
The Basal Position of Scaly Green Flagellates among the ... · among the Green Algae (Chlorophyta)...
14
Protist. Vol. 149,367-380, December 1998 © Gustav Fischer Verlag ORIGINAL PAPER Protist The Basal Position of Scaly Green Flagellates among the Green Algae (Chlorophyta) is Revealed by Analyses of Nuclear-Encoded SSU rRNA Sequences Takeshi Nakayama a , Birger Marin b , Harald D. Kranz c ,1, Barbara Surek b , Volker A. R. Huss c , Isao Inouye a , and Michael Melkonian b ,2 alnstitute of Biological Sciences, University of Tsukuba, Tsukuba, Ibaraki 305, Japan bBotanisches Institut, Lehrstuhll, Universitat zu Koln, Gyrhofstr. 15, D-50931 Koln, Germany clnstitut fOr Botanik und Pharmazeutische Biologie, Universitat Erlangen, Staudtstr. 5, D-91 058 Erlangen, Germany Submitted August 11, 1998; Accepted September 30, 1998 Monitoring Editor: Robert A. Andersen The prasinophytes comprise a morphologically heterogeneous assembly of mostly marine flagellates and coccoid taxa, which represent an important component of the nano- and picoplankton, and have previously figured prominently in discussions about the origin and phylogeny of the green plants. To evaluate their putative basal position in the Viridiplantae and to resolve the phylogenetic relation- ships among the prasinophyte taxa, we determined complete nuclear-encoded SSU rRNA sequences from 13 prasinophyte taxa representing the genera Cymbomonas, Halosphaera, Mamiella, Manto- niella, Micromonas, Pterosperma, Pycnococcus, and Pyramimonas. Phylogenetic analyses of SSU rRNA sequences using distance, parsimony and likelihood methods revealed four independent prasinophyte lineages (clades) which constitute the earliest divergences among the Chlorophyta. In order of their divergence these clades are represented by the genera Cymbomonas, Halosphaera, Pterosperma, Pyramimonas (clade I), Mamiella, Mantoniella, Micromonas (clade II), Pseudoscourfie/- dia (strain CCMP 717), Nephroselmis (clade III), and Tetraselmis, Scherffelia (clade IV). The coccoid Pycnococcus provasolii diverged after clade II, but before clade III. Since no other coccoid prasino- phyte taxa were analyzed in this study, the phylogenetic status of this taxon is presently unresolved. Our analyses provide further evidence for the basal phylogenetic position of the scaly green flagel- lates among the Chlorophyta and raise important questions concerning the class-level classification of the Chlorophyta. Introduction 1Current address: Abteilung Biochemie, Max-Planck-Institut fOr ZOchtungsforschung, Carl-v.-Linne-Weg 10, D- 50829 Ktiln, Germany 2Corresponding author; fax 49-2214705181 e-mail [email protected] Green plants, the Viridiplantae sensu Cavalier-Smith (1981; see also Sluiman 1985), represent a mono- phyletic lineage of eukaryotic organisms which comprises the green algae and the em bryophyte land plants. The unique type of plastid (Le. the Protist. Vol. 149,367-380, December 1998 © Gustav Fischer Verlag ORIGINAL PAPER Protist The Basal Position of Scaly Green Flagellates among the Green Algae (Chlorophyta) is Revealed by Analyses of Nuclear-Encoded SSU rRNA Sequences Takeshi Nakayama a , Birger Marin b , Harald D. Kranz c ,1, Barbara Surek b , Volker A. R. Huss c , Isao Inouye a , and Michael Melkonian b ,2 alnstitute of Biological Sciences, University of Tsukuba, Tsukuba, Ibaraki 305, Japan bBotanisches Institut, Lehrstuhll, Universitat zu Koln, Gyrhofstr. 15, D-50931 Koln, Germany clnstitut fOr Botanik und Pharmazeutische Biologie, Universitat Erlangen, Staudtstr. 5, D-91 058 Erlangen, Germany Submitted August 11, 1998; Accepted September 30, 1998 Monitoring Editor: Robert A. Andersen The prasinophytes comprise a morphologically heterogeneous assembly of mostly marine flagellates and coccoid taxa, which represent an important component of the nano- and picoplankton, and have previously figured prominently in discussions about the origin and phylogeny of the green plants. To evaluate their putative basal position in the Viridiplantae and to resolve the phylogenetic relation- ships among the prasinophyte taxa, we determined complete nuclear-encoded SSU rRNA sequences from 13 prasinophyte taxa representing the genera Cymbomonas, Halosphaera, Mamiella, Manto- niella, Micromonas, Pterosperma, Pycnococcus, and Pyramimonas. Phylogenetic analyses of SSU rRNA sequences using distance, parsimony and likelihood methods revealed four independent prasinophyte lineages (clades) which constitute the earliest divergences among the Chlorophyta. In order of their divergence these clades are represented by the genera Cymbomonas, Halosphaera, Pterosperma, Pyramimonas (clade I), Mamiella, Mantoniella, Micromonas (clade II), Pseudoscourfie/- dia (strain CCMP 717), Nephroselmis (clade III), and Tetraselmis, Scherffelia (clade IV). The coccoid Pycnococcus provasolii diverged after clade II, but before clade III. Since no other coccoid prasino- phyte taxa were analyzed in this study, the phylogenetic status of this taxon is presently unresolved. Our analyses provide further evidence for the basal phylogenetic position of the scaly green flagel- lates among the Chlorophyta and raise important questions concerning the class-level classification of the Chlorophyta. Introduction 1Current address: Abteilung Biochemie, Max-Planck-Institut fOr ZOchtungsforschung, Carl-v.-Linne-Weg 10, D- 50829 Ktiln, Germany 2Corresponding author; fax 49-2214705181 e-mail [email protected] Green plants, the Viridiplantae sensu Cavalier-Smith (1981; see also Sluiman 1985), represent a mono- phyletic lineage of eukaryotic organisms which comprises the green algae and the em bryophyte land plants. The unique type of plastid (Le. the
Transcript of The Basal Position of Scaly Green Flagellates among the ... · among the Green Algae (Chlorophyta)...

Protist. Vol. 149,367-380, December 1998 © Gustav Fischer Verlag
ORIGINAL PAPER
Protist
The Basal Position of Scaly Green Flagellatesamong the Green Algae (Chlorophyta) isRevealed by Analyses of Nuclear-Encoded SSUrRNA Sequences
Takeshi Nakayamaa, Birger Marinb, Harald D. Kranzc,1, Barbara Surekb, Volker A. R. Hussc,
Isao Inouyea , and Michael Melkonianb,2
alnstitute of Biological Sciences, University of Tsukuba, Tsukuba, Ibaraki 305, JapanbBotanisches Institut, Lehrstuhll, Universitat zu Koln, Gyrhofstr. 15, D-50931 Koln, Germanyclnstitut fOr Botanik und Pharmazeutische Biologie, Universitat Erlangen, Staudtstr. 5, D-91 058 Erlangen, Germany
Submitted August 11, 1998; Accepted September 30, 1998Monitoring Editor: Robert A. Andersen
The prasinophytes comprise a morphologically heterogeneous assembly of mostly marine flagellatesand coccoid taxa, which represent an important component of the nano- and picoplankton, and havepreviously figured prominently in discussions about the origin and phylogeny of the green plants. Toevaluate their putative basal position in the Viridiplantae and to resolve the phylogenetic relationships among the prasinophyte taxa, we determined complete nuclear-encoded SSU rRNA sequencesfrom 13 prasinophyte taxa representing the genera Cymbomonas, Halosphaera, Mamiella, Mantoniella, Micromonas, Pterosperma, Pycnococcus, and Pyramimonas. Phylogenetic analyses of SSUrRNA sequences using distance, parsimony and likelihood methods revealed four independentprasinophyte lineages (clades) which constitute the earliest divergences among the Chlorophyta. Inorder of their divergence these clades are represented by the genera Cymbomonas, Halosphaera,Pterosperma, Pyramimonas (clade I), Mamiella, Mantoniella, Micromonas (clade II), Pseudoscourfie/dia (strain CCMP 717), Nephroselmis (clade III), and Tetraselmis, Scherffelia (clade IV). The coccoidPycnococcus provasolii diverged after clade II, but before clade III. Since no other coccoid prasinophyte taxa were analyzed in this study, the phylogenetic status of this taxon is presently unresolved.Our analyses provide further evidence for the basal phylogenetic position of the scaly green flagellates among the Chlorophyta and raise important questions concerning the class-level classificationof the Chlorophyta.
Introduction
1Current address: Abteilung Biochemie, Max-Planck-Institut fOrZOchtungsforschung, Carl-v.-Linne-Weg 10, D - 50829 Ktiln,Germany2Corresponding author;fax 49-2214705181e-mail [email protected]
Green plants, the Viridiplantae sensu Cavalier-Smith(1981; see also Sluiman 1985), represent a monophyletic lineage of eukaryotic organisms whichcomprises the green algae and the embryophyteland plants. The unique type of plastid (Le. the
Protist. Vol. 149,367-380, December 1998 © Gustav Fischer Verlag
ORIGINAL PAPER
Protist
The Basal Position of Scaly Green Flagellatesamong the Green Algae (Chlorophyta) isRevealed by Analyses of Nuclear-Encoded SSUrRNA Sequences
Takeshi Nakayamaa, Birger Marinb, Harald D. Kranzc,1, Barbara Surekb, Volker A. R. Hussc,
Isao Inouyea , and Michael Melkonianb,2
alnstitute of Biological Sciences, University of Tsukuba, Tsukuba, Ibaraki 305, JapanbBotanisches Institut, Lehrstuhll, Universitat zu Koln, Gyrhofstr. 15, D-50931 Koln, Germanyclnstitut fOr Botanik und Pharmazeutische Biologie, Universitat Erlangen, Staudtstr. 5, D-91 058 Erlangen, Germany
Submitted August 11, 1998; Accepted September 30, 1998Monitoring Editor: Robert A. Andersen
The prasinophytes comprise a morphologically heterogeneous assembly of mostly marine flagellatesand coccoid taxa, which represent an important component of the nano- and picoplankton, and havepreviously figured prominently in discussions about the origin and phylogeny of the green plants. Toevaluate their putative basal position in the Viridiplantae and to resolve the phylogenetic relationships among the prasinophyte taxa, we determined complete nuclear-encoded SSU rRNA sequencesfrom 13 prasinophyte taxa representing the genera Cymbomonas, Halosphaera, Mamiella, Mantoniella, Micromonas, Pterosperma, Pycnococcus, and Pyramimonas. Phylogenetic analyses of SSUrRNA sequences using distance, parsimony and likelihood methods revealed four independentprasinophyte lineages (clades) which constitute the earliest divergences among the Chlorophyta. Inorder of their divergence these clades are represented by the genera Cymbomonas, Halosphaera,Pterosperma, Pyramimonas (clade I), Mamiella, Mantoniella, Micromonas (clade II), Pseudoscourfie/dia (strain CCMP 717), Nephroselmis (clade III), and Tetraselmis, Scherffelia (clade IV). The coccoidPycnococcus provasolii diverged after clade II, but before clade III. Since no other coccoid prasinophyte taxa were analyzed in this study, the phylogenetic status of this taxon is presently unresolved.Our analyses provide further evidence for the basal phylogenetic position of the scaly green flagellates among the Chlorophyta and raise important questions concerning the class-level classificationof the Chlorophyta.
Introduction
1Current address: Abteilung Biochemie, Max-Planck-Institut fOrZOchtungsforschung, Carl-v.-Linne-Weg 10, D - 50829 Ktiln,Germany2Corresponding author;fax 49-2214705181e-mail [email protected]
Green plants, the Viridiplantae sensu Cavalier-Smith(1981; see also Sluiman 1985), represent a monophyletic lineage of eukaryotic organisms whichcomprises the green algae and the embryophyteland plants. The unique type of plastid (Le. the

368 T. Nakayama et al.
chloroplast; for classification of plastid types seeMelkonian 1996) with chlorophylls a and b, two envelope membranes, stacked thylakoids and intraplastidial starch, and the equally unique type offlagellar transitional region (the 'stellate structure';Melkonian 1984) provide synapomorphic charactersfor the group. Ultrastructural and molecular analyses have resulted in the recognition of two major lineages within the Viridiplantae now generally recognized as the Streptophyta and the Chlorophyta (Bremer 1985; Sluiman 1985; recent reviews: Friedl1997; Huss and Kranz 1997; McCourt 1995; Melkonian and Surek 1995). Whereas the Streptophytacomprise an assemblage of green algal lineagespreviously classified as the Charophyceae sensuMattox and Stewart (1984) and additionally the embryophyte land plants (bryophytes, ferns and spermatophytes), the Chlorophyta contain all greenalgae except for the charophyte lineages. It is widelyheld that scaly green flagellates (their cell surface iscovered by non-mineralized organic scales, Mantonand Parke 1960; for a more recent review of theprasinophytes: Sym and Pienaar 1993) were presumably ancestral to both the Streptophyta and theChlorophyta. The main rationale for this assumptionhas been the fact that flagellate reproductive cells(zoospores, gametes) of some taxa in both phyla arecovered by a layer of square-shaped scales (40-50nm in diameter), which also occur as an underlayerin many prasinophytes but nowhere outside theViridiplantae. Previous phylogenetic analyses involving comparisons of complete nuclear-encodedSSU rRNA sequences of selected prasinophyte taxawith other green algae have supported this notion,as the scaly green flagellates have invariably occupied basal positions in the phylogenetic trees (Friedl1997; Huss and Kranz 1997; Melkonian and Surek1995; Melkonian et al. 1995; Steink6tter et al. 1994).However, until now only 7 out of the 17 recognizedgenera of prasinophytes have been incorporatedwithin a single molecular phylogenetic analysis(Melkonian et al. 1995) and thus the relationshipsamong the various genera and lineages of prasino-
phytes (for a review of different classificationschemes of prasinophytes see Sym and Pienaar1993) have remained largely unexplored. As to beexpected from a basal lineage, a previous analysisof SSU rRNA sequence comparisons involving fourgenera of prasinophytes (Steink6tter et al. 1994)demonstrated that the prasinophytes are not monophyletic but at most paraphyletic within the Chlorophyta, since two independent basal lineages wereidentified corresponding to the previously recognized orders Pseudoscourfieldiales and Chlorodendrales (Melkonian 1990a). Addition of three moreprasinophyte taxa (Le. Pterosperma, Mantoniella,and Mesostigma) to a phylogenetic analysis of 67taxa of Viridiplantae led to the tentative recognitionof two additional independent lineages of prasinophytes, one of which (Le. Mesostigma) occupied aposition within the Streptophyta (Melkonian et al.1995). It became apparent that in order to resolvephylogenetic relationships among prasinophytes, amore comprehensive analysis was required. Here,we present a detailed phylogenetic analysis of thephylogenetic position of prasinophytes among theChlorophyta based on sequence comparisons ofcomplete nuclear-encoded SSU rRNA from a totalof 12 genera (16 taxa) of prasinophytes. We identifyat least four independent basal lineages of prasinophytes, their order of divergence, and discuss putative trends in the early evolution of the Chlorophyta.
Results
To position the prasinophyte taxa within the SSUrRNA phylogeny of the Viridiplantae we first performed a global analysis of nuclear-encoded SSUrRNA sequences of 66 taxa of Viridiplantae (including both Streptophyta and Chlorophyta) using threeglaucocystophyte sequences as outgroups (Fig. 1).Results of distance and maximum parsimony analyses revealed that all prasinophyte taxa studied werepositioned with the Chlorophyta which formed amonophyletic lineage (supported by bootstrap val-
Figure 1. Phylogeny of the Viridiplantae based on nuclear-encoded SSU rRNA sequence comparisons inferredwith the neighbor joining and maximum parsimony method using a total of 1655 aligned positions. The phylogenetic tree shown is that of the neighbor joining (Kimura 1980; Saitou and Nei 1987) method (the distance that corresponds to 3% sequence divergence is indicated by the scale). The phylogeny is rooted with the glaucocystophytetaxa Cyanophora paradoxa, Glaucocystis nostochinearum, and Cyanoptyche gloeocystis. Bootstrap values of distance (neighbor joining; number of above the nodes; 100 replications) and weighted maximum parsimony (numberbelow the nodes; 100 replications) analyses using the identical dataset are indicated (only values >50% wererecorded). * the topology in the weighted parsimony analysis is different [the Mamiellales (clade II) diverged with a81 % bootstrap value directly after clade I; as in Fig. 2]. The prasinophyte taxa are printed in bold type. For furtherdetails see Results.
