
The Abundant NK Cells in Human Secondary Lymphoid … · The Abundant NK Cells in Human Secondary...
9
of August 6, 2018. This information is current as Receptors and Become Cytolytic Activation to Express Killer Cell Ig-Like Secondary Lymphoid Tissues Require The Abundant NK Cells in Human Moretta and Christian Münz Goodman, Barbara Morandi, William A. Muller, Alessandro Guido Ferlazzo, Dolca Thomas, Shao-Lee Lin, Kiera http://www.jimmunol.org/content/172/3/1455 doi: 10.4049/jimmunol.172.3.1455 2004; 172:1455-1462; ; J Immunol References http://www.jimmunol.org/content/172/3/1455.full#ref-list-1 , 13 of which you can access for free at: cites 33 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2004 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on August 6, 2018 http://www.jimmunol.org/ Downloaded from by guest on August 6, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of The Abundant NK Cells in Human Secondary Lymphoid … · The Abundant NK Cells in Human Secondary...

of August 6, 2018.This information is current as
Receptors and Become CytolyticActivation to Express Killer Cell Ig-LikeSecondary Lymphoid Tissues Require The Abundant NK Cells in Human
Moretta and Christian MünzGoodman, Barbara Morandi, William A. Muller, Alessandro Guido Ferlazzo, Dolca Thomas, Shao-Lee Lin, Kiera
http://www.jimmunol.org/content/172/3/1455doi: 10.4049/jimmunol.172.3.1455
2004; 172:1455-1462; ;J Immunol
Referenceshttp://www.jimmunol.org/content/172/3/1455.full#ref-list-1
, 13 of which you can access for free at: cites 33 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Abundant NK Cells in Human Secondary LymphoidTissues Require Activation to Express Killer Cell Ig-LikeReceptors and Become Cytolytic1
Guido Ferlazzo,2*† Dolca Thomas,‡ Shao-Lee Lin,* Kiera Goodman,* Barbara Morandi,†
William A. Muller,§ Alessandro Moretta,¶ and Christian Munz2*
Natural killer cells are important cytolytic cells in innate immunity. We have characterized human NK cells of spleen, lymphnodes, and tonsils. More than 95% of peripheral blood and 85% of spleen NK cells are CD56dimCD16� and express perforin, thenatural cytotoxicity receptors (NCRs) NKp30 and NKp46, as well as in part killer cell Ig-like receptors (KIRs). In contrast, NKcells in lymph nodes have mainly a CD56brightCD16� phenotype and lack perforin. In addition, they lack KIRs and all NCRexpression, except low levels of NKp46. The NK cells of tonsils also lack perforin, KIRs, NKp30, and CD16, but partially expressNKp44 and NKp46. Upon IL-2 stimulation, however, lymph node and tonsilar NK cells up-regulate NCRs, express perforin, andacquire cytolytic activity for NK-sensitive target cells. In addition, they express CD16 and KIRs upon IL-2 activation, andtherefore display a phenotype similar to peripheral blood NK cells. We hypothesize that IL-2 can mobilize the NK cells ofsecondary lymphoid tissues to mediate natural killing during immune responses. Because lymph nodes harbor 40% and peripheralblood only 2% of all lymphocytes in humans, this newly characterized perforin� NK cell compartment in lymph nodes and relatedtissues probably outnumbers perforin� NK cells. These results also suggest secondary lymphoid organs as a possible site of NKcell differentiation and self-tolerance acquisition. The Journal of Immunology, 2004, 172: 1455–1462.
N atural killer cells have been described as innate immuneeffectors, able to exert a prompt cytolytic activity againstinfected and tumor cells without prior stimulation (1, 2).
Accordingly, perforin, a potent cytolytic pore-forming protein, isconstitutively contained in cytoplasmic granules of almost all pe-ripheral blood NK cells (3), and these readily kill MHC class I-deficient targets (4). Ninety-five percent of the NK cells in bloodare CD56dimCD16�perforin� cells, while the remainder areCD56brightCD16�perforin� cells (3, 5). CD56dim NK cells fromblood can be attracted to peripheral tissues by several chemokinesreleased during inflammatory responses (6). In contrast, the mi-nority CD56brightCD16� NK cells from blood express receptorsfor homing to lymphoid tissues, such as CCR7, the receptor forCCL19 and CCL21 chemokines, and CD62L (6). Indeed, Caligiuriand coworkers (7) have recently found a CD56brightCD16� NKsubset in T cell areas of inflamed lymph nodes and could elicitIFN-� secretion of peripheral blood CD56brightCD16� NK cells by
stimulation with IL-2. Because lymph nodes contain 40% of alllymphocytes in humans, while blood harbors only 2% (8, 9), NKcells of secondary lymphoid organs might be the predominant NKcompartment in humans.
We present in this study a comprehensive analysis of human NKcells in secondary lymphoid organs, emphasizing their differencesfrom peripheral blood NK cells in inhibitory and activating recep-tors as well as cytolytic activity. We compared CD3�CD56� cellsfrom spleen, tonsils, and lymph nodes with peripheral blood NKcells. Most of the NK cells in spleen are similar to those in blood,but surprisingly, the cells in lymph nodes and tonsils differ fromthe major as well as minor NK cell subset in peripheral blood byphenotype and function. They are initially negative for CD16,killer cell Ig-like receptors (KIRs),3 perforin, and most natural cy-totoxicity receptors (NCRs). These markers and function, i.e.,CD16, perforin, KIRs, NCRs, and cytolytic activity, develop afterstimulation with IL-2. Therefore, unlike natural cytotoxicity inblood, the cytolytic function of the NK cells in lymphoid tissuesappears to require activation to be mobilized.
