
SYNAPTIC TRANSMISSION IINeurosceince 500 2009 Dr. Tim Murphy The chemistry of neurotransmitter...
54
SYNAPTIC TRANSMISSION II Neurosceince 500 2009 Dr. Tim Murphy The chemistry of neurotransmitter release. The early studies of Katz and others suggested that neurotransmitter release was quantized. Electron microscopy shows that nerve terminals contain 40-200 nm vesicles that were proposed to contain the transmitter packets or quanta. Evidence for vesicular release is from Douglas' experiment which showed that substances found in vesicles - chromogranins are co-released with norepinephrine. We will focus on: 1) storage and synthesis of neurotransmitters 2) production and recycling of synaptic vesicles 3) study and identification of synaptic vesicle proteins 4) molecular schemes for neurotransmitter release 5) modulation of neurotransmitter release in
-
Upload
della-crawford -
Category
Documents
-
view
219 -
download
2
Transcript of SYNAPTIC TRANSMISSION IINeurosceince 500 2009 Dr. Tim Murphy The chemistry of neurotransmitter...

SYNAPTIC TRANSMISSION II Neurosceince 500 2009Dr. Tim Murphy
The chemistry of neurotransmitter release.
The early studies of Katz and others suggested that neurotransmitter release was quantized. Electron microscopy shows that nerve terminals contain 40-200 nm vesicles that were proposed to contain the transmitter packets or quanta. Evidence for vesicular release is from Douglas' experiment which showed that substances found in vesicles - chromogranins are co-released with norepinephrine. We will focus on:1) storage and synthesis of neurotransmitters2) production and recycling of synaptic vesicles3) study and identification of synaptic vesicle proteins4) molecular schemes for neurotransmitter release5) modulation of neurotransmitter release in synaptic plasticity

Why store transmitters in vesicles?
• Concentration- (mediated by binding proteins and by H+ powered vesicular transporters)
• Protection from degradation - by proteases and esterases
• Allows for regulation• Provides a storage system

Most transmitters are simple molecules, derivations of amino acids or choline, packaged locally and found in 40-50 nm vesicles.
Aspartate-Glutamate -Glycine -GABA - glutamateSerotonin - tryptophanHistamine - histidineDopamine - tyrosineNorepinephrine - tyrosineAcetylcholine – choline
Neuropeptides Somatostatin, Enkephalin, etc are found in larger >100 nm dense core vesicles (DCVs). Made on ribosomes in the somatic region. Targeted to the nerve terminal for calcium
evoked exocytosis. http://synapses.mcg.edu/atlas/1_6_9.stm
DCV

Synaptic transmission, a specialized form of normal exocytosis.
• Issues with vesicle targeting, need for specificity and speed.
• Specific proteins on vesicles and targets guide the process.
• Molecular biology reveals homology in components from yeast to man.

Synaptic transmission is an adaptation of normal vesicle trafficking.

Cells are polar and need to traffic cargoesasymmetrically.
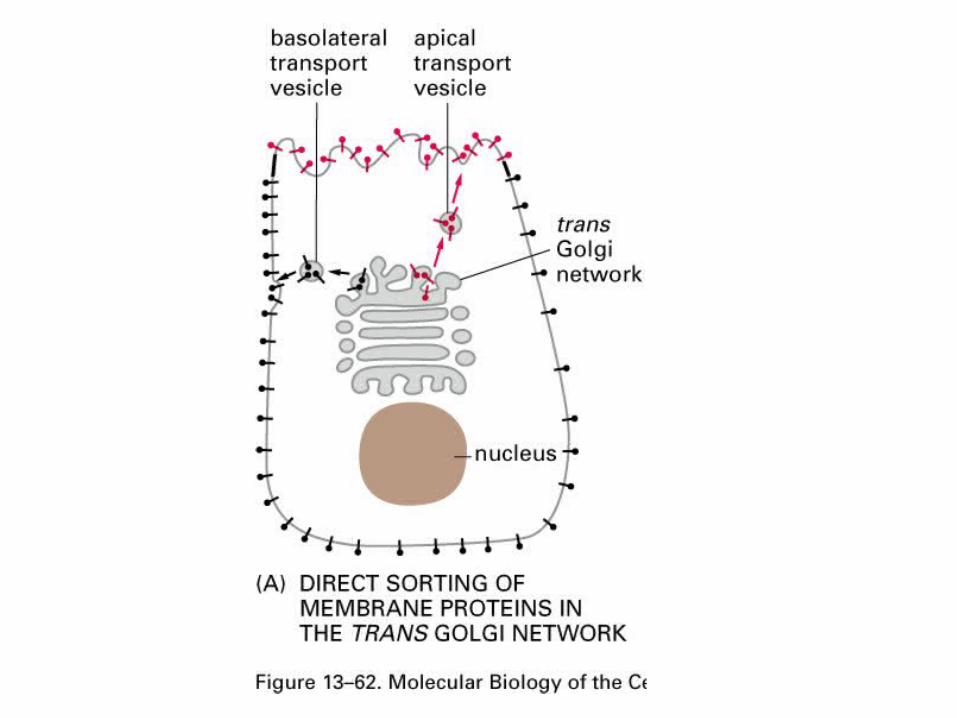

