
Temporally controlled targeted somatic mutagenesis in skeletal muscles of the mouse
6
LETTER Temporally Controlled Targeted Somatic Mutagenesis in Skeletal Muscles of the Mouse Michael Schuler, 1,2 Faisal Ali, 1 Elisabeth Metzger, 1 Pierre Chambon, 1,2 * and Daniel Metzger 1,2 * 1 Institut de Ge ´ ne ´ tique et de Biologie Mole ´ culaire et Cellulaire (IGBMC), Centre National de la Recherche Scientifique, Institut National de la Sante ´ et de la Recherche Me ´ dicale, Universite ´ Louis Pasteur, Colle ` ge de France, Illkirch-Cedex, France 2 Institut Clinique de la Souris (ICS), Illkirch-Cedex, France Received 4 January 2005; Accepted 8 January 2005 Summary: To generate temporally controlled targeted somatic mutations selectively and efficiently in skeletal muscles, we established a transgenic HSA-Cre-ER T2 mouse line in which the expression of the tamoxifen- dependent Cre-ER T2 recombinase is under the control of a large genomic DNA segment of the human skeletal muscle a-actin gene, contained in a P1-derived artificial chromosome. In this transgenic line Cre-ER T2 is selec- tively expressed in skeletal muscles, and Cre-ER T2 - mediated alteration of LoxP flanked (floxed) target genes is skeletal muscle-specific and strictly tamoxifen- dependent. HSA-Cre-ER T2 mice should be of great value to analyze gene function in skeletal muscles, and to establish animal models of human skeletal muscle dis- orders. genesis 41:165–170, 2005. c 2005 Wiley-Liss, Inc. Key words: skeletal muscle a-actin; cre/LoxP technology; Cre-ER T2 recombinase; tamoxifen; skeletal muscle Skeletal muscle is the most abundant tissue type in humans (Janssen et al., 2000). It is composed of highly dif- ferentiated myotubes, whose major function is to gener- ate motile force through contraction. The functional relevance of a variety of genes during myotube formation and homeostasis has been partially elucidated in germline knockout mice, revealing either impaired skeletal muscle development (Arnold and Braun, 2000; Buckingham et al., 2003; Cossu and Borello, 1999) or impaired skeletal muscle energy homeostasis (Kahn, 1994; Mauvais-Jarvis et al., 2002), and also providing mouse models for human muscular dystrophies (Allamand and Campbell, 2000; Buj-Bello et al., 2002; Cohn and Campbell, 2000). How- ever, germline null mutations often induce pre- or postna- tal lethality (Kastner et al., 1994; Branda and Dymecki, 2004; Metzger and Chambon, 2001), thus precluding the analysis of gene functions at later stages. Furthermore, as germline mutations do not discriminate between cell autonomous and noncell autonomous actions (Metzger and Chambon, 2001), it is difficult to determine the skele- tal muscle-specific function of widely expressed genes. To circumvent these problems, targeted somatic mutagenesis systems based on tissue- or cell-specific expression of the bacteriophage P1 Cre recombinase, which excises LoxP- flanked (floxed) DNA segments, have been developed (Lakso et al., 1992; Gu et al., 1994; Hennet et al., 1995). Several mouse lines with muscle-specific Cre expression have been established. Transgenic mice, expressing the Cre recombinase under the control of the muscle creatine kinase (MCK) promoter, excise floxed DNA segments in both skeletal and cardiac muscle (Bruning et al., 1998). When Cre expression was controlled by a 2.2-kb genomic segment of the human skeletal muscle a-actin (HSA) pro- moter, efficient recombination occurred mainly in skeletal muscles, but some recombination was also induced in car- diac muscle (Miniou et al., 1999; and our unpubl. obs.). Skeletal muscle-restricted Cre expression was obtained by inserting the Cre recombinase in the MLC1f locus (Bothe et al., 2000). However, none of these lines allowed skele- tal muscle-restricted, temporally controlled somatic muta- genesis. Temporal control of Cre activity was achieved in the mouse by fusing the Cre recombinase with mutated ligand binding domains of steroid receptors (Feil et al., 1996; Kellendonk et al., 1999; Schwenk et al., 1998; Tannour-Louet et al., 2002). Using transgenes with short promoter segments, cell-specific expression of chimeric proteins between Cre and mutated ligand binding domains of the human estrogen receptor a (ERa) (e.g., Cre-ER T s), whose activity is induced by the anti-estrogen tamoxifen (Tam), has allowed efficient spatiotemporally controlled somatic mutagenesis of floxed target genes in * Correspondence to: Daniel Metzger or Pierre Chambon, IGBMC and ICS, BP10142, 1 rue Laurent Fries, 67404 Illkirch-Cedex, France. E-mail: [email protected] or [email protected] Contract grant sponsors: Marie Curie Individual Fellowship (to M.S.), Boehringer Ingelheim Fellowship (to F.A.), Centre National de la Recherche Scientifique, Institut National de la Sante ´ et de la Recherche Me ´dicale, Colle `ge de France, Ho ˆpital Universitaire de Strasbourg, Association pour la Recherche sur le Cancer, Fondation pour la Recherche Me ´dicale, Human Frontier Science Program, Ministe `re de l’Education Nationale de la Recherche et de la Technologie. Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/gene.20107 ' 2005 Wiley-Liss, Inc. genesis 41:165–170 (2005)
-
Upload
michael-schuler -
Category
Documents
-
view
214 -
download
2
Transcript of Temporally controlled targeted somatic mutagenesis in skeletal muscles of the mouse

LETTER
Temporally Controlled Targeted Somatic Mutagenesisin Skeletal Muscles of the MouseMichael Schuler,1,2 Faisal Ali,1 Elisabeth Metzger,1 Pierre Chambon,1,2* and Daniel Metzger1,2*1Institut de Genetique et de Biologie Moleculaire et Cellulaire (IGBMC), Centre National de la Recherche Scientifique,Institut National de la Sante et de la Recherche Medicale, Universite Louis Pasteur, College de France,Illkirch-Cedex, France2Institut Clinique de la Souris (ICS), Illkirch-Cedex, France
Received 4 January 2005; Accepted 8 January 2005
Summary: To generate temporally controlled targetedsomatic mutations selectively and efficiently in skeletalmuscles, we established a transgenic HSA-Cre-ERT2
mouse line in which the expression of the tamoxifen-dependent Cre-ERT2 recombinase is under the controlof a large genomic DNA segment of the human skeletalmuscle a-actin gene, contained in a P1-derived artificialchromosome. In this transgenic line Cre-ERT2 is selec-tively expressed in skeletal muscles, and Cre-ERT2-mediated alteration of LoxP flanked (floxed) targetgenes is skeletal muscle-specific and strictly tamoxifen-dependent. HSA-Cre-ERT2 mice should be of great valueto analyze gene function in skeletal muscles, and toestablish animal models of human skeletal muscle dis-orders. genesis 41:165–170, 2005. �c 2005 Wiley-Liss, Inc.