Materials and MethodsSecondary lymphoid organ cell isolation and culture
Fifteen lymph nodes (from 10 donors), 8 palatine tonsils (from 5 donors),and 3 spleens were studied. The regional Organ Procurement Organizationprocured 5 human mesenteric lymph nodes and the autologous spleensfrom 3 individual brain-dead donors after obtaining informed consent fromappropriate individuals. Ten additional uninflamed, normal lymph nodeswere obtained from 7 patients who underwent surgical resections of cancermasses. Six tonsils from 4 different individuals were collected immediatelyafter surgery from patients undergoing tonsillectomy for chronic inflam-mation. All tissues were obtained as part of Institutional Review Board-approved protocols. Soon after their removal, tissues were dissociated toobtain single cell suspensions. Mechanically dissociated tissues were then
*Laboratory of Cellular Physiology and Immunology, The Rockefeller University,New York, NY 10021; †Istituto Nazionale per la Ricerca sul Cancro, Genova, Italy;‡Division of Nephrology, Department of Medicine, and §Department of Pathologyand Laboratory Medicine, Weill Medical College, New York, NY 10021; and ¶Di-partimento di Medicina Sperimentale, Sezione di Istologia, and Centro di Eccellenzaper le Ricerche Biomediche, Universita degli Studi di Genova, Genova, Italy
Received for publication June 12, 2003. Accepted for publication November19, 2003.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was funded by a Special Fellowship from the Leukemia and LymphomaSociety and by a grant from the Speaker’s Fund for Public Health Research, awardedby the City of New York (to C.M.); by a fellowship from the American Society ofTransplantation, Juvenile Diabetes Foundation (to D.T.); and by grants from AssociazioneItaliana per la Ricerca sul Cancro and from Ministero della Salute, Italy (to G.F.).2 Address correspondence and reprint requests to Drs. Guido Ferlazzo or Christian Munz,Laboratory of Cellular Physiology and Immunology, The Rockefeller University, 1230York Avenue, New York, NY 10021-6399. E-mail address: [email protected] [email protected]
3 Abbreviations used in this paper: KIR, killer cell Ig-like receptor; NCR, naturalcytotoxicity receptor; NTBA, NK-T-B Ag; ULBP, UL16-binding protein.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

filtered through a 100-�m nylon cell strainer to exclude undissociated frag-ments (BD Labware, Mountain View, CA). Debris and dead cells wereeliminated using a Ficoll-Hypaque discontinuous gradient. Single cell sus-pensions were then extensively washed and analyzed by flow cytometry orcryopreserved. Cells were also cultured for the indicated time periods inRPMI medium in the presence of 10% FCS and different doses of rIL-2(ranging from 500 to 5 IU/ml).
To evaluate IFN-� production by NK cells, tonsilar mononuclear cellswere cultured for 24 h in the presence of IL-12 (10 ng/ml), IL-2 (5 IU/ml),or both stimuli. Monensin (2 �M) was added to the cultures for the last 6 hof incubation.
mAbs, flow cytometry, and NK cell phenotypic analysis
Analysis of cell surface markers was performed using the following mAbsin direct immunofluorescence assays: allophycocyanin- or FITC-conju-gated anti-CD3, allophycocyanin-conjugated CD62L, PE- or CyChrome-conjugated anti-CD56, FITC-conjugated anti-CD16, anti-CD69, anti-HLA-DR, anti-CD8, and anti-CCR7, all from BD PharMingen (San Diego,CA), PE-conjugated anti-NKp30, NKp46, and NKp44 were from Immu-notech-Coulter (Marseille, France). CO202 mAb (anti-CD48), XA141 (anti-CD158a/p58.1 and anti-CD158h/p50.1), Y249 (anti-CD158b1/p58.2 andanti-CD158j/p50.2), AZZ158 (anti-CD158e1/e2/p70 and anti-CD158k/p140), Z176 (anti-p75-AIRM1), Z199 (anti-CD159a/NKG2A),ECM217/1 (anti-NKG2D), CO54 (anti-2B4), and ON56 (anti-NK-T-B Ag(NTBA)) were produced in A.M.’s laboratory or were kind gifts of L.Moretta (Istituto Giannina Gaslini, Genova, Italy). Direct immunofluores-cence staining was performed by diluting fluorochrome-labeled mAb with1 mg/ml human �-globulin (human therapy grade from Biotest srl, Milan,Italy), to block nonspecific FcR binding. Cells were then washed, and theflow cytometric analysis was performed. For indirect immunofluorescencestaining, nonspecific binding sites were saturated with human �-globulin,and then the relevant mAb was added and incubated for 30 min at 4°C.After extensive washing, FITC- or PE-conjugated isotype-specific goat anti-mouse Abs (Southern Biotechnology Associates, Birmingham, AL) wereadded and incubated for 30 min at 4°C. Negative controls included directlylabeled or unlabeled isotype-matched irrelevant mAbs. Cells were thenextensively washed and analyzed by flow cytometry. Perforin and IFN-�expression analysis on freshly isolated or activated NK cells was per-formed, respectively, by FITC-conjugated perforin Ab reagent set and al-lophycocyanin-conjugated anti-IFN-� (BD Biosciences-PharMingen) aftercells were fixed in 1% paraformaldehyde and permeabilized.
NK cell cytotoxicity assay
To evaluate the cytolytic activity after IL-2 stimulation, we used the NKcell-sensitive lymphoblastoid cell line 721.221, which does not expresssurface HLA class I molecules (10), as well as the T-lymphoblastoid cellline CCRF-CEM (referred in the text as CEM). The latter was used astarget because of its expression of UL16-binding protein (ULBP) 1 andULBP2 (11), two ligands of NKG2D, the only activating receptor ex-pressed by freshly isolated lymph node or tonsil NK cells. Cytotoxicityassays were performed, as previously described (12). Briefly, 106 targetcells were incubated with the dye molecule PKH26 (Sigma-Aldrich, St.Louis, MO), which inserts into plasma membranes due to its lipophilicaliphatic residue. Target cells were then incubated with secondary lym-phoid organ cells at different NK cell/target ratios. After 4 h, cells wereharvested; TO-PRO-3, a membrane-impermeable DNA stain, was added toeach culture (1 �M final concentration); and cells were finally analyzed byflow cytometry. PKH26 is excited at 551 nm, and TO-PRO-3, using theHe-Ne laser, at 633 nm. Thus, target cells are easily gated by PKH26 redfluorescence, while undamaged target cells can be distinguished from lysedcells by their ability to exclude the DNA stain after the coincubation witheffector cells. Background and maximum TO-PRO-3 staining were ob-tained by incubation with medium and detergent, respectively.
Proliferation assay by CFSE dilution
Mononuclear cells of tonsils and lymph nodes were labeled with 5 �MCFSE in PBS � 0.1% BSA for 10 min at 37oC. After washing twice withPBS � 0.1% BSA, cells were incubated with 100 IU/ml rIL-2 (Chiron,Emeryville, CA) for 7 days at 37oC. CFSE fluorescence (FL-1) was eval-uated on CD3�CD56� cells by four-color flow cytometry.
