA during the selection process. Thisinducible feature of the vector was, in retrospect, veryimportant in getting a reduction in the levels of thecorresponding protein during selection by IFN-y (Deisset al., 1995; Kissil et al., 1995).We have reported elsewhere on the isolation and charac-
terization of three novel genes which were rescued by thisstrategy and which were proved to function as positivemediators of cell death. One of these genes coded for anovel Ca2+/calmodulin-dependent serine/threonine kinasethat carries eight ankyrin repeats and the death domain(Deiss et al., 1995; Feinstein et al., 1995). The other twogenes directed the synthesis of a basic, proline-rich, 15 kDaprotein (Deiss et al., 1995) and of a 46 kDa protein thatcarries a P loop motif (Kissil et al., 1995). The fourthantisense cDNA that was rescued by this procedure willbe described in this work.
ResultsEstablishment of HeLa cell lines that express therescued antisense cDNA fragment whichcorresponds to cathepsin DThe gene rescue was performed as described previously(Deiss et al., 1995). Briefly, HeLa cells were transfectedwith the antisense cDNA library and subjected to thedouble selection of hygromycin B and IFN-y. After 28days the cells that survived in the presence of IFN-y werepooled and the episomal DNA was extracted from themand shuttled into bacteria. Each of the rescued pTKO-1vectors was tested in a second round of transfection andthose that conveyed some protection from IFN-y-inducedcell death were further analysed. One of the functionalclones carried a cDNA insert of 370 bp that was 98%identical to a fragment of human cathepsin D cDNA. Thefragment spanned from position 1203 to 1573 in thecathepsin D cDNA (Faust et al., 1985) and was orientedin the vector in the antisense direction.
This rescued episomal vector (pTKO- 1 -anti-cath-D) wasintroduced into HeLa cells to generate a few independenthygromycin B-resistant polyclonal populations that stably
DHFR-tl anti-cath-DIFN-2 1 7+ -t
sense -goa i a
anti sense 11
GAPDH -uiS i
Fig. 1. Transfected HeLa cells express a large excess of antisenseversus sense cathepsin D RNA. Northern analysis of total RNA(20 ,ug) prepared from DHFR- and anti-cath-D-transfected HeLa cellsat day 4 of culture in the absence (-) or presence (+) of recombinanthumam IFN-y (1000 U/ml). The blot was probed with the cDNAfragment carried by the pTKO-1 vector. The positions of the sense andantisense cathepsin D RNAs are indicated. The GAPDH mRNA levelswere used for the calibration of the RNA amounts in each slot.
express the antisense cathepsin D RNA. They were com-pared with hygromycin B-resistant polyclonal HeLa cellpopulations that were transfected, in parallel, with a controlvector, pTKO1-DHFR. The cDNA insert of the pTKO-1-anti-cath-D plasmid hybridized on a Northern blot to asingle endogenous 2.2 kb mRNA, as was expected for thecathepsin D mRNA transcript (Faust et al., 1985). In thepTKO1-anti-cath-D-transfected cells it also hybridized toa composite antisense 1.2 kb RNA (Figure 1). The latterconsisted of 370 bases of the cDNA insert and 800 basesof sequences derived from the expression cassette (Deissand Kimchi, 1991). The antisense cathepsin D RNA levelswere elevated 20-fold in response to IFN-y. This wasdue to the presence of an interferon-responsive enhancerelement in the pTKO1 expression vector. Consequently, alarge excess of antisense over sense RNA expression wasdetected during the IFN-y treatment (Figure 1). The otherpolyclonal cell populations that we constructed displayeda similar profile of antisense RNA expression during theIFN-y selection (not shown).
Antisense cathepsin D RNA and pepstatin Aprotect HeLa cells from IFN-rinduced cell deathFirst, the stable HeLa cells transfectants were tested fortheir growth sensitivity to IFN-y. We have previouslyshown that parental HeLa cells displayed a biphasic patternof response to IFN-y, in which cells first ceased toproliferate but remained viable, followed by massive celldeath with the cytological characteristics of PCD (Deisset al., 1995). One of the assays that was used to measurethe antisense RNA-mediated effects was based on neutralred dye uptake into viable cells. In the absence of IFN-y,both cell lines (DHFR- and anti-cathepsin D-transfectedcells) behaved the same and displayed identical growthcurves. This suggested that antisense RNA expression hadno effects on normal growth of cells (Figure 2Aa). Also,the extent of reduction in neutral red dye uptake duringthe first 4 days, corresponding to the cytostatic effects ofIFN-y (Deiss et al., 1995), was similar in both cell lines.This indicated that cathepsin D antisense RNA expressionalso did not interfere with the cell cycle inhibitory effectsof the cytokine. The difference between the two cell
3862

Cytokine-induced cell death involves cathepsin D
At
a C67 b ;i
~~~~c)e/v) ie (das
/A A A
_74j4t,N
H F H Aten~~.
e psta.i__+___
Fig...212.I _'.' i I ad . i -
Time (days)> Time (day-sX
en(pHFR tAati-cateatd - lIFNT-, + + + +
Pepstatin Al - + - + - +
(circles) were treated with IFN-y (1000 U/mi; filled symbols) or left untreated (open symbols). Viable cells were stained with neutral red and the dyeuptake was measured at k54o nm. Each point represents an average of quadruplicate determinations. (b) Two independent DHFR-transfectedpolyclonal cell populations (open and filled squares) and a pair of anti-cath-D-transfected polyclonal cell populations (open and filled circles) weretreated with IFN-y (1000 U/ml) or left untreated. The fraction of viable cells was determined by comparing neutral red dye uptake of IFN-y-treatedcells to non-treated cultures at the indicated time points. Each point represents the average ±- SE of quadruplicate determinations. (B) Regrowth ofviable cells after withdrawal of IFN-y (1000 U/ml) from DHFR and anti-cath-D transfectants. Cells were seeded at an initial density of 10 000 cells!cm2, treated with a combination of hygromycin B and IFN-y (1000 U/ml) for 2 weeks, washed and stained with crystal violet 7 days later.(C) Protection from IFN-y-induced cell death by pepstatin A. The HeLa cells (DHFR and anti-cath-D transfectants) were incubated for 8 days withIFN-y (1000 U/ml) either in the presence of pepstatin A (10A4 M in 0.2% DMS0) or in its absence (0.2% DMSO alone). The DHFR-transfectedcells were also tested for responsiveness to pepstatin A in the absence of IFN-y. Data are given as mean ±,- SE neutral red dye uptake fromquadruplicate samples. (D) Light microscopy of HeLa cells on day 8 of IFN-y-treatment: (a) DHFR transfectants with no inhibitor; (b) anti-cath-Dtstct - the p c p i A. M 2
3863
L.RDeiss et al.