368 T. Nakayama et al.
chloroplast; for classification of plastid types seeMelkonian 1996) with chlorophylls a and b, two envelope membranes, stacked thylakoids and intraplastidial starch, and the equally unique type offlagellar transitional region (the 'stellate structure';Melkonian 1984) provide synapomorphic charactersfor the group. Ultrastructural and molecular analyses have resulted in the recognition of two major lineages within the Viridiplantae now generally recognized as the Streptophyta and the Chlorophyta (Bremer 1985; Sluiman 1985; recent reviews: Friedl1997; Huss and Kranz 1997; McCourt 1995; Melkonian and Surek 1995). Whereas the Streptophytacomprise an assemblage of green algal lineagespreviously classified as the Charophyceae sensuMattox and Stewart (1984) and additionally the embryophyte land plants (bryophytes, ferns and spermatophytes), the Chlorophyta contain all greenalgae except for the charophyte lineages. It is widelyheld that scaly green flagellates (their cell surface iscovered by non-mineralized organic scales, Mantonand Parke 1960; for a more recent review of theprasinophytes: Sym and Pienaar 1993) were presumably ancestral to both the Streptophyta and theChlorophyta. The main rationale for this assumptionhas been the fact that flagellate reproductive cells(zoospores, gametes) of some taxa in both phyla arecovered by a layer of square-shaped scales (40-50nm in diameter), which also occur as an underlayerin many prasinophytes but nowhere outside theViridiplantae. Previous phylogenetic analyses involving comparisons of complete nuclear-encodedSSU rRNA sequences of selected prasinophyte taxawith other green algae have supported this notion,as the scaly green flagellates have invariably occupied basal positions in the phylogenetic trees (Friedl1997; Huss and Kranz 1997; Melkonian and Surek1995; Melkonian et al. 1995; Steink6tter et al. 1994).However, until now only 7 out of the 17 recognizedgenera of prasinophytes have been incorporatedwithin a single molecular phylogenetic analysis(Melkonian et al. 1995) and thus the relationshipsamong the various genera and lineages of prasino-
phytes (for a review of different classificationschemes of prasinophytes see Sym and Pienaar1993) have remained largely unexplored. As to beexpected from a basal lineage, a previous analysisof SSU rRNA sequence comparisons involving fourgenera of prasinophytes (Steink6tter et al. 1994)demonstrated that the prasinophytes are not monophyletic but at most paraphyletic within the Chlorophyta, since two independent basal lineages wereidentified corresponding to the previously recognized orders Pseudoscourfieldiales and Chlorodendrales (Melkonian 1990a). Addition of three moreprasinophyte taxa (Le. Pterosperma, Mantoniella,and Mesostigma) to a phylogenetic analysis of 67taxa of Viridiplantae led to the tentative recognitionof two additional independent lineages of prasinophytes, one of which (Le. Mesostigma) occupied aposition within the Streptophyta (Melkonian et al.1995). It became apparent that in order to resolvephylogenetic relationships among prasinophytes, amore comprehensive analysis was required. Here,we present a detailed phylogenetic analysis of thephylogenetic position of prasinophytes among theChlorophyta based on sequence comparisons ofcomplete nuclear-encoded SSU rRNA from a totalof 12 genera (16 taxa) of prasinophytes. We identifyat least four independent basal lineages of prasinophytes, their order of divergence, and discuss putative trends in the early evolution of the Chlorophyta.
Results
To position the prasinophyte taxa within the SSUrRNA phylogeny of the Viridiplantae we first performed a global analysis of nuclear-encoded SSUrRNA sequences of 66 taxa of Viridiplantae (including both Streptophyta and Chlorophyta) using threeglaucocystophyte sequences as outgroups (Fig. 1).Results of distance and maximum parsimony analyses revealed that all prasinophyte taxa studied werepositioned with the Chlorophyta which formed amonophyletic lineage (supported by bootstrap val-
Figure 1. Phylogeny of the Viridiplantae based on nuclear-encoded SSU rRNA sequence comparisons inferredwith the neighbor joining and maximum parsimony method using a total of 1655 aligned positions. The phylogenetic tree shown is that of the neighbor joining (Kimura 1980; Saitou and Nei 1987) method (the distance that corresponds to 3% sequence divergence is indicated by the scale). The phylogeny is rooted with the glaucocystophytetaxa Cyanophora paradoxa, Glaucocystis nostochinearum, and Cyanoptyche gloeocystis. Bootstrap values of distance (neighbor joining; number of above the nodes; 100 replications) and weighted maximum parsimony (numberbelow the nodes; 100 replications) analyses using the identical dataset are indicated (only values >50% wererecorded). * the topology in the weighted parsimony analysis is different [the Mamiellales (clade II) diverged with a81 % bootstrap value directly after clade I; as in Fig. 2]. The prasinophyte taxa are printed in bold type. For furtherdetails see Results.

Prasinophyte Ancestry of the Chlorophyta 369
Streptophyta
Chlorophyta
100100
100100
8083
61
100100
7281
92
*
.--------Sphagnum palustreMnium hornum
Hylocomium splendensFunaria hygrometrica
L- Marchantia polymorphaFossombronia pusilla
71 Pellia epiphylla95 L- Anthoceros agrestis
L I*OO~_--lC===G~in~g~ko biloba100 Pinusluchuensis
r__-jI~OO~_--i__....:Lychnothamnus barbatusL-__~I~__--1 100 Chara foetida
100 L-----Nitellaflexilis...--- Mesotaenium caldariorum
Mougeotia scalarisr---------- Genicularia spirotaeniaStaurastrum sp. M752
Cosmarium botrytisColeochaete scutataColeochaete orbicularis
L- Klebsormidium flaccidumL- Chlorokybus atmophyticus
~--- Mamiella gilva100 100 Mantoniella antarctica100 100 95 Mantoniella squamata
100 Micromonas pusilla.----- Cymbomonas sp.L..-----Halosphaera sp•.---- Pyramimonas disomata
Pyramimonas propulsaIUlL--- Pyramimonas olivacea
L..----Pyramimonas parkeaeL- Pterosperma cristatum
100 Dunaliella salina100 L- Asteromonas gracilis
L- Chlamydopodium starriiChlamydomonas reinhardtii
Volvox carteriHydrodictyon reticulatum
Pediastrum duplexScenedesmus abundansScenedesmus obliquus
...---- Neochloris aquaticaL- Characium hindakii
L..- Ankistrodesmus stipitatus...------Prototheca wickerhamii
riiVi1,,----- Nanochlorum eucaryotumChlorella vulgaris
1.- Oltmannsiellopsis viridisTrebouxia impressa
Trebouxia asymmetricaMyrmecia biatorellaeMyrmecia israelensis
L- Trebouxia magnaMicrothamnion kuetzingianum
Fusochloris perforatum1.- Leptosira terrestris
Acrosiphonia sp.Ulothrix zonata
Pseudendoclonium basiliense91'L--- Gloeotilopsis planctonica
100 Scherffelia dubia100 Tetraselmis striata
89 .-------- PseudoscourfutlJia marina CCMP 71798 Nephroselmis olivacea
1.- Pycnococcus provasolii
9298
I_~l~OO~C===::;:: Glaucocystis nostochinearumr 100 Cyanophora paradoxaL- Cyanoptyche gloeocystis
3%
Prasinophyte Ancestry of the Chlorophyta 369
Streptophyta
Chlorophyta
100100
100100
8083
61
100100
7281
92
*
.--------Sphagnum palustreMnium hornum
Hylocomium splendensFunaria hygrometrica
L- Marchantia polymorphaFossombronia pusilla
71 Pellia epiphylla95 L- Anthoceros agrestis
L I*OO~_--lC===G~in~g~ko biloba100 Pinusluchuensis
r__-jI~OO~_--i__....:Lychnothamnus barbatusL-__~I~__--1 100 Chara foetida
100 L-----Nitellaflexilis...--- Mesotaenium caldariorum
Mougeotia scalarisr---------- Genicularia spirotaeniaStaurastrum sp. M752
Cosmarium botrytisColeochaete scutataColeochaete orbicularis
L- Klebsormidium flaccidumL- Chlorokybus atmophyticus
~--- Mamiella gilva100 100 Mantoniella antarctica100 100 95 Mantoniella squamata
100 Micromonas pusilla.----- Cymbomonas sp.L..-----Halosphaera sp•.---- Pyramimonas disomata
Pyramimonas propulsaIUlL--- Pyramimonas olivacea
L..----Pyramimonas parkeaeL- Pterosperma cristatum
100 Dunaliella salina100 L- Asteromonas gracilis
L- Chlamydopodium starriiChlamydomonas reinhardtii
Volvox carteriHydrodictyon reticulatum
Pediastrum duplexScenedesmus abundansScenedesmus obliquus
...---- Neochloris aquaticaL- Characium hindakii
L..- Ankistrodesmus stipitatus...------Prototheca wickerhamii
riiVi1,,----- Nanochlorum eucaryotumChlorella vulgaris
1.- Oltmannsiellopsis viridisTrebouxia impressa
Trebouxia asymmetricaMyrmecia biatorellaeMyrmecia israelensis
L- Trebouxia magnaMicrothamnion kuetzingianum
Fusochloris perforatum1.- Leptosira terrestris
Acrosiphonia sp.Ulothrix zonata
Pseudendoclonium basiliense91'L--- Gloeotilopsis planctonica
100 Scherffelia dubia100 Tetraselmis striata
89 .-------- PseudoscourfutlJia marina CCMP 71798 Nephroselmis olivacea
1.- Pycnococcus provasolii
9298
I_~l~OO~C===::;:: Glaucocystis nostochinearumr 100 Cyanophora paradoxaL- Cyanoptyche gloeocystis
3%

370 T. Nakayama et al.
ues >90%) to the exclusion of the Streptophyta.Within the Chlorophyta the prasinophytes were paraphyletic forming several independent, early diverginglineages (Fig. 1). Since in the global analysis somenodes and the branching order of several prasinophyte lineages were not well supported by bootstrap
values, we refined the analysis using more positions(1699 compared to 1655 in the global analysis) and arestricted dataset (21 taxa of Chlorophyta, withstreptophyte taxa as outgroups). Results of maximum likelihood, maximum parsimony and distanceanalyses of nuclear encoded SSU rRNA sequences
93 Halosphaera sp.
99/98 Cymbomonas tetramitiformis
89100/99
Pterosperma cristatum
Pyramimonas olivacea clade I
Pyramimonas disomata
Pyramimonas parkeae
Micromonas pusilla
Mantoniella squamata
~~/100 clade IIMantoniella antarctica
100100/100
53*/65 Pyramimonas propulsa
Mamiella sp.100
100/100
100100/100
100100/100
Acrosiphonia sp. ]Ulvophyceae
Gloeotilopsis planctonica
Chlamydopodium starrii ]Chlorophyceae
Dunaliella salina
'---="-- Leptosira terrestris ] Trebouxiophyceae
Tetraselmis striata ]clade IV
Scherffelia dubia
63 82 Pseudoscourfieldia marina CCMP 717 ]95/87 99181 clade III
'------ Nephroselmis olivacea
7690190
6890/89
'-------- Pycnococcus provasolii
Coleochaete scutata
Klebsormidium flaccidum
3%
Figure 2. Phylogeny of the prasinophytes based on nuclear-encoded SSU rRNA sequence comparisons inferredwith the maximum likelihood, maximum parsimony and the neighbor joining method using a total of 1699 alignedpositions. The phylogenetic tree shown is that of the neighbor joining (Kimura 1980; Saitou and Nei 1987) method(the distance that corresponds to 3% sequence divergence is indicated by the scale). The phylogeny is rooted withthe streptophyte genera Coleochaete scutata and Klebsormidium f1accidum. Bootstrap values of maximum likelihood (above the nodes; 100 replications), weighted maximum parsimony (left number below the nodes; 100 replications), and distance (neighbor joining; right number below the nodes; 500 replications) analyses using the identical dataset are indicated (only values >50% were recorded; * different topology in weighted parsimony analysis, fordetails see Results).
370 T. Nakayama et al.
ues >90%) to the exclusion of the Streptophyta.Within the Chlorophyta the prasinophytes were paraphyletic forming several independent, early diverginglineages (Fig. 1). Since in the global analysis somenodes and the branching order of several prasinophyte lineages were not well supported by bootstrap
values, we refined the analysis using more positions(1699 compared to 1655 in the global analysis) and arestricted dataset (21 taxa of Chlorophyta, withstreptophyte taxa as outgroups). Results of maximum likelihood, maximum parsimony and distanceanalyses of nuclear encoded SSU rRNA sequences
93 Halosphaera sp.
99/98 Cymbomonas tetramitiformis
89100/99
Pterosperma cristatum
Pyramimonas olivacea clade I
Pyramimonas disomata
Pyramimonas parkeae
Micromonas pusilla
Mantoniella squamata
~~/100 clade IIMantoniella antarctica
100100/100
53*/65 Pyramimonas propulsa
Mamiella sp.100
100/100
100100/100
100100/100
Acrosiphonia sp. ]Ulvophyceae
Gloeotilopsis planctonica
Chlamydopodium starrii ]Chlorophyceae
Dunaliella salina
'---="-- Leptosira terrestris ] Trebouxiophyceae
Tetraselmis striata ]clade IV
Scherffelia dubia
63 82 Pseudoscourfieldia marina CCMP 717 ]95/87 99181 clade III
'------ Nephroselmis olivacea
7690190
6890/89
'-------- Pycnococcus provasolii
Coleochaete scutata
Klebsormidium flaccidum
3%
Figure 2. Phylogeny of the prasinophytes based on nuclear-encoded SSU rRNA sequence comparisons inferredwith the maximum likelihood, maximum parsimony and the neighbor joining method using a total of 1699 alignedpositions. The phylogenetic tree shown is that of the neighbor joining (Kimura 1980; Saitou and Nei 1987) method(the distance that corresponds to 3% sequence divergence is indicated by the scale). The phylogeny is rooted withthe streptophyte genera Coleochaete scutata and Klebsormidium f1accidum. Bootstrap values of maximum likelihood (above the nodes; 100 replications), weighted maximum parsimony (left number below the nodes; 100 replications), and distance (neighbor joining; right number below the nodes; 500 replications) analyses using the identical dataset are indicated (only values >50% were recorded; * different topology in weighted parsimony analysis, fordetails see Results).

from 16 prasinophyte and 5 other chlorophyte taxa(from the three classes Ulvophyceae, Trebouxiophyceae and Chlorophyceae) are summarized inFigure 2. The trees were rooted with the streptophytetaxa Co/eochaete scutata and Klebsormidium f1accidum as outgroups. We used C. scutata and K. f1accidum as outgroups because of their relatively shortbranch lengths. However, when other streptophytetaxa were used as outgroups (e.g. Cosmarium botrytis/Mesotaenium caldariorum or Nitella f1exilis/Charafoetida) the same tree topology as shown in Figure 2was obtained (results not shown).
