

The adaptive Immune response - ruhr-uni-bochum.de · 28 Somatic hypermutation Affinity maturing of...
29
Transcript of The adaptive Immune response - ruhr-uni-bochum.de · 28 Somatic hypermutation Affinity maturing of...
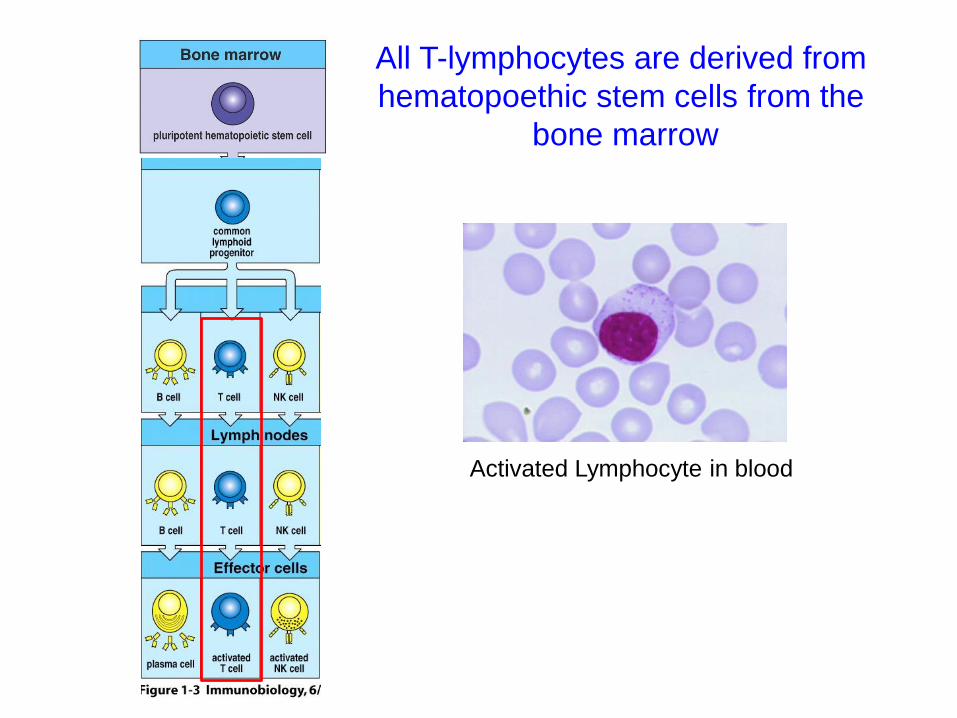
Activated Lymphocyte in blood
All T-lymphocytes are derived from
hematopoethic stem cells from the
bone marrow
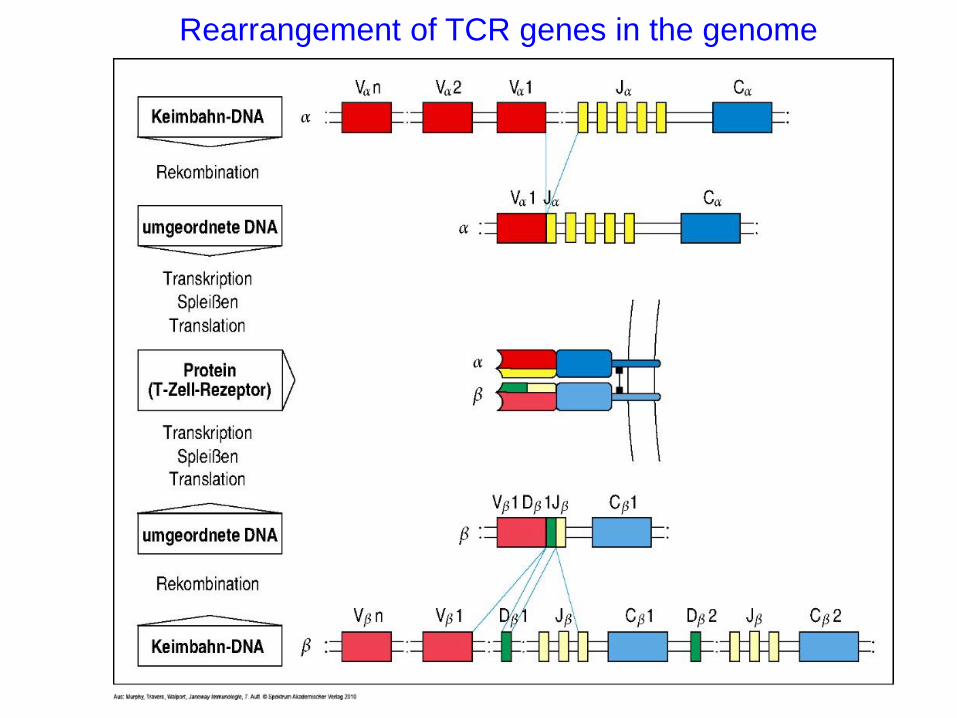
Rearrangement of TCR genes in the genome
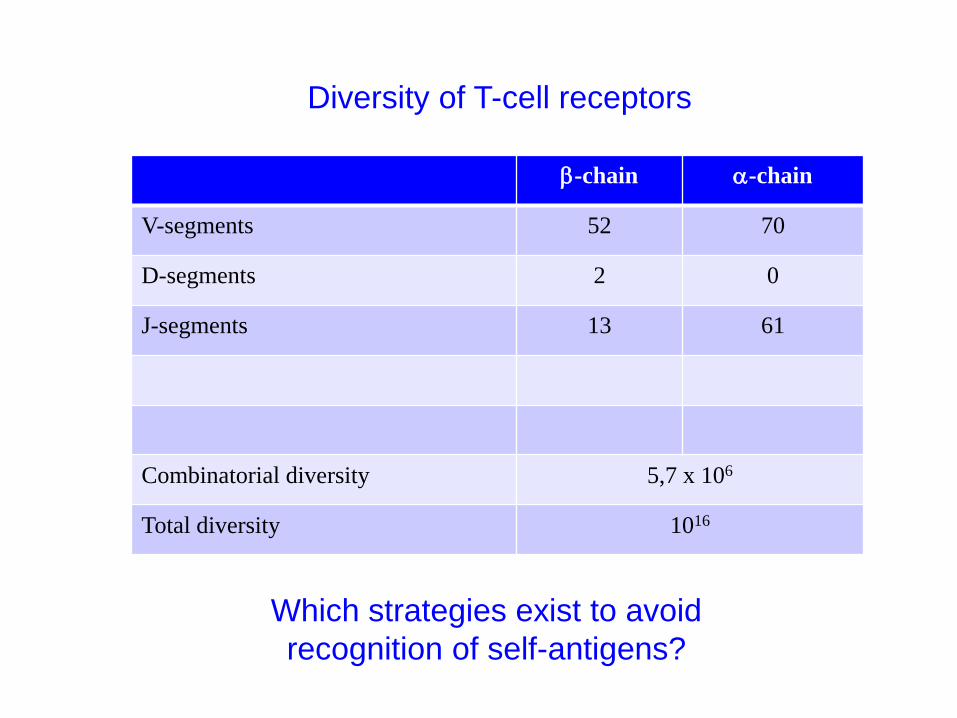
b-chain a-chain
V-segments 52 70
D-segments 2 0
J-segments 13 61
Combinatorial diversity 5,7 x 106
Total diversity 1016
Diversity of T-cell receptors
Which strategies exist to avoid
recognition of self-antigens?

