
TABLE OF CONTENTS - file.scirp.orgfile.scirp.org/pdf/JBC_SAMPLE-01-01-20091203111403.pdf · Journal...
47
Journal of Biophysical Chemistry, 2009, 1, 1-45 Copyright © 2009 SciRes. JBC TABLE OF CONTENTS Volume 1 Number 1 November 2009 Decapod crustacean chelipeds: an overview P. MARIAPPAN, C. BALASUNDARAM, B. SCHMITZ…………………………………………………………1 The structure of the nasal chemosensory system in squamate reptiles. 2. Lubricatory capacity of the vomeronasal organ S. J REHOREK, B. T. FIRTH, M. N. HUTCHINSON…………………………………………………………14 Enhanced expression of a calcium-dependent protein kinase from the moss Funaria hygrometrica under nutritional starvation D. MITRA, M. M. JOHRI………………………………………………………………………………………24 Domain III function of Mu transposase analysed by directed placement of subunits within the transpososome S. MARICONDA, S.-Y. NAMGOONG, K.-H. YOON, H. JIANG, R. M. HARSHEY…………………………32
Transcript of TABLE OF CONTENTS - file.scirp.orgfile.scirp.org/pdf/JBC_SAMPLE-01-01-20091203111403.pdf · Journal...

Journal of Biophysical Chemistry, 2009, 1, 1-45
Copyright © 2009 SciRes. JBC
TABLE OF CONTENTS
Volume 1 Number 1 November 2009 Decapod crustacean chelipeds: an overview
P. MARIAPPAN, C. BALASUNDARAM, B. SCHMITZ…………………………………………………………1
The structure of the nasal chemosensory system in squamate reptiles.
2. Lubricatory capacity of the vomeronasal organ
S. J REHOREK, B. T. FIRTH, M. N. HUTCHINSON…………………………………………………………14
Enhanced expression of a calcium-dependent protein kinase from the moss Funaria
hygrometrica under nutritional starvation
D. MITRA, M. M. JOHRI………………………………………………………………………………………24
Domain III function of Mu transposase analysed by directed placement of subunits
within the transpososome
S. MARICONDA, S.-Y. NAMGOONG, K.-H. YOON, H. JIANG, R. M. HARSHEY…………………………32

Journal of Biophysical Chemistry (JBC)
Journal Information
SUBSCRIPTIONS
The Journal of Biophysical Chemistry (Online at Scientific Research Publishing, www.SciRP.org) is published
quarterly by Scientific Research Publishing, Inc.,USA.
E-mail: [email protected]
Subscription rates: Volume 2 2009 Print: $50 per copy.
Electronic: free, available on www.SciRP.org.
To subscribe, please contact Journals Subscriptions Department, E-mail: [email protected]
Sample copies: If you are interested in subscribing, you may obtain a free sample copy by contacting Scientific
Research Publishing, Inc at the above address.
SERVICES
Advertisements
Advertisement Sales Department, E-mail: [email protected]
Reprints (minimum quantity 100 copies)
Reprints Co-ordinator, Scientific Research Publishing, Inc., USA.
E-mail: [email protected]
COPYRIGHT
Copyright© 2009 Scientific Research Publishing, Inc.
All Rights Reserved. No part of this publication may be reproduced, stored in a retrieval system, or transmitted, in
any form or by any means, electronic, mechanical, photocopying, recording, scanning or otherwise, except as
described below, without the permission in writing of the Publisher.
Copying of articles is not permitted except for personal and internal use, to the extent permitted by national
copyright law, or under the terms of a license issued by the national Reproduction Rights Organization.
Requests for permission for other kinds of copying, such as copying for general distribution, for advertising or
promotional purposes, for creating new collective works or for resale, and other enquiries should be addressed to
the Publisher.
Statements and opinions expressed in the articles and communications are those of the individual contributors and
not the statements and opinion of Scientific Research Publishing, Inc. We assumes no responsibility or liability for
any damage or injury to persons or property arising out of the use of any materials, instructions, methods or ideas
contained herein. We expressly disclaim any implied warranties of merchantability or fitness for a particular
purpose. If expert assistance is required, the services of a competent professional person should be sought.
PRODUCTION INFORMATION
For manuscripts that have been accepted for publication, please contact:
E-mail: [email protected]

Keywords. Allometry; autotomy; chela display; cheliped; claw; handedness; regeneration
Decapod crustacean chelipeds: an overview
PITCHAIMUTHU MARIAPPAN, CHELLAM BALASUNDARAM and BARBARA SCHMITZ
The structure, growth, differentiation and function of crustacean chelipeds are reviewed. In many decapod crusta-ceans growth of chelae is isometric with allometry level reaching unity till the puberty moult. Afterwards the same trend continues in females, while in males there is a marked spurt in the level of allometry accompanied by a sud-den increase in the relative size of chelae. Subsequently they are differentiated morphologically into crusher and cutter making them heterochelous and sexually dimorphic. Of the two, the major chela is used during agonistic encounters while the minor is used for prey capture and grooming. Various biotic and abiotic factors exert a negative effect on cheliped growth. The dimorphic growth pattern of chelae can be adversely affected by factors such as parasitic infection and substrate conditions. Display patterns of chelipeds have an important role in agonistic and aggressive interactions. Of the five pairs of pereiopods, the chelae are versatile organs of offence and defence which also make them the most vulnerable for autotomy. Regeneration of the autotomized chelipeds imposes an additional energy demand called “regeneration load” on the incumbent, altering energy allocation for somatic and/or reproductive processes. Partial withdrawal of chelae leading to incomplete exuvia-tion is reported for the first time in the laboratory and field in Macrobrachium species.
1. General morphology
Chelipeds of decapod crustaceans have attracted human curiosity and fired human imagination since Aristotle (Hopkins 1993) probably because they figure so promi-nently both in structure and function in the life of these animals. Crustaceans are mostly aquatic arthropods which breathe through gills, have two pairs of antennae, and numerous paired appendages on thorax and abdomen (Stebbing 1893; Schmitt 1965) that are grouped into cepha-lic, thoracic and abdominal appendages in relation to the body tagmata. The cephalic and thoracic regions are usu-ally fused to form a cephalothorax and the appendages are known as cephalo-thoracic appendages. Decapod append-ages are the best example of serial homology with a serial modification in basic structure from the first to the last walking leg (Wood and Wood 1932). With the exception of the antennules, which are uniramous, other appendages are basically biramous and possess a basal segmented protopod with a coxa and basis and may have lateral
(exites) or medial (endites) protrusions (Manton 1977; McLaughlin 1982). From the protopod arise the exopod and endopod. Of the two, the latter has undergone a variety of specialisations resulting in its transformation for vari-ous functions like sensory reception, feeding, walking, burrowing and swimming while the exopod is drastically reduced or may even be lost. This has further been facili-tated by mineralisation of the exoskeleton endowing rigi-dity and support to the appendages which are made flexible by the arthrodial membrane. All decapods usually have five pairs of well developed walking legs with exceptions in the sergistid family of the Dendrobranchiata, many of the Anomura, and a few Brachyura. In these animals the fifth or fourth and fifth pair of pereiopods are reduced in size for special func-tions. Occasionally both pairs may be vestigial or absent (McLaughlin 1982). Structural modifications of decapod appendages due to diversified functions and life style have been described in different groups (Tiegs and Manton 1958; Kaestner 1970; Schram 1978). The major modifications
Journal of Biophysical Chemistry, 2009, 1, 1-13
of the appendages have evolved essentially from the feeding habits of the groups (either for filtering or for predation). However, the functional and structural modifi-cations of crustacean chelipeds are not only due to feed-ing and locomotion (Dahl 1956; Bock and von Wahlert 1965; Manton 1977) but also change with environmental conditions (Smith and Palmer 1994) and species specific needs supplemented by hox genes (Averof and Patel 1997). Among decapods the chelate legs are unique with the first (Brachyura) or the second pair (among Macrura with first two chelate pereiopods) being the sole organ of offence and defence. The typical cheliped or ambulatory pereiopod com-prises of an exopod and endopod of which the latter is highly reduced or lost. The exopod comprises seven podomeres: (i) coxa, a short, stout cylinder, moving anterio- posteriorly in articulation with the sternum and epimeron, (ii) basis, a short cylinder with lesser average diameter than the coxa and articulated by hinge joints with the coxa, moving dorso-ventrally, (iii) ischium, larger and wider than the basis, fused immovably and curved upward, (iv) merus, (v) carpus, (vi) propodus, and (vii) dac-tyl. The basis and ischium when fused together form the basi-ischium (Lochhead 1961). Typically Caridean shrimps have chelate or subchelate first and second pairs of pereiopods; however, among the Processidae often only one pereiopod of the first pair is chelate, while the opposing member is simple. A very unique and exaggerated development of one chela of the first pair of pereiopod is characteristic of snapping shrimp of the family Alpheidae. After being cocked in the open position and building up tension, this large snapper claw (of up to half the animal’s size) closes rapidly forming a thin water jet; its high velocity (25 m/s) results in the formation of a small cavitation bubble (3⋅5 mm in dia-meter), that collapses with an extremely loud and short sound (up to 248 dB re 1 µPa at 1 cm distance for 240 ns) (Schmitz 2000). This signal is used for intraspecific, hydrodynamic communication (Herberholz and Schmitz 1998) as well as for territorial defence and to stun or even kill small prey (small shrimp, crabs or fish) (MacGinitie and MacGinitie 1949; Hazlett and Winn 1962; Schultz et al 1998). Marked asymmetry of the first pair of chelate pereiopods is also common in a number of other decapods like lobsters and fiddler crabs.
2. Growth
Crustacean growth is discontinuous since the highly min-eralised old exoskeleton is shed through a process known as ecdysis (= moulting) whenever a certain growth incre-ment is achieved, whereas growth in vertebrates is continuous. In decapods three types of allometry exist:
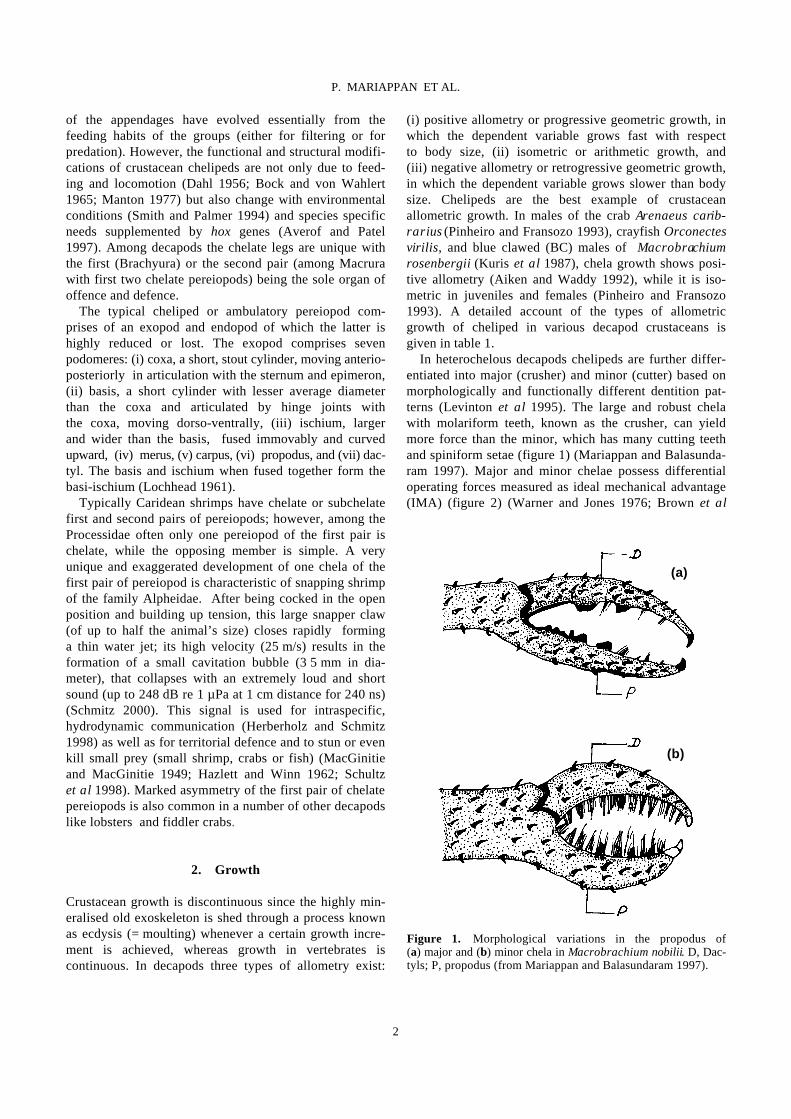
(i) positive allometry or progressive geometric growth, in which the dependent variable grows fast with respect to body size, (ii) isometric or arithmetic growth, and (iii) negative allometry or retrogressive geometric growth, in which the dependent variable grows slower than body size. Chelipeds are the best example of crustacean allometric growth. In males of the crab Arenaeus carib-rarius (Pinheiro and Fransozo 1993), crayfish Orconectes virilis, and blue clawed (BC) males of Macrobrachium rosenbergii (Kuris et al 1987), chela growth shows posi-tive allometry (Aiken and Waddy 1992), while it is iso-metric in juveniles and females (Pinheiro and Fransozo 1993). A detailed account of the types of allometric growth of cheliped in various decapod crustaceans is given in table 1. In heterochelous decapods chelipeds are further differ-entiated into major (crusher) and minor (cutter) based on morphologically and functionally different dentition pat-terns (Levinton et al 1995). The large and robust chela with molariform teeth, known as the crusher, can yield more force than the minor, which has many cutting teeth and spiniform setae (figure 1) (Mariappan and Balasunda-ram 1997). Major and minor chelae possess differential operating forces measured as ideal mechanical advantage (IMA) (figure 2) (Warner and Jones 1976; Brown et al
Figure 1. Morphological variations in the propodus of (a) major and (b) minor chela in Macrobrachium nobilii. D, Dac-tyls; P, propodus (from Mariappan and Balasundaram 1997).
(a)
(b)
2
P. MARIAPPAN ET AL.

1979). In male Uca and Alpheus , the major chela is used for aggressive and courtship displays, while the other is used for capture and manipulation of prey and grooming (Hazlett 1962; Nolan and Salmon 1970; Crane 1975). Many crustaceans have spatulate chelae which are used to
scrap algae from rocks (McLaughlin 1982). The atyid shrimps use brush and spiny setae on chelipeds to scrape up debris (Fryer 1960). In Macrobrachium australe the minor chela endowed with abundant bristles serves as a sort of net to catch prey while the major chela is used to pick up prey. Such a differential function among cheli-peds is also observed in Homarus spp. (Davis 1987). Chela size is also related to feeding habits. For instance, detritivorous crabs have small slender claws (Seed and Hughes 1995) while carnivorous counterparts like Ocy-pode spp. possess enlarged chelae to facilitate predation. In Macrobrachium nobilii, the robust second pair is used for prey capture and the slender first pair functions to deliver the food to the mouth (P Mariappan and C Balasundaram, unpublished data). The crustacean chelipeds, thus differentiated in size in otherwise bilaterally symmetrical organisms, provide a prominent example of asymmetry, which is referred to as cheliped laterality or handedness (Govind 1989). The presence of a crusher chela on the right or left side in many decapod crustaceans and deviation from a 1 : 1 ratio has been widely reported (table 2). In predatory Brachyura the presence of the major chela on the right side facilitates handling of asymmetric hard shelled molluscan prey, providing a possibility for coevolution of a predator–prey complex (Abbay-Kalio and Warner 1989; Seed and Hughes 1995). This concept however becomes untenable when the handedness changes (Ahmed 1978; Govind et al 1988). Reversal of handedness from crusher
Table 1. Allometric growth of crustacean chelipeds. Species
Sex
Allometric status
Source
Arenaeus cribrarius M + Pinheiro and Fransozo 1993 F –
Austropotamobius pallipes M + Grandjean et al 1997 F –
Cleistostoma kuwaitense M + Clayton 1990 F –
Liocarcinus depurator M + Muino et al 1999 F +
Macrophthalmus birtipes F + Barnes 1968 Macrophthalmus setosus F 0
Macrobrachium nobilii M + F 0
P Mariappan and C Balasunda-ram, unpublished
Macrobrachium rosenbergii M + Nagamine and Knight 1980 Orconectes propinquus M + Orconectes rusticus M + Garvey and Stein 1993 Orconectes virilis M +
Trapezia ferruginea M + Finney and Abele 1981 F 0 B + M, Male; F, female; B, berried; +, positive allometry; –, negative allometry; 0, isometry.
Figure 2. Ideal mechanical advantage (IMA) measurements of Macropipus depurator chelae (a) strong and (b) fast chela. The arrows show the direction through which forces F1 and F2 act. T, Tooth, N, notch (from Warner and Jones 1976).
(a)
(b)
3
P. MARIAPPAN ET AL.

to cutter and vice-versa or from pincer claw to snapper claw, when a chela is lost, has been well documented in some heterochelous crabs, lobsters, and snapping shrimp (Wilson 1903; Yamaguchi 1977; Mellon 1981; Govind 1989; Young et al 1994). In other species with plasticity in chela development into major or minor forms, the esta-blishment of laterality (handedness) is determined by eco-logical factors (Davis 1987; Smith and Palmer 1994; Goldstein and Noetzli 1997), and the reversal of handed-ness depends on the age of the animal (Cheung 1976). However, in species where there is no reversal, genetic factors determine laterality (Bush 1930; Yamaguchi 1977). Apart from functional differences, structural variations between crusher and cutter also have been elucidated (Ogonowski and Lang 1979; Ogonowski et al 1980). After autotomy, the resultant changes in the composition of chela muscles at the time of chela development, reversal, and regeneration are well documented in lob- sters (Homarus americanus), and snapping shrimp Alpheus heterochaelis (Stephens and Mellon 1979; Mellon and Stephens 1980; Govind and Lang 1981; Quigley and Mellon 1984; Govind et al 1987, 1988; Govind and Pearce 1988a, b, 1994; Govind 1989). In Gecarcinus lateralis, there is an attendant break-
down in claw muscle protein that occurs at moulting which allows the reduced claw to be drawn through the comparatively small foramen at the proximal end of the propus (Skinner 1966; Mykels and Skinner 1981). Sexual dimorphism in cheliped size has also been esta-blished in crabs (Crothers 1967), lobsters and crayfish (Snedden 1990), mantis shrimp (Schuster and Caldwell 1989), snapping shrimp (Read and Govind 1997), and freshwater prawns (Mariappan and Balasundaram 1997). Generally such a dimorphism between a cheliped pair (Darby 1934) is mainly based on size rather than form (Lee 1995) and when adjusted for size variations their functions are similar as in Ozius verreauzii (Hughes 1989). However in Alpheus heterochaelis the male pincer claw really differs in form from that of the female struc-turally (Read and Govind 1997). The development of a dimorphic pattern begins at the time of puberty moult (Hartnoll 1974; Pinheiro and Fransozo 1993, 1998), which is a prerequisite for functional sexual maturity (see e.g. Hyas lyratus, Stevens et al 1993). In some decapods the attainment of puberty moult is identified by the level of change in propodus length (e.g. Nephrops norvegicus, Farmer 1974). Differences in chela allometry are used to
Table 2. Handedness in decapod crustaceans. Species Handedness Source Calappa philargius R Ng and Tan 1985 Callinectes sapidus R Hamilton et al 1976 Carcinus maenas R Abby-Kalio and Warner 1989 Glabropilumnus laevimanus R Tweedie 1950 Globopilumnus globosus R Tweedie 1950 Heteropanope glabra R Tweedie 1950 Heterozius rotundifrons R Jones 1978 Macrobrachium nobilii R Mariappan and Balasundaram 1997 Menippe mercenaria R Cheung 1976 Necora puber R Norman and Jones 1991 Neopanope texana R Swartz 1972 Pilumnus hirtellus R Tweedie 1950 Uca lactea R Yamaguchi 1973, 1977 Uca vocans R Barnwell 1982 Uca tetragonon R Barnwell 1982 Uca formosensis R Barnwell 1982 Ocypode gaudichaudii L Trott 1987 Synalpheus brevicarpus L Herrick 1911 Alpheus dentipes – Dawes 1934 Alpheus heterochaelis – Young et al 1994 Chlorodopsis melanochira – Tweedie 1950 Homarus americanus – Herrick 1911 Macrobrachium australe – Davis 1987 Nephrops norvegicus – Farmer 1974 Ocypode quadrata – Haley 1969 Thalassina anomala – Pillai 1990 Uca formosensis – Shih et al 1999 Xantho exartus – Tweedie 1950 R, Right handed; L, left handed; –, equal distribution of right and left handed animals.
4
P. MARIAPPAN ET AL.

differentiate immature from mature phases in Pagurus prideauxi (Paulian 1936). Factors like feeding, mate-guarding, and fighting influence the development of such dimorphic patterns of chelipeds (Vermeij 1977; Hughes 1989). Parasites exert a remarkable negative effect on the growth of chelipeds in various crustaceans. Bopyrids, entoniscids and sacculinids are the common parasites known to affect the normal growth of chelipeds. Infection of a bopyrid Gyge branchialis on Upogebia littoralis and Probopyrus pandalicola on Palaemonetes, Ione thoracica on Callianassa laticauda, an entoniscid Entonella mono-ensis and a sacculinid Sacculina polygenea on Hemigrap-sus sanguinesus showed a significant reduction of chela size when compared to uninfected forms (Tucker 1930; Reverberi 1943; Morris 1948; Hartnoll 1960; Yamaguchi and Aratake 1997).
In Macrobrachium rosenbergii the development of polymorphic males is common in natural as well as com-munally cultured populations. These males are differenti-ated into (i) small males (SM), with delicate, clear or light pink claws and with a low ratio of claw to body length and much smaller than the other two morphotypes, (ii) orange-clawed males (OC) with non-spineous, often orange claws, having a higher claw to body length ratio, and (iii) blue-clawed males (BC) with blue, spineous claws and a high ratio of claw to body length. Small males can transform into blue-clawed males through orange-clawed forms in the absence of dominant BC males or when raised in isolation (Ra’anan and Cohen 1985; Kuris et al 1987). Among mature males of Pisa spp., Jassa fal-cata and Inachus leptochirus, even within the same age group there is a remarkable difference in the size and
Table 3. Variations in the percentage of limb loss in field populations of various decapod crustaceans. Species Category Per cent Source Atergatis flloridus M 41⋅30 Norman 1995 F 18⋅40 Callinectes sapidus – 24⋅80 Smith 1990a, b Cancer magister – 25⋅00 Shirley and Shirley 1988 Cancer magister – 45⋅00 Durkin et al 1984 Cancer pagurus M 13⋅20 Bennett 1973 F 9⋅90 Carcinus maenas M 12⋅50 Abello et al 1994 F 7⋅90 Carcinus maenas M* 1⋅70 Sekkelsten 1988 M** 17⋅90 Carcinus maenas M 53⋅30 McVean 1976 F 55⋅00 Chionoecetes bairdi J 34⋅60 Edwards 1972 M 43⋅00 F 23⋅00 Cyrtograpsus angulatus – 80⋅00 Spivak and Politis 1989 Homarus americanus M 44⋅40 Moriyasu et al 1999 F 61⋅30 Homarus americanus – 21⋅00 Estrella and Armstrong 1994 Homarus americanus M 40⋅00 Briggs and Mushacke 1979 F 30⋅20 Macrobrachium nobilii J 10⋅90 M 15⋅22
Mariappan and Balasundaram 1999b
F 22⋅30 Necora puber J 23⋅00 Norman and Jones 1991 M 32⋅80 F 28⋅80 Nephrops norvegicus M 62⋅00 Chapman and Rice 1971 F 41⋅00 Panulirus argus – 40⋅30 Davis 1981 Paralithodes camtschatica J 29⋅40 Edwards 1972 – 14⋅80 Paralithodes camtschatica M 15⋅30 Niwa and Kurata 1964 F 19⋅50 J, Juveniles; M, males; F, females; –, not categorised. Carapace width: *20–34⋅9, **65–79⋅9 mm.
5
P. MARIAPPAN ET AL.

