
TEMPERATURE COUPLING AND 'TRADE-OFF' PHENOMENA...
19
J. exp. Biol. 114, 17-35 (1985) 17 Printed in Great Britain © The Company of Biologists Limited 1985 TEMPERATURE COUPLING AND 'TRADE-OFF' PHENOMENA IN THE ACOUSTIC COMMUNICATION SYSTEM OF THE CRICKET, GRYLLUSBIMACULATUS DE GEER (GRYLLIDAE) BY JOHN A. DOHERTY* Max-Planck-Institut fur Verhaltensphysiologie, Abteilung Huber, D-8131 Seewiesen, Federal Republic ofGermanv Accepted 8 August 1984 . SUM M A R Y The effects of ambient temperature on stridulation (calling song) in males, and phonotaxis in females, were studied in the chirping cricket, Gryllus bimaculatus. In the male, temperature had the greatest effect on the syllable and chirp repetition rates. Both increased linearly with increas- ing temperature between 15 and 24°C; there was no effect of temperature on these temporal properties at higher temperatures (24-33 °C). Syllable duration, number of syllables per chirp and dominant frequency remained relatively unaffected by changes in temperature. Stridulation and phono- taxis were temperature coupled because the female at 15, 22 and 30°C responded best to synthetic songs with syllable and chirp repetition rates that matched these temporal properties of the male's calling song at the same temperature. The phonotactic behaviour of the female indicates that certain combinations of temporal properties in the male's calling Song improve the female's response at different temperatures, suggesting the presence of 'trade-off phenomena in phonotaxis and pattern recognition in G. bimaculatus. INTRODUCTION Acoustic communication among conspecifics requires that signal production in the sender be matched with signal recognition in the receiver. This matching between sender and receiver is complicated in poikilothermic animals that signal and respond over a range of ambient temperatures (e.g. Walker, 196Za,6, 1975; Schneider, 1977; Gerhardt, 1978, 1982, 1983). In some poikilotherms which produce signals that change with temperature, this problem has been solved by 'temperature coupling' (Gerhardt, 1978). This means that a change in temperature causes parallel shifts in signal generation and signal recognition, so that sender and receiver remain matched over the entire range of temperatures in which com- munication occurs. Temperature coupling is best expressed in communication systems that employ temporal signals and has been described in acoustic com- munication in crickets, grasshoppers and treefrogs, and in visual communication in •Present address: Section of Neurobiology and Behavior, Seeley G. Mudd Hall, Cornell University, Ithaca, NY 14853, U.S.A. Key words: Crickets, temperature effects, stridulation, phonotaxis, trade-off phenomena.
Transcript of TEMPERATURE COUPLING AND 'TRADE-OFF' PHENOMENA...

J. exp. Biol. 114, 17-35 (1985) 17Printed in Great Britain © The Company of Biologists Limited 1985
TEMPERATURE COUPLING AND 'TRADE-OFF' PHENOMENAIN THE ACOUSTIC COMMUNICATION SYSTEM OF THE
CRICKET, GRYLLUSBIMACULATUS DE GEER (GRYLLIDAE)
BY JOHN A. DOHERTY*
Max-Planck-Institut fur Verhaltensphysiologie, Abteilung Huber, D-8131Seewiesen, Federal Republic ofGermanv
Accepted 8 August 1984
. SUM M A R Y
The effects of ambient temperature on stridulation (calling song) inmales, and phonotaxis in females, were studied in the chirping cricket,Gryllus bimaculatus. In the male, temperature had the greatest effect onthe syllable and chirp repetition rates. Both increased linearly with increas-ing temperature between 15 and 24°C; there was no effect of temperatureon these temporal properties at higher temperatures (24-33 °C). Syllableduration, number of syllables per chirp and dominant frequency remainedrelatively unaffected by changes in temperature. Stridulation and phono-taxis were temperature coupled because the female at 15, 22 and 30°Cresponded best to synthetic songs with syllable and chirp repetition ratesthat matched these temporal properties of the male's calling song at thesame temperature. The phonotactic behaviour of the female indicates thatcertain combinations of temporal properties in the male's calling Songimprove the female's response at different temperatures, suggesting thepresence of 'trade-off phenomena in phonotaxis and pattern recognition inG. bimaculatus.
INTRODUCTION
Acoustic communication among conspecifics requires that signal production inthe sender be matched with signal recognition in the receiver. This matchingbetween sender and receiver is complicated in poikilothermic animals that signaland respond over a range of ambient temperatures (e.g. Walker, 196Za,6, 1975;Schneider, 1977; Gerhardt, 1978, 1982, 1983). In some poikilotherms whichproduce signals that change with temperature, this problem has been solved by'temperature coupling' (Gerhardt, 1978). This means that a change in temperaturecauses parallel shifts in signal generation and signal recognition, so that sender andreceiver remain matched over the entire range of temperatures in which com-munication occurs. Temperature coupling is best expressed in communicationsystems that employ temporal signals and has been described in acoustic com-munication in crickets, grasshoppers and treefrogs, and in visual communication in
•Present address: Section of Neurobiology and Behavior, Seeley G. Mudd Hall, Cornell University,Ithaca, NY 14853, U.S.A.
Key words: Crickets, temperature effects, stridulation, phonotaxis, trade-off phenomena.

18 J . A . DOHERTY
fireflies (e.g. Walker, 1957; Helversen & Helversen, 1981; Gerhardt, 1978, 1982,1983; Carlson, Copeland, Raderman & Bulloch, 1976).
Temperature coupling is not the only solution to the problem of maintainingeffective communication at different temperatures. In some species of acrididgrasshoppers, for example, the receiver evaluates properties of the signal (i.e. ratiosbetween syllable durations and pause durations) that are independent of temper-ature (see Helversen, 1972; Helversen, 1979; Helversen & Helversen, 1981;Skovmand & Pedersen, 1983).
The study of temperature coupling phenomena can provide new insights into theinterrelationships of the physiological mechanisms that underlie pattern generationand pattern recognition. Studies of interspecific hybridization in crickets andtreefrogs have provided indirect evidence that pattern generation and patternrecognition have common neural elements (see Hoy, 1978; Hoy, Hahn & Paul,1977; Doherty & Gerhardt, 1983, 1984). This idea has been termed the 'geneticcoupling' hypothesis, and the phenomenon of temperature coupling supports thishypothesis (Hoy, 1974).
