
T cell responses are initiated in the T cell areas of secondary lymphoid organs CD4+ Th1 CD4+ Th2...
24
T cell responses are initiated in the T cell areas of secondary lymphoid organs CD4+ Th1 CD4+ Th2 CD8+ Cytotoxic Memory T cells Effector T cells Proliferation and Differentiation r the last 25 years, dendritic cells (DC) have emer as the major APC involved in this process
-
Upload
meagan-flowers -
Category
Documents
-
view
221 -
download
2
Transcript of T cell responses are initiated in the T cell areas of secondary lymphoid organs CD4+ Th1 CD4+ Th2...

T cell responses are initiated in the T cell areas of secondary lymphoid organs
CD4+ Th1
CD4+Th2
CD8+Cytotoxic
Memory T cells
Effector T cells
Proliferation and Differentiation
Over the last 25 years, dendritic cells (DC) have emerged as the major APC involved in this process

T cell and APC priming
•Origin and heterogeneity of DCs•Maturation of DCs•T cell priming by DCs•Role of DCs in Tolerance induction

DCs progenitors are generated in the bone marrow. They give rise to circulating DCs precursors. Circulating DCs precursors enter nonlymphoid tissues as immature DCs. DC are scattered throughout all non lymphoid tissues where they reside in a resting, (so-called immature) state. In the absence of ongoing inflammatory and immune responses, they constantly migrate at low rate to draining lymph nodes.
DCs precursors
Immature DCs
DCs progenitors
Mature DCs
In inflammatory conditions, immature DC migrate to draining lymph nodes where after maturation (mature DC), they prime the rare circulating naïve antigen-specific lymphocytes.
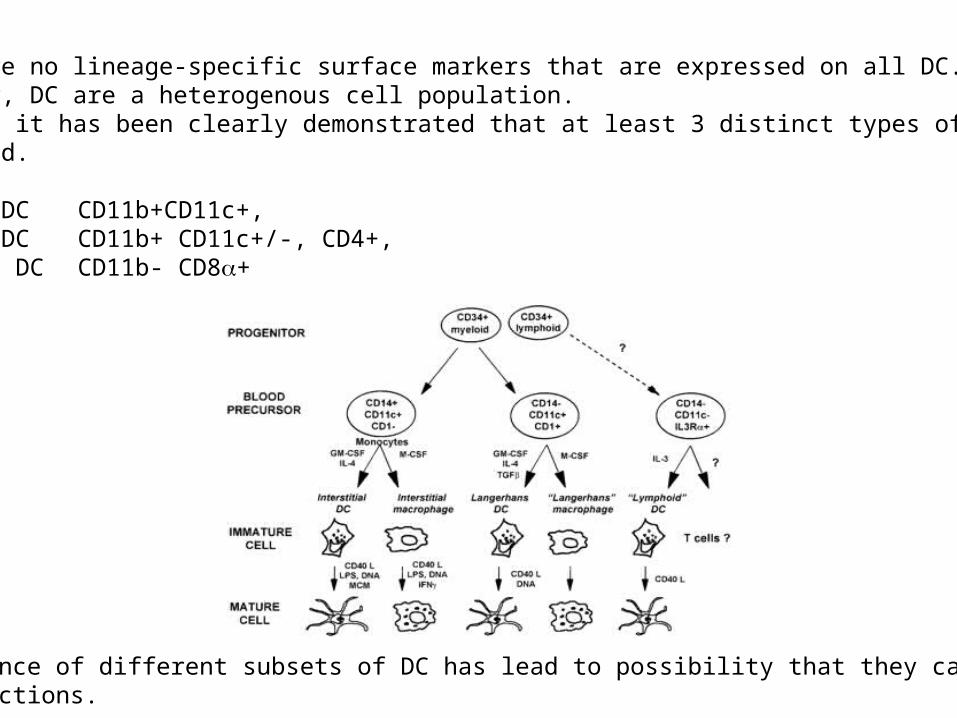
There are no lineage-specific surface markers that are expressed on all DC.Moreover, DC are a heterogenous cell population.However, it has been clearly demonstrated that at least 3 distinct types of DC can begenerated.Mice: myeloid DC CD11b+CD11c+, myeloid DC CD11b+ CD11c+/-, CD4+, lymphoid DC CD11b- CD8+
The existence of different subsets of DC has lead to possibility that they can performunique functions.