Selection of T-lymphocytes in the Thymus---Central Tolerance
Negative selection of T-lymphocytes binding to self antigens with high affinity
Cells binding to self antigens die by apoptosis
Positive selection on ability of binding to MHCI and MHCII molecules
Cells with no affinity to MHC die by apoptosis
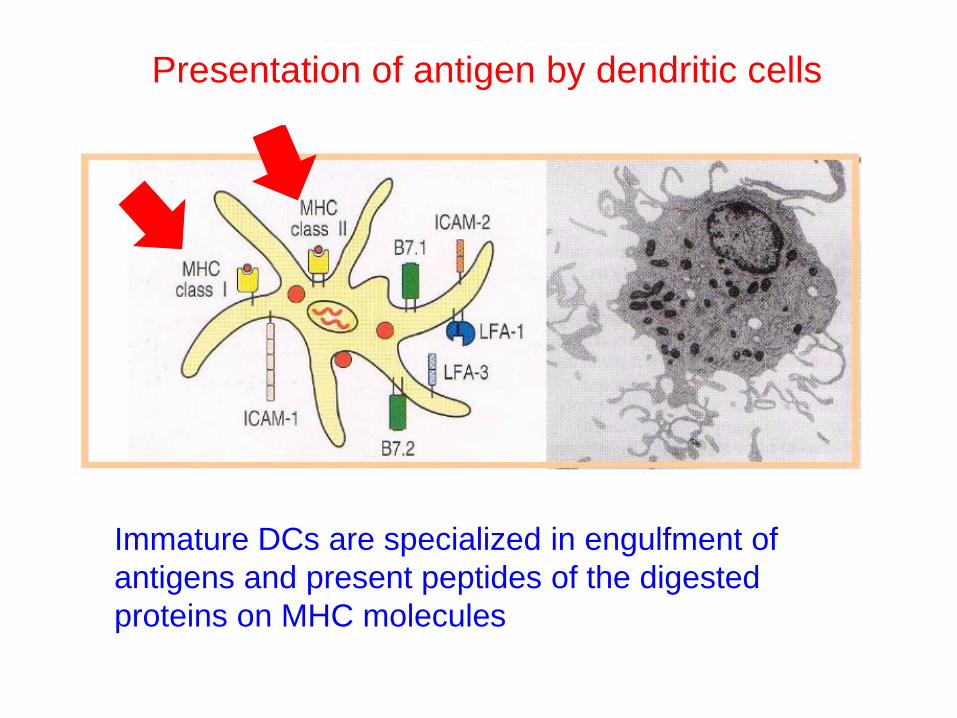
Immature DCs are specialized in engulfment of
antigens and present peptides of the digested
proteins on MHC molecules
Presentation of antigen by dendritic cells

MHC class I MHC class II
Expression by cell type all nucleated cells on antigen presenting
cells
Interaction with T-cell
subsets
CD8 cytotoxic T-cells CD4 T-helper cell
Molecular structure a-chain associated with
b2-microglobulin
a-chain associated with
b-chain
Peptides presented 8-10 amino acids larger than 13AA