In this paper, I report on how the cricket, Gryllus bimaculatus, changessignalling and phonotaxis at different temperatures and establishes sender-receivermatching. I provide new evidence for temperature coupling in a chirping cricketand discuss the interactions of temporal properties and 'trade-off phenomena thatunderlie calling song recognition and the elicitation of phonotaxis behaviour atdifferent temperatures. This paper is an extension of Walker's pioneering study onthe effects of temperature on acoustic communication in trilling tree crickets(Walker, 1957). The calling song of G. bimaculatus differs from that in treecrickets, as it is composed of discrete chirps that contain three to five sound pulses(syllables). These chirps are repeated in a continuous though variable temporalsequence, whereas the pulse (syllable) repetition rate within individual chirps ismore stereotyped. As in tree crickets., the calling song of G. bimaculatus and otherGryllids attracts conspecific females for mating. Furthermore, these cricketscommonly sing over a broad temperature range between 15 and 35 °C (Alexander,1957; Alexander & Meral, 1967; Walker, I962a,b). Finally, the recognition processin the female can be elucidated indirectly by studying phonotaxis behaviour on awalking compensator to playbacks of synthetic models of the calling song (seeWendler, Dambach, Schmitz & Scharstein, 1980; Weber, Thorson & Huber, 1981;Thorson, Weber. & Huber, 1982; Schmitz, Scharstein & Wendler, 1982).
MATERIALS AND METHODS
Male stridulation
The calling songs of single males of G. bimaculatus from laboratory cultures wererecorded 2-10 weeks after the final moult. Singing males were isolated in glass jarsand recorded in the dark in a temperature-controlled incubator (Memmert RO-8),after being acclimated to the incubator temperature for a period of 2 h to severaldays. Acclimation times had no systematic effects on the physical properties ofcalling song. Songs were recorded on an Akai 280D-SS tape recorder(9-5 cms"1), and the recording microphone (AKG) was suspended above the glass

Cricket stridulation and phonotaxis 19
jar within 10 cm of the male. Usually, spontaneous singing was recorded, althoughsometimes stridulation was initiated by playing back synthetic calling songs that hadphysical properties that were characteristic of the natural calling song. Air tem-perature was measured by a thermister that was suspended within 5 cm of thesinging male. Due to the small body sizes of these crickets, it was assumed that thetemperature of the surrounding air was the same as the body temperature of thesinging male (see Prestwich & Walker, 1981). All temperatures reported in thispaper were ±1°C.
The temporal and spectral properties of the calling song were analysed by usingeither a digital oscilloscope or a PDP 11/40 (DEC) computer. Syllable periods(SP), chirp periods (CP) and the number of syllables per chirp (SN) were digitizedand analysed by the computer (see Figs 2, 3 for definitions of temporal properties).The temporal properties of chirps with 3-5 syllables were pooled. The syllableduration (SD) of the third, fourth or fifth syllables of chirps was measured directlyfrom the oscilloscope (±2ms). The sinusoidal nature of calling songs enabled thedirect measurement of dominant frequency by taking the reciprocal of the timebetween successive zero crossings of the expanded wave form on the digitaloscilloscope (±10 Hz).
In some males, calling songs were recorded several times at the same tem-perature. The data from these recordings were pooled in individuals because therewere no differences in the songs that were recorded at different times at the sametemperature. The calling songs of some males were recorded while other males weresinging at the same time in the incubator. These data were pooled also because therewere no systematic differences in the calling songs of individuals which sang eitherin acoustic isolation or with other singing males.
Female phonotaxis
Adult virgin females from laboratory cultures were studied 2-12 weeks after thefinal moult. Within a week after the last moult, they were kept in isolation until theexperiments, and did not hear songs during this isolation period. Phonotactictracking was studied by using a feedback-controlled walking compensator (Kramerspherical treadmill) in an anechoic chamber (for details, see Weber et al. 1981;Thorson et al. 1982). Briefly, the female walked on top of a sphere and the changein the female's position was detected by a pulsed-infrared scanning system. Thissystem drove compensatory motors that moved the sphere in the direction oppositeto the direction of the female's movement. The compensatory movement kept thefemale on top of the sphere, enabling study of a freely walking female in anunchanging but controllable sound field. By analysing instantaneous velocity anddirection profiles of the female's movement (see Fig. 1), the tracking performanceto an acoustic stimulus could be quantified by calculating the percentage of thestimulus presentation time that the female clearly 'tracked' the stimulus (Weber etal. 1981; Thorson et al. 1982). A stimulus that was not tracked by a female wasdeemed unattractive, whereas a stimulus that was clearly tracked was consideredattractive. In this paper, clear tracking was defined as: (i) corrective meanderingabout the angular direction of the speaker (usually less than ±60°); (ii) femalefollowing of switches of the song between two loudspeakers with an angular
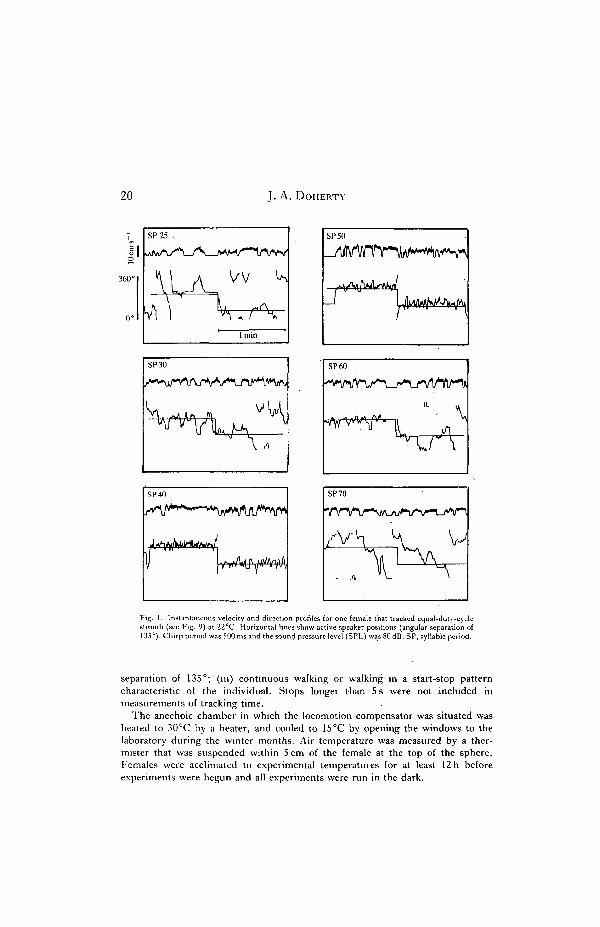
20 J. A. DoiIERTY
360
SP70
Fig. 1. Instantaneous velocity and direction profiles for one female that tracked equal-duty-cyclestimuli (see Fig. 9) at 22°C. Horizontal lines show active speaker positions (angular separation of135°). Chirp period was 500 ms and the sound pressure level (SPL) was80dB. SP, syllable period.
separation of 135°; (hi) continuous walking or walking in a start-stop patterncharacteristic of the individual. Stops longer than 5 s were not included inmeasurements of tracking time.