Feature of Immature DC =Antigen-capturing cell
Immature DC are very efficient in Ag uptake andprocessing.Antigens are taken up by receptor-mediated endocytosis(FcR, C-type lectin receptors,scavenger receptors etc) orby macropinocytosis and phagocytosis.MHC class II-rich compartments are abundant. These late endosomal structures contain theHLA-DM (H-2M in mice) products which enhance and edit peptide binding to MHC class II molecules.Secrete proteases that may degrade external Aginto peptides that can be loaded directly onto empty MHC class II on the surface of the DC.
Figure 1: Three pathways for loading major histocompatibility complex (MHC) class II molecules with peptides in dendritic cells. a, b, Antigen is captured by an antigen receptor, and together antigen and antigen receptor are internalized. In MHC-class-II-positive compartments, the antigen is processed, generating peptides, some of which will be presented on the surface of the dendritic cell in complex with an MHC class II molecule. a, Newly synthesized MHC class II molecules are transported from their site of synthesis through the Golgi into the class-II-positive compartments, where they meet their peptide cargo. b, Class II molecules may be recycled from the cell surface into the correct compartments, where again they are loaded with peptide. c, extracellular proteases may degrade external antigens, generating peptides that can be loaded directly onto empty MHC class II molecules on the surface of the dendritic cell.
Davoust J.and Banchereau J. Naked antigen-presenting molecules on dendritic cellsNature Cell Biology 2:E46-E48, 2000.

Exogenous Ag leads exclusively to presentation on MHC class II molecules.
How class I-restricted immune responses are generated by DC which take up exogenous Ags?
Exogenous Ags may also be presented by MHC class I molecules, a phenomenon referred to as cross-presentation.Presentation of endogenous Ags by MHC class I molecules:Ubiquitinated proteins, proteosome degradation, resulting peptides are transported to the ER by TAP and loaded on new MHC class I molecules, Golgi, plasma membrane.
Two main intracellular pathways for cross-presentation have been reported:• TAP-independent: requires the presence of MHC class I molecules in the endocytic pathway. Low endosomal ph allows peptides generated in endosomes to exchange with endogenous peptidesbound to recycling MHC class I molecules.• TAP-dependent: This pathway requires transport of internalized Ags from endocytic compartments to the cytosol. DCs have a unique endosome-to-cytosol transport pathway thatallows selective delivery of internalized Ags to the cytosol.

The Ag/pathogen induces the immature DC to undergo pheno-typic and functional changes that culminate in the complete transition from Ag-capturing cell to an APC (maturation).
What are the stimuli that activate immmature DCs?There are different views as to the nature of DC-activating stimuli.
These views need not be mutually exclusive
In 1989, Janaway postulated that: APCs possess receptors Pattern Recognition Receptors (PRRs) that recognize and are triggered by evolutionarily conserved moleculesexpressed by pathogens that are not found in higher eukaryotes. These molecules have essential roles in the biology of the invading agent. These molecules are called Pathogen Associated Molecular Patterns (PAMPs).

Reis e Sousa, C. Dendritic cells as sensors of infectionImmunity 14:495-498, 2001.