Key words: skeletal muscle a-actin; cre/LoxP technology;Cre-ERT2 recombinase; tamoxifen; skeletal muscle
Skeletal muscle is the most abundant tissue type inhumans (Janssen et al., 2000). It is composed of highly dif-ferentiated myotubes, whose major function is to gener-ate motile force through contraction. The functionalrelevance of a variety of genes during myotube formationand homeostasis has been partially elucidated in germlineknockout mice, revealing either impaired skeletal muscledevelopment (Arnold and Braun, 2000; Buckinghamet al., 2003; Cossu and Borello, 1999) or impaired skeletalmuscle energy homeostasis (Kahn, 1994; Mauvais-Jarviset al., 2002), and also providing mouse models for humanmuscular dystrophies (Allamand and Campbell, 2000;Buj-Bello et al., 2002; Cohn and Campbell, 2000). How-ever, germline null mutations often induce pre- or postna-tal lethality (Kastner et al., 1994; Branda and Dymecki,2004; Metzger and Chambon, 2001), thus precluding theanalysis of gene functions at later stages. Furthermore, asgermline mutations do not discriminate between cellautonomous and noncell autonomous actions (Metzgerand Chambon, 2001), it is difficult to determine the skele-tal muscle-specific function of widely expressed genes. Tocircumvent these problems, targeted somatic mutagenesis
systems based on tissue- or cell-specific expression of thebacteriophage P1 Cre recombinase, which excises LoxP-flanked (floxed) DNA segments, have been developed(Lakso et al., 1992; Gu et al., 1994; Hennet et al., 1995).Several mouse lines with muscle-specific Cre expressionhave been established. Transgenic mice, expressing theCre recombinase under the control of the muscle creatinekinase (MCK) promoter, excise floxed DNA segments inboth skeletal and cardiac muscle (Bruning et al., 1998).When Cre expression was controlled by a 2.2-kb genomicsegment of the human skeletal muscle a-actin (HSA) pro-moter, efficient recombination occurred mainly in skeletalmuscles, but some recombination was also induced in car-diac muscle (Miniou et al., 1999; and our unpubl. obs.).Skeletal muscle-restricted Cre expression was obtained byinserting the Cre recombinase in the MLC1f locus (Botheet al., 2000). However, none of these lines allowed skele-tal muscle-restricted, temporally controlled somatic muta-genesis. Temporal control of Cre activity was achieved inthe mouse by fusing the Cre recombinase with mutatedligand binding domains of steroid receptors (Feil et al.,1996; Kellendonk et al., 1999; Schwenk et al., 1998;Tannour-Louet et al., 2002). Using transgenes with shortpromoter segments, cell-specific expression of chimericproteins between Cre and mutated ligand bindingdomains of the human estrogen receptor a (ERa) (e.g.,Cre-ERTs), whose activity is induced by the anti-estrogentamoxifen (Tam), has allowed efficient spatiotemporallycontrolled somatic mutagenesis of floxed target genes in
* Correspondence to: Daniel Metzger or Pierre Chambon, IGBMC and
ICS, BP10142, 1 rue Laurent Fries, 67404 Illkirch-Cedex, France.
E-mail: [email protected] or [email protected]
Contract grant sponsors: Marie Curie Individual Fellowship (to M.S.),
Boehringer Ingelheim Fellowship (to F.A.), Centre National de la Recherche
Scientifique, Institut National de la Sante et de la Recherche Medicale,
College de France, Hopital Universitaire de Strasbourg, Association pour la
Recherche sur le Cancer, Fondation pour la Recherche Medicale, Human
Frontier Science Program, Ministere de l’Education Nationale de la
Recherche et de la Technologie.Published online in
Wiley InterScience (www.interscience.wiley.com).