ResultsSubsets of NK cells in human secondary lymphoid organs
Mononuclear cells isolated from lymph nodes, spleens, and tonsilswere analyzed for CD3�CD56� NK cells (Fig. 1). NK cells rep-
resented 5% of the total mononuclear cells isolated from unin-flamed lymph nodes (median value in 15 lymph nodes, range1–7%). In inflamed tonsils, the median value of CD56�CD3� cellswas 0.4% of the total cells (range 0.2–0.7% in six tonsils). Unin-flamed spleens showed a variable percentage of NK cells (7, 17,and 50% in three donors), generally higher than in peripheralblood. Interestingly, the CD56� bright subset of NK cells wasdominant in tonsils and lymph nodes (75% median value; Fig. 1),whereas in peripheral blood and spleen the majority of NK cellswere CD56dim (95 and 85%, respectively, median value; Fig. 1).Spleen consists primarily of red pulp, which in turn is rich in bloodvessels, so that splenic NK cells may primarily represent bloodcells with a minor contribution of the white pulp lymphoid tissueareas. Our data on the dominance of CD56bright NK cells in lymphnodes and tonsils parallel a prior report on human inflamed lymphnodes (7).
Because NK cells present in secondary lymphoid organs ap-peared different from peripheral blood, we performed a more com-plete phenotypic characterization. Representative experiments areshown in Fig. 2A and summarized in Table I. We found that NKcells in lymph nodes and tonsils were consistently negative for theFc�RIII (CD16). CD16 triggers NK recognition of Ab-opsonizedtarget cells during Ab-dependent cellular cytotoxicity (13, 14).Noteworthily, while the CD56dim subsets in the spleen and in theperipheral blood express comparable levels of CD16, bothCD56bright and CD56dim NK cells in lymph nodes and tonsils werenegative for CD16. A similar pattern of expression was detectableusing mAbs directed against the CD8 molecule, i.e., most CD56�
cells in tonsils and lymph nodes were CD8�. NK cell activationmarkers, such as HLA-DR and CD69, were slightly increased inthe six tonsils analyzed, but not in the lymph nodes. This confirmsthat our tonsils represent inflamed secondary lymphoid tissue,while our lymph nodes were uninflamed. Finally, the pan-leuko-cyte Ag CD48 was expressed at similar levels on all the NK cellsubsets analyzed. Therefore, the NK cells of lymph nodes are pri-marily not activated with high CD56, but low CD16, HLA-DR,and CD69 levels.
Because CD56bright NK cells in peripheral blood represent theonly NK cell subset expressing CCR7 and CD62L, we analyzedwhether lymph node or tonsil CD56bright NK cells express thesame receptors for homing to lymphoid tissues. As shown in Fig.2B, we confirmed the expression of CCR7 on peripheral bloodCD56bright NK cells. In contrast, surface expression of eitherCCR7 (Fig. 2B) or CD62L (data not shown) was not detectable onNK cells isolated from two lymph nodes and three tonsils. Only inone lymph node we detected 10% of NK cells expressing CCR7,
FIGURE 1. CD56bright NK cells are enriched in human secondary lym-phoid organs. Single mononuclear cell suspensions were obtained fromperipheral blood, lymph nodes, spleens, and tonsils. Cells were then ana-lyzed by flow cytometry using specific mAb against CD3 (allophycocyaninconjugated) and CD56 (PE conjugated). Numbers indicate the medianvalue of CD56-positive cell percentages in all the experiments (ranges areshown in brackets).
1456 CHARACTERIZATION OF NK CELLS IN SECONDARY LYMPHOID ORGANS
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

but not CD62L. Of note, this CCR7 expression was lost upon IL-2culture (data not shown).
We next analyzed NK cells of tonsils, lymph nodes, and spleensfor their expression of inhibitory receptor. The HLA class I-spe-
cific killer inhibitory and activating receptors (KIRs; CD158; KIR-DL; KIR-DS) were absent on NK cells of lymph nodes and tonsils,but detectable in peripheral blood and spleen (Fig. 3 and Table I).CD158a/h(p58.1/p50.1) and CD158e1/e2/k(p70/p140) were ex-pressed on subsets of CD56dim cells in blood and spleen, but notlymph nodes and tonsils. In all autologous sets of spleen andlymph nodes, CD158 expression was only found on spleen NKcells, but absent in lymph node NK cells (one example is shown inFig. 3). The polymorphic CD158 receptors recognize shared allelicdeterminants of polymorphic HLA class Ia molecules (15). Dif-ferences in their expression between individuals are the basis forNK recognition of allogeneic cells in humans. The HLA-E-specificCD94/CD159a(NKG2A) inhibitory receptor (16) was expressedon most of the CD56bright NK cells in lymph nodes and tonsils. Inspleen, subsets of CD56bright and CD56dim NK cells expressedCD94/CD159a(NKG2A). All CD56bright NK cells and a part ofCD56dim NK cells in blood were CD94/CD159a(NKG2A)�. Itsligand, HLA-E, is a nonclassical HLA class Ib molecule that pref-erentially binds leader sequences of MHC class I molecules, andits surface expression is therefore regulated by the overall HLAclass I level in cells (17). Finally, the inhibitory sialoadhesin fam-ily molecule p75/AIRM1 of unknown ligand specificity (18) washomogenously expressed on NK cells of all lymphoid organs (Fig.3 and Table I). Therefore, p75/AIRM-1 and CD94/NKG2A seemto be the predominant inhibitory receptors on NK cells in lymphnodes, whereas CD158 KIRs are scarce.
When we turned to activating receptors, we found expression ofNKG2D and the coreceptors 2B4 and NTBA on NK cells of allanalyzed organs (Fig. 4 and Table I). NKG2D binds the stress-induced MHC class I-like MICA/B surface Ags as well as thehuman CMV ULBPs (19, 20); 2B4 recognizes CD48 (21); andNTBA enhances NK activation after engagement of unknown li-gands to date (22). Surprisingly, NK cells of uninflamed lymphnodes showed no expression of NCRs, except for low levels ofNKp46. NCRs mediate tumor and dendritic cell recognition by NKcells (23, 24). In contrast, peripheral blood NK cells and NK cellsfrom spleen demonstrated NKp30 and NKp46 expression on allNK cells (Fig. 4). Tonsils, like lymph nodes, lacked expression ofNKp30, but expressed ex vivo NKp44 and NKp46 on some NKcells (Fig. 4 and Table I). NKp44, an activation-induced NCR, wasabsent on NK cells from spleen, peripheral blood, and lymphnodes. Together with the CD69 and HLA-DR expression (Fig. 2),this argues for activation by inflammation of our tonsilar NK cells.Interestingly, NKp44 expression in the tonsils was consistentlyhigher than the usually constitutive NCRs NKp30 and NKp46.This NK phenotype has not been previously observed, neither invitro nor ex vivo. The absence of NCRs on NK cells from tonsilsand lymph nodes does not reflect hereditary absence of these mol-ecules in the analyzed donors, and is not a result of loss during thepreparation of the cell suspension, because in the spleens, autolo-gous to our lymph node samples, NCR expression was conserved(Fig. 4). Furthermore, addition of tonsilar cell suspensions to pe-ripheral blood NK cells did not decrease NCR expression on bloodNK cells, arguing against proteases in tonsil and lymph node prep-arations that destroy NCRs (data not shown). Therefore, NCR ex-pression on lymph node NK cells is primarily restricted to lowlevels of NKp46.
