The Accumulation of p53 Abnormalities Is Associated With ...
8
The Accumulation of p53 Abnormalities Is Associated With Progression of Mucosa-Associated Lymphoid Tissue Lymphoma By Mingqing Du, Huaizheng Peng, Nalini Singh, Peter G. Isaacson, and Langxing Pan The genetic mechanisms underlying the genesis of low- grade mucosa-associated lymphoid tissue (MALT) lympho- mas and their transformation into high-grade lymphoma are poorly understood. p53 inactivation, commonly caused by mutation and allele loss, has been shown to play an im- portant role in the early development andlor the late disease progression of many human tumors including lymphoid ma- lignancies and, thus, may also be important in MALT lymphomagenesis. We examined 75 cases (48 low grade and 27 high grade) of MALT lymphoma for p53 allele loss and mutation as well as protein accumulation. DNA samples pre- pared from microdissected cell populations were used for the detection of p53 gene abnormalities. Loss of heterozy- gosity (LOH) of the gene was detected by polymerase chain reaction-based analysis of p53 CA repeat polymorphism, whereas p53 mutation was studied by single-strand confor- mation polymorphism analysis and direct sequencing. p53 expression was assessed by immunostaining with CM1 ALIGNANT LYMPHOMA derived from mucosa-as- sociated lymphoid tissue (MALT) accounts for the majority of extranodal lymphomas and arises from acquired MALT in several extranodal sites such as the stomach, sali- vary gland, and thyr~id.”~ Low-grade B-cell lymphoma usu- ally exhibits an indolent clinical but is capable of transforming into high-grade tumor! Although the clinical, pathologic, and immunophenotypic features of MALT lymphoma have been well documented,”3 the genetic basis underlying its development and high-grade transformation is poorly understood. p53, a transcription factor, acts as a cell cycle check point proteinandinducescellcyclearrestinthelate G1 phase or apoptosis after DNA di~nage.5.~ p53 inactivation, abolishing its tumor-suppressor activity, is the most common event in human malipancie~~,~ and contributes to both early tumor develop- mentl0.” and late disease pro~ssion.’’ The inactivation of p53 tumor-suppressoractivitycan be causedatthegenelevelby mutation,I3 deletion,I4 and arrangement'^ or at the protein level by binding to oncogenic proteins such as MDM2.I6 Among these, p53 mutation and allelic loss are the most common. Inactivation of p53 tumor-suppressor activity during tu- mor development is a process of accumulation of its genetic abnormalitie~.~,’~ p53 mutation can precede allele loss or vice versa.” In both circumstances, p53 function is only partially inactivated because only one allele of the p53 gene is affected. Mutant p53 loses tumor-suppressor activity and may also exert a dominant negative effect on the remaining wild-type; however, it is unable completely to suppress wild- type function.19-*’ p53 inactivation involving both alleles, usually by mutation in one allele and loss of the other, results in the complete loss of p53 function. These different p53 abnormalities have been shown to exhibit distinct tumori- genic potentials in p53 transgenic mice with increasing tu- morigenicity after one allele loss, one allele mutation, and inactivation of both alleles.” These different p53 gene abnor- malities may also be expected to be involved at different stages of human tumor development. M Blood, Vol86, No 12 (December 15), 1995: pp 4587-4593 polyclonal antibody. p53 allele loss and mutation, which re- sulted in the alteration in the amino acid sequence, were found in both low-grade (LOH, 3 of 44 [6.89/01; mutation, 9 of 48 [18.8%1) and high-grade (LOH, 6 of 21 [28.6%3; mutation, 9 of 27 [33.3%1) MALT lymphomas, particularly in the latter group. p53 staining was not observed in any low-grade tu- mors but in 6 high-grade cases that harbored missense mu- tations. There were also differences in the extent of p53 abnormalities, between low- and high-grade tumors. Of the 11 low-grade tumors showing p53 abnormalities, only 1 tu- mor showed the concomitance of p53 mutation and allele loss, whereas in high-grade tumors, 6 of 9 affected cases displayed both p53 mutation and allele loss. Our results sug- gest that p53 partial inactivation may play an important role in the development of low-grade MALT lymphomas, whereas complete inactivation may be associated with high- grade transformation. 0 1995 by The American Society of Hematology. Various frequencies of p53 mutation have been docu- mented in lymphoid malignan~ies.‘~-~~ p53 mutation is thought to be associated with high-grade transformation of follicular lymphoma” and chronic lymphocytic leukemia.” Recently, a high frequency of p53 mutation has been re- ported in marginal zone splenic B-cell lymphoma,’8 which is analogous to MALT lymphoma in its origin of cell lineage, immunophenotyping, histology, and genetic findings such as the absence of bcl-2 and bcl-l rear~-angement.’~.’~ Thus, inactivation of the p53 gene may also be important in the tumorigenesis of MALT lymphoma. Most previous p53 studies have failed to pay full attention to the analysis of both allele loss and mutation together in the same tumor g r o ~ p , ” ~ ’ ~ and thus the full potential of p53 inactivation in tumorigenesis may have been underestimated. In the present study, we examined p53 gene abnormalities, including both allele loss andmutation as well as protein accumulation in 75 cases of well-characterized low- and high-grade MALT lymphoma to determine the full potential of p53 inactivation in MALT lymphomagenesis. Our results suggest that p53 partial inactivation may be associated with the development of low-grade tumor, whereas complete in- activation may be important for high-grade transforma- tion. From the Department of Histopathology, University College Lon- Submitted June 7, 1995; accepted August 7, 1995. Address reprint requests to Peter G. Isaacson, DM, FRC Path, Department of Histopathology, University College London Medical School, Rockefeller Building, University Street, London WCIE SJJ, UK. The publication costsof this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1734 solely to indicate this fact. don Medical School, London, UK. 0 1995 by The American Society of Hematology. 0006-4971/95/86/2-0035$3.00/0 4587 For personal use only. on March 31, 2018. by guest www.bloodjournal.org From
Transcript of The Accumulation of p53 Abnormalities Is Associated With ...