shape of the chela (Sexton and Reid 1951; Hartnoll 1963). Season-induced cyclic changes in chela polymorphism has been reported in males of Orconectes propinquus (Stein 1976).
3. Autotomy
Autotomy refers to a reflex severance of one or more limbs in response to injury or its threat, which occurs al-ways in a predetermined breakage plane (Wood and Wood 1932; Robinson et al 1970; McVean 1982). A number of factors contribute to the prevalence of auto-tomy, which has been extensively studied and reviewed from time to time (Wood and Wood 1932; Bliss 1960; McVean 1982; Juanes and Smith 1995). Crustaceans widely practice self amputation of one or more limbs dur-ing inter- and intraspecific competition for limited re-sources like food, shelter, mate and also as a strategy to avoid predation and wound limitation (Wood and Wood 1932; Bliss 1960; McVean 1982). Apart from such biological reasons, commercial factors like intentional harvesting of chelipeds in species like Menippe merce-naria (Savage and Sullivan 1978), incidental damage by fishing gear (Kirkwood and Brown 1998), and culling of undesirable individuals (Kennelly et al 1990) are also responsible for the loss of chelipeds. In the polymorphic male population of M. rosenbergii, cheliped loss is a periodic event among the dominant blue-clawed males (bulls) on attaining a critical value of 1 : 2⋅8 ± 0⋅18 body length/chela length as a growth strategy (Schmalbach et al 1984). Males of M. nobilii (28%) (carapace length: 1⋅6–2⋅5 cm) resort to chela autotomy during exuviation even when reared individually under ideal laboratory conditions (Mariappan and Balasundaram 1999a); even multiple limb autotomy occurs in M. malcolmsonii in the field (P Mariappan and C Balasundaram, unpublished data). The limb loss varies from species to species (1⋅7% in Carcinus maenas, Sekkelsten 1988; 80% in Cyrtograpsus angulatus, Spivak and Politis 1989), within a species (C. maenas, 1⋅7%, Sekkelsten 1988; 55%, McVean 1976) and as a function of size within a species (Necora puber, 12% in juveniles and 38% in adults, Norman and Jones 1991) (table 3). To a certain extent temporal and geo-graphic variations also contribute to autotomy in a given population (Shirley and Shirley 1988; Smith 1990a). Though the autotomised animals get immediate advantage in terms of survival, in the long term the need to divert body resources for regeneration has an adverse effect on the regular energy budget. Further the injured animal becomes less dominant and remains more vulnerable to further attacks in a community; autotomy also limits its access to shelter, food gathering potential, and its abi- lity to find a mate (Kuris and Mager 1975; Sekkelsten
1988; Davenport et al 1992; Abello et al 1994; Smith 1995).
4. Regeneration
Crustaceans have the ability to replace lost limbs by means of regeneration, which is linked with moulting (Prizbram 1901; Bliss 1960; Skinner 1985). However, at any given time, in a wild population of Cancer magis-ter the proportion of animals with regenerating limbs (5%) is comparatively lower than that of animals with lost limbs (18%) (Shirley and Shirley 1988), suggesting an increased vulnerability of autotomised animals to preda-tion (McVean and Findlay 1979). In some species the process of limb regeneration affects the moult increment and moult interval but in others no such effect has been reported (Smith 1990b; Spivak 1990; Cheng and Chang 1993). Regeneration of a lost limb to its original size depends upon age and time of loss in a given moult cycle. Normally the lost limb regenerates within 2–3 moults, faster in juveniles than in adults (Skinner 1985; Smith 1990b).
5. Abnormalities in chelipeds
Abnormalities or malformation of chelipeds have been reported widely in various decapod crustaceans like lob-ster (Homarus americanus, Faxon 1881; H. gammarus and Nephrops norvegicus, cf. Shelton et al 1981), crayfish (Procambarus clarkii, Chokki and Ishihara 1994; Naka-tani et al 1997), crab (Geryon affinis granulatus, Oka-moto 1991; Macrophthalmus japonicus, Suzuki 1963), and the Japanese edible crab (Chionectes japonicus, Mo-toh 1971). Most of these claw abnormalities are mainly due to a lateral outgrowth in the propodus, which results es-pecially from abnormal wound healing following the damage of the propodus (Okamoto 1991; Nakatani et al 1992); this phenomenon could also be induced in the laboratory (Murayama et al 1994; Nakatani and Kitahara 1999).
6. Cheliped display
Communication in crustaceans often involves the display of antennae and chelipeds. The roles of the chelipeds in agonistic and aggressive interactions during inter- and intraspecific competition for a limited resource is well documented in the literature (Hazlett 1972; Salmon and Hyatt 1983). The possession of chelipeds plays a major role in acquisition and retention of shelters in Homarus americanus (O’Neill and Cobb 1979) and Macrobrachium nobilii (Balasundaram and Mariappan 1998). Different
6
P. MARIAPPAN ET AL.

kinds of acts or movements for cheliped presentation have been reported in various crustaceans (Liocarcinus depura-tor and Necora puber, Huntingford et al 1995; H. ameri-canus, Atema and Cobb 1980; Macrobrachium rosenbergii, Barki et al 1991; M. australiense, Lee and Fielder 1983). A detailed account of the use of chelipeds in communication is provided by Salmon and Hyatt (1983). Cheliped extension, meral spread, strike, lifting of claw, scissoring, thrust, cheliped striking, embrace, nip and push are the major events mediated by chelipeds (table 4 and figure 3).
7. Courtship and mating
The role of chelipeds in courtship display and the pres-ence of chelipeds as an aid in mate access have been extensively studied. In Uca pugilator there is a marked diffe-rence in display patterns between mature and immature males (Salmon et al 1978). Since chelae have a major role in displays during agonistic interactions, the degree of dominance is expressed by the type of chela morphometry. An animal with robust chelae has easy
access to mates during inter-male competition and also through sexual selection by females. Autotomy also plays a crucial role in mating since such handicaps lead to a negative effect on mate access as observed in small and medium sized males of Carcinus maenas (Sekkelsten 1988). Variations in mating and reproductive patterns are observed among polymorphic males of Macrobrachium rosenbergii. The dominant blue-clawed males (bulls, BC males) effectively court and protect mates (Ra’anan and Sagi 1985), while intermediate males (OC males) show reduced reproductive activity in the presence of BC males (Ra’anan and Cohen 1985). Submissive small males are also sexually less competent, but mate successfully in the absence of BC and OC males (Sagi 1984).
8. Implications of chelae for decapod culture
In communal culture of crustaceans, the possession of large crusher chelae triggers aggression between individu-als leading to physical damage of body parts (especially chelipeds) aggravating the rate of limb loss and mortality. Indeed chelipeds constitute 10–26% of the body weight in
Table 4. Cheliped mediated displays in decapod crustaceans. Act Description Cheliped extension2,3,5,6 Extension of chelae towards the opponent without contact
Cheliped presentation2 Ambulatory legs in walking position and both chelipeds in presentation position
Cheliped shaking3,5 Rapid oscillations of the dactyls while (without touching propodus) the claw is partly extended in the direction of another prawn
Complete lifting3,5 Lifting of the claws and anterior part of the body towards another individual
Incomplete lifting3 Similar to complete lifting, but the claws remain in the hori-zontal plane
Crouching6 Chelipeds are folded tightly against the body
Fending6 Outward swinging of one or both chelipeds
Grasping1,6 Seizing of another individual with thoracic appendages 3–5
Meral spread1,3 Outward spreading of the enlarged meri of the appendages
Nip3,4 One animal closes down the tips of its chelae on a body part of another animal
Push3,4,5 One animal pushes one of its chelae against a body part of another animal
Shielding6 Holding the chelipeds like a shield
Strike1,3 A blow delivered by one individual to another with the dac-tyls of one or both raptorial appendages
Scissoring3,5,6 Bringing the two claws together from the complete lifting position in a scissoring motion
Thrust3 Rapid simultaneous opening of the two claws in the direc-tion of another prawn
Sources: 1) Dingle 1969, 2) Hazlett and Bossert 1965, 3) Barki et al 1991, 4) Peebles 1979, 5) Lee and Fielder 1983, 6) Jachowski 1974.
7
P. MARIAPPAN ET AL.
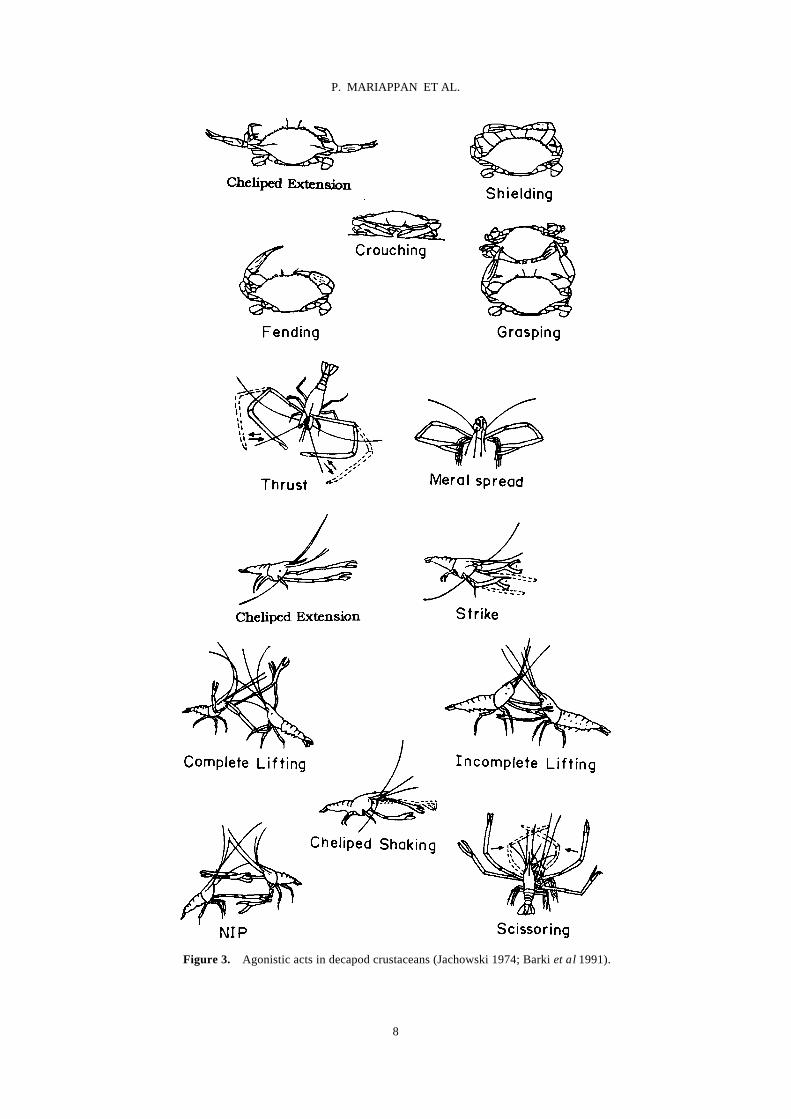
Figure 3. Agonistic acts in decapod crustaceans (Jachowski 1974; Barki et al 1991).
8
P. MARIAPPAN ET AL.

Macrobrachium nobilii (Mariappan and Balasundaram 1999a), 20% in Carcinus maenas and Liocarcinus hol-satus (Lee and Seed 1992), and 50% in Menippe merce-naria (Simonson and Steele 1981). In H. americanus, the possession of the crusher claw is essential for acquisition of limited resources, as well as establishment and mainte-nance of dominance hierarchies (O’Neill and Cobb 1979). In such cases the autotomised animal becomes subjugated and more subordinated during further attacks. In C. sapidus, the loss of chelipeds was shown to have not only a negative effect on foraging ability and prey handling time (Juanes and Hartwick 1990; Smith and Hines 1991), but also the incumbent has to channelise more metabolic energy for the regeneration of chelipeds. Thus in species like Callinectes sapidus (Ary et al 1987; Smith 1990b), the loss of chelipeds leads to a reduction in moult incre-ment due to energy diversion; such energy demand is called regeneration load (Skinner 1985), which may reduce reproductive output (Norman and Jones 1993; Luppi et al 1997). Chelotomy, dactylotomy and immobi-lisation of the dactyls have been shown to reduce the degree of cannibalism in H. americanus (Kendall et al 1982) and in M. rosenbergii (Karplus et al 1989; Diaz et al 1990). However the decreased survival rate due to forced severance of limbs and subsequent regeneration are major constraints that reduce the harvest size (Powell et al 1998).
9. Conclusion
Though autotomy, moulting and regeneration of chelipeds have been reviewed periodically, a collective perusal of literature attempted in this review reveals that the diverse functional and structural modifications of chelipeds are not only influenced by feeding and locomotion patterns, but also by environmental conditions and species-specific needs. A number of biotic and abiotic factors influence the development of chelae. The chelae are most vulnera-ble to autotomy and their regeneration imposes a regene-ration load in the regular energy budget of the animal resulting in a telling effect on the other regular somatic and reproductive processes. In aquaculture experimental removal of chelae minimizes aggressive interactions but the problem is recurrent due to regeneration potential and hence is of limited applicability. Since it takes more than one moult for total regeneration of the chelae, their use as a taxonomic character is doubtful.
Acknowledgements
Financial assistance from the Council of Scientific and Industrial Research, New Delhi, to PM in the form of a Senior Research Fellowship and University Grants Com-
mission, New Delhi, to CB in the form of a major research project is gratefully acknowledged. Thanks are also due to Dr S Prem Mathi Maran, Chennai, for line drawings.
References
Abby-Kalio N J and Warner G F 1989 Heterochely and handed-ness in the shore crab Carcinus maenas (L.) (Crustacea: Brachyura); Zool. J. Linn. Soc. 96 19–26
Abello P, Warman C G, Reid D G and Naylor E 1994 Chela loss in the shore crab, Carcninus maenas (Crustacea: Brach-yura) and its effect on mating success; Mar. Biol. 121 247–252
Ahmed M 1978 Development of asymmetry in the fiddler crab Uca cumulanta Crane, 1943 (Decapoda, Brachyura); Crusta-ceana 34 294–300
Aiken D E and Waddy S L 1992 The growth process in cray-fish; Rev. Aquat. Sci . 6 335–381
Ary R D, Bartell C K and Poirrier M A 1987 The effects of chelotomy on molting in the blue crab, Callinectes sapidus; J. Shellfish Res . 6 103–108
Atema J and Cobb J S 1980 Social behavior; in The biology and management of lobsters (eds) J S Cobb and B F Phillips (New York: Academic Press) vol 1, pp 409–450
Averof M and Patel N H 1997 Crustacean appendage evolution associated with changes in Hox genes expression; Nature (London) 388 682–686
Balasundaram C and Mariappan P 1998 Observations on the sheltering behaviour of Macrobrachium nobilii (Henderson and Matthai 1910); in Natl. Symp. Sustainable Aquaculture , Feb. 20–21, 1998, University of Delhi, New Delhi. Abstract No. 2
Barki A, Karplus I and Goren M 1991 Morphotype related dominance hierarchies in males of Macrobrachium rosenber-gii (Crustacea, Palaemonidae); Behaviour 117 145–160
Barnes R S K 1968 Relative carapace and chela proportions in some Ocypodid crabs (Brachyura, Ocypodidae); Crustaceana 14 131–136
Barnwell F H 1982 The prevalence of male right-handedness in the Indo-West Pacificfiddler crabs Uca vocans (Linnaeus) and U. tetragonon (Herbst) (Decapoda: Ocypodidae); J. Crust. Biol. 2 70–83
Bennett D B 1973 The effect of limb loss and regeneration on the growth of the edible crab, Cancer pagurus L.; J. Exp. Mar. Biol. Ecol. 13 45–53
Bliss D E 1960 Autotomy and regeneration; in The physiology of crustacea (ed.) T H Waterman (New York: Academic Press) vol 1, pp 561–589
Bock W J and von Wahlert G 1965 Adaptation and the form-function complex; Evolution 19 269–299
Briggs P T and Mushacke F M 1979 The American lobster and the pot fishery in the inshore waters of the south shore of Long Island, New York; N.Y. Fish Game J. 27 156–178
Brown S C, Cassuto S R and Loos R W 1979 Biomechanics of chelipeds in some decapod crustaceans; J. Zool. 188 143– 159
Bush S F 1930 Asymmetry and relative growth of parts in the two sexes of the hermit crab, Eupagurus prideauxi; Wilhelm Roux’ Arch. Entwicklungsmech. Org . 123 39–79
Chapman C J and Rice A L 1971 Some direct observations on the ecology and behaviour of the Norway lobster Nephrops norvegicus; Mar. Biol. 10 321–329
Cheng J-H and Chang E S 1993 Determinants of postmolt size
9
P. MARIAPPAN ET AL.

in the American lobster (Homarus americanus). 1. D sub(1) super(3) is the critical stage; Can. J. Fish. Aquat. Sci. 50 2106–2111
Cheung T S 1976 A biostatistical study of the functional consis-tency in the reversed claws of the adult male stone crabs, Menippe mercenaria (Say); Crustaceana 31 137–144
Chokki H and Ishihara T 1994 The second specimen of Pro-cambarus (Scapulicambarus) clarkii (Girard) bearing mal-formed chela; Bull. Owakidani Nat. Hist. Mus. Hakone 12 1–3 (in Japanese)
Clayton D A 1990 Crustacean allometric growth: a case for caution; Crustaceana 58 270–290
Crane J 1975 The fiddler crabs of the world (Ocypodidae: Genus Uca) (New Jersey: Princeton University Press)
Crothers J H 1967 The biology of the shore crab Carcinus mae-nas (L.); Field Stud. 2 407– 434
Dahl E 1956 Some crustacean relationships; in Bertil hanstrom: Zoological papers in honor of his sixty-fifth birthday (ed.) K G Wingstrand (Sweden: Lund Zool Inst) pp 138–147
Darby H H 1934 The mechanism of asymmetry in the Alphei-dae; Carnegie Inst. Washington Publ. 28 349–361
Davenport J, Spikes M, Thornton S M and Kelly B O 1992 Crab-eating in the diamond black terrapin Malaclemys terra-pin: dealing with dangerous prey; J. Mar. Biol. Assoc. U.K.; 72 835–848
Davis T A 1987 Laterality in Crustacea; Proc. Indian Natl. Sci. Acad . B53 47–60
Dawes B 1934 A study of normal and regenerative growth in pistol shrimp, Alpheus dentipes (Guèrin); Wilhelm Roux’ Arch. Entwicklungsmech. Org. 131 543–574
Diaz G G, Nakagawa H and Kasahara S 1990 Effect of propo-dus excision on growth and survival in giant freshwater prawn Macrobrachium rosenbergii; J. Fac. Appl. Biol. Sci. (Hiroshima Univ.) 29 19–24
Dingle H 1969 Statistical and information analysis of aggressive communication in the mantis shrimp Gonodactylus bredini Manning; Anim. Behav . 17 561–575
Durkin J T, Buchanan K D and Blahm T H 1984 Dungeness crab leg loss in the Columbia river estuary; Mar. Fish. Rev. 46 22–24
Edwards J S 1972 Limb loss and regeneration in two crabs: the king crab, Paralithodes camtschatica and the tanner crab Chionoecetes bairdi; Acta. Zool. 53 105–112
Estrella B T and Armstrong M P 1994 Massachusetts coastal commercial lobster trap sampling program May–November 1993; Mass. Div. Mar. Fish. 30
Farmer A S 1974 The development of external sexual characters of Nephrops norvegicus (L.) (Decapoda: Nephropidae); J. Nat. Hist. 8 241–255
Faxon W 1881 On some crustacean deformities; Bull. Mus. Comp. Zool . 8 257–274
Fryer G 1960 The feeding mechanism of some atyid prawns of the genus Caridina; Trans R. Soc. Edinburgh 54 335– 381
Finney W C and Abele L G 1981 Allometric variation and sexual maturity in the obligate coral commensal Trapezia fer-ruginea Latreille (Decapoda, Xanthidae); Crustaceana 41 113–130
Garvey J E and Stein R A 1993 Evaluating how chela size influences the invasion potential of an introduced crayfish (Orconectes rusticus); Am. Midl. Nat . 129 172–181
Goldstein J S and Noetzli C H 1997 Substrate variability as critical developmental factor in the claw asymmetry of the North American lobster, Homarus americanus; Today’s Aquaculture 6 4–5 and 11
Govind C K 1989 Asymmetry in lobster claws; Am. Sci . 77 468–474
Govind C K, Mellon, DeF and Quigley M M 1987 Muscle and muscle fiber type transformation in clawed crustaceans; Am. Zool. 27 1079–1098
Govind C K and Lang F 1981 Physiological identification and asymmetry of lobster claw closer motorneurons; J. Exp. Biol. 94 329–339
Govind C K and Pearce J 1988a Independent development of bilaterally homologous closer muscles in lobster claws; Biol. Bull. 175 430–433
Govind C K and Pearce J 1988b Remodeling of nerves during claw reversal in adult snapping shrimps; J. Comp. Neurol. 268 121–130
Govind C K and Pearce J 1994 Muscle remodelling in adult snapping shrimps via fat-fiber degeneration and slow-fiber genesis and transformation; Cell Tissue Res . 276 445–454
Govind C K, Pearce J and Potter D J 1988 Neural attrition following limb loss and regeneration in juvenile lobsters; J. Neurobiol. 15 4209–4222
Grandjean F, Romain D, Avila-Zarza C, Bramard M, Souty-Grosset C and Mocquard J P 1997 Morphometry, sexual di-morphism and size at maturity of the white-clawed crayfish, Austropotamobius pallipes pallipes (Lereboullet) from a wild French population at Deux-Sevres (Decapoda, Astacidea); Crustaceana 70 31–44
Haley S R 1969 Relative growth and sexual maturity of the Texas ghost crab, Ocypode quadrata (Fabr.) (Brachyura, Ocypodidae); Crustaceana 17 285–297
Hamilton P V, Nishimoto R T and Halusky J G 1976 Cheliped laterality in Callinectes sapidus (Crustacea: Portunidae); Biol. Bull . 150 393–401
Hartnoll R G 1960 Entionella monensis sp. nov., an entoniscis parasite of the crab Eurynome aspera (Pennant); J. Mar. Biol. Assoc. U.K . 39 101–107
Hartnoll R G 1963 The biology of Manx spider crabs; Proc. Zool. Soc. London 141 423– 496
Hartnoll R G 1974 Variations in growth pattern between some secondary sexual characters in crabs (Decapoda, Brachyura); Crustaceana 27 131–136
Hartnoll R G 1982 Growth; in The biology of crustacea (ed.) L G Abele (New York: Academic Press) vol. 2, pp 111–196
Hazlett B A 1962 Aspects of the biology of snapping shrimp (Alpheus and Syn apheus); Crustaceana 4 82–83
Hazlett B A 1972 Responses to agonistic postures by the spider crab Microphrys bicornutus; Mar. Behav. Physiol. 1 85–92
Hazlett B A and Bossert W H 1965 A statistical analysis of the aggressive communication systems of some hermit crabs; Anim. Behav . 13 357–373
Hazlett B A and Winn H E 1962 Sound production and associ-ated behavior of Bermuda crustaceans (Panulirus, Gonodac-tylus, Alpheus and Synalpheus); Crustaceana 4 25– 38
Herberholz J and Schmitz B 1998 Role of mechanosensory stimuli in intraspecific agonistic encounters of the snapping shrimp (Alpheus heterochaelis); Biol. Bull . 195 156–167
Herrick F H 1911 Natural history of American lobster; Bull.U.S. Bur. Fish. 29 149–408
Hopkins P M 1993 Regeneration of walking legs in the fiddler crab Uca pugilator; Am. Zool. 33 348–356
Hughes R N 1989 Foraging behaviour of a tropical crab, Ozius verreauxii; Proc. R.. Soc. London B237 201–212
Huntingford F A, Taylor A C, Smith, I P and Thorpe K E 1995 Behavioural and physiological studies of aggression in swimming crabs; J. Exp. Mar. Biol. Ecol. 193 21–39
10
P. MARIAPPAN ET AL.