In summary, we found four distinct NK subsets in tonsils andlymph nodes. Three of them, CD56brightCD159a�NCR�, CD56bright
CD159a�NCR�, and CD56dimCD16�NCR� NK cells, have not pre-viously been described, and one, CD56brightNKp44� NK cells, hadnever been isolated ex vivo. In addition, we detected an overall en-richment of CD56brightCD16� NK cells in spleen, lymph nodes, andtonsils, compared with blood. Although in peripheral blood less than
FIGURE 2. NK cell phenotype in secondary lymphoid organs. A, Cellsisolated from different lymphoid organs were analyzed by three-color flowcytometry using specific mAb against the indicated markers. The analysiswas performed after gating on CD3-negative cells, thus excluding contam-inating activated T cells expressing CD56. Note that the depicted lymphnode and spleen cell suspensions are from the same donor. B, CD56bright
NK cells isolated from tonsil (three experiments) do not express CCR7.Similar results were obtained in two of three lymph nodes, analyzed forCCR7 expression. In one lymph node, part of the NK cells (10%) ex-pressed CCR7, and this expression was lost upon IL-2 activation of NKcells. T cells isolated from peripheral blood, tonsil, or lymph node consis-tently showed partial expression of CCR7.
1457The Journal of Immunology
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
5% of CD56�CD3� mononuclear cells are CD56brightCD16�, theseamount to 15% in spleen and 75% in lymph nodes and tonsils.
Lymph node and tonsil NK cells up-regulate expression ofCD16, KIRs, NCRs, and perforin upon culture with IL-2, andconcomitantly become cytolytic
NCRs are surface receptors that play a major role in NK cell trig-gering during natural cytotoxic response, and CD16 mediates NKrecognition of Ab-opsonized targets. The finding of an impairedexpression of these activating receptors on both lymph node andtonsilar NK cells suggested a decreased cytolytic ability of thesecells. Indeed, lymph node and tonsilar NK cells lacked perforin,the main effector molecule in rapid natural cytotoxicity (Fig. 5A).Even NKp44-positive NK cells in tonsils had no detectable per-forin expression. This suggests that in these cells the expressedNCR molecules might trigger NK cell functions other than cyto-toxicity. To address whether these NK cells can acquire cytolyticfunctions after exposure to T cell-derived cytokines, we incubatedthem with IL-2 for up to 1 wk. After IL-2 activation, both lymphnode and tonsilar NK cells significantly up-regulated CD16 and allNCRs (Fig. 6). In addition, IL-2 induced KIR expression (Fig. 6).Therefore, IL-2 activation induced activating as well as inhibitoryreceptors on NK cells from lymph nodes and tonsils, and changedtheir phenotype to one reminiscent of the predominant CD16� NKsubset in peripheral blood. It is unlikely that these CD16/CD158�
NK cells represent the IL-2-driven expansion of a minor fractionof CD16� cells already present ex vivo, because NK cells largelyexpanded in number and up to 50% of the cells expressed CD16.Indeed, NK cells from tonsils and lymph nodes expanded 3- to
30-fold more than T cells in the same tissue (data not shown). Tofurther rule out the possibility that CD16�KIR� NK cells wereexpanded from a NK subset, which already expressed these mark-ers ex vivo, CD3�CD56�CD16� tonsil cells were sorted by flowcytometry to 99.5% purity and cultured in IL-2 for 1 wk. As shownin Fig. 6B, purified CD16� NK cells from tonsils expressed CD16and KIRs after IL-2 activation. We obtained CD16�KIR� NKsubpopulations of similar size to cultures from unseparated tonsiland lymph node cells. Finally, we also assessed the proliferation oftonsil (Fig. 6C) and lymph node (data not shown) NK cells uponstimulation with IL-2. CFSE dilution of CD3�CD56� cells wasevaluated after 1-wk activation with 100 IU/ml IL-2. Fig. 6Cshows that KIR� NK cells proliferated more vigorously and that,interestingly, KIRs are acquired by NK cells in secondary lym-phoid organs without many divisions. The subset of KIR� NKcells that underwent massive proliferation represented 35% of totalNK cells in the tonsil and 31% in the lymph node analyzed. Theseresults also further ruled out that KIR� NK cells observed after 1wk of culture in IL-2 represent the preferential proliferation andexpansion of pre-existing KIR� NK cells in the starting popula-tion, and suggest that lymphoid tissues might be a site for NK celldifferentiation.
