
Temporal Patterns of Gene Expression in the Antenna … · Temporal Patterns of Gene Expression in...
7
Copyright 8 1995 by the Genetics Society of America Temporal Patterns of Gene Expression in the Antenna of the Adult hsophilu mlunogaster Stephen L. Helfand, Kimberly J. Blake, Blanka Rogina, Mark D. Stracks, Alejandro Centurion and Boris Naprta Department of BioStructure and Function, School of Dental Medicine, University of Connecticut Health Center, Farmington, Connecticut 06030 Manuscript received December 7, 1994 Accepted for publication February 20, 1995 ABSTRACT The time course of gene expression in the adult fruit fly has been partially characterized by using enhancer trap and reporter gene constructs that mark 49 different genes. The relative intensity of the reporter protein in individual cells of the antennae was measured as a function of adult age. Most genes showed a graduated expression, and the intensity of expression had a reproducible and characteristic time course. Different genes displayed different temporal patterns of expression and more often than not the pattern of expression was complex. We found a number of genes having patterns that scaled with life span. In these cases the intensity of gene expression was found to be invariant with respect to biological time, when expressed as a fraction of the life span of the line. The scaling was observed even whenlifespan was varied as much as threefold. Such scaling serves to (1) further demonstrate that deterministic mechanisms such as gene regulation act to generate the temporal patterns of expression seen during adult life, (2) indicate that control of these regulatory mechanisms is linked to life span, and (3) suggest mechanisms by which this control is accomplished. We have concluded that gene expression in the adult fly is often regulated in a fashion that allows for graduated expression over time, and that the regulationitself is changing throughout adult life according to some prescribed program or algorithm. T HE temporal patterning of gene expression is widely recognized to play a central role during development (LAWRENCE 1992). Ithas been postulated that the regulation of gene expression may also play a role in the process of maturation and aging of the adult (FINCH 1990). Less is known about the regulation of gene expression in the maturing adult than during de- velopment. A number of studies have examined the regulation of gene expression in the mature aging ani- mal. These studies have both demonstrated a number of different genes that alter their level of expression during aging and have proposed molecular mecha- nisms for their regulation (RICHARDSON et al. 1987; SONG et al. 1991; SUPAKAR et al. 1993). The fruit fly, Drosophila melanogaster, which has played such a prominent part in elucidating the roleof gene expression during development, also possesses features that make it an excellent model system for examining gene expression during adult life and aging (ARKING and DUDAS 1989). Two such features are the fact that the adult fly consists almost entirely of postmitotic cells (BOZUCK 1972; IT0 and HOTTA 1992), eliminating the This work is dedicated to the memory of Joshua Bryan CorrespOndzng authoc Stephen L. Helfand, Department of BioStruc- ture and Function, School of Dental Medicine, University of Connect- icut Health Center, Farmington, CT 06030. E-mail: [email protected] Inouye Helfand. Genetics 140: 549-555 (June, 1995) confounding problems of cell replacement, and the availability of new techniques for examining gene ex- pression such as the enhancer trap and reporter gene techniques (O’KANE and GEHRING 1987; FREEMAN Despite extensive investigation, the control of the syn- thesis and degradation of macromolecules in the adult fly remains a matter of some controversy. This contro- versy may be due, at least in part, to limitations in the technology for examining macromolecules during adult life. Previous studies have had to concentrate on measuring changes in rate of synthesis or degradation of total RNA or protein in whole animals or tissues (SMITH et al. 1970; BAKER et al. 1985; LEVENBOOK 1986; WEBSTER 1986). Even studies focusing on a particular gene or gene product required the pooling of many cells. This pooling may have masked important aspects of gene regulation. New molecular genetic techniques such as the en- hancer trap and reporter gene techniques allow one to visualize the expression of single genes at the level of individual cells or specific organs in the whole animal (O’KANE and GEHRING 1987; FREEMAN 1991; RIESG~ ESCOVAR et al. 1992). These techniques provide a level of precision not previously available. In this paper we show the results of our studies using the enhancer trap and reporter gene techniques to examine gene expres- sion in the aging adult fly. 1991).
-
Upload
phungduong -
Category
Documents
-
view
226 -
download
7
Transcript of Temporal Patterns of Gene Expression in the Antenna … · Temporal Patterns of Gene Expression in...

