
Superoxide Dismutase - Journal of Biological Chemistry
12
THE JOURNAL or BIOLOGICAL CHEMISTRY Vol. 248, No. 10, Issue of 1IIay 25, pp. 3582-3592, 1973 Printed in U.S.A. Superoxide Dismutase ORGANELLE SPECIFICITY-* (Received for publication, October 12, 1972) RICH-~RD A. WEISIGER AND IRWIX FRIDOVICH From the Department of Biochemistry, Duke LTnivedy Medical Center, Durham, North Carolina 27710 SUMMARY Fresh chicken liver contains two types of superoxide dis- mutase, one of which is localized in the mitochondria while the other is found in the cytosol. The mitochondrial super- oxide dismutase was inactivated by treatment with mix- tures of chloroform and ethanol whereas the cytosol super- oxide dismutase was not. The cytosol superoxide dismutase was inhibited by cyanide, whereas the mitochondrial en- zyme was not. The cytosol superoxide dismutase was puri- fied to homogeneity and was found to contain copper and zinc and to be similar to the other cupro-zinc superoxide dismu- tases which have been isolated from diverse eukaryotes. In contrast, the superoxide dismutase present in chicken liver mitochondria, when isolated, was found to a manganoprotein which has a molecular weight of 80,000. It is composed of four subunits of equal size, which are not covalently joined. It contains 2.3 atoms of manganese per molecule and is strikingly similar to the superoxide dismutase previously isolated from bacteria (KEELE, B. B., JR., MCCORD, J. M., AND FRIDOVICH, I. (1970) J. Biol. Chem. 24.5, 6176-6181; VANCE, P. G., KEELE, B. B., JR., AND RAJAGOPALAN, K. V. (1972) J. Biol. Chem. 247, 47824786). Avimanganin which was isolated from chicken liver mitochondria (SCRUTTON, M. C. (1971) Biochemistry 10, 3897-3905) appears to be an in- active form of mitochondrial superoxide dismutase. The similarity between the mitochondrial superoxide dismutase and the corresponding enzymes from Escherichia coli and from Streptococcus mutans and the marked dissimilarity to the superoxide dismutase found in the cytosols of eukaryotes lends support to the theory that mitochondria have evolved from aerobic prokaryotes, which entered into an endocellular symbiosis with a protoeukaryote. Chicken liver was not unique in possessing distinct superoxide dismutases in cytosol and in mitochondrial compartments, since the same situation was shown to occur in pig heart. The chicken liver cytosol superoxide dismutase exhibited two peculiar properties. Thus when stored in deep, con- centrated solutions it underwent autoreduction to a yellow cuprous form which retained enzymatic activity, and it po- lymerized to forms which likewise retained activity. Both of these changes were reversible and appeared to be due to reactive sulfhydryl groups which could act as the endogenous reductant and could form intermolecular disulfide bridges. * This work was supported in part by Research Grant GM-10287 from the Xational Institutes of Health, Bethesda, Maryland. The cytosol superoxide dismutase consisted of a family of electrophoretically distinct forms which could readily be separated on a preparative scale by isoelectric focusing. These multiple forms were indistinguishable in size and in specific activity. The chemical basis of these dilIerences in charge was not explored. Superoxide dismutase catalyzes the reaction 02 + 0, + 2IIf + Hz02 + OS (4, 9-12) and appears to be ubiquitous among oxygen-metabolizing organisms and to be lacking in obligate anaerobes (13). This enzyme has been isolated from a wide range of eukaryotic sources including bovine erythrocytes and heart (4, 5), garden peas (6), Xaccharomyces cerevisiae (8), and Arezlrospora crassa (7) and in all of these was found to be a blue- green cupro-zinc protein. In contrast, the superoxide dismutase isolated from the prokaryotes Escherichia coli (1) and Slrepto- coccus mutans (2) was a pinkish manganoprotein. These two types of superoxide dismutase appeared to be completely unre- lated proteins which had evolved independently. During an investigation of the superoxide dismutase of chicken liver, it was observed that the enzyme from mitochondria n-as distinct from that present in the cytosol with respect to several properties including electrophoretic mobility, susceptibility to inhibition by cyanide, and to inactivation by mixtures of chloro- form plus ethanol. Since the cupro-zinc superoxide dismutases were known to be inhibited by cyanide whereas the mangano- superoxide dismutases were not (14), it appeared possible that the mitochondria of chicken liver contained the prokaiyotic type of superoside dismutase, whereas the cytosol contained the eukaryotic type. Previous isolations of the superoxide dismu- tases of eukaryotes had used a simple but rigorous procedure (4) which included treatment with the chloroform-ethanol mixture of Tsuchihashi (15). This treatment inactivates the prokaryotic type of superoxide dismutase so that, even had it been present in the mitochondria and therefore in the extracts prepared from the diverse eukaryotic sources (5, 6, 8)) it would have been inac- tivated and would have gone undetected. The purification and characterization of the superoxide dismutases present in the cytosol and in the mitochondria of chicken liver was therefore undertaken. This paper describes the discovery of the differ- ences between the mitochondrial and the rytosol superoxide dis- 3582 by guest on April 11, 2019 http://www.jbc.org/ Downloaded from
Transcript of Superoxide Dismutase - Journal of Biological Chemistry

THE JOURNAL or BIOLOGICAL CHEMISTRY Vol. 248, No. 10, Issue of 1IIay 25, pp. 3582-3592, 1973
Printed in U.S.A.
Superoxide Dismutase
ORGANELLE SPECIFICITY-*
(Received for publication, October 12, 1972)
RICH-~RD A. WEISIGER AND IRWIX FRIDOVICH
From the Department of Biochemistry, Duke LTnivedy Medical Center, Durham, North Carolina 27710
SUMMARY
Fresh chicken liver contains two types of superoxide dis- mutase, one of which is localized in the mitochondria while the other is found in the cytosol. The mitochondrial super- oxide dismutase was inactivated by treatment with mix- tures of chloroform and ethanol whereas the cytosol super- oxide dismutase was not. The cytosol superoxide dismutase was inhibited by cyanide, whereas the mitochondrial en- zyme was not. The cytosol superoxide dismutase was puri- fied to homogeneity and was found to contain copper and zinc and to be similar to the other cupro-zinc superoxide dismu- tases which have been isolated from diverse eukaryotes. In contrast, the superoxide dismutase present in chicken liver mitochondria, when isolated, was found to a manganoprotein which has a molecular weight of 80,000. It is composed of four subunits of equal size, which are not covalently joined. It contains 2.3 atoms of manganese per molecule and is strikingly similar to the superoxide dismutase previously isolated from bacteria (KEELE, B. B., JR., MCCORD, J. M., AND FRIDOVICH, I. (1970) J. Biol. Chem. 24.5, 6176-6181; VANCE, P. G., KEELE, B. B., JR., AND RAJAGOPALAN, K. V. (1972) J. Biol. Chem. 247, 47824786). Avimanganin which was isolated from chicken liver mitochondria (SCRUTTON, M. C. (1971) Biochemistry 10, 3897-3905) appears to be an in- active form of mitochondrial superoxide dismutase. The similarity between the mitochondrial superoxide dismutase and the corresponding enzymes from Escherichia coli and from Streptococcus mutans and the marked dissimilarity to the superoxide dismutase found in the cytosols of eukaryotes lends support to the theory that mitochondria have evolved from aerobic prokaryotes, which entered into an endocellular symbiosis with a protoeukaryote. Chicken liver was not unique in possessing distinct superoxide dismutases in cytosol and in mitochondrial compartments, since the same situation was shown to occur in pig heart.
The chicken liver cytosol superoxide dismutase exhibited two peculiar properties. Thus when stored in deep, con- centrated solutions it underwent autoreduction to a yellow cuprous form which retained enzymatic activity, and it po- lymerized to forms which likewise retained activity. Both of these changes were reversible and appeared to be due to reactive sulfhydryl groups which could act as the endogenous reductant and could form intermolecular disulfide bridges.
* This work was supported in part by Research Grant GM-10287 from the Xational Institutes of Health, Bethesda, Maryland.
The cytosol superoxide dismutase consisted of a family of electrophoretically distinct forms which could readily be separated on a preparative scale by isoelectric focusing. These multiple forms were indistinguishable in size and in specific activity. The chemical basis of these dilIerences in charge was not explored.