The Accumulation of p53 Abnormalities Is Associated With Progression of Mucosa-Associated Lymphoid Tissue Lymphoma
By Mingqing Du, Huaizheng Peng, Nalini Singh, Peter G. Isaacson, and Langxing Pan
The genetic mechanisms underlying the genesis of low- grade mucosa-associated lymphoid tissue (MALT) lympho- mas and their transformation into high-grade lymphoma are poorly understood. p53 inactivation, commonly caused by mutation and allele loss, has been shown to play an im- portant role in the early development andlor the late disease progression of many human tumors including lymphoid ma- lignancies and, thus, may also be important in MALT lymphomagenesis. We examined 75 cases (48 low grade and 27 high grade) of MALT lymphoma for p53 allele loss and mutation as well as protein accumulation. DNA samples pre- pared from microdissected cell populations were used for the detection of p53 gene abnormalities. Loss of heterozy- gosity (LOH) of the gene was detected by polymerase chain reaction-based analysis of p53 CA repeat polymorphism, whereas p53 mutation was studied by single-strand confor- mation polymorphism analysis and direct sequencing. p53 expression was assessed by immunostaining with CM1
ALIGNANT LYMPHOMA derived from mucosa-as- sociated lymphoid tissue (MALT) accounts for the
majority of extranodal lymphomas and arises from acquired MALT in several extranodal sites such as the stomach, sali- vary gland, and thyr~id .”~ Low-grade B-cell lymphoma usu- ally exhibits an indolent clinical but is capable of transforming into high-grade tumor! Although the clinical, pathologic, and immunophenotypic features of MALT lymphoma have been well documented,”3 the genetic basis underlying its development and high-grade transformation is poorly understood.
p53, a transcription factor, acts as a cell cycle check point protein and induces cell cycle arrest in the late G1 phase or apoptosis after DNA di~nage.5.~ p53 inactivation, abolishing its tumor-suppressor activity, is the most common event in human mal ipanc ie~~,~ and contributes to both early tumor develop- mentl0.” and late disease pro~ssion.’’ The inactivation of p53 tumor-suppressor activity can be caused at the gene level by mutation,I3 deletion,I4 and arrangement'^ or at the protein level by binding to oncogenic proteins such as MDM2.I6 Among these, p53 mutation and allelic loss are the most common.
Inactivation of p53 tumor-suppressor activity during tu- mor development is a process of accumulation of its genetic abnormalitie~.~,’~ p53 mutation can precede allele loss or vice versa.” In both circumstances, p53 function is only partially inactivated because only one allele of the p53 gene is affected. Mutant p53 loses tumor-suppressor activity and may also exert a dominant negative effect on the remaining wild-type; however, it is unable completely to suppress wild- type function.19-*’ p53 inactivation involving both alleles, usually by mutation in one allele and loss of the other, results in the complete loss of p53 function. These different p53 abnormalities have been shown to exhibit distinct tumori- genic potentials in p53 transgenic mice with increasing tu- morigenicity after one allele loss, one allele mutation, and inactivation of both alleles.” These different p53 gene abnor- malities may also be expected to be involved at different stages of human tumor development.
M
Blood, Vol86, No 12 (December 15), 1995: pp 4587-4593
polyclonal antibody. p53 allele loss and mutation, which re- sulted in the alteration in the amino acid sequence, were found in both low-grade (LOH, 3 of 44 [6.89/01; mutation, 9 of 48 [18.8%1) and high-grade (LOH, 6 of 21 [28.6%3; mutation, 9 of 27 [33.3%1) MALT lymphomas, particularly in the latter group. p53 staining was not observed in any low-grade tu- mors but in 6 high-grade cases that harbored missense mu- tations. There were also differences in the extent of p53 abnormalities, between low- and high-grade tumors. Of the 11 low-grade tumors showing p53 abnormalities, only 1 tu- mor showed the concomitance of p53 mutation and allele loss, whereas in high-grade tumors, 6 of 9 affected cases displayed both p53 mutation and allele loss. Our results sug- gest that p53 partial inactivation may play an important role in the development of low-grade MALT lymphomas, whereas complete inactivation may be associated with high- grade transformation. 0 1995 by The American Society of Hematology.
Various frequencies of p53 mutation have been docu- mented in lymphoid malignan~ies.‘~-~~ p53 mutation is thought to be associated with high-grade transformation of follicular lymphoma” and chronic lymphocytic leukemia.” Recently, a high frequency of p53 mutation has been re- ported in marginal zone splenic B-cell lymphoma,’8 which is analogous to MALT lymphoma in its origin of cell lineage, immunophenotyping, histology, and genetic findings such as the absence of bcl-2 and bcl-l rear~-angement.’~.’~ Thus, inactivation of the p53 gene may also be important in the tumorigenesis of MALT lymphoma.
Most previous p53 studies have failed to pay full attention to the analysis of both allele loss and mutation together in the same tumor g r o ~ p , ” ~ ’ ~ and thus the full potential of p53 inactivation in tumorigenesis may have been underestimated. In the present study, we examined p53 gene abnormalities, including both allele loss and mutation as well as protein accumulation in 75 cases of well-characterized low- and high-grade MALT lymphoma to determine the full potential of p53 inactivation in MALT lymphomagenesis. Our results suggest that p53 partial inactivation may be associated with the development of low-grade tumor, whereas complete in- activation may be important for high-grade transforma- tion.
From the Department of Histopathology, University College Lon-
Submitted June 7, 1995; accepted August 7, 1995. Address reprint requests to Peter G. Isaacson, DM, FRC Path,
Department of Histopathology, University College London Medical School, Rockefeller Building, University Street, London WCIE SJJ, UK.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. section 1734 solely to indicate this fact.
don Medical School, London, UK.
0 1995 by The American Society of Hematology. 0006-4971/95/86/2-0035$3.00/0
4587
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom

4588 DU ET AL
MATERIALS AND METHODS
Tissue samples. Seventy-five cases (paraffin-embedded tissues available in 63, frozen tissues available in 24) of B-cell MALT lymphoma were retrieved from the Department of Histopathology, University College London Medical School. These comprised 48 low-grade and 27 high-grade MALT lymphomas. Tumors originated from the stomach in 60 cases, salivary gland in 6 cases, small intes- tine in 5 cases, and thyroid in 4 cases. The diagnosis of low-grade MALT lymphoma was established by the characteristic histologic appearances’” together with immunophenotyping and clonal analysis of the Ig heavy chain gene. Of the 27 high-grade lymphomas, a low-grade MALT lymphoma component could be identified in 21, usually as one or more small foci present in paraffin sections from a minority of the blocks.
