
Sub-lethal Effects of Anthropogenic Contaminants on ...
72
ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2018 Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science and Technology 1692 Sub-lethal Effects of Anthropogenic Contaminants on Aquatic Invertebrates WARREN KUNCE ISSN 1651-6214 ISBN 978-91-513-0382-6 urn:nbn:se:uu:diva-355236
Transcript of Sub-lethal Effects of Anthropogenic Contaminants on ...

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2018
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1692
Sub-lethal Effects of AnthropogenicContaminants on AquaticInvertebrates
WARREN KUNCE
ISSN 1651-6214ISBN 978-91-513-0382-6urn:nbn:se:uu:diva-355236

Dissertation presented at Uppsala University to be publicly examined in Zootissalen,Evolutionsbiologiskt centrum, Norbyvägen 14-18, Uppsala, Friday, 7 September 2018 at10:00 for the degree of Doctor of Philosophy. The examination will be conducted in English.Faculty examiner: Doctor Jes Rasmussen (Department of Bioscience - Stream and WetlandEcology, Århus University).
AbstractKunce, W. 2018. Sub-lethal Effects of Anthropogenic Contaminants on Aquatic Invertebrates.Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Science andTechnology 1692. 70 pp. Uppsala: Acta Universitatis Upsaliensis. ISBN 978-91-513-0382-6.
Anthropogenic contaminants are considered to play a substantial role in the decline of freshwaterinvertebrate diversity. Sub-lethal effects of many of these contaminants on behaviour and life-history traits of aquatic invertebrates may contribute to their decline. As contaminants arerarely present in the environment alone, the effects of mixture exposures are highly relevant inassessing the risk these substances pose to the biota. This thesis focuses on sub-lethal effectsof exposure to aquatic pollutants, separately and in combination, on fresh-water invertebrates.To investigate the single and combined effects of pesticides, larvae of the midge, Chironomusriparius were exposed to a 1 hour pulse of two neonicotinoids and two pyrethroids. This shortexposure to environmentally relevant concentrations of pesticides decreased the survival anddelayed development in C. riparius. The combination of neonicotinoids and pyrethroids did notproduce synergistic effects; however, there was some indication of antagonism. Additionally,larvae of the damselfly, Coenagrion puella, were exposed for 14 hours to two environmentallyrelevant concentrations of pyrethroid pesticides, alone and in combination. Exposure to thepyrethroid, deltamethrin, reduced the larvae’s predatory ability. Combined exposure to bothdeltamethrin and esfenvalerate inhibited the Glutathione S-transferase detoxification pathwayand may have additive toxic effects on the larvae’s predatory ability. Microplastics areincreasingly gaining attention as an aquatic pollutant of major concern with respect to thetoxicity of the microplastics themselves as well as their capacity to adsorb persistent organicpollutants like pesticides. To investigate the effects of microplastics and a pyrethroid, alone andin combination, C. riparius larvae were raised in sediment spiked with two sizes of polystyrene-based latex microbeads and an environmentally relevant concentration of esfenvalerate undernormal and food-restricted conditions. Exposure to both sizes of microplastics and esfenvaleratelead to equally decreased emergence under food-restricted conditions. Additionally, exposureto esfenvalerate led to decreases in survival when food was scarce that did not occur whenmicroplastics were co-present. Antishistamines are also an emerging aquatic contaminant ofconcern with very little known about their biological effects on aquatic wildlife. Antihistaminescould potentially interfere with the histaminergic pathways and thus affect thermal tolerance andtemperature preference in aquatic invertebrates. The freshwater snail, Planorbarius corneus,was exposed for 24 hours to the antihistamine, diphenhydramine. This exposure increasedthermal tolerance and righting time, but did not affect temperature preference. The results ofthe investigation suggest that anthropogenic contaminates alone and/or in combination havesub-lethal effects on life history, behavior and physiology of aquatic invertebrates. Such sub-lethal effects have the potential to affect populations and community structure in the aquaticand terrestrial environment.
Keywords: ecotoxicology, sub-lethal, behavior, biomarkers, thermal tolerance, pesticides,pyrethroids, neonicotinoids, pharmaceuticals, antihistamines, microplastics, mixtures,freshwater, invertebrates, chironomids, damselflies, snails
Warren Kunce, Department of Ecology and Genetics, Animal ecology, Norbyvägen 18 D,Uppsala University, SE-752 36 Uppsala, Sweden.
© Warren Kunce 2018
ISSN 1651-6214ISBN 978-91-513-0382-6urn:nbn:se:uu:diva-355236 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-355236)

For Zoë


List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Kunce, W., Josefsson, S., Örberg, J., Johansson, F.
(2015). Combination effects of pyrethroids and neonico-tinoids on development and survival of Chironomus ripar-ius. Ecotoxicology and Environmental Safety, (122: 426—431).
II Kunce, W., Stoks, R., Johansson, F. (2017). Single and mixture impacts of two pyrethroids on damsel predatory behavior and physiological biomarkers. Aquatic Toxicolo-gy, (190: 70—77).
III Kunce, W., Varg, J. E., Outomuro, D., Svanbäck, R. and Johansson, F. (2018). Combined effects of microplastics and a pyrethroid on the benthic invertebrate, Chironomus Riparius. Manuscript.
IV Kunce, W. and Kührer, L.* (2018). Effects of acute anti-histamine exposure on thermal tolerance and behavior in the freshwater snail, Planorbarius corneus. Submitted manuscript.
* The authors contributed equally
Reprints were made with permission from Elsevier.


Contents
Introduction ................................................................................................... 11 Pyrethroids and Neonicotinoids ............................................................... 12 Microplastics ............................................................................................ 13 Antihistamines .......................................................................................... 14 Mixtures ................................................................................................... 15 Sub-lethal Effects ..................................................................................... 15
Objectives ..................................................................................................... 17
Methods ........................................................................................................ 18 Animals .................................................................................................... 18 Experiments .............................................................................................. 18 Chemical Analyses ................................................................................... 21 Life-history Traits .................................................................................... 22 Behavioral Assays .................................................................................... 23 Molecular Biomakers ............................................................................... 24 Thermal Tolerance ................................................................................... 24 Statistics ................................................................................................... 25
Results and Discussion ................................................................................. 27 Combination effects of pyrethroids and neonicotinoids (Paper I) ............ 27 Single and mixture impacts of two pyrethroids (Paper II) ....................... 30 Combined effects of microplastics and a pyrethroid (Paper III) .............. 34 Effects of an antihistamine (Paper IV) ..................................................... 38
Concluding Perspectives ............................................................................... 42
Lay Summary ................................................................................................ 45
Svensk Sammanfattning ................................................................................ 50
Acknowledgements ....................................................................................... 55
References ..................................................................................................... 58


Abbreviations
D E I T RSW PC1 PC2 GST MDA ETS MP CTmax Tpref RT
Deltamethrin Esfenvalerate Imidacloprid Thiacloprid Reconstituted soft water 1st Principal Component 2nd Principal Component Glutathione-S-transferase Malondialdehyde Electron transport system Microplastics Critical maximum temperature Temperature preference Righting time

10

11
Introduction
Why should we tolerate a diet of weak poisons, a home in insipid surround-ings, a circle of acquaintances who are not quite our enemies, the noise of motors with just enough relief to prevent insanity? Who would want to live in a world which is just not quite fatal?
― Rachel Carson, Silent Spring
The quality of the world’s freshwaters is deteriorating and they are losing biodiversity at an alarming rate (UNEP, 2016). Extinction rates of freshwater animals in well-studied ecosystems are estimated at 4% per decade (Ricciardi and Rasmussen, 1999). All of the driving forces behind this dis-turbing trend are linked to human activities (Vaughn, 2010). The release of anthropogenic contaminants in freshwaters is one of these forces (Dudgeon et al., 2006). Water quality legislation now controls the inputs of some con-taminants in parts of the world (Clean Water Act, 1972; European Commission, 2000). However, continued scientific efforts are required to identify and assess the risks of emerging pollutants as well as scrutinize es-tablished benchmarks for previously evaluated substances.
Restrictions on the first wave of mass-produced pesticides led to the de-velopment of newer generations of unregulated pesticides with greater non-mammalian specificity (Ensley, 2007a, 2007b). Contaminants such as plastic that were once considered inert are now under suspicion (Rochman et al., 2013). Advances in modern pharmaceuticals pose new challenges as these compounds are designed specifically to interact with physiological systems at very low concentrations (Deo and Halden, 2013). Rain, modern plumbing, dust and dumping all shepherd these advances of human civilization to the lakes and rivers were they come into contact with wildlife (Deo and Halden, 2013; Schulz, 2004; Wagner et al., 2014). Advances in waste water treat-ment technology are able to remove some, but far from all contaminants and implementation of such facilities around the world is still patchy and highly dependent on local economies and infrastructures (Bartelt-Hunt et al., 2009; Kay et al., 2018; Population Council, 2005; Weston et al., 2013). Conse-quently, waste water effluent is still a direct route of many anthropogenic contaminants into aquatic ecosystems. The now common practices of using biosolids from waste water treatment facilities as fertilizer and reclaimed water for irrigation deliver complex mixtures of sequestered pesticides,

12
pharmaceuticals, microplastics and other anthropogenic compounds to agri-cultural land where, along with pesticides that are applied directly to crops, they are susceptible to transport to aquatic ecosystems via wind and rain run-off (Topp et al., 2012; Walters et al., 2010; Weithmann et al., 2018; Weston et al., 2013). This thesis investigates adverse effects of four such contami-nants commonly detected in waste water effluent, biosolids and freshwater bodies: two neonicotinoid and two pyrethroid pesticides, polystyrene micro-plastics, and an antihistamine, on non-target aquatic invertebrates.
Pyrethroids and Neonicotinoids Two studies in this thesis contribute new knowledge on the biological effects of pesticide mixtures, knowledge that is important for improving the risk assessment of these chemicals. Pesticides are major contributors to the dete-rioration of aquatic biodiversity in freshwater ecosystems (Liess et al., 1999). An assessment of European monitoring sites found that 87% of the chemical contaminants that exceeded the acute risk threshold for inverte-brates were pesticides (Malaj et al., 2014). Pyrethroids and neonicotinoids are two classes of systemic pesticides that are known to be toxic to non-target aquatic organisms (Palmquist et al., 2012; Tennekes, 2010) and yet still occur regularly in surface waters above regulatory limits (Graaf et al., 2010). Adverse effects of exposure reported include loss of coordination and paralysis (Nyman et al., 2013; Samsøe-Petersen et al., 2001), feeding inhibi-tion (Anderson, 1982; Nyman et al., 2013; Reynaldi et al., 2006), reduced growth (Amweg et al., 2006), altered emergence patterns (Beketov, 2004; Schulz and Liess, 2001), reduced fecundity (Beketov and Liess, 2005; Forbes and Cold, 2005; Palmquist et al., 2008; Reynaldi and Liess, 2005) and acute mortality (Kreutzweiser et al., 2008; Osterberg et al., 2012).
Pyrethroids, such as deltamethrin and esfenvalerate investigated in this thesis, are neurotoxins that work by disrupting the votage-gated sodium channels of nerves which causes convulsions and paralysis (Soderlund, 2012). They were developed with greater selective toxicity to insects and other invertebrates as replacements for the organochlorine and organophos-phate pesticides of the mid 20th century, which were also highly toxic to mammals, notably the farmers that used them (Maund et al., 2012). Even though they are still in wide-use today, growing resistance of target pests to pyrethroids, led to the development of the neonicotinoids (Field et al., 2017). Similar to pyrethroids, neonicotinoids are also broad-spectrum neurotoxic insecticides with low toxicity to mammals. However the mechanism of ac-tion is different. Neonicotinoids, such as imidacloprid and thiacloprid inves-tigated in this thesis, interfere with the nicotinic acetylcholine receptors in the postsynaptic neurons (Jeschke et al., 2010; Tomizawa and Casida, 2005). Neonicotinoids were approved for use in the European Union in 2005 (Euro-

13
pean Commission, 2018a). However, use of imidacloprid has recently been restricted due to evidence of its toxicity to bees and thiacloprid has recently received a higher risk label based on its endocrine disrupting properties (Eu-ropean Commission, 2018a)
Microplastics Microplastics are an emerging contaminant of concern and data assessing their biological effects on aquatic organisms and interactions with other con-taminants is scarce. This thesis aims to contribute new knowledge on the biological effects of chronic exposure to microplastics, alone and in combi-nation with a pesticide. Microplastics include plastic particles that are small-er than 5 mm and come from manufacturing industries (both intentional products and byproducts) and the degradation of larger plastic products like water bottles, fishing equipment and synthetic clothing (Horton et al., 2017). Microplastics enter the aquatic environment directly via municipal and in-dustrial effluents (Blair et al., 2017; Horton et al., 2017), and atmospheric fallout (Dris et al., 2016) as well as indirectly by application of biosolids in agriculture (Mintenig et al., 2017; Weithmann et al., 2018). The estimated annual environmental load of microplastics in the European Union is be-tween 75000 and 300000 tonnes (European Commission, 2018b). Concen-trations of microplastics in freshwater sediments have been recorded in lake sediments as high as 2346 particles/kg (Su et al., 2016). The true concentra-tions might be higher since the methodologies used to count microplastics have been criticized as likely to be underestimations (Dris et al., 2015; Syberg et al., 2015). Microplastics are considered a persistent and accumu-lating pollutant since the rate of release is greater than the rate of degradation (Barnes et al., 2009).
Laboratory studies have shown that many different kinds of aquatic inver-tebrates with various different feeding mechanisms will ingest microplastics and field studies have documented microplastic ingestion in freshwater and marine environments (Boerger et al., 2010; Browne et al., 2008; Eriksson and Burton, 2003; Graham and Thompson, 2009; Lusher et al., 2013; Mur-ray and Cowie, 2011; Scherer et al., 2017; Setälä et al., 2014; Thompson et al., 2004). Known ecotoxicological effects of microplastic ingestion include altered feeding capacity (Besseling et al., 2013; Browne et al., 2013; Cole et al., 2015), induced immobilization (Rehse et al., 2016), reduced weight (Besseling et al., 2013), decreased reproductive fitness (Cole et al., 2015; Sussarellu et al., 2016), inhibited acetylcholinesterase activity (Oliveira et al., 2013) and disruption of lysosomal membrane integrity (von Moos et al., 2012).