populations became prominent later on during the IFN-y-induced cell death phase. In IFN-y-treated DHFR-trans-fected cells dye uptake dropped from day 4 on (Figure2Aa and Ab). Microscopic observations confirmed thatthis was due to massive cell death that eliminated almostall the viable adherent cells from the plates (Figure 2Da).Death was significantly (but not completely) inhibited byantisense cathepsin D RNA expression, as shown by thesustained values of the uptake (Figure 2Aa). Each of thetwo antisense cathepsin D RNA-expressing polyclonalpopulations displayed a significant increase in the fractionof cells that were stained by the vital dye during theIFN-y-induced cell death phase (Figure 2Ab).
Another way to measure protection from cell killingconsisted of counting the number of colonies that wereformed after releasing the cultures from long-term treat-ment with the cytokine. The reduced susceptibility of theantisense-transfected cells to cell killing by IFN-y wasmanifested by a 1-2 log increase in the number of cellsthat could form colonies following the removal of IFN-yfrom treated cell cultures (Figure 2B).
To further explore the participation of cathepsin D inIFN-y-induced PCD, the HeLa cells were incubated withpepstatin A, a specific inhibitor of aspartic proteases(Shields et al., 1991). Due to the fact that cathepsin D isthe major intracellular aspartic protease in cells, theoutcome of the intracellular effects of this pentapeptideare commonly attributed to specific inhibition of cathepsinD activity. Pepstatin A was added to the culture mediumat a final concentration of 10- M in 0.2% DMSO, inaccord with previous reports in which similar incubationprotocols led to effective intracellular concentrations ofthe drug (Shields et al., 1991). Pepstatin A had no effectson growing HeLa cells that were not treated with IFN-y.Addition of pepstatin A to the IFN-y-treated DHFR-transfected cells inhibited, to some extent, the killingprocess, as reflected by the elevated values of neutral reddye uptake (Figure 2C). The highest values of dye uptakethat could be measured in the presence of IFN-y wereobtained by applying the pepstatin A to the antisensecathepsin D RNA-expressing cells (Figure 2C). Micro-scopic examination of the IFN-y-induced cell cultures thatwere protected by the double treatment (antisense RNA +pepstatin A) revealed that the majority of cells displayedthe normal adherent phenotype, whereas only -20% ofthe cells had a round apoptotic morphology (Figure 2Db).This further indicated that the combined reduction of bothexpression and activity of this endoprotease was mosteffective in protecting cells from IFN-y-induced cell death.In summary, the antisense RNA and pepstatin A datasuggest an active role for cathepsin D in IFN-y-mediatedPCD.
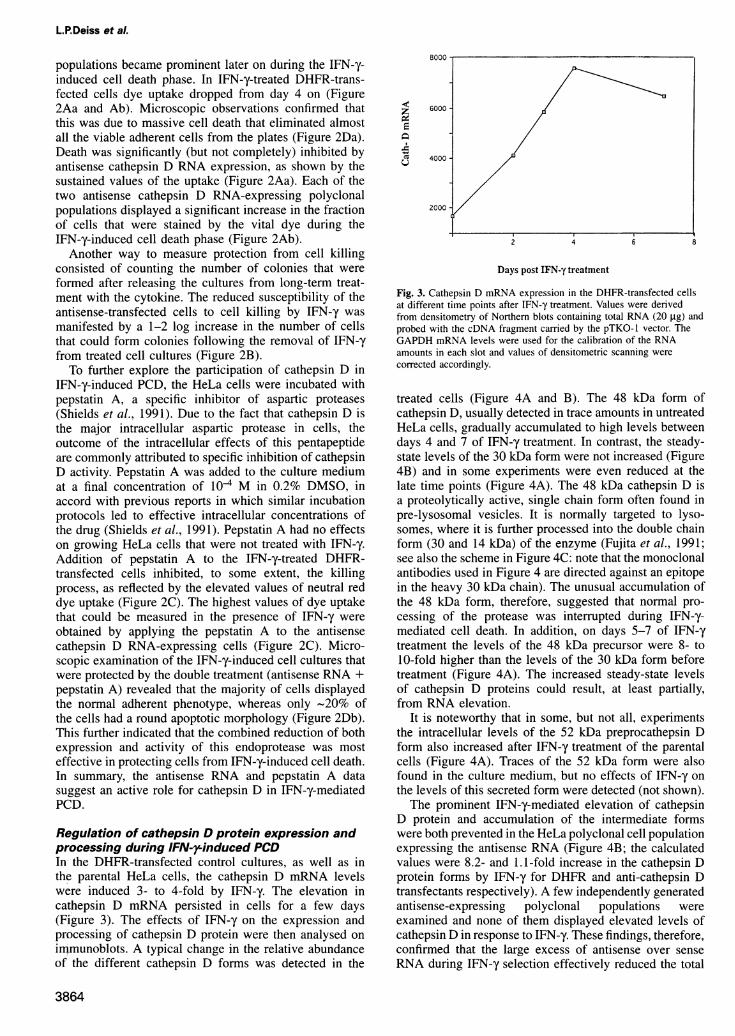
Regulation of cathepsin D protein expression andprocessing during IFN-rinduced PCDIn- the DHFR-transfected control cultures, as well as inthe parental HeLa cells, the cathepsin D mRNA levelswere induced 3- to 4-fold by IFN-y. The elevation incathepsin D mRNA persisted in cells for a few days(Figure 3). The effects of IFN-y on the expression andprocessing of cathepsin D protein were then analysed onimmunoblots. A typical change in the relative abundanceof the different cathepsin D forms was detected in the
8000
06000
'; 4000
2000
2 4 6 8
Days post IFN-y treatment
Fig. 3. Cathepsin D mRNA expression in the DHFR-transfected cellsat different time points after IFN-y treatment. Values were derivedfrom densitometry of Northern blots containing total RNA (20 ,ug) andprobed with the cDNA fragment carried by the pTKO-1 vector. TheGAPDH mRNA levels were used for the calibration of the RNAamounts in each slot and values of densitometric scanning werecorrected accordingly.