A natural extension of Janaway’s hypothesis is that DCs might be activated by signalsmade by other cells in response to PAMPs. Many tisssues may have their own PRR andproduce proinflammatory cytokines in response to infection.Examples: •Virally infected cells produce interferons in response to dsRNA. Interferonsact to activate DCs.• Keratinocytes can secrete TNF-a, IL-1 and IL-18 all of which activate Langerhans cellsa type of skin and mucosal DC. DCs express receptors for inflammatory cytokines (TNF-a,IFNs and IL-1.• In response to Taxoplasma gondii, endothelial cells produce chemokines that canactivate DCs.
The Danger HypothesisMatzinger, has proposed that DC activation does not need to involve PRRs, but insteadreceptors for self-molecules normally sequestered in intra-cellular compartments of healthy cells but are released when cells are stressed or die by necrosis (danger).Examples:•Mammalian heat-shock proteins (HSPs) can activate DCs and are released from necroticbut not from apoptotic cells. DCs express CD91 which binds to HSPs.•Other intracellular components shown to activate DCs include nucleotides (ATP)

Reis e Sousa, C. Dendritic cells as sensors of infectionImmunity 14:495-498, 2001.

Phenotypic and functional changes that occur during thematuration process of DCs.
• Antigen PresentationWhile maturing, DCs shut off antigen capture and class II synthesis. The MHC complexesaccumulate on the cell surface and acquire extremely long half-lives ( 100 hours).The peptide-MHC II complexes are retained for several days maximizing presentation of those Ags associated with the infectious organism that triggered the maturation process.
Hartgers F.C., Figdor C.G. and Adema G.J. Towards a molecular understanding of dendritic cell immunobiologyImmunol. Today 21:542-544, 2000.

• MigrationMaturing DCs migrate into the T cell area of lymphoid organs.
Immature Dcs express a variety of chemokine receptors (CCR1, CCR5, CXCR1 and CCR6)that participate in their recruitment to inflammed tissue and/or to allow their residency into non-lymphoid tissue.Exemple: Imm. Dcs express CCR6 the receptor for MIP-3a that is constitutively expressin liver and lungs. Upon maturation, there is a downregulation of receptors for chemokines produced at thesite of inflammation and upregulation of CCR7. SLC is a ligand for CCR7 and is expressedat high levels by HEVs in LNs and by stromal cells in T cell areas of many secondary lymphoid organs. ELC (other ligand of CCR7) made in T cell areas of lymphoid tissue.
SLC (6Ckine) and ELC (MIP-3b) act together to direct DC migration to T cell areas Of lymphoid tissue and to promote encounter with T cells.Adhesion molecules
Banchereau J. et al. Immunobiology of dendritic cellsAnn. Rev. Immunol. 18:767-811, 2000.

• Induction of costimulatory molecules
TCRMHC
CD4 or CD8
1
2
DC T cell
COSTIMULATION
Mature DCs can initiate primary T cell-mediated immuneresponses because they express high amounts of MHC complexes(signal 1) and costimulatory molecules (signal 2).

• Induction of survival molecules
Maturation stimuli upregulate the expression of molecules that can transduce survival signals.CD40 and RANK are ligands express by DCs that recognize, respectively, CD40-L and TRANCEexpressed on T cells. Mature DCs have a finite life expectancy, which affects their ability to stimulate T cells. There is evidence that once they have reach the LN, DCs survive for only a few days. It has been shown that activated T cells, expressing CD40-L and TRANCE, can trigger CD40 and RANK on DCs, thereby enhancing their viability.Thus, feedback signals from T cells can also provide signals that reinforce the maturation process.
Lanzavecchia A. and Sallusto F. Dynamics of T lymphocyte responses: intermediates, effectors, and memory cellsScience 290:92-97, 2000.

The encounter with an infectious organism requires not only that animmune response be initiated but also that it be of the appropriate class.
Examples:•Th2-dominated responses play a role in immunity to helminths
•Th1 responses are critical to eliminate many intracellular pathogenssuch as Listeria or Toxoplasma gondii.
Thus it is essential that the APC both alert T cells to the presence of infection and alsotransmit information about the nature of the infectious organism.Much of this information is transmitted by cytokines (signal 3).