Jachowski R L 1974 Agonistic behavior of the blue crab, Callinectes sapidus Rathbun; Behaviour 50 232–251
Jones M B 1978 Aspects of the biology of the big-handed crab, Heterozius rotundifrons (Decapoda: Brachyura), from Kai- koura, New Zealand; N.Z. J. Zool . 5 783– 794
Juanes F and Hartwick E B 1990 Prey size selection in dunge-ness crabs: the effect of claw damage; Ecology 71 744– 758
Juanes F and Smith L D 1995 The ecological consequences of limb damage and loss in decapod crustaceans: a review and prospectus; J. Exp. Mar. Biol. Ecol. 193 197–223
Kaestner A 1970 Invertebrate zoology (translated by H W Levi and L R Levi), vol 3, (New York: Wiley Interscience)
Karplus I, Samsonov E, Hulata G and Milstein A 1989 Social control of growth in Macrobrachium rosenbergii. I. The effect of claw ablation on survival and growth of communally raised prawns; Aquaculture 80 325–335
Kendall R A, Van Olst J C and Carlberg J M 1982 Effects of chelae immobilization on growth and survivorship for indi-vidually and communally raised lobsters, Homarus ameri-canus; Aquaculture 29 359–372
Kennelly S J, Watkins D and Craig J R 1990 Mortality of dis-carded spanner crabs, Ranina ranina (Linnaeus) in a tangle-net fishery-laboratory and field experiments; J. Exp. Mar. Biol. Ecol. 140 39–48
Kirkwood J M and Brown I W 1998 Effect of limb damage on the survival and burial time of discarded spanner crabs Ranina ranina (Linnaeus); Mar. Freshwater Res. 49 41–45
Kuris A M and Mager M 1975 Effect of limb regeneration on size increase a molt of the shore crabs Hemigrapsus oregonen-sis and Pachygrapsus crassipes; J. Exp. Zool. 193 353– 360
Kuris A M, Ra’anan Z, Sagi A and Cohen D 1987 Morphotypic differentiation of male Malaysian giant prawns, Macro-brachium rosenbergii; J. Crust. Biol. 7 219–237
Lee C L and Fielder D R 1983 Agonistic behaviour and the development of dominance hierarchies in the freshwater prawn, Macrobrachium australiense Holthuis, 1950 (Crusta-cea: Palaemonidae); Behaviour 83 1–17
Lee S Y 1995 Cheliped size and structure: the evolution of multi-functional decapod organ; J. Exp. Mar. Biol. Ecol. 193 161–176
Lee S Y and Seed R 1992 Ecological implications of the cheli-ped size in crabs: some data from Carcinus maenas and Lio-carcinus holsatus; Mar. Ecol. Prog. Ser. 84 151–160
Levinton J S, Judge M L and Kurdziel J P 1995 Functional dif-ferences between the major and minor claws of fiddler crabs (Uca, family Ocypodidae, Order Decapoda, Subphylum Crusta-cea): A result of selection or developmental constraint?; J. Exp. Mar. Biol. Ecol. 193 147–160
Lochhead J H 1961 Locomotion; in The physiology of crustacea (ed.) T H Waterman (New York: Academic Press) vol 2, pp 313–364
Luppi A T, Bas C C, Spivak E D and Anger K 1997 Fecundity of two grapsid crab species in the Laguna Mar Chiquita, Argentina; Arch. Fish. Mar. Res. 45 149–166
MacGinitie G E and MacGinitie N 1949 Natural history of marine animals (New York: McGraw Hill)
Manton S M 1977 The arthropods: habits, functional morphology and evolution (London, New York: Oxford University Press)
Mariappan P and Balasundaram C 1999a Molt related limb loss in Macrobrachium nobilii; Curr. Sci. 75 637–639
Mariappan P and Balasundaram C 1999b Prevalence of auto-tomy in field populations of Macrobrachium nobilii; Indian J. Fish. 46 61–66
Mariappan P and Balasundaram C 1997 Cheliped laterality in the freshwater prawn Macrobrachium nobilii (Henderson and Matthai 1910); Curr. Sci. 73 875–877
McLaughlin P A 1982 Comparative morphology of crustacean appendages; in The biology of crustacea (ed.) D E Bliss (New York: Academic Press) vol 2, pp 197–256
McVean A 1976 The incidence of autotomy in Carcinus mae-nas (L.); J. Exp. Mar. Biol. Ecol. 24 177–187
McVean A 1982 Autotomy; in The biology of crustacea (ed.) D E Bliss (New York: Academic Press) vol 4, pp 107–132
McVean A and Findlay I 1979 The incidence of autotomy in an estuarine population of the crab Carcinus maenas (L.); J. Mar. Biol. Assoc. U.K . 59 341–354
Mellon DeF Jr 1981 Nerves and the transformation of claw type in snapping shrimps; Trends Neurosci. 4 245–248
Mellon DeF Jr and Stephens P J 1980 Modifications in the arrangement of thick and thin filaments in transformed shrimp muscle; J. Exp. Zool. 213 173–179
Moriyasu M, Landsburg W, Wade E and Maynard D R 1999 The role of an estuary environment for regeneration of claws in the American lobster, Homarus americanus H. Milne Edwards, 1837 (Decapoda); Crustaceana 72 417–433
Morris J A 1948 Studies on the host-parasite relationship of Probopyrus pandalicola (Packard); Cathol. Univ. Am. Biol. Stud. 8 1–20
Motoh H 1971 Abnormalities found in the left cheliped of Japa-nese edible crab, Chionectes japonicus Rathbun; Res. Crust. 4–5 184–190
Muino R, Fernandez L, Gonzalez-Gurraiaran E, Freire J and Vilar J A 1999 Size at maturity of Liocarcinus depurator (Brachyura: Portunidae): a reproductive and morphometric study; J. Mar. Biol. Assoc. U.K. 79 295–303
Murayama O, Nakatani I and Nishita M 1994 Induction of lat-eral outgrowths on the chelae of the crayfish, Procambarus clarkii (Girard); Crust. Res. 23 69–73
Mykles D L and Skinner D M 1981 Preferential loss of thin filaments during molt-induced atrophy in crab claw muscle; J. Ultrastruct. Res. 75 314–325
Nagamine C M and Knight A W 1980 Development, matura-tion, and function of some sexually dimorphic structures of the Malaysian prawn, Macrobrachium rosenbergii (De Man) (Decapoda, Palaemonidae); Crustaceana 39 141–152
Nakatani I and Kitahara 1999 Induction of outgrowths at wounds on the cheliped of Procambarus clarkii (Decapoda, Cambari-dae); J. Crust. Biol. 19 1–7
Nakatani I, Okada Y and Yamaguchi T 1997 An extra claw on the first and on the third cheliped of the crayfish, Procamba-rus clarkii (Decapoda, Cambaridae); Crustaceana 70 788–798
Nakatani I, Yamaguchi T and Murayama O 1992 Abnormalities found in the chela of the crayfish, Procambarus clarkii (Gi-rard); Res. Crust. 21 207–209
Ng P K L and Tan L W H 1985 ‘Right handedness’ in the heterochelous calappoid and xanthoid crabs – suggestion for functional advantage; Crustaceana 49 98–100
Niwa K and Kurata H 1964 Limb loss and regeneration in the adult king crab Paralithodes camtschatica; Bull. Hokkaido Reg. Fish. Res. Lab . 28 51–55 (Transl. from Japanese by Fish. Res. Board Can. Transl. Ser. No. 1190, 1969)
Nolan B A and Salmon M 1970 The behavior and ecology of snapping shrimp (Crustacea: Alpheus heterochaelis and Alpheus norma nni); Forma Functio 2 289– 335
Norman C P 1995 Limb loss in the poisonous crab Atergatis floridus (Linnaeus) advantages of possessing toxins?; Crust. Res. 24 137–145
11
P. MARIAPPAN ET AL.

Norman C P and Jones M B 1991 Limb loss and its effect on handedness and growth in the velvet swimming crab Necora puber (Brachyura: Portunidae); J. Natl. Hist. 25 639– 645
Norman C P and Jones M B 1993 Reproduction ecology of the velvet swimming crab, Necora puber (Brachyura: Portuni-dae), at Plymouth; J. Mar. Biol. Assoc. U.K. 73 379–389
Ogonowski M M and Lang F 1979 Histochemical evidence for enzyme differences in crustacean fast and slow muscle; J. Exp. Zool . 207 143–151
Ogonowski M M, Lang F and C K Govind 1980 Histochemistry of lobster claw-closer muscles during development; J. Exp. Zool. 213 359–367
O’Neill D J and Cobb J S 1979 Some factors influencing the outcome of shelter competition in lobsters (Homarus ameri-canus); Mar. Behav. Physiol. 6 33–45
Okamoto K 1991 Abnormality found in the cheliped of Geryon affinis granulatus Sakai; Res. Crust. 20 63–65
Paulian R 1936 L’existence d’un stade critique dans la crois-sance relative de l’ Eupagurus prideauxi (Crustacée ano-moure); C.R. Seances Soc. Biol. Ses Fil . 121 435–437
Peebles J B 1979 The role of prior residence and relative size in competition for shelter by the Malaysian prawn Macro- brachium rosenbergii; Fish. Bull. 76 905–911
Pillai G 1990 Notes on the chelae of the mangrove lobster Tha-lassina anomala (Decapoda, Thalassinidae); Crustaceana 59 89–95
Pinheiro M A A and Fransozo A 1993 Relative growth of the speckled swimming crab Arenaeus cribrarius (Lamarck, 1818) (Brachyura, Portunidae), near Ubatuba, State of Sao Paulo, Brazil; Crustaceana 65 377–389
Pinheiro M A A and Fransozo A 1998 Sexual maturity of the speckled swimming crab Arenaeus cribrarius (Lamarck, 1818) (Decapoda, Brachyura, Portunidae), in the Ubatuba lit-toral, Sao Paulo State, Brazil; Crustaceana 71 434–452
Powell M L, Hammer H S and Watts S A 1998 Observations on the frequency of claw loss in the crayfish Procambarus clarkii; J. World Maricult. Soc. 29 485–490
Przibram H 1901 Experimentelle studien uber regeneration; Arch. Ent. Mech. Org . 11 321–345
Quigley M M and Mellon DeF Jr 1984 Changes in myofibrillar gene expression during fibre-type transformation in the claw closer muscles of the snapping shrimp Alpheus heterochelis; Dev. Biol . 106 262–265
Ra’anan Z and Cohen D 1985 The ontogeny of social structure and population dynamics in the freshwater prawn, Macro-brachium rosenbergii (de Man); in Crustacean issues II. Crustacean growth (eds) F M Schram and A Wenner (Rotter-dam: Balkema) pp 271–311
Ra’anan Z and Sagi A 1985 Alternative mating strategies in male morphotypes of the freshwater prawn Macrobrachium rosenbergii (de Man); Biol. Bull . 169 592–601
Read A T and Govind C K 1997 Regeneration and sex-biased transformation of the sexually dimorphic pincer claw in adult snapping shrimps; J. Exp. Zool. 279 356–366
Reverberi G 1943 Sul significato della “castrazione parassi-taria”. La trasformazione del sesso nei Crostacei parassiti da Bopiridi e da Rizocefali; Pubbl. Stn. Zool. Napoli 19 225–316
Robinson M H, Abele L G and Robinson B 1970 Attack auto- tomy: A defense against predators; Science 169 300–301
Sagi A 1984 Alternative reproduction strategies in male popu-lation of the freshwater prawn Macrobrachium rosenbergii, M.Sc. Thesis, Hebrew University, Jerusalem
Salmon M and Hyatt G W 1983 Communication; in The biology
of crustacea (ed.) D E Bliss (New York: Academic Press) vol. 7, pp 1–40
Salmon M, Hyatt G, McCarthy K and Costlow J D Jr 1978 Dis-play specificity and reproductive isolation in the fiddler crabs Uca panacea and U. pugilator. Z. Tierpsychol. 48 251–276
Savage T and Sullivan J R 1978 Growth and claw regeneration of the stone crab, Menippe mercenaria; Florida Mar. Res. Publ. 32 1–23
Schmalbach E A, Harpaz S, Kahan D, Galun R and Frankenberg E 1984 Periodic cheliped autotomy of the males of the Malaysian prawn Macrobrachium rosenbergii; Naturwissen-schaften 71 325–326
Schmitt W L 1965 Crustaceans (Ann Arbor: University of Michigan Press)
Schmitz B 2000 Sound production in Crustacea with special reference to the Alpheidae; in Physiology of the Crustacean nervous system (ed.) K Wiese (Springer-Verlag) (in press)
Schram F R 1978 Arthropods: A convergent phenomenon; Fieldiana 39 61–108
Schultz S, Wuppermann K and Schmitz B 1998 Behavioural interactions of the snapping shrimp (Alpheus heterochaelis) with conspecifics and sympatric crabs (Eurypanopeus depres-sus); Zool. Anal. Complex Syst. (Suppl I ) 101 85
Schuster S M and Caldwell R L 1989 Male defense of the breeding cavity and factors affecting the persistence of breed-ing pairs in the stomatopod, Gonodactylus bredini (Manning) (Crustacea: Hoplocarida); Ethology 82 192–207
Seed R and Hughes R N 1995 Criteria for prey size-selection in molluscivorous crabs with contrasting claw morphologies; J. Exp. Mar. Biol. Ecol. 193 177–195
Sekkelsten G I 1988 Effect of handicap on mating success in male shore crabs Carcinus maenas; Oikos 51 131–134
Sexton E W and Reid D M 1951 The life history of the multi-form species Jassa falcata (Montagu) (Crustacea, Amphi-poda) with a review of the bibliography of the species; J. Linn. Soc. London Zool . 57 29–88
Shelton P M J, Truby P R and Shelton R G J 1981 Naturally occurring abnormalities (Bruchdreifachbildungen) in the che-lae of three species of Crustacea (Decapoda) and a possible explanation; J. Embryol. Exp. Morphol . 63 285–304
Shih H-T, Mok H-K, Chang H-W and Lee S-C 1999 Morpho-logy of Uca formosensis, 1921 (Crustacea: Decapoda: Ocy-podidae), an endemic fiddler crab from Taiwan, with notes on its ecology; Zool. Stud . 38 164–177
Shirley S M and Shirley T C 1988 Appendage injury in dunge-ness crabs, Cancer magister, in Southeastern Alaska; Fish. Bull. 86 156–160
Simonson J L and Steele P 1981 Cheliped asymmetry in the stone crab, Menippe mercenaria, with notes on claw reversal and regeneration; Northeast Gulf Sci. 5 21–30
Skinner D M 1966 Breakdown and reformation of somatic muscle during the molt cycle of land crab, Gecarcinus later-alis; J. Exp. Zool. 163 115–124
Skinner D M 1985 Molting and regeneration; in The biology of crustacea (eds) D E Bliss and T H Mantel (New York: Aca-demic Press) vol 9, pp 43–143
Smith L D 1990a The frequency and ecological consequences of limb autotomy in the blue crab, Callinectes sapidus Rath-bun, Ph D thesis, University of Maryland, Maryland, USA
Smith, L D 1990b Patterns of limb loss in the blue crab, Callinectes sapidus Rathbun, and the effects of autotomy on growth; Bull. Mar. Sci. 46 23–36
Smith L D 1995 Effects of limb autotomy and tethering on juve-nile blue crab survival from cannibalism; Mar. Ecol. Prog. Ser. 116 65–74
12
P. MARIAPPAN ET AL.

Smith L D and Hines A H 1991 The effect of cheliped loss on blue crab Callinectes sapidus Rathbun foraging rate on soft-shell clams Mya arenaria L.; J. Exp. Mar. Biol. Ecol. 151 245–256
Smith L D and Palmer A R 1994 Effects of manipulated diet on size and performance of brachyuran crab claws; Science 264 710–712
Snedden W A 1990 Determinants of male mating success in the temperate crayfish Orconectes rusticus: chela size and sperm competition; Behaviour 115 100–113
Spivak E D 1990 Limb regeneration a common South American littoral crab Cyrtograpsus angulatus; J. Natl. Hist. 24 393–402
Spivak E D and Politis M A 1989 High incidence of limb auto-tomy in crab population from a coastal lagoon in the province of Buenos Aires, Argentina; Can. J. Zool. 67 1976– 1985
Stebbing T R R 1893 A history of Crustacea recent mala- costraca (London: Kegan Paul, Trench, Treubner and Co Ltd.)
Stein R A 1976 Sexual dimorphism in crayfish chelae: func-tional significance linked to reproductive activities; Can. J. Zool. 54 220–227
Stephens P J and Mellon DeF Jr 1979 Modification of structure and synaptic physiology in transformed shrimp muscle; J. Comp. Physiol. 132 97–108
Stevens B G, Donaldson W E, Haaga J A and Munk J E 1993 Morphometry and maturity of paired Tanner crabs, Chionoe-cetes bairdi, from shallow and deepwater environments; Can. J. Fish. Aquat. Sci. 50 1504–1516
Suzuki H 1963 An abnormality found in the cheliped of Mac-ropthalmus japonicus De Haan; Res. Crust. 1 51–53
Swartz R C 1972 Postlarval growth and reproduction in the
painted ghost crab Neopanope texana sayi, Ph D thesis, Col-lege of William and Mary,
Tiegs O W and Manton S M 1958 The evolution of the Arthro-poda; Biol. Rev . 33 255–337
Trott T J 1987 The prevalence of left-handedness in the painted ghost crab Ocypode gaudichaudii H. Milne Edwards and Lucas (Decapoda, Brachyura, Ocypodidae); Crustaceana 52 213–215
Tucker B W 1930 On the effects of an epicaridan parasite, Gyge branchialis, on Upogebia littoralis; Q. J. Microsc. Sci. (N.S. ) 74 1–118
Tweedie M W F 1950 The fauna of the Cocos-Keeling Islands, Brachyura and Stomatopoda; Bull. Raffles Mus . 22 105–148
Vermeij G J 1977 Patterns in claw size: the geography of crush-ing; Syst. Zool. 26 138–151
Warner G F and Jones A R 1976 Leverage and muscle type in crab chelae (Crustacea: Brachyura); J. Zool. 180 57–68
Wilson E B 1903 Notes on the reversal of asymmetry in the regeneration of chelae in Alpheus heterochelis; Biol. Bull. 4 197–210
Wood F D and Wood W H 1932 Autotomy in decapod Crusta-cea; J. Exp.. Zool. 62 1–55
Yamaguchi T 1973 Asymmetry and dimorphism of chelipeds in the fiddler crab, Uca lactea De Haan; Zool. Mag . 82 154–158
Yamaguchi T 1977 Studies on the handedness of the fiddler crab, Uca lactea; Biol. Bull. 152 424–436
Yamaguchi T and Aratake H 1997 Morphological modifications caused by Sacculina polygenea in Hemigrapsus sanguineus (De Haan) (Brachyura: Grapsidae); Crust. Res. 26 125–145
Young R E, Pearce J and Govind C K 1994 Establishment and maintenance of claw bilateral asymmetry in snapping shrimps; J. Exp. Zool. 269 319–326
13
P. MARIAPPAN ET AL.

Keywords. Harderian gland; nasolacrimal duct; squamate reptiles; vomeronasal organ
The structure of the nasal chemosensory system in squamate reptiles. 2. Lubricatory capacity of the vomeronasal organ
SUSAN J REHOREK , BRUCE T FIRTH and MARK N HUTCHINSON
.
The vomeronasal organ is a poorly understood accessory olfactory organ, present in many tetrapods. In mammals, amphibians and lepidosaurian reptiles, it is an encapsulated structure with a central, fluid-filled lumen. The morphology of the lubricatory system of the vomeronasal organ (the source of this fluid) varies among classes, being either intrinsic (mammalian and caecilian amphibian vomeronasal glands) or extrinsic (anuran and urodele nasal glands). In the few squamate reptiles thus far examined, there are no submucosal vomeronasal glands. In this study, we examined the vomeronasal organs of several species of Australian squamates using histological, histochemical and ultrastructural techniques, with the goal of determining the morphology of the lu-bricatory system in the vomeronasal organ. Histochemically, the fluid within the vomeronasal organ of all squamates is mucose-rous, though it is uncertain whether mucous and serous constituents constitute separate components. The vomeronasal organ produces few secretory granules intrinsically, implying an extrinsic source for the luminal fluid. Of three possible candidates, the Harderian gland is the most likely extrinsic source of this secretion.
1. Introduction
The vomeronasal organ is a nasal chemosensory structure found in most terrestrial vertebrates. It is embryologically derived from the olfactory placode, and is both morphologically and physiologi-cally similar to the main olfactory organ (see Halpern 1992 for review). Both systems consist of a chemosensory epithelium whose luminal aspects are bathed in a fluid, wherein odorant chemicals must dissolve prior to neural excitation (Getchell et al 1984a, b; Takami et al 1995). Variable dependence on either of these chemosensory systems has been documented within squa-mate reptiles (Halpern 1992; Schwenk 1993a, b; Cooper 1996). Snakes are acknowledged vomeronasal specialists, based on vari-ous morphological, neuroanatomical and behavioural features (see Halpern 1992 for review). Schwenk (1993a) and Dial and Schwenk (1996) proposed that gekkotan lizards may, in contrast, be olfac-tory specialists. However, evidence supporting this hypothesis is based on limited morphological, neuroanatomical and behavioural observations of some gekkotan species, as well as the
absence of snake-like vomeronasal behaviour (i.e., complex tongue-flicking). The morphology of the scincid lizard VNO has received some attention (Kratzing 1975; Halpern 1992). Though the vo-meronasal sensory capacity in scincid lizards is unknown, none of the features indicating snake-like vomeronasal specialization are present (i.e., complex tongue-flicking behaviour: Schwenk 1993b). The structure of the gekkotan vomernasal organ is similar to that of the scincid lizards (Gabe and Saint Giron 1976; Schwenk 1993b). Thus, though varying levels of nasal chemosensory de-pendence has been ascribed to snakes, skinks and gekkotans, there is little data on the morphology of the gekkotan vomeronasal or-gan. One aspect of the vomeronasal sense which has received little attention, is the lubricatory system. It is well ac-cepted that the lubricatory system in the main olfactory organ consists of the submucosal Bowman’s glands and sometimes the sustentacular cells (Andres 1969; Müller et al 1979; Getchell and Getchell 1992).The lubricatory system of the vomeronasal organ has not only received little attention but also appears to be vari
Journal of Biophysical Chemistry, 2009, 1, 14-23