Copyright 8 1995 by the Genetics Society of America
Temporal Patterns of Gene Expression in the Antenna of the Adult hsophilu mlunogaster
Stephen L. Helfand, Kimberly J. Blake, Blanka Rogina, Mark D. Stracks, Alejandro Centurion and Boris Naprta
Department of BioStructure and Function, School of Dental Medicine, University of Connecticut Health Center, Farmington, Connecticut 06030
Manuscript received December 7, 1994 Accepted for publication February 20, 1995
ABSTRACT The time course of gene expression in the adult fruit fly has been partially characterized by using
enhancer trap and reporter gene constructs that mark 49 different genes. The relative intensity of the reporter protein in individual cells of the antennae was measured as a function of adult age. Most genes showed a graduated expression, and the intensity of expression had a reproducible and characteristic time course. Different genes displayed different temporal patterns of expression and more often than not the pattern of expression was complex. We found a number of genes having patterns that scaled with life span. In these cases the intensity of gene expression was found to be invariant with respect to biological time, when expressed as a fraction of the life span of the line. The scaling was observed even when life span was varied as much as threefold. Such scaling serves to (1) further demonstrate that deterministic mechanisms such as gene regulation act to generate the temporal patterns of expression seen during adult life, (2) indicate that control of these regulatory mechanisms is linked to life span, and (3) suggest mechanisms by which this control is accomplished. We have concluded that gene expression in the adult fly is often regulated in a fashion that allows for graduated expression over time, and that the regulation itself is changing throughout adult life according to some prescribed program or algorithm.
T HE temporal patterning of gene expression is widely recognized to play a central role during
development (LAWRENCE 1992). It has been postulated that the regulation of gene expression may also play a role in the process of maturation and aging of the adult (FINCH 1990). Less is known about the regulation of gene expression in the maturing adult than during de- velopment. A number of studies have examined the regulation of gene expression in the mature aging ani- mal. These studies have both demonstrated a number of different genes that alter their level of expression during aging and have proposed molecular mecha- nisms for their regulation (RICHARDSON et al. 1987; SONG et al. 1991; SUPAKAR et al. 1993).
The fruit fly, Drosophila melanogaster, which has played such a prominent part in elucidating the role of gene expression during development, also possesses features that make it an excellent model system for examining gene expression during adult life and aging (ARKING and DUDAS 1989). Two such features are the fact that the adult fly consists almost entirely of postmitotic cells (BOZUCK 1972; IT0 and HOTTA 1992), eliminating the
This work is dedicated to the memory of Joshua Bryan
CorrespOndzng authoc Stephen L. Helfand, Department of BioStruc- ture and Function, School of Dental Medicine, University of Connect- icut Health Center, Farmington, CT 06030. E-mail: [email protected]
Inouye Helfand.
Genetics 140: 549-555 (June, 1995)
confounding problems of cell replacement, and the availability of new techniques for examining gene ex- pression such as the enhancer trap and reporter gene techniques (O’KANE and GEHRING 1987; FREEMAN
Despite extensive investigation, the control of the syn- thesis and degradation of macromolecules in the adult fly remains a matter of some controversy. This contro- versy may be due, at least in part, to limitations in the technology for examining macromolecules during adult life. Previous studies have had to concentrate on measuring changes in rate of synthesis or degradation of total RNA or protein in whole animals or tissues (SMITH et al. 1970; BAKER et al. 1985; LEVENBOOK 1986; WEBSTER 1986). Even studies focusing on a particular gene or gene product required the pooling of many cells. This pooling may have masked important aspects of gene regulation.
New molecular genetic techniques such as the en- hancer trap and reporter gene techniques allow one to visualize the expression of single genes at the level of individual cells or specific organs in the whole animal (O’KANE and GEHRING 1987; FREEMAN 1991; R I E S G ~ ESCOVAR et al. 1992). These techniques provide a level of precision not previously available. In this paper we show the results of our studies using the enhancer trap and reporter gene techniques to examine gene expres- sion in the aging adult fly.
1991).