Microdissection and DNA preparation. Paraffin (28 low-grade cases and 23 high-grade cases) and frozen (20 low-grade cases and 4 high-grade cases) sections were briefly stained with hematoxylin and eosin. Tumor and nontumor cell populations were separately microdissected from each case, as described previously.” The mi- crodissected cell population was digested with 200 &mL proteinase K in a 40 to 1 0 0 pL solution containing 10 mmol/L Tris HCI (pH 8.3). 50 mmol/L KC1 at 37°C for 16 hours. Proteinase K was inacti- vated at 95°C for I O minutes. Cell debris was pelleted by centrifuga- tion and the resulted supernatant was used for the subsequent genetic analysis of the p53 gene.
Detection of loss of heterozygosit?, (LOH) of the p53 gene. LOH of the p53 gene was detected by polymerase chain reaction (PCR)- based analysis of the p53 dinucleotide repeat (CA25) polymor- phism.” The CA repeat was PCR-amplificd from both tumor and nontumor DNA preparations of the same patient using primers: GT strand, AGGGATACTATTCAGCCCGAGGTG, and AC strand, ACTGCCACTCCTTGCCCCATTC (Oswell DNA Service, Depart- ment of Chemistry, Edinburgh University, Edinburgh, UK). One of the primers was end-labeled using [y-”P] and T4 polynucleotide kinase. PCR products (2 yL) were mixed with 4 yL sequence stop solution (98% formamide, I O mmol/L NaOH, 20 mmol/L EDTA, 0.05% bromophenol blue, and 0.05% xylene cyano1 FF), and the mixtures were denatured at 98°C for 5 minutes and separated on 6% sequencing gel. The gel was routinely dried and exposed to an x- ray film for I to 2 days.
Single-strand conformation polymorphism (SSCP) ana1.vsi.r. Be- cause the majority of p53 mutations occur in exon 5, 6, 7, and 8 of the gene,’.’ these exons were separately amplified by PCR using primers described previously.” SSCP analysis of these PCR products was performed using our recently described protocol, which used polyacrylamide agarose composite gel together with a background free silver staining method and showed a high sensitivity (97%) for p53 mutation detection.” Briefly, PCR products (2 to 5 yL) were mixed with sequence stop solution. The mixtures were denatured at 98°C for 5 minutes, cooled on ice, and loaded onto a composite gel containing 5% acrylamide (acry1amide:bis-acrylamide = 99:l) and 0.25% agarose with or without 10% glycerol. Electrophoresis was performed at 6 W for 12 to 18 hours at 4°C maintained by a refriger- ated circulator (Pharmacia, Uppsala, Sweden), and gel was subjected to silver staining.
Direct DNA sequencing. Samples for which mutation was sug- gested by SSCP were further analyzed by direct DNA sequencing using fmol DNA sequencing system (Promega, Southampton, UK).
Immunostaining of p53 protein. Immunostaining for p53 protein was performed in cases with paraffin-embedded tissues, which were composed of 37 low-grade and 26 high-grade tumours. For antigen retrieval, paraffin sections were treated in 0.01 moln sodium citrate buffer (pH 6.0) in a pressure cooker for 2 minutes before staining.25
G0015 G0078 N T N T
Fig 1. Detection of LOH of the p53 gene in MALT lymphomas. p53 CA repeat was amplified by PCR, in which one of the primer was end-labeled (see the Materials and Methods). The PCR products were denatured and separated on 6% sequence gel. N and T indicate non- tumor and tumor samples from the same patient. Case G0078 showed loss of one p53 allele in the tumor sample.
p53 protein was routinely immunostained using CM1 polyclonal antibody (I/5,000 dilution) for 16 hours at room temperature, fol- lowed by Biotinylated Swine antirabbit Ig (11250 dilution) for 40 minutes and StreptABComplexRIRP (111 00 dilution) for 40 minutes. Finally, sections were visualized by incubation with diaminobenzi- dine (DAB) and HzOz. All reagents used for the staining were pur- chased from Dako (High Wycombe, UK). For each experiment, positive (a breast carcinoma known to express mutant p53 protein) and negative controls (without the primary antibody) were included.
Immunostaining for p53 using more concentrated CM1 antibody (either U2.500 dilution for 16 hours or 111,OOO dilution for 1 hour) was additionally performed in some of the MALT lymphomas, which showed negative staining for p53 when CM1 antibody was used at 115.000 dilution.
RESULTS
LOH qf the p53 gene in MALT lymphoma. Of the 15 MALT lymphomas studied, nontumor DNA preparations were available from 74 cases. Among these cases, 65 (87.8%) were heterozygous for the p53 CA repeat and thus were informative for LOH analysis. Because the analysis was performed from DNA samples prepared from microdissected cell populations, this largely eliminated the interference from nontumor cells and LOH was easily determined by a direct comparison of the allelic density between tumor and nontu- mor samples of the same patient (Fig l ) . In total, LOH of the p53 gene was found in 9 (13.8%) cases, with a much higher frequency in high-grade (6/21 [28.6%]) than in low- grade (3/44 [6.8%]) tumors.
Mutation of the p53 gene in MALT lymphoma. Using PCR-SSCP analysis, mutations suggested by the altered elec- trophoretic mobility were identified in 21 cases and these
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom
p53 INACTIVATION IN MALT LYMPHOMA 4589
1 2
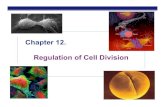
Fig 2. Detection of p53 mutation in MALT lymphomas by PCR-SSCP analysis. PCR products of p53 exon 6 were denatured, separated on polyacryl- amide-agarose gel, and shown by silver staining (see the Materials and Methods). Lane 1 and 10, placenta DNA as negative controls; lanes 2 through 8, individ- ual MALT samples; lane 9, A3/kawa cell line con- taining a missense mutation in exon 6'' used as a positive control. Lanes 4 (case G00151 and 6 (case 60114) showed migrating fragments different from the normal control.