Intracellular Location Endoplasmatic Reticulum Endosomes
Characteristics of MHC molecules
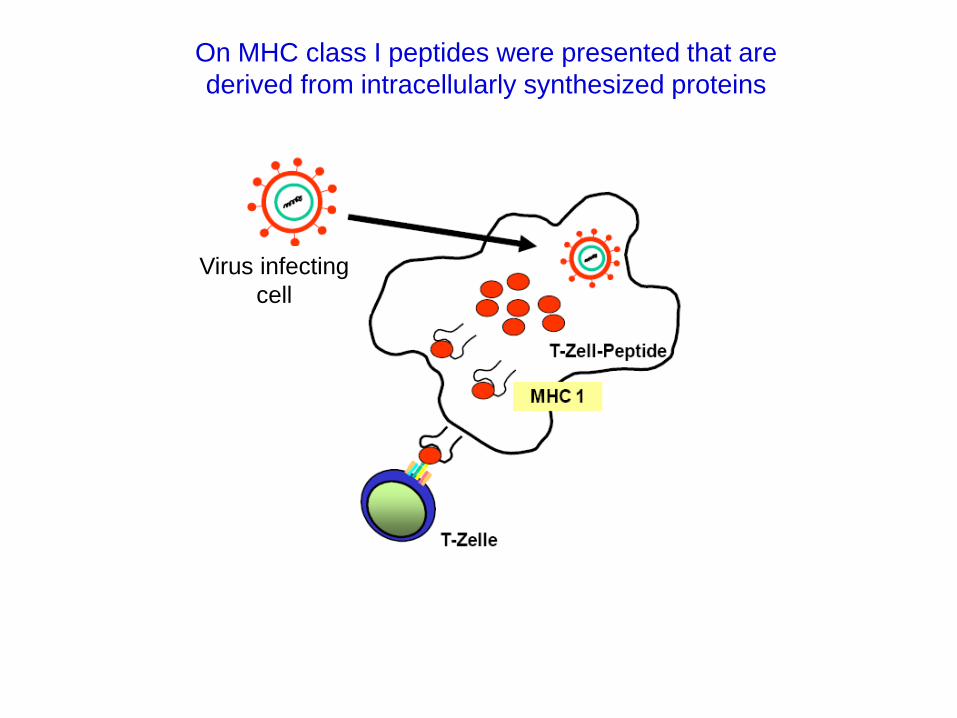
On MHC class I peptides were presented that are
derived from intracellularly synthesized proteins
Virus infecting
cell
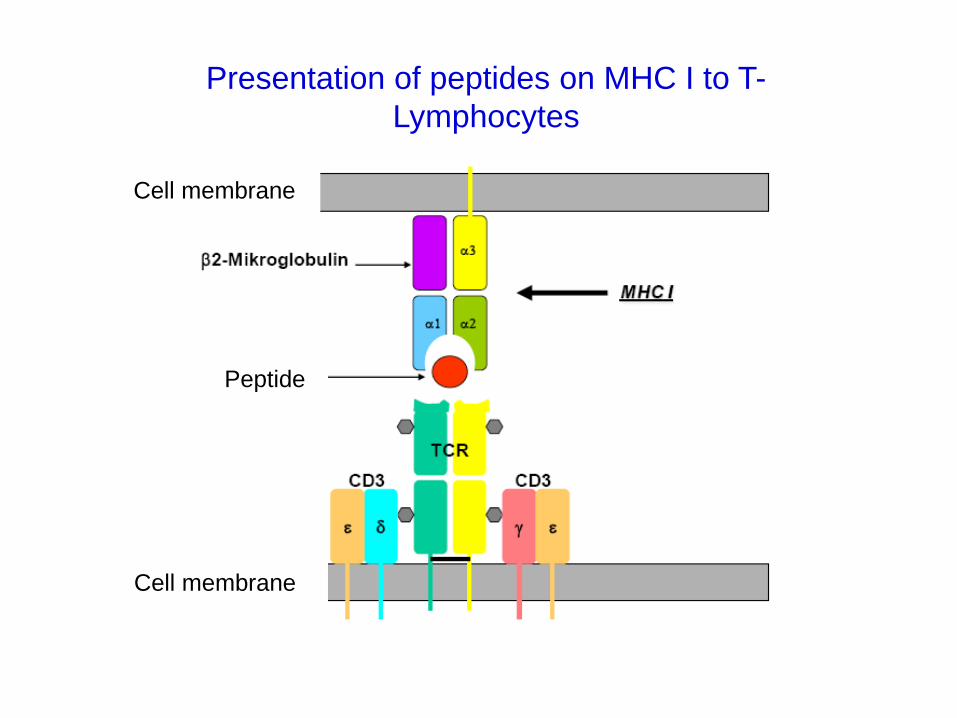
Presentation of peptides on MHC I to T-
Lymphocytes
Peptide
Cell membrane
Cell membrane
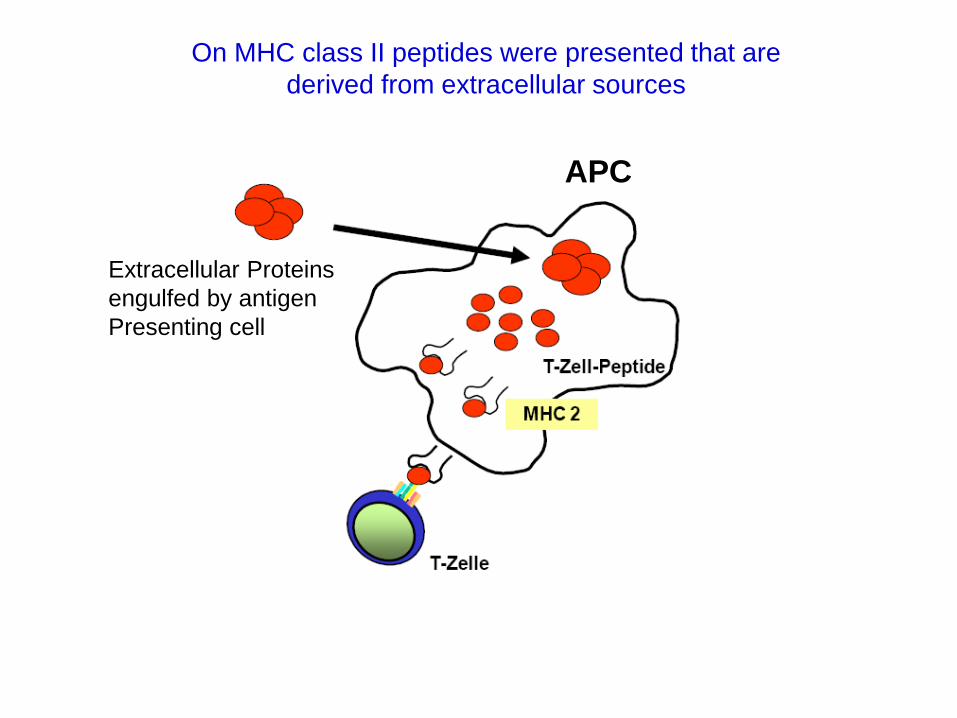
Extracellular Proteins
engulfed by antigen
Presenting cell
APC
On MHC class II peptides were presented that are
derived from extracellular sources
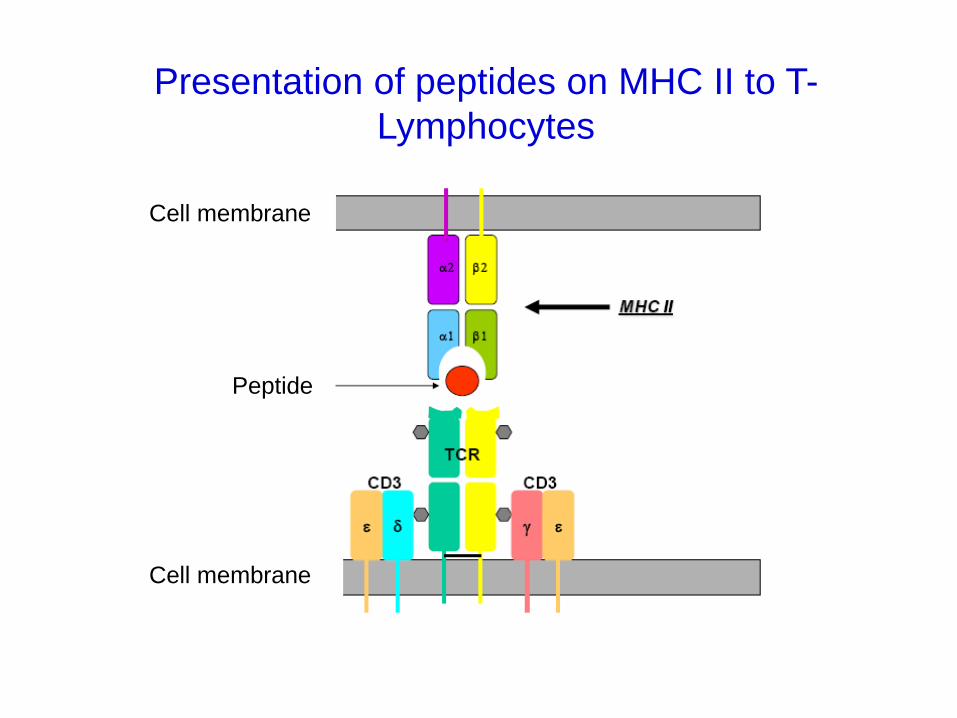
Presentation of peptides on MHC II to T-
Lymphocytes
Peptide
Cell membrane
Cell membrane
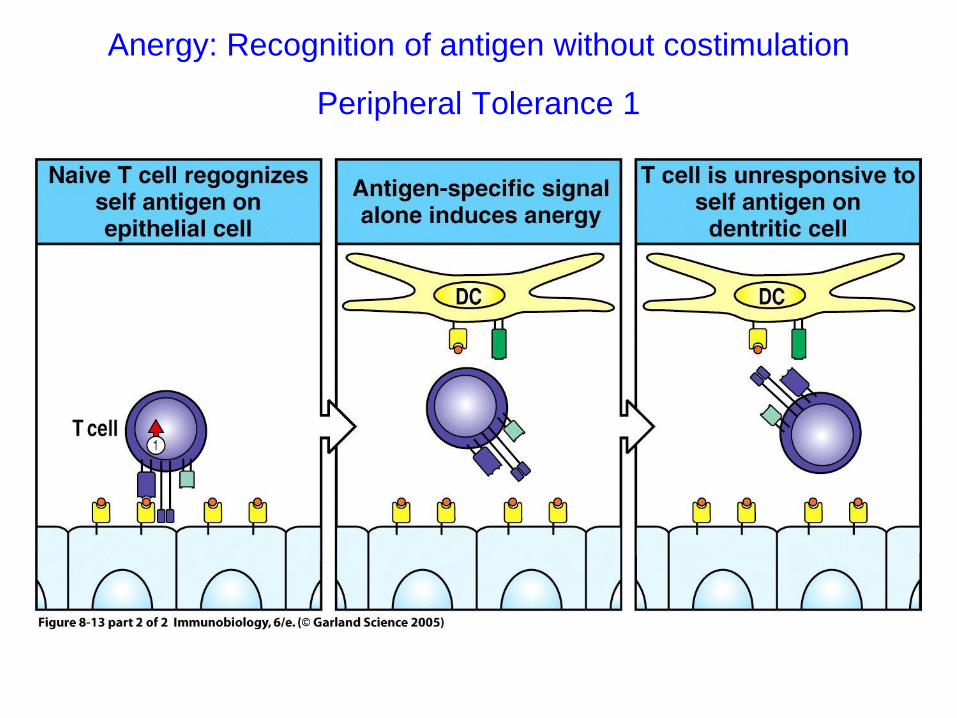
Anergy: Recognition of antigen without costimulation
Peripheral Tolerance 1
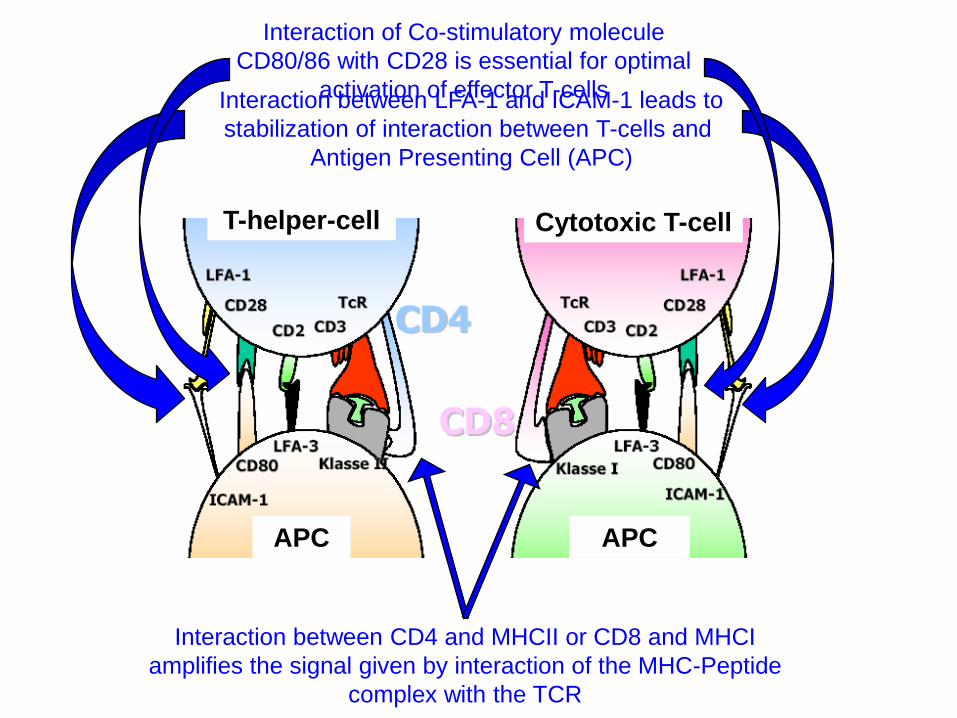