able within tetrapods. In mammals, for example, the vomeronasal lubricatory system consists of submucosal, seromucous vomero-nasal glands (see Adams 1992 for review), and the development of the vomeronasal organ is positively correlated to the presence of these glands (Cooper and Bhatnager 1976). This is not the case in squamate reptiles, in which no such glands are known (Kratzing 1975; Gabe and Saint Girons 1976). However, the absence of these glands does not seem to hinder the development of the vo-meronasal organ in squamates. This suggests that there is suffi-cient secretion for the squamate vomeronasal organ from other sources to compensate the absence of the intrinsic vomeronasal glands. Whether sufficient glandular material might be scattered throughout the vomeronasal organ in squamates is unknown, but seems unlikely (Bannister 1968; Altner et al 1970; Kratzing 1975; Gabe and Saint Girons 1976; Wang and Halpern 1980; Takami and Hirosawa 1987, 1990; Halpern 1992). However, most studies have been carried out on snake and scincid lizard species. The morphology of the vomeronasal organ in gekkotans has thus far only been reported in the survey of Gabe and Saint Girons (1976). This survey, carried out at the light microscopic level, showed some features in vomeronasal lubricatory system of gekkotans (presence of potential secretory material in the non-sensory epi-thelium) which were not shared with either scincid lizards or snakes. This has not since been verified with either other speci-mens or with ultrastructural analysis. Further examination of the gekkotan condition is thus warranted, as this potential difference in the vomeronasal lubricatory system may translate into func-tional differences in the vomeronasal system within squamate reptiles (akin to that potentially existing between snakes and mammals). There are several gekkotan taxa, each of which potentially vary in dependence on the vomeronasal sense. Of the three gekkotan taxa found in Australia, two (Diplodactylinae and Pygopodidae) are restricted to the Australasian region (Greer 1989). The legless pygopods possess many snake-like behavioural (i.e., oscillatory tongue-flicking) and morphological (i.e., relatively slender, slightly bifurcate tongue) characters. Both of these characters might indi-cate snake-like vomeronasal speciality (Schwenk 1993b). Py-gopods are most closely related to the fully limbed diplodactyline geckos (Kluge 1987). Gekkoninae, is a closely related sister taxon to the Diplodactylinae/ Pygopodidae taxa, also occurs in Australia (Kluge 1987). These were then compared to the vomeronasal or-gan of a scincid lizard (Morethia adelaidensis) and a snake (Pseu-donaja textilis). We thus aimed to determine not only whether the pygopod vomeronasal organ differed from that of geckos, but also to determine how vomeronasal organ morphology of geckos and pygopods compares to that of the scincid lizard and snake. Special attention was given to the lubricatory system.
2. Materials and methods
Adults from the following species were collected from the out-skirts of Adelaide, South Australia, during spring (September–November); (Gekkota) Gekkonidae (Geckos) Gekkoninae: Chris-tinus marmoratus (20), Diplodactylinae: Strophurus intermedius (5), Pygopodidae (flap-footed lizards): Delma molleri (20), (Scin-comorpha) Scincidae (skinks): M. adelaidensis (6), Serpentes (snakes) Elapidae: P. textilis (18). At least one of each sex per species was examined with each of the morphological techniques. All animals were sacrificed with an intraperitoneal injection of sodium pentobarbitol (Nembutal), decapitated, and the heads placed in fixative (see below). Either entire heads, or half heads (cut sagittally) of at least 1 specimen per species were fixed in 10% phosphate-buffered for-malin for at least 1 week, decalcified in 10% aqueous EDTA, em-bedded in paraffin, and sectioned serially (7 µm). Alternate slides were stained with haematoxylin-eosin, in order to maximize mate-rial for the species in which only a few specimens were obtained. Alternate slides of either full or half heads (not stained with haematoxylin-eosin) were tested histochemically for the presence of acidic mucosubstances and proteins. Neutral and acidic muco-substances were detected by the periodic acid-Schiff (PAS), and alcian yellow (at pH 2· 5) (Ravetto 1964) methods respectively. The mercury bromo- phenol blue (BPB) test was used to detect protein (Barka and Anderson 1965), with pronase digestion for control. For transmission electron microscopy, vomeronasal organs (at least 1 specimen per species) which had been dissected from the other side of the nasal capsule, were fixed for 4 h at room tempera-ture in 3% formaldehyde/3% gluteraldehyde in 0· 1 M phosphate buffer at pH 7· 4, and postfixed for 1 h in 1% osmium tetroxide, then dehydrated through a series of ethyl alcohols and embedded in epoxy resin. Grids with thin sections (0· 1 µm) were stained with 2% uranyl acetate and lead citrate and examined with a PHILIPS CM 100 transmission electron microscope.
3. Results
3.1 Histology
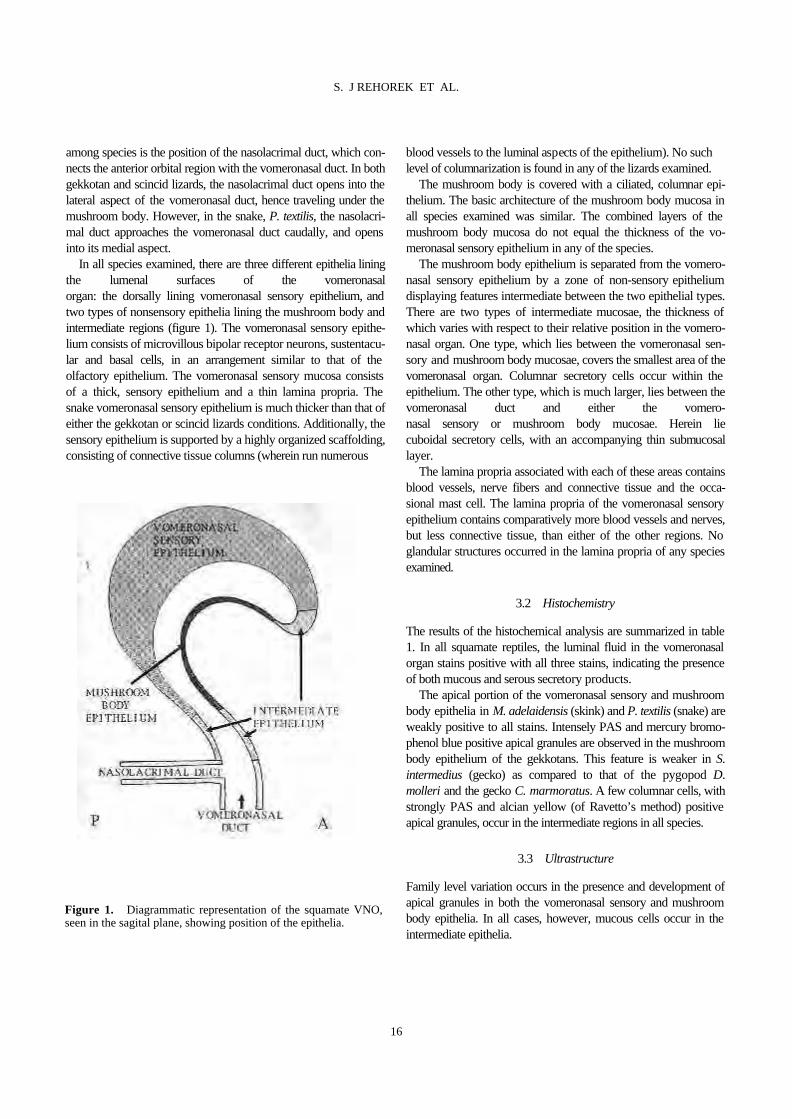
The squamate vomeronasal organ is a dome-shaped, bone- and cartilage-encased structure in the rostral floor of the nasal cavity. The mushroom body, a conch-like projection from the ventro-lateral aspect of the vomeronasal organ, projects into the lumen (figure 1). The vomeronasal duct connects the vomeronasal organ lumen with the mouth cavity. The vomeronasal organ appears to be in the same position and possesses roughly the same relative size in all species examined. Grossly, the only apparent difference
15
S. J REHOREK ET AL.
among species is the position of the nasolacrimal duct, which con-nects the anterior orbital region with the vomeronasal duct. In both gekkotan and scincid lizards, the nasolacrimal duct opens into the lateral aspect of the vomeronasal duct, hence traveling under the mushroom body. However, in the snake, P. textilis, the nasolacri-mal duct approaches the vomeronasal duct caudally, and opens into its medial aspect. In all species examined, there are three different epithelia lining the lumenal surfaces of the vomeronasal organ: the dorsally lining vomeronasal sensory epithelium, and two types of nonsensory epithelia lining the mushroom body and intermediate regions (figure 1). The vomeronasal sensory epithe-lium consists of microvillous bipolar receptor neurons, sustentacu-lar and basal cells, in an arrangement similar to that of the olfactory epithelium. The vomeronasal sensory mucosa consists of a thick, sensory epithelium and a thin lamina propria. The snake vomeronasal sensory epithelium is much thicker than that of either the gekkotan or scincid lizards conditions. Additionally, the sensory epithelium is supported by a highly organized scaffolding, consisting of connective tissue columns (wherein run numerous
blood vessels to the luminal aspects of the epithelium). No such level of columnarization is found in any of the lizards examined. The mushroom body is covered with a ciliated, columnar epi-thelium. The basic architecture of the mushroom body mucosa in all species examined was similar. The combined layers of the mushroom body mucosa do not equal the thickness of the vo-meronasal sensory epithelium in any of the species. The mushroom body epithelium is separated from the vomero-nasal sensory epithelium by a zone of non-sensory epithelium displaying features intermediate between the two epithelial types. There are two types of intermediate mucosae, the thickness of which varies with respect to their relative position in the vomero-nasal organ. One type, which lies between the vomeronasal sen-sory and mushroom body mucosae, covers the smallest area of the vomeronasal organ. Columnar secretory cells occur within the epithelium. The other type, which is much larger, lies between the vomeronasal duct and either the vomero- nasal sensory or mushroom body mucosae. Herein lie cuboidal secretory cells, with an accompanying thin submucosal layer. The lamina propria associated with each of these areas contains blood vessels, nerve fibers and connective tissue and the occa-sional mast cell. The lamina propria of the vomeronasal sensory epithelium contains comparatively more blood vessels and nerves, but less connective tissue, than either of the other regions. No glandular structures occurred in the lamina propria of any species examined.
3.2 Histochemistry
The results of the histochemical analysis are summarized in table 1. In all squamate reptiles, the luminal fluid in the vomeronasal organ stains positive with all three stains, indicating the presence of both mucous and serous secretory products. The apical portion of the vomeronasal sensory and mushroom body epithelia in M. adelaidensis (skink) and P. textilis (snake) are weakly positive to all stains. Intensely PAS and mercury bromo-phenol blue positive apical granules are observed in the mushroom body epithelium of the gekkotans. This feature is weaker in S. intermedius (gecko) as compared to that of the pygopod D. molleri and the gecko C. marmoratus. A few columnar cells, with strongly PAS and alcian yellow (of Ravetto’s method) positive apical granules, occur in the intermediate regions in all species.
3.3 Ultrastructure
Family level variation occurs in the presence and development of apical granules in both the vomeronasal sensory and mushroom body epithelia. In all cases, however, mucous cells occur in the intermediate epithelia.
Figure 1. Diagrammatic representation of the squamate VNO, seen in the sagital plane, showing position of the epithelia.
16
S. J REHOREK ET AL.
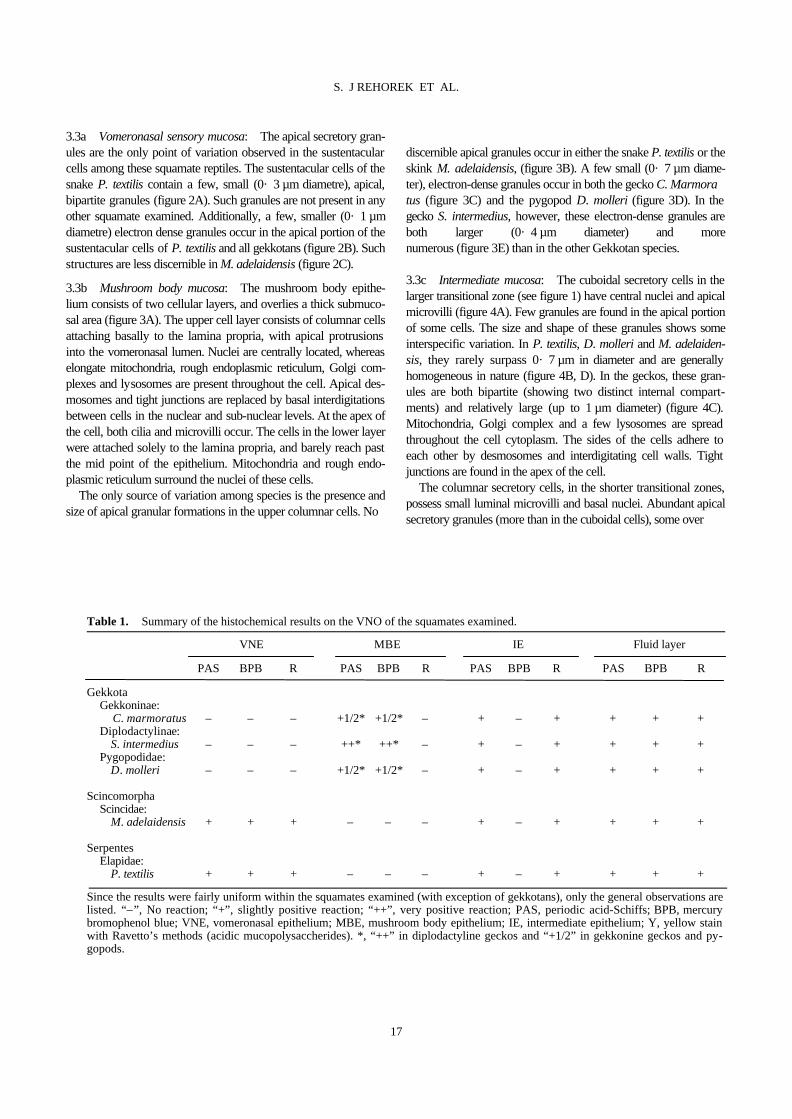
3.3a Vomeronasal sensory mucosa: The apical secretory gran-ules are the only point of variation observed in the sustentacular cells among these squamate reptiles. The sustentacular cells of the snake P. textilis contain a few, small (0· 3 µm diametre), apical, bipartite granules (figure 2A). Such granules are not present in any other squamate examined. Additionally, a few, smaller (0· 1 µm diametre) electron dense granules occur in the apical portion of the sustentacular cells of P. textilis and all gekkotans (figure 2B). Such structures are less discernible in M. adelaidensis (figure 2C). 3.3b Mushroom body mucosa: The mushroom body epithe-lium consists of two cellular layers, and overlies a thick submuco-sal area (figure 3A). The upper cell layer consists of columnar cells attaching basally to the lamina propria, with apical protrusions into the vomeronasal lumen. Nuclei are centrally located, whereas elongate mitochondria, rough endoplasmic reticulum, Golgi com-plexes and lysosomes are present throughout the cell. Apical des-mosomes and tight junctions are replaced by basal interdigitations between cells in the nuclear and sub-nuclear levels. At the apex of the cell, both cilia and microvilli occur. The cells in the lower layer were attached solely to the lamina propria, and barely reach past the mid point of the epithelium. Mitochondria and rough endo-plasmic reticulum surround the nuclei of these cells. The only source of variation among species is the presence and size of apical granular formations in the upper columnar cells. No
discernible apical granules occur in either the snake P. textilis or the skink M. adelaidensis, (figure 3B). A few small (0· 7 µm diame-ter), electron-dense granules occur in both the gecko C. Marmora tus (figure 3C) and the pygopod D. molleri (figure 3D). In the gecko S. intermedius, however, these electron-dense granules are both larger (0· 4 µm diameter) and more numerous (figure 3E) than in the other Gekkotan species. 3.3c Intermediate mucosa: The cuboidal secretory cells in the larger transitional zone (see figure 1) have central nuclei and apical microvilli (figure 4A). Few granules are found in the apical portion of some cells. The size and shape of these granules shows some interspecific variation. In P. textilis, D. molleri and M. adelaiden-sis, they rarely surpass 0· 7 µm in diameter and are generally homogeneous in nature (figure 4B, D). In the geckos, these gran-ules are both bipartite (showing two distinct internal compart-ments) and relatively large (up to 1 µm diameter) (figure 4C). Mitochondria, Golgi complex and a few lysosomes are spread throughout the cell cytoplasm. The sides of the cells adhere to each other by desmosomes and interdigitating cell walls. Tight junctions are found in the apex of the cell. The columnar secretory cells, in the shorter transitional zones, possess small luminal microvilli and basal nuclei. Abundant apical secretory granules (more than in the cuboidal cells), some over
Table 1. Summary of the histochemical results on the VNO of the squamates examined.
VNE MBE IE Fluid layer
PAS BPB R PAS BPB R PAS BPB R PAS BPB R
Gekkota Gekkoninae: C. marmoratus – – – +1/2* +1/2* – + – + + + + Diplodactylinae: S. intermedius – – – ++* ++* – + – + + + + Pygopodidae: D. molleri – – – +1/2* +1/2* – + – + + + + Scincomorpha Scincidae: M. adelaidensis + + + – – – + – + + + + Serpentes Elapidae: P. textilis + + + – – – + – + + + +
Since the results were fairly uniform within the squamates examined (with exception of gekkotans), only the general observations arelisted. “–”, No reaction; “+”, slightly positive reaction; “++”, very positive reaction; PAS, periodic acid-Schiffs; BPB, mercury bromophenol blue; VNE, vomeronasal epithelium; MBE, mushroom body epithelium; IE, intermediate epithelium; Y, yellow stain with Ravetto’s methods (acidic mucopolysaccherides). *, “++” in diplodactyline geckos and “+1/2” in gekkonine geckos and py-gopods.
17
S. J REHOREK ET AL.

2 µm in diameter, are their most prominent feature (figure 4E). The granules are homogenous, but vary in electron density among species.
4. Discussion
At the anatomical level, the vomeronasal organ of all the squa-mates studied exhibits some morphological variation. This includes
the route of the nasolacrimal duct and the columnarization of the vomeronasal sensory epithelium (both of which differentiates the snakes from the lizards) and the structure of the lubricatory sys-tem. The last of these, the structure of the lubricatory system, shows much variation. Thus, it is much harder to make generaliza-tion. Each facet of the vomeronasal lubricatory system will thus be discussed individually. Histochemically, both acidic mucopolysaccharides (based on reactivity to PAS and alcian yellow stains: Drury and Wallington 1980) and proteins (based on reactivity to mercury bromophenol blue: Barka and Anderson 1965) occur in the squamate luminal fluid. It is uncertain whether the two types of secretion form separate layers, are mixed together, or a combination of the two. A heterogeneous fluid layer, with two chemically distinct lamina, covers the sensory epithelia of the main olfactory organ of terres-trial vertebrates (Andres 1969; Müller et al 1979; Getchell and Getchell 1992). There is some evidence for such layering of fluid in the mammalian vomeronasal organ (Takami et al 1995). Though this layering appears to be important in the function of the main olfactory organ and the vomeronasal organ, the precise function of the fluid components is speculative. The most likely function for the fluid is as a medium for chemicals to dissolve before they can stimulate the neural components of the vomerona-sal sensory epithelium (Getchell and Getchell 1992; Getchell et al 1993). Additionally, the fluid may provide sustenance for the epithelia. The fluid may also contain stimulus binding proteins which transport the stimulus to the vomeronasal sensory epithe-lium, or enzymes which break down the complex chemicals to smaller units which would then bind to the vomeronasal receptor neurons. Further microchemical analyses of this fluid are required before either the laminous nature or the function of the vomeronasal fluid components can be ascertained. The fluid filling the lumen of the vomeronasal organ is produced by a vomeronasal lubricatory system. An intrinsic lubricatory system (i.e., secretory structures found either within or in close proximity to the chemosensory mucosae) is less well developed in squamate reptiles than in other tetrapods. Since a lubricatory sys-tem is needed to produce the luminal fluid, the possibility that there is an external source needs to be explored. In the remainder of this paper, evidence for intrinsic versus extrinsic sources of vomeronasal fluid is examined.
4.1 Intrinsic sources of luminal fluid
Intrinsic secretory structure for the vomeronasal organ may come in the form of distinct glandular masses (vomeronasal glands) or many scattered secretory cells within or in close proximity to the chemosensory mucosa. The composition of the intrinsic sources for the vomeronasal luminal fluid varies within tetrapods. The mammalian and caecilian amphibian vo-meronasal organs contain well developed vomeronasal glands (Cooper and Bhatnager 1976; Badenhorst 1978; Adams 1992). In
Figure 2. The apices of the vomeronasal sensory epithelia of P. textilis (A), S. intermedius (B) and M. adelaidensis (C). Note the presence of large granules in P. textilis (arrow heads), with smaller ones present in both P. textilis and S. intermedius (arrow heads). (Bar: 1 µm.)
18
S. J REHOREK ET AL.

anuran and urodele amphibians, however, the nasal glands in the adjoining nasal capsule are thought to be the source of the fluid (Dawley and Bass 1988; Døving et al 1993). In squamates, however, the vomeronasal organ has comparatively few intrinsic secretory structures, with no evidence of any glandular material development. Additionally, since the squamate vomeronasal duct lacks a connection to the nasal cavity (and thus cannot directly
receive fluid from the nasal gland), unlike the condition in other tetrapods, the nasal glands in squa- mates are an unlikely source of fluid for the vomeronasal organ. The paucity of intrinsic secretory structures is a striking feature of the squamate vomeronasal lubricatory system (Kratzing 1975; Gabe and Saint Girons 1976; Halpern 1992). Secretory granules are few and limited to mucous secretory cells in the intermediate region in all squamates examined and also to sustentacular (mu cous) and mushroom epithelial (serous) cells in the snake and gek
Figure 3. The mushroom body epithelia of P. textilis (A), M. adelaidensis (B), C. marmoratus (C), D. molleri (D) and S. inter- medius (E). Arrows point to apical e-dense granules. e, Epithelium, lp, lamina propria. (Bar: A = 5 µm; B–E = 2 µm).
19
S. J REHOREK ET AL.

kotans, respectively. Apical mucous secretory granules occur in the sustentacular cells of several other squamate species (Bannister 1968; Altner et al 1970; Gabe and Saint Girons 1976; Wang and Halpern 1980; Takami and Hiro- sawa 1990). Thus, the presence of mucous granules in the sustentacular cells is not unique to snakes and is unlikely to be associated with snake vomeronasal specialization. In addition to the geckos, iguanids and some other squamate
reptiles also possess a few apical secretory granules in the mush-room body epithelium (Gabe and Saint Girons 1976). Whether there is enough serous secretion produced to contribute signifi-cantly to the serous component of the luminal fluid is unknown. It is thus apparent that there is little (gekkotan) or no (snake and skinks) intrinsic source of serous secretion in the squamate VNO. Therefore, the source of the mucous and serous fluid in the squamate VNO lumen is unlikely to be intrinsic.
Figure 4. The intermediate epithelia of P. textilis (A and D), D. molleri (B) and S. interme-dius (C and E). A–D show the cuboidal epithelium, with higher magnifications in B–D indicat-ing structural diversity of the secretory granules. E shows the columnar epithelium (Bar: A and E = 5 µm; B–D = 1 µm).
20
S. J REHOREK ET AL.

4.2 Extrinsic sources of luminal fluid
The absence of any associated internal or nearby glands, or diffuse secretory structures, suggests that either the squamate vomerona-sal organ does not require a well developed lubricatory system or that there are alternative sources of fluid. The presence of mucous and serous components in the overlying fluid indicates that the fluid is necessary and that the squamate vomeronasal organ functions in a manner similar to that of other tetrapods. If this is the case, then the absence of the in-trinsic glandular structures, which is essential in the mammalian vomeronasal sense (Cooper and Bhatnager 1976; Takami et al 1995), implies that there is an extrinsic source for the fluid in the squamate VNO (Kratzing 1975). There are two criteria which a potential external source of fluid for the squamate vomeronasal luminal fluid must meet. First, the fluids must have ready access to the vomeronasal organ. Second, since the histo-chemical results indicate the presence of protein in the luminal fluid, the external source must be capable of producing serous secretions. Three extrinsic sources for the squamate vomeronasal organ have been suggested, including the salivary glands, secretions of the nasolacrimal duct, and the Harderian gland (Kratzing 1975; Halpern 1992; Rehorek 1997b). 4.2a Salivary glands: Although, in squamates, saliva is copi-ously produced by the salivary glands, and secreted into the mouth cavity, and it is possible for the fluids to flow into the vomeronasal organ via the vomeronasal duct, there is no experi-mental evidence linking the saliva in the mouth to the fluid layer of the VNO. Furthermore, the salivary glands of squamates produce both serous and mucous fluids (Saint Girons 1988). Since there is currently no evidence to either support or refute the role of the salivary glands in the vomeronasal sense, further studies are war-ranted. 4.2b Nasolacrimal duct: The nasolacrimal duct opens directly into the duct of the vomeronasal organ, or in the vicinity thereof, in all squamates with vomeronasal organs (Bellairs and Boyd 1950; Rehorek 1997a). Even when the vomeronasal organ is ab-sent, the nasolacrimal duct still opens in the same relative region (Bellairs and Boyd 1950; Slaby 1984). Thus, Kratzing (1975) proposed that the nasolacrimal duct may be a source of lubricant for the squamate vomeronasal organ. However, the nasolacrimal duct appears to possess few secretory granules (Saint Girons 1982; Rehorek 1997a). Thus, the nasolacrimal duct itself fails to meet one of the criteria. It is therefore unlikely that the nasolacrimal duct itself is a source of secretion for the vomeronasal organ. 4.2c Harderian gland: The Harderian gland is an enigmatic, ubiquitous, serous secreting structure, whose ducts open to the anterior portion of the orbit in squamates (Saint Girons 1982; Chieffi et al 1992; Rehorek 1997a, b). These ducts are closely associated with the proximal part of the nasolacrimal duct (Bellairs and Boyd 1947; Saint Girons 1982; Rehorek 1997b). Despite
some minor variations, the nasolacrimal duct and Harderian gland are associated directly or indirectly with the VNO in all squamate reptiles thus far examined (Bellairs and Boyd 1947; Saint Girons 1982; Rehorek 1992, 1997a, b). Thus, the Harderian gland meets both criteria (ready access to vomeronasal organ and serous secre-tory) of an external source for the vomeronasal luminal fluids. Of the three candidates, the Harderian gland is the most likely source of serous secretion for the fluid in the squamate vomerona-sal organ. Tracing studies have confirmed both the route and the presence of Harderian gland secretions (via the nasolacrimal duct) in the lumen of the squamate vomeronasal organ (Rehorek et al 1999). What the functional role of these secretions is, or even whether they are the sole contributors to the vomeronasal luminal fluid, is unknown, and further research is warranted.
5. Conclusions
The squamate vomeronasal organ has fewer intrinsic secretory structures than that of either amphibians or mammals. The fluid in the vomeronasal organ, particularly its serous compo-nent, is unlikely to derive solely from an intrinsic source, and would thus have to be derived, at least in part, from extrinsic sources.Much remains to be determined with respect to the lubri-catory system in the squamate vomeronasal organ. This includes not only the source of the fluid in the vomeronasal organ, but also the role of the secretory granules in the vomeronasal epithelia. This study supports previous morphological and tracing stud-ies suggesting that the Harderian gland plays a role in the vomero-nasal sense of squamate reptiles. The significance of this observation can be appreciated on several levels. If the Harderian gland functions in the squamate vomeronasal sense, then a 300 year old mystery may finally be solved. Therefore, examination of the squamate Harderian gland could lead to insights into the func-tion of the relatively understudied vomeronasal organ (physiologi-cal examinations of this structure are currently limited by its inaccessibility). Further studies of the VNO and Harderian gland interaction need to be carried out at both the comparative and molecular levels. Examination of this system in a variety of squa-mate and non-squamate tetrapods would establish the evolution-ary history of this unusual system (i.e., why or how an orbital gland came to be associated with a nasal chemosensory organ). At the molecular level, the precise role of the fluid in the squamate vomeronasal organ, and the relative contribution of the Harderian gland, would lead to a better understanding of the vomeronasal sense.
Acknowledgments
We thank Mr C M Leigh, R Murphy and G Hermanis, and the staff of Medical Illustrations and Photography, SUNY Health Science Center, Brooklyn, for technical assistance. We also thank Drs W J Hillenius, D G Homberger and K Schwenk for construc
21
S. J REHOREK ET AL.