3 4 5 6 7 8 9 1 0
mutations occurred in exon S in 7 cases, exon 6 in 8 cases. exon 7 in 4 cases, and exon 8 in 2 cases. Examples are shown in Fig 2.
To confirm the results obtained by SSCP analysis and to characterize the nature of these mutations, SSCP-positive cases were further analyzed by direct sequencing. The results are presented in Table 1 and an example is shown in Fig 3. Mutation was confirmed in all cases. In 18 cases, mutations lead to alterations in amino acid sequence, most frequently amino acid substitution caused by missense mutations (point mutation in 14 cases; tandem double mutations in 1 case) and less frequently truncated protein due to frame shift muta- tion (l-bp deletion in 2 cases) and nonsense mutation ( I
case). Mutations in the remaining three cases were silent and did not alter the amino acid sequence. The mutations identified showed a wide spectrum with a similar frequency of transition (10/20) and transversion (10/20) mutations. Four of 20 mutations, including 2 C+T, l G"A transitions and I G"T transversion, occurred at CpG dinucleotides. However, no predominant type of mutation was seen.
In summary, inactivating p53 mutation, which alters amino acid sequence, was found in l8 of 75 (24%) MALT lymphomas. Similar to LOH of the p53 gene, mutation is more frequent in high-grade (9/27 [33.3%]) than in low- grade (9148 [ 18.8%]) tumors.
Accunrulntiorl of p53 protein it1 MALT iwlpkotncr. Of
Table 1. Summary of MALT Lymphoma Cases Showing p53 Abnormalities
Case Malignant Grade Tumor Site p53 Staining LOH SSCP Codon Mutation Amino Acid Substitution
G0015 LG st - - Exon 6+ 193 CAT - CAC His - His G0318 LG st ND - Exon 6+ 197 GTG - GTA Val - Val G0078 LG st - + -
G0178 LG st ND + - G2098 LG s t - - Exon 6 + 196 CGA - TGA Arg - End
G0123 LG s t - - Exon 5+ 178 CAC - *AC Frameshift G0062 LG st - - Exon 5+ 160 ATG - CTG Met - Leu G0177 LG st - - Exon 6+ 197 GTG - ATG Val - Met
G0208 LG st - - Exon 5+ 165 CAG - CTG Gln - Leu G021 1 LG s t - - Exon 8+ 273 CGT + TGT Arg - Cys
G1181 LG SG - - Exon 5+ 165 CAG - CCG Gln - Pro
G2099 LG st - - Exon 5+ 165 CAG + CCG Gln - Pro G0525 LG Thy - + Exon 7+ 243 ATG - GTG Met - Val
G1201 HG s t - - Exon 7+ 253 G1200 HG s t - - Exon 6+ 206 G0168 HG s t - - Exon 6+ 194 G01 14 HG ST + - Exon 6+ 212 G0210 HG st - + Exon 5+ 158 G 1207 HG st + + Exon 8+ 274 G0016 HG s t + + Exon 5+ 151 G0399 HG s t + + Exon 6+ 192 G1918 HG st + + Exon 7+ 248 G 1205 HG SI + + Exon 7+ 254
Abbreviations: St, stomach; SG, salivary gland; Thy, thyroid; SI, small intestine; ND, not done. * Deletion.
ACC - ACT l T G - T*G CTT -CAT " r - l T A CGC - CTC GTT - GAT CCC - CAC CAG - CGG CGG - CAG ATC - GAC
Thr - Thr Frameshift Leu - His Phe - Leu Arg - Leu Val - Asp Pro - His Gln - Arg Arg - Gln Ile -Asp
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom
4590 DU ET AL
GO016 ‘T
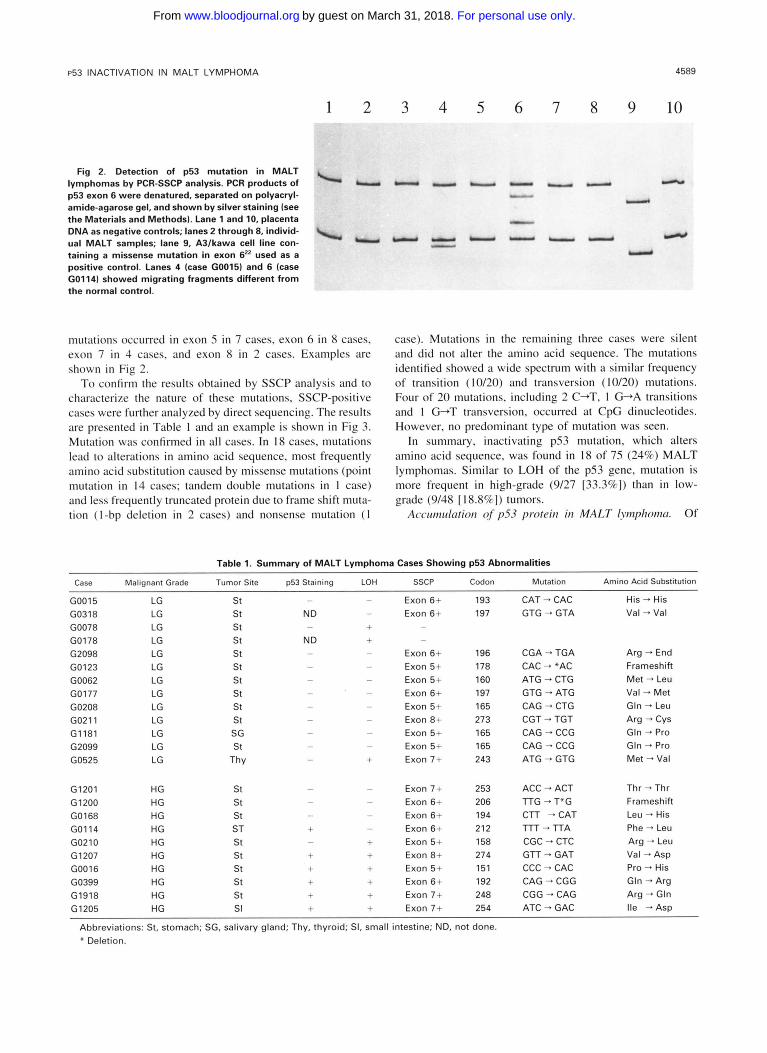
Fig 3. Direct sequencing of the PCR product from SSCP-positive cases. Case G0016 showed a missense mutation (CCC + CAC) in codon 151.