Because IL-2-activated NK cells of tonsils and lymph nodesacquired a phenotype similar to cytotoxic peripheral blood NKcells, we addressed whether they would also become cytolyticupon culture with IL-2. We investigated perforin expression andcytolytic ability before and after IL-2 activation. Perforin expres-sion was initially absent, but detectable as early as 24 h after IL-2administration. After 7 days of IL-2 stimulation, it was expressed
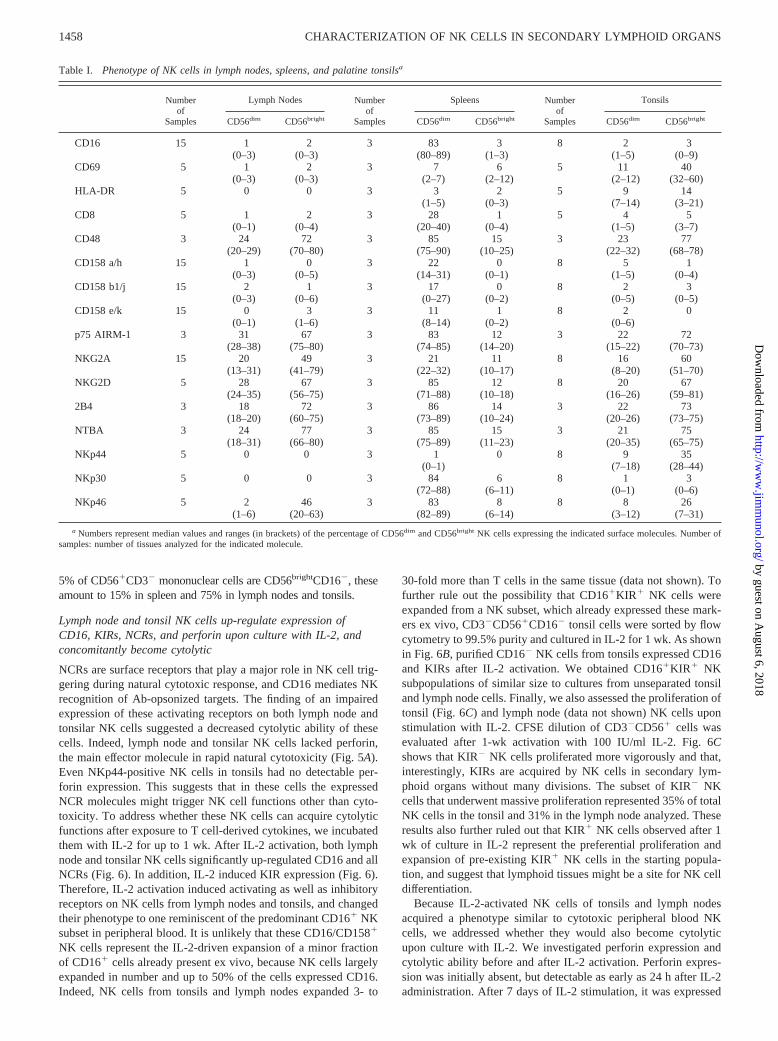
Table I. Phenotype of NK cells in lymph nodes, spleens, and palatine tonsilsa
Numberof
Samples
Lymph Nodes Numberof
Samples
Spleens Numberof
Samples
Tonsils
CD56dim CD56bright CD56dim CD56bright CD56dim CD56bright
CD16 15 1 2 3 83 3 8 2 3(0–3) (0–3) (80–89) (1–3) (1–5) (0–9)
CD69 5 1 2 3 7 6 5 11 40(0–3) (0–3) (2–7) (2–12) (2–12) (32–60)
HLA-DR 5 0 0 3 3 2 5 9 14(1–5) (0–3) (7–14) (3–21)
CD8 5 1 2 3 28 1 5 4 5(0–1) (0–4) (20–40) (0–4) (1–5) (3–7)
CD48 3 24 72 3 85 15 3 23 77(20–29) (70–80) (75–90) (10–25) (22–32) (68–78)
CD158 a/h 15 1 0 3 22 0 8 5 1(0–3) (0–5) (14–31) (0–1) (1–5) (0–4)
CD158 b1/j 15 2 1 3 17 0 8 2 3(0–3) (0–6) (0–27) (0–2) (0–5) (0–5)
CD158 e/k 15 0 3 3 11 1 8 2 0(0–1) (1–6) (8–14) (0–2) (0–6)
p75 AIRM-1 3 31 67 3 83 12 3 22 72(28–38) (75–80) (74–85) (14–20) (15–22) (70–73)
NKG2A 15 20 49 3 21 11 8 16 60(13–31) (41–79) (22–32) (10–17) (8–20) (51–70)
NKG2D 5 28 67 3 85 12 8 20 67(24–35) (56–75) (71–88) (10–18) (16–26) (59–81)
2B4 3 18 72 3 86 14 3 22 73(18–20) (60–75) (73–89) (10–24) (20–26) (73–75)
NTBA 3 24 77 3 85 15 3 21 75(18–31) (66–80) (75–89) (11–23) (20–35) (65–75)
NKp44 5 0 0 3 1 0 8 9 35(0–1) (7–18) (28–44)
NKp30 5 0 0 3 84 6 8 1 3(72–88) (6–11) (0–1) (0–6)
NKp46 5 2 46 3 83 8 8 8 26(1–6) (20–63) (82–89) (6–14) (3–12) (7–31)
a Numbers represent median values and ranges (in brackets) of the percentage of CD56dim and CD56bright NK cells expressing the indicated surface molecules. Number ofsamples: number of tissues analyzed for the indicated molecule.
1458 CHARACTERIZATION OF NK CELLS IN SECONDARY LYMPHOID ORGANS
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

in all tonsilar and lymph node NK cells (Fig.5A). IL-2-activatedtonsilar NK cells were also analyzed for their cytolytic capabilityagainst the classical HLA class I� NK cell target LCL 721.221 aswell as against CEM cells, a T lymphoblastoid cell line expressingthe NKG2D ligands ULBP1 and ULBP2. NK-mediated lysis ofCEM has been previously shown to be highly dependent onNKG2D (11), i.e., the single activating receptor consistently ex-pressed by all NK cells isolated from secondary lymphoid organs.To avoid allorecognition of CEM cells by tonsilar T cells,CD3�CD56� NK cells were sorted by flow cytometry before thecytotoxicity assays. Fig. 5, B and C, shows that IL-2-activatedtonsilar NK cells could exert a strong cytolytic activity, compara-ble to activated NK cells from blood. Freshly isolated tonsilar NKcells showed no detectable cytolytic function against both targetsat a 10:1 NK-target ratio (Fig. 5, B and C). To demonstrate that lowdoses of IL-2, which can be secreted by activated T cells, are ableto induce cytolytic properties and perforin expression of NK cellsin secondary lymphoid organs, tonsilar mononuclear cells werecultured with titrated doses of IL-2. Results shown in Fig. 7, A andB, show that as few as 5 IU/ml of IL-2 was sufficient to inducecytotoxic functions of lymphoid tissue NK cells.
In summary, our data indicate that NK cells of secondary lym-phoid organs can acquire cytolytic function and activating as wellas inhibitory receptors upon activation.
NK cells isolated from lymphoid tissues produce IFN-� afteractivation by IL-2 and IL-12
Peripheral blood CD56brightCD16� NK cells have been describedrecently to produce larger amounts of cytokines compared withtheir CD56lowCD16� counterpart (7). Thus, we investigatedwhether NK cells of secondary lymphoid organs could producecytokines such as IFN-�, early upon activation. As shown in Fig.7C, both IL-2 and IL-12 could promptly induce IFN-� productionby tonsil NK cells. As previously described for CD56highCD16�
NK cells, isolated from peripheral blood (7), slightly more NKcells produced IFN-� upon exposure to both cytokines comparedwith stimulations with the individual cytokines.
Therefore, while long-term (7-day) activation of NK cells insecondary lymphoid organs converts them to cytolytic effectors,short-term (1-day) activation stimulates the production of immu-nomodulatory cytokines by lymphoid tissue NK cells.
DiscussionIn this study we provide a phenotypic and functional characteriza-tion of NK cells located in secondary lymphoid organs, and dem-onstrate that they can acquire cytolytic functions as well as thecomplete NK cell receptor repertoire only upon activation.