APC APC
T-helper-cell Cytotoxic T-cell
Interaction between LFA-1 and ICAM-1 leads to
stabilization of interaction between T-cells and
Antigen Presenting Cell (APC)
Interaction between CD4 and MHCII or CD8 and MHCI
amplifies the signal given by interaction of the MHC-Peptide
complex with the TCR
Interaction of Co-stimulatory molecule
CD80/86 with CD28 is essential for optimal
activation of effector T-cells
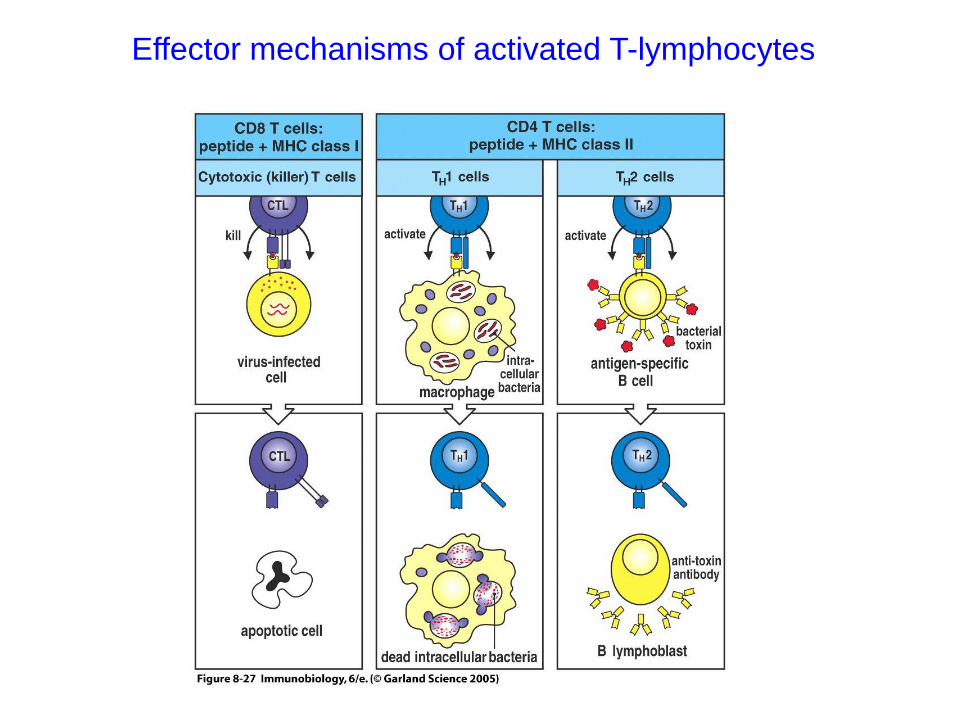
Effector mechanisms of activated T-lymphocytes

Th0
Th2
Th1
+
Mast/Baso
IL-4
DC
IL-12
+
IL-4
IFN-g
Differentiation of Th1 or Th2 lymphocytes
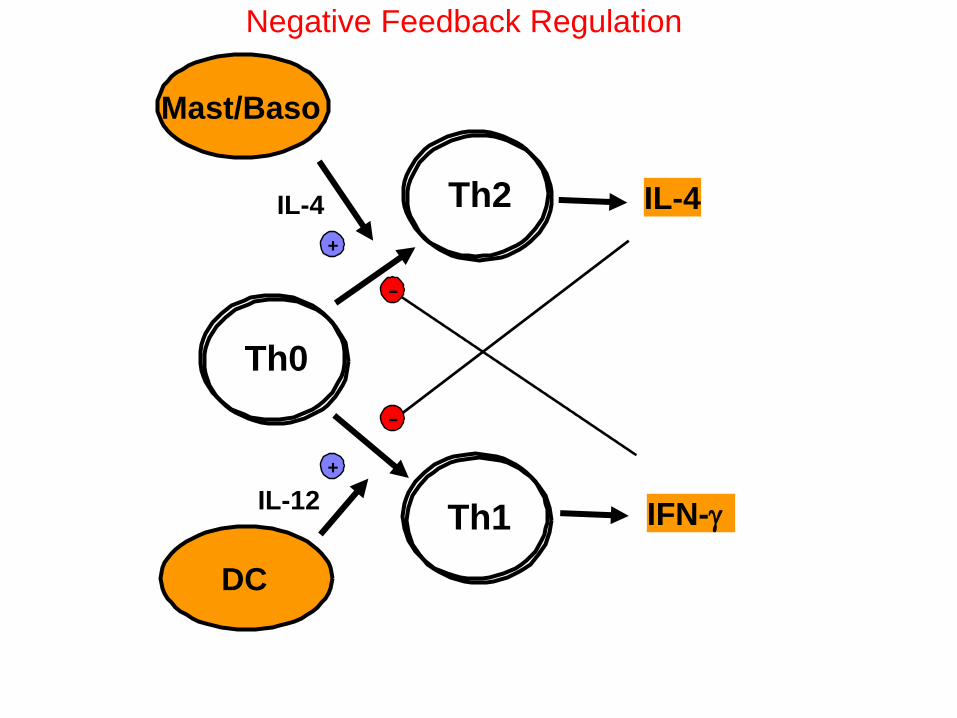
Th0
Th2
Th1
+
Mast/Baso
IL-4
DC
IL-12
+
IL-4
IFN-g
-
-
Negative Feedback Regulation
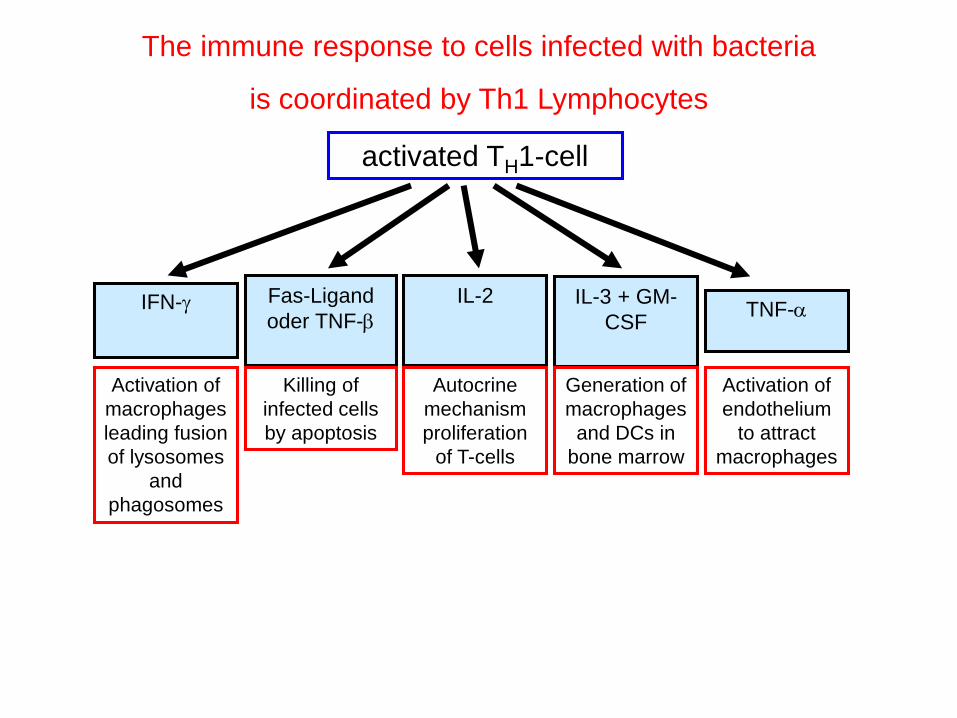
The immune response to cells infected with bacteria