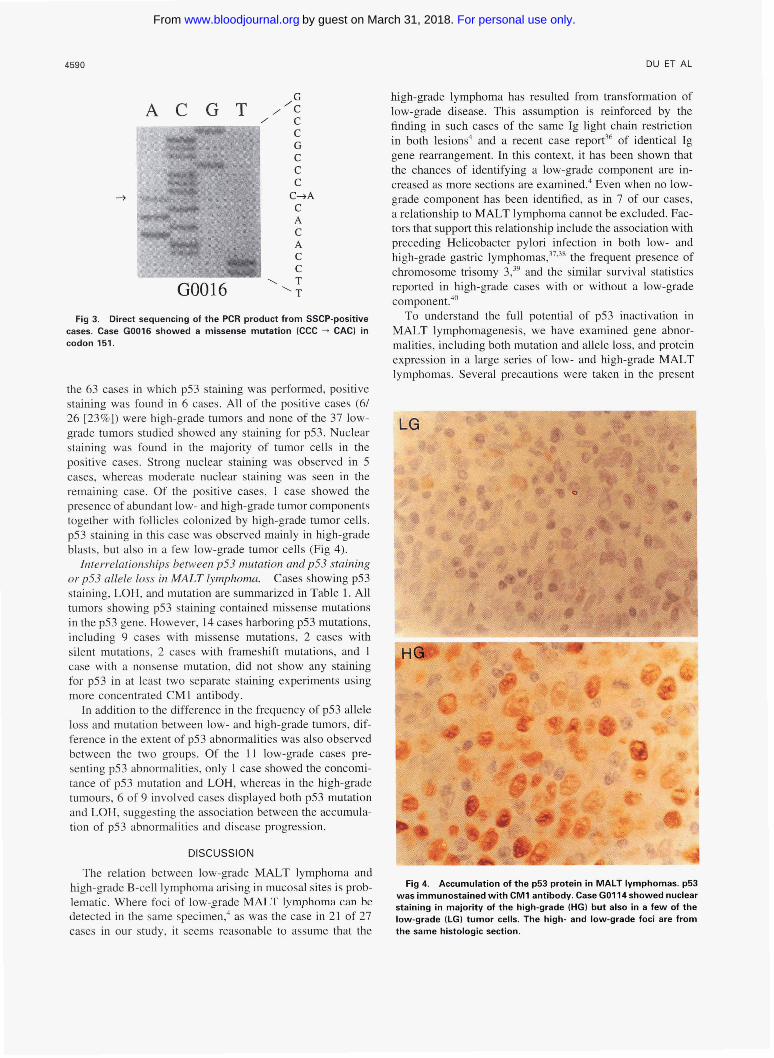
the 63 cases in which p53 staining was performed, positive staining was found in 6 cases. All of the positive cases (6/ 26 [23%]) were high-grade tumors and none of the 37 low- grade tumors studied showed any staining for p53. Nuclear staining was found in the majority of tumor cells in the positive cases. Strong nuclear staining was observed in 5 cases, whereas moderate nuclear staining was seen in the remaining case. Of the positive cases, 1 case showed the presence of abundant low- and high-grade tumor components together with follicles colonized by high-grade tumor cells. p53 staining in this case was observed mainly in high-grade blasts, but also in a few low-grade tumor cells (Fig 4).
Interrelationships between p53 mutation and p53 staining or p53 allele loss in MALT lymphoma. Cases showing p53 staining, LOH, and mutation are summarized in Table 1. All tumors showing p53 staining contained missense mutations in the p53 gene. However, 14 cases harboring p53 mutations, including 9 cases with missense mutations, 2 cases with silent mutations, 2 cases with frameshift mutations, and 1 case with a nonsense mutation, did not show any staining for p53 in at least two separate staining experiments using more concentrated CM1 antibody.
In addition to the difference in the frequency of p53 allele loss and mutation between low- and high-grade tumors, dif- ference in the extent of p53 abnormalities was also observed between the two groups. Of the 11 low-grade cases pre- senting p53 abnormalities, only 1 case showed the concomi- tance of p53 mutation and LOH, whereas in the high-grade tumours, 6 of 9 involved cases displayed both p53 mutation and LOH, suggesting the association between the accumula- tion of p53 abnormalities and disease progression.
DISCUSSION
The relation between low-grade MALT lymphoma and high-grade B-cell lymphoma arising in mucosal sites is prob- lematic. Where foci of low-grade MALT lymphoma rm he detected in the same specimen: as was the case in 21 of 27 cases in our study, it seems reasonable to assume that the
high-grade lymphoma has resulted from transformation of low-grade disease. This assumption is reinforced by the finding in such cases of the same Ig light chain restriction in both lesions4 and a recent case report36 of identical Ig gene rearrangement. In this context, it has been shown that the chances of identifying a low-grade component are in- creased as more sections are examined: Even when no low- grade component has been identified, as in 7 of our cases, a relationship to MALT lymphoma cannot be excluded. Fac- tors that support this relationship include the association with preceding Helicobacter pylori infection in both low- and high-grade gastric lymph0mas,3~”* the frequent presence of chromosome trisomy 3,39 and the similar survival statistics reported in high-grade cases with or without a low-grade component.40
To understand the full potential of p53 inactivation in MALT lymphomagenesis, we have examined gene abnor- malities, including both mutation and allele loss, and protein expression in a large series of low- and high-grade MALT lymphomas. Several precautions were taken in the present
R
Fig 4. Accumulation of the p53 protein in MALT lymphomas. p53 was immunostained with CM1 antibody. Case GOll4showed nuclear staining in majority of the high-grade (HGI but also in a few of the low-grade (LG) tumor cells. The high- and low-grade foci are from the same histologic section.