The first experimental evidence that DCs have the capacity to induce different types of T cell-mediated responses came from the observationthat DCs are the most relevant source of the Th1-polarizing cytokineIL-12.If all DCs produce IL-12, how could a Th-2 response ever be induced?
Distinct DC subsets elicit distinct Th responses:In mice:Lymphoid DCs Th1 (DC1) IL-12 +++++Myeloid DCs Th2 (DC2) IL-12 +/-
In humans:Lymphoid DCs Th2 (DC2) IL-12 +/-Myeloid DCs Th1 (DC1) IL-12 +++++Although human and mice are opposites in terms of the functionalclassification of DC subsets, studies showed the existence of a highIL-12-producing subset that induces Th1 responses and a low
IL-12-producing subset that induces Th2 responses.

The realization that distinct subsets of DCs can regulate immuneresponses differentially has been accompanied by the opposingrevelation that a single type of DC can make different cytokines depending on the microenvironment in which the DC has been activated or the pathogen.Examples:•DC1 stimulated in vitro in the presence of IL-10, TGF or PGE2can induce Th2 differentiation.• At the yeast stage, Candida albicans stimulates DCs to produce IL-12 and induce Th1 responses. At the hyphea stage, C. albicansstimulate DC to produce IL-4 and induce Th2 responses.
Thus, the type of T cell-response induced by DCs depends on the nature of the DC-activating stimulus, and not so much on their ontogeny.or,by modulating the production of IL-12 by DCs, different maturationstimuli play an instructive role for Th1 or Th2 responses.
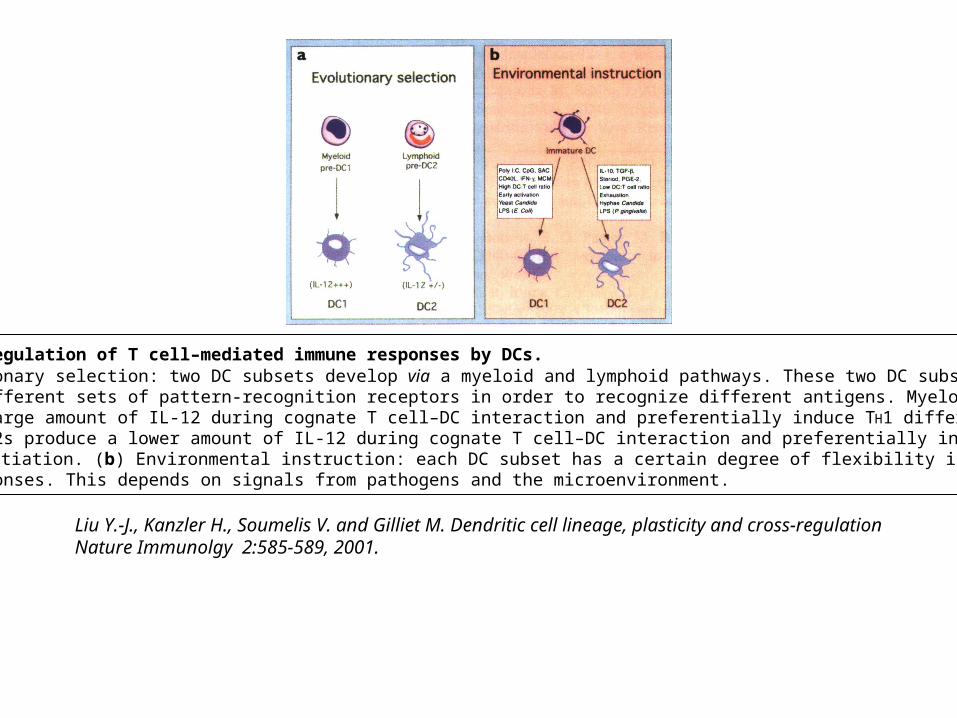
Liu Y.-J., Kanzler H., Soumelis V. and Gilliet M. Dendritic cell lineage, plasticity and cross-regulationNature Immunolgy 2:585-589, 2001.
Figure 1. Regulation of T cell–mediated immune responses by DCs. (a) Evolutionary selection: two DC subsets develop via a myeloid and lymphoid pathways. These two DC subsets express different sets of pattern-recognition receptors in order to recognize different antigens. Myeloid DC1s produce a large amount of IL-12 during cognate T cell–DC interaction and preferentially induce TH1 differentiation. Lymphoid DC2s produce a lower amount of IL-12 during cognate T cell–DC interaction and preferentially induce TH2 differentiation. (b) Environmental instruction: each DC subset has a certain degree of flexibility in directingT cell responses. This depends on signals from pathogens and the microenvironment.