Superoxide dismutase catalyzes the reaction 02 + 0, + 2IIf + Hz02 + OS (4, 9-12) and appears to be ubiquitous among oxygen-metabolizing organisms and to be lacking in obligate anaerobes (13). This enzyme has been isolated from a wide range of eukaryotic sources including bovine erythrocytes and heart (4, 5), garden peas (6), Xaccharomyces cerevisiae (8), and Arezlrospora crassa (7) and in all of these was found to be a blue- green cupro-zinc protein. In contrast, the superoxide dismutase isolated from the prokaryotes Escherichia coli (1) and Slrepto- coccus mutans (2) was a pinkish manganoprotein. These two types of superoxide dismutase appeared to be completely unre- lated proteins which had evolved independently.
During an investigation of the superoxide dismutase of chicken liver, it was observed that the enzyme from mitochondria n-as distinct from that present in the cytosol with respect to several properties including electrophoretic mobility, susceptibility to inhibition by cyanide, and to inactivation by mixtures of chloro- form plus ethanol. Since the cupro-zinc superoxide dismutases were known to be inhibited by cyanide whereas the mangano- superoxide dismutases were not (14), it appeared possible that the mitochondria of chicken liver contained the prokaiyotic type of superoside dismutase, whereas the cytosol contained the eukaryotic type. Previous isolations of the superoxide dismu- tases of eukaryotes had used a simple but rigorous procedure (4) which included treatment with the chloroform-ethanol mixture of Tsuchihashi (15). This treatment inactivates the prokaryotic type of superoxide dismutase so that, even had it been present in the mitochondria and therefore in the extracts prepared from the diverse eukaryotic sources (5, 6, 8)) it would have been inac- tivated and would have gone undetected. The purification and characterization of the superoxide dismutases present in the cytosol and in the mitochondria of chicken liver was therefore undertaken. This paper describes the discovery of the differ- ences between the mitochondrial and the rytosol superoxide dis-
3582
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3583
mutasea aud t’he purification and properties of the cytosol and of the mitochondrial superoxide dismutases.
The profound differences between the superoside dismutases of cytosol and of mitochondria and the great similarity between the mitochondrial and the bacterial superoxide dismutases would seem to bear upon the question of the origin of mitochon- dria. These results are therefore discussed in view of the theory that mitochondria evolved from aerobic prokaryotes which entered into an endocellular symbiotic relationship with a proto- eukaryote (16-20). The generality of these findings was tested by examining the superoxide dismutases present in the cytosol and mitochondria of pig heart.
XSTERIALS AND METHODS
Cytochrome c, type III, was purchased from Sigma. Frozen chicken livers and livers from freshly slaughtered chickens were obtained locally. Nicrogranular diethylaminoethylcellulose (DE-32) and carbosymethylcellulose (CM-52) were obtained from the Reeve =Ingel Company. Sephadex G-75 was a product of Pharmacia. Standard solutions of copper, zinc, and manga- nese were obtained from Fisher. Ammonium sulfate was the reagent grade from Baker. Xanthine oxidase, which was gen- erously provided by Dr. K. V. Ra,jagopalan of this department was prepared by a procedure which avoided exposure to proteoly- tic agents (21). Superoxide dismutase was assayed as previously described (4) but with the addition of 1 x 1O-3 M cyanide to in- hibit cytochrome c peroxidases and oxidases (22). In these assays it was found advisable to add the ferricytochrome c im- mediately prior to initiating the reaction, by the addition of santhine oxidase. This was to avoid modification of the cyto chrome c by cyanide. The xanthine oxidase, used in these assays, was protected against inhibition by cyanide by the pres- ence of xanthine (23). This level of cyanide did not inhibit the mitochondrial superoxide dismutase. Cyanide does not inhibit manganosuperoxide dismutase but does inhibit the corresponding cupro-zinc enzymes (14). Ultracent,rifugal analyses were per- formed by the method of sedimentation equilibrium (24) in 0.1 M KCl, 0.005 M potassium phosphate, 1 X low5 M EDTA, at pH 7.8 and 25”, with 1 yc fi-mercaptoethanol, in a Beckman model E analytical ultracentrifuge. Assessment of the purity of the en- zyme by disc gel electrophoresis was performed on 7.5% poly- acrylamide gels essentially as described by Davis (25) and the gels stained for protein were compared with duplicate gels which were stained for supcroside dismutase activity (22). Superoxide dis- mutase activity was localized on polyacrylamide gels by the pho- tochemical method previously described (22) but with the modification that the gel cylinders were suspended in 0.05 3% potassium phosphate, 1 x lo-* M EDTA at pl-I 7.8 and 25”, im- mediately prior to and during illumination. Cyanide-sensitive superoxide dismutase could be differentiated from cyanidc-insen- sitive superoxide dismutase on polyacrylamide elcctrophoreto- grams, by including 1 x 10e3 M cyanide in the developing rea- gents to suppress the activity of the sensitive enzyme. Elcctro- phoresis in acid urea gels was performed as described by Panyim and Chalkley (26). The molecular weight of the subunits of the enzyme was estimated by disc gel electrophoresis on 10% poly-
acrylamide gels in the presence of sodium dodecyl sulfate (27). Molecular weight standards used to calibrate the gels were trans- ferrin, 77,000; human albumin, 68,500; ovalbumin, 43,000; pep- sin, 35,000; carbonic anhydrase, 29,000; trypsin, 23,000; and bovine superoxide dismutase, 16,300. Manganese, copper, and zinc were quantit,ated by atomic absorption spectrophotometry with the use of a Perkia-Elmer model 107 which was calibrated
with st’andard solutions of salts of these metals. Protein was es- timated on the basis of its absorbance in the ultraviolet (28) or short ultraviolet (29) or by use of a biuret method (30). Crystal- line bovine serum albumin was used to standardize the biuret method. Results obtained from measurements of the absorb- ance of the purified superoxide dismutase in the short ultraviolet (29) were corrected by a factor of 1.57. They then coincided with the results of the ultraviolet biuret method (30). The reason for this correction is presented in the section dealing with the absorption properties of the mitochondrial superoxide dis- mutase. Optical spectra were recorded with a Cary model 15. Isoelectric focusing was performed at 1200 volts with an LKB 440.ml electrofocusing apparatus using a 170 ampholyte solu- tion. Duplicate samples for amino acid analyses were hydro- lyzed for 24, 36, and 48 hours at 110” in 6 M HCl plus 0.1% phenol under vacuum, and, after vacuum drying and resuspen- sion, the hydrolyzed amino acids were chromatographed. Total cysteine plus half-cystine was determined in duplicate according to the performic acid oxidation method of Hirs (31). Trypto- phan was measured by the spectrophotometric method of Edelhoch (32). Total cysteine was also determined by the di- methylsulfoxide method of Spencer and Wold (33). Chicken liver mitochondria were isolated by differential centrifugation in isotonic sucrose by the well documented method of Borst et al.
(34).
RESULTS
Alultiple Forms of Xuperoxide Dismutase in Chicken Liver- Soluble extracts of a single fresh chicken liver, prepared by ho- mogenization, ultrasonication, and centrifugation, contained at least. four electrophoretically distinct superoxide dismutases. These, which are shown in the drawing of Gel 1 of Fig. 1, may be referred to as A, B, C, D in order of increasing mobility. There were, in fact, some bands of activity beyond D but these were very faint and are represented by the shaded band in Gels I, 3, 5 and 7. Band A was unaffected by 1 mr\l cyanide whereas Bands B, C, and D were eliminated by cyanide. This is illustrated by Gel 2. On the other hand Bands B, C, and D were unaffected by the Tsuchihashi (15) mixture of chloroform and ethanol whereas Band A was eliminated by this treatment. This result is shown in Gel S. A soluble extract of isolated chicken liver mitochondria contained only Band A whereas the cytosol frac- tion of an isotonic sucrose homogenate of chicken liver contained Bands 13, C, and D but not Band A. This is shown by Gels 4
1 2 3 4 5 6 7
FIG. 1. Disc gel electrophoresis of various chicken liver prepa- rations at pH 8.9. The dark gels were stained for superoxide dismutase activity while the light gels were stained for protein as described in the text. Gel 1, crude homogenate; Gel d, crude homogenate plus cyanide; Gel 3, crude homogenate after treat- ment with chloroform and ethanol; Gel 4, extract from isolated mitochondria; Gel 5, cytosol fraction; Gel 6, pure mitochondrial superoxide dismutase; Gel 7, pure cytoplasmic superoxide dis- mutase.