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom

p53 INACTIVATION IN MALT LYMPHOMA 459 1
study to ensure the maximal detection of p53 abnormalities. First, microdissection was performed in each case to isolate both tumor and nontumor cell populations, from which p53 gene analysis was performed. This has been proven to be the crucial step because MALT lymphoma frequently coex- ists with reactive components and many of our samples con- tained only a small tumor mass. The DNA isolated from a whole tumor section or tissue may be adequate for SSCP analysis in some cases but is far below the requirement for direct sequencing and LOH analysis due to the presence of abundant nontumor cells (our unpublished results). Sec- ondly, a highly sensitive SSCP analysis using polyacryl- amide agarose composite gel and a background free silver staining method was used in the present study, which, in a previous was shown to detect 97% of p53 mutations. Finally, a highly polymorphic p53 CA repeat was used for detection of LOH that allowed us to analyze nearly 90% of our samples.
Our results show that frequent p53 mutation and allelic loss are associated with both low- (LOH, 6.8%; mutation, 18.8%) and high- (LOH, 28.6%; mutation, 33.3%) grade MALT lymphomas, particularly in the high-grade tumors. Our data also indicate that there is a significant difference in the extent of p53 abnormalities between low- and high- grade MALT lymphomas. In low-grade tumours, most p53- defective cases (10/1 l ) showed either one allele mutation or one allele loss, suggesting only partial loss of p53 function. Whereas in high-grade tumors, the majority (6/9) of affected cases exhibited both p53 mutation and allele loss, implying complete loss of p53 function. Thus, the extent of p53 inacti- vation is closely associated with the progression of MALT lymphoma. A partial inactivation of p53 function may be involved in the development of some low-grade MALT lymphomas, whereas a complete loss of p53 function may be important for high-grade transformation. In this aspect, the pathologic role of p53 inactivation in MALT lymphoma is different from that in follicular lymphoma in which p53 mutation has been suggested to be mainly associated with high-grade transformation” or that in splenic marginal zone B-cell lymphoma in which p53 inactivation appears to be involved with the development of low-grade tumors.”
The observation of distinct p53 gene abnormalities at the different stages of MALT lymphoma shows the nature of multistage tumorigenesis and highlights the fact that the loss of normal p53 function is a process of the accumulation of its genetic abnormalities. Our results also emphasizes the importance of analyzing both p53 mutation and allele loss to evaluate its full potential in tumorigenesis. Distinct p53 gene abnormalities (p53 allele loss, p53 mutation, or both) may cause different degree of p53 inactivation, exert differ- ent tumorigenic potentials, and thus play different roles dur- ing multistage tumorigenesis.
Mutant p53 protein frequently accumulates in cell nuclei due to its increased half-life and can be detected by immuno- chemical ~taining.”.~’ p53 staining frequently indicates mu- tation of the gene. In the present study, all cases showing p53 staining contained missense mutations in the gene. How- ever, 14 cases harboring p53 mutation did not show any p53
staining with CM1 antibody. Staining was not expected for 5 cases, including 2 cases in which the mutation was silent and another 3 cases in which mutations resulted in truncated protein products. The mechanism underlying the absence of p53 staining in the remaining 9 cases, in which missense mutations were found, is unclear. Lack of staining in tumors harboring p53 missense mutation has been reported pre- v i o u ~ l y . ” ~ , ~ ~ Two possibilities may account for this. First, the accumulation of mutant p53 may not reach the level detect- able by immunochemistry. This could be due to the insuffi- cient stabilization of some mutations or lack of sufficient protein expression?2.M Secondly, the antibody used may not recognize all mutant p53. However, this possibility is small in our case because p53 staining was performed using CM1 and antigen retrieval method, which together have been shown to be highly sensitive for the ~taining.~’ In addition, the CM1 antibody is polyclonal and theoretically should recognize all p53. Nevertheless, the use of immunostaining with CM1 antibody alone as a marker for p53 mutation in MALT lymphomas is limited.
Increasing expression of mutant p53 during tumor pro- gression has been observed previously?* This suggests that additional mechanisms other than gene mutation may affect the protein expressionM and the expression of mutant p53 may also be functionally important. Increasing evidence sug- gests that mutant p53 can exert dominant negative effects on the ~ i1d- type . l~~~’ Thus, the lack of mutant p53 expression in tumors harboring p53 mutation may indicate the absence of such dominant negative effects and represent a relatively weak model of mutation-mediated p53 inactivation. In keep- ing with this hypothesis, the majority of our cases not show- ing protein accumulation but with mutation were low grade and did not show loss of the other allele. Further studies following-up p53 expression and disease progression in these cases are necessary to understand the significance of mutant p53 expression in tumorigenesis.
Analysis of the p53 mutation spectrum in a tumor may show the possible etiology involved in its carcinogene~is.’.~~ For example, the high frequency of G+T transversion in the codon 249 in human hepatocellular carcinoma is related to alfatoxin B 1 -mediated mutagenesis,4’ whereas the predomi- nant transition mutation in the CpG island observed in colon cancer and lymphomas suggests the endogenous mutagene- sis4’ No predominant type of mutation in the p53 gene was found in MALT lymphoma and the mechanism underlying the diverse p53 mutations in this tumor remains unclear. Oxygen-reactive species, which are actively produced during inflammatory diseases49 and cause various types of mutation in DNA:’ may be important in the carcinogenesis of MALT lymphoma because this tumor is frequently derived from a background of inflammatory diseases.
Despite the progress in the understanding of clinical, pathologic, and immunophenotypic features of MALT lymphoma, the genetic mechanism underlying its develop- ment and high-grade transformation is not fully understood. This tumor does not show bcl-2 or bcl-l rea~~angement.’~,~~ Although c-myc rearrangement has been reported in 1 of 2 high-grade MALT tumors,53 we have been unable to detect
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom

4592 DU ET AL
c-myc rearrangement in a relatively large series of well- defined high-grade MALT cases (our unpublished results). Recently, a high frequency (56%) of chromosome trisomy 3 has been found in low-grade MALT lymphoma, which indicates its important role in the development of the tumor.54 However, trisomy of chromosome 3 does not appear to be important for the high-grade transformation because its fre- quency is lower (33%) in high-grade tumors.” Thus, the finding of p53 inactivation in MALT lymphoma represents the first gene identified so far to be frequently involved in its development as well as high-grade transformation.