The role of DCs in the induction of peripheral self-tolerance
Situation: During an infection there is extensive death of infectedcells. DCs are well-known to engulf dying cells and they should processand present self-antigens. How do DCs focus immunity on the infectious organism if they also present self-antigens?How come that T cells reactive to self-antigens are not activated duringan infection?

Experimental evidences that peripheral tolerance to tissue Ags involved bone-marrowderived cells in draining lymph nodes :• Influenza hemagglutinin (HA) expressed in many tissues as self-Ag. Injected TCR transgenic HA-specific CD4+T cells are anergized. Induction of anergy required bone-marrow derived cells that express the appropriate MHC. Anergy did not develop if only nonhematopoietic cells expressed theappropriate MHC. (Adler et al. J. Exp. Med. 187:1555, 1998)• OVA expressed in pancreatic islets. OVA is presented by bone-marrow derived cells and TCR transgenic OVA-specific CD8+T cells were tolerized by deletion. They showed that the tolerizing, bone marrow derived cellswere confined to the draining lymph nodes. (Kurts et al. J. Exp. Med. 184:923, 1996 and J. Exp. Med. 186:239, 1997).Somehow, self-Ags in peripheral tissues are transferred to bone-marrow derived cells in the draining LN and this tolerizes adult T cells.• DCs in afferent lymph carry apoptotic bodies derived from the intestinalepithelium. (Huang et al. J. Exp. Med.191:423, 2000)
DCs pick up apoptotic material from cells undergoing the turnovercharacteristic of most tissues and continually deliver samples of tissuesto the draining LN.

• DCs that phagocytose apoptotic cells are not induced to mature intostrong stimulator of T cell immunity.In contrast to DCs that phagocytosecells that are dying by necrosis.Sauter et al. J. Exp. Med.191:423, 2000.
• DCs only take up apoptotic cells when immature (Inaba et al. J. Exp. Med. 188:2163, 1998; Albert et al. J. Exp. Med. 188:1359, 1998)
• Apoptotic cells do not mature the DCs(Sauter et al. J. Exp. Med.191:423, 2000)
• Immature DCs are poor T cell stimulators(Schuler et al.J. Exp. Med.161:526, 1985.
Then, what is the immunological consequence of the presentation ofself-antigens derived from the processing of a large number of dying somatic cells in the LN?Null (only clearance) or induction of Self-tolerance?

Null ?
We know that bone-marrow derived cells within the LN can tolerize T cells to peptides (neo-self-Ags) synthesized inother tisues.
Induction of self-tolerance?
• Immature DCs do not express abundantly peptide-MHC complexeson their surface. Self-reactive T cells would not be able to recognizetheir ligand on immature DCs.
• Immature DCs lack CD40 and TRANCE-R that sustain viability for the 3-4 days needed before T cell tolerance becomes evident.

Hypothesis:Short-lived migratory DCs are processed and presented by LN resident DCs (lymphoid DCs). These DCs express high levelsof peptide MHC complexes and low levels of costimulatorymolecules.LDCs Anergy, deletion or regulatory T cells (Treg) that suppress the activation of self-reactive T cells.
Steinman R.M., Turley S., Mellman I., and Inaba K. The induction of tolerance by dendritic cells that have captured apoptotic cells. J. Exp. Med. 191:411-416, 2000.

SUMMARY
Lanzavecchia A. and Sallusto F. From synapses to immunological memory: the role of sustained T cell stimulationCurr. Opin. in Immunol. 12:92-98, 2000.


