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3584
and 5. Band A, which was due to the cyanide-insensitive super- oxide dismutase and which was eliminated by the chloroform- ethanol procedure, proved to be due to a manganoenzyme. Gel 6 contained a sample of this purified manganosuperoxide dismutase and was stained for protein rather than for activity. Bands B, C, and D proved to be due to a cupro-zinc superoxide dismutase present in the cytosol. This superoxide dismutase apparently exists as a group of electrophoretically distinct forms and thus gave rise to a family of bands. Gel 7 contained a sample of the purified chicken liver cupro-zinc superoside dismutase and was stained for protein rather than for activity. This pattern of bands was not modified by the presence of mercaptoethanol.
Purification of Cupro-zinc Superoxide Diswdase from Chicken
Liver-Unless stated otherwise, all manipulations were performed at O-4”. Five pounds of frozen chicken liver were homogenized
2.0-
0.5 -
FIG. 2. Chromatography on G-75. Pooled fractions from the CM-52 column were concentrated to a small volume over a PM- 10 ultrafiltration membrane and then diluted to 30 ml with 0.1
0 20 30 40 50 60 70 80 90
Fraction
M KCI, 0.005 M potassium phosphate, and 1OV M EDTA, pH 7.8, and then subjected to gel exclusion chromatography on a column (2.5 X 100 cm) of Sephadex G-75, which had been equilibrated with the diluting buffer. Fractions 70 to 87, which were of high specific activity, were pooled and concentrated. The small activity peak near Fraction 64 is probably due to a polymer of the superoxide dismutase (see text).
for 2 min with 4 liters of water in a gallon capacity stainless steel
liver during homogenization.
Waring Blendor, which was operated at top speed.
The slurry was clarified by centrifugation for 30 min at 13,000 x g.
The water
The supernatant solu- tion was mixed with 0.25 volume of ethanol and 0.15 volume of
was initially at room temperature, but was chilled by the frozen
chloroform and was stirred for 15 min, during which time its color changed to a light tan. It is this treatment with organic solvents which eliminates the mitochondrial superoxide dismu- tase and hence allows the use of whole frozen liver as a convenient source of the cytosol enzymes without interference from the mitochondrial enzyme. The suspension was then centrifuged for 15 min at 13,000 x g, and the supernatant solution was al- lowed to come to room temperature. Solid K~HPO1 (300 g per liter) was then added and, after stirring for 15 min at room tem- perature, the phases, which had been salted out, were allowed to separate. The less dense, ethanol-rich phase was collected and chilled and clarified by centrifugation. Two volumes of cold acetone were added under stirring, and the precipitate which formed was collected by centrifugation and was suspended in a minimal volume of 0.015 M sodium acetate buffer, whose pH was 5.0 and whose conductivity was 0.7 mmho. This enzyme solu- tion was then dialyzed against three changes, of 50 volumes each, of this acetate buffer over a period of 24 hours. The precipitate, which formed during dialysis, was removed by crntrifugation and the clear, brownish solution was stirred for 30 min with 100 g of CM-52 which had previously been equilibrated with the same acetate buffer. The ChI-52 was then collected on a 10. cm Buchner funnel and was washed with the 0.015 &c acetate buffer until the effluent had an absorbance of less than 0.010 at 280 nm. The CM-52 was then elutetl with 0.065 M sodium acetate buffer, at pH 5.0, until the fractions which were collected in tubes (13 x 150 mm) were no longer perceptibly greenish. All of the green fractions were pooled and were concentrated by ultrafiltration over a 150-mm iZmicon PM-10 membrane, down to a small volume of dark green liquid, which was then diluted to 30 ml with 0.1 M KCl, 0.005 M potassium phosphate, and 1 X
lo-5 M EDTA at pH 7.8. This material n-as subjected to gel exclusion chromatography on a column (2.5 x 100 cm) of Sepha- des G-75, which had been equilibrated with the same KCl- phosphate buffer. The enzyme eluted from this column as a major band plus a minor band. The specific superoxide dismu- tase activity was constant across the major band. This elution pattern is shown in Fig. 2. The active, blue-green material, which was obtained from t.his column, was pooled as shown in Fig. 2 and concentrated by ultrafiltration. The yield was 235 mg of enzyme, whose specific activity was 3295 units per mg.
Table I summarizes the results of each of the steps in this pro- cedure.
Criteria of Purity-As was shown in Fig. 1, this material was separable into several bands by electrophoresis on polyacryla- mide gel. All of these bands were, however, active. Thus, there were no protein components which did not exhibit super- oxide dismutase activity. In adtlitiou, the enzyme, while still fresh, cluted from Sephades G-75 in the form of a band of coil- stant specific activity as shown in Fig. 2. When this enzync
was brought to sedimentation equilibrium in the presence of 1 y0 mercaptoethanol, analysis of the protein distribution by the method of Yphantis (24) indicated that it was monodisperse with respect to sedimentation properties.
ilJolecular Weight-When the purified cytosol superoside dis- mutase was dissolved in 1 T; mclrcaptoetlianol, 0.1 M KU, 0.005
RI potassium phosphate, and 1 x 10-j M EDT.& at ~1% 7.8 and
then brought to sedimentation equilibrium in a centrifugal field, the resultant distribution of protein gave a linear Yphantis (24) plot. This is shown in Fig. 3. From the slope of the line in Fig. 3 and a measured solvent &nsity of 1.00554 and an assumed partial specific volume of 0.73, a molecular weight of 30,600 was calculated. In contrast, when similar operations wereperformed, but in the absence of mcrcaptoethanol, 1)olydispersity with rc- spect to sedimentation properties became apparent. Thus, as is shown in Fig. 4, at least three components were detectable, and their molecular weights were estimated to be 32,000, 48,000, and 78,000, respectively.’ Thcsc results suggested that this chicken liver cupro-zinc supcroxide diamutase had a tendency to polymerize by the formation of tlisulfide bridges. This was
1 These estimated molecular weights were derived from the Yphantis plots (Fig. 4), after correcting the gradient due to a given heavy component for the contribution made by the gradients of all lighter components. Thus the steep slope of a line segment representing the gradient of the heavy component was corrected by subtracting the slope of the line segment which represented the gradient of lighter components. Molecular weights were then based on these corrected slopes.
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3585
TABLE I Purijkation of chicken liver cytoplasmic superoxide dismutase
Preparation stage
Supernatant from homogenate.. . Supernatant from Tsuchihashi fractionation. Ethanolic phase Acetone precipitate resuspended and dialyzed. Aft,er chromatography on CM-52. After chromatography on G-75.
Volume
ml
3575 3740 1340 700 40 75
Protein con- centration
mg/nzl
65.5 21.5 8.96 5.57
20.0 3.13
FIG. 3. Equilibrium sedimentation of cytoplasmic superoxide dismutase. The enzyme, at a concentration of 1.0 mg per ml in 0.1 M KCl, 0.005 M potassium phosphate, 1 X 10m6 M EDTA, pH 7.8, and 1% p-mercaptoethanol, was equilibrated at a rotor speed of 34,000 rpm. Five fringes were read from an interference optics photograph and were averaged, and the In fringe displacement was plotted as a function of the square of the distance from the center of rotation. The calculated molecular weight, according to the method of Yphantis (24), is 30,600.