The anechoic chamber in which the locomotion compensator was situated washeated to 30°C by a heater, and cooled to 15°C by opening the windows to thelaboratory during the winter months. Air temperature was measured by a ther-mister that was suspended within 5 cm of the female at the top of the sphere.Females were acclimated to experimental temperatures for at least 12 h beforeexperiments were begun and all experiments were run in the dark.


Cricket stridulation and phonotaxis 23
3 0 -
•S 2 0 -
finn —^
•
700 —
J, 600-T3
Q .
S" 5 0 0 -
u
400 -
300 -
(
|
(_<
i
i
>9
i
1
t
1 •
A ,
— ( )|
1
« j
•
K
CD
• I I• • •CP
(i
ci
~i
oii
o
I•
D(
1
<1
)
w
1
)
I•
1•
1•
•o
-i[I- °
\ I r20 25 30
Temperature (°C)
Fig. 3
I I l l
- 2
1 1 115
1 1 1 120
1 1 1 1 1 1 125
Temperature (°C)Fig. 4
1 1 1 130
1 1 135
Dom
inan
t fre
quen
cy (
kHz)
I 1
I I
I I
I I
I I
I I
I I
I
3 "*
"S
_
P
»•
e e
e
o
BO
O
N
o 65
ea
•
a 8
p o o
«•
e
g —
Sylla
ble
dura
tion
(ms)
o
oI
I I
I
e t
»e e
*
a a
• o
o
t>
f5
OO
I B
&
n•a
B
o?

Cricket stridulation andphonotaxis IS
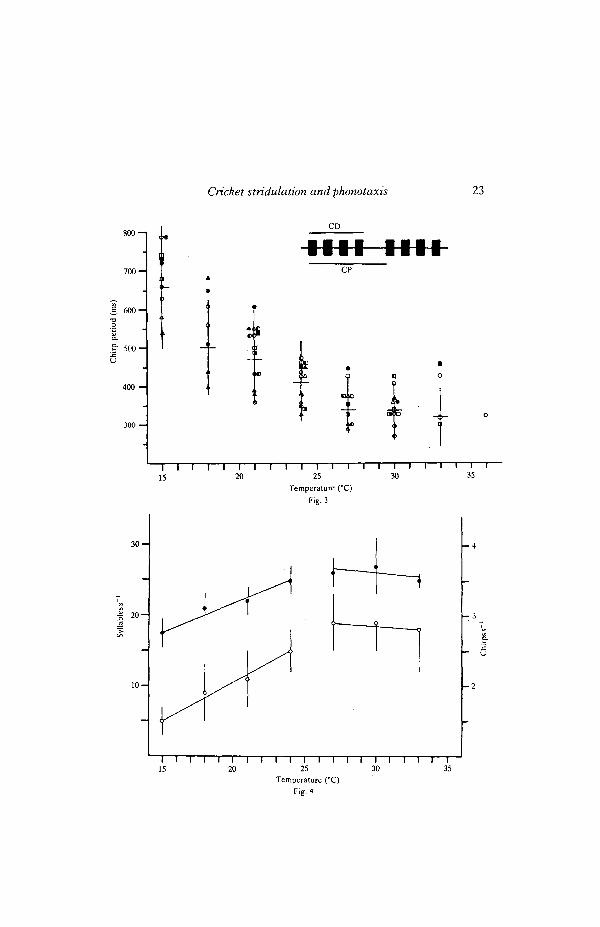
and 24°C; at higher temperatures, these temporal properties did not change withtemperature.
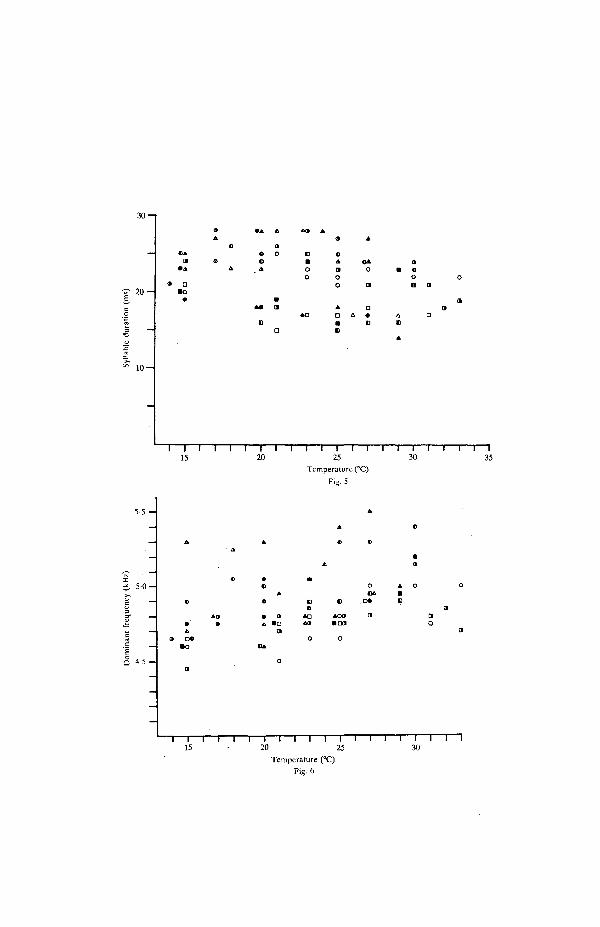
Compared with SP and CP, there were little or no effects of temperature onsyllable duration, dominant frequency and the number of syllables per chirp. Meansyllable durations of individuals ranged between 14 and 28 ms (Fig. 5) and meandominant frequencies of most individuals ranged between 4-6 and 5 0 kHz (Fig. 6).At 15, 22 and 30°C, the majority of chirps (about 70 %) contained 4 syllables. Thepercentage of 3-syllable chirps increased with increasing temperature from 10% at15°C to 25 % at 30°C, whereas the percentage of 5-syllable chirps decreased from20% at 15°C to 5% at 30°C, indicating a tendency of the male to shorten chirpduration by reducing the number of syllables per chirp.