14
Antihistamines Pharmaceuticals are biologically active chemicals developed for high effec-tiveness at low concentrations, not only for humans, but also livestock and pets. Many drug targets are conserved across phyla including aquatic wild-life species (Gunnarson et al., 2008). Antihistamines are a class of pharma-ceuticals recently identified as a risk to aquatic organisms (Kosonen and Kronberg, 2009; Kristofco and Brooks, 2017). Antihistamines, developed to reduce the symptoms of allergic reaction in mammals, target histamine re-ceptors and are some of the most widely used medications in the world (Galeotti et al., 1999; Simons and Simons, 2011). Histamines are highly conserved neurotransmitters found in the nervous tissues, gut, skin and im-mune tissues of invertebrates where they are known to play a role in a wide variety of biological processes such as circadian and feeding rhythms, learn-ing, and memory (Carpenter and Gaubatz, 1975; Green, 2008; Osborne and Patel, 1984). Few studies have been conducted thus far that investigate the ecotoxicological effects of antihistamines, but the available data indicates that some invertebrates are equally or more sensitive than vertebrates to an-tihistamines (Kristofco and Brooks, 2017). Known adverse effects of expo-sure in aquatic vertebrates and invertebrates include inhibited growth (Hoppe et al., 2012), inhibited feeding (Berninger et al., 2011), reduced reproductive output (Berninger et al., 2011; Meinertz et al., 2010) and increased boldness (Jonsson et al., 2014). Histaminergic pathways are also known to modulate temperature preference and thermal tolerance in Drosophila and antihista-mine exposure was shown to interfere with these functions (Hong et al., 2006). Antihistamine-induced changes in temperature preference, thermal tolerance and other physiological processes could have adverse effects on aquatic ectotherms, particularly as they face increased climatic variability (Katzenberger et al., 2014; Piasečná et al., 2015).
Antihistamines have previously been used as a tool in the exploration of histaminergic pathways in molluscs (Matsuo et al., 2016), but this thesis includes the first study to investigate the ecotoxicity of antihistamine expo-sure in a mollusc. The antihistamine tested, diphenhydramine, was one of the first antihistamines developed and is still the active ingredient in widely sold allergy tablets (DrugBank, 2017; Statista, 2017 ). It has been recorded at concentrations as high as 1.4-50 μg/L downstream from waste water treat-ment facilities (Bartelt-Hunt et al., 2009; Ferrer et al., 2004) and is highly persistent in sediments with no observed degradation after 3 years (Walters et al., 2010).

15
Mixtures Risk assessment of “no effect” concentrations must also take into considera-tion the reality of freshwaters as “chemical cocktails” in which animals are not exposed to one contaminant at a time, but complex mixtures (Kortenkamp, 2007; Relyea, 2009). Growing resistance to pesticides has led to the practice of applying mixtures for maximum crop protection and this practice is reflected in the pesticide patterns found in aquatic environments (Ahmad, 2007; Ansara-Ross et al., 2012; Gilliom, 2007). Monitoring studies of pesticide loads in freshwaters report that suites of different pesticides are present and several are present in concentrations over the permitted bench-mark values (Ansara-Ross et al., 2012; Gilliom, 2007; Graaf et al., 2010; Kreuger, 1998). Studies on pharmaceutical loading in freshwaters also detect the presence of multiple co-occuring substances (Rosi-marshall and Royer, 2012). Additionally, microplastics in the environment have been shown to adsorb multiple difference chemical contaminants up to 1 million times the concentrations found in the water (Teuten et al., 2009). The ingestion of microplastics is therefore, highly likely to co-occur with the ingestion of adsorbed substances, particularly in sediments where both microplastics and chemical substances accumulate.
Interactions of multiple present xenobiotics within an organism can affect the uptake, availability, metabolism and elimination of these substances (Coors and Frische, 2011; Lydy et al., 2004). Interactions can result in addi-tive, antagonistic or synergistic effects where the impact on the organism is quite different from single exposure effects (Anderson and Zhu, 2004; Cedergreen, 2014; Kepner, 2004; Orton et al., 2012). Ingestion of microplas-tics may increase chemical exposure by desorption of contaminants in the gut and even potentiate chemical toxicity, however current data is scarce and inconclusive (Browne et al., 2013; Kleinteich et al., 2018; Oliveira et al., 2013). Interestingly, there is some evidence to suggest that ingested micro-plastics can behave as cleaning agents, adsorbing chemicals found in the gut and thereby reducing the chemicals’ bioavailability (Koelmans et al., 2013). Additional data on combination effects of common pollutants for improved risk assessment is in demand by regulatory authorities (European Commis-sion, 2012)
Sub-lethal Effects For centuries, humans trusted that the solution to pollution was dilution. However, with a population of 7.6 billion and rising at some point there will not be enough freshwater to adequately dilute all that we put in it and some suggest that point has already arrived (Johnson et al., 2001; Population Council, 2005). Both pesticides and pharmaceuticals are specifically de-

16
signed for high physiological impact at very low concentrations and effects on non-target organisms have been observed at concentrations far below those measured in the environment (Caldwell et al., 2008). For dilution to be considered a solution there must be a concentration cut-off for a contami-nant at which no adverse effects of exposure occur. Traditional ecotoxico-logical risk assessment is based on mortality data; the lethal concentrations of a substance for select model species. However, even in the absence of direct mortality, lower concentrations of contaminants can have effects that are ecologically relevant (Fleeger et al., 2003; Relyea and Hoverman, 2006). Exposure to sublethal concentrations of contaminants have been shown to adversely impact non-target organisms through changes in mobility (Baatrup and Bayley, 1993; Fong et al., 2017), foraging behaviors (Ingram et al., 2015), predator-prey interactions (Bisesi et al., 2016; Janssens and Stoks, 2012; Trekels et al., 2011), life-history traits (Goedkoop et al., 2010; Janssens and Stoks, 2013), fecundity (Cole et al., 2015; Toumi et al., 2013), and other physiological functions (Froese et al., 2009; Hyne and Maher, 2003). Therefore, in order to determine if a contaminant is being sufficiently diluted in a freshwater environment (i.e. the measured concentration induces no adverse effects) it is important to investigate sublethal effects of expo-sure.

17
Objectives
This thesis investigates three types of anthropogenic contaminants ubiqui-tous in freshwater ecosystems: pesticides, microplastics and pharmaceuti-cals. The four papers included in this thesis test the hypothesis that these substances can affect the behavior and life-history traits of aquatic inverte-brates and consequently the biodiversity of freshwater ecosystems.
Papers I and II investigate the effects of environmentally relevant concentra-tions of pesticides, alone and in combination. First, paper I focuses on the effects of a brief pulse exposure to two neonicotinoids and two pyrethroids on the life-history traits of the freshwater midge, Chironomus riparius. Then pa-per II explores acute exposure effects of two pyrethroids on predatory behav-ior and sub-organismal biomarkers of the damselfly, Coenagrion puella.
Paper III investigates the effects of latex-based polystyrene microspheres alone and in combination with esfenvalerate, one of the pyrethroid insecti-cides investigated in papers I and II. This is a spiked-sediment exposure that tracks the life-history traits of C. riparius which spends its larval stage in the benthos.
Paper IV aims to initiate more focus on the potential for antihistamines to interfere with the thermoregulatory physiology and behavior of aquatic in-vertebrates. The freshwater snail, Planorbarius corneus, was exposed to high concentrations of the antihistamine, diphenhydramine, and then evalu-ated for effects on thermal tolerance, temperature preference and righting behavior.

18
Methods
Animals Chironomous riparius is a common, non-pestilent midge species approved by the Organization for Economic Cooperation and Development (OECD) for sediment and water toxicity assessments. The short-life cycle and benthic larval stage of the midge make it a good model organism for investigating effects of aquatic contaminants on life-history traits. An in-house culture of > 10 generations provided the C. riparius larvae used in papers I and III.
Coenegrion puella is a common, non-pestilent damselfly species that oc-curs in the agricultural landscapes of Europe. Even though damselflies are not a typical model organism, they have been used in some ecotoxicological assessments (Campero et al., 2007; Chang et al., 2009, 2007; Hardersen and Wratten, 1996). The aquatic larval stage is vulnerable to pesticide exposure and its predatory behaviors are easily observed in the laboratory. The C. puella larvae used in paper II were hatched from eggs oviposted in the labor-atory by wild females caught in tandem pairs at two ponds in Uppsala, Swe-den. The larvae were 5-weeks-old at the time of the study and the head width (distance between the eyes) ranged from 0.74-2.01 mm (mean with SE: 1.12 ± 0.02 mm).
Planobarius corneus is an air-breathing freshwater snail common across Europe. Various gastropod species have been previously used in ecotoxico-logical assessments including P. corneus (Cacciatore et al., 2013; Hellou et al., 2009; Khalil, 2015). Aquatic snails are appropriate for evaluating ther-mal tolerance and temperature preference in aquatic ecotherms due to proven methodologies developed for non-toxicological studies (Gerald, 2005; Jo-hansson, 2015). An in-house culture of > 3 generations provided the P. corneus egg clutches that were subsequently hatched and reared for use in the experiments in paper IV. Reproductively mature snails were used in the experiments with a shell height of approx. 9 mm and a weight of approx. 300 mg.
Experiments The purpose of paper I was to investigate the effects of two pyrethroid and two neonicotinoid pesticides alone and in combination on the life-history

19
traits of a benthic invertebrate. First-instar larvae of C. riparius were ex-posed to a 1 h pulse of sub-lethal, environmentally relevant concentrations of deltamethrin, esfenvalerate, imidacloprid and thiacloprid, singularly, in pairs (deltamethrin + esfenvalerate, imidacloprid + thiacloprid) and all together. Reconstituted soft water (RSW) comprised of NaHCO3 (48 mg/L), CaSO4·2H2O (30 mg/L), MgSO4·7H2O (61.37 mg/L) and KCl (2 mg/L)(Greenberg et al., 1985) was used in all experimental vessels during and after the 1 h pulse exposure and then vessels were topped up with deion-ized water over the course of the experiment. A 1 h pulse was selected to simulate the first hour of a rain event when pesticide concentrations from run-off and storm water are at a maximum (Brady et al., 2006). The larvae were reared for 28 days and then assessed for survival, development time, and emergence. Thereafter, adults were collected in swarm cages according to treatment for mating, and fecundity was assessed in terms of the number of eggs per egg rope produced.
The aim of paper II was to investigate the acute exposure effects of two pyrethroid pesticides, alone and in combination, on the predatory behavior and sub-organismal biomarkers of a damselfly. Five-week-old larvae of C. puella were exposed for 14 h to sub-lethal, environmentally relevant concen-trations of deltamethrin and esfenvalerate, singularly and in combination. The pesticides were dissolved in acetone and acetone concentrations were adjusted uniformly across treatments, including the controls. The larvae re-covered for 2 h in clean water, then individually observed for 10 minutes while fed artemia prey and scored for hunting behaviors and general activity (see table 1). The exposure, recovery and behavioral assays were conducted in RSW. After the assays, the larvae were frozen at -80° C for later analysis of molecular markers of toxicity: glutathione-S-transferase (GST), oxidative damage (MDA) and electron transport system (ETS). Additional larvae were also exposed and frozen for these analyses without undergoing behavioral assays.
The objective of paper III was to investigate the effects of a chronic sedi-ment exposure to microplastics alone and combination with a pyrethroid pesticide, on the life-history traits of a benthic invertebrate. First-instar lar-vae of C. riparius were reared in sediment spiked with 1.0 µm or 10.0 µm latex-based polystyrene microbeads, a sub-lethal environmentally relevant concentration of esfenvalerate and a combination of the 1.0 µm microbeads and esfenvalerate. The overlying water in the experimental vessels was dechlorinated, aerated tap water. The experiment was conducted twice due to an error in the amount of spinach added to the artificial sediment resulting in low nutrient conditions during the first trial. The results of the first trial pro-vide additional insight into the effects of exposure under stressful conditions that may occur in nature. The low nutrient trial is referred to as “Experiment

20
A” and the normal conditions trial is referred to as “Experiment B.” The larvae were assessed for survival after 10 days and then again after 34 days for Experiment A and 28 days for Experiment B. The life-history traits as-sessed were emergence, development time and sex ratio of emerged adults.
The aim of paper IV was to explore the potential for antihistamines to have toxic effects on the thermoregulatory physiology and behavior of aquatic invertebrates. Three experiments were conducted exposing mature P. corneus for 24 h to the antihistamine, diphenhydramine and then carrying out post-exposure assessments (see Table 1). The first experiment deter-mined thermal tolerance by observing each snail’s attachment to a glass sur-face as the temperature of the water was increased 1 ° C every 30 minutes. The second experiment evaluated temperature preference of each snail in an aluminum channel with a temperature gradient of 8 - 33 ° C over a period of 24 h. Finally the third experiment observed the time it took for each snail to right itself after placement on its right side. A pilot-study using dechlorinat-ed, aerated tap water during the exposure period resulted in >90% post-exposure immobility in all treatments that did not occur when culture water was used, therefore the diphenhydramine exposures for all three experiments were conducted with solutions prepared by step-wise dilution using water from the culture. However, the three assays themselves were conducted in dechlorinated, aerated tap water with no observed adverse effects