treated cells (Figure 4A and B). The 48 kDa form ofcathepsin D, usually detected in trace amounts in untreatedHeLa cells, gradually accumulated to high levels betweendays 4 and 7 of IFN-y treatment. In contrast, the steady-state levels of the 30 kDa form were not increased (Figure4B) and in some experiments were even reduced at thelate time points (Figure 4A). The 48 kDa cathepsin D isa proteolytically active, single chain form often found inpre-lysosomal vesicles. It is normally targeted to lyso-somes, where it is further processed into the double chainform (30 and 14 kDa) of the enzyme (Fujita et al., 1991;see also the scheme in Figure 4C: note that the monoclonalantibodies used in Figure 4 are directed against an epitopein the heavy 30 kDa chain). The unusual accumulation ofthe 48 kDa form, therefore, suggested that normal pro-cessing of the protease was interrupted during IFN-y-mediated cell death. In addition, on days 5-7 of IFN-ytreatment the levels of the 48 kDa precursor were 8- to10-fold higher than the levels of the 30 kDa form beforetreatment (Figure 4A). The increased steady-state levelsof cathepsin D proteins could result, at least partially,from RNA elevation.
It is noteworthy that in some, but not all, experimentsthe intracellular levels of the 52 kDa preprocathepsin Dform also increased after IFN-y treatment of the parentalcells (Figure 4A). Traces of the 52 kDa form were alsofound in the culture medium, but no effects of IFN-y onthe levels of this secreted form were detected (not shown).The prominent IFN-y-mediated elevation of cathepsin
D protein and accumulation of the intermediate formswere both prevented in the HeLa polyclonal cell populationexpressing the antisense RNA (Figure 4B; the calculatedvalues were 8.2- and 1.1-fold increase in the cathepsin Dprotein forms by IFN-y for DHFR and anti-cathepsin Dtransfectants respectively). A few independently generatedantisense-expressing polyclonal populations wereexamined and none of them displayed elevated levels ofcathepsin D in response to IFN-,y. These findings, therefore,confirmed that the large excess of antisense over senseRNA during IFN-y selection effectively reduced the total
3864

Cytokine-induced cell death involves cathepsin D
AIFN--r - + _
Bt ± -
days: 1 1 2 3 4 5 6 7
52 Kd --48 Kue-
1 5 1 5
r--- ------ --- -
_0 - 4 K c-
30 Kd -- -
SO(D ..--_ _ . - -_ -
CKii-Iv- .- -e r)Sr
_ _ - ---SOD
D 1 2 3
- - 48Kd
-03 J4 sIinecr'ia ir formT_l- law _~ - 3OKd
K.d-t.-14Kdi doutIe chain fiorm -- - - Vinclin
Fig. 4. Regulation of expression and processing of cathepsin D protease by IFN-y and TNF-ax. (A and B) Immunoblot analysis of cathepsin D formsbefore and after treatment with IFN-y (1000 U/ml). Cell lysates were prepared at the indicated time points from parental HeLa cells (A) and fromDHFR and anti-cath-D transfectants (B). Samples of 300 ,ug were fractionated on SDS-PAGE (12%), blotted to nitrocellulose and detected using theECL system (Amersham). The sizes of cathepsin D forms are shown. The same blots were reacted with polyclonal antibodies generated againstcopper/zinc superoxide dismutase (SOD) to correct for possible differences in protein amounts in each slot. (C) A scheme that depicts the differentsteps of cathepsin D processing as previuosly reported for rat cathepsin D (Fujita et al., 1991). (D) Immunoblot analysis of cathepsin D forms before(lane 1) and after treatment of U937 cells with TNF-ax (lanes 2 and 3, 24 and 48 h respectively).
levels of cathepsin D protein, as was expected. Thequestion as to why the residual levels that continued tobe expressed in these IFN-y-treated cells did not accumu-late as intermediate forms of cathepsin D is still open.
Cathepsin D aspartic protease mediates APO- 1iFasand the TNF-a-induced PCDWe studied whether cathepsin D protease is also involvedin other apoptotic systems triggered by the activation ofcell surface receptors that differ from the IFN-y receptor.The different HeLa cell transfectants were treated withthe agonistic anti-APO- 1 monoclonal antibody, in orderto determine whether cathepsin D mediates Fas/APO- 1-induced apoptosis. The parental and DHFR-transfectedcells were efficiently killed by anti-Fas/APO-1 antibodies.Cell death exhibited features characteristic of apoptosis,similar to the IFN-y effects. By 40 h ~70% of the cellsrounded up and detached from the plates (not shown) andthe uptake of neutral red dye was reduced accordingly(Figure 5A). The killing required a short pretreatment ofthe cells with a low dosage of IFN-y (50 U/ml), whichhad no effect by itself on cell viability. The low dosageof IFN-y sensitized the cells to killing by the agonisticantibody, due to elevation of Fas/APO-1 expression (Itohet al., 1991; further morphological and molecular detailson the system will be published elsewhere). Expressionof antisense cathepsin D RNA, or alternatively the additionof pepstatin A to the culture medium of the DHFR-transfected cells, substantially suppressed Fas/APO- 1-mediated cell death, resulting in an increased fraction ofviable cells (Figure SA). The latter indicated that cathepsinD is essential for Fas/APO-1-induced PCD.We also found that pepstatin A interfered with the
apoptotic process triggered in U937 histiocytic lymphomacells by tumour necrosis factor-a (TNF-ux) (Wright et al.,
1992). The killing in this system was very rapid andcharacterized by typical nuclear changes, such aschromatin condensation followed by fragmentation. DAPIstaining of U937 nuclei indicated that 6 h after TNF-axadministration approximately a third of the cell populationalready contained nuclei with typical fragmentedchromatin (Figure SB). Addition of pepstatin A to theculture showed a significant reduction in the number offragmented nuclei (Figure SB). Interestingly, the earlierstep of chromatin condensation seemed less susceptibleto the effect of pepstatin A. These data indicated thatcathepsin D endoprotease also mediates some criticalsteps along the apoptotic pathway which leads to U937cell death.