REFERENCES
1 . Isaacson P, Wright DH: Malignant lymphoma of mucosa-asso- ciated lymphoid tissue. A distinctive type of B-cell lymphoma. Can- cer 52:1410, 1983
2. Isaacson P, Spencer J: Malignant lymphoma of mucosa-associ- ated lymphoid tissue. Histopathology 11:445, 1987
3. lsaacson PG: Lymphomas of mucosa-associated lymphoid tis- sue (MALT). Histopathology 16:617, 1990
4. Chan JKC, Ng CS, Isaacson PG: Relationship between high- grade lymphoma and low-grade B-cell mucosa-associated lymphoid tissue lymphoma (MALToma) of the stomach. Am J Pathol 136:1153, 1990
5. Kastan MB, Onyekwere 0, Sidransky D, Vogelstein B, Craig RW: Participation of p53 protein in the cellular response to DNA damage. Cancer Res 51 :6304, 1991
6. Kuerbitz SJ, Plunkett BS, Walsh WV, Kastan MB: Wild-type p53 is a cell cycle checkpoint determinant following irradiation. Proc Natl Acad Sci USA 89:7491, 1992
7. Lane DP: p53, guardian of the genome. Nature 358:15, 1992 8. Hollstein M, Sidransky D, Vogelstein B, Harris C: p53 muta-
tions in human cancers, Science 253:49, 1991 9. Levine AJ, Perry ME, Chang A, Silver A, Dittmer D, Wu M,
Welsh D: The 1993 Walter Lecture: The role of the p53 tumour- suppresser gene in tumourigenesis. Br J Cancer 69:409, 1994
IO. Shirasawa S, Urabe K, Yanagawa Y, Toshitani K, Iwama T, Sasazuki T: p53 gene mutations in colorectal tumours from patients with familial polyposis coli. Cancer Res 51:2874, 1991
1 I . Malkin D, Li FP, Strong LC, Graumeni JF, Nelson CE, Kim DH, Kassel J, Gryka MA, Bischoff FZ, Tainsky MA, Friend SH: Germline p53 mutations in a familial syndrome of breast cancer, sarcomas, and other neoplasms. Science 250:1233, 1990
12. Lassam NJ, From L, Kahn HJ: Overexpression of p53 is a late event in the development of malignant melanoma. Cancer Res 53:2235, 1993
13. Nigro, JM, Baker SJ, Preisinger AC, Jessup JM, Hostetter R, Cleary K, Bigner SH, Davidson N, Baylin S, Devilee P, Glover T, Collin FS, Weston A, Modali R, Harris CC: Mutation of the p53 gene occurs in diverse tumour types. Nature 342:705, 1989
14. Caron de Fromental C, Soussi T: TP53 tumour suppressor gene: A model for investigating human mutagenesis. Genes Chro- mosom Cancer 4: I , 1992
15. Masuda H, Miller C, Koeffler HP, Battifora H, Cline MJ: Rearrangement of the p53 gene in human osteogenic sarcoma. Proc Natl Acad Sci USA 84:7716, 1987
16. Oliner JD, Kinzler KW, Meltzer PS, George DL, Vogelstein B: Amplification of a gene encoding a p53-associated protein in human sarcomas. Nature 358:80, 1992
17. Lane DP: p53 and human cancers. Br Med Bull 50582, 1994 18. Nakai H, Misawa S, Toguchida J, Yandel DW, Ishizalu K:
Frequent p53 gene mutations in blast crisis of chronic myelogenous
leukaemia, especially in myeloid crisis harbouring loss 01 a chromo- some 17p. Cancer Res 52:6588, 1992
19. Srivastava S, Wang S, Tong YA, Hao ZM, Chang EH: Domi- nant negative effect of a germ-line mutant p53: A step tostering tumourigenesis. Cancer Res 53:4452, 1993
20. Frebourg T, Sadelain M, Ng YS, Kassel J, Friend SH: Equal transcription of wild-type and mutant p53 using bicistronic vectors results in the wild-type phenotype. Cancer Res 54:878, 1994
2 I . Harvey M, Vogel H, Morris D, Bradley A, Bernstein A, Done- hower LA: A mutant p53 transgene accelerates tumour development in heterozygous but not nullizygous p53-deficient mice. Nat Genet 9:305, 1995
22. Gaidano G, Ballerini P, Gong JZ, Inghirami G, Neri A. New- comb EW, Magrath IT, Knowles DM, Dalla-Favera R: p53 mutation in human lymphoid malignancies associated with Burkitt lymphoma and chronic lymphocytic leukaemia. Proc Natl Acad Sci USA 88:5413. 1991
23. Sugimoto K, Toyoshima H, Sakai R, Miyagawa K, Hagiwara K, Hirai H, Ishikawa F, Takaku F: Mutations of the p53 gene in lymphoid leukemia. Blood 77: I 153, I991
24. Cesarman E, Chadburn A, Inghirami G, Gaidano G , Knowles DM: Structural and functional analysis of oncogenes and tumour suppressor genes in adult T-cell leukemidymphoma shows frequent p53 mutations. Blood 80:3205, 1992
25. Wada M, Bartram CR, Nakamura H, Hachiya M, Chen DL, Borenstein J. Miller CW, Ludwig L, Hansen-Hagge TE, Ludwig WD. Reiter A, Mizoguchi H, Koeffler HP: Analysis of p53 mutation in a large series of lymphoid hematologic malignancies of childhood. Blood 82:3163, 1993
26. lchikawa A, Hotta T, Saito H: Mutations of the p53 gene in B-cell lymphoma. Leuk Lymphoma 1 l :2l , 1993
27. Sander CA, Yano T, Clark HM, Harris C, Longo D, Jaffe ES. Raffeld M: p53 mutation is associated with progression in follicular lymphomas. Blood 82:1994, 1993
28. Baldini L, Francchiolla NS, Cro LM, Trecca D, Romitti L, Polli E, Maiolo AT, Neri A: Frequent p53 gene involvement in splenic B-cell leukaemidlymphomas of possible marginal zone ori- gin. Blood 84:270. 1994
29. Isaacson PG: Malignant lymphoma of the gastrointestinal tract, in lsaacson PG, Norton AJ (eds): Extranodal Lymphomas. New York. NY, Churchill Livingstone, 1994, p 15