21
Table 1. Overview of the exposure scenarios in papers I-IV.
Chemical Analyses It is standard practice in ecotoxicological studies to analytically quantify the concentrations of test substances in water and sediment exposures (OECD, 2000). The concentrations of pesticides in paper I were not analytically verified due to logistical restraints. However, the concentrations of deltame-thrin and esfenvalerate in water in paper II and the concentrations of esfen-valerate in sediment in paper III, were quantified at the Department of Aquatic Sciences and Assessment at the Swedish University of Agricultural Sciences in Uppsala, Sweden. The measurements of the four single expo-sures in paper II were 0.065 μg/L and 0.130 μg/L deltamethrin, and 0.069 μg/L and 0.130 μg/L esfenvalerate. The concentrations of the combination exposure were measured as 0.067 μg/L deltamethrin and 0.120 μg/L esfen-valerate. These concentrations fall within the ranges of deltamethrin and esfenvalerate detected in freshwaters (Brady et al., 2006; Feo et al., 2010; Lindström and Kreuger, 2015). The sediment concentrations of esfenvalerate
survival 1 Control 0 µg / Lemergence of adults 2 Deltamethrin 0.034 µg / Ldevelopment time 3 Esfenvalerate 0.065 µg / Leggs per egg rope 4 Imidacloprid 2.7 µg / L
5 Thiacloprid 2.3 µg / LDeltamethrin 0.034 µg / L Esfenvalerate 0.065 µg / LImidacloprid 2.7 µg / LThiacloprid 2.3 µg / L
Deltamethrin 0.034 µg / LEsfenvalerate 0.065 µg / LImidacloprid 2.7 µg / LThiacloprid 2.3 µg / L
Hunting Behaviours 1 Control 0 µg / Lhead orientation toward prey 2 Deltamethrin 0.23 µg / L
step towards prey 3 Deltamethrin 0.45 µg / Lstrike at prey 4 Esfenvalerate 0.23 µg / Lprey capture 5 Esfenvalerate 0.45 µg / L
General Activity Deltamethrin 0.23 µg / Lswimming bout Esfenvalerate 0.23 µg / L
walking boutMolecular Biomarkers
malondialdehyde (MDA)
10-day larval survival 1 Control -final survival 2 1.0 µm microplastics 14,219 particles/mL wet sediment
emergence of adults 3 10.0 µm microplastics 14,219 particles/mL wet sedimentdevelopment time 4 Esfenvalerate 0.07 µg/kg wet sediment
sex of emerged adults Esfenvalerate 0.07 µg/kg wet sediment 1.0 µm microplastics A - 14,219 particles/mL wet sediment
B - 21,328 particles/mL wet sediment
critical thermal maximum 1 Control 0 µg/Ltemperature preference 2 diphenhydramine 10 µg/L
righting time 3 diphenhydramine 100 µg/L4 diphenhydramine 1000 µg/L
Treatment Contaminants Nominal Concentrations
respiratory electron transport system (ETS)
glutathione-S-transferase (GST)
2
3
28/34 days sediment
Paper Organism Assessment ExposureNo. of
experiments
C. riparius larvae
I
6
IV 24 h water
C. riparius larvae
III
5
P. corneus
adults
8
7
61 h water 1
114 h waterC. puella larvae
II

22
in paper III were 0.053 μg/kg dry sediment for Experiment A and 0.064 μg/kg dry sediment for Experiment B. These concentrations were below the limit of quantification (5 μg/kg) and near the detection limit (0.03 μg/kg). The diphenhydramine measurements in paper IV were conducted at SWE-TOX in Södertälje, Sweden. Three concentrations of diphenhydramine plus a control were measured for each of the three experiments conducted. The range of concentrations chosen was selected for proof-of-principle rather than environmental relevance (Bartelt-Hunt et al., 2009). For the full table of measured diphenhydramine concentrations, see table 1 in paper IV.
Life-history Traits Effects of contaminant exposure on life history traits of C. riparius were assessed in papers I and III. Toxic effects of contaminants on life-history traits such as the development time and emergence rate of benthic inverte-brates can modify community compositions with ripple effects on ecosystem structure and function (Liess and Von Der Ohe, 2005; Sundbäck et al., 2007; Widenfalk et al., 2008). Pesticides have previously been shown to adversely affect the life-history traits of non-target midges (Forbes and Cold, 2005; Goedkoop et al., 2010; Kahl et al., 1997). However, as of yet, the biological impact of microplastic ingestion on aquatic invertebrates is largely unknown (Wagner et al., 2014).
In paper I, experimental vessels were checked daily for emergence (12 larvae/replicate, 10 replicates/treatment). The emerged adults were sexed and their development time (the no. of days from hatching to emergence as adults) recorded. Then the adults were placed in swarm cages according to treatment where they mated and females oviposted eggs ropes in petri dishes of RSW. The number of eggs per rope was estimated according to the pro-tocol in Benoit et al. (1997) (Benoit et al., 1997). Survival was defined as the number of adults that emerged as a proportion of the number of larvae at the beginning of the experiment.
In paper III, there were 26 replicates (experimental vessels) per treat-ment in Experiment A of which 10 were removed after 10 days in order to assess 10-day larval survival. Experiment B was smaller with 18 replicates per treatment of which 4 were removed after 10 days for the 10-day larval survival assessment. For both experiments, each replicate contained 12 lar-vae in accordance with the recommendation of 2 to 3 cm2 sediment surface area per larva (OECD, 2004). Ten-day larval survival was assessed by sifting through the sediment of each removed experimental vessel and counting the live larvae. Then the remaining experimental vessels were checked daily for emergence. The emerged adults were sexed and their development time (the no. of days from hatching to emergence as adults) recorded. Additionally, the sediment of the experimental vessels was also sifted at the end of the

23
experiments to count how many surviving larvae did not emerge as adults. Therefore the survival endpoint described in paper III includes both larvae and adults. Furthermore a second emergence endpoint was assessed that was the number of emerged adults as a percentage of the total survivors (adults + larvae remaining in the sediment).
Behavioral Assays Toxic inhibition of mobility and normal foraging behaviors of predators can change the life-history and fitness traits of both the predators and their prey with consequences for ecosystem structure and function (Maltby, 1999). Predation and activity behaviors of damselfly larvae were assessed in paper II. All observations were “blind,” coded to ensure impartiality and the num-ber of larvae per treatment was 15-16. After the exposure and recovery peri-od, each larva was observed for 10 min in a 3 L white plastic bucket (diame-ter 15 cm) with 200 mL of water and 150–200 freshly hatched Artemia nau-plii prey. Ten minutes has been shown to be a sufficiently long observation period for assessment toxicity-induced behavioral changes (Debecker et al., 2016; Stoks et al., 2005). All behaviors were scored quantitatively. Scored hunting behaviors were step toward prey, head orientation toward prey, cap-ture of prey and striking at prey (extension of mouthparts without successful capture). Scored general activity behaviors were bouts of swimming and walking.
Temperature preference and righting response of aquatic snails were as-sessed in paper IV. Preference for an optimal thermal environment is a fac-tor that influences the behavior of aquatic ectotherms (Barila et al., 1982; Martin and Huey, 2008; Piasečná et al., 2015) and the ability to migrate in response to climate changes, may be necessary for the survival of ectother-mic populations in habitats close to their thermal maximum (Eliason and Anttila, 2017). After exposure to diphenhydramine (18-21 snails/treatment), snails were placed one per channel at 21 °C in a temperature gradient (min:8 ±1 °C, max: 33± 1°C, 0.5°C/cm) for 24 hours. Snail location in the tempera-ture gradient was recorded once every 3 hours during the light hours of the 16:8 light:dark photoperiod. Using this design, temperature preference was determined by the means of the maximum and the minimal temperatures visited for each treatment as well as the mean overall temperatures from the 24-hour-period for each treatment.
The ability of an organism to right itself after placed on its back can be evaluated as evidence of toxicity affecting neuromuscular function, coordi-nation, orientation and stress response (Fonck et al., 2015; Orr et al., 2007; Schram et al., 2014). Self-righting in snails is also an important behavior for avoiding predation (Manríquez et al., 2013). Previous studies have shown that environmental stressors, including pharmaceuticals, can disrupt self-

24
righting in snails (Fong et al., 2017; Hellou et al., 2009; Schram et al., 2014). Before and after the exposure period, the snails, which have left-coiling shells, were flipped onto their right side with a stick. The time each snail took to completely right with its foot re-attached to the glass surface was recorded three times and the minimum and mean righting times for each treatment were calculated.
Molecular Biomakers Exposure to xenbiotics can induce changes in an organism on the molecular level. Molecular biomarkers are measurable biochemical indicators of sub-cellular effects of toxicity that can supplement the interpretation of observed organismal and population level effects (Hyne and Maher, 2003). The bi-omarkers measured in this thesis are Glutathione-S-transferase, the electron transport system and oxidative damage. Glutathione-S-transferase is a detox-ification enzyme upregulated in the presence of xenobiotics (Kostaropoulos et al., 2001). Measurement of the electron transport system is an estimation of energy consumption (De Coen and Janssen, 2003), and oxidative damage can result from xenobiotic-induced production of reactive oxygen species (Khalil, 2015). In paper II, C. puella larvae were analyzed for glutathione-S-transferase (GST), electron transport system (ETS) and oxidative damage (MDA) at the Catholic University of Leuven in Belgium. Three larvae were combined to provide the material for each sample. The samples were ho-mogenized in a buffer solution (100 mmol/L Tris-HCl, pH 8.5, 15% polyvi-nyl pyrrolidone, 153 lmol/L MgSO4 and 0.2% Triton X-100) which provid-ed supernatant for the analyses. The method for measuring GST activity followed the protocol from McLouglin et al. (2000) with protein content measurements conducted according to Bradford (1976) with results ex-pressed in μmol/min/μg protein (Bradford, 1976; McLoughlin et al., 2000). The ETS assay followed the protocol from Janssens et al. (2015) with results expressed in nmol O2/min/μg protein (Janssens et al., 2015). Oxidative damange was estimated as malondialdehyde formation which is a common biomarker for toxicity-induced lipid peroxidation and expressed in nmol MDA/mg fat. The oxidative damage assay followed the protocol of Janssens and Stoks (2014) for measuring malondialdehyde formation with lipid con-tent calculation according to the protocol of Bligh and Dyer (1959) (E.G.Bligh and W.J.Dyer, 1959; Janssens and Stoks, 2014).
Thermal Tolerance Some xenobiotics are known to affect the range of temperatures aquatic eco-therms can tolerate (Gordon, 2005) with consequences for populations al-

25
ready living in habitats at one end of their thermal range (Eliason and Anttila, 2017). The known capacity for antihistamines to alter maximum thermal tolerance in Drosophila, raises the question of antihistamine effects on the critical thermal maxima of aquatic invertebrates (Hong et al., 2006). In paper IV, P. corneus adults were evaluated for thermal tolerance after exposure to diphenhydramine, see Table 1 for concentrations. After exposure and a brief recovery period, snails were placed individually in glass jars filled with aerated, dechlorinated water. The jars were placed in a water bath at 21°C for a 30 minute acclimatization period followed by an increase in temperature of one degree every 30 minutes. A snail was considered to have reached maximum thermal tolerance when it lost attachment to the glass.
Statistics All statistical analyses were performed in R (R Core Team, 2016).
In paper I, effects of on survival, emergence (the number of larvae that be-came adults) and sex of chironomids were analyzed with binomial general-ized linear mixed effects models with treatment as a fixed effect and repli-cate vessel as a random effect. The treatment model was compared to the null model with one-way ANOVA with a significant difference between the models indicating the presence of an effect of treatment. Where an effect of treatment was found, a Tukey post-hoc test was conducted for pairwise comparisons to determine the effects of specific treatments compared to the controls and each other. Development time was analyzed by linear mixed effects models with treatment as a fixed effect and the replicate vessel as a random effect. The treatment models were compared to the null models with one-way ANOVA with a significant difference between the models indicat-ing the presence of an effect of treatment. The sexes were analyzed separate-ly due to dimorphic development rates.
Since the behavioral variables in paper II might be correlated, a principle component analysis was conducted on the four predation-related behaviors and the two general activity behaviors, which reduced the behaviors to two principal components which were used for further analysis. The effects of treatment on the first two principle components were thereafter analyzed with ANCOVA with larval size set as a covariate, and subsequent pairwise comparison when the ANCOVA showed significant effects of treatments. Effects on glutathione-S-transferase (GST), electron transport system (ETS) and oxidative damage (MDA) were analyzed with ANOVA. Where an effect of treatment was found, a Tukey test for pair-wise comparison was conduct-ed. Additional Dunnett tests were conducted for the first principal compo-nent, ETS and MDA.

26
In paper III, effects on survival, emergence and sex of chironomids were analyzed with binomial generalized linear mixed effects models with treat-ment as a fixed effect and replicate as a random effect. The treatment model was compared to the null model with one-way ANOVA with a significant difference between the models indicating the presence of an effect of treat-ment. Where an effect of treatment was found, a Tukey post-hoc test was conducted for pairwise comparisons to determine the effects of specific treatments compared to the controls and each other. An additional emer-gence analysis was included: the adult percentage of surviving individuals (adults + larvae remaining in the sediment) which was analyzed with one-way ANOVA. Effects of pro-longed exposure were also analyzed with an ANOVA of the generalized linear mixed effects for 10-day survival and survival at the end of the experiment followed by further analysis of the sin-gle esfenvalerate treatment. This further analysis was done with a one-way ANOVA comparing a general linear mixed model of a subset of the com-bined data for Experiment A or Experiment B, respectively, for only two categories (10 day esfenvalerate exposure and 34 day esfenvalerate expo-sure) to the null model. Development time was analyzed by linear mixed effects models with treatment as a fixed effect and the replicate vessel as a random effect. The treatment models were compared to the null models with one-way ANOVA with a significant difference between the models indicat-ing the presence of an effect of treatment. The sexes were analyzed separate-ly due to dimorphic development rates. Female development time was not analyzed in Experiment A due to low female emergence.
In paper IV, the effects of diphenhydramine on temperature preference, critical thermal maximum and righting time were assessed with one-way ANOVAs. Where a significant effect was found, a Jonckheere-Terpstra post-hoc test was conducted with manual step-down application to take into ac-count the apparent dose-response trend.