DOI: 10.1002/gene.20107
' 2005 Wiley-Liss, Inc. genesis 41:165–170 (2005)
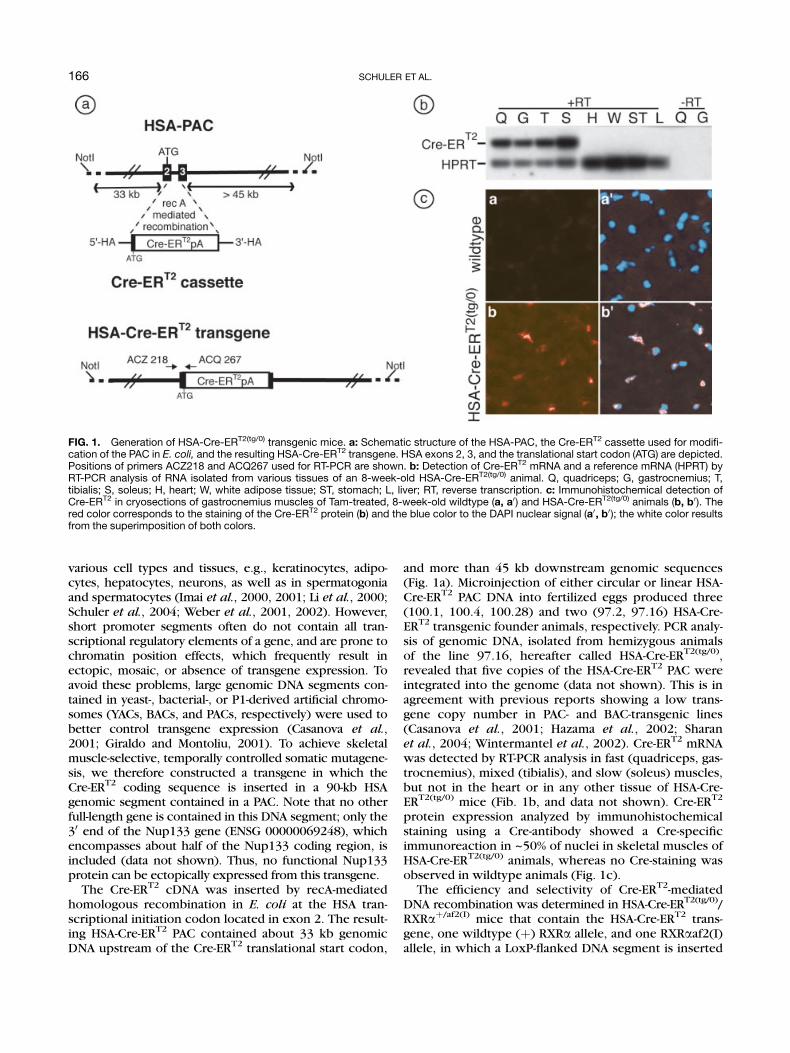
various cell types and tissues, e.g., keratinocytes, adipo-cytes, hepatocytes, neurons, as well as in spermatogoniaand spermatocytes (Imai et al., 2000, 2001; Li et al., 2000;Schuler et al., 2004; Weber et al., 2001, 2002). However,short promoter segments often do not contain all tran-scriptional regulatory elements of a gene, and are prone tochromatin position effects, which frequently result inectopic, mosaic, or absence of transgene expression. Toavoid these problems, large genomic DNA segments con-tained in yeast-, bacterial-, or P1-derived artificial chromo-somes (YACs, BACs, and PACs, respectively) were used tobetter control transgene expression (Casanova et al.,2001; Giraldo and Montoliu, 2001). To achieve skeletalmuscle-selective, temporally controlled somatic mutagene-sis, we therefore constructed a transgene in which theCre-ERT2 coding sequence is inserted in a 90-kb HSAgenomic segment contained in a PAC. Note that no otherfull-length gene is contained in this DNA segment; only the30 end of the Nup133 gene (ENSG 00000069248), whichencompasses about half of the Nup133 coding region, isincluded (data not shown). Thus, no functional Nup133protein can be ectopically expressed from this transgene.
The Cre-ERT2 cDNA was inserted by recA-mediatedhomologous recombination in E. coli at the HSA tran-scriptional initiation codon located in exon 2. The result-ing HSA-Cre-ERT2 PAC contained about 33 kb genomicDNA upstream of the Cre-ERT2 translational start codon,
and more than 45 kb downstream genomic sequences(Fig. 1a). Microinjection of either circular or linear HSA-Cre-ERT2 PAC DNA into fertilized eggs produced three(100.1, 100.4, 100.28) and two (97.2, 97.16) HSA-Cre-ERT2 transgenic founder animals, respectively. PCR analy-sis of genomic DNA, isolated from hemizygous animalsof the line 97.16, hereafter called HSA-Cre-ERT2(tg/0),revealed that five copies of the HSA-Cre-ERT2 PAC wereintegrated into the genome (data not shown). This is inagreement with previous reports showing a low trans-gene copy number in PAC- and BAC-transgenic lines(Casanova et al., 2001; Hazama et al., 2002; Sharanet al., 2004; Wintermantel et al., 2002). Cre-ERT2 mRNAwas detected by RT-PCR analysis in fast (quadriceps, gas-trocnemius), mixed (tibialis), and slow (soleus) muscles,but not in the heart or in any other tissue of HSA-Cre-ERT2(tg/0) mice (Fib. 1b, and data not shown). Cre-ERT2
protein expression analyzed by immunohistochemicalstaining using a Cre-antibody showed a Cre-specificimmunoreaction in ~50% of nuclei in skeletal muscles ofHSA-Cre-ERT2(tg/0) animals, whereas no Cre-staining wasobserved in wildtype animals (Fig. 1c).