Within the Chlorophyta the prasinophytes studiedformed five independent, early diverging lineages(Fig. 2). The earliest divergence comprising thegenera Halosphaera, Cymbomonas, Pterospermaand Pyramimonas (a total of 7 taxa) was supportedby high bootstrap values in all types of analyses andthus represents a monophyletic lineage (clade I).Within this clade two lineages of taxa could be distinguished, one comprising the genera Halosphaera andCymbomonas, the second the genera Pterospermaand Pyramimonas. Whereas the first lineage was invariably supported by high bootstrap values and ispresumably monophyletic, the second lineage wasonly weakly supported in neighbor-joining and maximum likelihood analyses. In unweighted maximumparsimony analyses five equally parsimonious treesof 1072 steps (consistency index [CI] = 0.57, retention index [RI] =0.64) were obtained (not shown), thesingle most parsimonious tree which resulted fromthe weighted analysis (CI = 0.80, RI = 0.83) placed theCymbomonas/Halosphaera lineage (supported by abootstrap value of 100%) within the Pyramimonas/Pterosperma lineage splitting the genus Pyramimonas (not shown). The phylogenetic relationshipsamong Pterosperma (the two different isolates of P.cristatum analyzed had identical SSU rRNA sequences) and the four species of Pyramimonas couldnot be resolved using SSU rRNA (Fig. 2).
Clade I is a sister group to all remaining taxa (including members of the Ulvophyceae, Trebouxiophyceae and Chlorophyceae; Fig. 2). The branchuniting the latter is supported by relatively highbootstrap values (>75%) in all methods of analysis.The next diverging lineage, represented by thegenera Mamiella, Mantoniella and Micromonas(clade II), is supported by bootstrap values of 100%in all methods of analysis and a long commonbranch length (Fig. 2). In clade II Mamiella divergesfirst and is well separated from a monophyleticlineage (100% bootstrap values in all analyses)comprising the two remaining genera Mantoniellaand Micromonas. Micromonas is resolved as a sisterto the two species of Mantoniella (Fig. 2).
Prasinophyte Ancestry of the Chlorophyta 371
A third lineage of prasinophytes is represented bythe single coccoid member of the group studied,namely Pycnococcus provasolii [in addition to theauthentic strain CCMP 1203 we also analyzed another coccoid prasinophyte (CCMP 1198 listed as'unid. coccoid' prasinophyte), the 18S rRNA sequence of which turned out to be identical to CCMP1203]. It diverges after clade II and is united with theremaining chlorophyte taxa by a branch which issupported by relatively high bootstrap values inmaximum parsimony and neighbor joining analyses(95% and 87% respectively), but a considerablylower value in maximum likelihood analyses (63%;Fig. 2). Since the P. provasolii sequence is characterized by a relatively long branch and the lineage iscurrently only represented by a single taxon we prefer to defer recognition of a separate clade for thislineage until additional SSU rRNA sequences havebeen determined from coccoid prasinophytes.
The two later diverging lineages of prasinophytes(clade III: Pseudoscourfieldiales; clade IV: Chlorodendrales) and their phylogenetic relationships tothe 'advanced' chlorophyte classes Ulvophyceae,Trebouxiophyceae (the zoosporine members of thisclass were formerly classified as the Microthamniales sensu Melkonian; Melkonian 1990b) andChlorophyceae have been previously described andextensively discussed by Steink6tter et al. (1994)and need not be dealt with here except to note thatmonophyly of clades III and IV respectively wasagain demonstrated by high (> 80%; clade III) orvery high (100%; clade IV) bootstrap values in allmethods of analysis. The clade uniting the Chlorodendrales (genera Tetraselmis and Scherffelia) withthe later diverging 'advanced' Chlorophyta [hererepresented by Acrosiphonia sp. and G/oeotilopsisplanctonica (both Ulvophyceae), Chlamydopodiumstarrii and Dunaliella salina (both Chlorophyceae),and Leptosira terrestris (Trebouxiophyceae)] waswell supported in all methods of analysis (100%bootstrap values), whereas the clade uniting thePseudoscourfieldiales with the later diverging lineages was only well supported in maximum parsimony and neighbor-joining analyses (bootstrapvalues of 90% and 89% respectively), whereas inmaximum likelihood analyses the bootstrap valuewas again lower (68%).
Comparisons of bootstrap values provide somemeasure of confidence in phylogenetic tree reconstructions. All four clades of prasinophytes werewell supported by bootstrap values in all methods ofanalysis; their order of divergence was also wellsupported in maximum parsimony and distanceanalyses, but less well supported in maximum likelihood analyses. A complementary method for as-
from 16 prasinophyte and 5 other chlorophyte taxa(from the three classes Ulvophyceae, Trebouxiophyceae and Chlorophyceae) are summarized inFigure 2. The trees were rooted with the streptophytetaxa Co/eochaete scutata and Klebsormidium f1accidum as outgroups. We used C. scutata and K. f1accidum as outgroups because of their relatively shortbranch lengths. However, when other streptophytetaxa were used as outgroups (e.g. Cosmarium botrytis/Mesotaenium caldariorum or Nitella f1exilis/Charafoetida) the same tree topology as shown in Figure 2was obtained (results not shown).
Within the Chlorophyta the prasinophytes studiedformed five independent, early diverging lineages(Fig. 2). The earliest divergence comprising thegenera Halosphaera, Cymbomonas, Pterospermaand Pyramimonas (a total of 7 taxa) was supportedby high bootstrap values in all types of analyses andthus represents a monophyletic lineage (clade I).Within this clade two lineages of taxa could be distinguished, one comprising the genera Halosphaera andCymbomonas, the second the genera Pterospermaand Pyramimonas. Whereas the first lineage was invariably supported by high bootstrap values and ispresumably monophyletic, the second lineage wasonly weakly supported in neighbor-joining and maximum likelihood analyses. In unweighted maximumparsimony analyses five equally parsimonious treesof 1072 steps (consistency index [CI] = 0.57, retention index [RI] =0.64) were obtained (not shown), thesingle most parsimonious tree which resulted fromthe weighted analysis (CI = 0.80, RI = 0.83) placed theCymbomonas/Halosphaera lineage (supported by abootstrap value of 100%) within the Pyramimonas/Pterosperma lineage splitting the genus Pyramimonas (not shown). The phylogenetic relationshipsamong Pterosperma (the two different isolates of P.cristatum analyzed had identical SSU rRNA sequences) and the four species of Pyramimonas couldnot be resolved using SSU rRNA (Fig. 2).
Clade I is a sister group to all remaining taxa (including members of the Ulvophyceae, Trebouxiophyceae and Chlorophyceae; Fig. 2). The branchuniting the latter is supported by relatively highbootstrap values (>75%) in all methods of analysis.The next diverging lineage, represented by thegenera Mamiella, Mantoniella and Micromonas(clade II), is supported by bootstrap values of 100%in all methods of analysis and a long commonbranch length (Fig. 2). In clade II Mamiella divergesfirst and is well separated from a monophyleticlineage (100% bootstrap values in all analyses)comprising the two remaining genera Mantoniellaand Micromonas. Micromonas is resolved as a sisterto the two species of Mantoniella (Fig. 2).
Prasinophyte Ancestry of the Chlorophyta 371
A third lineage of prasinophytes is represented bythe single coccoid member of the group studied,namely Pycnococcus provasolii [in addition to theauthentic strain CCMP 1203 we also analyzed another coccoid prasinophyte (CCMP 1198 listed as'unid. coccoid' prasinophyte), the 18S rRNA sequence of which turned out to be identical to CCMP1203]. It diverges after clade II and is united with theremaining chlorophyte taxa by a branch which issupported by relatively high bootstrap values inmaximum parsimony and neighbor joining analyses(95% and 87% respectively), but a considerablylower value in maximum likelihood analyses (63%;Fig. 2). Since the P. provasolii sequence is characterized by a relatively long branch and the lineage iscurrently only represented by a single taxon we prefer to defer recognition of a separate clade for thislineage until additional SSU rRNA sequences havebeen determined from coccoid prasinophytes.
The two later diverging lineages of prasinophytes(clade III: Pseudoscourfieldiales; clade IV: Chlorodendrales) and their phylogenetic relationships tothe 'advanced' chlorophyte classes Ulvophyceae,Trebouxiophyceae (the zoosporine members of thisclass were formerly classified as the Microthamniales sensu Melkonian; Melkonian 1990b) andChlorophyceae have been previously described andextensively discussed by Steink6tter et al. (1994)and need not be dealt with here except to note thatmonophyly of clades III and IV respectively wasagain demonstrated by high (> 80%; clade III) orvery high (100%; clade IV) bootstrap values in allmethods of analysis. The clade uniting the Chlorodendrales (genera Tetraselmis and Scherffelia) withthe later diverging 'advanced' Chlorophyta [hererepresented by Acrosiphonia sp. and G/oeotilopsisplanctonica (both Ulvophyceae), Chlamydopodiumstarrii and Dunaliella salina (both Chlorophyceae),and Leptosira terrestris (Trebouxiophyceae)] waswell supported in all methods of analysis (100%bootstrap values), whereas the clade uniting thePseudoscourfieldiales with the later diverging lineages was only well supported in maximum parsimony and neighbor-joining analyses (bootstrapvalues of 90% and 89% respectively), whereas inmaximum likelihood analyses the bootstrap valuewas again lower (68%).
Comparisons of bootstrap values provide somemeasure of confidence in phylogenetic tree reconstructions. All four clades of prasinophytes werewell supported by bootstrap values in all methods ofanalysis; their order of divergence was also wellsupported in maximum parsimony and distanceanalyses, but less well supported in maximum likelihood analyses. A complementary method for as-
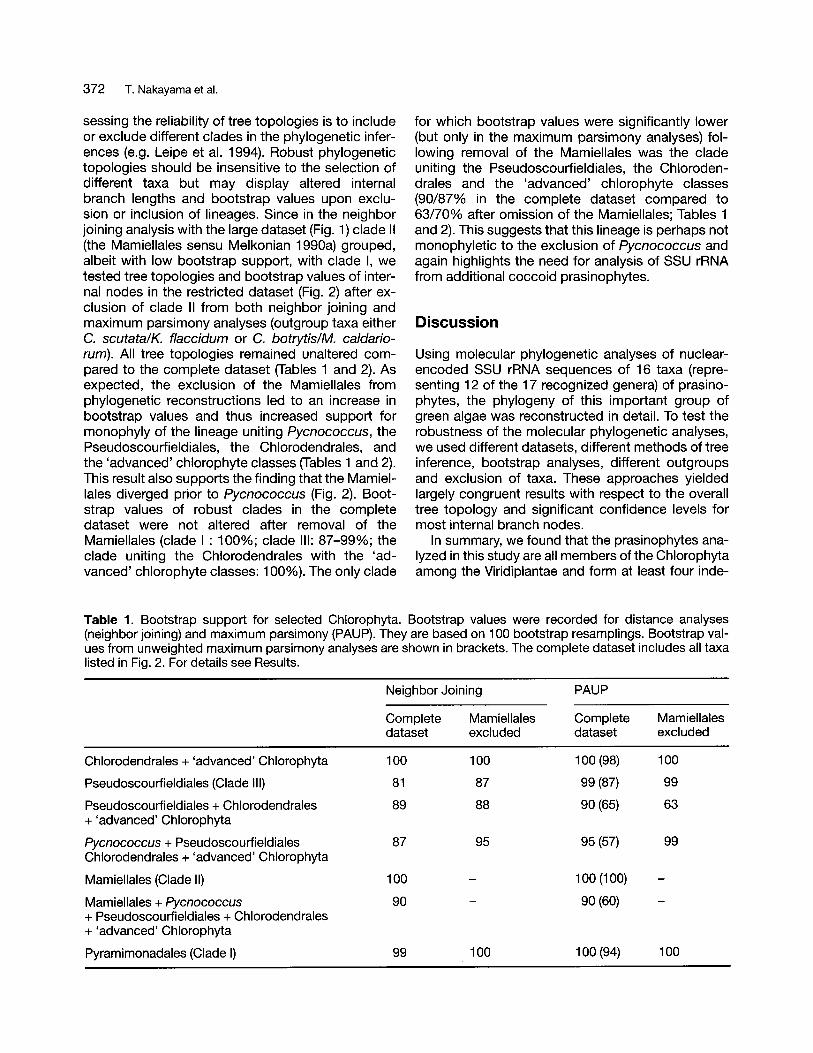
372 T. Nakayama et al.
sessing the reliability of tree topologies is to includeor exclude different clades in the phylogenetic inferences (e.g. Leipe et al. 1994). Robust phylogenetictopologies should be insensitive to the selection ofdifferent taxa but may display altered internalbranch lengths and bootstrap values upon exclusion or inclusion of lineages. Since in the neighborjoining analysis with the large dataset (Fig. 1) clade II(the Mamiellales sensu Melkonian 1990a) grouped,albeit with low bootstrap support, with clade I, wetested tree topologies and bootstrap values of internal nodes in the restricted dataset (Fig. 2) after exclusion of clade II from both neighbor joining andmaximum parsimony analyses (outgroup taxa eitherC. scutata/K. f1accidum or C. botrytis/M. caldariorum). All tree topologies remained unaltered compared to the complete dataset (Tables 1 and 2). Asexpected, the exclusion of the Mamiellales fromphylogenetic reconstructions led to an increase inbootstrap values and thus increased support formonophyly of the lineage uniting Pycnococcus, thePseudoscourfieldiales, the Chlorodendrales, andthe 'advanced' chlorophyte classes (Tables 1 and 2).This result also supports the finding that the Mamiellales diverged prior to Pycnococcus (Fig. 2). Bootstrap values of robust clades in the completedataset were not altered after removal of theMamiellales (clade I : 100%; clade III: 87-99%; theclade uniting the Chlorodendrales with the 'advanced' chlorophyte classes: 100%). The only clade
for which bootstrap values were significantly lower(but only in the maximum parsimony analyses) following removal of the Mamiellales was the cladeuniting the Pseudoscourfieldiales, the Chlorodendrales and the 'advanced' chlorophyte classes(90/87% in the complete dataset compared to63/70% after omission of the Mamiellales; Tables 1and 2). This suggests that this lineage is perhaps notmonophyletic to the exclusion of Pycnococcus andagain highlights the need for analysis of SSU rRNAfrom additional coccoid prasinophytes.
Discussion
Using molecular phylogenetic analyses of nuclearencoded SSU rRNA sequences of 16 taxa (representing 12 of the 17 recognized genera) of prasinophytes, the phylogeny of this important group ofgreen algae was reconstructed in detail. To test therobustness of the molecular phylogenetic analyses,we used different datasets, different methods of treeinference, bootstrap analyses, different outgroupsand exclusion of taxa. These approaches yieldedlargely congruent results with respect to the overalltree topology and significant confidence levels formost internal branch nodes.
In summary, we found that the prasinophytes analyzed in this study are all members of the Chlorophytaamong the Viridiplantae and form at least four inde-
Table 1. Bootstrap support for selected Chlorophyta. Bootstrap values were recorded for distance analyses(neighbor joining) and maximum parsimony (PAUP). They are based on 100 bootstrap resamplings. Bootstrap values from unweighted maximum parsimony analyses are shown in brackets. The complete dataset includes all taxalisted in Fig. 2. For details see Results.