The synapse has adapted forms of vesicle cycling found in all cells.

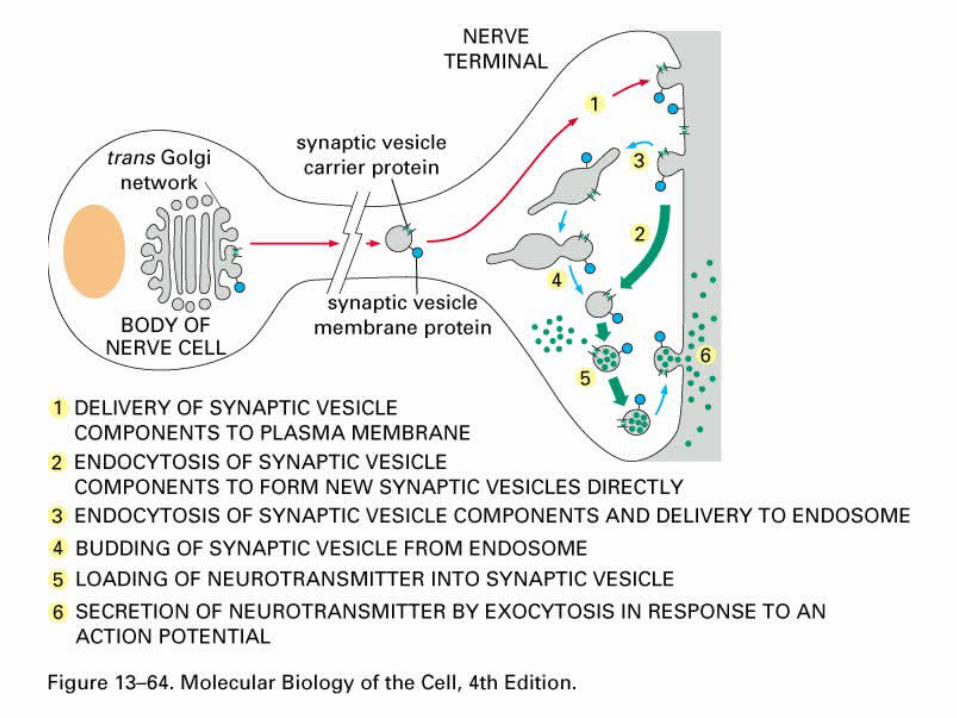
Production and recycling of synaptic vesicles.
• Synaptic vesicle components are made in the soma and move through the golgi, transported to the release sites by fast axonal transport (which ones?).
• Since vesicles are released by exocytosis need to be recycled or nerve the terminal would grow quite large, also need to conserve synaptic vesicle proteins.
• Recycling of synaptic vesicles - HRP in extracellular media (an enzyme that can give staining reaction) gets engulfed by endocytosis, visualized with EM and shown to go to: coated vesicles – clathrin endosome-sorting area then either to lysosome (destruction) or back into vesicle pool.

Markers of synaptic vesicle turnover
• HRP enzyme (horse radish peroxidase), tracking by stain viewed on EM.
• Membrane stains, FM-143 dyes.• Capacitance measurements real-time.• pH sensitive GFPs, linked to synaptobrevin
synaptophlurin, fluor quenched in vesicle by H+.• Antibodies to lumenal domain of synaptic vesicle
proteins.• Quantum dots (Zhang et al. 2009 Science).
See Harata et al. 2006 J. Neurochem. for Review of all methods except Q-dots.