Examination of the pattern of cathepsin D expressionin TNF-a-treated U937 cells revealed that it shared a fewcommon features with the HeLa cell system. The totallevels of cathepsin D proteins were significanly increased.Moreover, the proteolytically active 48 kDa intermediateform accumulated in TNF-oc-treated U937 cells, indicatingthat, again, processing into the double chain form wasinterrupted (Figure 4D). Yet, in contrast to the HeLa cellsystem, this conversion was not completely blocked anda mild increase in the 30 kDa form was detected as well.These data suggest a common pattern of changes in theexpression/processing of cathepsin D protein in a fewapoptotic systems.
Ectopic expression of cathepsin D is notcompatible with cell viabilityThe outcome of overexpression of cathepsin D was directlymeasured in HeLa cells by co-transfections with the lacZgene, used as a marker of gene expression. Cathepsin Dwas driven by the tetracycline-repressible promoter(Gossen and Bujard, 1992) and the P-galactosidase gene
3865

B
+ +_ +
fragmented nuclei
<1
33.7±8.3+
+
8.8±5.4
Fig. 5. Involvement of cathepsin D protease in Fas/APO- 1- and TNF-a-mediated cell death. (A) Suppression of Fas/APO- 1-mediated cell death byanti-cathepsin D RNA or by pepstatin A. The HeLa cells (DHFR and anti-cath-D transfectants, 20 000 cells/microtiter well) were exposed to anti-APO-1 antibodies for 40 h as described in Materials and methods. Pepstatin A (104 M in 0.2% DMSO) was added where indicated to the DHFRtransfectants 20 h before their exposure to the anti-APO- 1 antibodies. Viability was assessed by neutral red assay in quadruplicate samples; resultsare expressed as percentage dye uptake at the end of each treatment out of the total uptake in the corresponding control wells, which were notexposed to the antibodies (100% viability). (B) Pepstatin A interferes with TNF-a-induced apoptotic cell death in U937 cells. The cells were seededat a density of 2x105 cells/ml 24 h after their preincubation with pepstatin A (104 M in 1% DMSO) or with DMSO alone. Where indicated, TNF-(x(100 U/ml, 10 ng/ml) was added and 6 h later samples were cytospun on glass slides and stained with DAPI (0.5 .g/ml; Sigma). Microscopy wasperformed under fluorescent light conditions. Magnification X 1000. Nuclei with fragmented chromatin are indicated by arrows; empty arrowheadspoint to mitotic nuclei. Data are presented as the percentage ± SE of cells with a fragmented nuclear morphology. For each condition a minimum of400 cells in 14 separate fields were scored.
was driven by the CMV constitutive promoter. The mor-phology of lacZ-containing blue cells was determined 48 hpost-transfection, in cultures which were maintained in theabsence of tetracycline to allow cathepsin D transcription/translation. It was found that 70% of the lacZ-containingcells displayed a round apoptotic phenotype upon co-transfections with cathepsin D, whereas co-transfectionswith the control tetracycline vector displayed a backgroundof <20% apoptotic cells (Figure 6A-C). In order to furtherquantitate the effects of ectopic expression of cathepsin Don cells, in a second independent approach co-transfectionswere performed with vectors expressing the secretedalkaline phosphatase (SEAP) instead of lacZ. In theseexperiments we measured the outcome of tetracyclinewithdrawal on SEAP activity released by transfectantscarrying the cathepsin D gene. We found that activationof cathepsin D by tetracycline withdrawal significantlyreduced the SEAP activity secreted into the culture mediumaround 48 h post-transfection, as compared with the valuesobtained from the same population maintained in thepresence of tetracycline (Figure 6D). In contrast, tetra-cycline withdrawal had no effect on SEAP activity releasedby control cultures co-transfected with the empty vector.
Discussion
Cathepsin D has multiple functions in different cellularcompartments. The best known function is the classicallysosomal activity that participates in the proteolysis ofendocytosed proteins. In addition, cathepsin D has an
endosomal activity that is involved in the proteolyticactivation of secreted proteins, such as growth factors andhormones (Diment et al., 1989). It also has an extracellularactivity, carried by the secreted 52 kDa form, which hasbeen implicated in controlling the migration of metastaticcells (Mignatti and Rifkin, 1993) and which could alsobe involved in the control of cell migration during normaldevelopment. This endoprotease has been implicated insome pathological disorders, such as degenerative brainchanges, connective tissue diseases and muscular dys-trophy (Matus and Green, 1987; Kenessey et al., 1989;Johnson et al., 1991). In some of these disorders abnormalproteolytic activity of cathepsin D was correlated withincreased degradation of cytoskeletal elements such as
MAP-2. Suprisingly, the phenotype of recently describedmice deficient for cathepsin D reflected only a smallportion of these multiple functions. The latter may exclus-
L.P.Deiss et al.
A
2) -
0-i
Anti Fas/APO-1Anti-cath-DPepsatia]n A
3866
*...
E
.)