We have analyzed human spleens, lymph nodes, and tonsils incomparison with peripheral blood and described four distinct NKsubsets resident in secondary lymphoid organs. Three of them,CD56brightNKG2A�NCR�, CD56brightNKG2A�NCR�, andCD56dimCD16�NCR� NK cells, have not previously been found,and one, CD56brightNKp44� NK cells, had not been isolated exvivo. In addition, we detected an overall enrichment ofCD56brightCD16� NK cells in spleen, lymph nodes, and tonsils.
We showed that tonsil and lymph node NK cells that acquire acomplete KIR phenotype in the presence of IL-2 undergo minimalcell divisions, while a large part, but not all, of KIR� NK cellsproliferated strongly. This further confirmed that different subsetsof NK cells are harbored in these lymphoid tissues and that at leastpart of these NK cells might be already committed to differentiateinto classical, KIR�, cytolytic, NK cells. Because peripheral bloodKIR� NK cells do not acquire KIRs and CD16 by culture in IL-2,NK cell localization to secondary lymphoid organs might be aprerequisite for maturation into the classical cytolytic NK pheno-type, found predominantly in blood.
When analyzed for effector functions, NK cells of secondarylymphoid organs were able to promptly produce IFN-�, but wereunable to kill target cells. The acquisition of cytolytic function byNK cells in secondary lymphoid tissue is accompanied by the de
FIGURE 3. Inhibitory receptors on NK cells of secondary lymphoidorgans. See legend to Fig. 2A.
FIGURE 4. Activating receptors on NK cells of secondary lymphoidorgans. See legend to Fig. 2A.
1459The Journal of Immunology
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

novo expression of molecules related to this function, i.e., activat-ing and inhibitory surface receptors. We showed in vitro that lowlevels of IL-2 render lymphoid tissue NK cells competent for tar-get cell killing. In vivo, this effect could be initiated by IL-2 se-cretion of T cells (25) because NK cells colocalize with T cells insecondary lymphoid organs (7). This would imply that lymph nodeand tonsil NK cells may become cytolytic only after T cell prim-ing. However, recent findings in mice suggest that dendritic cellscould also secrete IL-2 upon maturation (26) and might be a can-didate for NK-activating cells in lymph nodes and tonsils. Morestudies of DCs in secondary lymphoid organs are needed to assessthis possibility. NK cells in secondary lymphoid organs, whichbecome cytolytic upon activation, could target infected or tumorcells that have infiltrated these tissues. In addition, they mightmediate an editing function of APCs (27). We and others havepreviously demonstrated that immature dendritic cells can be lysedby NK cells (24, 28). Therefore, NK cells in lymph nodes andtonsils that become cytolytic after activation could eliminate in-
FIGURE 5. IL-2-treated NK cells of lymph nodes and tonsils up-regu-late perforin and subsequently are cytolytic. A, Cells from peripheral bloodor secondary lymphoid organs were analyzed for perforin expression.Analysis shown is performed on CD3-negative cells. Tonsilar NK cellswere stimulated with IL-2 for the indicated time points before analysis. Band C, Upon IL-2 culture, NK cells were greatly expanded in numbers, andtheir cytolytic capabilities against LCL721.221 cells (B) and CEM (C)were evaluated. Unseparated tonsil cells (B) or sorted tonsilar NK cells (C)were used. The indicated E:T ratios (e:LCL721.221 and e:CEM) were cal-culated on the number of NK cells added to the cytotoxicity assays. Num-bers inside the dot plots indicate the percentage of lysed cells (in the smallquadrant) at the indicated E:T ratios. Background (Spont.) and maximumTO-PRO-3 stainings (Total) were obtained by incubation with medium anddetergent, respectively. Freshly isolated tonsilar NK cells showed no de-tectable cytolytic function at a 10:1 E:T ratio (Fresh).
FIGURE 6. NK cells of lymph nodes and tonsils up-regulate CD16, NCRs,and KIRs upon IL-2 activation. A, Expression of CD16; the NCRs NKp30,NKp46, and NKp44; as well as the KIRs CD158a/h (p58.1/p50.1) andCD158b1/j (p58.2/p50.2) was analyzed on NK cells from tonsils and lymphnodes prior to and after IL-2 activation for 7 days. FSC, forward scatter. B,Phenotype of tonsilar NK cells sorted by flow cytometry and cultured for 1 wkin the presence of IL-2 (100 IU). CTRL, irrelevant isotype-matched mAb. Thenumbers inside the dot plots indicate the percentages of positive cells obtainedafter culture in the presence of IL-2. Analyses were performed gating onCD3�CD56� cells for both unsorted and sorted cell cultures. C, Proliferationof tonsilar NK cells was evaluated by CFSE dilution after 1 wk of stimulationwith IL-2. KIR expression was evaluated before (day 0) and after (day 7)culture with IL-2. KIRs, combined Abs against both CD158a/h andCD158b1/j; CTRL, irrelevant isotype-matched mAbs. Analyses shown wereperformed after four-color flow cytometry by gating on CD3�CD56� NKcells and, at day 7, also gating on B cells as a control for CFSE dilution. KIRsare acquired without many divisions by NK cells, while highly proliferatingNK cells remain KIR�. Similar results were obtained with cells isolated froma normal, not inflamed, lymph node.
1460 CHARACTERIZATION OF NK CELLS IN SECONDARY LYMPHOID ORGANS
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

completely matured dendritic cells that are responsible for periph-eral tolerance induction via stimulation of regulatory T cells orinduction of T cell deletion (29). Furthermore, infected dendriticcells could also be targeted, especially after CMV-mediated MHCclass I down-regulation (30). Both mechanisms, targeting of im-mature and virus-infected dendritic cells by cytolytic NK cells,would ensure Ag presentation in the immunogenic context of ma-ture dendritic cells.