Modes of release and recycling.
• In neurons Heuser and Miller originally propose two pathways, coated pits and cisternae (endosome) for most synaptic transmission and non-coated pit pathways for times of high activity.
• However the situation in neurons may be more complicated and can include transient forms of fusion such as “kiss and run”.


T. Ryan PNAS 2002

Evidence for 2 modes of release, from ultrastructure snap-shots and capacitance changes (surface area).
Harata et al. 2006 J. Neurochem.
Frog neuromusc. quick-freeze EM Mast cell de-granulation capacitance.



SNARE Hypothesis• Synaptic vesicles have a specific protein that directs
them to a receptor on the plasma membrane (with the aid of other proteins SNAPs etc.).
• The vesicle associated SNAP receptor (v-SNARE) in neurons is believed to be synaptobrevin (VAMP).
• Target sites such as the plasma membrane (nerve terminal) would have a corresponding t-SNARE (syntaxin and SNAP-25).
• At sites other than the nerve terminal specific vesicle and target SNAREs would function to target vesicles to specific compartments.

Coiled-coil interactions.

A B
C D
Yeast
Yeast

Generic fusion mechanisminvolves a tightening of the SNAREs to push out H20.
Hey what about Ca2+?


Study and identification of synaptic vesicle proteins.
• Vesicles isolated by differential centrifugation - reach an equilibrium between buoyant force of vesicle and centrifugal force in a gradient.
• Study of synaptic vesicle proteins has helped delineate the players in neurotransmitter release.
• Sequence analysis indicates evolutionary similarities between yeast exocytosis and neurotransmitter release. Use of yeast temperature sensitive mutants for secretion (sec mutants). Shown that neuronal proteins can complement lost or defective yeast genes.


Synapsins (Ia, Ib, IIa, IIb) Vesicle associated proteins.
• Hypothesis: the binding of synapsin to synaptic vesicle proteins is regulated by phosphorylation, providing a connection between second messenger activated protein kinases and the modulation of release by regulation of vesicle availability.
• Candidate for regulating molecule in the release of vesicles from the filamentous actin/spectrin network they are stored in. Synapsin binds to cytoskeletal proteins including spectrin and actin.
• Binding of synapsin is weakened by phosphorylation - injecting dephosphosynapsin blocks neurotransmitter release while CaMK (active) increases it (Llinas experiment).

Synaptotagmin (P65) Ca2+ binding.• Single polypeptide, one transmembrane spanning region and large
cytoplasmic domain (Syt1,2 vesicle membrane, Syt3,6,7 plasma membrane).
• Protein kinase C like homology in carboxyl terminal (C2 domain), involved in calcium and phospholipid binding. Injection of C2 peptide can block release after the vesicle is docked (squid; Augustine lab).
• Forms tetramers with each binding calcium (possible) cooperatively consistent with 3-4 power of Ca dependence of release (Sudhof and others).
• In vitro studies it binds Ca cooperatively at concentrations in the physiological range (10-100 M) for release.
• In the absence of Ca2+ synaptotagmin may serve as a brake for release. Ca2+ removes the brake. May explain how release in nonneuronal cells occurs without synaptotagmin.

G. Augustine Curr Op. in Neurobiol.
Oligomerization of synaptotagmin and itsbinding to synapticprotein partners occursover similar Ca2+ concentrationrange as neurotransmitterrelease.

Copyright ©2003 Cell Press.Cell, Vol 112, 519-533, 21 February 2003
ReviewMembrane FusionReinhard Jahn 1, Thorsten Lang 1, and Thomas C. Südhof 2
Synaptotagmins (Syt 1,2,3,6,7) bind Ca and phospholipids possibly promoting membrane fusion.

Fernandez-Chacon et al. Nature 2001
Critical experiment was to the change affinity of Ca for synaptotagmin in vivo

R233Q
Fernandez-Chacon et al. Nature 2001
Knock in mutation of synaptotagmin with lower Ca sensitivity.

Knock in mutation of synaptotagmin with lower Ca sensitivityresults in lower release probability and facilitating responses.
Fernandez-Chacon et al. Nature 2001
*Note the reduction in wild-type current amplitude is not a consistent finding.
*

Knock in mutation of synaptotagmin with lower Ca sensitivityreduces average response amplitude but not RRP size.
Fernandez-Chacon et al. Nature 2001

Synaptophysin
• Abundant synaptic vesicle protein of unknown function.