is coordinated by Th1 Lymphocytes
activated TH1-cell
IFN-g
Activation of
macrophages
leading fusion
of lysosomes
and
phagosomes
Fas-Ligand
oder TNF-b
Killing of
infected cells
by apoptosis
IL-2
Autocrine
mechanism
proliferation
of T-cells
IL-3 + GM-
CSF
Generation of
macrophages
and DCs in
bone marrow
TNF-a
Activation of
endothelium
to attract
macrophages
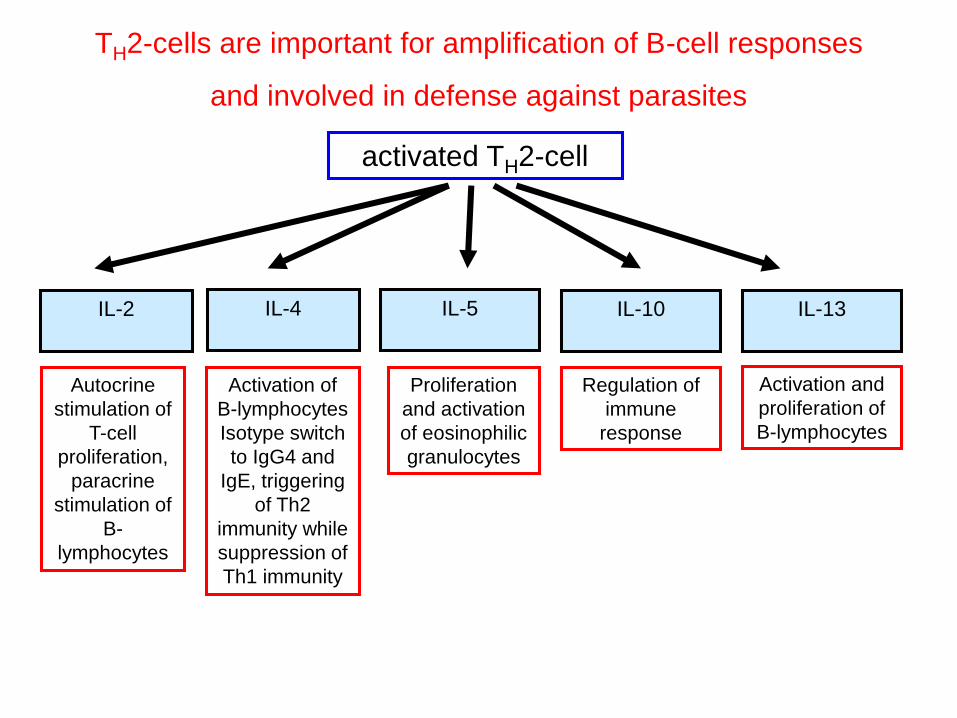
TH2-cells are important for amplification of B-cell responses
and involved in defense against parasites
activated TH2-cell
Autocrine
stimulation of
T-cell
proliferation,
paracrine
stimulation of
B-
lymphocytes
IL-4
Activation of
B-lymphocytes
Isotype switch
to IgG4 and
IgE, triggering
of Th2
immunity while
suppression of
Th1 immunity
Proliferation
and activation
of eosinophilic
granulocytes
Regulation of
immune
response