tive criticism of this manuscript. This work was funded by an Adelaide University postgraduate scholarship and the New York College of Osteopathic Medicine, of the New York Institute of Technology. Animals were collected in accordance with the regula-tions stipulated by the South Australian National Parks and Wild-life Act (permit numbers U2338-01, 02, and 03; C21465-03; Q010000-03, 04, and 05; Y12016-01, 02 and 03). Animals were kept in captivity for a brief time in accordance with the guidelines set by the University of Ade-laide Animal Ethics Committee application M/58/93.
References
Adams D R 1992 Fine structure of the vomeronasal and septal olfactory epithelia and of glandular structures; Microsc. Res. Tech. 23 86–97
Altner H, Müller W and Brachner I 1970 The ultrastructure of the vomeronasal organ in reptilia; Z. Zellforsch. 105 107–122
Andres K H 1969 Der olfaktorisches Saum der Katze; Z. Zellforsch. Mikrosk. Anat. 96 250–274
Badenhorst A 1978 The development and the phylogeny of the organ of Jacobson and the tentacular apparatus of Ichthyophis glutinosus (Linné); Ann. Univ. Stellen. Serie 2A 1 1–26
Bannister L H 1968 Fine structure of the sensory endings in the vomeronasal organ of the Slow worm Anguis fragilis; Nature (London) 217 275–276
Barka T and Anderson P J 1965 Histochemistry: Theory, practice and bibliography (New York: Harper and Row)
Bellairs A d’A and Boyd J D 1947 The lachrymal apparatus in lizards and snakes-1. The brille, the orbital glands, lachrymal canaliculi and origin of the lachrymal duct; Proc. Zool. Soc. London 117 81–108
Bellairs A d’A and Boyd J D 1950 The lachrymal apparatus in lizards and snakes-2. The anterior part of the lachrymal duct and its relationship with the palate and with the nasal and vomeronasal organs; Proc. Zool. Soc. London 120 269–310
Chieffi G, Chieffi-Baccari G, Di Matteo L, d’Istria M, Marmorino C, Minucci S and Varriale B 1992 The Harderian gland of amphibians and reptiles; in Harderian glands: Porphyrin metabolism, behavioural and endocrine effects (eds) S M Webb, R A Hoffman, M L Puig-Domingo and R Reiter (Berlin: Springer-Verlag) pp 91–108
Cooper J G and Bhatnager K P 1976 Comparative anatomy of the vomeronasal complex in bats; J. Anat. 122 571–601
Cooper W E Jr 1996 Preliminary reconstructions of nasal chemosensory evolution in squamata; Amphibia-Reptilia. 17 395–415
Dawley E M and Bass A H 1988 Organization of the vomeronasal organ in a Plethodontid salamander; J. Morphol. 198 243–255
Dial B E and Schwenk K 1996 Olfaction and predation detection in Coleonyx brevis (Squamata: Eublepharidae), with comments on the functional significance of buccal pulsing in Geckos; J. Exp. Zool. 276 415–424
Døving K B, Trotier D, Rosin J-F and Holley A 1993 Functional architecture of the vomeronasal organ of the frog (genus Rana); Acta Zool. 74 173–180
Drury R A B and Wallington E A 1980 Carleton’s histological technique (Oxford: Oxford University Press)
Gabe M and Saint Girons H 1976 Contribution a la morpholige
compareé des fosses nasales et de leur annexes chez les Lépidosauriens; Mém. Mus. Nat. Hist. Nat. A98 1–87 Getchell M L and Getchell T V 1992 Fine structural aspects of
secretion and extrinsic innervation in the olfactory mucosa; Microsc. Res. Tech. 23 111–127
Getchell M L, Rafols J A and Getchell T V 1984a Histological and histochemical components of the salamander olfactory mucosa: effects of isoproterenol and olfactory nerve section; Anat. Rec. 208 553–565
Getchell T V, Margolis F L and Getchell M L 1984b Perireceptor and receptor events in vertebrate olfaction; Prog. Neurobiol. 23 317–345
Getchell T V, Su Z and Getchell M L 1993 Mucous domains: microchemical heterogeneity in the mucociliary complex of the olfactory epithelium; The molecular basis of smell and taste transduction (Ciba Foundation Symposium 179) (Chichester: Wiley) pp 27–50
Greer A E 1989 The biology and evolution of Australian lizards (Norton: Surrey Beatty)
Halpern M 1992 Nasal chemical senses in reptiles: Structure and function; in Biology of the reptilia. Vol. 18. Physiology E (eds) C Gans and D Crews (Chicago: University of Chicago) pp 424–532
Kluge A G 1987 Cladistic relationships in the Gekkonoidea (Squamata; Sauria); Misc. Publ. Univ. Mich. 173 1–54
Kratzing J E 1975 The fine structure of the olfactory and vomeronasal organs of a lizard (Tiliqua scincoides scincoides); Cell Tissue Res. 156 239–252
Müller H, Drenkhahn D and Haase E 1979 Vergleichend quantitative und ultrastrukturelle Untersuchungen am Geruchs-organ von Hausaubenrassen; Z. Mikrosk. Anat. Forsch. 93 888–900
Ravetto C 1964 Alcian blue-Alcian yellow: a new method for the identification of different acid groups; J. Histochem. Cytochem. 12 44–45
Rehorek S J 1992 A comparative morphological analysis of Australian scincid Harderian glands, Honours thesis, Depart-ment of Anatomy and Histology, University of Adelaide, Adelaide, Australia
Rehorek S J 1997a A potential role of the squamate Harderian gland in vomerolfaction, Ph.D thesis, Department of Ana-tomical Sciences, University of Adelaide, Adelaide, Australia
Rehorek S J 1997b Squamate Harderian gland: An overview; Anat. Rec. 248 301–306
Rehorek S J, Hillenius W J, Quan W and Halpern M 1999 XXI meeting for the Association for Chemoreceptive Sciences, Sarasota, 14-18 April 1999
Saint Girons H 1982 Histologie compareé des glandes orbitaires des Lépidosauriens; Ann. Sci. Nat. Zool. Paris 4 171– 191
Saint Girons H 1988 Les glandes céphaliques exocrines des Reptiles. I – Données anatomiques et histologiques; Ann. Sci. Nat. Zool. Paris 9 221–255
Schwenk K 1993a Are geckoes olfactory specialists?; J. Zool. London 229 289–302
Schwenk K 1993b The evolution of chemoreception in squamate reptiles: a phylogenetic approach; Brain Behav. Evol. 41 124–137
Slaby O 1984 Morphogenesis of the nasal apparatus in a member of the genus Chameleon L. Morphogenesis of the nasal capsule,
22
S. J REHOREK ET AL.

the nasal epithelial tube and the organ of Jacobson in Sauropsida VIII; Folia Morphol. (Prague) 32 225–246
Takami S, Getchell M L and Getchell T V 1995 Resolution of sensory and mucoid glycoconjugates with terminal a-galactose residues in the mucomicrovillar complex of the vomeronasal sensory epithelium by dual confocal laser scanning microscopy; Cell Tissue Res. 280 211–216
Takami S and Hirosawa K 1987 Light microscopic observations of
the vomeronasal organ on Habu, Trimeresurus flavoviridis; Jpn. J. Exp. Med. 57 163–174
Takami S and Hirosawa K 1990 Electron microscopic ob- servations on the vomeronasal sensory epithelium of a crotaline snake, Trimerusurus flavoviridis; J. Morphol. 205 45–61
Wang D and Halpern M 1980 Light and electron microscopic observations on the normal structure of the vomeronasal organ of garter snakes; J. Morphol. 164 47–67
23
S. J REHOREK ET AL.

Keywords. Calcium-dependent protein kinase; degenerate primer; Funaria hygrometrica; nutrient; polymerase chain reaction; starvation ________________
Abbreviations used: CDPK, calcium dependent protein kinase; HMGR, 3-hydroxy-3-methylglutaryl-coenzyme A reductase; LCM, low calcium medium; LNM, low nitrate medium; LPM, low phosphate medium; LSM, low sulphate medium; MMG, minimal medium with glucose; NR, nitrate reductase; ORF, open reading frame; PCR, polymerase chain reaction; SnRK, sucrose non fer-menting (snf) related kinase; SPS, sucrose phosphate synthase.
Enhanced expression of a calcium-dependent protein kinase from the moss Funaria hygrometrica under nutritional starvation
DOYEL MITRA and MAN MOHAN JOHRI
Among the downstream targets of calcium in plants, calcium-dependent protein kinases (CDPKs) form an inter-esting class of kinases which are activated by calcium binding. They have been implicated in a diverse array of responses to hormonal and environmental stimuli. In order to dissect the role of CDPKs in the moss Funaria hygrometrica, a polymerase chain reaction (PCR)-based approach was adopted to clone the gene. Using degener-ate PCR primers against conserved regions of CDPKs, a 900 bp amplicon was obtained from the genomic DNA of Funaria. Southern hybridization under low stringency conditions indicated the presence of several CDPK related sequences in the Funaria genome. This observation is consistent with reports of multigene families of CDPKs in other plants. The 900 bp fragment was subsequently used to isolate a 2⋅2 kb partial genomic clone of the CDPK gene from Funaria. The genomic clone encodes an open reading frame (ORF) of 518 amino acids. Interestingly, unlike other CDPK genes from plants, the entire 1⋅5 kb ORF is not interrupted by introns. The deduced amino acid sequence of the Funaria gene shows extensive homology with CDPKs from higher plants, 73% identity with the Fragaria CDPK and 71% identity with CDPK isoform 7 of Arabidopsis. Phylogenetic analysis revealed that the Funaria CDPK is closer to the CDPKs from higher plants like strawberry and Arabidopsis as compared to those from lower plants such as the liverwort Marchantia, the green alga Chlamydomonas or another moss Tortula. Northern analysis shows enhanced expression of the CDPK transcript within 24–48 h of starvation for nitrogen, phosphorus or sulphur. So far the only other kinase which is known to be induced by nutrient starvation in plants is the wpk 4 which is a snf-1 related kinase (SnRKs). To our knowledge this is the first report that implicates a CDPK in the starvation response.
1. Introduction
Calcium is an important second messenger in plants that mediates signaling pathways elicited by hormonal as well as environmental stimuli (reviewed by Bush 1995). Cal-cium also plays a critical role in various developmental processes like pollen tube growth (Picton and Steer 1983)
and cytokinin induced bud formation in Funaria (Saun-ders and Hepler 1983). Among the downstream targets of calcium, the calcium dependent protein kinases (CDPKs) have emerged as the most predominant gene family in plants (Harmon et al 2000). CDPKs are a group of serine threonine kinases that have an N-terminal catalytic and a C-terminal calcium binding domain linked by an auto-
Journal of Biophysical Chemistry, 2009, 1, 24-31

inhibitory region. The kinase domain is preceded by a variable region, which differs both in sequence and size among different CDPK isoforms. The calcium binding region contains EF hand motifs and is also called the calmodulin-like domain. In the absence of calcium, the autoinhibitory region inhibits the kinase activity (Harmon et al 1994). The kinase is activated by the binding of cal-cium to the EF hands which brings about a conformational change that relieves the autoinhibition. Interestingly, CDPKs have so far been reported only in plants and protists but are absent in the completely sequenced genomes of Cenorhabditis elegans, Drosophlia melanogaster and Saccharomyces cerevisiae. The CDPK genes are highly conserved across species and are encoded by multigene families. There are about 40 CDPK isoforms in Arabidopsis, nine in maize and three in rice and soy-bean. The recombinant CDPK isoforms α, β and γ of soybean have been tested for their calcium binding affi-nity in the absence of substrates and found to have Kd values of 51, 1⋅4, and 1⋅6 µM, respectively (Lee et al 1998). Using isoform specific antibodies against the rice CDPKs, OsCDPK2 and OsCDPK11, it has been dem-onstrated that the two isoforms have different patterns of expression during seed development and in response to light (Frattini et al 1999). Such differences in substrate specificity, calcium binding affinity and tissue or sub-cellular localization of the various isoforms have been implicated to contribute to the multiplicity of function. CDPKs have been implicated in several aspects of plant growth and development (Harmon et al 2000). Studies involving the transient transfection of maize protoplasts with the Arabidopsis CDPK isoforms, CDPK1 and CDPK1a, have led to the establishment of their role in the stress response pathway (Sheen 1996). Recent work has implicated CDPKs in the Avr9 elicitor induced defense pathway (Romeis et al 2000). On the basis of induction of transcript levels, CDPKs have been impli-cated to play a role in mediating signalling elicited by the phytohormones such as, auxin (Botella et al 1996), ABA, cytokinin and gibberellin (Yoon et al 1999). Abiotic stresses like mechanical strain (Botella et al 1996), salt stress and dehydration (Urao et al 1994) have also been shown to induce transcription of CDPK genes in mung bean and Arabidopsis, respectively. Earlier work from our laboratory has shown the presence of multiple cal- cium dependent protein kinases in the protonema of Funaria (D’Souza and Johri 1999). In order to understand the role of CDPKs in the moss protonema we have iso-lated and characterized a partial genomic clone of a CDPK. The up-regulation of the transcript levels of this CDPK by nutrient starvation has been demonstrated by Northern analysis. To our knowledge, this is the first report that implicates a CDPK in the starvation response.
2. Materials and methods
2.1 Plant material and growth conditions
The experiments were carried out using the protonema of the moss Funaria hygrometrica Hedw. (Cell line J-2). The cultures were maintained in low-calcium medium (LCM) as described by Handa and Johri (1977). For the starvation experiments, cells were grown in media defi-cient in either of the nutrients such as nitrate, phosphate or sulphate. Before inoculating the LCM-grown cells into either of these media, they were washed with same medium in which they were inoculated. The low nitrate medium (LNM), low phosphate medium (LPM) and low sulphate medium (LSM) contain 0⋅5 mM nitrate, 24 µM phosphate and 8 µM sulphate, respectively. Cells grown in minimal medium with glucose (MMG) containing 8⋅4 mM nitrate, 1⋅8 mM phosphate and 1 mM sulphate served as control. The cells were inoculated at an initial density of 0⋅5 mg/ml in each of these media. During the first 48 h the cells grown in any of the media: LNM, LPM, LSM or MMG, grew at the same rate. This ensured that within 48 h, cul-tures grown in different media for the same duration of time, had identical cell densities. For the nutrient depriva-tion experiments, the cultures were harvested at identical cell densities of the treated and the control (MMG). After collection, the cells were frozen, ground in liquid nitrogen and stored at – 80°C till further use.
2.2 Extraction of DNA and polymerase chain reaction
Chloronema cells of Funaria were harvested at a density of 2 mg/ml and genomic DNA was prepared according to Dellaporta et al (1984) and subjected to CTAB purifica-tion. The polymerase chain reaction (PCR) conditions were as follows: 1⋅5 mM MgCl2, 0⋅2 mM dNTP, 100 pmole of each primer, 25 ng of Funaria genomic DNA, 2⋅5 U of Taq polymerase (Promega, Madison, USA)/100 µl reac-tion. The cycling conditions consisted of denaturation for 2 min at 94°C, followed by 45 cycles of denaturation for 45 s at 94°C, annealing at 54°C for 1 min and extension at 72°C for 1⋅5 min and then 12 min of extension at 72°C. The reactions were carried out using a thermocycler (Stratagene, USA). The sequences of the degenerate primers D1 and D2 were as follows: (D1) 5′ GGIGTIATGCA(T/C)(C/A)GIGA(T/C) (T/C)TIAA(A/G)CCIGA(A/G)AA 3′ (D2) 5′ GTIAT (A/G)AAICCIGAICC(A/G)TC(T/C) TT(A/G)TC 3′ These were obtained from Bangalore Genei. The PCR products were cloned in the pGEM-T vector from Promega, USA.
25
D. MITRA ET AL.

2.3 Southern hybridization
The genomic DNA of Funaria was digested with the enzymes XhoI, HindIII and EcoRI (Promega, USA), frac-tionated on a 0⋅7% agarose gel and transferred to a nylon membrane (Amersham, UK) (Sambrook et al 1989). The 900 bp PCR product was radiolabelled with α-P32 dATP (BRIT, India) by the random primer labelling kit (Amer-sham) and used as a probe. Southern hybridization was carried out at 65°C in a solution containing 6 × SSC, 5 × Denhardt’s solution, 0⋅5% SDS and 0⋅1 mg/ml of dena-tured salmon sperm DNA. The blot was washed in 0⋅1 × SSC and 0⋅1% SDS at 65°C for 15 min for the high strin-gency condition and in 0⋅5 × SSC and 0⋅1% SDS for 20 min at 55°C for the low stringency condition.
2.4 Northern hybridization
Total RNA was prepared from protonema grown under different nutrient levels using the RNeasy Plant minikit (Qiagen). The RNA was fractionated on a formaldehyde/ agarose (1⋅2% agarose) gel and blotted onto a nylon membrane (Sambrook et al 1989). Hybridization was carried out at 42°C in a solution containing 6 × SSC, 5 × Denhardt’s solution, 0⋅5% SDS, 50% formamide and 0⋅1 mg/ml of denatured salmon sperm DNA. The blot was washed in 2 × SSC, 0⋅1% SDS at 65°C for 20 min fol-lowed by a wash in 0⋅1 × SSC, 0⋅1% SDS at 65°C for 20 min. The Northern blots were scanned and the relative mRNA levels were estimated on the basis of signal inten-sities that were normalized with the values of the intensi-ties of the 28S rRNA of the ethidium bromide stained gels. The analysis was done using the image analysis software Adobe photoshop, version 5⋅5.
2.5 Genomic library construction and screening
Genomic DNA of Funaria was partially digested with Sau3AI and then size fractionated by sucrose gradient centrifugation in order to select fragments in the size range of 15 to 23 kb. Subsequently the DNA was partially filled-in with dGTP and dATP using Klenow polymerase (Amersham) and ligated to the λGEM-11 vector (Pro-mega). Plaques were screened by hybridization with the 900 bp PCR product as the probe (Sambrook et al 1989). Subclones of the inserts present in the positively hybridiz-ing plaques were made in the pGEM7Zf(+) vector (Promega, USA).
2.6 DNA sequencing
The genomic DNA clones were sequenced by an auto-mated DNA sequencer (model 377, Perkin Elmer) accord-
ing to the manufacturer’s instructions by Bangalore Genei.
3. Results
3.1 PCR amplification of a fragment of a CDPK gene
The degenerate primers D1 and D2 were designed against the conserved subdomains VIb and the third EF hand, corresponding to the sequences GVMHRDLKPEN and DKDGSGYIT, respectively. Using the above primers, three amplicons of approximate sizes 900, 800 and 600 bp, respectively, were obtained from the genomic DNA of Funaria of (figure 1). The present communication reports the analysis of only the 900 bp amplicon, referred to as P1. The annealing temperature of the polymerase chain reaction was varied from 44°C to 54°C and the P1 frag-ment could be amplified under all the conditions. Ampli-fication of P1 under high stringency conditions of PCR, namely at high temperatures such as 54°C, suggests that P1 represents authentic amplification of a CDPK gene. The P1 insert was cloned in the pGEM-T vector and sequen-ced. The sequence of P1 confirmed that it was a CDPK.
3.2 Genomic Southern analysis of Funaria CDPK
Southern hybridization of genomic DNA digested with XhoI, HindIII and EcoRI with the P1 probe was per-
Figure 1. PCR amplification of genomic DNA of Funaria with primers D1 and D2. Lane 1: negative control, PCR done without genomic DNA, lanes 2, 3, 4, 5, 6, 7: PCR done at annealing temperatures of 44, 46, 48, 50, 52 and 54°C, respec-tively. Lane 8: molecular weight markers. The 900 bp fragment is marked by an arrow.
26
D. MITRA ET AL.

formed under low and high stringency conditions of hybridization (figure 2). As expected, under high strin-gency conditions single bands were observed in all three digests, which confirmed the presence of the CDPK gene in the Funaria genome. Multiple bands were observed in the digests under low stringency conditions indicating the presence of several CDPK-related sequences in the genome. This observation is consistent with reports of multigene families of CDPKs in other plants.
3.3 Construction and screening of a genomic library of Funaria
Genomic DNA of Funaria was partially digested with Sau3AI and subjected to size fractionation by sucrose density gradient centrifugation. DNA Fragments in the size range of 15 to 23 kb were selected and ligated to the λGem-11 vector. Hence a genomic library of Funaria was constructed. In order to isolate the full-length gene, 250,000 plaques of this library were screened by hybridi-zation with the radiolabelled P1 probe. Three positive clones were identified and purified to homogeneity. These clones were subjected to restriction analysis and hybridi-zation to P1. The clone with the longest CDPK insert of 2⋅5 kb was subcloned using KpnI into the vector
pGEM7Zf(+) and sequenced. This led to the isolation of a 2⋅2 kb partial genomic clone of the Funaria CDPK.
3.4 Northern analysis of the Funaria CDPK
The expression pattern of the Funaria CDPK transcript was examined by Northern analysis. Chloronema cells were grown in MMG and as expected, an increase in the duration of growth was accompanied by a rise in cell den-sity of these cultures. Samples collected at different inter-vals of time were subjected to Northern analysis using the P1 probe. A 2⋅6 kb transcript was detected in all the lanes but interestingly the transcript levels showed a rise with an increase in the cell density (figure 5A). There was a 2⋅3-fold increase in the transcript level in the sample at the cell density of 22 mg/ml, compared to the level in the sample at the cell density of 2⋅6 mg/ml. Since increased cell density in these cultures leads to a reduction in the availability of nutrients, it is conceivable that the tran-script level was responding to the nutritional status of the medium. In order to check this possibility, cultures were grown in media, that were deficient in either of the three important macro-nutrients nitrogen, phosphorus and sul-phur, these media have been referred to as LNM, LPM and LSM, respectively. Consistent with our hypothesis, the transcript of the Funaria CDPK gene was found to be up-regulated when the cells were grown in LNM, LPM or LSM as compared to the MMG grown control cells (fig-ure 5B, C). In LNM, the transcript was up-regulated after 24 h of treatment whereas in LSM and LPM the effect was observed after 48 h. After 48 h of growth in the LNM, the FhCDPK transcript was induced by 1⋅6-fold as compared to the level in MMG-grown cells. Growth for 48 h in the LSM and LPM caused a 1⋅9-fold and 2⋅2-fold rise in the transcript levels, respectively, in comparison with tran-script levels in cells grown in MMG.
4. Discussion
Since CDPKs are highly conserved across species, a PCR based approach was adopted to clone the gene. The use of degenerate PCR primers designed against sequences in the kinase domain and in the calcium binding region ensured the amplification of only CDPKs. A 900 bp frag-ment of a CDPK gene was amplified from the genomic DNA of Funaria. This fragment (P1) was subsequently used to obtain the 2⋅2 kb partial clone from the genomic library of Funaria. This genomic clone encodes an ORF of 518 amino acids. The entire ORF was found not to be interrupted by introns, unlike the CDPK genes of isoforms 6, 9 and 19 in Arabidopsis or the maize CDPK gene which contain seve-ral introns. This gene has a 3′ untranslated region of 624 bp
Figure 2. Southern blot analysis. 15 µg of genomic DNA of Funaria was digested with XhoI, HindIII and EcoRI, electro-phoresed on a 0⋅7% agarose gel and transferred onto a nylon membrane. The membrane was probed with radiolabelled P1. The positions of the DNA molecular weight markers are indi-cated in kb. The blot was washed in 0⋅1 × SSC and 0⋅1% SDS at 65°C for 15 min for the high stringency condition (A) and in 0⋅5 × SSC and 0⋅1% SDS for 20 min at 55°C for the low strin-gency condition (B). The arrows indicate positions of single bands in the above three digests in A.
27
D. MITRA ET AL.

followed by a putative polyadenylation sequence, AATAAT. The ORF has all the features characteristic of CDPKs. It encodes 39 amino acids of the N-terminal variable region followed by the 11 kinase subdomains in the catalytic region and the four EF hands in the regula-tory region. The autoinhibitory region consists of 30 amino acids in contrast to the usual number of 31 in other
CDPKs. The significance of this observation is not understood. The deduced amino acid sequence of the Funaria CDPK shows extensive homology with other CDPKs namely, 73% identity with the Fragaria CDPK and 71% identity with CDPK isoform 7 of Arabidopsis (figure 3). It shows greater homology to the higher plant CDPKs
Figure 3. Comparison of the deduced amino acid sequences of CDPKs from Funaria hygro-metrica (FhCDPK, GenBank Acc. No. AF276999); Fragaria annanasa (FaCDPK, GenBank Acc. No. AAB88537) and the isoform 7 of Arabidopsis thaliana (AtCDPK7, GenBank Acc. No. AAB03247). The alignment was done using Clustal W version 1⋅8 (Thompson et al 1994) and coloured with BoxShade. The boundaries of the variable, kinase and the autoinhibitory domains are demarcated by arrows. The EF hand motifs in the Calmodulin like domain are marked by lines. Dashes represent gaps introduced to maximize the alignment. Black and gray shaded backgrounds indicate amino acid residues that are identical or conservative changes, respectively, in the three sequences.
28
D. MITRA ET AL.