550 S. L. Helfand et al.
MATERIALS AND METHODS Fly stocks: Forty of the 49 marked genes used in this study,
including all the lines shown in Figures 1-5, were enhancer trap lines containing the P-LmWconstruct in which a nuclear targeting sequence is associated with P-gal (BIER et al. 1989). These 40 lines were selected at random from a set of 100 lines that showed expression in the adult antenna. These 100 lines had themselves been obtained from a set of over 800 enhancer trap lines. The criteria for their selection was expression in- cluding but not limited to the adult antenna. These lines had all been backcrossed with the wl"* stock for over a dozen generations (BOYNTON and TULLY 1992). Molecular studies have shown that these are 40 different genes (S. HELFAND, unpublished results). The remaining nine marked genes con- sist of seven enhancer trap lines of known genes (KASSIS et al. 1992) and stocks containing reporter genes for the e l m promoter (YAO and WHITE 1994) and the cut promoter (JACK et al. 1991).
Fly culturing: All flies were kept in plastic vials containing a standard corn meal agar medium with several grains of yeast added (ASHBURNER 1989). Approximately 30 flies were in each vial; flies were passed to fresh vials every 7 days. All flies were cultured in humidified temperature-controlled environ- mental chambers at 25" throughout development. Adult flies were collected without anesthesia within 2 hr after eclosion from pupae and put in humidified temperaturecontrolled environmental chambers set at 18", 25", or 29". Male and female flies were aged together in the same vial.
Whole mount X-@ staining: For each time point, at least 10 male and 10 female fly heads were removed, fixed for 20 min in 1% glutaraldehyde in phosphate-buffered saline (1X PBS), and reacted with a standard X-gal solution (5-bromo- 4-chloro-3-indolyl-~-D-galactopyranoside, Boehringer-Manne- heim) (ASHBURNER 1989). For all but two lines, X-gal reactions were performed for 18 hr at 37". Reactions for two lines were terminated after 2 hr due to their high level of P- gal. After X-gal reaction, heads were rinsed twice with I X PBS and stored in 70% glycerol in 1 X PBS. Antennae were removed from heads and placed on microscope slides, cov- erslips were added, and photographs were taken with a X16 Leitz objective. Wherever possible, flies homozygous for the marked gene were used. While staining revealed the expected lower level in intensity in heterozygotes, spatial and temporal patterns of stain and life span studies demonstrated little, if any, difference between homozygotes and heterozygotes. In some of the lines, the large number of stained cells, combined with the use of low power views (X160), make it difficult to visualize the nuclear staining of individual cells. Under higher power (x~OO), the blue staining is seen to be associated with nuclei of individual cells.
Quantitation of p-gal expression: The amount of P-gal ex- pressed in the antennae of the marked gene stocks is too low to be detected by standard chromogenic assays such as chlorophenol red-P-D-galactopyranoside (CPRG) (GLASER and LIS 1990). We developed an optically based computer- assisted video microscopy system for the quantification of P- gal that used our X-gal reacted whole mounts. A detailed description of the computer-assisted video microscopy system will be published elsewhere. Briefly, images of each specimen were captured using a system consisting of a Leitz Orthoplan I1 microscope, a color Optronix video camera, a Macintosh Quadra 800 computer with RasterOps Video Capture Board, and IPLab Spectrum software (Signal Analytics, Inc.). Images of 220 antennae from a particular time point were captured. The reliability of the system is such that capture of the same image at different times shows <2% difference in values. The IPLab spectrum software was used to determine the relative intensity of X-gal staining by first acting as a spectrophotome-
ter defining the range of blue to be examined, then as a densitometer, measuring the relative intensity contributed by that color, and finally determining the area of each antennae. The relative staining of each antenna was determined by divid- ing the amount of staining by the area. Average values and SEM at each time point were calculated using Microsoft Excel then plotted with curves fit using KaleidaGraph.
Life span: Life span studies were carried out using methods of collection and culture noted above except that the flies were passed into fresh vials every other day at which time the number of dead males and females were recorded. Over 400 male and female flies were scored for each life span study. Male and female flies were aged together in the same vial. All lines tested appeared to have normal life spans as compared to our wild-type controls. Mean and maximum life spans for animals homozygous and heterozygous for the marked genes were similar.
RESULTS
The level of expression of some genes in the adult fly changes over the life span of the animal: Enhancer trap and reporter gene techniques provide an indirect means of determining the transcriptional activity of a particular gene by measuring the amount of the re- porter gene protein, p-galactosidase (p-gal), being ex- pressed. The level of expression of p-gal was examined in whole mount adult antennae at sequential time points throughout the adult life span for 49 different genes. The selection of these 49 different genes was based solely upon their expression in adult antennae. The adult antenna was chosen due to its ready perme- ability to reagents such as X-gal, accessibility to visual inspection and experimental manipulation, and its na- ture as a well defined tissue consisting entirely of post- mitotic cells (BOZUCK 1972; ITO and HOTTA 1992; RIESGO-ESCOVAR et al. 1992).