Effect of temperature on female phonotaxis
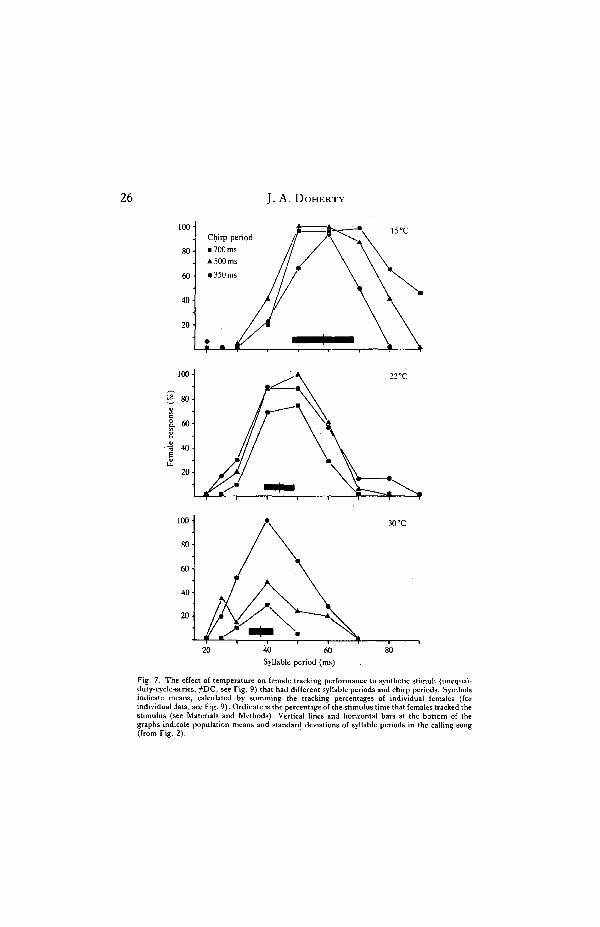
If temperature coupling occurs in G. bimaculatus, then the effect of temperatureon the tracking behaviour of females should parallel that on the calling song ofmales. That is, the SPs and CPs that elicit optimal tracking by females should shiftfrom longer to shorter values with increasing temperature to match the effect oftemperature on these temporal properties in the calling song. This appeared tooccur in G. bimaculatus and the results are summarized in Fig. 7 (individual dataare presented in Figs 8, 9). At 15°C, females tracked synthetic calling songs withSPs that ranged from 40 to 80 ms and some even tracked songs with 90ms. SPs;tracking was best when the SP was between 50 and 70 ms. This range of besttracking corresponded with the range of SPs in calling song at the same temperature(means of individual males at 15 °C ranged from 50 to 70 ms). At 22°C, femalestracked songs with SPs that ranged from 30 to 60 ms, and the best tracking occurredwhen the SP was 40—50 ms, which was similar to the range of SPs in calling songs ofmales at the same temperature (means of individual males ranged from 40 to 55 msat 21 °C). The range of SPs that elicited tracking in females shifted furtherdownward at 30°C. At this high temperature, females tracked songs with SPs thatranged from 30 to 50 ms and the best tracking occurred when the SP was 40 ms.The majority of males at 30°C produced calling songs with mean SPs that rangedfrom 36 to 42 ms.
Temperature also affected the range of CPs tracked by females (Fig. 8).Synthetic songs with CPs greater than 700 ms were tracked better at 15°C than at30°C; some females even tracked songs with CPs as long as 4 s. Among individualfemales, tracking performance at 15°C was least variable to song s1 with CPs thatranged from about 400 to 900 ms. This 'optimal' range of CPs overlapped the rangeof CPs in calling song at the same temperature (see Fig. 3 and bar in Fig. 8). At30°C, no females tracked songs with CPs greater than 900 ms; the majority failed to
Fig. 5. The effect of temperature on syllable duration in the calling song. Symbols are means ofindividuals (same individuals as in Figs 2, 3, 6). Sixteen syllables were measured per individual,and the standard deviations of individuals ranged between 0 3 and 3 8 ms.
Fig. 6. The effect of temperature on dominant (carrier) frequency of the calling song. Symbols aremeans of individuals (same individuals as in Figs 2, 3, 5). Eight measurements were averaged perindividual, and the standard deviations of individual means ranged between 10 and 400 Hz.
26 J. A. Doi-IERTY
15 X
20 40 60
Syllable period (ms)
Fig. 7. The effect of temperature on female tracking performance to synthetic stimuli (unequal-duty-cycle-series, ^DC, see Fig. 9) that had different syllable periods and chirp periods. Symbolsindicate means, calculated by summing the tracking percentages of individual females (forindividual data, see Fig. 9). Ordinate is the percentage of the stimulus time that females tracked thestimulus (see Materials and Methods). Vertical lines and horizontal bars at the bottom of thegraphs indicate population means and standard deviations of syllable periods in the calling song(from Fig. 2).

Cricket stridulation and phonotaxis 27
100-1
I 50f-
*• • a15 °C
a a D
8"
200 400 600 800 1000 1200 1600 2000 2500 4000
100-1
50-
30 °C
T * f T200 400 600 800 1000 1200 1600 2000 2500 4000
Chirp period (ms)
Fig. 8. The effect of temperature on female tracking performance to synthetic stimuli withdifferent chirp periods. Symbols indicate tracking performances (percentage of stimulus time thestimulus was tracked) of individual females at 15 and 30 °C. All stimuli had 4 syllabic chirps and20ms syllable durations. Syllable period was 60ms at 15°C and 40ms at 30°C, so the chirpdurations at these two temperatures were different (200ms at 15°C and 140ms at 30°C). Verticallines and horizontal bars at the bottoms of the graphs indicate population means and standarddeviations of chirp periods in the calling song (from Fig. 3).
track songs with CPs greater than 500-600 ms. The variability of tracking per-formance among females at 30°C was minimal to songs with CPs that rangedbetween 250 and 450 ms. This range was comparable to the range of CPs in callingsongs at 30°C (see Fig. 3 and bar in Fig. 8).