Figure 5. Expression of Funaria CDPK mRNA. About 45 µg of total RNA was electrophoresed on a 1⋅2% agarose gel con-taining formaldehyde and transferred onto a nylon membrane.(A) The cultures were collected after 21, 31, 55 and 95 h of growth in MMG when the cell densities were 2⋅6, 4, 11 and 22 mg/ml, respectively. (B) The cultures were grown in LN and MMG media for 2, 12, 24 and 48 h and the cell densities were0⋅6, 1⋅2, 1⋅7 and 3⋅5 g/ml, respectively. (C) The cultures were grown in LP, LS and MMG media for 24 and 48 h and the cell densities were 1⋅7 and 3⋅6 mg/ml, respectively. The blots were probed with P1. Ethidium bromide staining of rRNA are pre-sented to demonstrate equal loading in all lanes. The 2⋅6 kb transcript of the Funaria CDPK is marked by an arrow.
than to the green alga Chlamydomonas (50% identity, GenBank S57488), or the liverwort, Marchantia (64% identity; GenBank BAA81749) or the moss, Tortula (59% identity; GenBank AAB70706) enzymes. The kinase and the autoinhibitory regions in the protein encoded by the Funaria CDPK show 84% and 73% identity with the respective regions in the Fragaria enzyme, while the sequence identity in the calmodulin like domain is only 60% between the two CDPKs. A comparison of the deduced amino acid sequences of the kinase domains of
the Funaria CDPK with various members of the CDPK superfamily and a few other kinases is presented in figure 4. It is clear from the dendrogram that the Funaria CDPK shows maximum homology to the CDPKs from the angio-sperms such as strawberry and Arabidopsis. Studies by Knight et al (1995) have shown that the stress response pathway is well conserved between the moss Physcomi-trella and the cereal wheat. These two lines of evidence strongly suggest that the signaling mechanisms are well conserved between the mosses Funaria and Physcomitrella
Figure 4. Dendrogram showing the phylogenetic relationships between the Funaria CDPK and other CDPKs, calmodulin dep-endent kinases (CaMK), calcium calmodulin dependent kinases (CCaMK), CDPK related kinases (CRK) and wpk4. The dedu-ced amino acid sequence of the kinase domains of these mole-cules were compared and aligned using Clustal W and the dendrogram was generated on the basis of the alignment using the TreeView software (Page 1996). The kinases shown in the dendrogram are as follows: Picea mariana CDPK (PmCDPK), Zea mays CDPK1 (ZMCDPK1), Arabidopsis thaliana CDPK (AK1), Tortula ruralis CDPK (TrCDPK), Glycine max CDPKβ (GmCDPKβ), Marchantia polymorpha CDPK-B (MpCDPKB), Marchantia polymorpha CDPK-A (MpCDPKA), Arabidopsis thaliana CDPK6 (AtCDPK6), Arabidopsis thaliana CDPK 7 (AtCDPK7), Fragaria ananassa CDPK (FaCDPK), Funaria hygrometrica CDPK (FhCDPK), Dunaliella tertiolecta CDPK1 (DtCDPK1), Chlamydomonas eugametos CDPK (CeCDPK), Zea mays CRK (ZmCRK), Malus domesticus CaMK (MdCaMK), Lilium longiflorum CCaMK (LlCCaMK), Plasmodium falci-parum CDPK2 (PfCDPK2), Rattus norvegicus CaMK (RnCaMK).
29
D. MITRA ET AL.

and higher plants like Arabidopsis and wheat. So it would be interesting to study if CDPK isoforms from Arabi-dopsis, particularly AtCDPK7, respond to starvation. Mosses provide the unique opportunity of a morphologi-cally simple system, which seems to be endowed with molecular mechanisms matching the complexity of those in angiosperms. Our findings further highlight the impor-tance of mosses as model systems to study signaling pathways. Interestingly, the Funaria CDPK has a lysine (at over-all position 364, figure 3) instead of the very well con-served glutamic acid at position 12 of the calcium binding loop in the first EF hand. In the Plasmodium CDPK, an E to K transition at an identical position in the first EF hand has been shown to drastically reduce the calcium affinity and kinase activity of the enzyme (Zhao et al 1994). As the consequences of such changes in the EF hands can not be extrapolated from one system to another (Harmon et al 2000), the consequence of this change on the calcium binding property of the Funaria CDPK remains to be understood. Northern analysis indicates that the expression of the Funaria CDPK transcript is up-regulated by nutrient dep-rivation. The only other kinase, which is induced by nutri-ent starvation in plants, is the wpk 4 (wheat protein kinase 4) from wheat, which belongs to the SnF-1 related kinase 3 (SnRK3) family. The transcript of wpk4 is also induced by light and cytokinin (Sano and Youssefian 1994). The Funaria CDPK was found to respond differently and the transcript levels were unaffected by darkness and cyto-kinin (data not shown). Recent work of Ikeda et al (1999) has shown that wpk4 can phosphorylate and thereby inac-tivate 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGR). HMGR catalyses a key step in the isoprenoid biosynthetic pathway. Nutritional starvation is known to lead to a down-regulation of anabolic processes. The delay in the up-regulation (24 to 48 h) of the FhCDPK transcript levels suggest that it is more likely to be associated with the acclimation response rather than the starvation sensing mechanism. Similar to the role of wpk4 in inactivation of HMGR, it is plausible that the FhCDPK also plays a role in down-regulating anabolic pathways in the acclimation response to nutrient starva-tion. In this context, it may be pointed out that the spinach CDPK has been shown to inactivate NR (nitrate reduc-tase) by phosphorylation under in vitro conditions (Doug-las et al 1998). Calcium dependent phosphorylation and subsequent inactivation of the spinach sucrose phosphate synthase (SPS) has also been demonstrated (McMichael et al 1995). NR and SPS catalyse key steps in the nitrogen assimilation and sucrose synthesis pathways, respectively. Against the backdrop of these observations, we would like to propose that the Funaria CDPK could be involved in the down regulation of NR and SPS during starvation conditions.
Acknowledgements
We thank Prof. B J Rao and Prof. S Sharma for fruitful discussions and critical reading of the manuscript.
References
Botella J R, Arteca J M, Somodevilla M and Arteca R N 1996 Calcium-dependent protein kinase gene expression in res-ponse to physical and chemical stimuli in mungbean (Vigna radiata); Plant Mol. Biol. 30 1129–1137
Bush D 1995 Calcium regulation in plant cells and its role in signaling; Annu. Rev. Plant Physiol. Plant Mol. Biol. 46 95–122
Dellaporta S L, Wood J and Hicks J B 1984 Molecular biology of plants (Cold Spring Harbor: Cold Spring Harbor Labora-tory Press)
Douglas P, Moorhead G, Hong Y, Morrice N and MacKintosh C 1998 Purification of a nitrate reductase kinase from Spina-cea oleracea leaves and its identification as a calmodulin–domain protein kinase; Planta 206 435–442
D’Souza J S and Johri M M 1999 Ca2+ dPKs from the pro-tonema of the moss Funaria hygrometrica. Effect of indole–acetic acid and cultural parameters on the activity of a 44 kDa Ca2+ dPK; Plant Sci. 145 23–32
Frattini M, Morello L and Brevario D 1999 Rice calcium–dependent protein kinase isoforms OsCDPK2 and OsCDPK11 show differential responses to light and different expression patterns during seed development; Plant Mol. Biol. 41 753–764
Handa A K and Johri M M 1977 Cyclic adenosine 3′5′-mono-phosphate in moss protonema; Plant Physiol. 59 490–496
Harmon A C, Gribskov M and Harper J 2000 CDPKs – a kinase for every Ca2+ signal; Trends Plant Sci. 5 154–159
Harmon A C, Yoo B-C and McFaffery C 1994 Pseudosubstrate inhibition of CDPK, a protein kinase with a calmodulin-like domain; Biochemistry 33 7278–7287
Ikeda Y, Koizumi N, Kusano T and Sano H 1999 Sucrose and Cytokinin Modulation of WPK4, a Gene Encoding a SNF1-Related Protein Kinase from Wheat; Plant Physiol. 121 813–820
Knight C D, Sehgal A, Atwal K, Wallace J, Cove D J, Coates D, Quatrano R S, Bahadur S, Stockley P G and Cuming A C 1995 Molecular responses to Abscissic acid and stress are conserved between moss and cereals; Plant Cell 7 499–506
Lee J Y, Yoo B C and Harmon A C 1998 Kinetic and calcium-binding properties of three calcium-dependent protein kinase isoenzymes from soybean; Biochemistry 37 6801–6809
McMichael R W, Bachmann M and Huber S C 1995 Spinach leaf sucrose-phosphatase synthase and nitrate reductase are phosphorylated by multiple protein kinases in vitro; Plant Physiol. 108 1077–1082
Page R D M 1996 TREEVIEW: An application to display phy-logenetic trees on personal computers; Comput. Appl. Biosci. 12 357–358
Picton J M and Steer M W 1983 Evidence for the role of Ca2+ ions in tip extension of pollen tubes; Protoplasma 115 11–17
Romeis T, Piedras P and Jones J D 2000 Resistance gene-dependent activation of a calcium-dependent protein kinase in the plant defense response; Plant Cell 12 803–816
Sambrook J, Fritsch E and Maniatis T 1989 Molecular cloning: A laboratory manual (Cold Spring Harbor: Cold Spring Har-bor Laboratory Press)
30
D. MITRA ET AL.

Sano H and Youssefian S 1994 Light and nutritional regulation of transcripts encoding a wheat protein kinase homolog is mediated by cytokinins; Proc. Natl. Acad. Sci. USA 91 2582–2586
Saunders M J and Hepler P K 1983 Calcium antagonists and calmodulin inhibitors block cytokinin-induced bud formation in Funaria; Dev. Biol. 1 41–49
Sheen J 1996 Ca2+-dependent protein kinases and stress signal transduction in plants; Science 274 1900–1902
Thompson J D, Higgins D G and Gibson T J 1994 CLUSTALW: improving the sensitivity of progressive multi-ple sequence alignment through sequence weighting, posi-tions-specific gap penalties and weight matrix choice; Nucleic Acids Res. 22 4673–4680
Urao T, Katagiri T, Mizoguchi T, Yamaguchi-Shinozaki K, Hayashida N and Shinozaki K 1994 Two genes that encode Ca(2+)-dependent protein kinases are induced by drought and high salt stress in Arabidopsis thaliana; Mol. Gen. Genet. 244 331–340
Yoon G M, Cho H S, Ha H J, Liu J R and Lee H S 1999 Charac-terization of NtCDPK1, a calcium–dependent protein kinase gene in Nicotiana tabacum and the activity of its encoded protein; Plant Mol. Biol. 39 991–1001
Zhao Y, Pokutta S, Maurer P, Lindt M, Franklin R M and Kap-pes B 1994 Calcium-binding properties of a calcium-dependent protein kinase from Plasmodium falciparum and the significance of individual calcium-binding sites for kinase activation; Biochemistry 33 3714–3721
31
D. MITRA ET AL.

Keywords. DNA transposition; MuA transposase; phage Mu; transpososome assembly
Domain III function of Mu transposase analysed by directed placement of subunits within the transpososome
SUSANA MARICONDA, SOON-YOUNG NAMGOONG, KI-HOON YOON, HONG JIANG and RASIKA M HARSHEY
Assembly of the functional tetrameric form of Mu transposase (MuA protein) at the two att ends of Mu depends on interaction of MuA with multiple att and enhancer sites on supercoiled DNA, and is stimulated by MuB pro-tein. The N-terminal domain I of MuA harbours distinct regions for interaction with the att ends and enhancer; the C-terminal domain III contains separate regions essential for tetramer assembly and interaction with MuB protein (IIIα and IIIβ, respectively). Although the central domain II (the ‘DDE’ domain) of MuA harbours the known catalytic DDE residues, a 26 amino acid peptide within IIIα also has a non-specific DNA binding and nuclease activity which has been implicated in catalysis. One model proposes that active sites for Mu transposition are assembled by sharing structural/catalytic residues between domains II and III present on separate MuA monomers within the MuA tetramer. We have used substrates with altered att sites and mixtures of MuA proteins with either wild-type or altered att DNA binding specificities, to create tetrameric arrangements wherein specific MuA sub-units are nonfunctional in II, IIIα or IIIβ domains. From the ability of these oriented tetramers to carry out DNA cleavage and strand transfer we conclude that domain IIIα or IIIβ function is not unique to a specific subunit within the tetramer, indicative of a structural rather than a catalytic function for domain III in Mu transposition.
1. Introduction
The DNA cleavage and joining reactions of transposition are carried out within a tetrameric unit of the Mu transpo-sase (MuA protein) (figure 1A; reviewed in Mizuuchi, 1992; Lavoie and Chaconas 1995). While MuA can bind six att sites (L1-L3 at the left or attL end, and R1-R3 at the right or attR end), and three internal enhancer sites (O1-O3) on Mu DNA, the MuA tetramer footprints on only three of the att sites (L1, R1 and R2; figure 1B), and only two specific phosphodiester bonds (adjacent to L1 and R1) are cleaved on opposite DNA strands. The result-ing 3′ OH groups are joined or strand transferred to two phosphodiesters placed 5 bp apart on two strands of target DNA. The enhancer interacts with the L and R ends early in the Mu transposition reaction to form a nucleoprotein
complex LER (Watson and Chaconas 1996; see figure 1A), interactions within which lead to formation of a sta-ble type 0 complex in which the Mu transposase (MuA protein) assumes its active tetrameric form, catalyzing the subsequent cleavage (type I complex) and joining (type II complex) reactions (see Chaconas et al 1996). The MuA tetramer is eventually dislodged by the action of a host protein ClpX, followed by replication of Mu (Kruklitis et al 1996; Levchenko et al 1997). The MuA monomer (its normal form in solution; Kuo et al 1991) is a multi-domain protein (Nakayama et al 1987; figure 1C). The amino-terminal domain I contains the recognition motifs for two types of DNA sites – the enhancer and att sites (see Chaconas et al 1996). The cen-tral domain II contains a triad of ‘DDE’ residues present within IIα (refered to here as the DDE domain), that are
Journal of Biophysical Chemistry, 2009, 1, 32-45
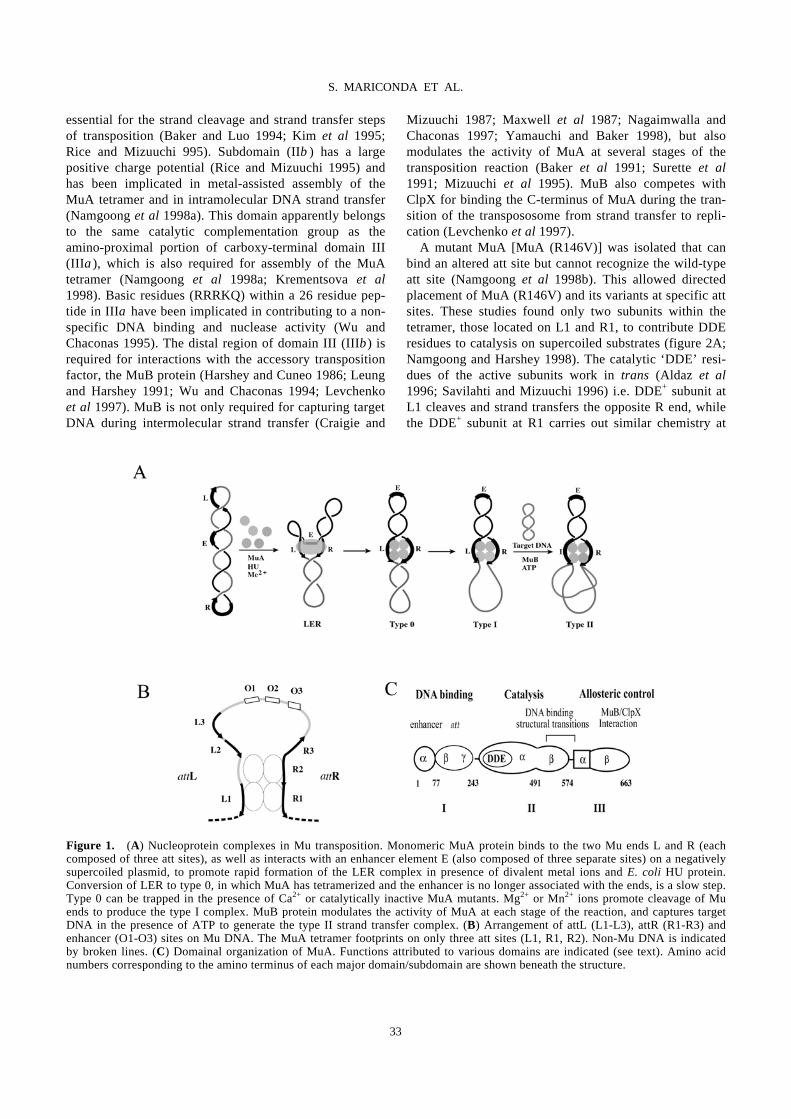
essential for the strand cleavage and strand transfer steps of transposition (Baker and Luo 1994; Kim et al 1995; Rice and Mizuuchi 995). Subdomain (IIβ ) has a large positive charge potential (Rice and Mizuuchi 1995) and has been implicated in metal-assisted assembly of the MuA tetramer and in intramolecular DNA strand transfer (Namgoong et al 1998a). This domain apparently belongs to the same catalytic complementation group as the amino-proximal portion of carboxy-terminal domain III (IIIα), which is also required for assembly of the MuA tetramer (Namgoong et al 1998a; Krementsova et al 1998). Basic residues (RRRKQ) within a 26 residue pep-tide in IIIα have been implicated in contributing to a non-specific DNA binding and nuclease activity (Wu and Chaconas 1995). The distal region of domain III (IIIβ) is required for interactions with the accessory transposition factor, the MuB protein (Harshey and Cuneo 1986; Leung and Harshey 1991; Wu and Chaconas 1994; Levchenko et al 1997). MuB is not only required for capturing target DNA during intermolecular strand transfer (Craigie and
Mizuuchi 1987; Maxwell et al 1987; Nagaimwalla and Chaconas 1997; Yamauchi and Baker 1998), but also modulates the activity of MuA at several stages of the transposition reaction (Baker et al 1991; Surette et al 1991; Mizuuchi et al 1995). MuB also competes with ClpX for binding the C-terminus of MuA during the tran-sition of the transpososome from strand transfer to repli-cation (Levchenko et al 1997). A mutant MuA [MuA (R146V)] was isolated that can bind an altered att site but cannot recognize the wild-type att site (Namgoong et al 1998b). This allowed directed placement of MuA (R146V) and its variants at specific att sites. These studies found only two subunits within the tetramer, those located on L1 and R1, to contribute DDE residues to catalysis on supercoiled substrates (figure 2A; Namgoong and Harshey 1998). The catalytic ‘DDE’ resi-dues of the active subunits work in trans (Aldaz et al 1996; Savilahti and Mizuuchi 1996) i.e. DDE+ subunit at L1 cleaves and strand transfers the opposite R end, while the DDE+ subunit at R1 carries out similar chemistry at
Figure 1. (A) Nucleoprotein complexes in Mu transposition. Monomeric MuA protein binds to the two Mu ends L and R (each composed of three att sites), as well as interacts with an enhancer element E (also composed of three separate sites) on a negatively supercoiled plasmid, to promote rapid formation of the LER complex in presence of divalent metal ions and E. coli HU protein. Conversion of LER to type 0, in which MuA has tetramerized and the enhancer is no longer associated with the ends, is a slow step. Type 0 can be trapped in the presence of Ca2+ or catalytically inactive MuA mutants. Mg2+ or Mn2+ ions promote cleavage of Mu ends to produce the type I complex. MuB protein modulates the activity of MuA at each stage of the reaction, and captures target DNA in the presence of ATP to generate the type II strand transfer complex. (B) Arrangement of attL (L1-L3), attR (R1-R3) and enhancer (O1-O3) sites on Mu DNA. The MuA tetramer footprints on only three att sites (L1, R1, R2). Non-Mu DNA is indicated by broken lines. (C) Domainal organization of MuA. Functions attributed to various domains are indicated (see text). Amino acid numbers corresponding to the amino terminus of each major domain/subdomain are shown beneath the structure.
33
S. MARICONDA ET AL.
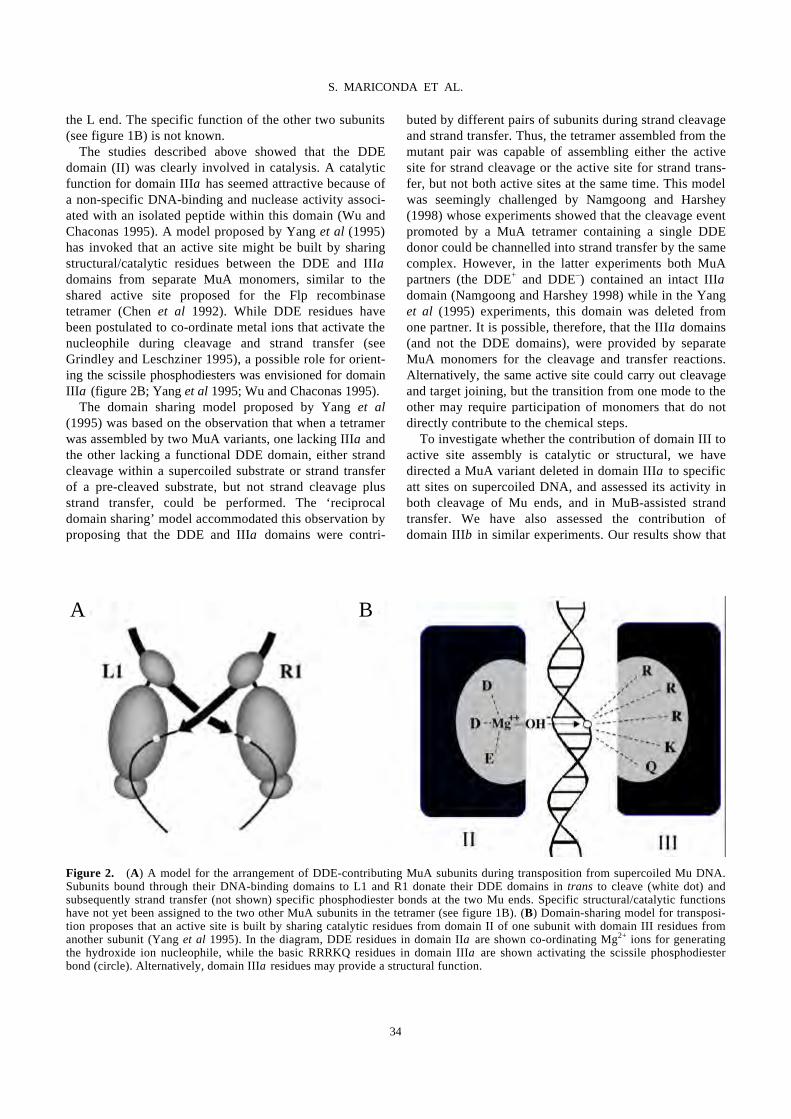
the L end. The specific function of the other two subunits (see figure 1B) is not known. The studies described above showed that the DDE domain (II) was clearly involved in catalysis. A catalytic function for domain IIIα has seemed attractive because of a non-specific DNA-binding and nuclease activity associ-ated with an isolated peptide within this domain (Wu and Chaconas 1995). A model proposed by Yang et al (1995) has invoked that an active site might be built by sharing structural/catalytic residues between the DDE and IIIα domains from separate MuA monomers, similar to the shared active site proposed for the Flp recombinase tetramer (Chen et al 1992). While DDE residues have been postulated to co-ordinate metal ions that activate the nucleophile during cleavage and strand transfer (see Grindley and Leschziner 1995), a possible role for orient-ing the scissile phosphodiesters was envisioned for domain IIIα (figure 2B; Yang et al 1995; Wu and Chaconas 1995). The domain sharing model proposed by Yang et al (1995) was based on the observation that when a tetramer was assembled by two MuA variants, one lacking IIIα and the other lacking a functional DDE domain, either strand cleavage within a supercoiled substrate or strand transfer of a pre-cleaved substrate, but not strand cleavage plus strand transfer, could be performed. The ‘reciprocal domain sharing’ model accommodated this observation by proposing that the DDE and IIIα domains were contri-
buted by different pairs of subunits during strand cleavage and strand transfer. Thus, the tetramer assembled from the mutant pair was capable of assembling either the active site for strand cleavage or the active site for strand trans-fer, but not both active sites at the same time. This model was seemingly challenged by Namgoong and Harshey (1998) whose experiments showed that the cleavage event promoted by a MuA tetramer containing a single DDE donor could be channelled into strand transfer by the same complex. However, in the latter experiments both MuA partners (the DDE+ and DDE–) contained an intact IIIα domain (Namgoong and Harshey 1998) while in the Yang et al (1995) experiments, this domain was deleted from one partner. It is possible, therefore, that the IIIα domains (and not the DDE domains), were provided by separate MuA monomers for the cleavage and transfer reactions. Alternatively, the same active site could carry out cleavage and target joining, but the transition from one mode to the other may require participation of monomers that do not directly contribute to the chemical steps. To investigate whether the contribution of domain III to active site assembly is catalytic or structural, we have directed a MuA variant deleted in domain IIIα to specific att sites on supercoiled DNA, and assessed its activity in both cleavage of Mu ends, and in MuB-assisted strand transfer. We have also assessed the contribution of domain IIIβ in similar experiments. Our results show that
Figure 2. (A) A model for the arrangement of DDE-contributing MuA subunits during transposition from supercoiled Mu DNA. Subunits bound through their DNA-binding domains to L1 and R1 donate their DDE domains in trans to cleave (white dot) and subsequently strand transfer (not shown) specific phosphodiester bonds at the two Mu ends. Specific structural/catalytic functions have not yet been assigned to the two other MuA subunits in the tetramer (see figure 1B). (B) Domain-sharing model for transposi-tion proposes that an active site is built by sharing catalytic residues from domain II of one subunit with domain III residues from another subunit (Yang et al 1995). In the diagram, DDE residues in domain IIα are shown co-ordinating Mg2+ ions for generating the hydroxide ion nucleophile, while the basic RRRKQ residues in domain IIIα are shown activating the scissile phosphodiester bond (circle). Alternatively, domain IIIα residues may provide a structural function.
A B
34
S. MARICONDA ET AL.