Neighbor Joining PAUP
Complete Mamiellales Complete Mamiellalesdataset excluded dataset excluded
Chlorodendrales + 'advanced' Chlorophyta 100 100 100 (98) 100
Pseudoscourfieldiales (Clade III) 81 87 99 (87) 99
Pseudoscourfieldiales + Chlorodendrales 89 88 90 (65) 63+ 'advanced' Chlorophyta
Pycnococcus + Pseudoscourfieldiales 87 95 95 (57) 99Chlorodendrales + 'advanced' Chlorophyta
Mamiellales (Clade II) 100 100 (100)
Mamiellales + Pycnococcus 90 90 (60)+ Pseudoscourfieldiales + Chlorodendrales+ 'advanced' Chlorophyta
Pyramimonadales (Clade I) 99 100 100 (94) 100
372 T. Nakayama et al.
sessing the reliability of tree topologies is to includeor exclude different clades in the phylogenetic inferences (e.g. Leipe et al. 1994). Robust phylogenetictopologies should be insensitive to the selection ofdifferent taxa but may display altered internalbranch lengths and bootstrap values upon exclusion or inclusion of lineages. Since in the neighborjoining analysis with the large dataset (Fig. 1) clade II(the Mamiellales sensu Melkonian 1990a) grouped,albeit with low bootstrap support, with clade I, wetested tree topologies and bootstrap values of internal nodes in the restricted dataset (Fig. 2) after exclusion of clade II from both neighbor joining andmaximum parsimony analyses (outgroup taxa eitherC. scutata/K. f1accidum or C. botrytis/M. caldariorum). All tree topologies remained unaltered compared to the complete dataset (Tables 1 and 2). Asexpected, the exclusion of the Mamiellales fromphylogenetic reconstructions led to an increase inbootstrap values and thus increased support formonophyly of the lineage uniting Pycnococcus, thePseudoscourfieldiales, the Chlorodendrales, andthe 'advanced' chlorophyte classes (Tables 1 and 2).This result also supports the finding that the Mamiellales diverged prior to Pycnococcus (Fig. 2). Bootstrap values of robust clades in the completedataset were not altered after removal of theMamiellales (clade I : 100%; clade III: 87-99%; theclade uniting the Chlorodendrales with the 'advanced' chlorophyte classes: 100%). The only clade
for which bootstrap values were significantly lower(but only in the maximum parsimony analyses) following removal of the Mamiellales was the cladeuniting the Pseudoscourfieldiales, the Chlorodendrales and the 'advanced' chlorophyte classes(90/87% in the complete dataset compared to63/70% after omission of the Mamiellales; Tables 1and 2). This suggests that this lineage is perhaps notmonophyletic to the exclusion of Pycnococcus andagain highlights the need for analysis of SSU rRNAfrom additional coccoid prasinophytes.
Discussion
Using molecular phylogenetic analyses of nuclearencoded SSU rRNA sequences of 16 taxa (representing 12 of the 17 recognized genera) of prasinophytes, the phylogeny of this important group ofgreen algae was reconstructed in detail. To test therobustness of the molecular phylogenetic analyses,we used different datasets, different methods of treeinference, bootstrap analyses, different outgroupsand exclusion of taxa. These approaches yieldedlargely congruent results with respect to the overalltree topology and significant confidence levels formost internal branch nodes.
In summary, we found that the prasinophytes analyzed in this study are all members of the Chlorophytaamong the Viridiplantae and form at least four inde-
Table 1. Bootstrap support for selected Chlorophyta. Bootstrap values were recorded for distance analyses(neighbor joining) and maximum parsimony (PAUP). They are based on 100 bootstrap resamplings. Bootstrap values from unweighted maximum parsimony analyses are shown in brackets. The complete dataset includes all taxalisted in Fig. 2. For details see Results.
Neighbor Joining PAUP
Complete Mamiellales Complete Mamiellalesdataset excluded dataset excluded
Chlorodendrales + 'advanced' Chlorophyta 100 100 100 (98) 100
Pseudoscourfieldiales (Clade III) 81 87 99 (87) 99
Pseudoscourfieldiales + Chlorodendrales 89 88 90 (65) 63+ 'advanced' Chlorophyta
Pycnococcus + Pseudoscourfieldiales 87 95 95 (57) 99Chlorodendrales + 'advanced' Chlorophyta
Mamiellales (Clade II) 100 100 (100)
Mamiellales + Pycnococcus 90 90 (60)+ Pseudoscourfieldiales + Chlorodendrales+ 'advanced' Chlorophyta
Pyramimonadales (Clade I) 99 100 100 (94) 100

pendent clades representing early divergences in thephylogeny of the Chlorophyta. The analyses withsome confidence also resolved the phylogenetic relationships between these four clades of prasinophytes, Le. their sequence of divergence in the tree.Our results thus support an earlier study based on amuch more limited dataset (Steinkotter et al. 1994)which concluded that the Prasinophyceae sensuMoestrup and Throndsen (1988) are not monophyletic (at most paraphyletic) and represent independent, early diverging lineages within the Chlorophyta. The study by Steinkotter et al. (1994) includedtaxa from only two of the prasinophyte orders,namely the Pseudoscourfieldiales and the Chlorodendrales sensu Melkonian (1990a). An extension ofthis study to members of the orders Mamiellales andPyramimonadales (sensu Melkonian 1990a) whichhave previously figured prominently in discussionsabout the phylogenetic origin of the Viridiplantae (e.g.Friedl 1997; Kantz et al. 1990; Manton 1965; Mattoxand Stewart 1984; Melkonian 1982, 1984, 1990a;Melkonian and Surek 1995; Moestrup 1982, 1991;Moestrup and Throndsen 1988; Norris 1980; O'Kelly1992; O'Kelly and Floyd 1984; Sym and Pienaar1993) seemed highly appropriate.
Paraphyly of the prasinophytes
The 16 taxa analyzed formed a paraphyletic assemblage. This is to be expected from a basal group
Prasinophyte Ancestry of the Chlorophyta 373
of organisms (Sluiman 1985). It is now clear thatmost of the diagnostic characters which have beenused to define the prasinophytes as a single taxonomic entity (Le. cell surface scales, flagellar pit,parabasal Golgi bodies, extrusomes, long parallelbasal bodies, persistent interzonal spindle) are plesiomorphic and also occur in other groups of greenalgae (recent review: Sym and Pienaar 1993). Nounique character exists that unites all prasinophytetaxa to the exclusion of other Viridiplantae or members of other algal phyla (the only character thatcomes close to meeting this criterion are the flagellar T-hairs, which among the flagellate prasinophytetaxa are absent only from the genera Mesostigma,Micromonas and Pterosperma; Marin and Melkonian 1994; Moestrup and Throndsen 1988). Finally,taxa have been included in the Prasinophyceae(sensu Moestrup and Throndsen 1988) which clearlyshare apomorphic character states with 'advanced'chlorophyte classes, Le. a basal apparatus-associated 'phycoplast' system of microtubules during cytokinesis (see discussions in Mattox and Stewart1984 and Steinkotter et al. 1994). We now have evidence that the prasinophytes as presently conceived are even polyphyletic rather than paraphyletic since one prasinophyte genus (Mesostigma) is apparently a member of the Streptophyta(Melkonian et al. 1995; Marin and Melkonian, unpublished observations). There is thus no rationalreason to retain either the Prasinophyceae sensu
Table 2. Bootstrap support for selected Chlorophyta. Bootstrap values were recorded for distance analyses(neighbor joining) and maximum parsimony (PAUP). They are based on 100 bootstrap resamplings. Bootstrap values from unweighted maximum parsimony analyses are shown in brackets. As outgroup taxa for these analysesthe zygnematalean algae Cosmarium botrytis and Mesotaenium caldariorum were chosen. Ingroup taxa are thoseof Fig. 2. For details see Results.
Neighbor Joining PAUP
Complete Mamiellales Complete Mamiellalesdataset excluded dataset excluded
Chlorodendrales + 'advanced' Chlorophyta 100 100 100 (97) 100
Pseudoscourfieldiales (Clade III) 90 88 99 (90) 99
Pseudoscourfieldiales + Chlorodendrales 89 87 87 (61) 70+ 'advanced' Chlorophyta
Pycnococcus + Pseudoscourfieldiales 73 88 91 (52) 99Chlorodendrales + 'advanced' Chlorophyta
Mamiellales (Clade II) 100 100 (100)
Mamiellales + Pycnococcus 47 76 (44)+ Pseudoscourfieldiales + Chlorodendrales+ 'advanced' Chlorophyta
Pyramimonadales (Clade I) 98 100 99 (89) 100
pendent clades representing early divergences in thephylogeny of the Chlorophyta. The analyses withsome confidence also resolved the phylogenetic relationships between these four clades of prasinophytes, Le. their sequence of divergence in the tree.Our results thus support an earlier study based on amuch more limited dataset (Steinkotter et al. 1994)which concluded that the Prasinophyceae sensuMoestrup and Throndsen (1988) are not monophyletic (at most paraphyletic) and represent independent, early diverging lineages within the Chlorophyta. The study by Steinkotter et al. (1994) includedtaxa from only two of the prasinophyte orders,namely the Pseudoscourfieldiales and the Chlorodendrales sensu Melkonian (1990a). An extension ofthis study to members of the orders Mamiellales andPyramimonadales (sensu Melkonian 1990a) whichhave previously figured prominently in discussionsabout the phylogenetic origin of the Viridiplantae (e.g.Friedl 1997; Kantz et al. 1990; Manton 1965; Mattoxand Stewart 1984; Melkonian 1982, 1984, 1990a;Melkonian and Surek 1995; Moestrup 1982, 1991;Moestrup and Throndsen 1988; Norris 1980; O'Kelly1992; O'Kelly and Floyd 1984; Sym and Pienaar1993) seemed highly appropriate.
Paraphyly of the prasinophytes
The 16 taxa analyzed formed a paraphyletic assemblage. This is to be expected from a basal group
Prasinophyte Ancestry of the Chlorophyta 373
of organisms (Sluiman 1985). It is now clear thatmost of the diagnostic characters which have beenused to define the prasinophytes as a single taxonomic entity (Le. cell surface scales, flagellar pit,parabasal Golgi bodies, extrusomes, long parallelbasal bodies, persistent interzonal spindle) are plesiomorphic and also occur in other groups of greenalgae (recent review: Sym and Pienaar 1993). Nounique character exists that unites all prasinophytetaxa to the exclusion of other Viridiplantae or members of other algal phyla (the only character thatcomes close to meeting this criterion are the flagellar T-hairs, which among the flagellate prasinophytetaxa are absent only from the genera Mesostigma,Micromonas and Pterosperma; Marin and Melkonian 1994; Moestrup and Throndsen 1988). Finally,taxa have been included in the Prasinophyceae(sensu Moestrup and Throndsen 1988) which clearlyshare apomorphic character states with 'advanced'chlorophyte classes, Le. a basal apparatus-associated 'phycoplast' system of microtubules during cytokinesis (see discussions in Mattox and Stewart1984 and Steinkotter et al. 1994). We now have evidence that the prasinophytes as presently conceived are even polyphyletic rather than paraphyletic since one prasinophyte genus (Mesostigma) is apparently a member of the Streptophyta(Melkonian et al. 1995; Marin and Melkonian, unpublished observations). There is thus no rationalreason to retain either the Prasinophyceae sensu
Table 2. Bootstrap support for selected Chlorophyta. Bootstrap values were recorded for distance analyses(neighbor joining) and maximum parsimony (PAUP). They are based on 100 bootstrap resamplings. Bootstrap values from unweighted maximum parsimony analyses are shown in brackets. As outgroup taxa for these analysesthe zygnematalean algae Cosmarium botrytis and Mesotaenium caldariorum were chosen. Ingroup taxa are thoseof Fig. 2. For details see Results.
Neighbor Joining PAUP
Complete Mamiellales Complete Mamiellalesdataset excluded dataset excluded
Chlorodendrales + 'advanced' Chlorophyta 100 100 100 (97) 100
Pseudoscourfieldiales (Clade III) 90 88 99 (90) 99
Pseudoscourfieldiales + Chlorodendrales 89 87 87 (61) 70+ 'advanced' Chlorophyta
Pycnococcus + Pseudoscourfieldiales 73 88 91 (52) 99Chlorodendrales + 'advanced' Chlorophyta
Mamiellales (Clade II) 100 100 (100)
Mamiellales + Pycnococcus 47 76 (44)+ Pseudoscourfieldiales + Chlorodendrales+ 'advanced' Chlorophyta
Pyramimonadales (Clade I) 98 100 99 (89) 100

374 T. Nakayama et al.
Moestrup and Throndsen 1988 (emended byMoestrup 1991) or for that matter the Micromonadophyceae sensu Mattox and Stewart 1984 (the Micromonadophyceae included flagellates with a persistent interzonal spindle; this assemblage is conceptually different from the Prasinophyceae because itexcludes the thecate prasinophytes but includesgenera lacking scales such as Pedinomonas or having non-prasinophyte-type scales such as Monomastix).
Four independent lineages of prasinophytes
All methods of phylogenetic inference used in thisstudy resolved the prasinophyte taxa into at leastfour independent clades (for a discussion of the position of Pycnococcus provasolii see below) whichare designated here as clades I-IV.
Clade I. This well-supported clade representingthe genera Halosphaera, Cymbomonas, Pterosperma and Pyramimonas emerged from all phylogenetic analyses as the earliest divergence within theChlorophyta. Based on a number of synapomorphiccharacters these taxa have always been regardedas relatively closely related. They share a helicalstructure (transitional helix or coiled fiber) in theflagellar transitional region (this structure is foundnowhere else in the Viridiplantae), nine longitudinalrows of Iimulus-shaped flagellar scales, a striatedfiber (termed duct fiber) associated with the 1dmicrotubular flagellar root, a characteristic system IIfiber/nucleus/microbody/chloroplast association,and a unique type of flagellar tip hair, the PI-hair(Inouye et al. 1990; Marin and Melkonian 1994;Moestrup and Hori 1989; Sym and Pienaar 1993).These genera were previously grouped into either asingle family (Halosphaeraceae; Moestrup andThrondsen 1988) or two closely related families(Pterospermataceae and Pyramimonadaceae; Melkonian 1990a). All members of this clade (to whichone should probably add the genera Tasmanites andPrasinopapilla which were not included in this study;Sym and Pienaar 1993) in the motile state have atleast four flagella, a cruciate flagellar root systemand a complex scaly cell covering. Surprisingly themonophyly of this clade was not revealed in amolecular phylogenetic study of rbcl sequences ofdiverse prasinophytes (Daugbjerg et al. 1995). On afiner taxonomic scale the monophyly of the generaHalosphaera and Cymbomonas to the exclusion ofPterosperma and Pyramimonas as shown in thisstudy was somewhat unexpected. Both, Halosphaera and Cymbomonas show common traits inthe structure of their flagellar basal apparatuses(Hori et al. 1985; Inouye, unpublished observations),
but they have not previously been allied in a formalway. The phylogenetic relationships between thegenera Pterosperma and Pyramimonas as well asamong species of Pyramimonas could not be resolved using SSU rRNA sequences.
Clade II. This very well supported clade representing the genera Mamiella, Mantoniella and Micromonas in all phylogenetic analyses (except for theneighbor joining analysis of the global dataset,where it was sister to clade I; Fig. 1) emerged as thesecond divergence within the Chlorophyta. Clade IIhas previously been recognized as a distinct orderwithin the prasinophytes, namely the Mamiella/es(Melkonian 1990a; Moestrup 1984; Moestrup andThrondsen 1988). In his emendation of this orderMoestrup (1991) added the coccoid Pycnococcusprovasolii (as 'Omega 48', = CCMP 1203; see Guillard et al. 1991) as a separate familiy to the order, aplacement which is not supported by the present investigation nor by a previous phylogenetic analysisusing partial rbcl sequences (Daugbjerg et al.1995). And whereas Moestrup and Throndsen(1988) and Moestrup (1991) hesitated to include thescale-less Micromonas pusilla in the Mamiellales,molecular phylogenetic analyses have beyond reasonable doubt positioned Micromonas within theMamiellales (Daugbjerg et al. 1995; Kantz et al. 1990and this study). The Mamiellales (to which Dolichomastix and the flagella-less genus Bathycoccusshould be added; Daugbjerg et al. 1995; Egeland etal. 1995; Eikrem and Throndsen 1990; Throndsenand Zingone 1997) are characterized by a number ofstructural features most of which can now be regarded as 'loss of' characters: lack of an underlayerof square-shaped scales, lack of microtubular flagellar roots attached to basal body no. 2, near-identical spiderweb-like scales on flagellum (if present)and cell body. These characters are clearly apomorphic within the Mamiellales, but they also occur as'loss of' characters in other green algal lineages (e.g.loss of square-shaped scales from the flagellar andcell surfaces of reproductive cells of most 'advanced' Chlorophyta, and loss of microtubular flagellar roots from basal body no. 2 in reproductivecells of several lineages of the Streptophyta). Theonly true synapomorphy of the Mamiellales knownto us is the presence of two rows of distal subunitson flagellar T-hairs (Marin and Melkonian 1994). It ispossible though, that a specific signature of chloroplast pigments may also characterize this group(e.g. the presence of the xanthophyll 'uriolide'; thexanthophyll prasinoxanthin is, however, not restricted to the Mamiellales but occurs also in Pycnococcus provasolii and other coccoid prasinophytes;Fawley 1992; Foss et al. 1984). Furthermore, we can
374 T. Nakayama et al.
Moestrup and Throndsen 1988 (emended byMoestrup 1991) or for that matter the Micromonadophyceae sensu Mattox and Stewart 1984 (the Micromonadophyceae included flagellates with a persistent interzonal spindle; this assemblage is conceptually different from the Prasinophyceae because itexcludes the thecate prasinophytes but includesgenera lacking scales such as Pedinomonas or having non-prasinophyte-type scales such as Monomastix).