Figure 1 shows representative genes for each of four different temporal patterns of expression. Line 302 de- picts the class of genes whose pattern of expression does not change during adult life. Line 1059 is one example from a class of genes in which the level of expression begins slowly in early life, reaches a peak in midlife, and then gradually declines. Line 1020 in Fig- ure 1 and line 1085 in Figure 2 are two representatives from a class of genes that do not begin expression until several days after the adult emerges from the pupal case. Another class of genes, which dramatically reduces expression well before the end of adult life, is illustrated by line 2216.
The relative frequency of each of these different classes of temporal patterns of expression is shown in Table 1. Of the 49 genes examined 80% show marked changes in their level of expression during their adult life span, indicating that gene expression in the adult fly is a more dynamic process than generally appreciated.
Temporal patterns of expression suggest that gene regulation remains active during adult life: In addition to documenting the dynamic nature of gene expression in the adult, these studies also provide evidence that

302 1 I
1059 S
Gene Expression During Aging
R
1020 (+
2216 il Days 1
r
5 10 20-25 35-40 50
55 1
1 - 100 pm
60 FIGURE 1.-Photomicrographs ofwhole mount adult antennae from enhancer trap lines 302,1059,1020, and 2216 at different
ages reacted with X-gal to reveal blue staining in the nuclei of cells that are expressing @-gal. Each photograph represents a mica1 examde from over 60 different antennae examined for each time point. Days listed on the bottom are from the time of' the adults' emergence from the pupal case.
the expression of some genes in the adult is regulated in a systematic manner throughout adult life. Although the expression of different marked genes may show different temporal motifs, under similar conditions each gene's temporal pattern of expression is stereo- typic (Figure 1). This fact is further illustrated in Fig- ures 2 and 3,line 1085, which show qualitative and quan- titative analyses of the level of p-gal expression throughout the life span for one of our marked genes. The quantitative data confirms the conclusions drawn from the qualitative inspection of our in situ studies, showing that each gene has its own temporal signature pattern of expression and that these patterns are often complex. Stereotypic temporal patterns of expression suggest a deterministic or nonrandom aspect to the control of gene expression during adult life.
Altering life span shows that the temporal patterning of gene expression scales to life span: To gain insight into the mechanisms underlying the regulation of gene expression during adult life, we examined the temporal pattern of gene expression under conditions that alter life span. Figures 2-5 show that when ambient tempera- ture is used to alter life span, gene expression for line
1085 changes in a manner that is a function of the l ie span of the adult fly.
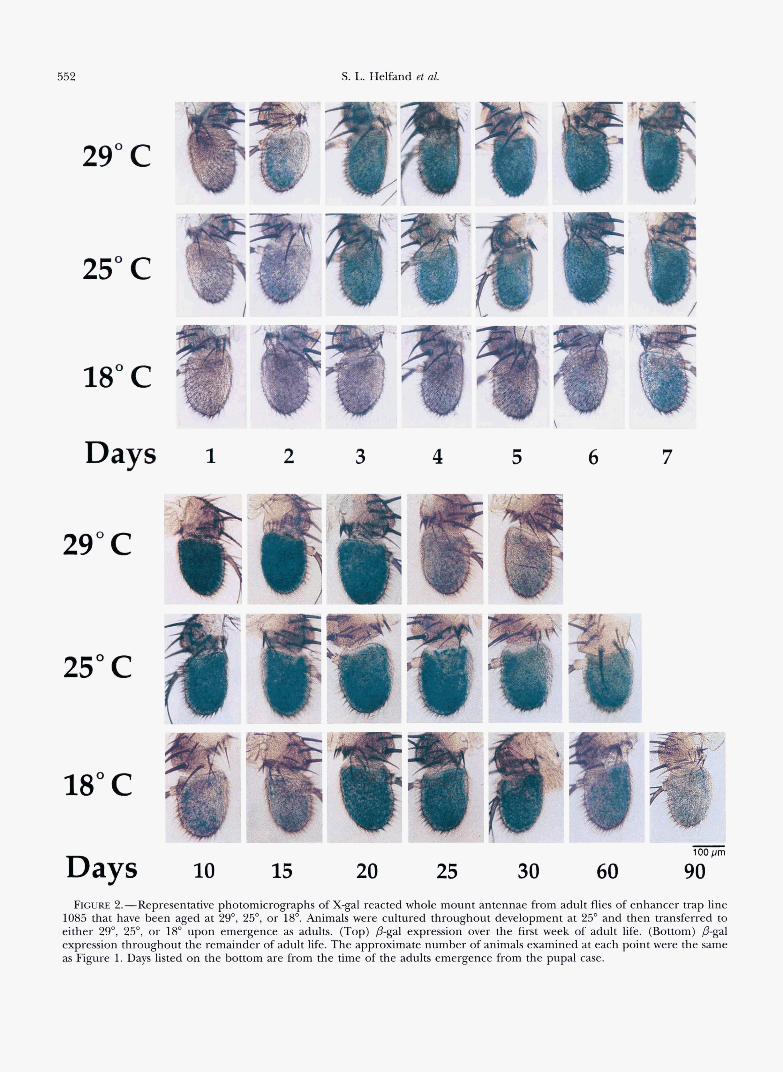
Figure 2 shows the level of p-gal being expressed in representative antenna for line 1085 cultured through- out adult life at three different temperatures, while Figure 3 shows the quantitative results of these experi- ments. Figure 4 shows survivorship curves demonstrat- ing the threefold change in life span seen when the adult is cultured at different ambient temperatures. The maximum life spans are 45, 72, and 135 days at 29") 25") and 18") respectively.