unlike the DDE contribution, where two unique subunits of MuA (see figure 2A) provide DDE residues in trans for both cleavage and strand transfer, no unique position could be identified for the IIIα contribution. These results could be extended to include domain IIIβ function (MuB interaction domain) as well. Our studies also shed light on the original observation by Yang et al (1995) which prompted the ‘reciprocal domain sharing’ model (see above).
2. Materials and methods
2.1 DNA substrates and proteins
pL1**, pR2** and pJMM plasmids have been described (Namgoong and Harshey 1998; Jiang et al 1999). Plasmid pL1**-R2** was constructed by appropriate exchange of restriction fragments between pL1** and pR2**. MuA (E392A), MuA (R146V) and MuA (∆560-663) have been described (Kim et al 1995; Namgoong et al 1998a, b). MuA (∆609-663) was constructed by PCR mutagenesis procedures similar to those described for MuA (∆560-663) (Namgoong et al 1998a). The R146V mutation was moved into these variants by appropriate restriction fragment exchange. Heart muscle kinase (HMK) recognition tags were engineered at the N-termini of MuA and its derivatives by addition of the sequence MGSRRASV (Li et al 1989; the underlined sequence is the kinase recognition pentapeptide) before the MuA start codon (at the NdeI site on the pET vector) by PCR metho-dology. Wild-type and variant forms of MuA, as well as MuB and HU proteins were purified as described by Yang et al (1995). The kinase-tagged proteins required a slight modi-fication in the purification procedure: SP-sepharose fast-flow column was substituted for phosphocellulose and the pH at this step was maintained at 7⋅0.
2.2 Radiolabelling HMK-tagged proteins
Type I reactions with HMK-tagged proteins were per-formed in 20 µl solutions containing 25 mM Hepes (pH 7⋅8), 10 mM MgCl2, 100 mM NaCl, 1 mM DTT, at 30°C for 20 min, followed by addition of 1 µg of heparin to remove loosely bound protein from the DNA. Labelling reactions were done by addition of 1 µl (γ-32P) ATP (10 µCi) and 1u HMK (Sigma P2645), at 30°C for 5 min. Free (γ-32P) ATP was removed by passing the reaction mixture over a Centri-sep gel filtration spin column (Prince-ton Separation). After agarose gel electrophoresis, the gel was dried and exposed to film on a BioRad Molecular Imager.
2.3 Mu DNA cleavage and strand transfer
Type I cleavage reactions and type II strand transfer reac-tions were carried out as described by Namgoong et al (1998a).
2.4 Determination of Mu end cleavage
Two methods were employed to detect cleavage at the left and right Mu ends. Primer extension analysis was carried out as described by Namgoong and Harshey (1998), while end-labelling of appropriately digested restriction frag-ments with (α-32P) cordycepin phosphate and terminal nucleotidyl transferase was carried out as described by Jiang et al (1999).
3. Results
3.1 Domain IIIα function of MuA subunits at L1 or R2 sites is not required for DNA cleavage or strand transfer
3.1a Domain IIIα function of the L1 subunit in cleavage of R1: Under normal reaction conditions, subunits con-tributing DDE domains occupy specific positions within the MuA tetramer (see figure 2A). If IIIα domains par-ticipate in catalysis (see figure 2B), the expectation is that the positions of subunits contributing these domains will also be specific. This section describes the IIIα function of subunits at L1 and R2 only (see figure 1B). This is because the altered att site functions poorly at R1 due to positional effects (Namgoong and Harshey 1998). Three separate arrangements of MuA tetramers were configured as shown in figures 3 and 4. The double aster-isk denotes presence of the altered att site to which the R146V DNA-binding variant specifically binds. Note that although the plasmids used in this study contain all six att sites (see figure 1B), for clarity only those on which the tetramer footprints are indicated. Note also that MuA with a wild-type att DNA-binding domain can also recognize altered att sites; therefore, in all reactions containing a MuA (R146V) variant plus a second protein, the substrate was pre-incubated with the R146V-containing protein prior to the addition of its partner protein, ensuring that the altered sites were selectively blocked from binding to the protein without the R146V substitution. In the first set of experiments (figure 3), a domain IIIα
– DDE+ mutant [MuA (R146V) (∆560-663)] was placed at L1 and a DDE– IIIα+ mutant [MuA (E392A)] at all other sites on plasmid pL1** (figure 3A). Generation of a type I complex (figure 3B, lane 4) indicated that cleavage had occurred in this equimolar mixture of proteins, but not in controls with either protein alone (figure 3B, lanes 2 and 3; the DDE– protein can assemble an uncleaved type 0
35
S. MARICONDA ET AL.
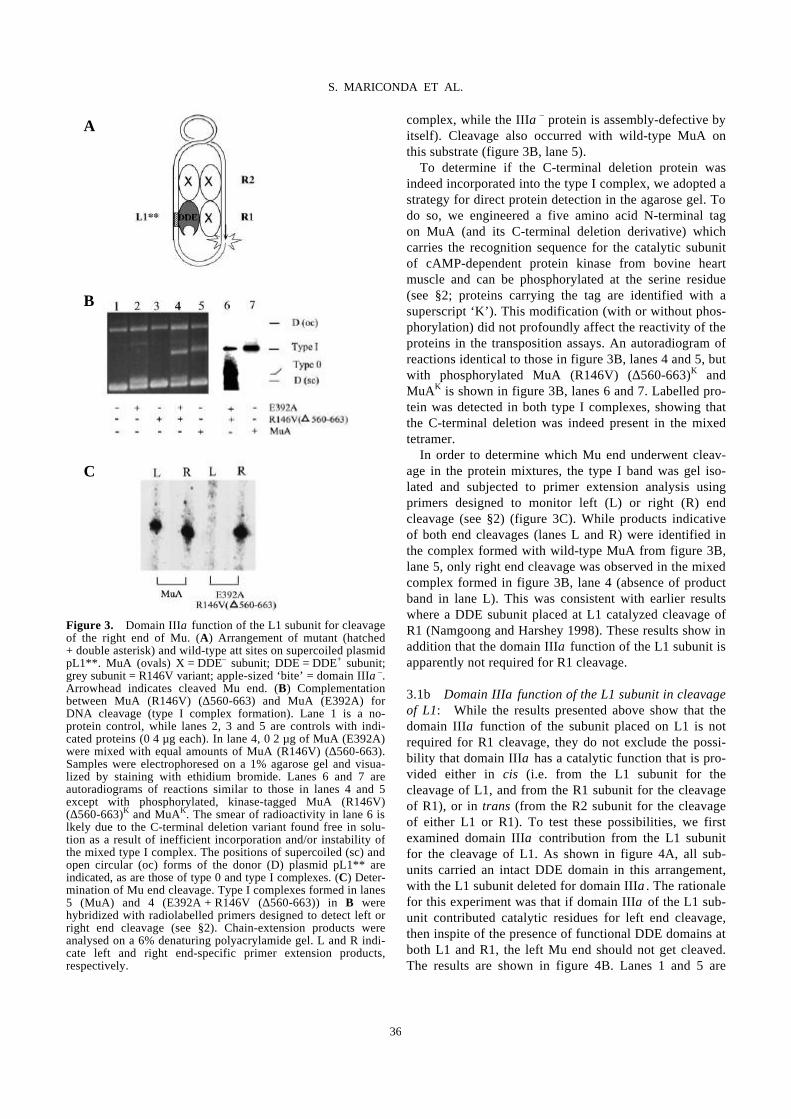
complex, while the IIIα – protein is assembly-defective by
itself). Cleavage also occurred with wild-type MuA on this substrate (figure 3B, lane 5). To determine if the C-terminal deletion protein was indeed incorporated into the type I complex, we adopted a strategy for direct protein detection in the agarose gel. To do so, we engineered a five amino acid N-terminal tag on MuA (and its C-terminal deletion derivative) which carries the recognition sequence for the catalytic subunit of cAMP-dependent protein kinase from bovine heart muscle and can be phosphorylated at the serine residue (see §2; proteins carrying the tag are identified with a superscript ‘K’). This modification (with or without phos-phorylation) did not profoundly affect the reactivity of the proteins in the transposition assays. An autoradiogram of reactions identical to those in figure 3B, lanes 4 and 5, but with phosphorylated MuA (R146V) (∆560-663)K and MuAK is shown in figure 3B, lanes 6 and 7. Labelled pro-tein was detected in both type I complexes, showing that the C-terminal deletion was indeed present in the mixed tetramer. In order to determine which Mu end underwent cleav-age in the protein mixtures, the type I band was gel iso-lated and subjected to primer extension analysis using primers designed to monitor left (L) or right (R) end cleavage (see §2) (figure 3C). While products indicative of both end cleavages (lanes L and R) were identified in the complex formed with wild-type MuA from figure 3B, lane 5, only right end cleavage was observed in the mixed complex formed in figure 3B, lane 4 (absence of product band in lane L). This was consistent with earlier results where a DDE subunit placed at L1 catalyzed cleavage of R1 (Namgoong and Harshey 1998). These results show in addition that the domain IIIα function of the L1 subunit is apparently not required for R1 cleavage. 3.1b Domain IIIα function of the L1 subunit in cleavage of L1: While the results presented above show that the domain IIIα function of the subunit placed on L1 is not required for R1 cleavage, they do not exclude the possi-bility that domain IIIα has a catalytic function that is pro-vided either in cis (i.e. from the L1 subunit for the cleavage of L1, and from the R1 subunit for the cleavage of R1), or in trans (from the R2 subunit for the cleavage of either L1 or R1). To test these possibilities, we first examined domain IIIα contribution from the L1 subunit for the cleavage of L1. As shown in figure 4A, all sub-units carried an intact DDE domain in this arrangement, with the L1 subunit deleted for domain IIIα. The rationale for this experiment was that if domain IIIα of the L1 sub-unit contributed catalytic residues for left end cleavage, then inspite of the presence of functional DDE domains at both L1 and R1, the left Mu end should not get cleaved. The results are shown in figure 4B. Lanes 1 and 5 are
Figure 3. Domain IIIα function of the L1 subunit for cleavage of the right end of Mu. (A) Arrangement of mutant (hatched + double asterisk) and wild-type att sites on supercoiled plasmid pL1**. MuA (ovals) X = DDE– subunit; DDE = DDE+ subunit; grey subunit = R146V variant; apple-sized ‘bite’ = domain IIIα–. Arrowhead indicates cleaved Mu end. (B) Complementation between MuA (R146V) (∆560-663) and MuA (E392A) for DNA cleavage (type I complex formation). Lane 1 is a no-protein control, while lanes 2, 3 and 5 are controls with indi-cated proteins (0⋅4 µg each). In lane 4, 0⋅2 µg of MuA (E392A) were mixed with equal amounts of MuA (R146V) (∆560-663). Samples were electrophoresed on a 1% agarose gel and visua-lized by staining with ethidium bromide. Lanes 6 and 7 are autoradiograms of reactions similar to those in lanes 4 and 5 except with phosphorylated, kinase-tagged MuA (R146V) (∆560-663)K and MuAK. The smear of radioactivity in lane 6 is lkely due to the C-terminal deletion variant found free in solu-tion as a result of inefficient incorporation and/or instability of the mixed type I complex. The positions of supercoiled (sc) and open circular (oc) forms of the donor (D) plasmid pL1** are indicated, as are those of type 0 and type I complexes. (C) Deter-mination of Mu end cleavage. Type I complexes formed in lanes 5 (MuA) and 4 (E392A + R146V (∆560-663)) in B were hybridized with radiolabelled primers designed to detect left or right end cleavage (see §2). Chain-extension products were analysed on a 6% denaturing polyacrylamide gel. L and R indi-cate left and right end-specific primer extension products, respectively.
A
B
C
36
S. MARICONDA ET AL.
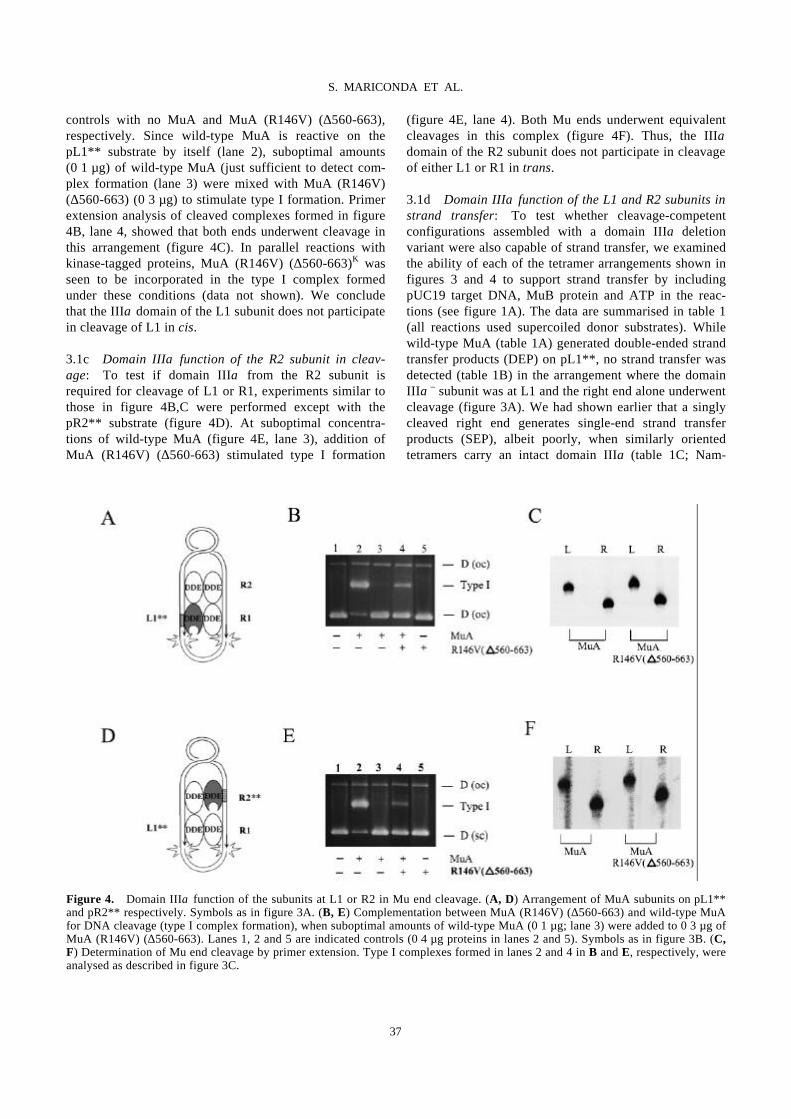
controls with no MuA and MuA (R146V) (∆560-663), respectively. Since wild-type MuA is reactive on the pL1** substrate by itself (lane 2), suboptimal amounts (0⋅1 µg) of wild-type MuA (just sufficient to detect com-plex formation (lane 3) were mixed with MuA (R146V) (∆560-663) (0⋅3 µg) to stimulate type I formation. Primer extension analysis of cleaved complexes formed in figure 4B, lane 4, showed that both ends underwent cleavage in this arrangement (figure 4C). In parallel reactions with kinase-tagged proteins, MuA (R146V) (∆560-663)K was seen to be incorporated in the type I complex formed under these conditions (data not shown). We conclude that the IIIα domain of the L1 subunit does not participate in cleavage of L1 in cis. 3.1c Domain IIIα function of the R2 subunit in cleav-age: To test if domain IIIα from the R2 subunit is required for cleavage of L1 or R1, experiments similar to those in figure 4B,C were performed except with the pR2** substrate (figure 4D). At suboptimal concentra-tions of wild-type MuA (figure 4E, lane 3), addition of MuA (R146V) (∆560-663) stimulated type I formation
(figure 4E, lane 4). Both Mu ends underwent equivalent cleavages in this complex (figure 4F). Thus, the IIIα domain of the R2 subunit does not participate in cleavage of either L1 or R1 in trans. 3.1d Domain IIIα function of the L1 and R2 subunits in strand transfer: To test whether cleavage-competent configurations assembled with a domain IIIa deletion variant were also capable of strand transfer, we examined the ability of each of the tetramer arrangements shown in figures 3 and 4 to support strand transfer by including pUC19 target DNA, MuB protein and ATP in the reac-tions (see figure 1A). The data are summarised in table 1 (all reactions used supercoiled donor substrates). While wild-type MuA (table 1A) generated double-ended strand transfer products (DEP) on pL1**, no strand transfer was detected (table 1B) in the arrangement where the domain IIIα
– subunit was at L1 and the right end alone underwent cleavage (figure 3A). We had shown earlier that a singly cleaved right end generates single-end strand transfer products (SEP), albeit poorly, when similarly oriented tetramers carry an intact domain IIIa (table 1C; Nam-
Figure 4. Domain IIIα function of the subunits at L1 or R2 in Mu end cleavage. (A, D) Arrangement of MuA subunits on pL1** and pR2** respectively. Symbols as in figure 3A. (B, E) Complementation between MuA (R146V) (∆560-663) and wild-type MuA for DNA cleavage (type I complex formation), when suboptimal amounts of wild-type MuA (0⋅1 µg; lane 3) were added to 0⋅3 µg of MuA (R146V) (∆560-663). Lanes 1, 2 and 5 are indicated controls (0⋅4 µg proteins in lanes 2 and 5). Symbols as in figure 3B. (C, F) Determination of Mu end cleavage by primer extension. Type I complexes formed in lanes 2 and 4 in B and E, respectively, were analysed as described in figure 3C.
37
S. MARICONDA ET AL.

goong and Harshey 1998). Comparison of the results in B and C might suggest that domain IIIa plays a role in strand transfer. However, when the tetramer arrangement in figure 4A was tested for strand transfer, in spite of the fact that L1 position was still occupied by a domain IIIa
– subunit, double-end strand transfer was detected (table 1D). Thus, domain IIIa is not required for strand transfer of either end. Strand transfer reactions performed using the MuA subunit arrangement shown in figure 4D, also showed the presence of double-ended strand transfer
products (table 1E). Thus, domain IIIa function at R2 is also not required for the strand transfer of either end. In summary, the data in figures 3, 4 and table 1 suggest that domain IIIα does not provide a catalytic function for cleavage or strand transfer of Mu ends. The finding that IIIα function of the L1 subunit is required only when the right end alone has undergone cleavage but not when both ends are cleaved (compare table 1B and D), is indicative of a structural rather than a catalytic impediment to strand transfer in these mixed tetramers.
3.2 A domain IIIα€– subunit is preferentially
excluded from R1 when randomly mixed with a domain IIIα+ partner
Yang et al (1995) had observed that mixtures of MuA (E392A) and MuA (∆560-663) on wild-type substrates could perform cleavage but not strand transfer. In light of the results in table 1B where singly cleaved R end com-plexes do not support strand transfer when a IIIα– subunit is at L1, we wondered whether only a single end had un-dergone cleavage in the protein mixtures tested by Yang et al (1995). We therefore repeated their experiment (figure 5), and analysed the type I complexes by end-labelling, a procedure that allows a better quantitation of the proportion of left and right end cleavages (see Jiang et al 1999). The donor plasmid pJMM carries all wild-type att sites (Jiang et al 1999). Wild-type MuA showed efficient type I complex formation on this substrate (figure 5A, lane a). While, MuA (∆560-663) and MuA (E392A) showed no type I activity on their own (figure 5A, lanes b and f, res-pectively), mixtures of these two proteins in various indi-cated molar ratios gave rise to the type I product (figure 5A, lanes c–e). DNA from type I complexes formed in lanes a and d was extracted as described by Jiang et al (1999) and digested with BamHI and XbaI in one case, and with BamHI and AatII in the other (figure 5B). The 3′-hydroxyl ends generated from the digestion as well as those produced by cleavage of Mu ends were radio-actively labelled with 32P and fractionated by electropho-resis in denaturing polyacrylamide gels. The diagnostic bands for left end cleavage are LC1 and LC2; those for right end cleavage are RC1 and RC2. For more details, the reader may refer to Jiang et al (1999). No cleavage was detected at the Mu ends in control DNA not reacted with transposase (figure 5C, lanes 1 and 4). The type I complex formed by wild-type MuA (figure 5A, lane a) had undergone efficient cleavage at both L and R ends (figure 5C, lanes 2 and 5; LC1, LC2 and RC1 in lane 2 and RC1, RC2 and LC1 in lane 5). By compari-son, the type I product from the d reaction shown in figure 5A showed nearly exclusive cleavage at the right end
Table 1. Strand transfer proficiency of oriented MuA tetra-mers incorporating a domain IIIα– subunit at L1 or R2 on
supercoiled donor substrates.
38
S. MARICONDA ET AL.

(RC1 with undetectable LC1 and LC2 in lane 3; RC1 and RC2 in lane 6). Note that the disappearance of the RU1 band in lanes 5 and 6 is indicative of complete right end cleavage. The results in figure 5 show that the domain IIIα– sub-unit preferentially occupies L1 in this protein mixture. These results explain why Yang et al (1995) failed to observe strand transfer by this pair of proteins, since the combination of a singly cleaved right end and a domain IIIα– subunit at L1 (figure 3) is structurally impaired in transposition (table 1B). The data in figure 5C also show that for the L1 end to have undergone cleavage (albeit poorly), a domain IIIα– DDE+ subunit was likely posi-tioned at R1 in a small proportion of complexes.
To summarize the results thus far (figures 3–5 and table 1), cleavage at L1 does not depend on an intact domain IIIα at L1, R1 or R2. Cleavage at R1 does not depend on an intact domain IIIα at L1 or R2. While we could not assess if the domain IIIα of the R1 subunit is required for cleavage of R1, such a function is unlikely given the results with the L1 subunit. Strand transfer of cleaved left and right ends does not depend on domain IIIα function of the subunits at L1 or R2. While domain IIIα does not participate in the catalysis of transposition, it appears to provide a structural role; this role must be critical at the R1 site since a IIIα€
– subunit is preferentially excluded from R1 when randomly mixed with a domain IIIα+ partner.
Figure 5. Domain IIIα– subunit is excluded from R1 on wild-type substrates. (A) Complementation between MuA (∆560-663) and MuA (E392A) for type I complex formation on pJMM donor substrate (lanes c–e). Lanes a, b and f are indicated controls (0⋅4 µg protein each). Symbols as in figure 3B. (B) End-labelling strategy to assay left and right end cleavages in the type I complex. The top and bottom strands of the Mu genome are represented by unfilled and filled bars, respectively. The diamonds indicate the strand cleavage positions. Double digestion with BamHI-XbaI or BamHI-AatII, followed by 3′ end labelling (indicated by the asterisk) would give rise to the indicated radioactive products. They can be revealed by electrophoresis in denaturing polyacrylamide gels (see C). Uncleaved attL generates an LU doublet consisting of a 95 nt fragment from the bottom strand, which has the same length as the fragment from the top strand; uncleaved attR generates RU1 from the top strand, which differs in length by 8 nt from the corresponding bottom strand fragment RU2. The products specific to left and right end cleavages are denoted by LC and RC, respectively. (C) The type I product from reactions in lanes a and d of A, were analysed by the strategy outlined in B and are shown in lanes 2/5 and 3/6 respectively. Lanes 1 and 4 represent the substrate DNA that was not treated with MuA. B/X and B/A stand for BamHI-XbaI and BamHI-AatII restriction digestions.
39
S. MARICONDA ET AL.