FIG. 4. Equilibrium sedimentation of cytoplasmic superoxide dismutase in the absence of p-mercaptoethanol. The enzyme at a concentration of about 1.5 mg per ml in 0.1 M KCl, 0.005 M po- tassium phosphate, and 1 X 10-S M EDTA, pH 7.8, was equil- ibrated at a rotor speed of 24,000 rpm, and the In of the absorb- ance at 280 nm plotted against the square of the distance from t.he center of rotation. A portion of the plot gives the straight line shown in A which corresponds to a molecular weight of 32,000. When the enzyme was allowed to reach a new equi- librium at 12,000 rpm, two new straight line segments were plotted as shown in B and C. Their slopes’ correspond to mo- lecular weights of 48,000 and 78,000, respectively. These results indicate that the enzyme tends to form polymers in the absence of p-mercaptoethanol.
verified by gel exclusion chromatography. Thus, a sample of the purified enzyme, which had been stored frozen for at least 1 month, was equilibrated with 0.1 51 KCl, 0.005 M potassium phosphate, and 1 x lo-” M EDTA at pH 7.8 and then chromato- graphed on a column (2.5 X 100 cm) of Sephadex G-75 which
Total protein Total units Specific
activity Yield
g x 10-g cdmg %
234.0 6.00 26.0 100 80.4 3.78 47.0 62 12.0 2.65 222 44 3.90 1.95 503 32 0.800 1.20 1500 20 0.235 0.773 3295 12.7
1.0 1.8 8.5
19.7 57.7
126.7
FIG. 5. Chromatography of purified cytoplasmic superoxide dismutase on G-75. Purified enzyme which had been stored frozen for about a month was subjected to gel exclusion chro- matography on a Sephadex G-75 column equilibrated to 0.1 ~5 KCl, 0.005 M potassium phosphate, and 1 X lo+ M EDTA, pH 7.8. At least three molecular weight fractions are present as shown by the triangles. This is a sample which had previously eluted on this same column as a single peak. When an identical sample was run on the same column with the addition of 1% mercaptoethanol to the buffer, the two early eluting peaks were largely converted to the lowest molecular weight form, as shown by the circles. Thus, the polymerization was probably due to the formation of disulfide bridges.
had been equilibrated with the same buffer. The elution profile showed three bands of superoxide dismutase activity. When the entire procedure was repeated, but in the presence of 1% mercaptoethanol, only a single peak of activity was obtained. These results are illustrated in Fig. 5. It is apparent that this superoxide dismutase does polymerize during storage and that the polymers, which retain activity, may be dissociated again by treatment with mercaptoethanol.
X&units-When chicken liver cytosol enzyme was subjected to polyacrylamide gel electrophoresis in the presence of 152 sodium dodecyl sulfate and 0.1 M mercaptoethanol (27) a major band of molecular weight 16,800 and a minor band of molecular weight 31,600 were observed. These molecular weights were estimated from comparisons of their mobility with the mobilities of standards as described under “Materials and Methods.” Sodium dodecyl sulfate, per se, appeared to accentuate the tendency of this enzyme to polymerize so that electrophoresis in 1 y0 sodium dodecyl sulfate, but in the absence of mercaptoetha- nol, gave multiple, diffuse, and faint bands. It may be inferred that this cytosol superoxide dismutase was composed of two sub- units of identical size but whether or not these subunits were joined by disulfide bridges remains uncertain.
d4etaZs-&4tomic absorption spectrophotometry demonstrated that the purified cytosol superoxide dismutase contained 0.30% copper and 0.25% zinc. ‘This calculates to 0.9 atom of copper
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3586
TABLE II
Amino acid composition of cytoplasmic chicken liver
superoxide dismutasea
Residues per mole of enzyme (nearest
integer) Amino acid A mine acid content b
-I. ?>%oles/nzole enzyme
Lysine ......................... 19.5 Histidine. ...................... 14.2 Arginine ....................... 8.4 Aspartic acid. .................. 31.6 Threonine ...................... 17.6c Serine .......................... 14.1c Glntamic acid. ................. 23.0 Proline ......................... 11.5 Glycine ........................ 50.1 Alanine. ....................... 21.5 Valine ......................... 28.2’1 hlethioninc ..................... 4.0 Isoleucine ...................... 14.1” Lencine ........................ 15.8’” Tyrosine ....................... 2.0 Phenylalanine .................. 7.8
Cysteic acid .................... 14.3” Tryptophan .................... 0’
Total residues. ...............
20 14
8 32 18 14
23 12
50 22 28
4
14 16
2
8 14
0
299
0” ’ 2 ’ 8 ’ ’ ’ ’ r ’ ’ 400 450 500 550 600 650 i nanometers
60
FIG. 6. Visible absorption spectra of cytoplasmic superoxide dismutase. Freshly isolated enzyme at a concentration of 84.4 mg per ml in 0.05 M potassium phosphate-l X lo-” M EDTA, pH 7.8, was examined in a I-cm cuvette. This curve is marked oxidized. When a trace of sodium dithionite was added to the cell and the spectrum was taken again, the curve marked reduced was seen. When a sample of the fresh enzyme was examined with the use of reduced enzyme as the blank, the curve marked difference was obtained. The spectrum of autoreduced enzyme was identical with enzyme reduced by dithionite.
00 270 290 310 330 350 370 nanometers
FIG. 7. Ultraviolet absorption spectrum of cytoplasmic super- oxide dismutase. Enzyme at a concentration of 2.2 mg per ml in 0.05 M potassium phosphate-l X lo-4 M EDTA, pH 7.8, was examined in a l-cm cell.
and 0.8 atom of zinc per subunit. Manganese, which would have been measurable at 0.01 atom per subunit, was not detected.
Optical Spectra-The purified cytosol enzyme exhibited a
broad absorption band whose maximum was at 670 nm (E, =
233 liters per mole cm). This absorption band, which is associ- ated with the Cu++ prosthetic group, was bleached within a few seconds by such reductants as mercaptoethanol, cysteine, NazSz02, or ascorbate. The reduced enzyme retained its activity and was rather stable in air. Reoxidation in air was a very slow process and occurred over a period of hours. The reduced en- zyme could, however, be rapidly reoxidized by ferricyanide. Fig. 6 compares the visible absorption spectra of the superoside
dismutase, as isolated, with that of the enzyme after reduction
with mercaptoethanol and also presents the difference spectrum. The ultraviolet absorption spectrum of the chicken liver
cytosol enzyme is shown in Fig. 7. This spectrum is similar to that seen with other cupro-zinc superoxide dismutases (4-8) and
probably reflects both an absence of tryptophan (28) and ab- sorbance by the ligand field of the metal prosthetic groups (35-
a Triplicate samples were hydrolyzed 24, 48, and 72 hours.
b All calculations based on molecular weight of 30,400. c Values were extrapolated to zero time at hydrolysis. d Values were extrapolated to infinite time of hydrolysis.
8 Triplicate samples hydrolyzed with dimethylsulfoxide by the method of Spencer and Wold (23).
/ From interpretation of ultraviolet absorption spectrum
37). The molar extinction coefficient at 258 nm was 8,750 liters per mole cm.
Amino Acid Composition of Cytosol Enzyme-Amino acid analyses were performed after acid hydrolysis as described under “Materials and Methods.” The results of such analyses were summarized in Table II. No attempts were made to analyze directly for tryptophan. Its absence II-as inferred from the ultraviolet absorption spectrum and was not unexpected (28).
Butoreduction of Superoxide Dismutase-When a concentrated (250 mg per ml) solution of superoxide dismutase in 0.05 _II potassium phosphate-l x 10e4 M EDTA, pH 7.8, was allowed to stand at 4”, those portions of the solution which were more than 0.75 cm from the air interface turned yellow over a period of weeks. This yellow enzyme had an absorption spectrum which was identical with that of the superoxide dismutase which had been reduced with mercaptoethanol. The blue-green and the autoreduced yellow materials exhibited identical activity. When they were assayed for cuprous copper, by t.he 2,2’-biquinoline method (38), the blue-green material was found to have only 6% of its copper in this reduced form, whereas the yellow ma- terial had 88% of its copper in the cuprous state. Reduction of the blue-green superoxide dismutase with mercaptoethanol con- verted all of its copper to the cuprous form.
Freshly isolated superoxide dismutase was titrated with p-
chloromercuriphenyl sulfonate in 0.05 M potassium phosphate-l x 10m4 M EDTA, pH 7.8, and was found to contain 1.2 f 0.2
sulfhydryl groups per subunit. In contrast, solutions which had been stored for several weeks, either frozen or at 4”, were found to contain 0.2 to 0.3 sulfhydryls group per subunit. The aged enzyme could be restored to a sulfhydryl titer of 1 per sub-
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3587
unit by treatment with 0.1 y0 cyst&u.? followed by anaerobic dialysis. It appears likely that the sulfhydryl group, which in the fresh enzyme was reactive toward mercurials, can account for autoreduction of the Cu++ and for the polymerization of this enzyme. This reactive sulfhydrgl group is apparently not cribical for the enzymatic activity, since neither aging nor mer- curials affected the level of superoxide dismutase activity.