Four independent lineages of prasinophytes
All methods of phylogenetic inference used in thisstudy resolved the prasinophyte taxa into at leastfour independent clades (for a discussion of the position of Pycnococcus provasolii see below) whichare designated here as clades I-IV.
Clade I. This well-supported clade representingthe genera Halosphaera, Cymbomonas, Pterosperma and Pyramimonas emerged from all phylogenetic analyses as the earliest divergence within theChlorophyta. Based on a number of synapomorphiccharacters these taxa have always been regardedas relatively closely related. They share a helicalstructure (transitional helix or coiled fiber) in theflagellar transitional region (this structure is foundnowhere else in the Viridiplantae), nine longitudinalrows of Iimulus-shaped flagellar scales, a striatedfiber (termed duct fiber) associated with the 1dmicrotubular flagellar root, a characteristic system IIfiber/nucleus/microbody/chloroplast association,and a unique type of flagellar tip hair, the PI-hair(Inouye et al. 1990; Marin and Melkonian 1994;Moestrup and Hori 1989; Sym and Pienaar 1993).These genera were previously grouped into either asingle family (Halosphaeraceae; Moestrup andThrondsen 1988) or two closely related families(Pterospermataceae and Pyramimonadaceae; Melkonian 1990a). All members of this clade (to whichone should probably add the genera Tasmanites andPrasinopapilla which were not included in this study;Sym and Pienaar 1993) in the motile state have atleast four flagella, a cruciate flagellar root systemand a complex scaly cell covering. Surprisingly themonophyly of this clade was not revealed in amolecular phylogenetic study of rbcl sequences ofdiverse prasinophytes (Daugbjerg et al. 1995). On afiner taxonomic scale the monophyly of the generaHalosphaera and Cymbomonas to the exclusion ofPterosperma and Pyramimonas as shown in thisstudy was somewhat unexpected. Both, Halosphaera and Cymbomonas show common traits inthe structure of their flagellar basal apparatuses(Hori et al. 1985; Inouye, unpublished observations),
but they have not previously been allied in a formalway. The phylogenetic relationships between thegenera Pterosperma and Pyramimonas as well asamong species of Pyramimonas could not be resolved using SSU rRNA sequences.
Clade II. This very well supported clade representing the genera Mamiella, Mantoniella and Micromonas in all phylogenetic analyses (except for theneighbor joining analysis of the global dataset,where it was sister to clade I; Fig. 1) emerged as thesecond divergence within the Chlorophyta. Clade IIhas previously been recognized as a distinct orderwithin the prasinophytes, namely the Mamiella/es(Melkonian 1990a; Moestrup 1984; Moestrup andThrondsen 1988). In his emendation of this orderMoestrup (1991) added the coccoid Pycnococcusprovasolii (as 'Omega 48', = CCMP 1203; see Guillard et al. 1991) as a separate familiy to the order, aplacement which is not supported by the present investigation nor by a previous phylogenetic analysisusing partial rbcl sequences (Daugbjerg et al.1995). And whereas Moestrup and Throndsen(1988) and Moestrup (1991) hesitated to include thescale-less Micromonas pusilla in the Mamiellales,molecular phylogenetic analyses have beyond reasonable doubt positioned Micromonas within theMamiellales (Daugbjerg et al. 1995; Kantz et al. 1990and this study). The Mamiellales (to which Dolichomastix and the flagella-less genus Bathycoccusshould be added; Daugbjerg et al. 1995; Egeland etal. 1995; Eikrem and Throndsen 1990; Throndsenand Zingone 1997) are characterized by a number ofstructural features most of which can now be regarded as 'loss of' characters: lack of an underlayerof square-shaped scales, lack of microtubular flagellar roots attached to basal body no. 2, near-identical spiderweb-like scales on flagellum (if present)and cell body. These characters are clearly apomorphic within the Mamiellales, but they also occur as'loss of' characters in other green algal lineages (e.g.loss of square-shaped scales from the flagellar andcell surfaces of reproductive cells of most 'advanced' Chlorophyta, and loss of microtubular flagellar roots from basal body no. 2 in reproductivecells of several lineages of the Streptophyta). Theonly true synapomorphy of the Mamiellales knownto us is the presence of two rows of distal subunitson flagellar T-hairs (Marin and Melkonian 1994). It ispossible though, that a specific signature of chloroplast pigments may also characterize this group(e.g. the presence of the xanthophyll 'uriolide'; thexanthophyll prasinoxanthin is, however, not restricted to the Mamiellales but occurs also in Pycnococcus provasolii and other coccoid prasinophytes;Fawley 1992; Foss et al. 1984). Furthermore, we can

now with confidence resolve the sequence of divergence of the three genera Mamiella, Mantoniella andMieromonas. As expected, Mamiella is the earliestdivergence in the clade with the genera Mantoniellaand Mieromonas as later divergences and displaying a sister group relationship. The same topology(but without significant bootstrap support) was obtained in the analyses of Daugbjerg et al. (1995)using the 1094-basepair fragment of rbel. Aspresently conceived the three genera of the Mamiellales represent a reduction series in cell size andflagellar number. In Mamiella the web-like flagellarscales have a distinct adnate spine reminiscent ofthe limulus-type flagellar scales in clade I (Moestrup1990; Sym and Pienaar 1993) from which they presumably evolved, whereas in Mantoniella flagellarand cell body scales have become much more similar to each other; finally all scales have been lost inthe minute Mieromonas. The principal conclusionfrom the sequence of divergences of clades I and IIis that the ancestor of clade II (Le. the Mamiellales)almost certainly had an underlayer of squareshaped scales (this occurs in both clade I and in thesister group of the Chlorophyta, the Streptophyta)which was lost during the early evolution of clade IIperhaps related to the diminution of cell size (as discussed in Melkonian and Surek 1995; see alsoDaugbjerg et al. 1995).
Clades III and IV. These two clades representingthe orders Pseudoscourfieldiales (clade III) andChlorodendrales (clade IV) sensu Melkonian (1990a)have been previously recognized in a phylogeneticanalysis using SSU rRNA sequence comparisons bySteink6tter et al. (1994). The present study has corroborated these findings using a much larger datasetof prasinophyte taxa. Whereas the Chlorodendralesare supported by bootstrap values of 100% in allmethods of analysis and branch off close to thenodes separating the 'advanced' lineages of Chlorophyta (Le. Ulvophyceae, Trebouxiophyceae andChlorophyceae), clade III diverges before the Chlorodendrales and is well separated from the latter by along branch common to the Chlorodendrales and the'advanced' classes of Chlorophyta. Clade III is supported by lower bootstrap values (>80%) than cladeIV. A morphological synapomorphy of clade III is thepresence of three microtubular flagellar roots (1 s, 1dand 2d; terminology: Moestrup and Hori 1989; in thegenus Pseudoseourfieldia the 2d root is little developed). In clade IV the unique cell covering (a periplastof fused scales commonly known as the theca; review: Becker et al. 1994) and the presence ofrhizankyrae (Melkonian and Preisig 1986) can becited as morphological synapomorphies. Both cladesare further distinguished from other prasinophytes by
Prasinophyte Ancestry of the Chlorophyta 375
the presence of rod-shaped 'double' scales on theflagellar surface (in Pseudoseourfieldia also on thecell body; Moestrup and Throndsen 1988). This scaletype is clearly an apomorphic character which presumably evolved in the ancestor of a lineage unitingthe Pseudoscourfieldiales, the Chlorodendrales andthe 'advanced' classes of Chlorophyta. The rodshaped scales were then presumably lost after the divergence of the Chlorodendrales but before the radiation of the 'advanced' groups of Chlorophyta. Concerning the number of gains and losses of this character during evolution, we note that our phylogeneticanalysis is much more parsimonious (one gain andone loss) than a previous analysis using the 1094basepair fragment of rbel (2-3 gains and 3-4 losses;Daugbjerg et al. 1995).
The phylogenetic status of Pycnococcusprovasolii
Over the last 15 years, the great diversity and abundance of marine picoplanktonic algae has becomeevident and sparked off an enormous amount of research (summarized by Fogg 1991; 1995, andStockner 1988). Based on their pigment signature,Hooks et al. (1988) distinguished four distinctgroups of eukaryotic picoplankters, two of whichhad pigments found only in prasinophytes. Foss etal. (1986) had previously shown that four clones ofcoccoid picoplankters contained both the chlorophyll c-like pigment Mg-DVP (Mg-2,4-divinyl phaeoporphyrin as monomethyl ester) and the xanthophyllprasinoxanthin, and thus resembled members of theMamiellales (Ricketts 1970). When Guillard et al.(1991) described some of these clones as a newgenus Pyenoeoeeus, they and also Moestrup (1991)placed this genus as a separate family in theMamiellales. Fawley (1992) investigated the pigmentcomposition of Pseudoseourfieldia marina (strain K0017; Moestrup and Throndsen 1988), which wasmost similar to P. provasolii but differed (e.g. in theabsence of the xanthophyll 'uriolide') from that ofthe Mamiellales. Two further coccoid prasinophytes(strains CCMP 1202 and CCMP 1194) had pigmentsignatures which differed from P. provasolii but werevery similar to the Mamiellales (Fawley 1992). It isthus likely that the coccoid scale-less prasinophytesconstitute several (at least two) phylogeneticlineages. All coccoid prasinophytes investigated todate share the presence of prasinoxanthin with theMamiellales and P. marina. Our phylogenetic analyses indicate that prasinoxanthin has presumablyevolved in the ancestor of a lineage that unites allChlorophyta to the exclusion of clade I. Further, theydo not lend support to the previous assignment of P.
now with confidence resolve the sequence of divergence of the three genera Mamiella, Mantoniella andMieromonas. As expected, Mamiella is the earliestdivergence in the clade with the genera Mantoniellaand Mieromonas as later divergences and displaying a sister group relationship. The same topology(but without significant bootstrap support) was obtained in the analyses of Daugbjerg et al. (1995)using the 1094-basepair fragment of rbel. Aspresently conceived the three genera of the Mamiellales represent a reduction series in cell size andflagellar number. In Mamiella the web-like flagellarscales have a distinct adnate spine reminiscent ofthe limulus-type flagellar scales in clade I (Moestrup1990; Sym and Pienaar 1993) from which they presumably evolved, whereas in Mantoniella flagellarand cell body scales have become much more similar to each other; finally all scales have been lost inthe minute Mieromonas. The principal conclusionfrom the sequence of divergences of clades I and IIis that the ancestor of clade II (Le. the Mamiellales)almost certainly had an underlayer of squareshaped scales (this occurs in both clade I and in thesister group of the Chlorophyta, the Streptophyta)which was lost during the early evolution of clade IIperhaps related to the diminution of cell size (as discussed in Melkonian and Surek 1995; see alsoDaugbjerg et al. 1995).
Clades III and IV. These two clades representingthe orders Pseudoscourfieldiales (clade III) andChlorodendrales (clade IV) sensu Melkonian (1990a)have been previously recognized in a phylogeneticanalysis using SSU rRNA sequence comparisons bySteink6tter et al. (1994). The present study has corroborated these findings using a much larger datasetof prasinophyte taxa. Whereas the Chlorodendralesare supported by bootstrap values of 100% in allmethods of analysis and branch off close to thenodes separating the 'advanced' lineages of Chlorophyta (Le. Ulvophyceae, Trebouxiophyceae andChlorophyceae), clade III diverges before the Chlorodendrales and is well separated from the latter by along branch common to the Chlorodendrales and the'advanced' classes of Chlorophyta. Clade III is supported by lower bootstrap values (>80%) than cladeIV. A morphological synapomorphy of clade III is thepresence of three microtubular flagellar roots (1 s, 1dand 2d; terminology: Moestrup and Hori 1989; in thegenus Pseudoseourfieldia the 2d root is little developed). In clade IV the unique cell covering (a periplastof fused scales commonly known as the theca; review: Becker et al. 1994) and the presence ofrhizankyrae (Melkonian and Preisig 1986) can becited as morphological synapomorphies. Both cladesare further distinguished from other prasinophytes by
Prasinophyte Ancestry of the Chlorophyta 375
the presence of rod-shaped 'double' scales on theflagellar surface (in Pseudoseourfieldia also on thecell body; Moestrup and Throndsen 1988). This scaletype is clearly an apomorphic character which presumably evolved in the ancestor of a lineage unitingthe Pseudoscourfieldiales, the Chlorodendrales andthe 'advanced' classes of Chlorophyta. The rodshaped scales were then presumably lost after the divergence of the Chlorodendrales but before the radiation of the 'advanced' groups of Chlorophyta. Concerning the number of gains and losses of this character during evolution, we note that our phylogeneticanalysis is much more parsimonious (one gain andone loss) than a previous analysis using the 1094basepair fragment of rbel (2-3 gains and 3-4 losses;Daugbjerg et al. 1995).
The phylogenetic status of Pycnococcusprovasolii
Over the last 15 years, the great diversity and abundance of marine picoplanktonic algae has becomeevident and sparked off an enormous amount of research (summarized by Fogg 1991; 1995, andStockner 1988). Based on their pigment signature,Hooks et al. (1988) distinguished four distinctgroups of eukaryotic picoplankters, two of whichhad pigments found only in prasinophytes. Foss etal. (1986) had previously shown that four clones ofcoccoid picoplankters contained both the chlorophyll c-like pigment Mg-DVP (Mg-2,4-divinyl phaeoporphyrin as monomethyl ester) and the xanthophyllprasinoxanthin, and thus resembled members of theMamiellales (Ricketts 1970). When Guillard et al.(1991) described some of these clones as a newgenus Pyenoeoeeus, they and also Moestrup (1991)placed this genus as a separate family in theMamiellales. Fawley (1992) investigated the pigmentcomposition of Pseudoseourfieldia marina (strain K0017; Moestrup and Throndsen 1988), which wasmost similar to P. provasolii but differed (e.g. in theabsence of the xanthophyll 'uriolide') from that ofthe Mamiellales. Two further coccoid prasinophytes(strains CCMP 1202 and CCMP 1194) had pigmentsignatures which differed from P. provasolii but werevery similar to the Mamiellales (Fawley 1992). It isthus likely that the coccoid scale-less prasinophytesconstitute several (at least two) phylogeneticlineages. All coccoid prasinophytes investigated todate share the presence of prasinoxanthin with theMamiellales and P. marina. Our phylogenetic analyses indicate that prasinoxanthin has presumablyevolved in the ancestor of a lineage that unites allChlorophyta to the exclusion of clade I. Further, theydo not lend support to the previous assignment of P.