Figure 2 (top) shows that when the animals have been cultured from the time of eclosion at 29", the expression of the gene marked in line 1085 is first seen on day 2 of adult life but is not apparent until day 7 when the animals have been cultured at 18". Figure 2 (bottom) shows that peak expression of the gene marked in line 1085 also shifts at these Merent temper- atures. This can be seen more readily in Figure 3 where the peak level appears to be reached by day 15 at 29", day 25 at 25") and not until after day 35 at 18". Taken together, these findings reveal that when viewed from the perspective of chronological or calendar time the
552 S. L. Helfand et al.
z 29" C
L
i
._ " F
25" C 1 I
18" C I
Days 1 2 3 4 5 6 7
29OC
- 25" C
18" C -
10 15 20 25 30 60 90 100 pm
Days FIGURE 2.-Representative photomicrographs of X-gal reacted whole mount antennae from adult flies of enhancer trap line
1085 that have been aged at 29", 25", or 18". Animals were cultured throughout development at 25" and then transferred to either 29", 25", or 18" upon emergence as adults. (Top) &gal expression over the first week of adult life. (Bottom) @gal expression throughout the remainder of adult life. The approximate number of animals examined at each point were the same as Figure 1. Days listed on the bottom are from the time of the adults emergence from the pupal case.

Gene Expression During Aging
TABLE I
Temporal patterns of gene expression seen during adult life
% of total
Type of pattern genes examined No. of genes
Constitutive expression" 10 20 Changing expression* 39 80 Increasing from low level 27 55
Off at eclosion 7 14 On from eclosion 20 41
Decreasing from high initial level 12 25 Total 49"
a Constitutive expression indicates that the amount of @-gal expression remained at the same, often high level, from the beginning of adult life, emergence of the adult from the pupal case, through the maximum life span of the line.
'Changing expression indicates that the amount of O-gal expression changed several fold, often 6 to 10-fold, during adult life, either increasing from a low level or decreasing from an initial higher level.
'The 49 genes examined are a subset of genes found to have expression in the antenna of the adult fly. Not all genes that show expression in the adult antenna have been included in this study.
timing of gene expression for animals cultured at the three different temperatures changes dramatically.
However, when the level of gene expression is ex- pressed as a proportion of life span as is shown in Figure 5, the timing and intensity of gene expression is seen to remain nearly the same despite a threefold change in life span. This shows that for the gene marked in
140 1 ,120 .r( 0 2 1 0 0
8 0
6 0
4 0 cu
.r( 2 20
0 2 0 4 0 6 0 8 0 100 120 140 Age (in days)
FIGURE 3.-Quantitative analysis of the effect of altering lifespan by means of ambient temperature on the temporal pattern of expression of @-gal in the antenna of adult flies from line 1085, 29" (O), 25" (A), and 18' (0). A computer- assisted video system was used to determine the intensity of the X-gal reaction in the same specimens as described in Figure 2. At least 20 different antennae were sampled for each point. Error bars are SEM. The curves were fit to the points for each temperature using a third degree polynomial equation.