IL-2 IL-5 IL-10 IL-13
Activation and
proliferation of
B-lymphocytes

But in the last years several new
subpopulations of T-helper cells were
described….
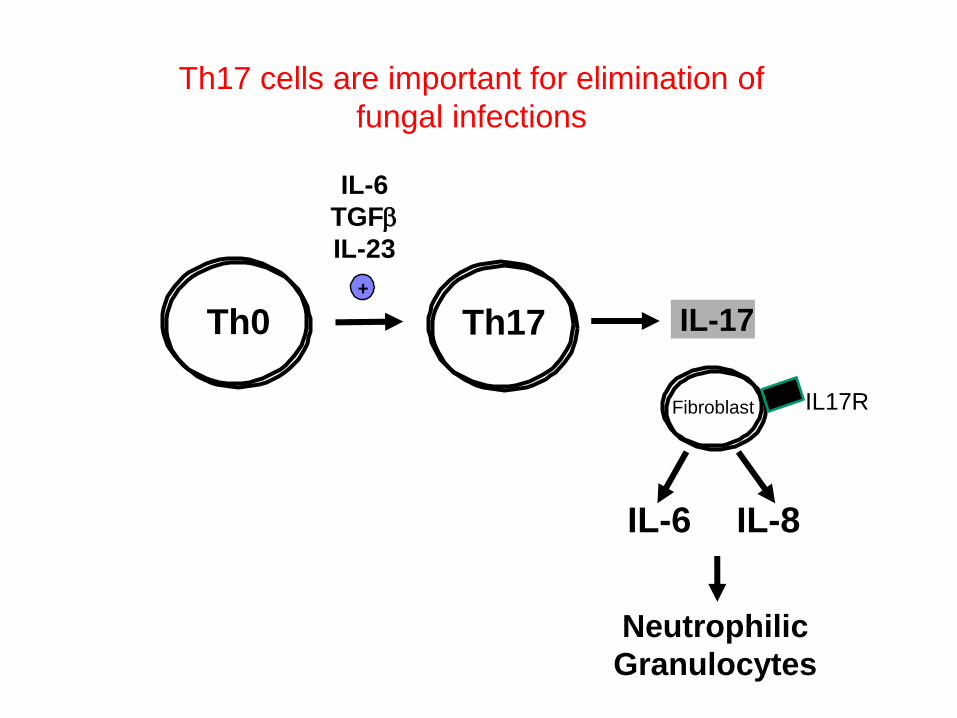
Th0 Th17 IL-17+
IL-6
TGFb
IL-23
IL-8IL-6
Neutrophilic
Granulocytes
Th17 cells are important for elimination of
fungal infections
Fibroblast IL17R
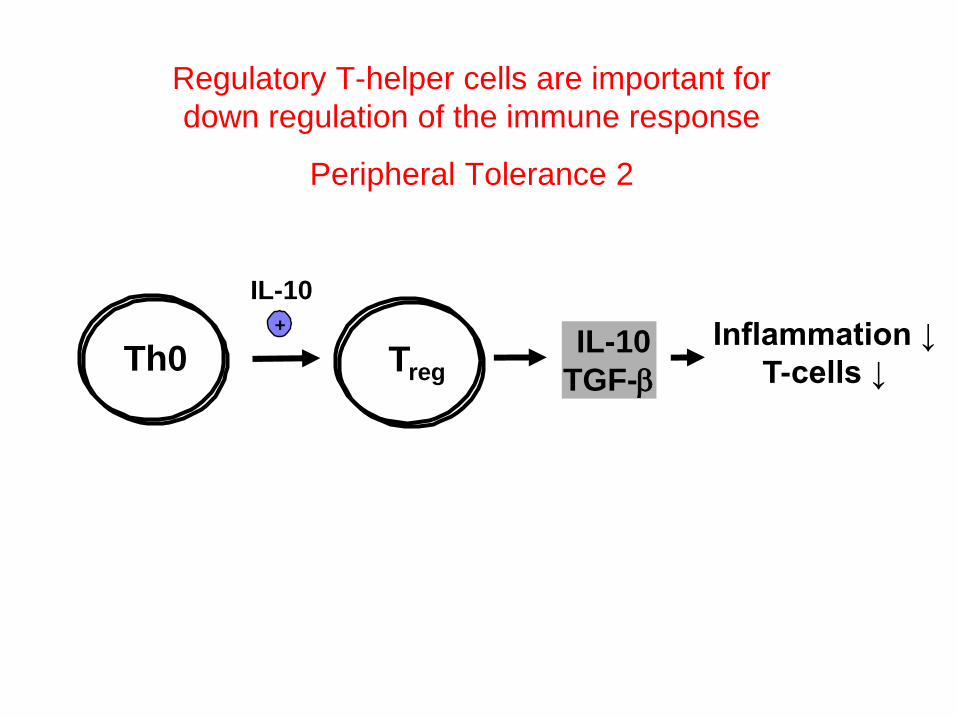
Th0 TregIL-10
TGF-b
+
IL-10
Regulatory T-helper cells are important for
down regulation of the immune response
Peripheral Tolerance 2
Inflammation ↓
T-cells ↓

O´Shea&Paul 2010. SCIENCE 327:1098
23
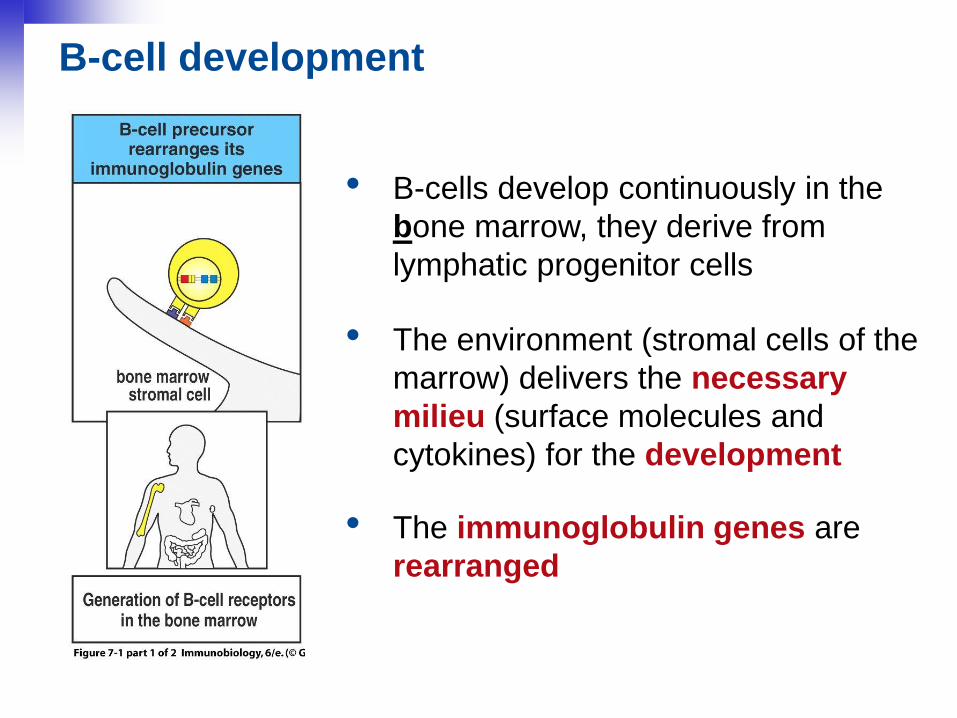
• B-cells develop continuously in the
bone marrow, they derive from
lymphatic progenitor cells
B-cell development
• The environment (stromal cells of the
marrow) delivers the necessary
milieu (surface molecules and