27
Results and Discussion
Combination effects of pyrethroids and neonicotinoids (Paper I) Larvae of C. riparius were exposed to a 1-h pulse of esfevalerate, deltame-thrin, imidacloprid and thiacloprid (singularly, in pairs and a mixture of all four) and evaluated for effects on life-history traits (Table 1).
Figure 1. Survival, defined as the proportion of C. riparius larvae that emerged within 28 days, after a 1 h pulse exposure to two pyrethroid and two neonicotinoid pesticides (for concentrations see Table 1). The letters above the bars denote signifi-cant differences according to a Tukey post-hoc test for pairwise comparison. Error bars denote standard error.
Single exposure to the pyrethroid, esvenvalerate, and to the neonicotinoid, thiacloprid, decreased survival from 94% in the control to 79% (Tukey, p < 0.03) and 68% (Tukey, p < 0.0001), respectively (Fig. 1). Exposure to a four-pesticide cocktail also decreased survival to 72% (Tukey, p < 0.001) (Fig. 1). However, neither the combined exposure to the two pyrethroids nor

28
the two neonicotinoids decreased survival compared to the control (Fig. 1). Adverse effects on survival may be due to starvation caused by pesticide-induced anorexia. Both pyrethroids and neonicotinoids are known to induce neurotoxic effects on mobility that lead to feeding inhibition (Landrum et al., 2002; Nyman et al., 2013).
These results indicate that while a 1-h pulse exposure to environmentally relevant concentrations of pesticides can impact survival, there does not appear to be a risk for additive or synergistic effects with combined exposure to these selected pesticides. However, the greater adverse impact of esfen-valerate and thiacloprid single exposures on survival compared to when combined with the other neonicotinoid, imidacloprid, or pyrethroid, deltame-thrin, raises the possibility of antagonism. Mixture antagonism of pyre-throids has previously found in effects on survival of Chironomus dilutes, although the mechanism for the antagonism remains unknown (Hasenbein et al., 2015). Differing sensitivity to different neonicotinoids is attributed to differences in metabolic capacity rather than target site specificity; that is to say that some species are able to metabolize some neonicotinoids better than others and then excrete them before they act on neurological targets (Manjon et al., 2018). Antagonistic effects of pesticide combinations have previously been shown to result from one of the compounds inducing the upregulation of metabolic enzymes more effectively than the other resulting in improved metabolism and excretion of both compounds (Anderson and Zhu, 2004). Xenobiotic chemicals go through three phases of metabolism and the phase I functionalized metabolites can also be toxic or even more toxic than the par-ent compound (Macherey and Dansette, 2015; Simon-Delso et al., 2015), If a pesticide’s metabolites are more toxic in C. riparius compared to the parent compound, then another possible mechanism of antagonism could be the occupation of metabolic enzymes by the additional xenobiotics in the mix-ture resulting in a reduction in the rate of the compound’s biotransformation to its more potent metabolites. Elucidation of this mechanism of antagonism is worth further investigation.

29
Figure 2. Development time, defined as the number of days from hatching to adult emergence, after a 1 h pulse exposure to two pyrethroid and two neonicotinoid pes-ticides (for concentrations see table 1). Fitted values provided by a generalized mixed effects model. The letters above the bars denote significant differences within each sex according to a Tukey post-hoc test for pairwise comparison. Error bars denote standard error.
Evidence of antagonism is also present in the effects of the pyrethroids on development time. The esfenvalerate single exposure delayed female devel-opment time by 1.6 days from the 23.7 ± 0.3 days observed in the control to 25.3 ± 0.4 days (Tukey, p < 0.001). In contrast, development time in the combined pyrethroid exposure was 24.8 ± 0.4 days and the pyrethroid-neonicotinoid cocktail was 23.1 ± 0.4 days; neither of which were statisti-cally different from the control (Tukey, p = 0.109, p = 0.703) (Fig. 2). How-ever, male development time was only affected by the combined pyrethroid exposure, which resulted in a 1.3 day delay from the 19.8 ± 0.3 days ob-served in the control to 21.1 ± 0.4 days (Tukey, p < 0.001) (Fig.2). The ne-onicotinoids had no effects on development time for females or males (Tuk-ey, all p > 0.05). These findings suggest that pyrethroid toxicity may have greater consequences for larval development compared to neonicotinoids. This finding is in keeping with results reported by Nicholls et al. (2017) and Stoughton et al. (2008) which found no effect of low neonicotinoid exposure on the development rate of the bee, Osmia bicornis or the midge, Chirono-mus tentas, respectively (Nicholls et al., 2017; Stoughton et al., 2008). Alt-hough higher concentrations of imidacloprid did delay development in C. tentas (Stoughton et al., 2008). Since, the neonicotinoids did decrease sur-vival there is the possibility that decreased population density may have

30
influenced the development rate by increasing per capita food availability and reduced competition (Ristola et al., 2009). The literature is currently inconclusive with regard to whether pyrethroids delay or hasten development rates in midges, with studies providing evidence of both (Forbes and Cold, 2005; Samsøe-Petersen et al., 2001). The faster development time in the cocktail exposure (D+E+I+T) compared to the pyrethroid exposures suggests an antagonism between the pyrethroids and neonicotinoid that has not been previously reported.
No effects of the pesticides alone or in combination were found on female C. riparius fecundity in terms of the number of eggs per egg rope (ANOVA, F8,233. = 1.66, p > 0.1). Findings from Forbes and Cold (2005) and Shi et al. (2011) similarly showed no effect of neonicotinoids on fecundity in midges or aphids (Forbes and Cold, 2005; Shi et al., 2011). Pyrethroids, on the other hand, have been suggested to indirectly reduce fecundity in midges (Goedkoop et al., 2010) and shown to directly reduce fecundity in two cope-pods (Chandler, 1990). Egg viability and success of the subsequent genera-tion would be prudent to investigate as well before conclusively ruling out adverse effects of pyrethroid and neonicotinoid exposure on C. riparius fit-ness.
Single and mixture impacts of two pyrethroids (Paper II) Larvae of C. puella were exposed for 14 h to esfenvalerate and deltamethrin (singularly and combined) and evaluated for effects on predation behaviors, general activity and three molecular biomarkers (see Table 1).

31
Figure 3. The effects of a 14 h sub-lethal pyrethroid exposure on the mean predation behaviors (PC1) and general activity (PC2) of damselfly larvae expressed as per-centages relative to the control values which were set as equivalent to 100%. The letters above the bars denote significant differences according to a Tukey post-hoc test for pairwise comparison (n = 14-15; Tukey, p < 0.05). Error bars denote stand-ard error.

32
The first two principal components (PC1 and PC2) accounted for 84% of the variation in the behavioral data. PC1 was primarily explained by the four predation-related behaviors (52%) and PC2 by the two general activity be-haviors (29%), see table 1 for the scored behaviors. Pyrethroid exposure markedly decreased predatory behaviors (PC1, ANOVA, F5,84 = 7.121, p < 0.001, Fig. 3). Compared to the amount of predation behaviors observed in control larvae, the single exposures to the two concentrations of deltamethrin and the combined exposure to both pyrethroids resulted in decreases of 75 ± 3% (p = 0.001), 67 ± 2% (p = 0.004) and 88% ± 2% (p < 0.0001), respec-tively. The single esfenvalerate exposures did not result in reduced predatory behaviors, supporting other findings that have shown esfevenalerate to be less potent compared to deltamethrin in inhibiting feeding (Ishaaya et al., 1983). The pyrethroids had no effect on the general activity of the damselfly larvae (PC2, ANOVA, F5,84 = 2.117, p = 0.071; Fig. 3) indicating that the pyrethroid-induced inhibition of normal feeding behavior observed in other invertebrates as well (Ishaaya et al., 1983; Rasmussen et al., 2013) is a toxic effect separate from impaired mobility and possibly more sensitive. This is supported by other studies that found pyrethroid-induced feeding inhibition coinciding with induction of hyperactivity (Huang et al., 2014; Rasmussen et al., 2013; Viran et al., 2003). While we did not find hyperactivity per se, the damselfly larvae exposed to the highest concentration of esfenvalerate were significantly more active than those exposed to the same concentration of deltamethrin (Tukey, p =0.037; Fig. 3). However, the behavioral assay was designed to quantify changes in mobility that would be symptomatic of tem-porary paralysis (knockdown) commonly seen in terrestrial invertebrates exposed to pyrethroids (Baatrup and Bayley, 1993; Kumar et al., 2011) and therefore duration of locomotive activities were not recorded (e.g. two bouts of walking were scored equally regardless of the amount of time spent walk-ing per bout) which means the possibility of esfenvalerate-induced hyperac-tivity cannot be ruled out.

33
Figure 4. The effects of a 14 h sub-lethal pyrethroid exposure on activity of gluta-thione S-transferase (GST), oxidative damage to lipids (MDA) and activity of the electron transport system (ETS) in damselfly larvae, expressed as percentages rela-tive to the control values which were set as equivalent to 100%. The letters above the bars denote significant differences of GST activity (n = 5-10; Tukey, p < 0.05); the * denotes a significant difference for MDA (Dunnett, p = 0.0415). No significant differences in ETS were found (ANOVA, p > 0.05; all Dunnett p values > 0.05). Error bars denote standard error.
Deltamethrin’s relative potency compared to esfenvalerate may be explained by less efficient metabolism of deltamethrin in C. puella. Only the two sin-gle esfevalerate exposures and the lower deltamethrin concentration expo-sure induced production of glutathione S-transferase (GST) by 52 ± 11%, 45 ± 7% and 42 ± 8% compared to the control (Tukey, p = 0.004, p = 0.018, and p = 0.052). GST induction in these three exposure categories was also significantly higher compared to the combination exposure (Tukey, p = 0.002, p = 0.006, and p = 0.017; Fig. 4). GST is an enzyme known to metab-olize pyrethroids in invertebrates albeit with significant variation between even closely related species (Ganesh et al., 2003; Gowland et al., 2002; Rodrigues et al., 2015b; Rumpf et al., 1997). The lack of GST induction after combined exposure to both pyrethroids suggests a toxic effect on the detoxification pathways themselves. Compromised metabolism of the pyre-throids would result in increased toxicity which is evident in the 88% de-crease in predation behaviors compared to a non-significant effect on preda-tion behaviors when GST was induced (Fig. 3).

34
The lowest, single exposure to esfenvalerate also increased oxidative damage to lipids (MDA, Dunnett, p = 0.042) whereas the other single expo-sures and combined exposure did not (all p values > 0.05; Fig. 4). Although the power of this experimental design was not strong enough to detect signif-icant changes in oxidative damage in all of the single exposure groups, the data does suggest that deltamethrin may also induce oxidative damage to lipids but not when combined with esfenvalerate (Fig. 4). This in contrast to an in-vitro study of pyrethroid-induced oxidative stress in human cell lines which showed dose-additivity (Romero et al., 2015). A study on fish sup-ports deltamethrin-induced oxidative stress and suggests that deltamethrin-induced reductions in activity are a result of resource re-allocation to detoxi-fication and antioxidant protection (Oliveira et al., 2012). Another fish study also showed deltamethrin-induced oxidative stress and antioxidant defense mechanisms but also significant variation between tissue-types (Sayeed et al., 2003). The absence of a combined effect on MDA may be connected to the unknown mechanism that prevented GST induction in the combination exposure, or possibly the combination exposure triggered an antioxidant mechanism not measured. Since we only measured oxidative damage to lipids, further research measuring damage to other tissues such as proteins or DNA, should be conducted before ruling out the possibility of oxidative damage induced by combined exposure.
Metabolizing xenobiotics and protecting against oxidative damage are costly processes that have shown to coincide with reduced growth and re-productive output (Maltby, 1999), and therefore it is interesting to estimate energy consumption. An organism’s energy consumption can be estimated with an assay that measures the activity of electron transport system (ETS) (De Coen and Janssen, 2003). Despite the evidence of GST induction and oxidative damage, we found no effect of pyrethroid exposure on ETS activi-ty (ANOVA, F5,34 = 0.502, p = 0.773; all Dunnett comparison p values > 0.50; Fig. 4).
Combined effects of microplastics and a pyrethroid (Paper III) Larvae of C. riparius were reared in sediment spiked with latex-based poly-styrene microspheres and esfenvalerate, singularly and in combination (Ta-ble 1). The experiment was conducted twice, once under low food conditions (Experiment A) and once under normal conditions (Experiment B). Effects of chronic exposure on life-history traits were evaluated. The results of the two experiments are not compared directly with each other as they were conducted at different times and with some methodological differences.

35
Figure 5. The effects of a chronic spiked-sediment exposure to two sizes of micro-plastic beads (MP: 1.0 µm and 10.0 µm) and esfenvalerate on the emergence of C. riparius larvae after (a) 34 days in low nutrient sediment and (b) 28 days in high nutrient sediment. Emergence is expressed as (a, b) percentage of the number of larvae at the start of each experiment and (a', b') adult percentage of the surviving individuals. The letters above the bars denote significant differences according to a Tukey post-hoc test for pairwise comparison (n = 26; Tukey, p < 0.05). Error bars denote standard error. No effects on emergence were found in the high nutrient sed-iment (n = 18; ANOVA, χ2 = 4.132, Df = 4, p = 0.389).