In summary, there was a good match between the range of SPs of synthetic
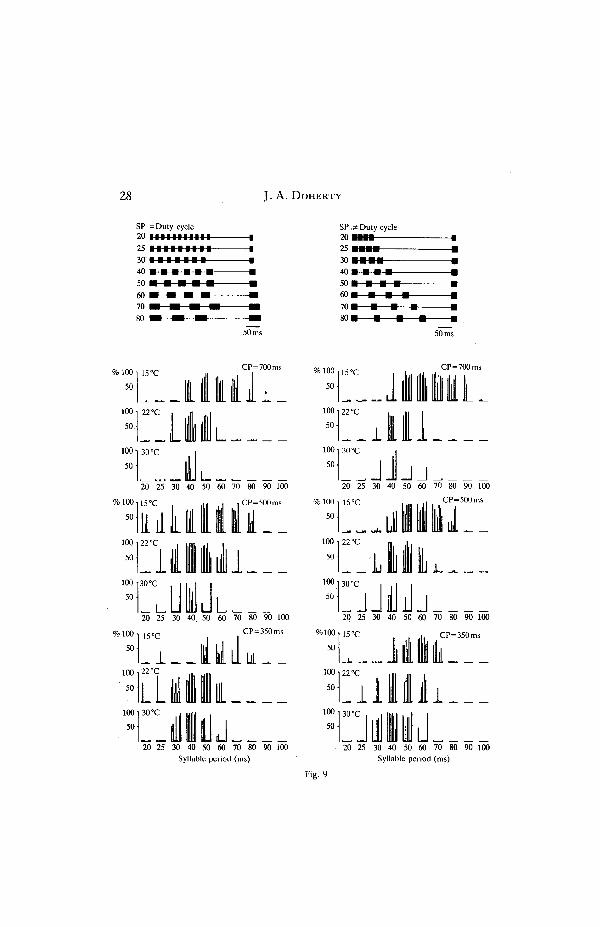
28 J. A. DOHERTY
SP =Duty cycle20 HIIIIHIII
25 • • • • • • • • •30 • • • • • • •40 • • • • • •50
60
70
80
50 ms
SP ,*Duty cycle2025
30
40 I
50 1
6 0 |
70 1
m—m-50 ms
%100
50
100
50
%100
50
100
50
100
%100
50
100
50
100
50
15 °C
22 °C
30 °C
CP=700 ms
L L
20 25 30 40 50 60 70 80 90 100
CP=500 ms
30 °CI I II.I I
I20 25 30 40. 50 60 70 80 90 100, , o ^ CP=350ms15 C
22 °C
L30 °C
20 25 30 40 50 60 70 80 90 100
Syllable period (ms)
%100
50
100
50
100
50
%100
50
100
50
100
%100
50
100
50
100
50
15 °C
22 °C
30 °
CP=700 ms
iiiiiiL
JJ20 25 30 40 50 60 70 80 90 100
15 °C CP=500ms
22 °C
_L30 °C
J20 25 30 40 50 60 70 80 90 10015 °C
22 °C
30
1
CP=350 ms
1
20 25 30 40 50 60 70 80 90 100
Syllable period (ms)
Fig. 9 :

Cricket stridulation and phonotaxis 29
calling song that elicited optimal phonotaxis by females and the range of SPs innatural calling songs emitted by males at the same temperature (Figs 7, 9). Thismatch between sender and receiver was less evident for CP, although at hightemperatures, songs with long CPs were not tracked well compared to songs withshorter CPs (Figs 7, 8, 9). Females appeared to track songs with short CPs equallywell at 15 and 30°C, but one must realize that the songs at these two temperatureshad optimal SPs (SPs adjusted to these temperatures, see Materials and Methods).Differences in tracking performances to songs with different CPs may have beenmore apparent at the margins of the SP-range; that is, if only 'marginally-attractive'syllable periods were used (i.e. at 40ms SP at 15°C).
Female phonotaxis and the interaction of calling song properties
Interaction of chirp period and syllable period
Chirp period affected the range of syllable periods that females tracked atdifferent temperatures (Fig. 7). In general, the range of SPs tracked by females wasbroad if the CP was within the range of CPs of calling song at this temperature, andnarrower when the CP was at the margins or even outside this natural range. CPhad little effect on the tracking of songs with SPs in the centre of the female'sresponse range, whereas this effect was most evident at the margins of the SP range.This is seen best when one compares female tracking at 15 and 30°C (Fig. 9). At15 °C, the range of SPs tracked by females was broad (40-80 ms) when songs hadlonger CPs of 500 and 700 ms. This range contracted to 50-60 ms ( = DC stimuli)and to 50-70 ms (^DC stimuli) when the CP was shortened to 350 ms. At 30°C, therange of SPs tracked by females was broad (30-50 ms) when stimuli had a short CPof 350 ms (characteristic of calling song at 30°C) and was narrower (40ms) whenstimuli had longer CPs of 500 and 700 ms. At both temperatures, the trackingperformances of most females\to songs with optimal SPs remained unchanged,regardless of what CP was used. In some females, however, there was a clearreduction in the tracking performance to songs with optimal SPs when CPs wereshort (15°C) and long (30°C) (see Fig. 9).
Interaction of syllable period and chirp duration
Female tracking performance to songs with different SPs also appeared to bedependent upon chirp duration (or the number of syllables per chirp, but see J. A.Doherty, in preparation). As for CP, the interaction between SP and chirp duration
Fig. 9. The effect of temperature on tracking performances of seven females to synthetic stimuliwith different syllable periods (SP) and chirp periods (CP). The graphs on the left summarize theresponses of individuals to equal-duty-cycle stimuli (top left) and the graphs on the rightsummarize the responses to unequal-duty-cycle stimuli (top right). The timing relationship for aCP of 350 ms is shown in the diagrams of the synthetic stimuli. The ordinates of the graphs indicatethe percentages of the stimulus times that females, tracked the stimulus (see Methods). Barsindicate the tracking scores of individual females, and the responses of individuals can be followedover different SPs, CPs and stimulus paradigms. For example, the responses of female number 3can be followed by looking at the third bar from the left for each stimulus. Closed circles indicate a0 % tracking score. No bars or points indicate that the female was not tested with the stimulus.Playback levels of stimuli were 80dB SPL, except for the following females: 4 (70dB at 22 and30°C), 5 (85 dB), 6 (85 dB at 15 and 22°C), 7 (85 dB).

30 J. A. DOHERTY
(CD) was most evident at the margins of the SP range; CD had little effect ontracking performance to songs with SPs in the middle of this range. The interactionbetween SP and CD (or SN) is seen best when the results from the =DC and^DCexperiments are compared (Fig. 9). At 15°C and with a CP of 700 ms, most femalestracked songs with SPs ranging from 40 to 70 ms in the = DC experiment, and from50 to 80 ms in the^DC experiment. The differences in these results were in thetracking of stimuli with 40 ms and 80 ms SPs. The differences in the trackingperformance to stimuli with 80 ms SPs may have been due to different syllablenumbers (3 versus 4) and different syllable durations (40 ms versus 20 ms) in thestimuli (Fig. 9). Stimuli with 40 ms SPs in the =DC and ^DC experiments differedonly in the number of syllables per chirp (6 versus 4 syllables, see stimuli in Fig. 9),so females apparently tracked longer duration chirps (or chirps with greater SNs)better than shorter duration chirps at 15°C. Furthermore, when one comparestracking performances to songs with shorter SPs in the = DC and^DC experiments(15°C, CP 500ms, SPs 20-30 ms), some females tracked longer duration chirpsbetter than shorter duration chirps that had the same SP.
Further comparisons of =DC and ^DC experiments at 22°C reveal that chirpswith longer durations elicited better tracking than shorter duration chirps (Fig. 9).When the CP was 500 ms or 350 ms and the SP was 30 ms, females tracked longerduration chirps (7 syllables, =DC stimulus) better than shorter duration chirps (4syllables, ^DC stimulus). This improved tracking performance was probably notdue to differences in total sound energy because the sound energy per chirp wasgreater in the shorter duration chirp (^DC stimulus, 67 % duty cycle) compared tothat in the longer duration chirp ( = DC stimulus, 50% duty cycle).