The efficiency and selectivity of Cre-ERT2-mediatedDNA recombination was determined in HSA-Cre-ERT2(tg/0)/RXRaþ/af2(I) mice that contain the HSA-Cre-ERT2 trans-gene, one wildtype (þ) RXRa allele, and one RXRaaf2(I)allele, in which a LoxP-flanked DNA segment is inserted
FIG. 1. Generation of HSA-Cre-ERT2(tg/0) transgenic mice. a: Schematic structure of the HSA-PAC, the Cre-ERT2 cassette used for modifi-cation of the PAC in E. coli, and the resulting HSA-Cre-ERT2 transgene. HSA exons 2, 3, and the translational start codon (ATG) are depicted.Positions of primers ACZ218 and ACQ267 used for RT-PCR are shown. b: Detection of Cre-ERT2 mRNA and a reference mRNA (HPRT) byRT-PCR analysis of RNA isolated from various tissues of an 8-week-old HSA-Cre-ERT2(tg/0) animal. Q, quadriceps; G, gastrocnemius; T,tibialis; S, soleus; H, heart; W, white adipose tissue; ST, stomach; L, liver; RT, reverse transcription. c: Immunohistochemical detection ofCre-ERT2 in cryosections of gastrocnemius muscles of Tam-treated, 8-week-old wildtype (a, a0) and HSA-Cre-ERT2(tg/0) animals (b, b0). Thered color corresponds to the staining of the Cre-ERT2 protein (b) and the blue color to the DAPI nuclear signal (a0, b0); the white color resultsfrom the superimposition of both colors.
166 SCHULER ET AL.

in the RXRa locus (Mascrez et al., 1998). Scanning ofSouthern blots showed that ~50% of the floxedRXRaaf2(I) alleles were converted into RXRaaf2(II)alleles in skeletal muscles of Tam-treated (1 mg for 5 con-secutive days) HSA-Cre-ERT2(tg/0)/RXRaþ/af2(I) animals. Incontrast, no excision of the floxed RXRaaf2(I) allele wasdetected in any other tissue of Tam-treated animals,including heart and visceral (stomach) muscle, or in anytissue of oil-treated HSA-Cre-ERT2(tg/0)/RXRaþ/af2(I) ani-mals (Fig. 2-I, and data not shown). A more sensitive, butnonquantitative PCR-based assay followed by Southernblot hybridization confirmed that floxed DNA excisionoccurred exclusively in skeletal muscles of theTam-treated animal (Fig. 2-II, -III), thus demonstratingthat Cre-ERT2 is selectively expressed in skeletal musclesand that its activity is tightly regulated by Tam. Similarrecombination efficiencies were observed after ad-ministration of 0.1 mg Tam for 5 consecutive days (datanot shown). Recombination efficiencies of 50% inskeletal muscle were previously reported in MCK-Cre(Andrechek et al., 2002; Bruning et al., 1998; Grobetet al., 2003), MLC1f-Cre (Bothe et al., 2000), and HSA-Cre animals (Buj-Bello et al., 2002; Miniou et al., 1999).As the average number of myonuclei within skeletalmuscles accounts for ~50% of the total number of nuclei(Schmalbruch and Hellhammer, 1977), the above resultsindicate that Tam-dependent recombination occurred inalmost all myonuclei.
To analyze the recombinase activity at the cellularlevel, HSA-Cre-ERT2(tg/0) mice were bred with ACZLtg/0
Cre-reporter animals, in which expression of b-galactosi-dase is induced after Cre-mediated recombination (Akagiet al., 1997). In oil-treated, double transgenic HSA-Cre-ERT2(tg/0)/ACZLtg/0 mice no X-gal staining was observed(Fig. 3, oil), confirming that the Cre-ERT2 recombinaseactivity is strictly Tam-dependent. After Tam-treatment,
b-galactosidase activity was readily detected in all skele-tal muscle fibers, whereas no Cre-ERT2 mediated recom-bination was detected in heart muscle or in visceralmuscle of the stomach (Fig. 3, Tam).