553
IO0
80
7 60
5 4 0
20
0
' E UY
0 20 40 6 0 80 100 120 140
Age (in days) FIGURE 4.-Survivorship curves for adult flies from line
1085 cultured at 29" (n), 25" (A), and 18" (0). Over 400 male and female flies were scored at each temperature. Maxi- mum life span of line 1085 was 45 days at 29", 72 days at 25", and 135 days at 18".
line 1085, the temporal pattern of gene expression scales with respect to life span. Twelve other genes tested also show their temporal pattern of expression to scale with life span (S. HELFAND, unpublished results).
DISCUSSION
Genetic elements are known to play a major role in determining a species' life span. Although the relation- ship between the genome and life span has been well documented, the mechanisms underlying this relation- ship are not understood (FINCH 1990). One model for
140
.r( g 1 2 0
3 1 0 0 Y
p!
cu 6 0 X
.H iZ' 4 0
80
0
v1 j 2 0
0 0 20 40 6 0 8 0 100
% Life Span FIGURE 5.-Intensity of the X-gal reaction plotted against
percentage life span. The percentage life span for each value was determined by dividing the day for each point (from Figure 3) by the maximum life span of line 1085 at that tem- perature (from Figure 4). The curves were fit to the points for each temperature 29" (O), 25" (A) , and 18" (0) using a third degree polynomial equation. SEM are the same as those in Figure 3 but have been left out here for clarity.

554 S. L. Helfand et al.
how the genome may affect life span emphasizes the contribution of nonrandom or programmed changes in gene expression during adult life (FINCH 1990). Such a model depends on the demonstration of genes whose expression is regulated during adult life.
The qualitative and quantitative analyses of gene ex- pression described in this paper show that many differ- ent genes (40 out of 49 examined) dramatically alter their level of expression during adult life and that these alterations are not random. Each gene has its own re- producible temporal signature pattern of expression over the life span of the animal. The documentation of these stereotypic temporal patterns of expression indi- cates that deterministic mechanisms, such as gene regu- lation, direct gene expression during adult life.
The existence of temporal patterns of gene expres- sion during adult life raises the question of the relation- ship between these patterns of gene expression and life span. We found a number of genes whose pattern of expression scaled with life span. In these cases the inten- sity of gene expression is found to be invariant with respect to biological time. Although the chronological or calendar timing of the level of gene expression changes with alterations in life span (Figures 2 and 3), the timing of the level of gene expression as a function of the percentage of the animals life span, i.e., age in biological time, remains to a certain extent fixed (Fig- ure 5). The curves are nearly superimposable. These results indicate that, at least in some instances, the con- trol of gene regulation is linked to life span.
An alternative interpretation of these results is that there is no linkage between the control of gene regula- tion and life span. The method used for altering life span, ambient temperature, could be affecting gene expression through a direct chemical-metabolic or ther- modynamic effect on the rate of gene expression of that gene. Ambient temperature would then be affecting the expression of the gene independently of its effect on life span. We have found, however, that when a manipu- lation other than ambient temperature is used to alter the life span of the adult fly the timing of gene expres- sion also shows scaling to life span. Shaker and Hypmki- netic are two single gene mutations that shorten life span in association with an increase in physical activity and general metabolic rate (TROUT and WLAN 1970). When gene expression is examined in the background of either of these two mutants, the temporal pattern of expression for several different genes changes in a simi- lar manner to what is seen with ambient temperature. The temporal patterns of gene expression scale with life span (S. HELFAND, unpublished results). The finding of scaling, both when life span was varied by ambient temperature or by genetic mutation, strengthens the interpretation that there is a linkage between the con- trol of gene expression and life span.
Some characteristics of this link between the control of gene regulation and life span may be predicted from
the type of scaling observed. Such scaling strongly im- plies the existence of gene control that is keyed to life span through a mechanism like an odometer that mea- sures distance traveled along a prescribed course. Su- perficially, an odometer model has features that are consistent with the “rate-of-living” hypothesis. The rate- of-living hypothesis, however, does not distinguish be- tween mechanisms that are due to random events us. those due to nonrandom events. In fact, random events, such as the accumulation of molecular damage due to reactive oxygen species, is one of the presently favored mechanisms used to support the rate-of-living hypothe- sis (SOHAL 1986; O m and SOW 1994). The odometer model, or course-distance traveled model, implies that nonrandom mechanisms are involved in linking the control of gene regulation to life span. Some step-by- step procedure or algorithm is being used to determine the level of gene expression. Using this algorithm, the level of gene expression is adjusted to fit its own prede- termined pathway of expression.