DISCUSSION
The effect of temperature on calling song
Syllable and chirp repetition rates
Temperature affected syllable (pulse) repetition rate (PR) and chirp repetitionrate (CR) in the calling song of G. bimaculatus only below 24°C (Fig. 4). The effectof temperature on PR was small compared to what has been reported in trillingspecies of crickets (Walker, 1962a,b, 1963). The PR in the calling song of trillingcrickets rose linearly between 15 and 35 °C, and PRs of individual species rangedbetween 25 and lOOpulsess"1 (see Walker, 19626). In G. bimaculatus, mean PRsof individuals were considerably lower than those of trilling crickets at comparabletemperatures and ranged from 14 to 33pulsess~' (Fig. 4). Similar ranges of PRshave been reported in other chirping species of Gryllus, including G. pennsyl-vanicus (Alexander, 1957) and G. campestris (Kutsch, 1969; Kriechbaum, 1983).These findings agree with those of Walker (19626), which showed that the effect oftemperature on pulse rate was greater in species that had higher PRs at a giventemperature.
The effect of temperature on chirp repetition rate in the calling song of G.bimaculatus was comparable to that on CR in other chirping crickets. In G.bimaculatus, mean CRs of individuals ranged from 12 to 3 9 chirps s"1 between 15and 33°C (Fig. 4). This range of CRs was similar to that in the chirping tree cricket,

Cricket stridulation and phonotaxis 31
Oecanthus fultoni; CRs of individuals ranged from about 18 to 3-8chirpss~'between 15 and 32°C (Walker, 1962a,6). In the field cricket, G. campestris, CRsranged from about 2-5 to 41chirpss~1 between 20 and 30°C (Kutsch, 1969).
The PR and CR in the calling songs of many species of crickets and katydidsincrease linearly with increasing temperature over the entire temperature rangeexperienced by these animals (Walker, 1962a,6, 1963, 1975). In G. bimaculatus,the effects of temperature on PR and CR were also linear, although this linearitywas not observed over the entire temperature range. Instead, the linear effects oftemperature on PR and CR were evident only between 15 and 24°C and thesetemporal properties reached a maximum and did not change with temperatureabove 24°C (Fig. 4). Similar results have been reported in other chirping species ofGryllus, such as G. pennsylvanicus and G. campestris (Alexander, 1957; Kutsch,1969; Kriechbaum, 1983), although temperature-independence of calling songtemporal properties at high temperatures is not universal in chirping crickets. In thechirping tree cricket, O. fultoni, for example, the effects of temperature on PR andCR were linear between 18 and 32°C (Walker, 1962a). Indeed, the linear corre-lation of CR with temperature is so good that this cricket has been called the'thermometer cricket' because one can determine the air temperature by countingthe chirps in a specified time (see Walker, 1962a, for earlier references).
The mechanisms underlying temperature-dependence in trilling and chirpingtree crickets and temperature-independence in G. bimaculatus and other chirpingspecies of Gryllus at high temperatures remain unclear. At high temperatures,sources for rate saturation (i.e. no further increase in PR and CR) in the callingsongs of chirping Gryllus may include mechanical as well as physiological con-straints on song pattern generation. More insights into this problem await theresults of studies on the mechanisms underlying stridulation at differenttemperatures.
Syllable duration and dominant frequency
Temperature had little or no effect on syllable duration in the calling song of G.bimaculatus (Fig. 5). Comparable results have been reported in trilling species oftree crickets {Anurogryllus and Oecanthus sp., Prestwich & Walker, 1981) and inthe chirping species, G. campestris (Kutsch, 1969). These results indicate that thewing closing phase during stridulation is little affected by changes in temperatureand that the increase of PR results primarily from shortening the silent, openingphase of the wings. This interpretation is supported by the results of studies onwing movements and electromyogram activity during stridulation at different tem-peratures (Kutsch, 1969; Volleth, 1981).
Compared with other temporal properties (PR, CR), there was little or no effectof temperature on dominant frequency in the calling song of G. bimaculatus (Fig.6). This result may be due to dominant frequency being determined by mechanicalresonance of the wings, which is also relatively unaffected by temperature (Koch,1980; C. J. H. Elliott and U. T. Koch, in preparation). Variable effects oftemperature on dominant frequency have been described in several subfamilies ofcrickets (see Walker, 19626; Volleth, 1981; Prestwich & Walker, 1981). In G.rubens and A. arboreus, this effect was slight and comparable to that in G.

32 J. A. DOHERTY
bimaculatus. Stronger influences of temperature on dominant frequency occurredin some species of Oecanthus and Nemobius, but in other species, the dominantfrequency reached a plateau at high temperatures (Walker, 19626).
The effect of temperature on phonotaxis
The effect of temperature on female phonotaxis paralleled the effect of tem-perature on male stridulation in G. bimaculatus. As far as syllable repetition rateand chirp repetition rate were concerned, the female's response was matched to themale's signal at temperatures between 15 and 30°C. This matching of sender andreceiver at different temperature has been termed 'temperature coupling'(Gerhardt, 1978), and these results agree with those in studies of othertemperature-coupled communication systems. Temperature coupling has beendemonstrated in acoustic communication in tree crickets, grasshoppers and tree-frogs (e.g. Walker, 1957; Helversen, 1972; Helversen & Helversen, 1981;Skovmand & Pedersen, 1983; Gerhardt, 1978, 1982, 1983; H. C. Gerhardt and J.A. Doherty, in preparation), and in visual communication in fireflies (e.g. Carlsonet al. 1976). In all of these studies, the effects of temperature on signal generation inthe sender are paralleled by similar effects of temperature on signal recognition inthe receiver. Temperature coupling usually occurs over the entire temperaturerange, although in some cases, coupling appears to break down at higher tem-perature (see Gerhardt, 1978; Skovmand & Pedersen, 1983).
Signal production and recognition systems
The existence of temperature coupling is consistent with the predictions of thegenetic coupling hypothesis (Hoy, 1974). This hypothesis states that there areneural elements (individual neurones or networks of neurones) common to bothpattern generation and pattern recognition. These common elements would bespecified by the same genes and may exist in the form of neural filters-or sensory-motor templates (Hoy, 1978). Besides temperature coupling phenomena, indirectevidence for genetic coupling also comes from studies of interspecific hybridizationin crickets and treefrogs (Hoy et al. 1977; Doherty & Gerhardt, 1983, 1984).Evidence against genetic coupling comes primarily from studies of interspecifichybridization in acridid grasshoppers (Helversen & Helversen, 1975a,b; for areview see Eisner & Popov, 1978).