376 T. Nakayama et al.
provasolii to the Mamiellales made on the basis ofthe presence of Mg-DVP and prasinoxanthin [MgDVP is a known precursor to chlorophyll a and hasbeen found in taxa of clade I, in other algal phyla(e.g. in cryptophytes; Schimek et al. 1994) and evenin photosynthetic bacteria (Jones 1963)]. Whether P.provasolii is a representative of a fifth indepedentlineage among the prasinophytes or should perhapsbe included in clade III (see Results) cannot be decided at present and must await the determinationof SSU rRNA sequences from additional strains ofcoccoid prasinophytes and subsequent phylogenetic analyses. Adding new SSU rRNA sequencesfrom taxa of clades II and III to the analyses has neither disrupted the monophyly of each clade nor thesequence of divergence of clades !-IV (Marin andMelkonian, unpublished observations). Here we onlynote that the close phylogenetic relationship previously deduced from pigment signatures and rbclsequences between P. provasolii and Pseudoscourfieldia marina (Daugbjerg et al. 1995; Fawley 1992)has not been borne out by our analysis. However,phylogenenetic analyses based on SSU rRNA sequences (Kantz et al. 1990; Steinkotter et al. 1994;this study) used a different strain of P. marina(CCMP 717). Most likely the two strains currentlydesignated P. marina (CCMP 717 and K-0017) donot refer to the same taxon (genus).
Classification of the prasinophytes and theChlorophyta
As the prasinophytes are at least paraphyletic(Steinkotter et al. 1994; this study) and most likelyeven polyphyletic (Marin and Melkonian, unpublished observations) they should no longer be recognized as a single taxonomic entity (see above). In thepresent study we distinguish four clades of taxawhich have traditionally been recognized as prasinophytes. These represent basal lineages within theChlorophyta, whereas the genus Mesostigma is amember of the Streptophyta (Marin and Melkonian,unpublished observations; Melkonian et al. 1995).
How do the four clades compare with previousclassification schemes of the prasinophytes? Theclassification scheme advanced by Moestrup andThrondsen and emended by Moestrup (Moestrup1991; Moestrup and Throndsen 1988) is no longertenable as their order Chlorodendrales is polyphyletic including members of clades I, III and IV.The Chlorodendrales sensu Moestrup and Throndsen was apparently based on a single character(presence of an underlayer of square-shapedscales) which is now regarded as a plesiomorphy ofthe whole group Viridiplantae. In their study of flag-
ellar hair ultrastructure in the prasinophytes Marinand Melkonian (1994) already indicated that theChlorodendrales sensu Moestrup and Throndsenappeared heterogeneous with respect to flagellarhair types. The four clades correspond most closelyto the classification scheme proposed by Melkonian(1990a) who distinguished four orders of prasinophytes: the Pyramimonadales (clade I), the Mamiellales (clade II; this order is equivalent to the Mamiellales sensu Moestrup and Throndsen), the Pseudoscourfieldiales (clade III), and the Chlorodendrales sensu Melkonian and Preisig (clade IV). Thesingle notable exception is Mesostigma viride whichwas placed (as a separate family) in the Pyramimonadales by Melkonian (1990a) but based on SSUrRNA sequence comparisons is a member of theStreptophyta (Marin and Melkonian, unpublishedobservations). The four clades of prasinophytes andtheir sequence of divergence are also supported bysynapomorphic morphological characters (see discussion above). In a future revised classification ofthe Chlorophyta the four clades of prasinophyteswill likely attain a more prominent taxonomic statusthan presently perceived. Another question thatneeds to be addressed is whether a strictly hierarchical classification should be applied to the Chlorophyta. If such an approach is adopted it would haveto take into account that several of the later diverging 'advanced' groups of green algae are currentlyrecognized at the class level, i.e. the Ulvophyceae,the Chlorophyceae, and the Trebouxiophyceae(Friedl 1997). Some authors have also consideredthe presence of additional classes in the Chlorophyta (van den Hoek et al. 1995; Moestrup 1991).Again their formal status can only be assessed in arevised phylogeny that takes into account the basalposition of four independent lineages of scaly greenflagellates within the Chlorophyta.
Methods
The origins of the prasinophyte strains used in thisstudy and accession numbers of SSU rRNA sequences (EMBUGenBank) are summarized in Table3. Cultures not available from other sources can beobtained from the authors. Total genomic DNA wasisolated by phenol/chloroform extraction (Kranz etal. 1995; Sambrook et al. 1989) or by the CTAB protocol (Surek et al. 1994). Nuclear-encoded rRNAgenes of Cymbomonas tetramitiformis, Halosphaerasp., Mamiella sp., Mantoniella antarctica, Pterosperma cristatum (Yokohama strain), Pyramimonasdisomata, P. olivacea, P. parkeae, and P. propulsawere amplified by polymerase chain reaction (PCR)
376 T. Nakayama et al.
provasolii to the Mamiellales made on the basis ofthe presence of Mg-DVP and prasinoxanthin [MgDVP is a known precursor to chlorophyll a and hasbeen found in taxa of clade I, in other algal phyla(e.g. in cryptophytes; Schimek et al. 1994) and evenin photosynthetic bacteria (Jones 1963)]. Whether P.provasolii is a representative of a fifth indepedentlineage among the prasinophytes or should perhapsbe included in clade III (see Results) cannot be decided at present and must await the determinationof SSU rRNA sequences from additional strains ofcoccoid prasinophytes and subsequent phylogenetic analyses. Adding new SSU rRNA sequencesfrom taxa of clades II and III to the analyses has neither disrupted the monophyly of each clade nor thesequence of divergence of clades !-IV (Marin andMelkonian, unpublished observations). Here we onlynote that the close phylogenetic relationship previously deduced from pigment signatures and rbclsequences between P. provasolii and Pseudoscourfieldia marina (Daugbjerg et al. 1995; Fawley 1992)has not been borne out by our analysis. However,phylogenenetic analyses based on SSU rRNA sequences (Kantz et al. 1990; Steinkotter et al. 1994;this study) used a different strain of P. marina(CCMP 717). Most likely the two strains currentlydesignated P. marina (CCMP 717 and K-0017) donot refer to the same taxon (genus).
Classification of the prasinophytes and theChlorophyta
As the prasinophytes are at least paraphyletic(Steinkotter et al. 1994; this study) and most likelyeven polyphyletic (Marin and Melkonian, unpublished observations) they should no longer be recognized as a single taxonomic entity (see above). In thepresent study we distinguish four clades of taxawhich have traditionally been recognized as prasinophytes. These represent basal lineages within theChlorophyta, whereas the genus Mesostigma is amember of the Streptophyta (Marin and Melkonian,unpublished observations; Melkonian et al. 1995).
How do the four clades compare with previousclassification schemes of the prasinophytes? Theclassification scheme advanced by Moestrup andThrondsen and emended by Moestrup (Moestrup1991; Moestrup and Throndsen 1988) is no longertenable as their order Chlorodendrales is polyphyletic including members of clades I, III and IV.The Chlorodendrales sensu Moestrup and Throndsen was apparently based on a single character(presence of an underlayer of square-shapedscales) which is now regarded as a plesiomorphy ofthe whole group Viridiplantae. In their study of flag-
ellar hair ultrastructure in the prasinophytes Marinand Melkonian (1994) already indicated that theChlorodendrales sensu Moestrup and Throndsenappeared heterogeneous with respect to flagellarhair types. The four clades correspond most closelyto the classification scheme proposed by Melkonian(1990a) who distinguished four orders of prasinophytes: the Pyramimonadales (clade I), the Mamiellales (clade II; this order is equivalent to the Mamiellales sensu Moestrup and Throndsen), the Pseudoscourfieldiales (clade III), and the Chlorodendrales sensu Melkonian and Preisig (clade IV). Thesingle notable exception is Mesostigma viride whichwas placed (as a separate family) in the Pyramimonadales by Melkonian (1990a) but based on SSUrRNA sequence comparisons is a member of theStreptophyta (Marin and Melkonian, unpublishedobservations). The four clades of prasinophytes andtheir sequence of divergence are also supported bysynapomorphic morphological characters (see discussion above). In a future revised classification ofthe Chlorophyta the four clades of prasinophyteswill likely attain a more prominent taxonomic statusthan presently perceived. Another question thatneeds to be addressed is whether a strictly hierarchical classification should be applied to the Chlorophyta. If such an approach is adopted it would haveto take into account that several of the later diverging 'advanced' groups of green algae are currentlyrecognized at the class level, i.e. the Ulvophyceae,the Chlorophyceae, and the Trebouxiophyceae(Friedl 1997). Some authors have also consideredthe presence of additional classes in the Chlorophyta (van den Hoek et al. 1995; Moestrup 1991).Again their formal status can only be assessed in arevised phylogeny that takes into account the basalposition of four independent lineages of scaly greenflagellates within the Chlorophyta.
Methods
The origins of the prasinophyte strains used in thisstudy and accession numbers of SSU rRNA sequences (EMBUGenBank) are summarized in Table3. Cultures not available from other sources can beobtained from the authors. Total genomic DNA wasisolated by phenol/chloroform extraction (Kranz etal. 1995; Sambrook et al. 1989) or by the CTAB protocol (Surek et al. 1994). Nuclear-encoded rRNAgenes of Cymbomonas tetramitiformis, Halosphaerasp., Mamiella sp., Mantoniella antarctica, Pterosperma cristatum (Yokohama strain), Pyramimonasdisomata, P. olivacea, P. parkeae, and P. propulsawere amplified by polymerase chain reaction (PCR)

Prasinophyte Ancestry of the Chlorophyta 377
Table 3. Prasinophyte strains used in this study, their origins and accession numbers of SSU rRNA sequences.CCMP = Provasoli-Guillard National Center for Culture of Marine Phytoplankton, West Boothbay Harbor, Maine,USA; NIES = Microbial Culture Collection. National Institute for Environmental Studies, Tsukuba, Japan
Taxon
Coccoid unidentified prasinophyteCymbomonas tetramitiformis SchillerHalosphaera sp.Mamiella sp.Mantoniella antarctica MarchantMicromonas pusilla (Butcher) Manton et ParkePterosperma cristatum SchillerPterosperma cristatum SchillerPycnococcus provasolii GuillardPyramimonas disomata ButcherPyramimonas olivacea N.CarterPyramimonas parkeae Norris et PearsonPyramimonas propulsa Moestrup et Hill
Strain
CCMP 1198Shizugawa, Miyagi, JapanShizugawa, Miyagi, JapanShizugawa, Miyagi, JapanLUtzow Holmbukta, AntarcticaCCMP489Yokohama, Kanagawa,JapanNIES 221CCMP 1203Changi, SingaporeShizugawa, Miyagi, JapanHachijo Is., Tokyo, JapanNIES 251 ('P. aft. amyliferaj
Accession no.
AJ010406AB017126AB017125AB017129AB017128AJ010408AB017127AJ010407X91264AB017121AB017122AB017124AB017123
using oligonucleotide primers 'SR1' (5'-TACCTGGTTGATCCTGCCAG-3') and 'SR12' (5'-CCTTCCGCAGGTTCACCTAC-3'). The remaining taxa wereamplified as described previously (Huss and Sogin1990; Surek et al. 1994). Complete SSU rRNA sequences were determined for both strands by solidphase sequencing according to Surek et al. (1994)or after a second PCR using the following six pairsof primers 'SR1 '/'SR3' (5'-AGGCTCCCTGTCCGGAATC-3'), 'SR2'/'SR5' (5'-CATTCAAATTTCTGCCCTATC-3' / 5'-ACTACGAGCTTTTTAACTGC-3'),'SR4'/'SRT (5'-AGCCGCGGTAATTCCAGCT-3' / 5'TCCTTGGCAAATGCTTTCGC-3'), 'SR6'/'SR9' (5'GTCAGAGGTGAAATTCTTGG-3' / 5'-AACTAAGAACGGCCATGCAC-3'), 'SR8'/,SR11' (5'-GGATTGACAGATTGAGAGCT-3' / 5' -CGCTTACTAGGAATTCCTCG-3'), 'SR10'/'SR12' (5'-AGGTCTGTGATGCCCTTAGA-3'). These PCR-products were purifiedusing polyethylene glycol (Sambrook et al. 1989)and directly sequenced with the Dye TerminatorCycle Sequencing Kit on an DNA autosequencermodel 373A from Applied Biosystems.
SSU rRNAs were manually aligned and only regions which could be unambiguously aligned wereused in the phylogenetic analyses (the alignment isavailable from the authors upon request). A datasetwith 1655 positions was used as input for distanceand maximum parsimony analyses of 66 taxa ofViridiplantae (with 3 glaucocystophyte taxa as outgroups) to position the prasinophyte taxa among theViridiplantae, whereas a smaller dataset of 21 taxaof Chlorophyta (with two taxa of the Streptophyta asoutgroups) with 1699 positions was used as inputfor distance, maximum parsimony, and maximumlikelihood analyses to evaluate the position of the
prasinophyte taxa within the Chlorophyta and relative to each other. Distance analyses were performed with the PHYLIP package (PHYLIP version3.5c; Felsenstein 1993), DNAdist was used to generate distance matrices according to the two-parameter model of Kimura (1980), which were converted to phylogenetic trees using the neighbor joining method (Saitou and Nei 1987). Maximum parsimony analyses were implemented with the programPAUP (version 3.1.1.; Swofford 1993). Analyseswere done using a weighting scheme (rescaled consistency index over an interval of 1-1000) for eachsite within the aligned dataset (Hillis et al. 1994).Random additions of the weighted data were analyzed using the heuristic search option with abranch-swapping algorithm (tree bisection-reconnection). For maximum likelihood analyses, thefastDNAml program (version 1.0; Olsen et al. 1994)was used with the global search option and jumbledtaxon addition; a transition/transversion ratio of 2.0was used since analyses with the observed ratio(1.3) did not change the results. All phylogeneticanalyses were rooted with different outgroups (seeResults for details). Bootstrap analyses (100 replications for maximum parsimony and maximum likelihood analyses, 100 or 500 replications for neighborjoining analyses; Felsenstein 1985) were used to assess the stability of monophyletic groups. The following reference sequences (with accession numbers) were used in the phylogenetic analyses: Acrosiphonia sp. (U03757), Ankistrodesmus stipitatus(X561 00), Anthoceros agrestis (X80984), Asteromonas gracilis (M95614), Chara foetida (X70704),Characium hindakii (M63000), Chlamydomonas reinhardtii (M32703), Chlamydopodium (Characium) starrii
Prasinophyte Ancestry of the Chlorophyta 377
Table 3. Prasinophyte strains used in this study, their origins and accession numbers of SSU rRNA sequences.CCMP = Provasoli-Guillard National Center for Culture of Marine Phytoplankton, West Boothbay Harbor, Maine,USA; NIES = Microbial Culture Collection. National Institute for Environmental Studies, Tsukuba, Japan
Taxon
Coccoid unidentified prasinophyteCymbomonas tetramitiformis SchillerHalosphaera sp.Mamiella sp.Mantoniella antarctica MarchantMicromonas pusilla (Butcher) Manton et ParkePterosperma cristatum SchillerPterosperma cristatum SchillerPycnococcus provasolii GuillardPyramimonas disomata ButcherPyramimonas olivacea N.CarterPyramimonas parkeae Norris et PearsonPyramimonas propulsa Moestrup et Hill
Strain
CCMP 1198Shizugawa, Miyagi, JapanShizugawa, Miyagi, JapanShizugawa, Miyagi, JapanLUtzow Holmbukta, AntarcticaCCMP489Yokohama, Kanagawa,JapanNIES 221CCMP 1203Changi, SingaporeShizugawa, Miyagi, JapanHachijo Is., Tokyo, JapanNIES 251 ('P. aft. amyliferaj
Accession no.
AJ010406AB017126AB017125AB017129AB017128AJ010408AB017127AJ010407X91264AB017121AB017122AB017124AB017123
using oligonucleotide primers 'SR1' (5'-TACCTGGTTGATCCTGCCAG-3') and 'SR12' (5'-CCTTCCGCAGGTTCACCTAC-3'). The remaining taxa wereamplified as described previously (Huss and Sogin1990; Surek et al. 1994). Complete SSU rRNA sequences were determined for both strands by solidphase sequencing according to Surek et al. (1994)or after a second PCR using the following six pairsof primers 'SR1 '/'SR3' (5'-AGGCTCCCTGTCCGGAATC-3'), 'SR2'/'SR5' (5'-CATTCAAATTTCTGCCCTATC-3' / 5'-ACTACGAGCTTTTTAACTGC-3'),'SR4'/'SRT (5'-AGCCGCGGTAATTCCAGCT-3' / 5'TCCTTGGCAAATGCTTTCGC-3'), 'SR6'/'SR9' (5'GTCAGAGGTGAAATTCTTGG-3' / 5'-AACTAAGAACGGCCATGCAC-3'), 'SR8'/,SR11' (5'-GGATTGACAGATTGAGAGCT-3' / 5' -CGCTTACTAGGAATTCCTCG-3'), 'SR10'/'SR12' (5'-AGGTCTGTGATGCCCTTAGA-3'). These PCR-products were purifiedusing polyethylene glycol (Sambrook et al. 1989)and directly sequenced with the Dye TerminatorCycle Sequencing Kit on an DNA autosequencermodel 373A from Applied Biosystems.