cytokines) for the development
• The immunoglobulin genes are
rearranged
24
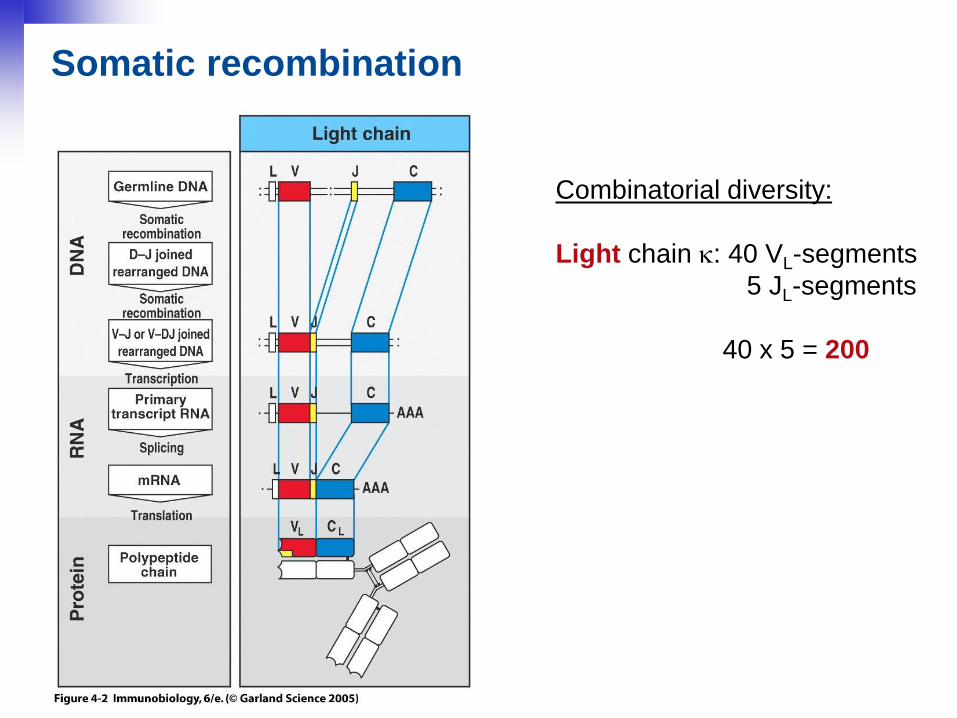
Somatic recombination
Combinatorial diversity:
Light chain : 40 VL-segments
5 JL-segments
40 x 5 = 200
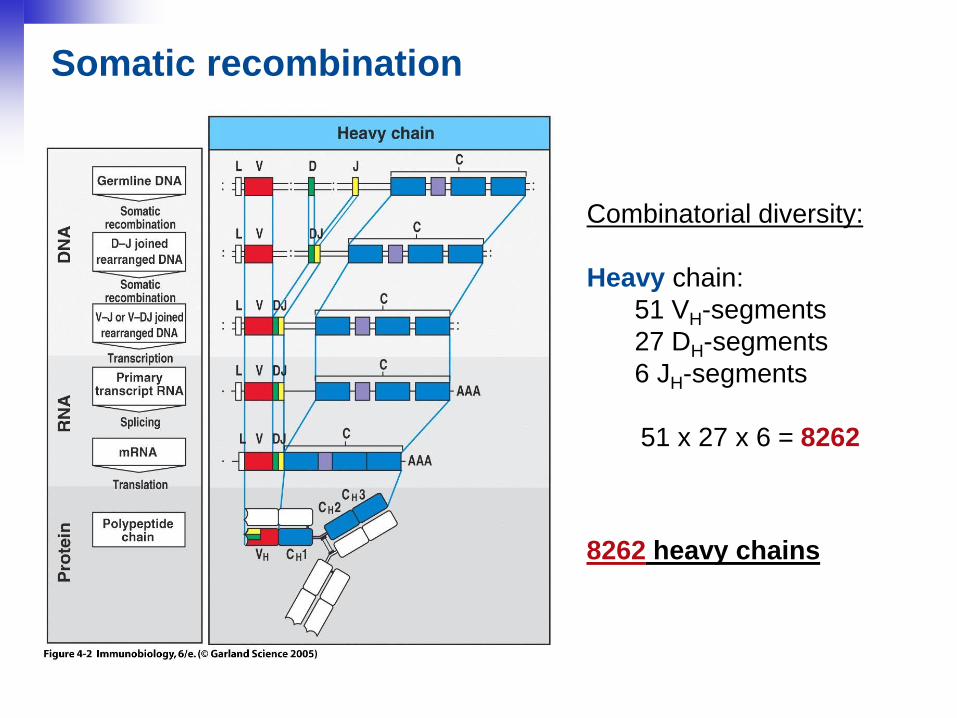
25
Combinatorial diversity:
Heavy chain:
51 VH-segments
27 DH-segments
6 JH-segments
51 x 27 x 6 = 8262
8262 heavy chains
Somatic recombination

26
200 light chains x 8.262 heavy chains = 1,6 x 106
Somatic recombination
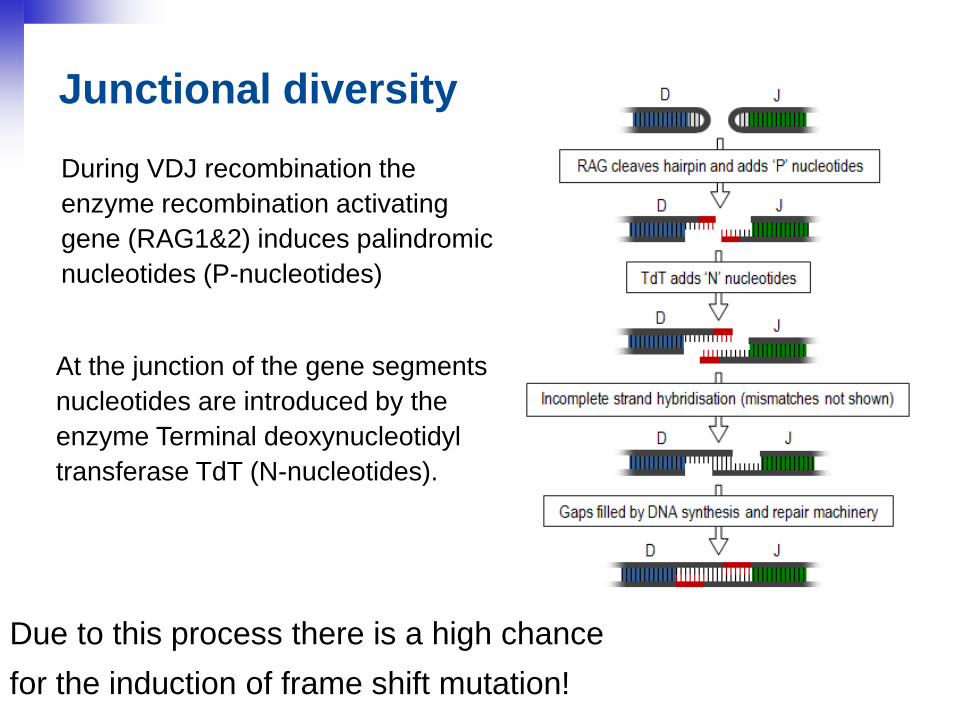
27
Junctional diversity
At the junction of the gene segments
nucleotides are introduced by the
enzyme Terminal deoxynucleotidyl
transferase TdT (N-nucleotides).
Due to this process there is a high chance
for the induction of frame shift mutation!
During VDJ recombination the
enzyme recombination activating
gene (RAG1&2) induces palindromic
nucleotides (P-nucleotides)

28
Somatic hypermutation
Affinity maturing of the antigen binding sites takes place in the
secondary lymphatic organs (only in activated B-cells).
Mutations occur in the whole variable region of the antibody.
Mutations appear in certain regions (hot spots).
The mechanism of the somatic hypermutation is mostly
unknown.
B cells become apoptotic when somatic hypermutation
resulted in reduced binding affinity of the antibody (during
affinity maturation)

29
Summary
Antibodies can be raised against almost every molecule
The antibody diversity is based on three different
mechanisms:
1) Combinatorial diversity
(Somatic recombination)
2) Junctional diversity
(Inaccurate junction)
3) Somatic hypermutation
(Affinity maturation in germinal centres)


