At present, neither the results of hybridization studies nor the temperaturecoupling phenomena observed in behaving animals provides conclusive evidence foror against the existence of common neural elements in sender and receiver (seeDoherty & Gerhardt, 1984). More direct morphological and physiological analysesare needed of the neural elements involved with pattern generation and patternrecognition. Studies of the neural correlates of temperature coupling phenomenamay provide new insights. As expressed by Gerhardt (1978), temperature couplingcould result either from temperature having a single effect on the neural networkcommon to both pattern generation and pattern recognition, or from temperatureaffecting completely different neural networks in the same way. Considering theaccuracy of sender-receiver matching at different temperatures in several diverse

Cricket stridulation and phonotaxis 33
communication systems, the common network idea is the most parsimonious inexplaining temperature coupling phenomena. Obviously, the falsification of eitherone of these possibilities will require new investigations into how and where in theanimal's nervous system temperature is affecting pattern generation and patternrecognition.
Simplicity versus complexity in calling song recognition
A male cricket can convey individual and species information to the female viaseveral temporal and spectral properties of its calling song. The results ofbehavioural studies make it clear that some properties of the calling song are moreimportant than others in triggering recognition and eliciting phonotaxis in femalecrickets (e.g. Zaretky, 1972; Hill, 1974a,6; Popov & Shuvalov, 1977; Pollack &Hoy, 1979, 1981; Weber et al. 1981; Thorson et al. 1982; Stout, DeHaan &McGhee, 1983). Furthermore, within the auditory pathway, neurones have beenrecorded and partly identified that encode spectral and temporal properties of thecalling song (for reviews see Eisner & Popov, 1978; Huber, 1983; Boyan, 1984),although elements involved in the recognition process have not been identified untilrecently (Schildberger, 1984).
Recent results obtained by studying females of G. campestris on a walkingcompensator show that the syllable repetition rate of around 30 Hz is the 'chief -and in some cases both necessary and sufficient - property' of the calling song thattriggers recognition and phonotaxis in females (Thorson et al. 1982). Thorson etal. formulated the '30-Hz hypothesis' for calling song recognition, which states thatthe 30-Hz syllable modulation is the only feature required for recognition, as long asthe carrier frequency is audible. This hypothesis reflects 'simplicity' in calling songrecognition in chirping crickets because it implies that as long as the 30-Hzmodulation is present other temporal properties, such as chirp duration and chirprepetition rate, are not essential in the recognition process.
In G. bimaculatus, calling song recognition appears to be more complex (see alsoPopov, Shuvalov, Svetlogorskaya & Markovich, 1974; Stout et al. 1983). Theattractiveness of the calling song, as expressed by the tracking performance of thefemale on a locomotion compensator, was dependent upon the evaluation of severaltemporal properties. That is, female tracking of stimuli with 'unattractive' syllableperiods (i.e. at the margins of the effective range for tracking) was turned on or offby changing the values of supposedly 'non-essential' temporal properties, such aschirp period and chirp duration (Figs 7, 9). The influence of different temporalproperties on the attractiveness of the calling song in G. bimaculatus reflects whatcould be called a 'trade-off, which expresses the relative weightings of combinedparameters of the calling song in the recognition process. Hints for such trade-offphenomena 'in the recognition process in G. bimaculatus are evident at all tem-peratures (Figs 7, 9; see also J. A. Doherty, in preparation). If one temporalproperty is 'unattractive', the attractiveness of the entire stimulus can be maintainedby raising the attractiveness of other temporal properties. Trade-off effects infemales may well serve to 'buffer' the recognition process against differences in thecalling song that may result from males singing at different temperatures, singing atdifferent chirp repetition rates, or from different amounts of environmental degra-

34 J. A. DOHERTY
dation of the physical properties of the calling song during signal transmissionbetween sender and receiver.
This research was supported by a postdoctoral stipendium from the Max-Planck-Society. I would like to thank especially Professor Franz Huber for his help andencouragement throughout this research. I also wish to thank Drs T. Weber andH.-U. Kleindienst for technical help and valuable discussions and criticisms.Professor F. Huber and Drs T. Weber and J. Thorson provided useful commentson the manuscript. I also thank H. Bamberg for help with the figures and L. Simonfor typing the manuscript.
R E F E R E N C E S
ALEXANDER, R. D. (1957). The taxonomy of the field crickets of the Eastern United States (Orthoptera:Gryllidae: Acheta). Ann. enl. Soc. Am. 50, 584-602.
ALEXANDER, R. D. & MERAL, G. H. (1967). Seasonal and daily chirping cycles in the northern spring and fallfield crickets, Gryllus veletis and G. penmylvanicus. OhioJ. Sci. (ti, 200-209
BOYAN, G. S. (1984). Neural mechanisms of auditory information processing by identified interneurones inorthoptera. J. Insect Physiol. 30, 27-41.
CARLSON, A. D., COPELAND, J.. RADERMAN, R. & BULLOCH, A. G. M. (1976). Role of interflash intervals in
a firefly courtship (Photinus macdermotti). Anim. Behav. 24, 786-792.DOHERTY, J. A. & GERHARDT, H. C. (1983). Hybrid treefrogs: vocalizations of males and selective
phonotaxisof females. Science, N.Y. 220, 1078-1080.DOIIISR i'Y, J. A. & GERHAROT, H. C. (1984). Acoustic communication in hybrid treefrogs: sound production
by males and selective phonotaxis by females. J. comp. Physio!. 154, 319-330.ELSNER, N. & POPOV, A. V. (1984). Neuroethology of acoustic communication. Adv. Insect Phvsiol. 13,
229-335.GERHARDT, H. C. (1978). Temperature coupling in the vocal communication system of the gray treefrog,
Hyla versicolor. Science, N.Y. 199, 992-994.GERHARDT, H. C. (1982). Sound pattern recognition in some North American treefrogs (Anura: Hylidac):
implications for mate choice. Am. Zool. 22, 581-595.GERHARDT, H. C. (1983). Acoustic communication in treefrogs. Verh. dt. zool. Ges. 1983, 25-35.HELVERSEN, D. VON (1972). Gesang des Mannchens und Lautschema des Weibchens bei der
Feldheuschrecke Chorthippus biguttulus (Orthoptera, Acrididae).7- comp. Physiol. 81, 381-422.HELVERSEN, D. VON & HELVERSEN, O. VON (1975a). Verhaltensgenetische Untersuchungen am akustischen
Kommunikationssystem der Fcldheuschrecken. I. Der Gesang von Artbastarden zwischen Chorthippusbiguttulus und Chorthippus mollis. J. comp. Physiol. 104, 273-299.
HELVERSEN, D. VON & HELVERSEN, O. VON (19756). Verhaltensgenetische Untersuchungen am akustischenKommunikationssystem der Feldheuschrecken. II. Das Lautschema von Artbastarden zwischen Chorthip-pus biguttulus und Chorthippus mollis. J. comp. Physiol. 104, 301-323.
HELVERSEN, D. VON & HELVERSEN, O. VON (1981). Korrespondenz zwischen Gesang auslosendem Schemabei Feldheuschrecken. Nova Ada Acad. Caesar. Leop. Carol. Leopoldina 51, 449-462.