3.3 At least two subunits must encode IIIβ function for MuB-mediated strand transfer
The MuB protein interacts with MuA through domain IIIβ and modulates its efficiency at several stages in the trans-position pathway prior to strand transfer (see Chaconas et al 1996). In addition, MuB is responsible for binding
target DNA and transporting it to the MuA tetramer for strand transfer. In all of the experiments described above, the IIIα– subunit is also deleted in domain IIIβ (see figure 1C). Thus, one can see that domain IIIβ function at either L1 or R2 is not required for MuB-assisted strand transfer (see table 1). A similar conclusion was reached in earlier experiments using complementing mixtures of proteins carrying a domain IIIβ deletion and/or a DDE– mutation (Mizuuchi et al 1995). These experiments did not involve the use of oriented tetramers as reported here. Mizuuchi et al (1995) concluded that not all four MuA subunits need to interact with MuB and that the position of MuB-interacting subunits is not unique. To determine the minimal number of subunits required for MuB interaction, we set up several different arrange-ments of MuA tetramers (table 2). These included placing a single wild-type (DDE+) subunit at either at L1 or R2 with domain IIIβ – subunits at the other sites, diagonal placement of two wild-type or two IIIβ – subunits at L1-R2, as well as diagonal combinations of domain IIIβ – and DDE– subunits. The data for two of these arrangements are shown in figure 6, while those for all of them are summarised in table 2. Experiments shown in figure 6 test strand transfer from plasmid pL1** when a single functional domain IIIβ + subunit, MuA (R146V), was placed at L1 and MuA (∆609-663) (domain IIIβ –) subunits were at the other sites (figure 6A). Strand transfer reactions were similar to cleavage reactions described under figures 3–5, except they included pUC19 target DNA, MuB protein and ATP. SDS was added to the reactions prior to electrophoresis, in order to dissociate DNA-protein complexes. Therefore, the type I complex would be detected as the open circular plasmid in this assay [Donor(oc)]. While wild-type MuA generated double-ended strand transfer products that migrated as a series of distinct bands (reflecting the dis-tribution of topoisomers retained in the Mu sequence of the donor plasmid) just above the open circular form of the target DNA (DEP, lane 5), no strand transfer products were detected with a mixture of MuA (R146V) and MuA (∆609-663) (figure 6B, lane 4). Controls with either no protein, or with MuA (R146V) and MuA (∆609-663) alone are shown in figure 6B, lanes 1–3. The increase in the open circular form of the donor plasmid in lanes 3 and 4 is indicative of Mu end cleavage. These results are tabu-lated in table 2B. Similar results were seen with this pro-tein mixture on the pR2** substrate (table 2C). Thus, it appears that a single domain IIIβ + subunit cannot support MuB-mediated strand transfer from the L1 or R2 positions. When two MuA (R146V) (domain IIIβ +) subunits were placed at L1 and R2 on the pL1**-R2** substrate, with MuA (∆609-663) (domain IIIβ –) subunits at the other sites (figure 6C), strand transfer was observed with the
Table 2. Strand transfer proficiency of oriented tetramers incorporating a domain IIIβ – subunit at L1, R1 and R2
on supercoiled donor substrates.
40
S. MARICONDA ET AL.
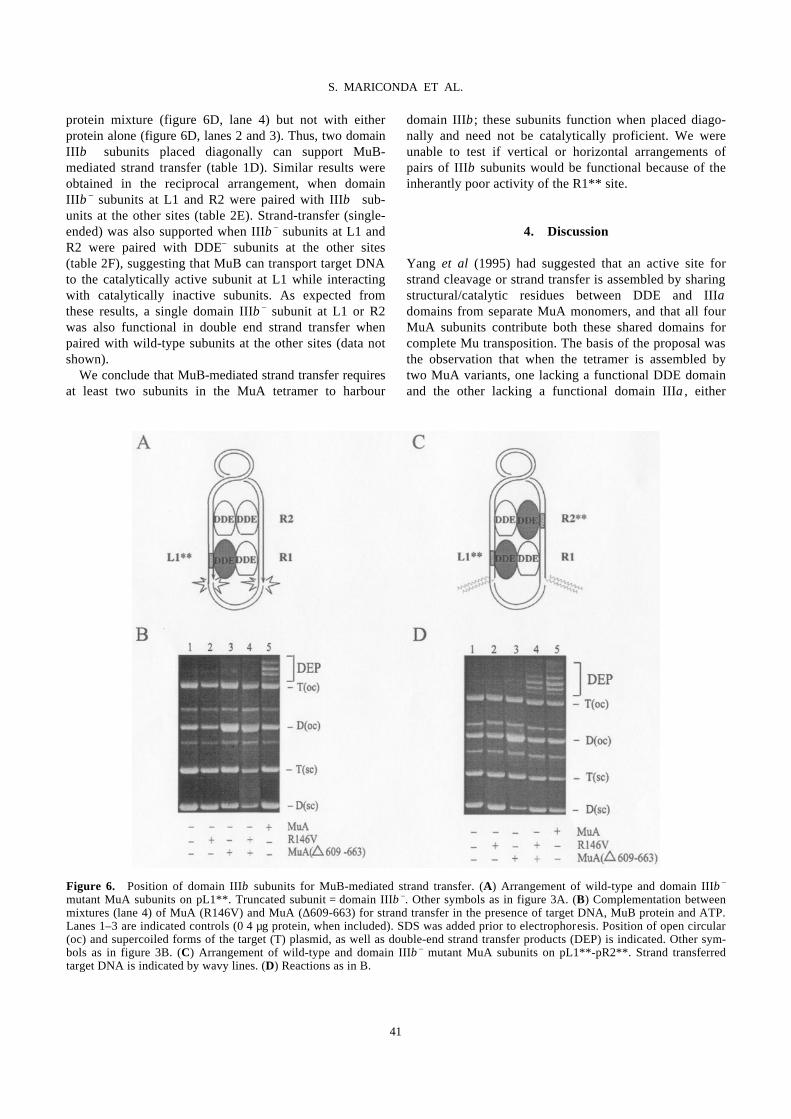
protein mixture (figure 6D, lane 4) but not with either protein alone (figure 6D, lanes 2 and 3). Thus, two domain IIIβ + subunits placed diagonally can support MuB-mediated strand transfer (table 1D). Similar results were obtained in the reciprocal arrangement, when domain IIIβ – subunits at L1 and R2 were paired with IIIβ + sub-units at the other sites (table 2E). Strand-transfer (single-ended) was also supported when IIIβ – subunits at L1 and R2 were paired with DDE– subunits at the other sites (table 2F), suggesting that MuB can transport target DNA to the catalytically active subunit at L1 while interacting with catalytically inactive subunits. As expected from these results, a single domain IIIβ – subunit at L1 or R2 was also functional in double end strand transfer when paired with wild-type subunits at the other sites (data not shown). We conclude that MuB-mediated strand transfer requires at least two subunits in the MuA tetramer to harbour
domain IIIβ; these subunits function when placed diago-nally and need not be catalytically proficient. We were unable to test if vertical or horizontal arrangements of pairs of IIIβ subunits would be functional because of the inherantly poor activity of the R1** site.
4. Discussion
Yang et al (1995) had suggested that an active site for strand cleavage or strand transfer is assembled by sharing structural/catalytic residues between DDE and IIIα domains from separate MuA monomers, and that all four MuA subunits contribute both these shared domains for complete Mu transposition. The basis of the proposal was the observation that when the tetramer is assembled by two MuA variants, one lacking a functional DDE domain and the other lacking a functional domain IIIα, either
Figure 6. Position of domain IIIβ subunits for MuB-mediated strand transfer. (A) Arrangement of wild-type and domain IIIβ – mutant MuA subunits on pL1**. Truncated subunit = domain IIIβ –. Other symbols as in figure 3A. (B) Complementation between mixtures (lane 4) of MuA (R146V) and MuA (∆609-663) for strand transfer in the presence of target DNA, MuB protein and ATP. Lanes 1–3 are indicated controls (0⋅4 µg protein, when included). SDS was added prior to electrophoresis. Position of open circular (oc) and supercoiled forms of the target (T) plasmid, as well as double-end strand transfer products (DEP) is indicated. Other sym-bols as in figure 3B. (C) Arrangement of wild-type and domain IIIβ – mutant MuA subunits on pL1**-pR2**. Strand transferred target DNA is indicated by wavy lines. (D) Reactions as in B.
41
S. MARICONDA ET AL.

strand cleavage within a supercoiled substrate or strand transfer of a precleaved substrate, but not strand cleavage plus strand transfer, can be performed. The inspiration for the shared active model was derived from experiments in the Flp recombination system, where a monomer of Flp bound to its recognition site orients an adjacent phos-phodiester for nucleophilic attack by the active site tyro-sine derived in trans from a second Flp monomer (Chen et al 1992). In the Mu active site, DDE residues were pro-posed to provide the nucleophile while IIIα residues could orient the scissile phosphodiester (figure 2B; Yang et al 1995). The demonstration of a non-specific nuclease activity associated with domain IIIα heightened this speculation (Wu and Chaconas 1995). Analysis of trans-position from linear substrates under altered reaction con-ditions (e.g. inclusion of DMSO in the reaction; Yang et al 1996), was not inconsistent with the Yang et al (1995) proposal. Using oriented tetramers, recent experiments on super-coiled Mu substrates have shown that unlike the results derived from linear substrates, only two DDE+ subunits (located at L1 and R1 and acting in trans) sequentially carry out strand cleavage followed by strand transfer (fig-ure 2A; Namgoong and Harshey 1998). (The trans action of DDE was also observed on linear substrates employing two right ends; Aldaz et al 1996; Savilahti and Mizuuchi 1996.) To reconcile the results of Namgoong and Harshey (1998) with those of Yang et al (1995), both of which were performed under normal reaction conditions using supercoiled substrates, two possibilities suggested them-selves. The first was that although the same DDE domains are used during cleavage and strand transfer, perhaps dif-ferent IIIα domains are required during these two steps. The second was that the IIIα deletion variant used in the experiments of Yang et al (1995) might be responsible for the different results since this domain was intact in the experiments of Namgoong and Harshey (1998). Support for the former scenario would be consistent with a cata-lytic function, while that for the latter would indicate a structural role for domain IIIα. The present study was undertaken to dissect the importance of domain to IIIα to transposition by designing experiments that would distin-guish between the two possibilities.
4.1 No unique MuA subunit contributes domain IIIα function to strand cleavage or strand transfer
The first reported study using the altered specificity variant MuA(R146V) (Namgoong and Harshey 1998) provided convincing evidence that a single DDE+ MuA subunit placed at a particular att site functioned in a specific manner, and thus did not undergo rearrange- ment during assembly on supercoiled substrates. Inter-
pretation of the data in this study is based on this assumption. The results presented in figures 3–5 and table 1 show that a MuA subunit missing domain IIIα can function from L1 and R2 (two of the three ‘core’ att sites L1, R1, R2), without affecting double-end cleavage or strand transfer; domain IIIα function of the R1 subunit is also not required for L1 cleavage. Thus, there appears to be no critical position for the functioning of domain IIIα in transposition. The simplest interpretation of these results is that domain IIIα does not contribute residues to the catalytic pocket of the transposition active site, since if that had been the case, we would have expected such a contribution to be exquisitely position-specific. Several observations suggest that a more plausible function for this domain is to impart structural integrity to the trans-pososome during assembly, cleavage and transition from the cleavage to strand transfer mode. For example, all mutants in this domain are affected in assembly of the MuA tetramer (see figures 3–4; Baker et al 1991; Wu and Chaconas 1995; Krementsova et al 1998; Namgoong et al 1998a). Secondly, singly-cleaved right end complexes containing a domain IIIα deleted subunit fail to mature into strand transfer products (table 1B) and are unstable (our unpublished data). The latter results provide an explanation for why Yang et al (1995) observed cleavage but not strand transfer with a complementing pair of DDE+ IIIα– and DDE– IIIα+ variants on wild-type super-coiled substrates. We have shown in this study that the right end of Mu undergoes preferential cleavage under these conditions (figure 5). That this same complementing pair of MuA variants could strand transfer pre-cleaved linear substrates (Yang et al 1995), is consistent with our finding that doubly cleaved ends can undergo strand trans-fer even when one subunit carries a domain IIIα deletion (table 1D). Domain IIIα−deleted subunits positioned at R1 on pre-cleaved R1-R2 oligonucleotide substrates have been shown to support strand transfer as well (Aldaz et al 1996). Also, pre-cleaved Mu ends are able to rescue MuA assembly defects manifested on uncleaved substrates (Surette et al 1991; Namgoong et al 1994; Kim et al 1995; Wu and Chaconas 1995). The MuB protein used in strand transfer assays from linear substrates also assists in suppression of assembly defects (Namgoong et al 1998a). Although all of these results favour a structural role for domain IIIα, we cannot rigorously exclude the possibility that there is an inherent flexibility in the catalytic contri-bution of the domain IIIα domain, and that alternate modes of protein-DNA associations may become manifest when one subunit in the tetramer is deleted for domain IIIα. Such alternate associations were indeed observed in the contribution of DDE domains on supercoiled vs linear substrates (Namgoong and Harshey 1998). However, in contrast to linear substrates, we imagine that a supercoiled
42
S. MARICONDA ET AL.

substrate must pose constraints on the number of protein-DNA arrangements that can be catalytically fruitful. Another line of evidence against a catalytic role for domain IIIα comes from the experiments of Wu and Chaconas (1995). These authors observed that when the domain IIIα peptide harbouring the non-specific nuclease activity carried multiple mutations in a stretch of basic amino acid residues, it simultaneously lost both DNA binding and nuclease activities; however, when these mutations were transfered to the full length MuA protein, the resultant MuABAN protein was primarily defective in assembly. The mutant could be rescued for assembly and cleavage (at approximately half the efficiency of the wild-type protein), by the addition of either 10% dimethyl sul-foxide (DMSO) or MuB protein (plus ATP). On a pre-cleaved substrate, the strand transfer activities of MuABAN and wild-type MuA were indistinguishable (Wu and Chaconas 1995). These results argue strongly against a catalytic role for domain IIIα in either DNA cleavage or strand transfer under the reaction conditions employed in vitro.
4.2 Active sites and the role of domain IIIα in transposition
The MuA transposase tetramer mediates two strand breakage-joining reactions via hydrolysis followed by transesterification. In principle, four active sites could each carry out one cleavage or one joining reaction. Alternatively, two active sites could sequentially carry out two cleavages followed by two joining reactions. The results obtained in this study, combined with those of Namgoong and Harshey (1998), are most easily accom-modated by the latter scenario. Since only mutants in the DDE residues of MuA have thus far found to be uncondi-tionally defective in transposition (Baker and Luo 1994; Kim et al 1995), the ‘active sites’ for Mu transposition are likely constituted solely of DDE residues. Domain IIIα residues appear not to contribute directly to the catalytic pockets, but rather to the structural integrity/flexibility of the transpososome. The relevance of the nuclease activity associated with domain IIIα is not clear at present, but it appears not to be involved in the chemistry of transposi-tion in vitro, since mutants without this activity can be rescued for transposition under altered reaction conditions (Wu and Chaconas 1995). The recently solved structure of the Tn5 synaptic complex show that DDE residues are likely the sole catalytic determinants of the active site of this transposase (Davies et al 2000). If only two subunits of MuA perform the entire chemis-try of transposition, what is the role of the other two sub-units? It is likely that they contribute structurally or allosterically to the catalytic competence of the tetramer,
since the catalytic prowess of MuA is manifested only within the context of the tetramer. Alternatively, or in addition, these subunits may be involved in initial target binding. While there is no direct evidence for involvement of any particular MuA domain in target recognition, the large positive charge potential of domain IIβ inferred from the crystal struture of domain II (Rice and Mizuuchi 1995), combined with a non-specific DNA-binding acti-vity associated with this domain (Nakayama et al 1987), have led to speculations for a target-binding function for domain IIβ. However, although several mutations in this domain impair intramolecular DNA strand transfer, MuB-assisted intermolecular strand transfer is unaffected (Kre-mentsova et al 1998; Namgoong et al 1998a). The role of MuB is discussed in more detail in the next section. Tn5 transposition is catalyzed by a transposase dimer which, like the Mu reaction, acts in trans (Davies et al 2000). A two-active site model has also been proposed for Tn10 transposition, even though this reaction involves four single-strand breaks (two at each end) and two strand joinings (Bolland and Kleckner 1996). The oligomeric state of the Tn10 transposition apparatus is not yet known. In contrast, there is evidence that in Tn7 transposition, which like Tn10 proceeds by a double-strand cleavage and single-strand joining reactions, four separate active sites perform four DNA cleavage reactions (Sarnovsky et al 1996). It is surmised that the Tn7 transpososome consists of at least two subunits each of two distinct pro-teins, TnsA and TnsB. While TnsA mediates cleavage at the 5′ ends, TnsB mediates cleavage and joining at the 3′ ends. Each of these proteins encodes a DDE motif (Sar-novsky et al 1996). Whether the same subunits of TnsB sequentially mediate the cleavage and joining of 3′ ends, as appears to be the case for MuA, is not as yet known. In summary, domain IIIα of MuA appears to play a role in transpososome assembly, but is unlikely to be involved in catalysis of transposition. While a catalytic role for DDE residues has been demonstrated for two subunits (those at L1 and R1; Namgoong and Harshey 1998) within the MuA tetramer, no specific catalytic function can yet be assigned to the other two subunits. These subunits may play an entirely structural role, might be involved in ini-tial target recognition/binding, or may play a role in post-transposition events such as Mu DNA replication (Levchenko et al 1995; Kruklitis et al 1996).
4.3 Specificity of domain IIIβ in MuB-assisted strand transfer
The MuB protein plays several roles in Mu transposition. It is not only an allosteric effector, stimulating the assem-bly, cleavage and strand transfer activities of MuA, but also captures target DNA and influences target-site selec-
43
S. MARICONDA ET AL.

tion (see Chaconas et al 1996). Residues in domain IIIβ of MuA are essential for interaction with MuB protein, and thus for MuB function. The quaternary structure of the active form(s) of MuB is not known. We have examined the specificity of domain IIIβ func-tion in MuB-assisted intermolecular strand transfer, and found that (i) a single IIIβ domain+ subunit at L1 or R2 is not sufficient, and (ii) two IIIβ domains are sufficient, but their positions are not unique (figure 6 and table 2; we were only able to test diagonal placements). The non-specific nature of the function of domain IIIβ subunits has also been demonstrated in an earlier study (Mizuuchi et al 1995), which did not involve the use of oriented tetramers or determine the minimal number of IIIβ + subunits nece-ssary for function. While the non-unique positioning of IIIβ subunits can be rationalized for the allosteric-effector function of MuB, it is perhaps harder to rationalize for the target-transporter function. In the former case, a confor-mational change in a pair of MuA subunits could con-ceivably be propagated to the other two subunits by protein-protein interactions. In the latter case, the target DNA is likely delivered to specific sites on MuA, since, although MuB binds target DNA and influences target selection, the sequence of the selected sites is the same in the presence of absence of MuB (Mizuuchi and Mizuuchi 1993). How can specific delivery of target DNA be achieved from a non-specific position? Our results would argue that once MuB docks (non-specifically) on the MuA tetramer, it simply serves as an anchor to increase the local concentration of target DNA for specific capture by the catalytical MuA subunits.
Acknowledgements
This work was supported primarily by funds from the National Institutes of Health (GM33247). Partial support was provided by the Robert Welch Foundation (F-1351).
References
Aldaz H, Schuster E and Baker T A 1996 The interwoven architecture of the Mu transposase couples DNA synapsis to catalysis; Cell 85 257–269
Baker T A and Luo L 1994 Identification of residues in the Mu transposase essential for catalysis; Proc. Natl. Acad. Sci. USA 91 6654–6658
Baker T A, Mizuuchi M and Mizuuchi K 1991 MuB protein allosterically activates strand transfer by the transposase of phage Mu; Cell 65 1003–1013
Bolland S and Kleckner N 1996 The three chemical steps of Tn10/IS10 transposition involve repeated utilization of a single active site; Cell 84 223–233
Chaconas G, Lavoie B D and Watson M A 1996 DNA transposition – jumping gene machine, some assembly required; Curr. Biol. 7 817–820
Chen J-W, Lee J and Jayaram M 1992 DNA cleavage in trans by the active site tyrosine during Flp recombination: switch-ing protein partners before exchanging strands; Cell 69 647–658
Craigie R and Mizuuchi K 1987 Transposition of Mu DNA: joining of Mu to target DNA can be uncoupled from cleavage at the ends of Mu; Cell 51 493–501
Davies D R, Goryshin I Y, Reznikoff W S and Rayment I 2000 Three-dimensional structure of the Tn5 synaptic complex transposition intermediate; Science 289 77–85
Grindley N D and Leschziner A E 1995 DNA transposition: from a black box to a colour monitor; Cell 83 1063–1066
Harshey R M and Cuneo S 1986 Carboxyl-terminal mutants of phage Mu transposase; J. Genet. 65 159–174
Jiang H, Yang J-Y and Harshey R M 1999 Criss-crossed inter-actions between the enhancer and the att sites of phage Mu during DNA transposition; EMBO J. 18 3845–3855
Kim K, Namgoong S-Y, Jayaram M and Harshey R M 1995 Step-arrest mutants of phage Mu transposase. Implications in DNA-protein assembly, Mu end cleavage, and strand transfer; J. Biol. Chem. 270 1472–1479
Krementsova E, Giffin M J, Pincus D and Baker T A 1998 Mutational analysis of the Mu transposase. Contributions of two distinct regions of domain II to recombination; J. Biol. Chem. 273 31358–31365
Kruklitis R, Welty D J and Nakai H 1996 ClpX protein of Escherichia coli activates bacteriophage Mu transposase in the strand transfer complex for initiation of Mu DNA syn-thesis; EMBO J. 15 935–944
Kuo C F, Zou A, Jayaram M, Getzoff E and Harshey R M 1991 DNA-protein complexes during attachment-site synapsis in Mu DNA transposition; EMBO J. 10 1585–1591
Lavoie B D and Chaconas G 1995 Transposition of phage Mu DNA; Curr. Top. Microbiol. Immunol. 204 83–102
Leung P and Harshey R M 1991 Two mutations of phage Mu transposase that affect strand transfer or interactions with B protein lie in distinct polypeptide domains; J. Mol. Biol. 219 189–199
Levchenko I, Luo L and Baker T A 1995 Disassembly of the Mu transposase tetramer by the ClpX chaperone; Genes Dev. 9 2399–2408
Levchenko I, Yamauchi M and Baker T A 1997 ClpX and MuB interact with overlapping regions of Mu transposase: impli-cations for control of the transposition pathway; Genes Dev. 11 1561–1572
Li B L, Langer J A, Schwartz B and Pestka S 1989 Creation of phosphorylation sites in proteins: construction of a phospho-rylatable human interferon alpha; Proc. Natl. Acad. Sci. USA 86 558–562
Maxwell A, Craigie R and Mizuuchi K 1987 B protein of bacteriophage Mu is an ATPase that preferentially stimulates intermolecular DNA strand transfer; Proc. Natl. Acad. Sci. USA. 84 699–703
Mizuuchi K 1992 Polynucleotidyl transfer reactions in trans-positional DNA recombination; Annu. Rev. Biochem. 61 1011–1051
Mizuuchi M, Baker T A and Mizuuchi K 1995 Assembly of phage Mu transpososomes: cooperative transitions assisted by protein and DNA scaffolds; Cell 83 375–385
Mizuuchi M and Mizuuchi K 1993 Target site selection in transposition of phage Mu; Cold Spring Harbor Symp. Quant. Biol. 58 515–523
Naigamwalla D Z and Chaconas G 1997 A new set of Mu DNA transposition intermediates: alternate pathways of target capture preceding strand transfer; EMBO J. 16 5227–5234
44
S. MARICONDA ET AL.

Nakayama C, Teplow D B and Harshey R M 1987 Structural domains in phage Mu transposase: identification of the site-specific DNA-binding domain; Proc. Natl. Acad. Sci. USA 84 1809–1813
Namgoong S-Y and Harshey R M 1998 The same two monomers within a MuA tetramer provide the DDE domains for the strand cleavage and strand transfer steps of trans-position; EMBO J. 17 3775–3785
Namgoong S-Y, Jayaram M, Kim K and Harshey R M 1994 DNA-protein cooperativity in the assembly and stabilization of Mu strand transfer complex. Relevance of DNA phasing and att site cleavage; J. Mol. Biol. 238 514–527
Namgoong S-Y, Kim K, Saxena P, Jayaram M, Giedroc D P and Harshey R M 1998a Mutational analysis of domain IIβ of bacteriophage Mu transposase: domains IIα and IIβ belong to different catalytic complementation groups; J. Mol. Biol. 275 221–232
Namgoong S-Y, Sankaralingam S and Harshey R M 1998b Altering the DNA-binding specificity of Mu transposase in vitro; Nucleic Acids Res. 26 3521–3527
Rice P and Mizuuchi K 1995 Structure of the bacteriophage Mu transposase core: a common structural motif for DNA trans-position and retroviral integration; Cell 82 209–220
Sarnovsky R J, May E W and Craig N L 1996 The Tn7 transposase is a heteromeric complex in which DNA breakage and joining activities are distributed between different gene products; EMBO J. 15 6348–6361
Savilahti H and Mizuuchi K 1996 Mu Transpositional Recom-
bination: donor DNA cleavage and strand transfer in trans by the Mu transposase; Cell 85 271–280
Surette M G, Harkness T and Chaconas G 1991 Stimulation of the Mu A protein-mediated strand cleavage reaction by the Mu B protein, and the requirement of DNA nicking for stable type 1 transpososome formation. In vitro transposition char-acteristics of mini-Mu plasmids carrying terminal base pair mutations; J. Biol. Chem. 266 3118–3124
Watson M A and Chaconas G 1996 Three-site synapsis during Mu DNA transposition: A critical intermediate preceding engagement of the active site; Cell 85 435–445
Wu Z and Chaconas G 1994 Characterization of a region in phage Mu transposase that is involved in interaction with the Mu B protein; J. Biol. Chem. 269 28829–28833
Wu Z and Chaconas G 1995 A novel DNA binding and nuclease activity in domain III of Mu transposase: evidence for a catalytic region involved in donor cleavage; EMBO J. 14 3835–3843
Yamauchi M and Baker T A 1998 An ATP-ADP switch in MuB controls progression of the Mu transposition pathway; EMBO J. 17 5509–5518
Yang J-Y, Jayaram M and Harshey R M 1996 Positional information within the Mu transposase tetramer: catalytic contributions of individual monomers; Cell 85 447–455
Yang J-Y, Kim K, Jayaram M and Harshey R M 1995 Domain sharing model for active site assembly within the Mu a tetramer during transposition: the enhancer may specify domain contributions; EMBO. J. 14 2374–2384
45
S. MARICONDA ET AL.


