Cytokine-induced cell death involves cathepsin D
C 80 -
-- 60ae )
o
0
0~
o20-cS 2C
0 control vector
B.4
*
* 4> D
44~
r .c
ei #
-
.0
4.AL)
co
wC,)
i 0-
100
80-
60-
40-
20-
0 control vector
Fig. 6. Ectopic expression of cathepsin D reduces cell viability. (A and B) X-Gal staining of HeLa cells co-transfected with lacZ (driven by theCMV promoter) and either with the cathepsin D cDNA (driven by a tetracycline-repressible promoter) or with the control vector. In both cases cellswere cultured in the absence of tetracycline and stained after 48 h with X-Gal solution for 3 h. Light microscopic micrographs are shown.Magnification X200. Examples of normal blue stained cells (A) and of apoptotic blue stained cells (B) are indicated by arrows. (C) The frequencyof blue cells with an apoptotic rounded morphology was assessed by counting 800 total blue cells from eight different fields coming from duplicatetransfections (described in A and B). (D) Assessment of secreted alkaline phosphatase (SEAP) in the growth medium of HeLa cells co-transfectedwith SEAP and with either the control vector or the above mentioned cathepsin D vector; each transfection was divided into two plates, one ofwhich was immediately supplemented with tetracycline (1.5 ,ug/ml). SEAP activity secreted into the growth medium during the last 5 h of incubationwas determined 48 h after transfections. Data on SEAP activity were obtained in duplicate from four experiments. The values give the percentage ofSEAP activity measured in the absence of tetracycline out of total activity produced in the presence of tetracyline.
ively refer to functions which were rate limiting andnot compensated for by other mechanisms. Thus thehomozygous knockout mice exhibited progressive atrophyof the intestinal mucosa, probably due to cell migrationabnormalities. They also displayed a profound destructionof lymphoid cells, probably due to deficiencies in theendosomal processing of secreted death-protective growthfactors. Lysosomal bulk proteolysis, however, was main-tained in these mice, suggesting that the main well-known function of this protease was masked by some
compensatory mechanisms (Saftig et al., 1995).A novel facet in the function of cathepsin D is docu-
mented here for the first time, using cell cultures ofepithelial and haematopoietic origin that have beenexposed to a well-defined external stress. We propose thatcathepsin D mediates a regulated type of programmed celldeath, initiated by various cytokines. The first clue was
provided by the rescue of a cathepsin D cDNA fragmentas a functional element in a genetic screen set to isolate
positive mediators of IFN-y-induced apoptosis in HeLacells. This suggested that cathepsin D must be a ratelimiting step in the commitment or execution of PCD, atleast in the model cell system in which the gene rescue
was performed. The involvement of this protease in celldeath was then further established and extended to othercytokines, including Fas/APO-1 ligand and TNF-a, withthe help of a second functional tool, i.e. the cathepsin Dpentapeptide inhibitor. It was found that the inactivationof cathepsin D by pepstatin A protected cells fromcytokine-induced cell death in a fashion similar to theprotection conferred by antisense RNA expression. InHeLa cells the combined reduction of both expressionand activity of this endoprotease was most effectivein protecting cells from IFN-y-induced cell death. Thecathepsin D connection to cell death was then furthersupported by the finding that expression and processingof the endogenous protease were both regulated duringPCD. Either the increased protein expression levels per se
3867
(7
40 14
.-,. -1 rI...~~~~~~~~~~~1 le.
-F
Tiz.
cath-D vector
,/ I
k.
.4.V 4; gbdh
irw
s"
-Arvi

L.RDeiss et al.
or the abnormal accumulation of the intermediate activeform of cathepsin D could be relevant to the process.Finally, another line of experiments showed that ectopicexpression of the cathepsin D gene induced the death ofHeLa cells in the absence of any external stimulus. Thelatter data are consistent with a role of cathepsin D as apositive mediator of programmed cell death, but definitelydo not provide by themselves independent evidence, dueto the non-specific effects that could accompany theoverexpression of a protease.
It will be interesting to determine in the future theprecise intracellular localization of cathepsin D duringcell death. For example, to find out whether it continuesto be localized in vesicular compartments or may bereleased into the cytosol. The block in processing into thedouble chain form of cathepsin D could result fromalterations in targeting to lysosomes during cell death orfrom some leakage into the cytoplasm from endosomalvesicles. Determination of the intracellular localization ofcathepsin D will be important for studying the proteinsubstrates during the death process. Also, it will beinteresting to define the spectrum of apoptotic systems inwhich cathepsin D functions as a positive mediator. Forinstance, the involvement of cathepsin D could berestricted to apoptotic systems which are exclusivelytriggered by the interaction of cytokines with their cognatereceptors or, alternatively, may be a common target ofseveral external and internal stimuli. Systems in whichcell death is triggerd by p53 activation, DNA damageagents, steroids, cytokine deprivation or deregulated c-mycshould be examined in this respect.Members of the ced-3/ICE family of cysteine proteases
have thus far been implicated in various Fas/APO- 1-mediated apoptotic systems (Enari et al., 1995; Los et al.,1995). This emerged from the finding that transfectionswith crmA, a cowpox viral gene that inhibits the ICEprotease, protected cells from Fas/APO-1-mediated celldeath. In addition, the tetrapeptide YVAD, which specific-ally inhibits the ICE family of proteases, also interferedwith Fas/APO-1-mediated programmed cell death. Therelevance of other proteases in some well-defined apoptoticprocesses, e.g. granzyme B, which mediates cell death bycytotoxic T lymphocytes (Heusel et al., 1994), has alsobeen previously documented. Obviously the functionalposition of cathepsin D with respect to the ICE-likeproteases along the Fas/APO- 1-induced apoptotic path-ways must be pursued in the future. Altogether, we proposethat this aspartyl protease should be added to the growinglist of proteases that mediate programmed cell death.
Materials and methodsCell lines and cytokine treatmentThe HeLa human epithelial carcinoma cells were grown in DMEM(BioLab) and the U937 human histiocytic cells in RPMI (BioLab).Growth medium was supplemented with 10% fetal calf serum (BioLab),4 mM glutamine, 100 U/ml penicillin and 0.1 mg/ml streptomycin.Recombinant human interferon-y (3X 107 U/ml) was purchased fromPeproTech. Recombinant TNF-oc (5x 106 U/ml) was a kind gift of CetusCorporation. Hybridoma supernatant of anti-APO-1 antibodies (5 jgIgG3/ml; Trauth et al., 1989) was kindly provided by P.H.Krammer.HeLa cells were first pretreated with 50 U/ml IFN-y for 20 h and werethen exposed to the anti-APO-l antibody at 38°C for an additional 40 h(50 ng/ml).