HELVERSEN, O. VON (1979). Angeborenes Erkennen akustischer Schlussclreize. Verh. dt. zool. Ges. 1979,42-59.
HILL, K. G. (1974a). Acoustic communication in the Australian field crickets Teleogryllus commodus and T.oceanicus (Orthoptera Gryllidae). Ph.D. thesis, University of Melbourne.
HILL, K. G. (19746). Carrier frequency as a factor in phonotactic behavior of female crickets (Teleogrylluscommodus). J. comp. Physiol. 93, 7-18.
HOY, R. R. (1974). Genetic control of acoustic behavior in crickets. Am. Zool. 14, 1067-1080.HOY, R. R. (1978). Acoustic communication in crickets: a model system for the study of feature detection.
Fedn Proc. Fedn Am. Socs exp. Biol. 37, 2316-2323.HOY, R. R., HAHN, J. & PAUL, R. C. (1977). Hybrid cricket auditory behavior: evidence for genetic coupling
in animal communication. Science, N.Y. 195, 82-83.HUBER, F. (1983). Neural correlates of orthopteran and cicada phonotaxis. In Neurobiology and Behavioral
Physiology: Roots and Growing Points, (eds F. Huber & H. Markl), pp. 108-135. New York: Springcr-Verlag.
KOCH, U. T. (1980). Analysis of cricket stridulation using minature angle detectors. J. comp. Physiol. 136,247-256.

Cricket stridulation and phonotaxis 35
KRIECHBAUM, W. (1983). Zeitstruktur des Lockgesanges bei Gtyllus campestris L. Diplomarbeit, UniversitatTubingen.
Kui'SCH, W. (1969). Ncuromuskulare Aktivitat bei verschiedenen Verhaltensweisen von drei Grillcnartcn.Z. vergl. Physiol. 63, 335-378
POLLACK, G. S. & HOY, R. R. (1979). Temporal patterns as a cue for species-specific calling song recognitionin crickets. Science, N.Y. 204, 429-432.
POLLACK, G. S. & HOY, R. R. (1981). Phonotaxis to individual rhythmic components of a complex cricketcalling song. JT. comp. Physiol. 144, 367-373.
POPOV, A. V. & SHUVALOV, V. F. (1977). Phonotactic behavior of crickets. J. comp. Physiol. 119, 111-126.Popov, A. V., SHUVALOV, V. F., SVETLOGORSKAYA, I. D. & MARKOVICH, A. M. (1974). Acoustic behaviour
and auditory system in insects. In Mechanoreception Symposium, (ed. J. Schwartzkopff), pp. 281-306.Abh. RheinischWestfal Akad Wiss 53.
PRESTWICH, K. N. & WALKER, T. J. (1981). Energetics of singing in crickets: effect of temperature in threetrilling species (Orthoptera: Gryllidae). J. comp. Physiol. 143, 199-212.
SCHILDBERGER, K. (1984). Temporal selectivity of identified auditory neurons in the cricket brain. J. comp.Physiol. 155, 171-185.
SCHMITZ, B., SCHARSTEIN, H. & WENDLEK, G. (1982). Phonotaxis in Gryllus campestris L. (Orthoptera,Gryllidae). I. Mechanisms of acoustic orientation in intact female crickets. J. comp. Physiol. 148, 431-444.
SCHNEIDER, H. (1977). Acoustic behavior and physiology of vocalization in the European tree frog Hylaarborea (L.). In The Reproductive Biology of Amphibians, (eds D. H. Taylor & S. I. Guttman), pp.295-335. New York: Plenum Press.
SKOVMAND, O. & PEDERSEN, S. B. (1983). Song recognition and song pattern in a shorthorned grasshopper.J . comp. Physiol. 153, 393-401.
STOUT, J. F., DEHAAN, C. H. & MCGHEE, R. W. (1983). Attractiveness of the male Acheta domesticuscalling song to females. I. Dependence on each of the calling song features. J . comp. Physiol. 153, 509-521.
THORSON, J., WEBER, T . & HUBER, F. (1982). Auditory behavior of the cricket. I I . Simplicity of calling-songrecognition in Gryllus, and anomalous phonotaxis at abnormal carrier frequencies, j?. comp. Phvsiol. 146,361-378.
VOLLETH, M. (1981). Fliigelbewegung und Gesang bei Laubheuschrecken und Grillen (Orthoptera,Ensifera) in Abhangigkeit von der Temperatur. Diplomarbeit, Universitat Erlangen.
WALKER, T. ] . (1957). Specificity in the response of female tree crickets (Orthoptera, Gryllidae,Oecanthinae) to calling songs of the males. Ann. ent. Soc. Am. 50, 626-636.
WALKER, T . J. (1962a). The taxonomy and calling songs of United States tree crickets (Orthoptera:Gryllidae: Oecanthinae). I. The genusNeoxabea and the niveus and varicomis groups of the Oecanthus.Ann. ent. Soc. Am. 55, 303-322.
WALKER, T. J. (19626). Factors responsible for intraspecific variation in the calling songs of crickets.Evolution 16, 407-428.
WALKER, T. J. (1963). The taxonomy and calling songs of United States tree crickets (Orthoptera: Gryllidae:Oecanthinae). I I . The nigricomis group of the genus Oecanthus. Ann. ent. Soc. Am. 56, 772-789.
WALKER, T . J. (1975). Effects of temperature on rates in poikilotherm nervous system. Evidence from thecalling songs of meadow katydids (Orthoptera: Tettigoniidae: Orchelimum) and reanalysis of publisheddata. J . comp. Physiol. 101, 57-69.
WEBER, T., THORSON, J. & HUBER, F. (1981). Auditory behavior of the cricket. I. Dynamics of compensatedwalking and discrimination paradigms on the Kramer treadmill. J. comp. Physiol. 141, 215-232.
WENDLER, G., DAMBACH, M., SCHMITZ, B. & SCHARSTEIN, H. (1980). Analysis of the acoustic orientation
behavior in crickets Gryllus campestris (L.). Naturwissenschaften 67, 99-100.ZARETSKY, M. D. (1972). Specificity of the calling song and short term changes in the phonotactic response
by female cricket Scapsipedus marginatus (Gryilidae). J. comp. Physiol. 79, 153-172.











![[45 ] THE QUANTITATIVE NUTRITIONAL …jeb.biologists.org/content/jexbio/33/1/45.full.pdf · Quantitative nutritional requirements of Drosophila melanogaster 47 spores, and the fluctuations](https://static.fdocuments.in/doc/165x107/5ac1f6ec7f8b9a4e7c8db233/45-the-quantitative-nutritional-jeb-nutritional-requirements-of-drosophila.jpg)