36
Figure 6. The effects of a chronic spiked-sediment exposure to two sizes of micro-plastic beads (1.0 µm and 10.0 µm) and esfenvalerate on the survival of C. riparius larvae after 10 days (white bars) and combined larvae and emerged adults (shaded bars) after (a) 34 days in low nutrient sediment and (b) 28 days in high nutrient sed-iment. Star (*) denotes a significant difference in 34-day survival between treat-ments exposed in low nutrient sediment (Tukey, p = 0.017). Circle (¤) denotes a significant difference between 10 day larval survival and 34 day survival after expo-sure to esfenvalerate only in the low nutrient sediment (a: ANOVA, Df = 1, χ2 = 4.69, p = 0.030, b: ANOVA, Df = 1, χ2 = 3.19, p = 0.074). Error bars denote stand-ard error.
Microplastics and esfenvalerate had no effects on emergence, survival, de-velopment time, or sex of C. riparius in high nutrient sediment. Mean emer-gence as a percent of the number of larvae in each treatment at the beginning of the experiment ranged from 44.5% to 48.2% in the contaminant treat-

37
ments compared to 58.3% in the control (ANOVA, χ2 = 4.132, Df = 4, p = 0.389, Fig.5b). Mean emergence as a percent of the number of surviving individuals (adults + larvae) after 34 days ranged from 88.1% to 98.6% in the contaminant treatments compared to 89.6% in the control (ANOVA: F4,40 = 0.639, p = 0.638, Fig. 5b'). This finding is in line with a previous spiked-water exposure which also found no effect on chironomid emergence at a similarly low concentration of esfenvalerate (Rodrigues et al., 2015a). Mean larval survival at 10 days ranged from 45.8% to 77.1% across the treatments (ANOVA, χ2 = 5.611, Df = 4, p = 0.230, Fig. 6b).Mean survival at 28 days ranged from 46.5% to 64.3% across treatments (ANOVA, χ2 = 4.132, Df = 4, p = 0.389). Mean development time ranged from 20.2 to 22.7 days for males and 22.6 to 24.9 days for females with no significant effects of treatment detected (males: ANOVA χ2= 3.8878, Df = 4, p = 0.4214; females: ANOVA χ2= 4.168, Df = 4, p = 0.3837). This finding is in contrast to other pyrethroid exposure studies which found delays in female development time including Paper I (Maul et al., 2008; Rodrigues et al., 2015a). The percentage of males emerged ranged from 39.9% to 57.8% in the contaminant treatments com-pared to 57.7% in the control and no significant effect of treatment on sex was found (ANOVA, χ2= 5.608, Df = 4, p = 0.230). Previous studies on ef-fects of pesticides, including esfenvalerate, have found skewed sex ratios as a result of exposure (Agra and Soares, 2009; Rodrigues et al., 2015a). Male midges are thought to be more susceptible to pyrethroid toxicity due to their more rapid development (Goedkoop et al., 2010). Our findings, however, suggest no such male-specific toxicity.
Effects of the contaminants on life-history traits were found when food was insufficient providing an impression of how these contaminants can affect benthic larvae under sub-optimal conditions. The artificial sediment across all treatments including the control contained both organic and inor-ganic natural particulate matter < 1 mm in size ensuring that any affects ob-served in the microplastic spiked-sediments are not merely effects of expo-sure to small particulate matter. Mean emergence as a percent of the number of larvae in each treatment at the beginning of the experiment ranged from 3.4% to 5.9% in the contaminant treatments compared to 16.8% in the con-trol (ANOVA, χ2 = 29.957, Df = 4, p > 0.001, Fig.5a). Mean emergence as a percent of the number of surviving individuals (adults + larvae) after 34 days ranged from 4.9% to 12.2% in the contaminant treatments compared to 25.7% in the control (ANOVA F4,82 = 2.463, p =0.051, Fig. 5a'). Esfen-valerate, 1.0 µm microplastics and 10 µm microplastics, and a combination of esfevalerate and 1.0 µm microplastics all significantly decreased emer-gence compared to the control treatment (Tukey, all p values < 0.05, Fig. 5a and 5a'). This indicates that exposure to microplastics is stressful to the lar-vae with an energy cost similar to exposure to a low concentration of a pyre-throid. Ingestion of microplastics has previously been shown to increase

38
oxidative stress in sandworms and induce dietary constraints in amphipods (Browne et al., 2013; Straub et al., 2017).
Mean larval survival in the low nutrient sediment at 10 days ranged from 70.0% to 84.2% across the treatments (ANOVA, χ2 = 2.368, Df = 4, p = 0.669, Fig. 6a). Mean survival at 34 days ranged from 62.7% to 79.9% across treatments (ANOVA, χ2 = 11, Df = 4, p = 0.0266). Survival at 34 days was significantly lower in the 10.0 µm microplastic treatment (67.6%) com-pared to the 1.0 µm microplastic treatment (79.9%) (Tukey, p = 0.017). Esfenvalerate significantly decreased larval survival by 13.7% over time when food was insufficient, from 84.2% at 10 days to 70.5% at 34 days (ANOVA, Df = 1, χ2 = 4.69, p = 0.030), but not when combined with 10.0 µm microplastics which saw a decrease in survival over time of only 7.5% from 76.9% at 10 days to 69.4% at 34 days (Fig. 6a). Detoxification of pesti-cides is an energetically costly process (Rodrigues et al., 2015b). It is rea-sonable then that chronic exposure to a sub-lethal concentration of a pesti-cide can decrease survival if the organisms are experiencing a nutrition defi-cit. The absence of a similar effect in the combination treatment suggests that the microplastics may have adsorbed the esfenvalerate, decreasing its bioavailability. This microplastic “cleaning” effect has been previously re-ported in experimental and theoretical studies (Koelmans et al., 2013; Oliveira et al., 2013). Mean male development time ranged from 23.3 in the control up to 27 days in the 10.0 µm microplastics treatment, however no effect of treatment was detected (ANOVA χ2= 3.612, Df= 4, p= 0.4611). Low emergence rates in the low nutrient sediment experiment affected the power to detect differences in male emergence, and female emergence rates were too low to allow for any analysis of development time. The percentage of males emerged ranged from 81.3% to 100% in the contaminant treatments compared to 87.6% in the control and no effect of treatment on sex was de-tected (Df = 4, χ2 = 5.608, p = 0.230). The high survival but low emergence in low nutrient conditions highlights the importance of sufficient food re-sources for normal development, and female development in particular due to the higher energy demands of egg production (Goedkoop et al., 2010).
Effects of an antihistamine (Paper IV) Adult P. corneus were exposed for 24 h to three concentrations of the anti-histamine, diphenhydramine, and then evaluated for effects on temperature preference (Tpref), critical thermal maximum (CTmax) and righting time (RT).

39
Figure 7. Mean critical thermal maximum (CTmax °C) of P. corneus after 24 hours exposure to diphenhydramine. The p-value denotes a significant effect of diphenhy-dramine compared to the control (Jonchkheere-Terpstra; n = 21-23 snails per treat-ment). The error bars denote standard error.
The CTmax of P. corneus was determined to be 37.7 °C. Thermal tolerance increased after exposure to diphenhydramine concentrations of 9.58, 97.0 and 949 µg/L by 0.1, 0.7 and 2.2 °C respectively (Fig. 7). The increase in CTmax found in the highest concentration exposure was significant (Jonck-heere-Terpstra, p = 0.003, n = 21-23 snails per treatment). An increase in thermal tolerance after exposure to a xenobiotic rather than a decrease is an unusual finding in an aquatic ectotherm. However, thermal tolerance is not a commonly assessed endpoint and the available data are on fish, not inverte-brates (Gordon, 2005). To our knowledge, this is the first study to investigate the effects of a pharmaceutical on thermal tolerance in an invertebrate. A study on the effects of anti-depressants in snails did find adverse effects on pedal neurons which manifested as increased foot detachment from the sub-strate (Ford and Fong, 2016), however all of the snails in our study attached to the glass substrate after exposure to diphenhydramine without any signs of toxicity in that regard. Since significant increases in thermal tolerance were not found at the lower, environmentally relevant concentrations for freshwa-ters and sediments (Bartelt-Hunt et al., 2009; Ferrer et al., 2004), it is unlike-ly that diphenhydramine in the environment has a significant impact on thermal tolerance of wild P. corneus populations. However, even though these results are not alarming, they do indicate that further investigation into the effects of antihistamines on thermal tolerance in aquatic ectotherms is warranted.
40
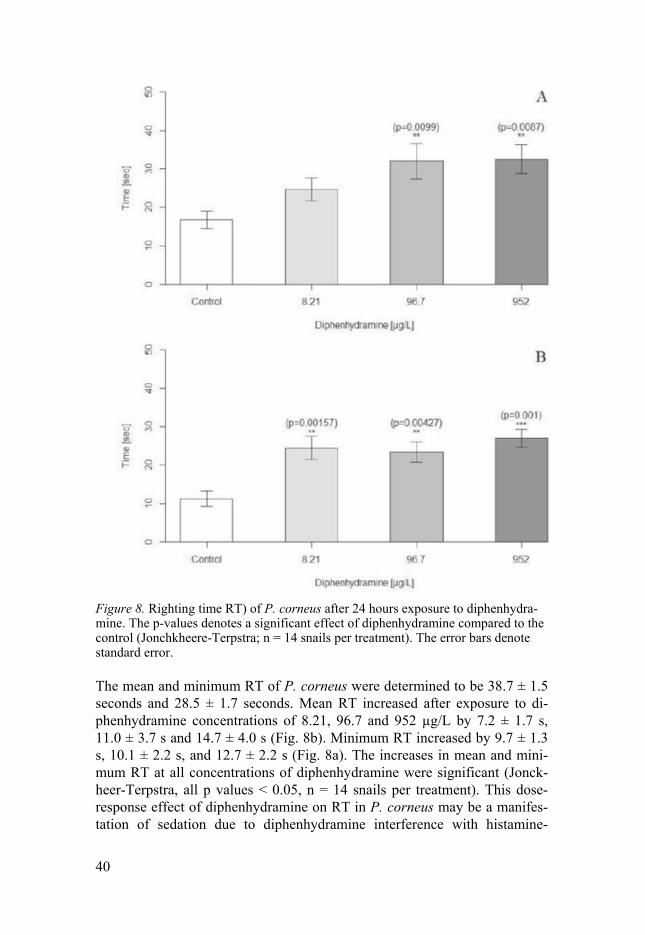
Figure 8. Righting time RT) of P. corneus after 24 hours exposure to diphenhydra-mine. The p-values denotes a significant effect of diphenhydramine compared to the control (Jonchkheere-Terpstra; n = 14 snails per treatment). The error bars denote standard error.
The mean and minimum RT of P. corneus were determined to be 38.7 ± 1.5 seconds and 28.5 ± 1.7 seconds. Mean RT increased after exposure to di-phenhydramine concentrations of 8.21, 96.7 and 952 µg/L by 7.2 ± 1.7 s, 11.0 ± 3.7 s and 14.7 ± 4.0 s (Fig. 8b). Minimum RT increased by 9.7 ± 1.3 s, 10.1 ± 2.2 s, and 12.7 ± 2.2 s (Fig. 8a). The increases in mean and mini-mum RT at all concentrations of diphenhydramine were significant (Jonck-heer-Terpstra, all p values < 0.05, n = 14 snails per treatment). This dose-response effect of diphenhydramine on RT in P. corneus may be a manifes-tation of sedation due to diphenhydramine interference with histamine-

41
regulated sleep-wake cycles (Green, 2008). Histamine-signaling genes with thermoregulatory functions identified in Drosophila melanogaster include the hisCl1 gene (Hong et al., 2006). As the hisCl1 gene is also expressed in circadian clock neurons (Díaz et al., 2006) there may be a mechanistic link between the observed changes in thermal tolerance and righting time in P. corneus. The anti-depressant, fluoxetine, which also has antihistamingeric properties, has also been shown to increase RT in snails at even lower con-centrations and shorter exposure periods (Fong et al., 2017). Since we found effects at the lowest concentration of diphenhydramine tested, we recom-mend further testing to determine the lowest observed effect concentration.
The Tpref for P. corneus was determined to be 21 ° C. The Tpref did not change after diphenhydramine exposure. Toxicity could have manifested as preference for a lower or higher temperature or as absence of preference (a wider range), effects which were found in Drosophila after exposure to anti-histamines (Hong et al., 2006). However, our findings offer no indication that histaminergic pathways play a role in mediating temperature preference in P. corneus.

42
Concluding Perspectives
In conclusion, this thesis provides evidence that sub-lethal exposures to sin-gle and combined anthropogenic contaminants can have adverse effects on aquatic invertebrates. Paper I showed that a brief water exposure to envi-ronmentally relevant concentrations of neonicotinoids and pyrethroids can have lasting adverse effects on the life-history traits of a benthic inverte-brate. This short 1 h exposure to pesticides decreased survival and delayed development of larvae of the midge, C. riparius. The combination of neon-icotinoids and pyrethroids did not produce synergistic effects; however, there was some indication of antagonism. Subsequently, Paper II showed that a water exposure to environmentally relevant concentrations of pyre-throids can adversely affect the predatory ability and detoxification pathways of a benthic invertebrate. A 14 h exposure to deltamethrin reduced predatory ability in the larvae of the damselfly, C. puella, and combined exposure with esfenvalerate may have additive toxicity on predation behavior. Additional-ly, single exposure to each pyrethroid induced production of Glutathione S-transferase while combined exposure did not. Then Paper III showed that exposure to 1.0 and 10.0 µm polystyrene-based latex microbeads and envi-ronmentally relevant concentration of esfenvalerate in sediment can have adverse effects on life-history traits of a benthic invertebrate under low nu-trient conditions. Exposure to both sizes of microplastics and esfenvalerate led to equally decreased emergence when food was limited. Additionally, exposure to esfenvalerate resulted in decreased survival when food was lim-ited but not when microplastics were co-present. Finally, Paper IV showed that a water exposure to an antihistamine can alter thermal tolerance and righting behavior in an aquatic gastropod. The 24 h exposure to diphenhy-dramine increased thermal tolerance and delayed righting in the freshwater snail, P. corneus.
Reduced larval survival, delayed development, decreased emergence, in-hibited predation behavior, induction and inhibition of detoxification en-zymes, increased thermal tolerance and delayed righting time are all adverse effects with the potential to affect population dynamics of aquatic inverte-brate communities. For example, inhibition of predation behavior and growth rate might affect fecundity in an organism which ultimately might affect their abundance. Two of the three species investigated have a terrestri-al adult stage with important roles to play in terrestrial community structures as well. Most of these effects occurred at environmentally relevant concen-

43
trations indicating that current contamination of freshwaters is high enough to affect invertebrate populations in nature and consequently the biodiversity of freshwater ecosystems as well as the adjacent terrestrial ecosystems.
Non-negligible effects of exposure to pyrethroids and neonicotinoids on non-target organisms after exceptionally short, single exposure periods are important findings for informing the appropriate risk assessment of pesti-cides in the natural environment where exposure to aquatic biota happens frequently and the spikes in concentrations can last longer. In the environ-ment, mixture exposure is the norm and the resulting effects are difficult to predict. Mixture toxicity is complicated and depends on largely unknown interactions with the detoxification pathways unique to each species. When sub-lethal effects are rarely tested and mixture toxicity is difficult to predict, regulation of pesticides based on “no observed effect concentrations” are likely to be overly optimistic. While further studies investigating the combi-nation effects of pesticides on sub-lethal endpoints would help refine the risk assessment of commercial pesticides, it may be more prudent to investigate solutions that would indirectly reduce the need for synthetic plant protection products. For example, while pesticides are largely understood as a solution to the problem of feeding the human population, the current rate of food waste is another variable on the same side of that equation. Similarly, rec-ommending further research that would ultimately support tighter pesticide regulations is one approach to reducing their use in ornamental gardens, but encouraging the public to choose native plant species adapted to the local pests may also be an effective route to the same goal. And finally, advances in CRISPR-based genomic tools with great potential to reduce malaria vec-tors, could replace the widespread use of high concentrations of pyrethroids and neonicotinoids in regions vulnerable to malaria.
The evidence is growing that plastic pollution, once considered harmless and biologically inert, is in fact a threat to wildlife. Ingestion of microplas-tics carries a subtle cost to the organism that can be amplified under stress. The silver-lining is that microplastics have the potential to act as cleaning agents reducing the bioavailability of co-occurring chemical contaminants. More evidence is coming out that microplastic pollution is primarily fibers and fragments. This highlights the need for further investigation into the biological effects of those shapes in addition to the bead shape commonly found in personal care products and industrial production. Further research on microplastic ingestion in other benthic invertebrates as well is recom-mended for a robust risk assessment of microplastic pollution in the benthos. Efforts to dramatically reduce the input of plastics to aquatic ecosystems are already underway however, further research is very necessary to appropriate-ly guide these efforts. A world-wide ban on plastic microbeads in personal care products, for example, while well-intended, would be an inefficient step toward improved aquatic ecosystem health if detrimental effects on wildlife are primarily caused by fibers from synthetic clothing and fishing nets.