For DAPI staining, U937 cells were cytospun on glass slides and
fixed with 3% paraformaldehyde. The cells were washed in phosphate-buffered saline (PBS) and stained with DAPI (0.5 jig/ml; Sigma) for10 min, washed in PBS, drained and mounted in Mowiol.
Rescue of antisense cDNAs and transfectionsTransfection of the antisense cDNA library into HeLa cells was doneby the standard calcium phosphate technique. The pTKO- I vectorscarrying antisense cDNA fragments were isolated as described (Deissand Kimchi, 1991; Deiss et al., 1995). A few secondary polyclonalHeLa cell populations expressing the cathepsin D antisense cDNAfragment from the pTKO-1 vector were generated. This was performedby transfection of subconfluent monolayers of HeLa cells with 40 jig ofthe corresponding plasmid (named pTKO-1-anti-cath-D). HeLa cellswere transfected in parallel with a control vector, pTKO- 1-DHFR (Deissand Kimchi, 1991). Pools of 104 independent stable clones were generatedfrom each transfection. The stable transfectants were continously grownin the presence of 200 jig/mI hygromycin B (Calbiochem).
Neutral red dye uptake assayThe HeLa cells were cultivated in 96-well microtiter plates at an initialnumber of 15 000 or 20 000 cells/well and were treated with eitherIFN-y or anti-APO-1 antibodies respectively or were left untreated.Where indicated, pepstatin A (pepA) (Sigma) or DMSO were added tothe culture medium. The culture medium and drugs were replaced every3-4 days. Viable cells were stained with neutral red (Sigma) as detailedbefore (Wallach, 1984). The dye uptake was measured in quadruplicatesat X540 nm using an automated Micro-Elisa auto-reader.
RNA analysisTotal cellular RNA was extracted using Tri-ReagentTM (MolecularResearch Center Inc.). Samples of 20 ,ug total RNA were processed onNorthern blots as previously described in detail (Yarden and Kimchi,1986). DNA fragments used as probes were purified from agarose gelswith the Geneclean kit (BIO 101 Inc.). The fragments were labelledwith 5 jiCi [ct-32P]dCTP (3000 Ci/mmol; Amersham) using a RandomPriming kit (Boehringer).
Protein analysisCells were extracted in RIPA (10 mM Tris-HCI, pH 7.2, 150 mM NaCI,1% Triton X-100, 0.1% SDS, 1% deoxycholate and 5 mM EDTA)containing a mixture of protease and phosphatase inhibitors (1 mMphenylmethylsulfonyl fluoride, 4 jig/ml aprotenin, 100 ,ug/ml leupeptin,1.5 jtg/ml pepstatin A, 2 jig/ml antipain, 2 jig/mi chymostatin, 0.1 mMNaVO3 and 0.1 mM NaF). Protein concentration was determined usinga protein assay reagent (Bio-Rad). Aliquots of 300 jig of the celllysates were fractionated by SDS-PAGE (12%). The proteins were thenelectroblotted onto a nitrocellulose membrane and blots were incubatedin blocking solution [10% skimmed milk and 0.05% Tween-20 (Sigma)in PBS] for 2 h at room temperature and then reacted with antibody-containing solution for 18 h at 4°C. The washed membranes wereincubated with peroxidase-conjugated second antibodies, either goatanti-mouse IgG [IgG(H+L) chains; Jackson Immuno Research Labora-tories] at a 1:10 000 dilution or protein A conjugated to horseradishperoxidase (Amersham) at a 1:10 000 dilution. Detection of the boundantibodies was carried out using ECL detection reagents (Amersham).The anti-cathepsin D monoclonal antibodies (EURO/DPC UK) wereused at 1:5 dilution; these antibodies recognize an epitope in the30 kDa heavy chain. Polyclonal antibodies against copper/zinc superoxidedismutase were used at a 1:250 dilution. These antibodies were kindlyprovided by Y.Groner.
Transient transfectionsCathepsin D cDNA insert (2176 bp, a SalI-EcoRI fragment containingthe full-length coding sequences and flanking non-coding regions;see Faust et al., 1985) was subcloned into the tetracycline-controlledexpression vector (pSBC-TtA) (Dirks et al., 1993) The vector (40 jig)was transiently transfected into a HeLa cell clone (HtTA- 1) that expressesthe tetracycline transactivator gene by the standard calcium phosphatetechnique (2 x 105 cells were seeded in 9 cm plates 18-20 h prior totransfection). An empty tetracycline promoter-containing vector wasused as a control in the assays. In order to exclusively follow thetransfected cells, these constructs were co-transfected with either theCMV-,B-galactosidase fusion (Clontech) or with the SEAP geneexpressed from the SV40 promoter (the pSBC-2 vector) (Dirks et al.,1993). The molar ratio was 6:1 in favour of the tetracycline vectors.Each transfection was divided into two plates, one of which was
3868

Cytokine-induced cell death involves cathepsin D
immediately supplemented with tetracycline (1.5 gg/ml). All theenzymatic activities were assessed 48 h after transfections.
fl-Galactosidase staining and determination of SEAP activityTo detect lacZ expression, cells were fixed with 3% paraformaldehydefor 5 min, rinsed twice with PBS and stained for 3 h in X-Gal buffercontaining 77 mM Na2HPO4, 23 mM NaH2PO4, 1.3 mM MgCl,,1 mg/ml X-Gal, 3 mM K3Fe(CN)6, 3 mM K4Fe(CN)6. The reaction wasstopped with 70% ethanol. Photography was under phase microscopyusing Kodak Ektachrome 160T. For the SEAP activity assay the mediumof transfected cells was changed 5 h before assay. Aliquots of 100 ,ulmedium were removed from the transfected plates and heated at 650Cfor 5 min. The medium was then clarified by centrifugation at 14000 gfor 2 min. The medium aliqouts were adjusted to 1 X SEAP assay buffercontaining 2 M diethanolamine, pH 9.8, 1 M MgCI2 and 20 mML-homoarginine. Aliquots of 20 gl 120 mM p-nitrophenylphosphatedissolved in water were then added to each mixture. The reactionmixtures were then incubated for 30 min at 37°C. The hydrolysis ofp-nitrophenylphosphate was measured at 405 nm.