Synaptobrevin (VAMP-1 and 2, vesicle associated membrane protein).
• Functions as a vesicle SNARE.• Role in release is aided by use of toxins. Tetanus
toxin and botulinum B,D,F, and G (have protease activity) cleave synaptobrevin selectively, and block release.
• Homologous to yeast proteins BOS1 and SNC1, which have been shown to complement yeast which are mutants in vesicular transport.

Soluble proteins.• NSF - N-ethylmaleimide sensitive factor: isolated
based on its ability to restore function to a N-ethylmaleimide treated golgi prep. NSF has ATP hydrolysis activity and is associated with breaking down SNARE complexes after exocytosis.
• SNAPs-soluble NSF attachment proteins. • Nsec-1/munc-18 and munc 13, syntaxin binding
proteins likely bound to syntaxin when it is not in the SNARE complex, involved in priming mechanism role unclear.

ATP is not actually required for release once vesicles are docked, but is thought to break down the SNARE complexes to promoterecycling.

Plasma membrane proteins.
• Syntaxin putative t-SNARE, binds to N and P/Q-type calcium channels (Sheng et al. 1994) and SNAP-25 and synaptotagmin cleaved by botulinum toxin C1. Critical for localizing Ca channel near release site.
• SNAP-25 peripheral membrane protein binds to syntaxin, cleaved by botulinum toxin A and E. Forms a SNARE complex with syntaxin and synaptobrevin.

Rizo and Sudhof 2002 Nature Rev. Neurosci.

Rizo and Sudhof 2002 Nature Rev. Neurosci.

Release animation.
• T. Galli, V. Haucke, N. R. Gough, Synaptic vesicle fusion followed by clathrin-mediated endocytosis. Sci. STKE 2003, tr3 (2003) shockwave animation of ves. fusion.

Mechanisms of short-term presynaptic plasticity
• Two major mechanisms: increasing the action potential evoked calcium transient or basal calcium levels (residual calcium hypothesis-Zucker).
• Elevated basal calcium concentration leads to increased release probability (a high affinity effect mediated by nM levels of calcium).

Modulation of neurotransmitter release in synaptic plasticity
• Presynaptic modulation of transmission by effecting release machinery or calcium influx. Simplest mechanism is to modulate presynaptic calcium current.
• Mechanism for post-tetanic potentiation may involve a saturation of calcium buffer or mitochondrial uptake systems resulting in higher levels of intracellular calcium and more release (residual calcium theory). Longer time course of modulation (several minutes) may involve calcium dependent or independent vesicle mobilization steps.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
A single presynaptic Neuron can have varied transmitter release at its different synapses.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
A single postsynaptic Neuron can have varied transmitter release at its different inputs.
2 nA

Steven’s lab (Schikorski et al.).

Presynaptic regulation.
Atwood and Karunanithi2002 Nature Rev. Neurosci.
Typically a small number of active zone for CNS synapses likely less than 5, while could be hundreds for the neuromuscular junction.
(quantal content, number of vesicles released per AP)

Atwood and Karunanithi2002 Nature Rev. Neurosci.
Presynaptic regulation.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
Presynaptic regulation.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
Presynaptic regulation.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
Presynaptic regulation.

Postsynaptic regulation.
Atwood and Karunanithi2002 Nature Rev. Neurosci.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
The number of receptors will alsoPlay a role.

Atwood and Karunanithi2002 Nature Rev. Neurosci.
Kiss and run, versus full fusion of vesicles.

Readings• Fundamental Neuroscience 1st Ed. Chapters 7 and 8 (for
Neurochem also), 2nd Ed. Chapters 7 and 8., 3rd Ed. Chapters 7 and 8.
• Molecular Biol. Of the Cell 4th Ed. Chapter 13.• Harold L. Atwood & Shanker Karunanithi Diversification of
synaptic strength: presynaptic elements. Nature Reviews Neuroscience 3, 497 -516 (2002). Advanced review comprehensive.
• T. Galli, V. Haucke, N. R. Gough, Synaptic vesicle fusion followed by clathrin-mediated endocytosis. Sci. STKE 2003, tr3 (2003) shockwave animation of ves. fusion.
• T. Sudhof The synaptic vesicle cycle. Annu. Rev. 2004 Vol. 27: 509-547


