44
This thesis adds to the growing body of evidence that pharmaceuticals in municipal waste water effluent are a risk to the health and well-being of aquatic animals. Further investigations into how different pharmacological compounds affect different aquatic organisms would be of substantial benefit to a prioritization process that could provide direction for regulatory efforts and the development of new water treatment technologies. These investiga-tions should explore non-traditional endpoints based on the compounds’ bio-active targets and physiology of the test species. Additionally, research relat-ed to climate-change-related multiple stressor scenarios, should look not only at how temperature stress affects the toxicity of xenobiotics, but also how xenobiotics affect an organism’s ability to respond to temperature fluc-tuations. Finally, the toxicity of pharmaceutical pollution is important to take into account in scientific evaluations of the capacity of aquatic ecosystems to adjust to a rapidly changing climate.
We have come a long way since Rachel Carson’s publication of “Silent Spring” in 1962 which first criticized the mantra that “the solution to pollu-tion is dilution” and spawned the entire discipline of ecotoxicology. Despite ingeniously inventing new contaminants as quickly as we ban the old ones, there is world-wide consensus now that freshwater is precious and freshwa-ter ecosystems should be conserved as much as possible. It is my hope that this thesis has contributed in some small ways to increasing what is possible.

45
Lay Summary
“Save the bees!”
“Beat the bead!”
“The fish are on drugs!”
The media have been doing their part to raise awareness about the dangers of pesticides, microplastics and pharmaceuticals in the environment. No one with a Twitter account hasn’t heard that pesticides are causing bee colony collapse, seen a pic of a sea turtle eating a plastic bag or downloaded an app to find out if their toothpaste had microplastics, or been made to feel a twinge of guilt that their birth control pills and anti-depressants are poison-ing the oceans. Environmentalism is seldom subtle. Environmental science, on the other hand, cares a great deal about subtlety. Science wants to know, what exactly is going on?
Throughout its first few decades in the latter half of the 20th century, the new discipline of ecotoxicology, sought to quantify exactly how much of a human-made chemical we could safely release into the environment without killing everything there. Traditional ecotoxicology studies took a handful of non-target species which would stand-in for everything people weren’t try-ing to kill on purpose and exposed them to increasing amounts of a single chemical and recorded how many died. Later came reproductive studies and chronic exposure studies and eventually tests on a wider variety of species along with concern about mixture interactions and a push to acknowledge the relevance of non-lethal effects as well. Until the 21st century, the contam-inants on an ecotoxicologist’s radar were mostly heavy metals from the in-dustrial era and chemical weapons – those used against fellow humans as well as those used against the natural world, creatures that seemed hell-bent on eating our crops and taking up residence in our carpets. Then came the first reports of garbage patches in the oceans, whale stomachs full of trash and the realization that we piss out a good amount about of all the drugs we take. Ecotoxicology began to feel a bit more like veterinary medicine, the wildlife equivalents of “what happens when Fluffy eats her plastic ball?” and “will Fluffy die if I give her an aspirin?” and as more monitoring surveys reported on environmental conditions, the more ominous the questions be-came, like “what happens if Fluffy eats a plastic ball everyday of her life?”

46
and “what happens to Fluffy if she eats a tiny bit of aspirin, birth control pills, anti-depressants, cancer drugs and allergy medications….everyday?” At the same time, we’re still using heavy metals in industry and we’re still deploying chemical weapons on a daily basis. Despite increased regulation, those concerns have not faded away.
This thesis enters a context in which humanity’s trash -- chemical, physi-cal and pharmacological—is still regularly ending up in lakes and rivers all around the world. The burden is on the ecotoxicologists to prove whether these substances do or do not cause harm to those fragile ecosystems. It is up to us to decide what kind of harm to look for and how loud the alarm should be sounded, if at all. There are still many who loudly proclaim that plastic is inert and therefore harmless, and that the solution to pollution is dilution. Are those assertions true? This thesis looks at what happens when ecologi-cally beneficial, non-biting midges, damselflies and snails are exposed in the lab to contaminants that they may be exposed to in nature.
The first study of this thesis was a simulation of a storm event. The con-centrations of pesticides in rivers and lakes peak during the first hour of a heavy rainstorm. This is because the rain hits plants and soils that have been sprayed and then the run-off carries away some of those pesticides down storm drains or drainage ditches where it eventually ends up in the rivers and the lakes. When non-biting midges hatch from their eggs, they are tiny little red worm-like larvae less than 1 millimeter long. They grow over a period of a few weeks until they briefly form pupae before transforming into their flying, adult form. They mate in swarms and will die a natural death within a few weeks if nothing kills them (to eat for the most part, but perhaps also out of annoyance). This short life-span makes it easier to study the effects of contaminant exposure over an entire life, compared to other species that live longer. When the midge larvae were only 2-days-old, we squirted tiny amounts of four different kinds of really common pesticides (imidacloprid, thiacloprid, deltamethrin and esfenvalerate) from two major pesticide types (neonicotinoids and pyrethroids) into the water. The amounts used were within the range that has been reported to be present in freshwaters in nature. Some larvae were only exposed to one pesticide, some got a combination of two and the rest were exposed to all four. After an hour, we poured off that pesticide contaminated water and gave them fresh clean water. Then we kept track of how many made it to adulthood and how long it took them.
We found that this short 1-hour-long exposure to concentrations of pesti-cides similar to what is found in nature, killed some of the midge larvae and delayed the pace of development to adulthood for some others. The pesti-cides can interfere with the larvae’s nerves making it difficult for them to move for a while. If they can’t move, they can’t eat, and that initial loss of nutrition could weaken them to the point that they can’t recover at all and they starve to death. It could also mean they need more time than usual to grow before their bodies are ready for transformation. The good news is that

47
the combination of exposure to all four pesticides together wasn’t worse than the exposure to only one. In fact, some combinations weren’t as bad as expo-sure to only one. Why would that happen? There are specialized detoxifica-tion systems inside all animals that are responsible for handling toxins that enter the body. Sometimes these systems make a toxin worse before they make it better. This isn’t on purpose, it’s because natural detoxification sys-tems have not had the chance to evolve to handle the new toxins that we create. Although, adaption is happening and we call that “developing re-sistance.” In the case where a toxin gets turned into something worse before it gets turned into something better, then that whole process is slowed down if the system is handling another toxin at the same time. Additionally, some toxins are more readily recognized than others and trigger an amping up of the detoxification system (the animal is therefore “more resistant” to these toxins). Greater resistance to one toxin can lead to greater resistance to both toxins when they are both present. One toxin that isn’t immediately and effi-ciently recognized as such by the detoxification system has more time to inflict toxic damage to the animal then if a more recognizable toxin was present at the same time to trigger the system. Once the system is kicked into gear, both toxins may then be eliminated more efficiently.
The second study of the thesis looked at how tiny amounts of two pesti-cides can affect behavior. Damselflies, closely related to dragonflies, are brightly-colored, winged insects usually found around the banks of ponds and streams. They deposit their eggs into the stems of marshy plants where the offspring hatch into wingless larvae with six legs and a tail that live un-derwater on the muddy bottom for one to two years before transforming into their air-breathing, flying adult form to mate. Both the larvae and adults are adept predators feeding primarily on smaller insects and crustaceans. We hatched damselfly larvae in the laboratory where we then exposed them for 14 hours to either a high concentration or a low concentration of two differ-ent pyrethroid pesticides (esfenvalerate and deltamethrin) or a combination of both pesticides. Then we watched each individual larva for 10 minutes in a bucket filled with freshly hatched crustacean prey and counted how many times the larva performed different actions. The actions were grouped into two categories: those associated with hunting and those associated with gen-eral movement. We then euthanized the larvae and ran tests on their bodies for molecular signs of toxicity. We looked for increased oxidative damage to fat molecules, increased energy use and increased detoxification activity.
We found that the damselfly larvae that were exposed to deltamethrin didn’t hunt as much as the ones that weren’t exposed to the pesticides. It wasn’t just that the larvae tried to hunt but weren’t able to. They didn’t even try very much. The larvae that were exposed to both deltamethrin and esfen-valerate seemed to try even less, but we weren’t able to observe enough lar-vae to say with certainty that both pesticides at the same time were worse than just deltamethrin. Why weren’t they hunting? The most obvious answer

48
would be temporary paralysis. Both of these pesticides can cause that in insects; that’s how they function as insecticides. However, the larvae walked and swam like normal so general paralysis couldn’t be the answer. Other studies have also showed that exposure to pyrethroids can make insects stop eating. Whatever the mechanism is, it’s not good news for wild populations of damselfly larvae or their food webs. When predators stop eating and their populations shrink, there can be ripple effects throughout the ecosystem. Interestingly, we found that each pesticide by itself triggered the damselfly detoxification system in terms of upregulating the detoxification enzyme, glutathione-S-transferase, but that didn’t happen when the larvae were ex-posed to both pesticides. Since that group of larvae was the worst at hunting, they were definitely experiencing toxic effects so why weren’t their detoxifi-cation systems working harder? We can’t say for sure, but one idea is that the pyrethroid combination caused damage to the detoxification system itself which prevented it from clearing out the pesticides resulting then in even less eating.
The third study of this thesis was made up of two experiments that pri-marily investigated microplastics and compared the effects of microplastics to the effects of a pesticide. We also combined the microplastics and pesti-cide to see what would happen. Midge larvae (same species as in the first study) were raised in lab-made dirt that had been spiked either with 1.0 or 10.0 micron-sized plastic (polystyrene-based latex) beads, or esfenvalerate (the pesticide) or a combination of the 1.0 micron beads and esfenvalerate. After a few weeks we realized there weren’t as many adult midges emerging as there ought to have been. It turned out we hadn’t mixed enough food into the dirt at the beginning which resulted in an opportunity to see what hap-pens when exposure to these contaminants occurs under stressful conditions that are actually more likely to be the case out in nature. Of course, we also did a second experiment in which food was plentiful. We found that when conditions were ideal, none of the contaminants appeared to affect the midg-es. However, when food was scarce, both sizes of microplastics and the esfevalerate all reduced the number of larvae that made it to adulthood. Ad-ditionally, fewer larvae survived under those conditions when exposed to esfenvalerate, but not when microplastics were mixed in as well. This study shows that substances that may appear harmless are in fact stressing the an-imals, a fact that becomes apparent when resources are limited. The silver-lining here is that the microplastics could be acting like cleaning agents by soaking up the esfenvalerate.
The fourth and final study of this thesis aims to initiate a conversation about the potential for antihistamine pollution to interfere with the ther-moregulatory physiology and behavior of aquatic wildlife. We exposed freshwater snails to a range of concentrations of the popular antihistamine, diphenhydramine, for 24 hours. Then we looked for changes in the highest temperature the snails could tolerate, which temperature they preferred to be

49
in the most and how long it took them to get back up on their foot after they were placed on their side. Exposure to diphenhydramine resulted in the snails tolerating a higher temperature and taking much longer time to right themselves. Antihistamines in humans work by blocking the receptors that receive histamines, which are neurotransmitters. Histamines and their recep-tors are not only found in humans, but other animals as well, including in-sects and snails. In humans histamine pathways are not only involved in immune response (e.g. allergic reactions) but also regulating wakefulness which is why one of the side effects of disphenhydramine is sleepiness. There is evidence from other studies that suggest this is true for other ani-mals as well. Therefore, the snails we exposed to diphenhydramine may have struggled to stand back up because they were sedated. Some interesting studies on fruit flies showed that the same histamine-signaling gene that is expressed in circadian clock neurons (i.e. involved in regulating wakeful-ness) also has thermoregulatory functions. Our experiment found no changes in temperature preference in the snails after they were exposed to diphenhy-dramine, however they did tolerate higher temperatures. This study provides evidence that contaminants should be reviewed for their potential to effect thermogulatory physiology of aquatic animals as this form of toxicity may have consequences for populations already living in habitats at one end of their thermal range.
In conclusion, this thesis provides evidence that sub-lethal concentrations of pollutants can harm the insects and snails living in freshwater environ-ments. Reduced larval survival, delayed development, decreased emergence, inhibited predation behavior, induction and inhibition of detoxification en-zymes, increased thermal tolerance and delayed righting time are all harmful effects with the potential to change the population dynamics of freshwater ecosystems. Two of the three species investigated have a terrestrial adult stage with important roles to play in terrestrial community structures as well. Most of the observed harmful effects of exposure occurred at environmental-ly relevant concentrations indicating that current contamination of freshwa-ters is high enough to affect wildlife populations and consequently the biodi-versity of freshwater ecosystems as well as the adjacent terrestrial ecosys-tems. We have come a long way since Rachel Carson’s publication of “Si-lent Spring” in 1962 which first criticized the mantra that “the solution to pollution is dilution” and spawned the entire discipline of ecotoxicology. Despite ingeniously inventing new contaminants as quickly as we ban the old ones, there is world-wide consensus now that freshwater is precious and freshwater ecosystems should be conserved as much as possible. It is my hope that this thesis has contributed in some small ways to increasing what is possible.