AcknowledgementsWe thank Dr S.Korenfeld for providing the full-length cDNA clone ofhuman cathepsin D and Dr P.Krammer for the anti-APO-1 monoclonalantibodies and for helpful discussions. We also thank Dr Groner foranti-SOD antibodies. This work was supported by The Israel ScienceFoundation administered by The Israel Academy of Sciences andHumanities. A.K. is an incumbent of the Helena Rubinstein Chair ofCancer Research.
ReferencesBoldin,M.P., Varfolomeev,E.E., Pancer,Z., Mett,L., Camonis,J.H. and
Wallach,D. (1995) A novel protein that interacts with the death domainof Fas/APO1 contains a sequence motif related to the death domain.J. Biol. Chem., 270, 7795-7798.
Chinnalyan,A.M., O'Rourke,K., Tewari,M. and Dixit,V.M. (1995) FADD,a novel death domain-containing protein, interacts with the deathdomain of Fas and initiates apoptosis. Cell, 81, 505-512.
Deiss,L.P. and Kimchi,A. (1991) A genetic tool used to identifythioredoxin as a mediator of a growth inhibitory signal. Science, 252,117-120.
Deiss,L.P., Feinstein,E., Berissi,H., Cohen,O. and Kimchi,A. (1995)Identification of a novel serine/threonine kinase and a novel 15-kDprotein as potential mediators of gamma interferon-induced cell death.Genes Dei'., 9, 15-30.
Diment,S., Martin,K.J. and Stahl,P.D. (1989) Cleavage of parathyroidhormone in macrophage endosomes illustrates a novel pathway forintracellular processing of proteins. J. Biol. Chem., 264, 13403-13416.
Dirks,W., Wirth,M. and Hauser,H. (1993) Dicistronic transcription unitsfor gene expression in mammalian cells. Gene, 128, 247-259.
Ellis,R.J., Yuan,J.Y. and Horvitz,H.R. (1991) Mechanisms and functionsof cell death. Annu. Rev. Cell. Biol., 7, 663-698.
Enari,M., Hug,H. and Nagata,S. (1995) Involvement of an ICE-likeprotease in Fas-mediated apoptosis. Nature, 375, 78-81.
Evan,G.I., Wyllie,A.H., Gilbert,C.S., Littlewood,T.D., Land,H.,Brooks,M., Waters,C.M., Penn,L.Z. and Hancock,D.C. (1992)Induction of apoptosis in fibroblasts by c-myc protein. Cell, 69,119-128.
Faucheu,C. et al. (1995) A novel human protease similar to theinterleukin-I beta converting enzyme induces apoptosis in transfectedcells. EMBO J., 14, 1914-1922.
Faust,P.L., Kornfeld,S. and Chirgwin,J.M. (1985) Cloning and sequenceanalysis of cDNA for human cathepsin D. Proc. Natl Acad. Sci. USA,82, 4910-4914.
Feinstein,E., Wallach,D., Boldin,M., Varfolomeev,E. and Kimchi,A.(1995) The death domain: a module shared by proteins with diversecellular functions. Trends Biochem. Sci., 20, 342-344.
Fernandes,A.T., Litwack,G. and Alnemri,E.S. (1994) CPP32, a novelhuman apoptotic protein with homology to Caenorhabditis eleganscell death protein Ced-3 and mammalian interleukin- 1 beta-convertingenzyme. J. Biol. Chem., 269, 30761-30764.
Fernandes,A.T., Litwack,J. and Alnemri,E.S. (1995) Mch2, a newmember of the apoptotic Ced-3/Ice cysteine protease gene family.Cancer Res., 55, 2737-2742.
Fujita,H., Tanaka,Y., Noguchi,Y., Kono,A., Himeno,M. and Kato,K.
(1991) Isolation and sequencing of a cDNA clone encoding rat liverlysosomal cathepsin D and the structure of three forms of matureenzymes. Biochem. Biophys. Res. Commun., 179, 190-196.
Gossen,M. and Bujard,H. (1992) Tight control of gene expression inmammalian cells by tetracycline-responsive promoters. Proc. NatlAcad. Sci. USA, 89, 5547-5551.
Grether,M.E., Abrams,J.M., Agapite,J., White,K. and Steller,H. (1995)The head involution defective gene of Drosophila melanogasterfunctions in programmed cell death. Genes Dev., 9, 1694-1708.
Heu,H., Xiong,J. and Goeddel,D.V. (1995) The TNF receptor 1-associatedprotein TRADD signals cell death and NF-KB activation. Cell, 81,495-504.
Heusel,J.W., Wesselschmidt,R.L., Shresta,S., Russell,J.H. and Ley,T.J.(1994) Cytotoxic lymphocytes require granzyme B for the rapidinduction of DNA fragmentation and apoptosis in allogeneic targetcells. Cell, 76, 977-987.
Itoh,N., Yonehara,S., Ishii,A., Yonehara,M., Mizushima,S.,Sameshima,M., Hase,A., Seto,Y. and Nagata,S. (1991) The polypeptideencoded by the cDNA for human cell surface antigen Fas can mediateapoptosis. Cell, 66, 233-243.
Johnson,G.V., Litersky,J.M. and Whitaker,J.N. (1991) Proteolysis ofmicrotubule-associated protein 2 and tubulin by cathepsin D.J. Neurochem., 57, 1577-1583.
Kenessey,A., Banay,S.M., DeGuzman,T. and Lajtha,A. (1989) Increasein cathepsin D activity in rat brain in aging. J. Neurosci. Res., 23,454-456.
Kimchi,A. (1992) Cytokine triggered molecular pathways that controlcell cycle arrest. J. Cell Biochem., 50, 1-9
Kissil,J.L., Deiss,L.P., Bayewitch,M., Raveh,T., Khaspekov,G. andKimchi,A. (1995) Isolation of DAP-3: a novel mediator of interferon-gamma-induced cell death. J. Biol. Chem., 270, 27932-27936.