Isolation of Mitochondrial Superoxide Dismutase--Unless stated otherwise all steps were performed at 4”. Fresh chicken liver (30 pounds) was homogenized with 40 liters of 0.25 M sucrose-O.01 nr K,HP04 with the use of a 4-liter stainless steel Waring Blendor. Each portion of liver and sucrose was homog- enized for 10 s at low speed followed by 20 s at high speed. The homogenate was centrifuged at 2,000 X g for 8 min. The super- natant was filtered through double cheesecloth, was diluted with 3 volumes of 0.9cG KCl, and was then passed at a flow rate of 200 ml per min through a refrigerated Sharples supercentrifuge operating at 45,000 rpm. It was necessary to change centrifuge bowls at least twice during this procedure. The mitochondrial paste (810 g) was scraped from the centrifuge bowls, and a small sample was examined at magnification of 1,000 times under phase contrast. This material proved to be composed almost entirely of mitochondria with only small inclusions of cell debris or of nuclei. The mitochondria were then frozen and thawed by suspension in 5 volumes of 0.05 M potassium phosphate, 1 X lo-* M EDTA at pH 7.8, which was initially at 25”. This sus- pension was then exposed, in 400.ml batches, to ultrasonic disruption for 5 min at 4” under a Rranson sonifier operated at, a power output of 125 watts. The sonicate was clarified by centrifugation for 60 min at 27,000 X g, and the supernatant solution was then heated to 60” for 2 min. This heating was done by immersing 800.ml portions in 2-liter Erlenmeyer flasks iu a 70” water bath under continual agitation. The copious precipitate which formed during heating was removed by cen- trifugation for 15 min at 27,000 X g. Streptomycin sulfat,e was added to the supernatant to a final concentration of 2.57, and, after incubation at room temperature for 30 min, the suspension was clarified by centrifugation. Solid (NH&SO4 was added (383 g per liter) to bring the solution to 60% of saturation and after stirring for at least 4 hours at 24”, the precipitate was re- moved by centrifugation and was discarded. The supernatant, solution was brought to 90% saturation with (NHJSO, by addition of 213 g per liter of the solid salt and was stirred at 24” for at least 4 hours. The precipitate was collected by centrifu- gation and was suspended in a minimal volume of 0.05 nl potas- sium phosphate, 1 X lo-* M EDTA at pH 7.8. This suspension was dialyzed versus several changes of 0.005 M potassium phos- phate, 1 x 10e5 11 EDT& pH 7.8, over a period of 24 hours and was then clarified by centrifugation. The supernatant solution was passed through a column (2.5 X 15 cm) of DE-32 which had been equilibrated with the dialysis buffer. The column was then washed with this buffer until the effluent contained less than 1 unit (4) of superoside dismutase activity per 50 ~1. The effluent was pooled and concentrated by ultrafiltration over an Amicon P;11-10 membrane to a total volume of 50 ml. It was then dialyzed against 0.04 M potassium acetate, pH 5.5, and was adsorbed onto a column (2.5 x 25 cm) of CM-52 which had been equilibrated with the same buffer. The column was then washed with that buffer until a small peak of protein was eluted. One liter of a linear gradient of potassium acetate (0.04 to 0.20 M) at pH 5.5 was then applied, and the fractions of highest specific activity were pooled and concentrated by ultrafiltration over an Amicon PYI-10 membrane. Fig. 8 presents the results of t,his
FIG. 8. Chromatography on CM-52. The mitochondrial prepa- ration was equilibrated by dialysis with 0.04 M potassium acetate, pH 5.5, and then applied to a column (2.5 X 25 cm) of CM-52 equilibrated with the same buffer. The column was washed with this buffer until eight fractions (20 ml) had been collected and then 1000 ml of a linear gradient of this same buffer was ap- plied. A---A, absorbance at 280 nm; O---O, superoxide dismutase activity of the fractions collected; - - -, conductivity of the fractions collected. Tubes 14 to 18 were pooled for further purification.
0.4
JO.3 2 -0.2 2
% z
f”’ I
0 50 60 70 80 90 100 110 120 130
Fraction
FIG. 9. Chromatography on G-200. The pooled fractions from the CM-52 column (Fig. 1) were concentrated on a PM-10 mem- brane and dialyzed to equilibrium against 0.10 M KCI, 0.05 M potassium phosphate, and 1 X 10e4 M EDTA, pH 7.8. They were then subjected to gel exclusion chromatography on a column (2.5 X 120 cm) of Sephadex G-200 which had been equilibrated with t,his buffer. Fractions (7.5 ml each) were collected and Fractions 72 to 83 were pooled and characterized. O-O, absorbance at 280 nm; O--O, activity; A--A, manganese content.
chromat,ographic fractionation. The concentrated enzyme so lut,ion, which was visibly pink, was dialyzed against 0.1 M KCI, 0.05 nr potassium phosphate, 1 x 10e4 M EDTA, pH 7.8, and then subjected to gel exclusion chromatography on a column (2.5 X 120 cm) of Sephadex G-200, which had been equilibrated with this buffer. Fig. 9 presents the results of this procedure. The enzyme eluted as a single peak of constant specific activity. The final yield was 52.2 mg with a specific activity of 3400 units of superoxide dismutase per mg of protein and a recovery of 407,. The results of the individual stages of the purification procedure are summarized in Table III.
Estimation of Purity-Electrophoresis at pH 8.9 on poly- acrylamide gel revealed a single band of protein which coincided with the band of activity. This activity was not diminished perceptibly by 1 X 10M3 M cyanide. Polyacrylamide gel electro- phoresis in acid urea (26) after incubation with 1 y. P-mercapto- ethanol also gave a single band of protein. When the treatment with 6.mercaptoethanol was omitted, polyacrylamide electro- phoresis in acid urea (26) showed an additional minor band whose
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3588
TABLE III Purification of chicken liver mitochondrial superoxide dismutase
Purification Stage VOlLlitle Protein* Specific Total TOtal Yield Purification (ml) cone . Activity Units Protein (%) FaCtOr
@T/mu Wmg) (ml)
Supernatantfrom 4400 57.1 1.75 4.4x105 251,240 100 1.0 sonicate
Heat Step
Streptomycin Sulfate
3800 35.3 2.75 3.7x105 134,140 84.1 1.6
3800 19.3 5.08 3.7x105 73,340 84.1 2.9
60% tiniw Sulfate
4560 11.3 7.58 4.1x105 51.528 93.2 4.5
90% Anxronium Sulfate. Precipi- tate Redissolved
435 9.1 115 4.6~10~ 3,959 104.5 66
Dialysis and Centrifugation
600 3. 7 I 164 3.6~10~ 2220 81.8 94
After Passing through DE-32
940 1.0 376 3.5x105 940 79.7 214
After Ccl~xx? Ci-XCM-C- 8.70 ' 10.47 2420
2.21x105 91.1 50.1
1383 1 I
tcqraphy on CM-52
After Column Chrcano- 5.64 12.93 3400 1.77x105 52.2 40.1 1941 ,tcqraphy on G-200 ,
* Assuming E:tm = 1.0 at 280 nm. After column steps, the ultraviolet biuret method was used (30).
FIG. 10. Equilibrium sedimentation of mitochondrial super- oxide dismutase from chicken liver. Superoxide dismutase at a concentration of 1.0 mg per ml in 0.10 M KCI, 0.005 M pOtaSSiUm phosphate, and 1 X lCt+~ EDTA, pH 7.8, was brought to equilib- rium at a rotor speed of 34,000 rpm. Five fringes of an interfer- ence optics photograph were read and averaged, and In fringe displacement was plotted as a function of the square of the dis- tance from the center of rotation.
intensity seemed to increase with the age of the sample. Elec- trophoresis on polyacrylamide gels in the presence of 1 y0 sodium dodecyl sulfate (27) also gave a single band of protein. As shown in Fig. 9, gel exclusion chromatography on Sephadex C-200 gave a single peak of constant manganese to protein ratio. Finally, equilibrium ultracentrifugation indicated that the ma- terial was homogeneous with respect to its sedimentation proper- ties. Th.is is illustrated in Fig. 10. All of these criteria indi- cated that the mitochondrial enzyme had been purified to homogeneity. The minor band, which was seen in acid urea gels, suggests that air oxidation of mitochondrial superoxide dismutase can modify its electrophoretic mobility under these conditions and that this change was reversible by reduction with @-mercaptoethanol.