The validity of our use of P-gal as a marker of gene expression during adult life requires that the changes in the level of ,&gal observed reflect gene regulation and not just general alterations in cell physiology or changes in cell populations. There is little change in somatic cell populations in the adult D. melunogaster in general and in the antennae in particular. The cells of the adult antennae are postmitotic (BOZUCK 1972; IT0 and HOTTA 1992), and there is little loss due to cell death. Less than l0-15% of the cells in the third seg- ment of the antennae are lost over the life span of the adult fly (S. HELFAND, unpublished results). We have found that flies containing an Rh1 opsin-lucZ construct (MISMER and RUBIN 1987) or an hsp70-ZucZ construct (LIS et al. 1983), as well as a number of our enhancer trap lines, such as line 302 (Figure l ) , all continue to express &gal up to their maximal life span, demonstra- ting that for most cells the machinery for the synthesis of P-gal is present and active throughout the fly’s adult life (S. HELFAND, unpublished results). Finally, the rate of degradation of @gal in adult cells is on the order of several hours (S. HELFAND, unpublished results). Since our experiments examine time points that are 21 day apart, this rate of degradation is well within the time scale needed for our studies.
The predictable, almost fixed, relationship between the temporal pattern of gene expression and the life span of the animal indicate that these marked genes are better predictors of age than calendar time itself. They may thus be useful as biomarkers of aging as they appear to reflect physiological rather than chronologi- cal age. The scaling of gene expression to conform with the life span of the animal suggests that specific mechanisms, such as the odometer model, link the con- trol of gene expression to life span. It is likely that these mechanisms are also involved in the process of aging and the determination of longevity. The coupling of

Gene Expression During Aging 555
the use of molecular biomarkers we have presented in this paper with standard Drosophila molecular and genetic techniques provides a means for identifying and isolating molecular and genetic determinants involved in regulating gene expression during adult maturation and aging in Drosophila. A more detailed analysis of these mechanisms should help to shed light on the elements involved in determining longevity.
The authors thank ELTON KATZ for his many insightful and gener- ous contributions. We also thank MARVIN TANZER, MARTIN RAFF, JO- SEPH JACK, and WILLIAM UPHOLT for discussions and critical com- ments on the manuscript, BRJAN GRIMWADE and GREG FITZGERALD for technical help, TIM TULLY for providing his enhancer trap muta- genesis for our screening, and JUDY KASSIS, GERALD RUBIN, and ROSE- MARIE REINKE for fly stocks. This work was supported by grants from the National Science Foundation (IBN-9122097), American Federa- tion for Aging Research, Sandoz Foundation for Gerontological Re- search, University of Connecticut Health Center Research Advisory Committee (S.L.H.), a National Institutes of Health NRSA Training Grant 5T32 ARO4370-14 (K.J.B.), and The Patrick and Catherine Weldon Donaghue Medical Research Foundation (B.R.).
LITERATURE CITED
ARKING, R., and S. P. DUDAS, 1989 Review of genetic investigations into the aging processes of Drosophila. J. A m . Geriatr. SOC. 37: 757-773.
ASHBURNER, M., 1989 Drosophila: A Laboratmy Handbook. Cold Spring Harbor Press, Cold Spring Harbor, NY.
BAKER, G. T., M. JACOBSON, and G. MOKRYNSKI, 1985 Agzng in Dr* sophila. CRC Press, Boca Raton, FL.
BIER, E., H. VASSIN, S. SHEPHERD, K. LEE, IL MCCALL et al., 1989 Searching for pattern and mutation in the Drosophila genome with a P-hcZ vector. Genes Dev. 3 1273-1287.
BOYNTON, S., and T. TULLY, 1992 latheo, a new gene involved in associative learning and memory in Drosophila mlanogaster, identi- fied from P element mutagenesis. Genetics 131: 655-672.