SSU rRNAs were manually aligned and only regions which could be unambiguously aligned wereused in the phylogenetic analyses (the alignment isavailable from the authors upon request). A datasetwith 1655 positions was used as input for distanceand maximum parsimony analyses of 66 taxa ofViridiplantae (with 3 glaucocystophyte taxa as outgroups) to position the prasinophyte taxa among theViridiplantae, whereas a smaller dataset of 21 taxaof Chlorophyta (with two taxa of the Streptophyta asoutgroups) with 1699 positions was used as inputfor distance, maximum parsimony, and maximumlikelihood analyses to evaluate the position of the
prasinophyte taxa within the Chlorophyta and relative to each other. Distance analyses were performed with the PHYLIP package (PHYLIP version3.5c; Felsenstein 1993), DNAdist was used to generate distance matrices according to the two-parameter model of Kimura (1980), which were converted to phylogenetic trees using the neighbor joining method (Saitou and Nei 1987). Maximum parsimony analyses were implemented with the programPAUP (version 3.1.1.; Swofford 1993). Analyseswere done using a weighting scheme (rescaled consistency index over an interval of 1-1000) for eachsite within the aligned dataset (Hillis et al. 1994).Random additions of the weighted data were analyzed using the heuristic search option with abranch-swapping algorithm (tree bisection-reconnection). For maximum likelihood analyses, thefastDNAml program (version 1.0; Olsen et al. 1994)was used with the global search option and jumbledtaxon addition; a transition/transversion ratio of 2.0was used since analyses with the observed ratio(1.3) did not change the results. All phylogeneticanalyses were rooted with different outgroups (seeResults for details). Bootstrap analyses (100 replications for maximum parsimony and maximum likelihood analyses, 100 or 500 replications for neighborjoining analyses; Felsenstein 1985) were used to assess the stability of monophyletic groups. The following reference sequences (with accession numbers) were used in the phylogenetic analyses: Acrosiphonia sp. (U03757), Ankistrodesmus stipitatus(X561 00), Anthoceros agrestis (X80984), Asteromonas gracilis (M95614), Chara foetida (X70704),Characium hindakii (M63000), Chlamydomonas reinhardtii (M32703), Chlamydopodium (Characium) starrii

378 T. Nakayama et al.
(saccatum) (M84319), Chlorella vulgaris (X13688),Chlorokybus atmophyticus (M95612), Coleochaeteorbicularis (M95611), Coleochaete scutata (X68825),Cosmarium botrytis (X79498), Cyanophora paradoxa(X68483), Cyanoptyche gloeocystis (AJ007275),Dunaliella salina (M84320), Fossombronia pusilla(X78341), Funaria hygrometrica (X74114), Fusochloris perforata (M62999), Genicularia spirotaenia(X74753), Ginkgo biloba (016448), Glaucocystisnostochinearum (X70803), Gloeotilopsis planctonica(Z28970), Hydrodictyon reticulatum (M74497), Hylocomium splendens (X95477), Klebsormidium f1accidum (M95613), Leptosira (Pleurastrum) terrestris(terrestre) (Z28973), Lychnothamnus barbatus(U81272), Mantoniella squamata (X73999), Marchantia polymorpha (X75521), Mesotaenium caldariorum (X75763), Microthamnion kuetzingianum(Z28974), Mnium hornum (X80985), Mougeotia scalaris (X70705), Myrmecia biatorellae (Z28971), Myrmecia israelensis (M62995), Nanochlorum eucaryotum (X06245), Neochloris aquatica (M62861),Nephroselmis olivacea (X74754), Nitella f1exilis(U05261), Oltmannsiellopsis viridis (086495), Pelliaepiphylla (X8021 0), Pediastrum duplex (M62997),Pinus luchuensis (038246), Prototheca wickerham;;(X56099), Pseudendoclonium basiliense (Z47996),Pseudoscourfieldia marina (X75565), Scenedesmusabundans (X73995), Scenedesmus obliquus (X561 03),Scherffelia dubia (X68484), Sphagnum palustre(y11370), Staurastrum sp. (X74752), Tetraselmis striata (X70802), Trebouxia asymmetrica (Z21553), Trebouxia impressa (Z21551), Trebouxia magna(Z21552), UJothrix zonata (Z47999), Volvox carteri f.nagariensis (X53904).
Acknowledgements
BM, BS and MM would like to thank Dr.ChristophSensen (Institute for Marine Biosciences, Halifax) forhelp with bootstrap analyses of maximum likelihoodtrees and Ute Powalowski for keeping stock culturesin Cologne. TN would like to thank Dr. MasanobuKawachi (National Institute of Environmental Sciences, Tsukuba) for providing the culture of Mantoniella antarctica and Dr. Masaya Sato (Universityof Tokushima) for advice on the sequencing technique. This study was supported by the DeutscheForschungsgemeinschaft.
References
Becker B, Marin B, Melkonian M (1994) Structure,composition and biogenesis of prasinophyte cell coverings. Protoplasma 181: 233-244
Bremer K (1985) Summary of green plant phylogenyand classification. Cladistics 1: 369-385
Cavalier-Smith T (1981) Eukaryote kingdoms: seven ornine? BioSystems 14: 461-481
Daugbjerg N, Moestrup 0, Arctander P (1995) Phylogeny of genera of Prasinophyceae and Pedinophyceae (Chlorophyta) deduced from molecular analysis of the rbcl gene. Phycol Res 43: 203-213
Egeland ES, Johnsen G, Eikrem W, Throndsen J, Liaaen-Jensen S (1995) Pigments of Bathycoccus prasinos (Prasinophyceae): methodological and chemosystematic implications. J Phycol 31: 554-561
Eikrem W, Throndsen J (1990) The ultrastructure ofBathycoccus gen. nov. and B. prasinos sp. nov.,a nonmotile picoplanktonic alga (Chlorophyta,Prasinophyceae) from the Mediterranean and Atlantic. Phycologia 29: 344-350
Fawley MW (1992) Photosynthetic pigments of Pseudoscourfieldia marina and select green flagellates andcoccoid ultraphytoplankton: Implications for the systematics of the Micromonadophyceae (Chlorophyta). JPhycoI28:26-31
Felsenstein J (1985) Confidence limits on phylogenies:an approach using the bootstrap. Evolution 39: 783-791
Felsenstein J (1993) PHYLIP (Phylogeny InferencePackage) Manual Version 3.5c. Department of Genetics, University of Washington, Seattle
Fogg GE (1991) Tansley review no. 30. The phytoplanktonic ways of life. New Phytol 118: 191-232
Fogg GE (1995) Some comments on picoplankton andits importance in the pelagic ecosystem. Aquat MicrobEcoI9:33-40
Foss P, Guillard RRL, Liaaen-Jensen S (1984) Prasinoxanthin - a chemosystematic marker for algae. Phytochemistry 23: 1629-1633
Foss P, Guillard RRL, Liaaen-Jensen S (1986) Carotenoids from eucaryotic ultraplankton clones (Prasinophyceae). Phytochemistry 25: 119-124
Friedl T (1997) The Evolution of the Green Algae. InBhattacharya D (ed) The Origin of Algae and Plastids.Springer Verlag, Wien, pp 87-101
Guillard RRL, Keller MD, O'Kelly CJ, Floyd GL, (1991)Pycnococcus provasolii gen. et sp. nov., a coccoidprasinoxanthin containing phytoplankter from the western North Atlantic and Gulf of Mexico. J Phycol 27:39-47
Hillis DM, Huelsenbeck JP, Swofford DL (1994) Hobgoblin of phylogenetics? Nature 369: 363-364
Hoek C van den, Mann DG, Jahns HM (1995) Algae.An Introduction to Phycology. Cambridge UniversityPress, Cambridge
Hooks CE, Bidigare RR, Keller MD, Guillard RRL(1988) Coccoid eukaryotic marine ultraplankters with
378 T. Nakayama et al.
(saccatum) (M84319), Chlorella vulgaris (X13688),Chlorokybus atmophyticus (M95612), Coleochaeteorbicularis (M95611), Coleochaete scutata (X68825),Cosmarium botrytis (X79498), Cyanophora paradoxa(X68483), Cyanoptyche gloeocystis (AJ007275),Dunaliella salina (M84320), Fossombronia pusilla(X78341), Funaria hygrometrica (X74114), Fusochloris perforata (M62999), Genicularia spirotaenia(X74753), Ginkgo biloba (016448), Glaucocystisnostochinearum (X70803), Gloeotilopsis planctonica(Z28970), Hydrodictyon reticulatum (M74497), Hylocomium splendens (X95477), Klebsormidium f1accidum (M95613), Leptosira (Pleurastrum) terrestris(terrestre) (Z28973), Lychnothamnus barbatus(U81272), Mantoniella squamata (X73999), Marchantia polymorpha (X75521), Mesotaenium caldariorum (X75763), Microthamnion kuetzingianum(Z28974), Mnium hornum (X80985), Mougeotia scalaris (X70705), Myrmecia biatorellae (Z28971), Myrmecia israelensis (M62995), Nanochlorum eucaryotum (X06245), Neochloris aquatica (M62861),Nephroselmis olivacea (X74754), Nitella f1exilis(U05261), Oltmannsiellopsis viridis (086495), Pelliaepiphylla (X8021 0), Pediastrum duplex (M62997),Pinus luchuensis (038246), Prototheca wickerham;;(X56099), Pseudendoclonium basiliense (Z47996),Pseudoscourfieldia marina (X75565), Scenedesmusabundans (X73995), Scenedesmus obliquus (X561 03),Scherffelia dubia (X68484), Sphagnum palustre(y11370), Staurastrum sp. (X74752), Tetraselmis striata (X70802), Trebouxia asymmetrica (Z21553), Trebouxia impressa (Z21551), Trebouxia magna(Z21552), UJothrix zonata (Z47999), Volvox carteri f.nagariensis (X53904).
Acknowledgements
BM, BS and MM would like to thank Dr.ChristophSensen (Institute for Marine Biosciences, Halifax) forhelp with bootstrap analyses of maximum likelihoodtrees and Ute Powalowski for keeping stock culturesin Cologne. TN would like to thank Dr. MasanobuKawachi (National Institute of Environmental Sciences, Tsukuba) for providing the culture of Mantoniella antarctica and Dr. Masaya Sato (Universityof Tokushima) for advice on the sequencing technique. This study was supported by the DeutscheForschungsgemeinschaft.
References
Becker B, Marin B, Melkonian M (1994) Structure,composition and biogenesis of prasinophyte cell coverings. Protoplasma 181: 233-244
Bremer K (1985) Summary of green plant phylogenyand classification. Cladistics 1: 369-385
Cavalier-Smith T (1981) Eukaryote kingdoms: seven ornine? BioSystems 14: 461-481
Daugbjerg N, Moestrup 0, Arctander P (1995) Phylogeny of genera of Prasinophyceae and Pedinophyceae (Chlorophyta) deduced from molecular analysis of the rbcl gene. Phycol Res 43: 203-213
Egeland ES, Johnsen G, Eikrem W, Throndsen J, Liaaen-Jensen S (1995) Pigments of Bathycoccus prasinos (Prasinophyceae): methodological and chemosystematic implications. J Phycol 31: 554-561
Eikrem W, Throndsen J (1990) The ultrastructure ofBathycoccus gen. nov. and B. prasinos sp. nov.,a nonmotile picoplanktonic alga (Chlorophyta,Prasinophyceae) from the Mediterranean and Atlantic. Phycologia 29: 344-350
Fawley MW (1992) Photosynthetic pigments of Pseudoscourfieldia marina and select green flagellates andcoccoid ultraphytoplankton: Implications for the systematics of the Micromonadophyceae (Chlorophyta). JPhycoI28:26-31
Felsenstein J (1985) Confidence limits on phylogenies:an approach using the bootstrap. Evolution 39: 783-791
Felsenstein J (1993) PHYLIP (Phylogeny InferencePackage) Manual Version 3.5c. Department of Genetics, University of Washington, Seattle
Fogg GE (1991) Tansley review no. 30. The phytoplanktonic ways of life. New Phytol 118: 191-232
Fogg GE (1995) Some comments on picoplankton andits importance in the pelagic ecosystem. Aquat MicrobEcoI9:33-40
Foss P, Guillard RRL, Liaaen-Jensen S (1984) Prasinoxanthin - a chemosystematic marker for algae. Phytochemistry 23: 1629-1633
Foss P, Guillard RRL, Liaaen-Jensen S (1986) Carotenoids from eucaryotic ultraplankton clones (Prasinophyceae). Phytochemistry 25: 119-124
Friedl T (1997) The Evolution of the Green Algae. InBhattacharya D (ed) The Origin of Algae and Plastids.Springer Verlag, Wien, pp 87-101
Guillard RRL, Keller MD, O'Kelly CJ, Floyd GL, (1991)Pycnococcus provasolii gen. et sp. nov., a coccoidprasinoxanthin containing phytoplankter from the western North Atlantic and Gulf of Mexico. J Phycol 27:39-47
Hillis DM, Huelsenbeck JP, Swofford DL (1994) Hobgoblin of phylogenetics? Nature 369: 363-364
Hoek C van den, Mann DG, Jahns HM (1995) Algae.An Introduction to Phycology. Cambridge UniversityPress, Cambridge
Hooks CE, Bidigare RR, Keller MD, Guillard RRL(1988) Coccoid eukaryotic marine ultraplankters with

four different HPLC pigment signatures. J Phycol 24:571-580
Hori T, Inouye I, Horiguchi T, Boalch GT (1985) Observations on the motile stage of Halosphaera minorOstenfeld (Prasinophyceae) with special reference tothe cell structure. Bot Mar 28: 529-537
Huss VAR, Kranz HD (1997) Charophyte Evolution andthe Origin of Land Plants. In Bhattacharya 0 (ed) Originsof Algae and their Plastids. Springer Verlag, Wien, pp103-114
Huss VAR, Sogin ML (1990) Phylogenetic position ofsome Chlorella species within the Chlorococcalesbased upon complete small-subunit ribosomal RNA sequences. J Mol Evol 31: 432-442
Inouye I, Hori T, Chihara M (1990) Absolute configuration analysis of the flagellar apparatus of Pterospermacristatum (Prasinophyceae) and consideration of itsphylogenetic position. J Phycol 26: 329-344
Jones OTG (1963) Magnesium-2,4-divinylphaeoporphyrin as monomethylester, a protochlorophyll-like pigment produced by Rhodopseudomonas sphaeroides.Biochem J 89: 182-189
Kantz TS, Theriot EC, Zimmer EA, Chapman RL(1990) The Pleurastrophyceae and Micromonadophyceae: a cladistic analysis of nuclear rRNA sequencedata. J Phycol26: 711-721
Kimura M (1980) A simple method for estimating evolutionary rates of base substitution through comparativestudies of sequence evolution. J Mol Evol 16: 111-120
Kranz HD, Miks 0, Siegler M-L, Capesius I, SensenCW, Huss VAR (1995) The origin of land plants: phylogenetic relationships among charophytes, bryophytes,and vascular plants inferred from complete smail-subunit ribosomal RNA gene sequences. J Mol Evol 41:74-84
Leipe DO, Wainright PO, Gunderson JH, Porter 0,Patterson OJ, Valois F, Himmerich S, Sogin ML(1994) The stramenopiles from a molecular perspective:16S-like rRNA sequences from Labyrinthuloides minutaand Cafeteria roenbergensis. Phycologia 33: 369-377