NK cells in secondary lymphoid organs could also play a majorrole during in vivo IL-2 treatment aimed at enhancing patient re-sistance against malignancies. Draining lymph nodes are usuallythe first metastatic sites of a spreading cancer. The NK cells thatwe describe in these regions represent, altogether, a large pool ofeffector cells that can potentially target tumor cells and control
their lymphatic dissemination. Thus, it is tempting to speculate thatlymph node NK cells can acquire cytolytic function upon exoge-nous IL-2 administration. This could partially explain the benefi-cial effects observed in some patients treated with rIL-2 for ad-vanced cancers (31). Furthermore, low-dose IL-2 therapy results inrecruitment of CD56brightCD16� NK cells to peripheral blood,amounting to up to 70% of lymphocytes after 12 wk of treatment(32). These cells are not cycling and, therefore, probably representNK cells mobilized from extravascular tissues (33). The NK cellsof secondary lymphoid tissues, described in our study, could sig-nificantly contribute to this increase of peripheral blood NK cellsduring IL-2 therapy. Consistent with this hypothesis, we finddown-regulation of CCR7 on lymph node NK cells of one sampleafter IL-2 treatment (data not shown). Most NK cells in tonsils andlymph nodes, however, are already CCR7 and CD62L negativeafter isolation and, therefore, not anchored by these chemokinesand adhesion molecules in secondary lymphoid tissues. BothCCR7� and CCR7� NK cells in lymph nodes could, however, bemobilized to peripheral blood after IL-2 activation. An interestingpossibility is that therapy with s.c. injections of DCs may lead tothe activation of NK cells through the induction of IL-2-producingT cells or through the production of IL-2 and other cytokinesby DCs.
Because lymph nodes harbor 40% and peripheral blood only 2%of all 5 � 1011 lymphocytes in the human body (8, 9), lymph nodeNK cells outnumber peripheral blood NK cells by 10:1 (NK inblood, 10% (8); NK in lymph nodes, 5%). Taking into account thatNK cells from spleen (15% of total lymphocytes) and blood (2%of total lymphocytes) are similar, these organs together contain�1 � 1010 perforin� (95% in blood and 85% in spleen) and �2� 109 perforin� (5% in blood and 15% in spleen) NK cells.Lymph nodes alone contain �1 � 1010 perforin� NK cells, andother lymphoid organs such as tonsils also contribute to the per-forin� NK cell pool. Therefore, perforin� NK cells probably out-number perforin� classical NK cells in humans. NK activation by,for example, IL-2 is required to induce natural cytotoxicity in theseabundant NK cells of secondary lymphoid tissues.
It remains to be clarified whether the presence of such an abun-dant noncytolytic, but cytokine-secreting, NK cell subset in sec-ondary lymphoid organs might play relevant immunoregulatoryroles, for example, for T cell polarization, in the very early phasesof the immmune response.
In conclusion, we have characterized NK cells in the secondarylymphoid organs, spleen, lymph nodes, and tonsils. Althoughblood and spleen contain similar NK subsets, we found that tonsilsand lymph nodes contain mainly NKG2D�CD159a�CD158�
NCR� NK, which are different from the NK cells that predominatein blood. Upon IL-2 secretion by T cells (and possibly by dendriticcells), they up-regulate activating and inhibitory receptors as wellas perforin and acquire the ability to kill NK-sensitive targets.Cytolytic NK cells might control infected and tumor cells infil-trating secondary lymphoid organs, as well as edit APCs such asdendritic cells. The abundant NK compartment of lymph nodesand related tissues can be targeted in immune therapies, as shownwith IL-2 infusions, to mobilize its cytolytic capacity againsttumors.
AcknowledgmentsWe thank Ralph M. Steinman for critically reading the manuscript. Wegratefully acknowledge Svetlana Mazel and Tamara Shengelia for flowcytometric cell sorting.
References1. Trinchieri, G. 1989. Biology of natural killer cells. Adv. Immunol. 47:187.
FIGURE 7. Low doses of IL-2 are sufficient to induce perforin expres-sion, cytolytic activity, and IFN-� secretion by NK cells of secondarylymphoid organs. A and B, Perforin expression (A) and killing of NK sen-sible targets (B) by tonsilar NK cells were assessed after stimulation withthe indicated IL-2 concentrations (IU/ml). Perforin staining was performedby three-color flow cytometry. Perforin expression was analyzed forCD3�CD56� NK cells and CD3� T cells after 7 days of IL-2 stimulation(A). Cytotoxicity of tonsilar mononuclear cells with and without CD56�
cell depletion was evaluated after 7 days of IL-2 activation (B). Percentlysis of target cells LCL721.221 (221) and CEM (CEM). E:T ratios werecalculated on the number of NK or T cells (CD56�-depleted cultures),which were added to the cytotoxicity assays. The ratio displayed for CD56-depleted cultures represents the T cell to target ratio present in all assays(NK-T cell � 1:3.5 in IL-2-activated tonsil cultures). C, Tonsilar NK cellscan produce large amounts of IFN-� after stimulation for 24 h with 10ng/ml IL-12 (IL-12), 5 IU/ml IL-2 (IL-2), or both (IL-12 � IL-2). Analysiswas performed on CD3-negative cells. Numbers shown inside dot plotsindicate percentages of IFN-�-producing NK cells.
1461The Journal of Immunology
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

2. Bloom, B. R. 1982. Natural killers to rescue immune surveillance? Nature300:214.
3. Jacobs, R., G. Hintzen, A. Kemper, K. Beul, S. Kempf, G. Behrens,K. W. Sykora, and R. E. Schmidt. 2001. CD56bright cells differ in their KIRrepertoire and cytotoxic features from CD56dim NK cells. Eur. J. Immunol.31:3121.
4. Karre, K., H. G. Ljunggren, G. Piontek, and R. Kiessling. 1986. Selective rejec-tion of H-2-deficient lymphoma variants suggests alternative immune defensestrategy. Nature 319:675.
5. Cooper, M. A., T. A. Fehniger, S. C. Turner, K. S. Chen, B. A. Ghaheri,T. Ghayur, W. E. Carson, and M. A. Caligiuri. 2001. Human natural killer cells:a unique innate immunoregulatory role for the CD56bright subset. Blood 97:3146.
6. Campbell, J. J., S. Qin, D. Unutmaz, D. Soler, K. E. Murphy, M. R. Hodge,L. Wu, and E. C. Butcher. 2001. Unique subpopulations of CD56� NK and NK-Tperipheral blood lymphocytes identified by chemokine receptor expression rep-ertoire. J. Immunol. 166:6477.
7. Fehniger, T. A., M. A. Cooper, G. J. Nuovo, M. Cella, F. Facchetti, M. Colonna,and M. A. Caligiuri. 2003. CD56bright natural killer cells are present in humanlymph nodes and are activated by T cell derived IL-2: a potential new link be-tween adaptive and innate immunity. Blood 101:3052.
8. Westermann, J., and R. Pabst. 1992. Distribution of lymphocyte subsets andnatural killer cells in the human body. Clin. Invest. 70:539.
9. Trepel, F. 1974. Number and distribution of lymphocytes in man: a critical anal-ysis. Klin. Wochenschr. 52:511.
10. Shimizu, Y., and R. DeMars. 1989. Production of human cells expressing indi-vidual transferred HLA-A, -B, -C genes using an HLA-A, -B, -C null human cellline. J. Immunol. 142:3320.