BOZUCK, A. N., 1972 DNA synthesis in the absence of somatic cell division associated with ageing in Drosophila subobscura. Exp. Ger- ontol. 7: 147-156.
FINCH, C. E., 1990 Longevity, senescence, and the p o m e . University of Chicago Press, Chicago.
FREEMAN, M., 1991 First, trap your enhancer. Curr. Biol. 1: 378- 381.
GLASER, R. L., and J. T. LIS, 1990 Multiple, compensatory regulatory elements specify spermatocyte-specific expression of the Lhsoph- ila mlanogasterhsp26 gene. Mol. Cell. Biol. 1 0 131-137.
ITO, IC, andY. HOTTA, 1992 Proliferation pattern of postembryonic
neuroblasts in the brain of Drosophila mlanogaster. Dev. Biol. 149:
JACK, J., D. DORSEIT, Y. DELOTTO and S. LIU, 1991 Expression of the cut locus in the Drosophila wing margin is required for cell type specification and is regulated by a distant enhancer. Devel- opment 113 735-747.
KASSIS, J. A., E. NOLL, E. P. VANSICKLE, W. F. ODENWALD and N. PERRIMON, 1992 Altering the insertional specificity of a Drosoph- ila transposable element. Proc. Natl. Acad. Sci. USA 89: 1919- 1923.
LAWRENCE, P. A,, 1992 The Making of a Hy. The Genetics of Animal Design. Blackwell Scientific Publications, Oxford.
LEVENBOOK, L., 1986 Protan Synthesis in Relation to Insect Aging: An Overview. Springer-Verlag, New York.
LIS, J. T., J. A. SIMON and C. A. SUTTON, 1983 New heat shock puff and &gal activity resulting from tranformation of Drosophilawith an hsp70-lac Z hybrid gene. Cell 35: 403-410.
MISMER, D., and G. M. RUBIN 1987 Analysis of the promoter of the ninaE opsin gene in Drosophila mlanogaster. Genetics 116: 565- 578.
O’KANE, IC, and W. GEHRING, 1987 Detection in situ of genomic regulatory elements in Drosophila. Proc. Natl. Acad. Sci. USA
ORR, W. C., and R. S. SOHAL, 1994 Extension of life-span by overex- pression of superoxide dismutase and catalase in Drosophila mela- nogaster. Science 263: 1128- 1130.
RICHARDSON, A., J. A. BUTLER, M. S. RUTHERFORD, I. SEMSEI, M.-Z. Gu et al., 1987 Effect of age and dietary restriction on the expression of a-Globulin. J. Biol. Chem. 262: 12821-12825.
RIESGO-ESCOVAR, J., C. WOODARD, P. GAINES and J. CARLSON, 1992 Development and organization of the Drosophila olfactory system: An analysis using enhancer traps. J. Neurobiol. 23: 947-964.
SMITH, J. M., A. N. BOZUCK and S. T E B B U ~ , 1970 Protein turnover in adult Drosophila. J. Insect Physiol. 16: 601-613.
SOHAL, R. S., 1986 The rate of living theory: a contemporary inter- pretation. in Insect Aging: Strategies and Mechanisms. Springer-Ver- lag, Berlin.
SONG, C. S., T. R. RAO, W. F. DEMYAN, M. A. MANCINI, B. CHATTERJEE et al., 1991 Androgen receptor messenger ribonucleic acid (mRNA) in the rat liver: changes in mRNA levels during matura- tion, aging and calorie restriction. Endocrinology 128: 349-356.
SUPAKAR, P. C., C. S. SONG, M. H. JUNG, M. A. SLOMCZYNSKA, J.-M. KIM et al., 1993 A novel regulatory element associated with age- dependent expression of the rat androgen receptor gene. J. Blol. Chem. 268: 26400-26408.
TROUT, W. E., and W. D. KAPLAN, 1970 A relation between longevity, metabolic rate, and activity in Shaker mutants of Drosophila mela- nogaster. Exp. Geront. 5: 83-92.
WEBSTER, G. C., 1986 E f f t of Aging on the Components of the Protein Synthesis System. Springer-Verlag, New York.
YAO, K. M., and K. WHITE, 1994 Neural specificity of elav expression: defining a Drosophila promoter for directing expression to the nervous system. J. Neurochem. 63: 41-51.
134-148.
84: 9123-9127.
Communicating editor: V. G. FINNERTY


