50
Svensk Sammanfattning
"Rädda bina!"
"Bort med plasten!"
"Fisken använder droger!"
Medierna gör sin del för att öka medvetenheten om farorna med bekämp-ningsmedel, mikroplast och läkemedel i miljön. Har du ett Twitter-konto kan du inte ha undgått att höra att bekämpningsmedel gör att bikolonier kollap-sar, sett en bild på en havssköldpadda som äter en plastpåse eller laddat ner en app för att ta reda på om tandkrämen innehåller mikroplast, eller känt skuld för att p-piller och antidepressiva medel förgiftar haven. Miljöaktiv-ismen är sällan subtil. Miljövetenskap, å andra sidan, bryr sig mycket om subtilitet. Vetenskapen vill veta: exakt vad är det som händer?
Under sina första decennier som nytt forskningsområdet, i senare delen av 1900-talet, försökte ekotoxikologin bestämma exakt hur mycket av våra tillverkade kemikalier vi skulle kunna släppa ut i miljön utan att döda allt där. Traditionella ekotoxikologiska studier tog en handfull arter som fick representera allt som människan inte avsiktligt ville döda och utsatte dem för ökande mängder av en enda kemikalie och registrerade hur många som dog. Senare kom reproduktionsstudier och studier om kronisk exponering, så småningom följt av tester på en större mängd arter, farhågor över bland-ningsinteraktioner och ansträngningar för att erkänna att icke-dödliga effek-ter också är relevanta. Fram till 2000-talet var föroreningarna på en ekotoxi-kologs radar mestadels tungmetaller från den industriella eran och kemiska vapen – såväl de som användes mot medmänniskor som de som användes mot arter i naturen som verkade fast beslutna att äta våra grödor och bosätta sig i våra mattor. Sedan kom de första rapporterna om flytande sopområden i oceanerna, valmagar fulla av avfall och insikten att vi pissar ut en rejäl mängd av alla läkemedel vi tar. Ekotoxikologin började kännas mer som veterinärmedicin, vildmarksvarianten av "vad händer när Fluffy äter sin plastboll?" och "kommer Fluffy att dö om jag ger henne en aspirin?". När fler övervakningsstudier rapporterade om förhållandena i miljön kom de olycksbådande frågorna som "vad händer om Fluffy äter en plastboll varje dag under sitt liv?" och "vad händer med Fluffy om hon får i sig lite aspirin, p-piller, antidepressiva medel, cancermedicin och allergimedicin ... varje

51
dag?". Samtidigt fortsätter vi att använda tungmetaller i industrin och vi fortsätter att använda kemiska vapen dagligen, trots ökad reglering har dessa farhågor inte försvunnit.
Denna avhandling kommer i ett sammanhang där mänsklighetens skräp - kemiskt, fysiskt och farmakologiskt - fortfarande regelbundet hamnar i sjöar och floder över hela världen. Ansvaret ligger på ekotoxikologerna att bevisa om dessa ämnen orsakar eller inte orsakar skador på de bräckliga ekosyste-men. Det är upp till oss att bestämma vilken typ av skada vi ska leta efter och hur högt larmet ska låta, om det alls ska låta. Det finns fortfarande många som högt förkunnar att plast är inert och därför ofarligt, och att lös-ningen på föroreningsproblemet är att späda ut det mer. Är dessa påståenden sanna? Den här avhandlingen tittar på vad som händer när ekologiskt viktiga fjädermyggor, flicksländor och sniglar exponeras i laboratoriet för förore-ningar som de kan utsättas för i miljön.
Den första studien i den här avhandlingen var en simulering av en storm-händelse. Koncentrationerna av bekämpningsmedel i floder och sjöar ökar under den första timmen efter en kraftig regnskur. Detta beror på att regnet faller på besprutad jord och växter och sedan för med sig besprutningsmedlet via stormdräneringar och diken för att slutligen hamna i floder och sjöar. När myggor kläcks från sina ägg, är de små röd-lila larver på mindre än 1 milli-meter. Efter en period av tillväxt bildar det puppor för att sedan omvandlas till sin flygande, vuxna form. De parar sig i svärmar och dör naturligt inom några veckor om ingenting dödar dem innan dess (vanligtvis för att ätas, men kanske också av irritation). Deras korta livslängd gör det lätt att studera ef-fekterna av exponering för föroreningar under en hel livstid jämfört med arter som lever längre. När mygglarverna var 2 dagar gamla utsatte vi dem för små mängder av fyra olika typer av vanligt förekommande bekämp-ningsmedel (imidakloprid, tiacloprid, deltametrin och esfenvalerat) från två grupper av pesticider (neonicotinoider och pyretroider) i vattnet. Mängderna vi använde var inom de värden som rapporterats vara närvarande i sötvatten i naturen. Vissa larver utsattes för endast ett bekämpningsmedel, andra fick en kombination av två och resten var utsatta för alla fyra. Efter en timme hällde vi bort det förorenade vattnet och gav dem rent vatten. Efter det höll vi reda på hur många som överlevde till vuxen ålder och hur lång tid det tog dem att utvecklas.
Vi fann att en kort tids exponering (1 timme) för bekämpningsmedel av en koncentration som liknar den som finns i naturen, dödade några av lar-verna och förlängde utvecklingstiden för andra. Bekämpningsmedlen kan störa larvernas nervsystem vilket gör det svårt för dem att röra sig under en period. Om de inte kan röra sig, kan de inte äta, vilket kan försvaga dem så till den grad att de inte kan återhämta sig och svälter ihjäl. Det kan också innebära att de behöver mer tid än vanligt att växa innan deras kroppar är redo för metamorfos. En god nyhet är att exponeringen av en kombination av alla fyra bekämpningsmedel tillsammans inte var värre än exponeringen för

52
endast ett. Faktum är att vissa kombinationer inte var lika dåliga som expo-nering för endast ett medel. Varför blir det så? Det finns specialiserade av-giftningssystem inom alla djur som står för hanteringen av de gifter som kommer in i kroppen. Ibland gör dessa system ett gift värre innan de gör det bättre. Det beror på att de naturliga avgiftningssystemen inte har haft en chans att utvecklas för att hantera de nya gifter som vi skapar. Anpassning sker och vi kallar det att "utveckla resistens". I fallet när ett gift blir omvand-lat till något värre innan det omvandlas till något bättre gör det hela proces-sen långsammare om systemet samtidigt hanterar ett annat gift. Dessutom är vissa gifter lättare känna igen än andra och utlöser starkare respons i avgift-ningssystemet (djuret är därför "mer resistent" mot dessa gifter). Större re-sistens mot ett gift kan leda till större motstånd mot båda gifterna när båda är närvarande. Ett gift som inte omedelbart och effektivt känns igen av avgift-ningssystemet har mer tid att åstadkomma skada hos djuret än om ett gift som systemet känner igen också är närvarande och utlöser systemet. När systemet väl har startats kan båda gifterna elimineras mer effektivt.
Den andra undersökningen i avhandlingen undersökte hur små mängder av två bekämpningsmedel som kan påverka beteenden. Flicksländor, nära besläktade med trollsländor, är ljust färgade insekter med vingar som vanlig-en finns runt strandkanten vid dammar och strömmande vatten. De lägger sina ägg i stammar av vattenväxter och de kläcks till vinglösa larver med sex ben som lever under vattnet på leriga bottnar i ett till två år innan de om-vandlas till vuxna flygande individer för att para sig. Både larver och vuxna är skickliga rovdjur som främst äter mindre insekter och kräftdjur. Vi kläckte flicksländelarver inne på labb där vi i 14 timmar utsatte dem för antingen en hög eller låg koncentration av två olika pyretroid-bekämpningsmedel (esfen-valerat och deltametrin) eller en kombination av båda pesticiderna. Sedan undersökte vi varje flicksländelarvs beteende i 10 minuter i närvaro av by-tesdjur. Beteendena var indelade i två kategorier: de som hör till jakt och de som hör till allmän rörelse. Vi avlivade sedan larverna och testade deras kroppar för molekylära tecken på toxicitet. Vi letade efter ökad oxidativ skada på fettmolekyler, ökad energianvändning och ökad avgiftningsaktivi-tet.
Flicksländelarver som utsattes för deltametrin jagade inte lika mycket som de som inte utsattes för bekämpningsmedel. Det var inte bara så att lar-verna försökte men inte kunde jaga, de försökte nästan inte alls. Larverna, som utsattes för både deltametrin och esfenvalerat, försökte ännu mindre, men vi kunde inte observera tillräckligt med larver för att med säkerhet säga att båda bekämpningsmedel samtidigt var värre än bara deltametrin. Varför jagade de inte? Det mest uppenbara svaret skulle vara tillfällig förlamning. Båda dessa bekämpningsmedel kan orsaka förlamning hos insekter, det är så de fungerar som insektsbekämpningsmedel. Dock både gick och simmade larverna som vanligt vilket utesluter allmän förlamning. Andra studier har också visat att exponering för pyretroider kan göra att insekter slutar äta.

53
Oavsett mekanismen är det inte bra för naturliga populationer av flickslände-larver eller deras födovävar. När rovdjur slutar äta och deras populationer minskar, kan det ge följdeffekter i hela ekosystem. Intressant nog visade vi att varje pesticid i sig utlöste avgiftningssystem hos flicksländorna när det gäller uppreglering av avgiftningsenzymet, glutation-S-transferas, men det hände inte när larverna utsattes för båda bekämpningsmedlen samtidigt. Ef-tersom denna grupp av larver jagade minst, upplevde de definitivt toxiska effekter, men varför jobbade deras avgiftningssystem inte hårdare? Det går inte att säga säkert, men en förklaring skulle kunna vara att pyretroid-kombinationen orsakade skador på avgiftningssystemet i sig självt, vilket förhindrade det från att rensa ut bekämpningsmedlen som då resulterade i ännu mindre födosök.
Den tredje studien i denna avhandling bestod av två experiment som i första hand undersökte effekter av mikroplast och jämförde effekterna av mikroplast mot effekterna av ett bekämpningsmedel. Vi kombinerade också mikroplast och bekämpningsmedel för att se vad som skulle hända. Fjäder-myggslarver (samma art som i den första studien) föddes upp i labbtillverkat bottenmaterial som antingen hade tillsatts med 1,0 eller 10,0 mikrometer stora sfäriska pärlor av mikroplast (polystyrenbaserade latex) eller esfenvale-rat (bekämpningsmedel) eller en kombination av 1,0 mikrometer pärlor och esfenvalerat. Efter några veckor insåg vi att det inte fanns så många vuxna fjädermyggor som det borde ha varit. Det visade sig att vi inte hade blandat tillräckligt med mat i bottenmaterialet vilket resulterade i att vi kunde under-söka vad som händer när exponering för dessa föroreningar sker under stres-sande förhållanden som faktiskt är mer naturliga. Naturligtvis gjorde vi också ett andra experiment där de fick rikligt med mat. När förhållandena var ideala, fanns det ingen effekt av föroreningarna på myggorna. Men när maten var begränsande, minskade båda storlekarna av mikroplast och esfe-valerat antalet larver som klarade sig till vuxen ålder. Dessutom överlevde färre larver under födobegränsning vid närvaro av esfenvalerat, medan ingen effekt på överlevnad hittades i närvaro av mikroplast. Denna studie visar att ämnen som kan verka ofarliga faktiskt stressar djuren, ett faktum som up-penbaras när resurserna är begränsade. Den ljusa sidan är att mikroplasten kan fungera som rengöringsmedel genom att suga upp esfenvaleratet.
Den fjärde och sista studien i denna avhandling syftar till att initiera en diskussion om huruvida antihistaminföroreningar kan störa värmereglering-en hos vattenlevande organismer. Vi utsatte sötvattenslevande snäckor för olika koncentrationer av det ofta använda antihistaminpreparatet di-fenhydramin under 24 timmar. Sedan studerade vi om det skett någon för-ändring av den högsta vattentemperatur som snäckorna kunde tolerera samt vilken temperatur de föredrog, liksom hur lång tid det tog för dem att komma “på fötter” igen efter att de lagts på sidan. Exponering för di-fenhydramin resulterade i att snäckorna tolererade högre temperaturer och även att de tog mycket längre tid på sig för att komma “på rätt köl” igen. Hos

54
människor fungerar antihistaminer genom att de blockerar receptorer för neurotransmittorn histamin. Histaminer och deras receptorer finns inte bara hos människor, utan även hos andra djur, exempelvis insekter och snäckor. Mänskliga histaminer är inte enbart involverade i immunförsvaret (t.ex. vid allergiska reaktioner) utan påverkar även graden av vakenhet, vilket är en av anledningarna till att intag av difenhydramin leder till trötthet. Andra studier har visat att detta skulle kunna vara fallet även för andra djur. Följaktligen skulle en orsak till att snäckorna hade svårt att “resa sig” efter exponering av difenhydramin kunna vara att de var påverkade av drogen. Studier på banan-flugor har visat att en viss gen som påverkar vakenhet även har värmeregle-rande funktioner. Våra experiment på snäckor påvisade emellertid inte någon förändring av deras temperaturpreferens efter exponering av difenhydramin, även om de kunde uthärda högre temperaturer. Denna studie presenterar belägg för att miljögifter kan påverka värmeregleringen hos vattenlevande djur, vilket kan ha negativa konsekvenser för populationer som redan lever under förhållanden som gränsar till vad de är anpassade till.
Sammanfattningsvis visar denna avhandling att relativt låga, inte dödliga, koncentrationer av vissa föroreningar kan vara skadliga för sötvattenslev-ande insekter och snäckor. Studierna visar på flera effekter som har potential att förändra populationsdynamiken i sötvattensekosystem, såsom minskad överlevnad hos larver, fördröjd utveckling, minskad överlevnad till vuxen-stadiet, försämrat predationsbeteende, både uppreglering och inhibering av avgiftningsenzymer, ökad värmetolerans och försämrad förmåga att “komma på rätt köl”. Två av de studerade arterna är landlevande som färdigutveck-lade, och fyller viktiga funktioner även i detta stadium. Merparten av alla observerade skadliga effekter som orsakades av miljögifter i experiment inträffade vid sådana koncentrationer som regelmässigt uppmäts i naturen, vilket följaktligen kan påverka den biologiska mångfalden i ekosystem både i sötvatten och på land. Vi har kommit en bra bit på väg sedan publiceringen av Rachel Carsons bok Tyst vår (svensk upplaga 1963), som kritiserade den allmänna inställningen att “föroreningsproblem kan lösas genom utspäd-ning”, och som ledde till utvecklingen av ekotoxikologin. Trots att nya mil-jögifter utvecklas i samma takt som gamla förbjuds, är de flesta idag överens om att sötvatten är en livsviktig resurs och att sötvattensekosystem bör beva-ras i största möjliga omfattning. Jag hoppas att denna avhandling kan bidra till detta.