Molecular Weight-Mitochondrial superoxide dismutase at a
concentration of 1.0 mg per ml was brought to equilibrium in a centrifugal field, and the resultant distribution of protein was analyzed by the method of Yphantis (24). As shown in Fig. 10 the graph of In fringe displacement versus square of the distance from the center of rotation was linear. From the slope of this line and an assumed partial specific volume of 0.730 a molecular weight of 79,400 was calculated. This is exactly twice the mo- lecular weight previously reported for the bacterial superoxide dismutase (1, 2).
Mitochondrial superoxide dismutase was incubated in 1 u/;, sodium dodecyl sulfate with and without 0.1 M P-mercapto- ethanol and was then subjected to electrophoresis in polyacryl- amide gels (27). Proteins of known subunit weight were used to calibrate these gels. Chicken liver mitochondrial superoxide dismutase was found to be composed of subunits whose weight was 20,200 f 500 in the presence of /Z-mercaptoethanol and 18,500 f 500 in its absence. The subunits of the corresponding mitochondrial and bacterial (1, 2) enzymes are therefore identical in size. It is clear that the mitochondrial enzyme is composed of four subunits of identical weight, which are not covalently linked. The fact that P-mercaptoethanol caused an apparent 10% incresise in the molecular weight of the mitochondrial super- oxide dismutase subunits may be significant. Thus, intrachain disulfide bridges would cause a more compact and cor- respondingly more mobile subunit, and the mercaptan would then cause a decrease in mobility, which would be seen as an apparent increase in molecular weight. We tentatively con- clude on this basis that thrre are iutrachain disulfide bridges in the mitochondrial superoxide dismutase.
d4etal Content of Mitochondrial Enzyme-Analyses for man- ganese revealed 0.148c?0 manganese. This calculates to 2.13 manganese per molecule of weight 79,400. This analysis was repeated on several batches of the enzyme and was reproduced
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3589
nanometers
FIG. 11. Visible absorption spectrum of mitochondrial super- oxide dismutase. Enzyme at a concentration of 4.3 mg per ml was examined in 0.05 M potassium phosphate-l X lo-4 M EDTA, pH 7.8. The light path was 1.0 cm.
01
%oo I I
220 240 260 280 300 320 nanometers
FIG. 12. Ultraviolet absorption spectra of mitochondrial super- oxide dismutase. Curve 1 shows the enzyme at a concentration of 0.43 mg per ml in 0.05 M potassium phosphate-l X lo-* M EDTA, pH 7.8, with a l-cm light path. Curve 2 shows the enzyme at a concentration of 0.022 mg per ml in the same buffer.
X40%. The purified enzyme was also analyzed for copper and zinc with negative results. There could not have been more than 0.04 copper or zinc per molecule of the enzyme or they would have been detected.
Optical Spectra-The absorption spectrum of the mitochondrial superoxide dismutase was similar to that previously seen2 with thr bacterial enzyme (1, 2). Thus, as shown in Fig. 11, there was a broad absorption band with a maximum at 475 nm and a shoulder close to 600 nm. The extinction coefficient at 475 nm was 292 per subunit, which may be compared with correspond- ing coefficients of 200 and 280 for the E. coli (1) and the S. mutans (2) enzymes, respectively. These bacterial enzymes also ex- hibitcd shoulders in their absorption curves at 600 nm.
The mitochondrial superoxide dismutase also exhibited an absorption maximum in the ultraviolet at 252 nm, whose molar extinction coefficient was 144,000 1~~~ cm-l. The ratio of ab- sorbances at 280 nm and at 260 nm was 2.60. In the short ultra- violet region there was an unusual absorption shoulder at 225 nm. These results are shown in Fig. 12. This absorption at
2 It should be noted that the visible absorption maximum of Escherichia coli superoxide dismutase was correctly stated (1) to be at 473 nm, whereas the visible absorption spectrum actually shown (1) ‘was displaced inadvertently toward shorter wave length.
'200 I 210 I 220 I 230 I 240 250 26 nanometers
FIG. 13. Far ultraviolet spectra of several superoxide dismu- tases and of bovine serum albumin. --, mitochondrial super- oxide dismutase from chicken liver; . ..., Streptococcus mutans superoxide dismutase; ----, Escherichia coli superoxide dismutase; ---1 chicken liver superoxide dismutase from the cytoplasm; -.-., bovine serum albumin. Spectra were run in 0.05 M potas- sium phosphate-l X lo-” M EDTA, pH 7.8, with the use of l-cm cells at convenient concentrations, and the absorbances of each protein were multiplied by whatever factor was required to make the absorbance at 202.5 nm equal 1.0.
225 nm was of some interest since it is not characteristic of poly- peptides (39, 40) and since it would obviously interfere with the determination of protein concentration by the methods of Murphy and Kies (29), which is based upon the difference in absorbances at 215 nm and 225 nm and which was used in pre- vious st,udies of the manganosuperoxide dismutase from E. coli (I) and X. mutans (2). Fig. 13 shows that the superoxide dis- mutases from E. coli and from S. mutans also show an absorption shoulder at 225 nm, whereas the cupro-zinc superoxide dis- mutase from chicken liver cytosol does not. Bovine serum albumin showed this absorption shoulder to a small degree, possibly due to adsorbed metals. The ratio of the absorbances at 215 nm to those at 225 nm were 2.03 for the cupro-zinc super- oxide dismutase of chicken liver cytosol, 1.80 for bovine serum albumin, 1.39 for the S. mutans superoxide dismutase, and 1.31 for the superoxide dismutases from E. coli and from chicken liver mitochondria.
This shoulder at 225 nm leads to a systematic underestima- tion of the protein concentration, when the method of Murphy and Kies (29) is applied to solutions of the purified mangano- supcroside dismutases. On this basis it appears that the spe- cific activities and the manganese contents previously reported for the superoside dismutascs from E. coli (1) and from X. mu- tans (2) were overestimated. If we may extrapolate from the results obtained with the mitochondrial enzyme, this overesti- mation would have been by a factor of 1.6. When corrected for this source of error the manganese contents of the bacterial superoxide dismutase become virtually identical with that now reported for the mitochondrial enzyme. Thus, both contain 0.6 atom of manganese per subunit.
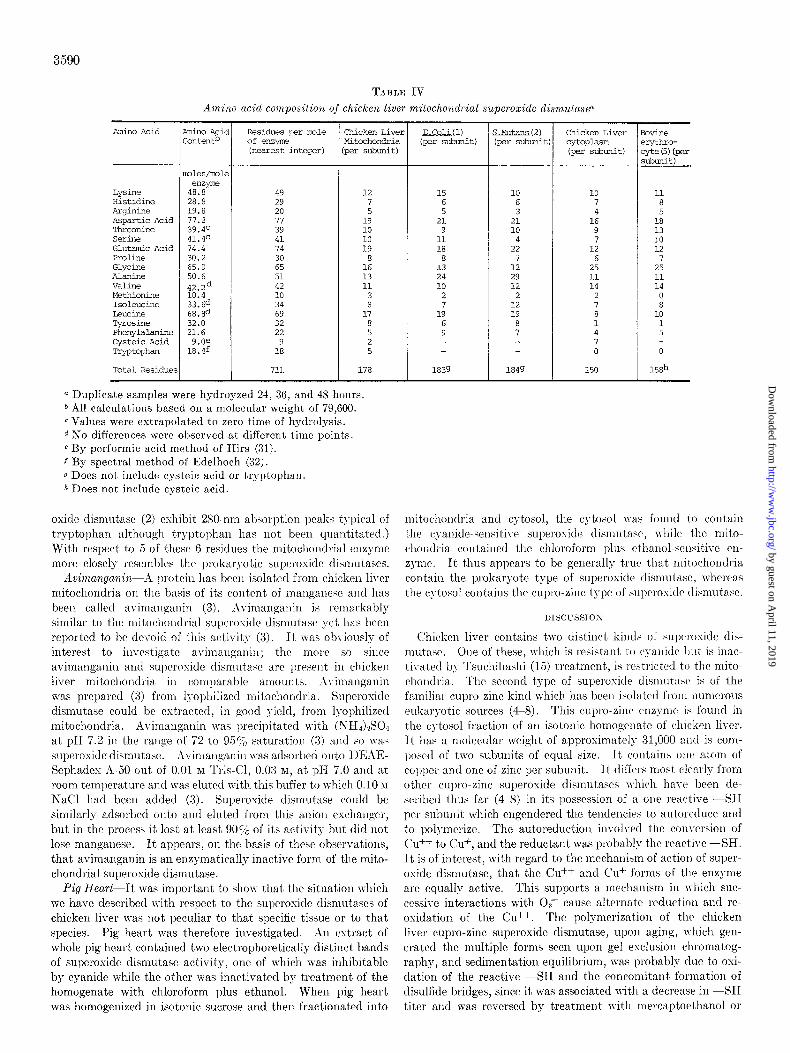
Amino Acid Andysis--The amino acid composition of mito- chondrial superoxide dismutase is given in Table IV and com- pared to several previously reported compositions of prokaryotic and eukaryotic superoxide dismutases. For 6 residues (glu- tamate, glycine, alanine, leucine, tyrosine, and tryptophan’l clear differences exist between the prokaryotic and eukaryotic values. (E. coli superoside dismutase (1) and S. mutans super-
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from
3590
TABLE IV Amino ucid composition of chicken liver mitochondrial supwoxide dismutase~
Amino Acid
Lyslne Hisbdine hrqinine Aspartic Acid Thremine SeClX Glutamic Acid PrOline Glycine Alanine Valine Methionine ISCJl~UCine Leucine Tyrosine Phenvlalanine Cyst& Acid Tryptophan
Total Residue:
tin0 Acic ,ontentb
Dles/lmle enzym?