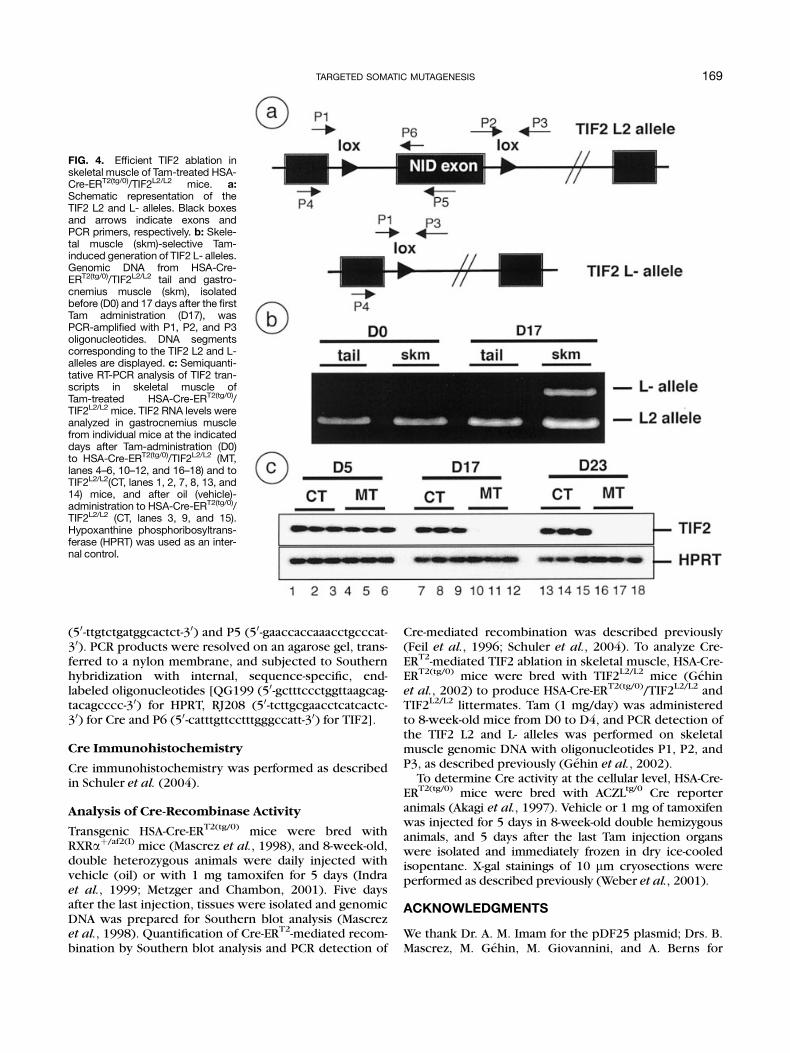
To demonstrate that genes can be efficiently ablatedin skeletal muscles in a temporally controlled manner,HSA-Cre-ERT2(tg/0) mice were bred with mice bearingfloxed transcriptional intermediary factor 2 (TIF2) L2alleles (TIF2L2/L2 mice, Gehin et al., 2002), to obtainHSA-Cre-ERT2(tg/0)/TIF2L2/L2 premutant mice, as well asTIF2L2/L2 control littermates. In agreement with theresults described above (Fig. 2), Tam administration(D0–D4) to premutant mice induced the conversion of~50% of the floxed TIF2 L2 alleles into L- alleles in skele-tal muscle (Fig. 4b, and data not shown). As expected,skeletal muscle TIF2 mRNA levels were similar invehicle-treated premutant mice and in Tam-treatedTIF2L2/L2 mice at D0, D17, and D23 (Fig. 4c). However,whereas TIF2 transcripts were not significantlydecreased in skeletal muscle of Tam-treated premutantmice at D5, they were reduced by more than 10-fold atD17, and were almost undetectable by D23, thus demon-strating that Tam efficiently induced Cre-ERT2-mediatedTIF2 ablation in skeletal muscle.
Taken together, the present results establish that theHSA-Cre-ERT2 mouse line allows efficient, temporallycontrolled ablation of floxed genes in a skeletal muscle-restricted manner. Similar recombination specificitiesand efficiencies were observed in the other HSA-Cre-ERT2 lines (100.1, 100.4, 100.28, and 97.2; data notshown), indicating that PAC-transgenes are reproduciblyexpressed in transgenic mice using either circular or lin-ear transgene DNA. In contrast, out of five transgenicmouse lines in which the 2.2-kb HSA promoter wasused to control Cre-ERT2 expression, only two linesexpressed Cre-ERT2 in skeletal muscle, and these lines
FIG. 2. Cre-ERT2-mediated excision of floxed DNA. Analysis of genomic DNA extracted from the indicated tissues of 8-week-old HSA-Cre-ERT2(tg/0)/RXR�þ/af2(I) animals treated either with oil or Tam. (I) Southern blot analysis of the wildtype [þ], floxed [af2(I)] and excised[af2(II)] RXR� alleles. (II) Ethidium bromide-stained agarose gel of PCR products of wildtype and excised af2(II) RXR� alleles. (III) Selectivedetection of the RXR�af2(II) allele after Southern hybridization of the PCR products shown in (II) with a radiolabeled loxP-specificoligonucleotide.
167TARGETED SOMATIC MUTAGENESIS

also expressed Cre-ERT2 in cardiomyocytes (data notshown). Thus, the use of large genomic DNA fragmentsis clearly recommended to obtain predictable transgeneexpression.
In summary, we established mouse lines in which Cre-ERT2 expression is controlled by HSA genomic sequencescontained in a 90 kb PAC clone. These HSA-Cre-ERT2
transgenic mouse lines allow efficient temporally con-trolled activation or ablation of floxed genes selectively inskeletal muscle. They should be useful tools to study genefunction in skeletal muscles, and to establish animalsmodels for muscular dystrophies or metabolic diseases.