11. Pende, D., P. Rivera, S. Marcenaro, C. C. Chang, R. Biassoni, R. Conte,M. Kubin, D. Cosman, S. Ferrone, L. Moretta, and A. Moretta. 2002. Majorhistocompatibility complex class I-related chain A and UL16-binding proteinexpression on tumor cell lines of different histotypes: analysis of tumor suscep-tibility to NKG2D-dependent natural killer cell cytotoxicity. Cancer Res.62:6178.
12. Lee-MacAry, A. E., E. L. Ross, D. Davies, R. Laylor, J. Honeychurch,M. J. Glennie, D. Snary, and R. W. Wilkinson. 2001. Development of a novelflow cytometric cell-mediated cytotoxicity assay using the fluorophores PKH-26and TO-PRO-3 iodide. J. Immunol. Methods 252:83.
13. Lanier, L. L., J. J. Ruitenberg, and J. H. Phillips. 1988. Functional and biochem-ical analysis of CD16 antigen on natural killer cells and granulocytes. J. Immunol.141:3478.
14. Perussia, B. 1998. Fc receptors on natural killer cells. Curr. Top. Microbiol.Immunol. 230:63.
15. Colonna, M., E. G. Brooks, M. Falco, G. B. Ferrara, and J. L. Strominger. 1993.Generation of allospecific natural killer cells by stimulation across a polymor-phism of HLA-C. Science 260:1121.
16. Braud, V. M., D. S. Allan, C. A. O’Callaghan, K. Soderstrom, A. D’Andrea,G. S. Ogg, S. Lazetic, N. T. Young, J. I. Bell, J. H. Phillips, et al. 1998. HLA-Ebinds to natural killer cell receptors CD94/NKG2A, B and C. Nature 391:795.
17. Braud, V., E. Y. Jones, and A. McMichael. 1997. The human major histocom-patibility complex class Ib molecule HLA-E binds signal sequence-derived pep-tides with primary anchor residues at positions 2 and 9. Eur. J. Immunol.27:1164.
18. Falco, M., R. Biassoni, C. Bottino, M. Vitale, S. Sivori, R. Augugliaro,L. Moretta, and A. Moretta. 1999. Identification and molecular cloning of p75/AIRM1, a novel member of the sialoadhesin family that functions as an inhibitoryreceptor in human natural killer cells. J. Exp. Med. 190:793.
19. Bauer, S., V. Groh, J. Wu, A. Steinle, J. H. Phillips, L. L. Lanier, and T. Spies.1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285:727.
20. Cosman, D., J. Mullberg, C. L. Sutherland, W. Chin, R. Armitage, W. Fanslow,M. Kubin, and N. J. Chalupny. 2001. ULBPs, novel MHC class I-related mole-cules, bind to CMV glycoprotein UL16 and stimulate NK cytotoxicity throughthe NKG2D receptor. Immunity 14:123.
21. Parolini, S., C. Bottino, M. Falco, R. Augugliaro, S. Giliani, R. Franceschini,H. D. Ochs, H. Wolf, J. Y. Bonnefoy, R. Biassoni, et al. 2000. X-linked lym-phoproliferative disease: 2B4 molecules displaying inhibitory rather than acti-vating function are responsible for the inability of natural killer cells to killEpstein-Barr virus-infected cells. J. Exp. Med. 192:337.
22. Bottino, C., M. Falco, S. Parolini, E. Marcenaro, R. Augugliaro, S. Sivori,E. Landi, R. Biassoni, L. D. Notarangelo, L. Moretta, and A. Moretta. 2001.NTB-A, a novel SH2D1A-associated surface molecule contributing to the inabil-ity of natural killer cells to kill Epstein-Barr virus-infected B cells in X-linkedlymphoproliferative disease. J. Exp. Med. 194:235.
23. Moretta, A., C. Bottino, M. Vitale, D. Pende, C. Cantoni, M. C. Mingari,R. Biassoni, and L. Moretta. 2001. Activating receptors and coreceptors involvedin human natural killer cell-mediated cytolysis. Annu. Rev. Immunol. 19:197.
24. Ferlazzo, G., M. L. Tsang, L. Moretta, G. Melioli, R. M. Steinman, and C. Munz.2002. Human dendritic cells activate resting NK cells and are recognized via theNKp30 receptor by activated NK cells. J. Exp. Med. 195:343.
25. Watson, J., and D. Mochizuki. 1980. Interleukin 2: a class of T cell growthfactors. Immunol. Rev. 51:257.
26. Granucci, F., C. Vizzardelli, N. Pavelka, S. Feau, M. Persico, E. Virzi,M. Rescigno, G. Moro, and P. Ricciardi-Castagnoli. 2001. Inducible IL-2 pro-duction by dendritic cells revealed by global gene expression analysis. Nat. Im-mun. 2:882.
27. Moretta, A. 2002. Natural killer cells and dendritic cells: rendezvous in abusedtissues. Nat. Rev. Immunol. 2:957.
28. Piccioli, D., S. Sbrana, E. Melandri, and N. M. Valiante. 2002. Contact-depen-dent stimulation and inhibition of dendritic cells by natural killer cells. J. Exp.Med. 195:335.
29. Steinman, R. M., D. Hawiger, and M. C. Nussenzweig. 2003. Tolerogenic den-dritic cells. Annu. Rev. Immunol. 21:685.
30. Raftery, M. J., M. Schwab, S. M. Eibert, Y. Samstag, H. Walczak, and G. Schon-rich. 2001. Targeting the function of mature dendritic cells by human cytomeg-alovirus: a multilayered viral defense strategy. Immunity 15:997.
31. Rosenberg, S. A. 2001. Progress in human tumor immunology and immunother-apy. Nature 411:380.
32. Caligiuri, M. A., C. Murray, M. J. Robertson, E. Wang, K. Cochran, C. Cameron,P. Schow, M. E. Ross, T. R. Klumpp, R. J. Soiffer, et al. 1993. Selective mod-ulation of human natural killer cells in vivo after prolonged infusion of low doserecombinant interleukin 2. J. Clin. Invest. 91:123.
33. Fehniger, T. A., E. M. Bluman, M. M. Porter, E. Mrozek, M. A. Cooper,J. B. VanDeusen, S. R. Frankel, W. Stock, and M. A. Caligiuri. 2000. Potentialmechanisms of human natural killer cell expansion in vivo during low-dose IL-2therapy. J. Clin. Invest. 106:117.
1462 CHARACTERIZATION OF NK CELLS IN SECONDARY LYMPHOID ORGANS
by guest on August 6, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