48.8 28.8 19.8 77.2 39.4c 41.4c 74.4 30.2 65.0 50.6 42.2d 10.4 33.64 68.8d 32.0 21.6
9.Of 18.4f
Residues per mle Chicken Liver of emyE Mitcchondria klearest iwqer) per subunit)
49 29 20 77 39 41 74 30 65 51 42 10 34 69 32 22
9 18
711
12 7 5
19 10 10 19
8 16 13 11
3 8
17
2 5
i78 1839
0 Duplicate samples were hydroyzed 24, 38, and 48 hours. b All calculations based on a molecular weight of 79,GOO. c Values were extrapolated to zero time of hydrolysis. d No differences were observed at different time points. 6 By performic acid method of Hirs (31). f By spectral method of Edelhoch (32). g Does not include cysteic acid or tryptophan. h Does not include cysteic acid.
oxide dismutasc (2) cshibit 280-am absorption peaks typical of tryptophan although tryptophan has not been quantitated.) With respect to 5 of thcsc 6 residues the mitochondrial enzyme more closely resemblrs the 1)rokaryotic superoside tlismutases.
Avimanganin-X lnotein has been isolated from chicken liver mitochondria on the basis of its content of mangancec and has been called a\-imallgallin (3). .i\imanganin is remarkably similar to the mitocahondrial sul)crositlc dismutase yet has been reported to bc devoid of this activity (3). It was obviously of interest to invcstigatc avimaugallill; the more so since avimanganin and superoxide dismutasic arc present in chicken liver mitochondria in c*omparable amounts. ;1vimanganin was prepared (3) from lyophilized mitochondria. Superosicle dismutase could bc cstractetl, in good yield, from lyophilizctl mitochondria. Avimanganin was precipitated lvith (XI-I&S04 at pII 7.2 in the range of 72 to 957; saturation (3) and so was superoxide dismutase. llvirnallgallill was adsorbed oIlto l)EhE- Sephades h-50 out of 0.01 M T&Cl, 0.03 M, at $1 7.0 and at room teml)crature and was elutcd with this buffer to which 0.10 JI NaCl had been added (3). Superoxide dismutase could be similarly adsorbed onto and eluted from this anion exchallger, but in the process it lost at lrast SOLi’ of its activity but did not lose manganese. It appears, on the basis of these observations, that avimanganin is an enzymatically inactive form of the mito- chondrial superoxide dismutase.
Pig Heart-It was important to shop that the situation which we have described with respect to the superoxide dismutascs of chicken liver was not peculiar to that specific tissue or to that species. Pig heart was therefore investigated. Xn extract of whole pig heart contained two electrophoretically distinct bands of superoxide dismutasc activity, one of which n-as inhibitable by cyanide while the other was inactivated by treatment of the homogenate with chloroform plus ethanol. When pig heart was homogenized in isotonic sucrose and then fractionated into
E&a. ( 1) (per submit)
15 6 5
21 9
11 18
8 13 24 10
2 7
19 6 9
10 6 3
21 10
4 22
'I 12 29 12
2 12 19
1849
Chicken Liver cytopl&Xl (per subunit)
10 7 4
16 9 7
12 6
25 11 14
2 7 8 1 4 7 0
150
bvine ?rythKo- *e(5) (per xbunit)
11 8 5
18 13 10 12
7 25 11 14
0 8
10 1 5
0
lsEh
mitochondria and cytosol, the cytosol was found to contain the c:!allide-seasitiv~ superosidc dismutase, while the mito- chondria contained the chloroform plus ethanol-sensitive cii- z)-me. It thus appears to be generally true that mitochondria contain the prokaryotc type of superoxide rlismutasc, whcrca.s the cytosol contains the cupro-zinc type of sul)rrosidc tlismutase.
Chicken liver contains two distinct kinds of supcrosiclc dis- mutasc. One of these, which is resistant to ryanidr but is inac- tivatod by Tsuchihashi (15) treatment, is restricted to the mito- chondria. The second type of superoxide dismutasr is of the familiar cupro-zinc kind which has been i,solatetl from numerous eukaryotic sources (4-8). This cupro-zinc enzyme is foulld in the cytosol fraction of an isotonic homogrnate of chickrn liver. It has a molecular weight of approximately 31,000 and is com- l)osrtl of two subunits of equal size. It contains onr atom of copper and one of zinc per subunit. It differs most rlrarly from othrr cupro-zinc superoxide dismutases which ha\-c, been de- scribed thus far (4-8) in its possession of a on(’ reactive --SH per subunit which engendered the tendencies to autoreduce and to polymerize. The autoreduction involved the conversion of vu++ to CL?, and the reductant was probably the reactive --SH. It is of interest, with regard to the mechanism of action of super- oxide dismutase, that the Cu++ and Cu+ forms of the enzyme are equally active. This supports a mechanism in n-hich suc- ccssivc interactions with 02 cause alternate reduction and re- oxidation of the CL?. The polgmcrization of the chicken liver cupro-zinc superoxidc dismutase, upon aging, which gen- erated the multiple forms seen upon gel exclusion chromatog- raphy, and sedimentation equilibrium, was probably due to osi- dation of the reactive -SH and the concomitant formation of disulfide bridges, since it was associated with a decrease in --SH titer and was reversed by treatment with mercaptoethanol or
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3591
cysteine. The chicken liver cytosol superoxide dismutare ap- parently exists as a family of electrophorctically discrete forms. These forms were readily separated during electrophoresis ou polyacrylamide gel as shown in Fig. 1. These multiple forms could also be separated on a preparative scale by isoelectric focusing. Thus, 500 mg of superoxide dismutase was mixed with 440 ml of 1 y0 pH 5 to 8 LKB Ampholine, and this solution was electrofocused for 24 hours in an LKB electrofocusing ap- paratus according to the directions in the LKB manual. Several sharply focused green bands were seen. These eluted at, pH 6.75, 6.69, 6.37, 6.26, 5.80, and 5.35. ,211 of these bauds were active. It is not known whether these multiple forms are truly isozymes or whether they represent the product of a single gene variably modified by amidation or by some other post-transla- tional change.
It is apparent from the results shown in Fig. 1 that the elec- trophoretic properties of the cupro-zinc superoside dismutase were unmodified by a purification procedure which included treatment with chloroform plus ethanol. &sags of activity
likewise demonstrated that the Tsuchihashi procedure was with- out effect on this enzyme. These results are in conflict with the assertion by Hartz and Deutsch (41) that treatment of cupro- zinc superoxide dismutase with chloroform plus et’hanol yields a modified protein.