MATERIALS AND METHODS
Generation of Transgenic HSA-Cre-ERT2 Mice
E. coli DH10B containing the recombinant HSA-PAC(no. 26786) was obtained after a custom library screen(RPC-1 human PAC library, K. Osoegawa and P. de Jong,Roswell Park Cancer Institute) performed at IncyteGenomics (St. Louis, MO) using the primers: 50-ccaagc-ccaccactccacacgactc-30 and 50-tcccctcccaactcctgccctacc-t-30. Exons 2 and 3 of the HSA gene were partiallyreplaced by the Cre-ERT2 coding sequence and thebovine PGK polyadenylation site by recA-based recombi-nation in E. coli (Imam et al., 2000). Briefly, the 50 and 30homology arms (HA) were amplified by PCR from HeLacell genomic DNA with the primers ABi 248 (50-aaaaag-gaggcaagacagtc-30), ABi 249 (50-cccacgatggacgggaacac-30) and ABi 12 (50-cggtatgggtcagaaagatt-30), ABi 13 (50-gaggcgtagagggacagcac-30), respectively. The amplified 50-HA (827 bp) and 30-HA (282 bp) were inserted in theEcoRI and PmeI restriction sites of pBVCre-ERT2 (Schuleret al., 2004) located upstream and downstream of theCre-ERT2 coding sequence, respectively. The 3.6-kbblunt-ended SalI DNA segment of this plasmid wascloned into the HindIII digested, blunt-ended pDF25plasmid (Imam et al., 2000), resulting in pDF-HSA-Cre-ERT2. HSA-PAC containing E. coli were transformed withpDF-HSA-Cre-ERT2 and integration of pDF-HSA-Cre-ERT2
into the PAC was selected by growth on agar plates sup-plemented with kanamycin and chloramphenicol(25 mg/ml, each) at 438C. Targeted clones were grownon agar plates containing kanamycin (25 mg/ml) andstreptomycin (200 mg/ml) at 438C, to select for the lossof the pDF vector DNA (Imam et al., 2000). After verifi-cation by Southern blot analysis, HSA-Cre-ERT2-PAC DNAwas digested by NotI and fractionated on a 15% sucrosegradient. Two picoliters of the purified, linear 90 kbHSA-Cre-ERT2 NotI-DNA segment or the circular HSA-Cre-ERT2 DNA were microinjected into fertilized eggs ata concentration of 2 ng/ml to generate transgenic mice(Feil et al., 1996). Breeding and maintenance of micewere performed according to institutional guidelines.Transgenic HSA-Cre-ERT2 animals were identified bySouthern blot analysis of genomic DNA isolated from tailbiopsies, digested by BamHI, and hybridized after gelelectrophoresis with a radiolabeled Cre-specific probe(Weber et al., 2001).
RT-PCR Analysis
Total RNA was isolated using RNA-Solv Reagent (OmegaBiotek, Doraville, GA). Five mg of total RNA were con-verted to cDNA with SuperScript II reverse transcriptase(Invitrogen Life Technologies, La Jolla, CA). Two ml ofthe reverse transcriptase reaction (1:20 diluted) wereamplified for 25 cycles with 558C annealing tempera-tures and the following primers : HPRT, QG197 (50-gtaat-gatcagtcaacgggggac-30) and QG198 (50-ccagcaagcttg-caaccttaacca-30); Cre, ACZ218 (50-gagccgagagtagcagttgt-30) and ACQ267 (50-ggcaaacggacagaagcatt-30), TIF2, P4
FIG. 3. X-Gal staining of 10-mm cryosections of fast (quadriceps),mixed (tibialis), heart and visceral (stomach) muscles isolated fromHSA-Cre-ERT2(tg/0)/AZCLtg/0 animals, 5 days after the last oil or Taminjection. Scale bar ¼ 50 mm.
168 SCHULER ET AL.
(50-ttgtctgatggcactct-30) and P5 (50-gaaccaccaaacctgcccat-30). PCR products were resolved on an agarose gel, trans-ferred to a nylon membrane, and subjected to Southernhybridization with internal, sequence-specific, end-labeled oligonucleotides [QG199 (50-gctttccctggttaagcag-tacagcccc-30) for HPRT, RJ208 (50-tcttgcgaacctcatcactc-30) for Cre and P6 (50-catttgttcctttgggccatt-30) for TIF2].
Cre Immunohistochemistry
Cre immunohistochemistry was performed as describedin Schuler et al. (2004).
Analysis of Cre-Recombinase Activity
Transgenic HSA-Cre-ERT2(tg/0) mice were bred withRXRaþ/af2(I) mice (Mascrez et al., 1998), and 8-week-old,double heterozygous animals were daily injected withvehicle (oil) or with 1 mg tamoxifen for 5 days (Indraet al., 1999; Metzger and Chambon, 2001). Five daysafter the last injection, tissues were isolated and genomicDNA was prepared for Southern blot analysis (Mascrezet al., 1998). Quantification of Cre-ERT2-mediated recom-bination by Southern blot analysis and PCR detection of
Cre-mediated recombination was described previously(Feil et al., 1996; Schuler et al., 2004). To analyze Cre-ERT2-mediated TIF2 ablation in skeletal muscle, HSA-Cre-ERT2(tg/0) mice were bred with TIF2L2/L2 mice (Gehinet al., 2002) to produce HSA-Cre-ERT2(tg/0)/TIF2L2/L2 andTIF2L2/L2 littermates. Tam (1 mg/day) was administeredto 8-week-old mice from D0 to D4, and PCR detection ofthe TIF2 L2 and L- alleles was performed on skeletalmuscle genomic DNA with oligonucleotides P1, P2, andP3, as described previously (Gehin et al., 2002).
To determine Cre activity at the cellular level, HSA-Cre-ERT2(tg/0) mice were bred with ACZLtg/0 Cre reporteranimals (Akagi et al., 1997). Vehicle or 1 mg of tamoxifenwas injected for 5 days in 8-week-old double hemizygousanimals, and 5 days after the last Tam injection organswere isolated and immediately frozen in dry ice-cooledisopentane. X-gal stainings of 10 mm cryosections wereperformed as described previously (Weber et al., 2001).