Kumar,S., Kinoshita,M., Noda,M., Copeland,N.G. and Jenkins,N.A.(1994) Induction of apoptosis by the mouse Nedd2 gene, whichencodes a protein similar to the product of the Caenorhabditis eleganscell death gene ced-3 and the mammalian IL-1 beta-convertingenzyme. Genes Dev., 8, 1613-1626.
Laster,S.M., Wood,J.G. and Gooding,L.R. (1988) Tumor necrosis factorcan induce both apoptic and necrotic forms of cell lysis. J. Immunol.,141, 2629-2634.
Lin,J.K. and Chou,C.K. (1992) In vitro apoptosis in the hepatoma cellline induced by transforming growth factor-beta. Cancer Res., 52,385-388.
Los,M. et al. (1995) Requirement of an ICE/CED-3 protease for Fas/APO-1-mediated apoptosis. Nature, 375, 81-83.
Matus,A. and Green,G.D. (1987) Age-related increase in a cathepsin Dlike protease that degrades brain microtubule-associated proteins.Biochemistry, 26, 8083-8086.
Mignatti,P. and Rifkin,D.B. (1993) Biology and biochemistry ofproteinases in tumor invasion. Physiol. Rev., 73, 161-195.
Miura,M., Zhu,H., Rotello,R., Hartwieg,E.A. and Yuan,J. (1993)Induction of apoptosis in fibroblasts by IL-1 beta-converting enzyme,a mammalian homolog of the C. elegans cell death gene ced-3. Cell,75, 653-660.
Novelli,F., Di Pierro,F., Francia di Celle,P., Bertini,S., Affaticati,P.,Garotta,G. and Forni,G. (1994) Environmental signals influencingexpression of the IFN-gamma receptor on human T cells controlwhether IFN-gamma promotes proliferation or apoptosis. J. Immunol.,152, 496-504.
Raff,M.C. (1992) Social controls on cell survival and cell death. Nature,356, 397-400.
Saftig,P., Hetman,M., Schmahl,W., Weber,K., Heine,L. and Peters,C.(1995) Mice deficient for the lysosomal proteinase cathepsin D exhibitprogressive atrophy of the intestinal mucosa and profound destructionof lymphoid cells. EMBO J., 14, 3599-3608.
Schwartz,L.M. and Osborne,B.A. (1993) Programmed cell death,apoptosis and killer genes. Immunol. Today, 14, 582-590.
Shields,P.P., Gonzales,T.A., Charles,D., Gilligan,J.P. and Stern,W. (1991)Accumulation of pepstatin in cultured endothelial cells and its effecton endothelial processing. Biochem. Biophys. Res. Commun., 177,1006-1012.
Stanger,B.Z., Leder,P., Lee,T.H., Kim,E. and Seed,B. (1995) RIP: anovel protein containing a death domain that interacts with Fas/APO-1 (CD95) in yeast and causes cell death. Cell, 81, 513-523.
Steller,H. (1995) Mechanisms and genes of cellular suicide. Science,267, 1445-1449.
Suda,T. and Nagata,S. (1994) Purification and characterization of theFas-ligand that induces apoptosis. J. Exp. Med., 179, 873-879.
3869

L.P.Deiss et al.
Tewari.M. and Dixit,V.M. (1995) Fas- and tumor necrosis factor-inducedapoptosis is inhibited by the poxvirus crm11A gene product. J. Biol.Clheo7i., 270, 3255-3260.
Tewari,M.. Quan.L.T.. O'Rourke,K., Desnoyers,S., Zeng,Z.. Beidler,D.R.. Poirier,G.G.. Salvesen.G.S. and Dixit.V.M. (1995) Yama/CPP32,B. a mammalian homolog of CED-3. is a CrmA-inhibitableprotease that cleaves the death substrate poly(ADP-ribose) polymerase.Cell, 81, 1-20
Trauth,B.C., Klas,C., Peters,A.M., Matzku,S., Moller,P., Falk,W.,Debatin,K.M. and Krammer.P.H. (1989) Monoclonal antibody-mediated tumor regression by induction of apoptosis. Science, 245,301-305.
Wallach,D. (1984) Preparations of lymphotoxin induce resistance to theirown cytotoxic effect. J. I/o17oti1naol., 132, 2464-2469.
Wang.L., Miura.M.. Bergeron,L.. Zhu,H. and Yuan,J. (1994) ic/i-i, anIce/cel-3-related gene. encodes both positive and negative regulatorsof programmed cell death. Cell. 78, 739-750.
White,K., Grether.M.E., Abrams,J.M., Young,L., Farrell,K. and Steller,H.(1994) Genetic control of programmed cell death in Drosophillia [seecomments]. Science. 264, 677-683.
Wright,S.C., Kumar.P., Tam,A.W.. Shen.N., Varma,M. and Larrick.J.W.(1992) Apoptosis and DNA fragmentation precede TNF-inducedcytolysis in U937 cells. J. Cell Bioclhen.. 48, 344-355.
Wyllie,A.H. (1995) The genetic regulation of apoptosis. Curi: Opin.Geliet. Dec.. 5, 97-104.
Yarden,A. and Kimchi,A. (1986) Tumor necrosis factor reduces c-ivi-cexpression and cooperates with interferon-gamma in HeLa cells.Science. 234, 1419-1421.
Yonish-Rouach,E., Resnitzky,D., Lotem,J., Sachs,L., Kimchi,A. andOren,M. (1991) Wild-type p53 induces apoptosis of myeloid leukaemiccells that is inhibited by interleukin-6. Nature, 352, 345-347.
Yuan,J. and Horvitz,H.R. (1992) The C. elegcanis cell death gene ced-4encodes a novel protein and is expressed during the period of extensiveprogrammed cell death. Det elopinent, 116, 309-320.
Yuan,J., Shaham,S., Ledoux,S., Ellis,H.M. and Horvitz,H.R. (1993) TheC. eleg(ants cell death gene ced-3 encodes a protein similar tomammalian interleukin-l beta-converting enzyyme. Cell, 75. 641-752.
Receivedl oni Februar-Y 29, 1996, revisedl oai April 11, 1996
3870


