The mitochondria of chicken liver have been found to contain a superoxidc dismutase which is strikingly similar to that pre- viously isolated from bacteria (I, 2) and is totally unlike that previously isolated from a wide range of eukaryotes (4-S). The mitochondrial superoxide dismutase was not previously detected in eukaryotes because the purification procedure, which was developed originally for the superoxide dismutase of erythro- cytes (4)) was applied to these eukaryotes with apparent success. It was not then appreciated that the Tsuchihashi procedure (15), which was appropriate for the purification of the cupro- zinc enzyme, caused complete loss of the manganoenzyme. l’re- vious isolations of superoxidc dismutase from eukargotes simply eliminated the mitochondrial enzyme by treatment with chloro- form-ethanol. Pig heart mitochondria also contain a super- oxide dismutase which is similar to the manganoenzyme and unlike the cupro-zinc enzyme in being inactivated by the Tsuchihashi procedure and in beiug unaffected by cyanide. It appears likely that the mitochondria of all eukargotes will prove to contain the manganosuperoxide dismutase whereas the cyto- sols of these eukaryotes will prove to contain the cupro-zinc superoxide dismutasc.
Why should the mitochondrial and the bacterial enzymes be so similar to each other and so different from the corresponding enzyme present in the cytosol of eukaryotcs? It appears most likely that present day mitochondria and bacteria share a com- mon ancestor which was already distinct from the ancestor of present day eukaryotes at the time when the accumulation of atmospheric oxygen called forth the evolution of superoxide dismutase. This result lends support to the theory that mito- chondria have descended from aerobic prokaryotes which en- tered into an endocellular symbiosis with a protoeukaryote. This theory has already received a great deal of attention and appears to be well supported on the basis of other data (16-20) although it has recently been re-examined and questioned (42). It is of course possible to state that the resemblance between the mitochondrial and the prokaryotic superoxide dismutases is the result of convergent evolution. This would however ne- cessitate the existence of similar selection pressures upon free
living bacteria, as upon mitochondria, over the long reaches of the evolutionary time scale, and this is scarcely imaginable. It will be important to inquire whether t’he manganosuperoxide dismutasc, found in the mitochondrion, is or is not actually synthesized under the direction of the mitochondrial gcnome. There are other intriguing questions raised by the differences between the mitochondrial and the cytosol supcroside dismu- tases. Thus, what is the functional significance of this retention of the mailganosuperoxide dismutase by mitochondria? Another way to state this is, why cannot the mitochondrion make do with the cupro-zinc enzyme produced by the nuclear genome? These questioi~s are currently under active consideration.
Acl;?zo~ledgmenls-The authors arc indebted to Dr. Howard Steinman and Dr. Ross Tyc for help in performing the amino acid analyses.
REFERENCES
1. Knmr:, B. B., JIL, MCCORD, J. M., AND Fl~IDOvICH, I. (1970) J. Biol. Chem. 245, 6176-6181
2. VANCE, P. G., KEELE, B. B., JR., AND R.IJAGOPAL.~N, K. V. (1972) J. Biol. Chem. 247, 4782-4786
3. SCRUTTON, M. C. (1971) Biochemistry 10, 3897-3905 4. MCCOILD, J. M., AND FRIDOVICH, I. (1969) J. Biol. Chem. 244,
6049-6055 5. KEELE, B. B., JR., MCCORD, J. M., AND FKIDOVICH, I. (1971)
J. Biol. Chem. 246, 2875-2880 6. SAWADA, Y., OIIYAMA, T., AND YAMAZAIiI, I. (1972) Biochim.
Biochim. Biophys. Acta 268, 305-312 7. MISRA, H. P., AND FRIDOVICH, I. (1972) J. Biol. Chem. 247,
3410-3414 8. GOSCIR’, S. A., .~ND FRIDOVICH, I. (1972) &o&m. Biophys.
Acta 289, 276~283 9. MASSEY, V., STIUCI~LAND, S., MAYHE~~, S. G., Ho\\-ELL, L. G.,
EXGLE:, P. C., MATTHEWS, 1~. G., SCHUX\N, M., AND SULLI- VAN, P. 8. (1969) Biochem. Biophys. Res. Commun. 36, 891- 897
10. BALLOU, D., PALMER, G., AND M~SSIGY, T’. (1969) Biochem. Biophys. Res. Commun. 36, 898-904
11. ROTILIO, G., BRAY, R. C., AND FINLDEN, II. hI. (1972) Biochim. Biophys. A eta 268, 605609
12. KLUG, D., RABANI, J., AND FRIDOVICI-I, I. (1972) J. BioZ. Chem. 247, 4839-4842
13. MCCORD, J. M., KISI~;LIC, B. B., JR., AXD FIIIDOVICH, I. (1971) Proc. Sat. Acad. Sci. U. S. A. 68, 102-1-1027
14. BEAUCIUMP, C. 0. (1973) Ph.D. dissertation, Duke University 15. TSUCHIH.LSHI, M. (1923) Biochem. 2. 140, 63-112 16. ROOD-L-X, D., AXO WILKIE, D. (1968) The Biogenesis of Mifo-
chondria, Methuen, London 17. BOARDMAX, X. K., LINNANE, A. W., .&ND SMILLIE, R. M., eds
(1971) Autonomy rind Biogenesis of Mifochondria and Chloro- plasts, American Elsevier New York
18. COHEN, S. S. (1970) Amer. Sci. 58, 281-289 19. SCHNEPF, IX., AND BROKEN, R. M., JR. (1972) in Results and
Problems in Cell Differentiation, (REINERT, J., AND UR- SPRUNG, H., eds) Vol. 2, pp. 299-322, Springer-Verlag, Berlin
20. MAE~GULI~, L. (1970) The Origin of &‘ukaryotic Cells, Yale Uni- versity Press, New Haven
21. BR.IDY, F. 0. (1969) Ph.D. dissertation, Duke University 22. BE.&UCHAMP, C. O., AND FRIDOVICH, I. (1971) Anal. Biochem.
44, 276-287 23. DIXON, M., AND KEILIN, D. (1935) Proc. Roy. Sot. Ser. B
Biol. Sci. 119, 159 24. YPHAXTIS, D. A. (1964) Biochemistry 3, 297-317 25. DAVIS. B. J. (1964) Ann. N. Y. Acad. Sci. 121. 404-427 26. PANY&, S., A&D C&LKLEY, R. (1969) Arch. B&&hem. Biophys.
130, 337-346 27. WEBER, K., AND OSBORN, M. (1969) J. Biol. Chem. 244, 4406-
4412 28. WETLAUFER, D. B. (1962) Advan. Protein Chem. 17, 303-390 29. MURPHY, J. B., AND KIES, M. W. (1960) Biochim. Biophys.
Acta 45, 382-384
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3592
30. ELLMAN, G. L. (1962) Anal. B&hem. 3, 40-48 31. HIRS, C. H. W. (1956) J. Biol. Chem. 219, 611-621 32. EDELHOCH, H. (1967) Biochemistry 6, 1948-1954 33. SPENCER, R. L., AND WOLD, F. (1969) Anal. Biochem. 32, 185-
190 34. BORST, P., RUTTENBERG, G. J. C. M., BND KROON, A. A. (1967)
Biochim. Biophys. Acta 149, 140-155 35. BANNISTER, J., BANNISTER, W., AND WOOD, E. (1971) Eur. J.
Biochem. 18, 178-186 36. ROTILIO, G., CALABRESE, L., BOSSA, F., BARRA, D., AGRO, A.
F., AND MONDOVI, B. (1972) Biochemistry 11, 2182-2192
37. WESER, U., BARTH, G., DJERASSI, C., HARTMAN, H., KRAUSS, P., VOELCKER, G., VOELTER, W., AND VOETSCH, W. (1972) Biochim. Biophys. Acta 278, 28-44
38. FELSENFELD, G. (1960) Arch. Biochem. Biophys. 3’7, 247-251 39. GOLDFARB, A. R., SAIDEL, L. J., AND MOSOVITCH, E. (1951) J.
Biol. Chem. 193, 397-404 40. RICKLI, E. E., GHAZANF.4~, S. A. S., GIBBONS, B. H., AND
EDSA~,, J. ?I’. (1964) J. &oZ. Chem: 239, 106g1078 41. HARTZ. J. W.. AND DEUTSCH. H. F. (1972) J. Biol. Chem. 247.
704317050 ’ ~ I
42. R~FF, R. A., AND MAHLER, H. R. (1972) Science 177, 575-582
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Richard A. Weisiger and Irwin FridovichSuperoxide Dismutase: ORGANELLE SPECIFICITY
1973, 248:3582-3592.J. Biol. Chem.
http://www.jbc.org/content/248/10/3582Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/248/10/3582.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 11, 2019
http://ww
w.jbc.org/
Dow
nloaded from


