ACKNOWLEDGMENTS
We thank Dr. A. M. Imam for the pDF25 plasmid; Drs. B.Mascrez, M. Gehin, M. Giovannini, and A. Berns for
FIG. 4. Efficient TIF2 ablation inskeletal muscle of Tam-treated HSA-Cre-ERT2(tg/0)/TIF2L2/L2 mice. a:Schematic representation of theTIF2 L2 and L- alleles. Black boxesand arrows indicate exons andPCR primers, respectively. b: Skele-tal muscle (skm)-selective Tam-induced generation of TIF2 L- alleles.Genomic DNA from HSA-Cre-ERT2(tg/0)/TIF2L2/L2 tail and gastro-cnemius muscle (skm), isolatedbefore (D0) and 17 days after the firstTam administration (D17), wasPCR-amplified with P1, P2, and P3oligonucleotides. DNA segmentscorresponding to the TIF2 L2 and L-alleles are displayed. c: Semiquanti-tative RT-PCR analysis of TIF2 tran-scripts in skeletal muscle ofTam-treated HSA-Cre-ERT2(tg/0)/TIF2L2/L2 mice. TIF2 RNA levels wereanalyzed in gastrocnemius musclefrom individual mice at the indicateddays after Tam-administration (D0)to HSA-Cre-ERT2(tg/0)/TIF2L2/L2 (MT,lanes 4–6, 10–12, and 16–18) and toTIF2L2/L2(CT, lanes 1, 2, 7, 8, 13, and14) mice, and after oil (vehicle)-administration to HSA-Cre-ERT2(tg/0)/TIF2L2/L2 (CT, lanes 3, 9, and 15).Hypoxanthine phosphoribosyltrans-ferase (HPRT) was used as an inter-nal control.
169TARGETED SOMATIC MUTAGENESIS

mice; Drs. E. Greiner and G. Schutz for helpful discus-sions; B. Chofflet, P. Andre, and the animal facility stafffor excellent technical assistance; and the secretary fortyping the manuscript.
LITERATURE CITED
Akagi K, Sandig V, Vooijs M, Van der Valk M, Giovannini M, Strauss M,Berns A. 1997. Cre-mediated somatic site-specific recombinationin mice. Nucleic Acids Res 25:1766–1773.
Allamand V, Campbell KP. 2000. Animal models for muscular dystro-phy: valuable tools for the development of therapies. Hum MolGenet 9:2459–2467.
Andrechek ER, Hardy WR, Girgis-Gabardo AA, Perry RL, Butler R,Graham FL, Kahn RC, Rudnicki MA, Muller WJ. 2002. ErbB2 isrequired for muscle spindle and myoblast cell survival. Mol CellBiol 22:4714–4722.
Arnold HH, Braun T. 2000. Genetics of muscle determination anddevelopment. Curr Top Dev Biol 48:129–164.
Bothe GW, Haspel JA, Smith CL, Wiener HH, Burden SJ. 2000. Selectiveexpression of Cre recombinase in skeletal muscle fibers. genesis26:165–166.
Branda CS, Dymecki SM. 2004. Talking about a revolution: the impactof site-specific recombinases on genetic analyses in mice. Dev Cell6:7–28.
Bruning JC, Michael MD, Winnay JN, Hayashi T, Horsch D, Accili D,Goodyear LJ, Kahn CR. 1998. A muscle-specific insulin receptorknockout exhibits features of the metabolic syndrome of NIDDMwithout altering glucose tolerance. Mol Cell 2:559–569.
Buckingham M, Bajard L, Chang T, Daubas P, Hadchouel J, Meilhac S,Montarras D, Rocancourt D, Relaix F. 2003. The formation of skele-tal muscle: from somite to limb. J Anat 202:59–68.
Buj-Bello A, Laugel V, Messaddeq N, Zahreddine H, Laporte J, PellissierJF, Mandel JL. 2002. The lipid phosphatase myotubularin is essen-tial for skeletal muscle maintenance but not for myogenesis inmice. Proc Natl Acad Sci U S A 99:15060–15065.
Casanova E, Fehsenfeld S, Mantamadiotis T, Lemberger T, Greiner E,Stewart AF, Schutz G. 2001. A CamKIIalpha iCre BAC allows brain-specific gene inactivation. genesis 31:37–42.
Cohn RD, Campbell KP. 2000. Molecular basis of muscular dystrophies.Muscle Nerve 23:1456–1471.
Cossu G, Borello U. 1999. Wnt signaling and the activation of myogene-sis in mammals. EMBO J 18:6867–6872.
Feil R, Brocard J, Mascrez B, LeMeur M, Metzger D, Chambon P. 1996.Ligand-activated site-specific recombination in mice. Proc NatlAcad Sci U S A 93:10887–10890.
Gehin M, Mark M, Dennefeld C, Dierich A, Gronemeyer H, Chambon P.2002. The function of TIF2/GRIP1 in mouse reproduction is dis-tinct from those of SRC-1 and p/CIP. Mol Cell Biol 22:5923–5937.
Giraldo P, Montoliu L. 2001. Size matters: use of YACs, BACs and PACsin transgenic animals. Transgen Res 10:83–103.
Grobet L, Pirottin D, Farnir F, Poncelet D, Royo LJ, Brouwers B, ChristiansE, Desmecht D, Coignoul F, Kahn R, Georges M. 2003. Modulatingskeletal muscle mass by postnatal, muscle-specific inactivation ofthe myostatin gene. genesis 35:227–238.
Gu H, Marth JD, Orban PC, Mossmann H, Rajewsky K. 1994. Deletionof a DNA polymerase b gene segment in T cells using cell type-spe-cific gene targeting. Science 265:103–106.
Hazama M, Watanabe D, Suzuki M, Mizoguchi A, Pastan I, Nakanishi S.2002. Different regulatory sequences are required for parvalbumingene expression in skeletal muscles and neuronal cells of trans-genic mice. Brain Res Mol Brain Res 100:53–66.