30. lsaacson PG, Matures E, Burke M, Catovsky D: The histopa- thology of splenic lymphoma with villous lymphocytes. Blood 84:3828, 1994
3 I . Pan LX, Diss TC, Peng HZ, Isaacson PG: Clonality analysis of defined B-cell populations in archival tissue sections using micro- dissection and polymerase chain reaction. Histopathology 24:323, 1994
32. Jones MH, Nakamura Y : Detection of loss of heterozygosity at the human TP53 locus using a dinucleotide repeat polymorphism. Genes Chromosom Cancer 5:89, 1992
33. Murakami Y, Hayashi K, Sekiya T: Detection of aberrations of the p53 alleles and the gene transcript in human tumor cell lines by single-strand conformation polymorphism analysis. Cancer Res 5 1 :3356, 1991
34. Peng HZ, Du MQ, Ji JX, Isaacson PG, Pan LX: High resolu- tion SSCP analysis using polyacrylamide agarose composite gel and a background silver staining method. Biotechniques 19:410, 1995
35. Norton AJ, Jordan S, Yeomans P: Brief high temperature heat denaturation (pressure cooking): A simple and effective method of antigen retrieval for routinely processed tissues. J Pathol 173:371, 1994
36. Montalban C, Manzanal A, Castrillo JM, Escribano L, Bellas C: Low grade gastric B-cell MALT lymphoma progressing into high
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom

p53 INACTIVATION IN MALT LYMPHOMA 4593
grade lymphoma. Clonal identity of the two stages of the tumour, unusual bone involvement and leukaemic dissemination. Histopa- thology 27:89, 1995
37. Wotherspoon AC, Ortiz Hidalgo C, Falzon MR, Isaacson PG: Helicobacter pylori-associated gastritis and primary B-cell gastric lymphoma. Lancet 338:1175, 1991
38. Parsonnet J, Hansen S, Rodriguez L, Gelb AB, Warnke RA, Jellum E, Orentreich N, Vogelman JH, Friedman GD: Helicobacter pylori infection and gastric lymphoma. N Engl J Med 330:1267, 1994
39. Wotherspoon AC, Finn T, Isaacson PG. Numerical abnormal- ities of chromosome 3 and 7 in lymphomas of mucosa associated lymphoid tissue and the spleen marginal zone. Lab Invest 70: 124A, 1994
40. Cogliatti SB, Schmid U, Schumacher U, Eckert F, Hansmann ML, Hedderich J, Takahashi H, Lennert K: Primary B-cell gastric lymphoma: A clinicopathological study of 145 patients. Gastroenter- ology 101:1159, 1991
41. Rotter V: p53, a transformation-related cellular encoded pro- tein, can be used as a biochemical marker for the detection of primary mouse tumour cells. Proc Natl Acad Sci USA 80:2613, 1983
42. Wynford-Thomas D: p53 in tumour pathology: Can we trust immunocytochemistry? J Pathol 166:329, 1992
43. Dix B, Robbin P, Carrello S, House A, Iacopetta B: Compari- son of p53 mutation and protein overexpression in colorectal carcino- mas. Br J Cancer 70585, 1994
44. Hall PA, Lane DP: p53 in tumour pathology: Can we trust immunohistochemistry?-Revisited! J Pathol 172:1, 1994
45. Baas IO, Mulder JWR, Offerhaus GJO, Vogelstein B, Hamil-
ton SR: An evaluation of six antibodies for immunohistochemistry of mutant p53 gene product in archival colorectal neoplasms. J Pathol 1725, 1994
46. Harris CC: p53: At the crossroads of molecular carcinogenesis and risk assessment. Science 262:1980, 1993
47. Bressac B, Kew M, Wands J, Ozturk M: Selective G to T mutations of p53 gene in hepatocellular carcinoma from Southern Africa. Nature 350:429, 1991
48. Rideout WM 3d, Coetzee-GA, Olumi AF, Spruck CH, Jones PA: 5-Methylcytosine as an endogenous mutagen in the p53 tumour suppressor gene. Princess Takamatsu Symp 22:207, 1991
49. Weitzman SA, Gordon LI: Inflammation and cancer: Role of phagocyte-generated oxidants in carcinogenesis. Blood 76:655, 1990
50. McBride TJ, Preston BD, Loeb LA: Mutagenic spectrum re- sulting from DNA damage by oxygen radicals. Biochemistry 30:307, 1991
51. Pan LX, Diss TC, Cunningham D, Isaacson PG: The hcl-2 gene in primary B-cell lymphoma of mucosa-associated lymphoid tissue (MALT). Am J Pathol 135:7, 1989
52. Wotherspoon AC, Pan L, Diss TC, Isaacson PG: A genotypic study of low grade B-cell lymphomas, including lymphomas of mu- cosa associated lymphoid tissue (MALT). J Pathol 162:135, 1990
53. Krieken JHJM, Raffeld M, Raghoebier S, Jaffe ES, van Om- men GJB, Kluin PM: Molecular genetics of gastrointestinal non- Hodgkin’s lymphomas: Unusual prevalence and pattern of c-myc rearrangements in aggressive lymphomas. Blood 76:797, 1990
54. Wotherspoon AC, Finn TM, Isaacson PG: Trisomy 3 in low grade B cell lymphomas of mucosa associated lymphoid tissue (MALT). Blood 85:2000, 1995
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom

1995 86: 4587-4593
M Du, H Peng, N Singh, PG Isaacson and L Pan progression of mucosa-associated lymphoid tissue lymphomaThe accumulation of p53 abnormalities is associated with
http://www.bloodjournal.org/content/86/12/4587.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on March 31, 2018. by guest www.bloodjournal.orgFrom