Manton I (1965) Some phyletic implications of flagellarstructure in plants. In Preston RO (ed) Advances inBotanical Research, Vo1.2. Academic Press, pp 1-34
Manton I, Parke M (1960) Further Observations onSmall Green Flagellates with Special Reference to Possible Relatives of Chromulina pusilla Butcher. J Mar BioiAssoc UK 39: 275-298
Marin B, Melkonian M (1994) Flagellar hairs in prasinophytes (Chlorophyta): ultrastructure and distribution onthe flagellar surface. J Phycol 30: 659-678
Mattox KR, Stewart KD (1984) Classification of theGreen Algae: a Concept Based on Comparative Cytology. In Irvine OEG John OM (eds) Systematics of theGreen Algae. Academic Press, London, pp 29-72
Prasinophyte Ancestry of the Chlorophyta 379
McCourt RM (1995) Green algal phylogeny. TrendsEcol Evol 10: 159-163
Melkonian M (1982) Structural and evolutionary aspects of the flagellar apparatus in green algae and landplants. Taxon 31: 255-265
Melkonian M (1984) Flagellar Apparatus Ultrastructurein Relaton to Green Algal Classification. In Irvine OEGJohn OM (eds) Systematics of the Green Algae - Systematics Association Special Volumes, NO.27. Academic Press, London, pp 73-120
Melkonian M (1990a) Phylum Chlorophyta: ClassPrasinophyceae. In Margulis L, Corliss JO, MelkonianM, Chapman 0 (eds) Handbook of Protoctista. Jonesand Bartlett Publishers, Boston, pp 600-607
Melkonian M (1990b) Chlorophyte Orders of UncertainAffinities: Order Microthamniales. In Margulis L, CorlissJO, Melkonian M, Chapman OJ (eds) Handbook of Protoctista. Jones and Bartlett Publishers, Boston, pp652-654
Melkonian M (1996) Systematics and Evolution of theAlgae: Endocytobiosis and Evolution of the Major AlgalLineages. In Behnke H-O, LOttge U, Esser K, KadereitJW, Runge M (eds) Progress in Botany Vol 57. SpringerVerlag, Berlin, pp 281-311
Melkonian M, Marin B, Surek B (1995) Phylogeny andEvolution of the Algae. In Arai R, Kato M, Ooi Y (eds)Biodiversity and Evolution. The National Science Museum Foundation, Tokyo, pp 153-176
Melkonian M, Preisig HR (1986) A light and electronmicroscopic study of Scherffelia dubia, a new memberof the scaly green flagellates (Prasinophyceae). Nord JBot 6: 235-256
Melkonian M, Surek B (1995) Phylogeny of the Chlorophyta: congruence between ultrastructural and molecular evidence. Bull Soc Zool Fr 120: 191-208
Moestrup 0 (1982) Flagellar structure in algae: a review, with new observations particularly on the Chrysophyceae, Phaeophyceae (Fucophyceae), Euglenophyceae, and Reckertia. Phycologia 21: 427-528
Moestrup 0 (1984) Further studies on Nephroselmisand its allies (Prasinophyceae); II. Mamiella gen. nov.,Mamiellaceae fam. nov., Mamiellales ord. nov. Nord JBot 4: 109-121
Moestrup 0 (1990) Scale structure in Mantoniella squamata, with some comments on the phylogeny of thePrasinophyceae (Chlorophyta). Phycologia 29: 437-442
Moestrup 0 (1991) Further studies of presumedly primitive green algae, including the description of Pedinophyceae Class.Nov. and Resultor Gen.Nov. J Phycol27: 119-133
Moestrup 0, Hori T (1989) Ultrastructure of the flagellar apparatus in Pyramimonas octopus (Prasinophyceae) II. Flagellar roots, connecting fibres, and numbering of individual flagella in green algae. Protoplasma148: 41-56
four different HPLC pigment signatures. J Phycol 24:571-580
Hori T, Inouye I, Horiguchi T, Boalch GT (1985) Observations on the motile stage of Halosphaera minorOstenfeld (Prasinophyceae) with special reference tothe cell structure. Bot Mar 28: 529-537
Huss VAR, Kranz HD (1997) Charophyte Evolution andthe Origin of Land Plants. In Bhattacharya 0 (ed) Originsof Algae and their Plastids. Springer Verlag, Wien, pp103-114
Huss VAR, Sogin ML (1990) Phylogenetic position ofsome Chlorella species within the Chlorococcalesbased upon complete small-subunit ribosomal RNA sequences. J Mol Evol 31: 432-442
Inouye I, Hori T, Chihara M (1990) Absolute configuration analysis of the flagellar apparatus of Pterospermacristatum (Prasinophyceae) and consideration of itsphylogenetic position. J Phycol 26: 329-344
Jones OTG (1963) Magnesium-2,4-divinylphaeoporphyrin as monomethylester, a protochlorophyll-like pigment produced by Rhodopseudomonas sphaeroides.Biochem J 89: 182-189
Kantz TS, Theriot EC, Zimmer EA, Chapman RL(1990) The Pleurastrophyceae and Micromonadophyceae: a cladistic analysis of nuclear rRNA sequencedata. J Phycol26: 711-721
Kimura M (1980) A simple method for estimating evolutionary rates of base substitution through comparativestudies of sequence evolution. J Mol Evol 16: 111-120
Kranz HD, Miks 0, Siegler M-L, Capesius I, SensenCW, Huss VAR (1995) The origin of land plants: phylogenetic relationships among charophytes, bryophytes,and vascular plants inferred from complete smail-subunit ribosomal RNA gene sequences. J Mol Evol 41:74-84
Leipe DO, Wainright PO, Gunderson JH, Porter 0,Patterson OJ, Valois F, Himmerich S, Sogin ML(1994) The stramenopiles from a molecular perspective:16S-like rRNA sequences from Labyrinthuloides minutaand Cafeteria roenbergensis. Phycologia 33: 369-377
Manton I (1965) Some phyletic implications of flagellarstructure in plants. In Preston RO (ed) Advances inBotanical Research, Vo1.2. Academic Press, pp 1-34
Manton I, Parke M (1960) Further Observations onSmall Green Flagellates with Special Reference to Possible Relatives of Chromulina pusilla Butcher. J Mar BioiAssoc UK 39: 275-298
Marin B, Melkonian M (1994) Flagellar hairs in prasinophytes (Chlorophyta): ultrastructure and distribution onthe flagellar surface. J Phycol 30: 659-678
Mattox KR, Stewart KD (1984) Classification of theGreen Algae: a Concept Based on Comparative Cytology. In Irvine OEG John OM (eds) Systematics of theGreen Algae. Academic Press, London, pp 29-72
Prasinophyte Ancestry of the Chlorophyta 379
McCourt RM (1995) Green algal phylogeny. TrendsEcol Evol 10: 159-163
Melkonian M (1982) Structural and evolutionary aspects of the flagellar apparatus in green algae and landplants. Taxon 31: 255-265
Melkonian M (1984) Flagellar Apparatus Ultrastructurein Relaton to Green Algal Classification. In Irvine OEGJohn OM (eds) Systematics of the Green Algae - Systematics Association Special Volumes, NO.27. Academic Press, London, pp 73-120
Melkonian M (1990a) Phylum Chlorophyta: ClassPrasinophyceae. In Margulis L, Corliss JO, MelkonianM, Chapman 0 (eds) Handbook of Protoctista. Jonesand Bartlett Publishers, Boston, pp 600-607
Melkonian M (1990b) Chlorophyte Orders of UncertainAffinities: Order Microthamniales. In Margulis L, CorlissJO, Melkonian M, Chapman OJ (eds) Handbook of Protoctista. Jones and Bartlett Publishers, Boston, pp652-654
Melkonian M (1996) Systematics and Evolution of theAlgae: Endocytobiosis and Evolution of the Major AlgalLineages. In Behnke H-O, LOttge U, Esser K, KadereitJW, Runge M (eds) Progress in Botany Vol 57. SpringerVerlag, Berlin, pp 281-311
Melkonian M, Marin B, Surek B (1995) Phylogeny andEvolution of the Algae. In Arai R, Kato M, Ooi Y (eds)Biodiversity and Evolution. The National Science Museum Foundation, Tokyo, pp 153-176
Melkonian M, Preisig HR (1986) A light and electronmicroscopic study of Scherffelia dubia, a new memberof the scaly green flagellates (Prasinophyceae). Nord JBot 6: 235-256
Melkonian M, Surek B (1995) Phylogeny of the Chlorophyta: congruence between ultrastructural and molecular evidence. Bull Soc Zool Fr 120: 191-208
Moestrup 0 (1982) Flagellar structure in algae: a review, with new observations particularly on the Chrysophyceae, Phaeophyceae (Fucophyceae), Euglenophyceae, and Reckertia. Phycologia 21: 427-528
Moestrup 0 (1984) Further studies on Nephroselmisand its allies (Prasinophyceae); II. Mamiella gen. nov.,Mamiellaceae fam. nov., Mamiellales ord. nov. Nord JBot 4: 109-121
Moestrup 0 (1990) Scale structure in Mantoniella squamata, with some comments on the phylogeny of thePrasinophyceae (Chlorophyta). Phycologia 29: 437-442
Moestrup 0 (1991) Further studies of presumedly primitive green algae, including the description of Pedinophyceae Class.Nov. and Resultor Gen.Nov. J Phycol27: 119-133
Moestrup 0, Hori T (1989) Ultrastructure of the flagellar apparatus in Pyramimonas octopus (Prasinophyceae) II. Flagellar roots, connecting fibres, and numbering of individual flagella in green algae. Protoplasma148: 41-56

380 T. Nakayama et al.
Moestrup 0, Throndsen J (1988) Light and electronmicroscopical studies on Pseudoscourfieldia marina, aprimitive scaly green flagellate (Prasinophyceae) withposterior flagella. Can J Bot 66: 1415-1434
Norris RE (1980) Prasinophytes. In Cox ER (ed)Phytoflagellates. Elsevier/North-Holland, New York,Amsterdam, Oxford, pp 85-145
O'Kelly CJ (1992) Flagellar apparatus architecture andthe phylogeny of "green" algae: chlorophytes, euglenoids, glaucophytes. In Menzel 0 (ed) The cytoskeletonof the algae. CRC Press, Boca Raton, Ann Arbor, London, Tokyo, pp 315-345
O'Kelly CJ, Floyd GL (1984) Flagellar apparatus absolute orientations and the phylogeny of the green algae.BioSystems 16: 227-252
Olsen GJ, Matsuda H, Hagstrom R, Overbeek R(1994) fastDNAml: A tool for construction of phylogenetic trees of DNA sequences using maximum likelihood. Comput Appl Biosci 10: 41-48
Ricketts TR (1970) The pigments of the Prasinophyceaeand related organisms. Phytochemistry 9: 1835-1842
Sambrook J, Fritsch EF, Maniatis T (1989) MolecularCloning: a Laboratory Manual. Cold Spring HarborLaboratory Press, Plainview NY
Saitou N, Nei M (1987) The neighbor-joining method: anew method for reconstructing phylogenetic trees. MolBioi Evol 4: 406-425
Schimek C, Stadnichuk IN, Knaust R, Wehrmeyer W(1994) Detection of chlorophyll c1 and magnesium-2,4-
divenylpheoporphyrin a5 monomethylester in cryptophytes. J Phycol30: 621-627
Sluiman HJ (1985) A cladistic evaluation of the lowerand higher green plants (Viridiplantae). Plant Syst Evol149: 217-232
Steinkotter J, Bhattacharya 0, Semmelroth I, BibeauC, Melkonian M (1994) Prasinophytes form independent lineages within the Chlorophyta: evidence from ribosomal RNA sequence comparisons. J Phycol 30:340-345
Stockner JG (1988) Phototrophic picoplankton: anoverview from marine and freshwater ecosystems. LimnoIOceanogr33:765-775
Surek B, Beemelmanns U, Melkonian M, Bhattacharya 0 (1994) Ribosomal RNA sequence comparisons demonstrate an evolutionary relationship between Zygnematales and charophytes. Plant Syst Evol191: 171-181
Swofford OL (1993) PAUP: phylogenetic analysis usingparsimony, version 3.1.1. Illinois Natural History Survey,Champaign
Sym SO, Pienaar RN (1993) The Class Prasinophyceae. In Round FE, Chapman OJ (eds) Progress inPhycological Research Vol 9. Biopress Ltd, Bristol, pp281-376
Throndsen J, Zingone A (1997) Dolichomastix tenuilepis sp. nov., a first insight into the microanatomy ofthe genus Dolichomastix (Mamiellales, Prasinophyceae,Chlorophyta). Phycologia 36: 244-254
380 T. Nakayama et al.
Moestrup 0, Throndsen J (1988) Light and electronmicroscopical studies on Pseudoscourfieldia marina, aprimitive scaly green flagellate (Prasinophyceae) withposterior flagella. Can J Bot 66: 1415-1434
Norris RE (1980) Prasinophytes. In Cox ER (ed)Phytoflagellates. Elsevier/North-Holland, New York,Amsterdam, Oxford, pp 85-145
O'Kelly CJ (1992) Flagellar apparatus architecture andthe phylogeny of "green" algae: chlorophytes, euglenoids, glaucophytes. In Menzel 0 (ed) The cytoskeletonof the algae. CRC Press, Boca Raton, Ann Arbor, London, Tokyo, pp 315-345
O'Kelly CJ, Floyd GL (1984) Flagellar apparatus absolute orientations and the phylogeny of the green algae.BioSystems 16: 227-252
Olsen GJ, Matsuda H, Hagstrom R, Overbeek R(1994) fastDNAml: A tool for construction of phylogenetic trees of DNA sequences using maximum likelihood. Comput Appl Biosci 10: 41-48
Ricketts TR (1970) The pigments of the Prasinophyceaeand related organisms. Phytochemistry 9: 1835-1842
Sambrook J, Fritsch EF, Maniatis T (1989) MolecularCloning: a Laboratory Manual. Cold Spring HarborLaboratory Press, Plainview NY
Saitou N, Nei M (1987) The neighbor-joining method: anew method for reconstructing phylogenetic trees. MolBioi Evol 4: 406-425
Schimek C, Stadnichuk IN, Knaust R, Wehrmeyer W(1994) Detection of chlorophyll c1 and magnesium-2,4-
divenylpheoporphyrin a5 monomethylester in cryptophytes. J Phycol30: 621-627
Sluiman HJ (1985) A cladistic evaluation of the lowerand higher green plants (Viridiplantae). Plant Syst Evol149: 217-232
Steinkotter J, Bhattacharya 0, Semmelroth I, BibeauC, Melkonian M (1994) Prasinophytes form independent lineages within the Chlorophyta: evidence from ribosomal RNA sequence comparisons. J Phycol 30:340-345
Stockner JG (1988) Phototrophic picoplankton: anoverview from marine and freshwater ecosystems. LimnoIOceanogr33:765-775
Surek B, Beemelmanns U, Melkonian M, Bhattacharya 0 (1994) Ribosomal RNA sequence comparisons demonstrate an evolutionary relationship between Zygnematales and charophytes. Plant Syst Evol191: 171-181
Swofford OL (1993) PAUP: phylogenetic analysis usingparsimony, version 3.1.1. Illinois Natural History Survey,Champaign
Sym SO, Pienaar RN (1993) The Class Prasinophyceae. In Round FE, Chapman OJ (eds) Progress inPhycological Research Vol 9. Biopress Ltd, Bristol, pp281-376
Throndsen J, Zingone A (1997) Dolichomastix tenuilepis sp. nov., a first insight into the microanatomy ofthe genus Dolichomastix (Mamiellales, Prasinophyceae,Chlorophyta). Phycologia 36: 244-254


