Hennet T, Hagen FK, Tabak LA, Marth JD. 1995. T-cell-specific deletionof a polypeptide N-acetylgalactosaminyl-transferase gene by site-directed recombination. Proc Natl Acad Sci U S A 92:12070–12074.
Imai T, Chambon P, Metzger D. 2000. Inducible site-specific somaticmutagenesis in mouse hepatocytes. genesis 26:147–148.
Imai T, Jiang M, Chambon P, Metzger D. 2001. Impaired adipogenesisand lipolysis in the mouse upon selective ablation of the retinoidX receptor alpha mediated by a tamoxifen-inducible chimeric Cre
recombinase (Cre-ERT2) in adipocytes. Proc Natl Acad Sci U S A98:224–228.
Imam AM, Patrinos GP, de Krom M, Bottardi S, Janssens RJ, Katsantoni E,Wai AW, Sherratt DJ, Grosveld FG. 2000. Modification of humanbeta-globin locus PAC clones by homologous recombination inEscherichia coli. Nucleic Acids Res 28:E65.
Indra AK, Warot X, Brocard J, Xiao JM, Chambon P, Metzger D. 1999.Temporally controlled site-specific mutagenesis in the basal layerof the epidermis: comparison of the recombinase activity of thetamoxifen-inducible Cre-ERT and Cre-ERT2 recombinases. NuclAcids Res 27:4324–4327.
Janssen I, Heymsfield SB, Baumgartner RN, Ross R. 2000. Estimation ofskeletal muscle mass by bioelectrical impedance analysis. J ApplPhysiol 89:465–471.
Kahn CR. 1994. Banting Lecture. Insulin action, diabetogenes, and thecause of type II diabetes. Diabetes 43:1066–1084.
Kastner P, Grondana J, Mark M, Gansmuller A, LeMeur M, Decimo D,Vonesch JL, Dolle P, Chambon P. 1994. Genetic analysis of RXRadevelopmental function: convergence of RXR and RAR signallingpathways in heart and eye morphogenesis. Cell 78:987–1003.
Kellendonk C, Tronche F, Casanova E, Anlag K, Opherk C, Schutz G.1999. Inducible site-specific recombination in the brain. J Mol Biol285:175–182.
Lakso M, Sauer B, Mosinger B Jr, Lee EJ, Manning RW, Yu SH, MulderKL, Westphal H. 1992. Targeted oncogene activation by site-spe-cific recombination in transgenic mice. Proc Natl Acad Sci U S A89:6232–6236.
Li M, Indra AK, Warot X, Brocard J, Messaddeq N, Kato S, Metzger D,Chambon P. 2000. Skin abnormalities generated by temporallycontrolled RXRa mutations in mouse epidermis. Nature 407:633–636.
Mascrez B, Mark M, Dierich A, Ghyselinck NB, Kastner P, Chambon P.1998. The RXRa ligand-dependent activation function 2 (AF-2) isimportant for mouse development. Development 125:4691–4707.
Mauvais-Jarvis F, Kulkarni RN, Kahn CR. 2002. Knockout models areuseful tools to dissect the pathophysiology and genetics of insulinresistance. Clin Endocrinol 57:1–9.
Metzger D, Chambon P. 2001. Site- and time-specific gene targeting inthe mouse. Methods 24:71–80.
Metzger D, Indra AK, Li M, Chapellier B, Calleja C, Ghyselinck NB,Chambon P. 2003. Targeted conditional somatic mutagenesis inthe mouse: temporally controlled knock out of retinoid receptorsin epidermal keratinocytes. Methods Enzymol 364:379–408.
Miniou P, Tiziano D, Frugier T, Roblot N, Le Meur M, Melki J. 1999.Gene targeting restricted to mouse striated muscle lineage.Nucleic Acids Res 27:e27.
Schmalbruch H, Hellhammer U. 1977. The number of nuclei in adultrat muscles with special reference to satellite cells. Anat Rec189:169–175.
Schuler M, Dierich A, Chambon P, Metzger D. 2004. Efficient temporallycontrolled targeted somatic mutagenesis in hepatocytes of themouse. genesis 39:167–172.
Schwenk F, Kuhn R, Angrand PO, Rajewsky K, Stewart AF. 1998. Tem-porally and spatially regulated somatic mutagenesis in mice.Nucleic Acids Res 26:1427–1432.
Sharan SK, Pyle A, Coppola V, Babus J, Swaminathan S, Benedict J,Swing D, Martin BK, Tessarollo L, Evans JP, Flaws JA, Handel MA.2004. BRCA2 deficiency in mice leads to meiotic impairment andinfertility. Development 131:131–142.
Tannour-Louet M, Porteu A, Vaulont S, Kahn A, Vasseur-Cognet M.2002. A tamoxifen-inducible chimeric Cre recombinase specifi-cally effective in the fetal and adult mouse liver. Hepatology35:1072–1081.
Weber P, Metzger D, Chambon P. 2001. Temporally controlled targetedsomatic mutagenesis in themouse brain. Eur J Neurosci 14:1777–1783.
Weber P, Cammas F, Gerard C, Metzger D, Chambon P, Losson R, MarkM. 2002. Germ cell expression of the transcriptional co-repressorTIF1beta is required for the maintenance of spermatogenesis inthe mouse. Development 129:2329–2337.
Wintermantel TM, Mayer AK, Schutz G, Greiner EF. 2002. Targetingmammary epithelial cells using a bacterial artificial chromosome.genesis 33:125–130.
170 SCHULER ET AL.


