55
Acknowledgements
Funding for this research was provided by the Oscar Lili Lamm Memorial Fund, the Uppsala University Zoological Foundation and the Swedish Re-search Council.
Many thanks to my main supervisor, Frank Johansson. Taking on an inter-disciplinary PhD student is no trivial endeavor, particularly one as extensive-ly interdisciplinary as me. Thank you for lengthening the leash in the begin-ning and for eventually ditching it altogether. By the end I felt like a free-ranging dolphin and you were the fisherman who was always happy to see me and throw me some of your catch. Although perhaps a bird analogy would be more appropriate! Thank you for accepting me for who I am as an environmental scientist and as a person. You’re so darn accepting, in fact, that being the first transgender student to receive a PhD from this depart-ment, feels utterly banal. That wouldn’t have been possible 20 years ago. Thank you for making the world a better place.
Jan Örberg, my co-supervisor, for letting me talk my way into the ecotoxi-cology master program back in 2009 (despite not having the pre-requisites), for taking me ice skating on the lake and for stepping up to help supervise my PhD studies when your added expertise was needed. You are Uppsala to me as much as the castle, the bicycle paths and the apple trees.
Sarah Josefsson, my co-supervisor, for your consistent and kind advice and discerning input. I have greatly appreciated your encouragement and compe-tency.
Ane Laugen, for preparing me for this PhD, for believing in me and for be-ing a very dear friend.
Dan Brabander, for teaching me that pairing compassion with science is a worthy endeavor and for providing the solid foundation on which this thesis was built.
Ingrid Ahensjö, for always being able and willing to tell me which direction is up! That is to say, no matter how confused or lost I got, you would be

56
there; an omnipresent, ever-glowing lamp post, always able to help me find whatever path I was looking for.
Richard Svanbäck and Javier Varg, for such an exciting and fruitful collabo-ration on the microplastics projects. You helped me turn a wild idea into actual, real science.
Gunilla Engström, for your endless support in the laboratory and for your friendship. You are the world’s best listener. I wouldn’t have been able to do this without you.
Annette Berger, for keeping bureaucratic headaches to a minimum with skill and ability that I hope no one ever takes for granted.
Mats, Jacob and Anssi for keeping it real. You helped show me that life and science can co-exist. I can’t picture the lunch room without you guys and I’m pretty sure the smell of coffee will always conjure up your faces. Thank you for keeping your doors open to me and my many random questions.
Elisabeth, for being so genuine and always making me laugh. I should also thank you for the several dozens of times you have given me advice or helped me solve a problem, but there are few people in the world who can make me chuckle as quickly or as often as you and that has meant a great deal to me.
Magnus and Rike for teaching me about snails and for inspiring me to see thermal tolerance and thermal regulation as traits that may be vulnerable to toxicity.
Lukas, for the insane hours and monumental effort you put into investigating the effects of diphenhydramine on those snails. You never failed to impress me.
Bea, for the “Summer of Tadpoles” and all your mentoring.
My team of last-minute translators who made sure I had a suitable “svensk sammanfattning” in this thesis: Sara, Sarah, Richard, Per and Elisabeth.
Murielle, Eryn, Johanna, Will and Yvonne for incorporating feminism and compassion into life at EBC making such a positive impact on the whole place. Thank you also for all the emotional labor and tissues for the days I cried through.

57
Per, for caring about me. Your warmth and consideration has made all the difference.
Tom, for bringing sunshine into our office even on the darkest of days
Kai, for being the best table tennis coach! I had wanted to learn my entire life and you made that dream happen.
The rest of my fabulous PhD cohort, for your inspirational camaraderie and support.
Brian and Leanne, for being the two people I met at the department who probably know me better than I know myself. Thank you for your lasting friendships.
Gijs, for the deep and meaningful discussions.
Sofia, Texas, and Ling, for keeping me anchored in the world outside Uni.
Dad, for all your love and support.
And Zoë, thank you for your patience. The thesis is done now. You will never have to watch this much TV again. Let’s go have some fun!

58
References
Agra, A.R., Soares, A.M.V.M., 2009. Effects of two insecticides on survival, growth and emergence of chironomus riparius meigen. Bull. Environ. Contam. Toxicol. 82, 501–504. doi:10.1007/s00128-009-9658-z
Ahmad, M., 2007. Potentiation/Antagonism of pyrethroids with organophosphate insecticides in Bemisia tabaci (Homoptera: Aleyrodidae). J. Econ. Entomol. 100, 886–93. doi:10.1603/0022-0493(2007)100[886:AOPWOI]2.0.CO;2
Amweg, E.L., Weston, D.P., You, J., Lydy, M.J., 2006. Pyrethroid insecticides and sediment toxicity in urban creeks from California and Tennessee. Environ. Sci. Technol. 40, 1700–1706. doi:10.1021/es051407c
Anderson, R.L., 1982. Toxicity of Fenvalerate and Permethrin to Several Nontarget Aquatic Invertebrates. Environ. Entomol. 11, 1251–1257. doi:https://doi.org/10.1093/ee/11.6.1251
Anderson, T.D., Zhu, K.Y., 2004. Synergistic and antagonistic effects of atrazine on the toxicity of organophosphorodithioate and organophosphorothioate insecti-cides to Chironomus tentans ( Diptera : Chironomidae ). Pestic. Biochem. Phys-iol. 80, 54–64. doi:10.1016/j.pestbp.2004.06.003
Ansara-Ross, T., Wepener, V., van den Brink, P., Ross, M., 2012. Pesticides in South African fresh waters. African J. Aquat. Sci. 37, 1–16. doi:10.2989/16085914.2012.666336
Baatrup, E., Bayley, M., 1993. Effects of the Pyrethroid Insecticide Cypermethrin on the Locomotor Activity of the Wolf Spider Pardosa amentata: Quantitative Analysis Employing Computer-Automated Video Tracking. Ecotoxicol. Envi-ron. Saf. doi:10.1006/eesa.1993.1046
Barila, T.Y., Stauffer, J.R.J., Hocutt, C.H., 1982. Temperature preference and avoid-ance response in the common shiner, Notropis cornutus (Mitchell). Arch. fur Hydrobiol. 96, 112–19.
Barnes, D.K.A., Galgani, F., Thompson, R.C., Barlaz, M., 2009. Accumulation and fragmentation of plastic debris in global environments. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 364, 1985–1998.
Bartelt-Hunt, S.L., Snow, D.D., Damon, T., Shockley, J., Hoagland, K., 2009. The occurrence of illicit and therapeutic pharmaceuticals in wastewater effluent and surface waters in Nebraska. Environ. Pollut. 157, 786–791. doi:10.1016/j.envpol.2008.11.025
Beketov, M.A., 2004. Comparative sensitivity to the insecticides deltamethrin and esfenvalerate of some aquatic insect larvae (Ephemeroptera and Odonata) and Daphnia magna. Russ. J. Ecol. 35, 200–204. doi:10.1023/B:RUSE.0000025972.29638.46
Beketov, M.A., Liess, M., 2005. Acute contamination with esfenvalerate and food limitation: chronic effects on the mayfly, Cloeon dipterum. Environ. Toxicol. 24, 1281–1286. doi:10.1897/04-256R1.1
Benoit, D. a., Sibley, P.K., Juenemann, J.L., Ankley, G.T., 1997. Chironomus tentans life-cycle test: Design and evaluation for use in assessing toxicity of

59
contaminated sediments. Environ. Toxicol. Chem. 16, 1165–1176. doi:10.1002/etc.5620160611
Berninger, J.P., Du, B., Connors, K.A., Eytcheson, S.A., Kolkmeier, M.A., Prosser, K.N., Valenti, T.W., Chambliss, C.K., Brooks, B.W., 2011. Effects of the anti-histamine diphenhydramine on selected aquatic organisms. Environ. Toxicol. Chem. 30, 2065–2072. doi:10.1002/etc.590
Besseling, E., Wegner, A., Foekema, E.M., Van Den Heuvel-Greve, M.J., Ko-elmans, A.A., 2013. Effects of microplastic on fitness and PCB bioaccumulation by the lugworm Arenicola marina (L.). Environ. Sci. Technol. 47, 593–600. doi:10.1021/es302763x
Bisesi, J.H., Sweet, L.E., van den Hurk, P., Klaine, S.J., 2016. Effects of an antide-pressant mixture on the brain serotonin and predation behavior of hybrid striped bass. Environ. Toxicol. Chem. 35, 938–945. doi:10.1002/etc.3114
Blair, R.M., Waldron, S., Phoenix, V., Gauchotte-Lindsay, C., 2017. Micro- and Nanoplastic Pollution of Freshwater and Wastewater Treatment Systems. Springer Sci. Rev. 5, 19–30. doi:10.1007/s40362-017-0044-7
Boerger, C.M., Lattin, G.. L., Moore, S.L., Moore, C.J., 2010. Plastic ingestion by planktivorous fishes in the North Pacific Central Gyre. Mar. Pollut. Bull. 60, 2275–8. doi:10.1016/j.marpolbul.2010.08.007
Bradford, M.M., 1976. A rapid and sensitive method for the for the quantitation of microgram quantities of protein utilizing the principle of protein dye-binding. Anal. Biochem. 72, 248–254. doi:10.1016/0003-2697(76)90527-3
Brady, J.A., Wallender, W.W., Werner, I., Fard, B.M., Zalom, F.G., Oliver, M.N., Wilson, B.W., Mata, M.M., Henderson, J.D., Deanovic, L.A., Upadhaya, S., 2006. Pesticide runoff from orchard floors in Davis , California , USA : A com-parative analysis of diazinon and esfenvalerate. Agric. Ecosyst. Environ. 115, 56–68. doi:10.1016/j.agee.2005.12.009
Browne, M.A., Dissanayake, A., Galloway, T.S., Lowe, D.M., Thompson, R.C., 2008. Ingested Microscopic Plastic Translocates to the Circulatory System of the Mussel, Mytilus edulis (L.). Environ. Sci. Technol. 42, 5026–31. doi:10.1021/es800249a
Browne, M.A., Niven, S.J., Galloway, T.S., Rowland, S.J., Thompson, R.C., 2013. Microplastic Moves Pollutants and Additives to Worms, Reducing Functions Linked to Health and Biodiversity. Curr. Biol. 23, 2388–2392. doi:10.1016/j.cub.2013.10.012
Cacciatore, L.C., Guerrero, N. V., Cochón, A.C., 2013. Cholinesterase and carboxy-lesterase inhibition in Planorbarius corneus exposed to binary mixtures of az-inphos-methyl and chlorpyrifos. Aquat. Toxicol. 128, 124–134. doi:10.1016/j.aquatox.2012.12.005
Caldwell, D.J., Mastrocco, F., Hutchinson, T.H., Länge, R., Heijerick, D., Janssen, C., Anderson, P.D., Sumpter, J.P., 2008. Derivation of an Aquatic Predicted No-Effect Concentration for the Synthetic Hormone, 17α-Ethinyl Estradiol. Envi-ron. Sci. Technol. 42, 7046–7054.
Campero, M., Slos, S., Ollevier, F., Stoks, R., 2007. Sublethal pesticide concentra-tions and predation jointly shape life history: Behavioral and physiological mechanisms. Ecol. Appl. 17, 2111–2122. doi:10.1890/07-0442.1
Carpenter, D.O., Gaubatz, G.L., 1975. H1and H2 histamine receptors on Aplysia neurones. Nature 254, 343–344. doi:10.1038/254343a0
Cedergreen, N., 2014. Quantifying synergy: A systematic review of mixture toxicity studies within environmental toxicology. PLoS One 9. doi:10.1371/journal.pone.0096580

60
Chandler, G.T., 1990. Effects of sediment-bound residues of the pyrethroid insecti-cide fenvalerate on survival and reproduction of meiobenthic copepods. Mar. Environ. Res. 29, 65–76. doi:10.1016/0141-1136(90)90028-M
Chang, X., Zhai, B., Liu, X., Wang, M., 2007. Effects of temperature stress and pesticide exposure on fluctuating asymmetry and mortality of Copera annulata (selys) (Odonata: Zygoptera) larvae. Ecotoxicol. Environ. Saf. 67, 120–127. doi:10.1016/j.ecoenv.2006.04.004
Chang, X., Zhai, B., Wang, B., Sun, C., 2009. Effects of the mixture of avermectin and imidacloprid on mortality and developmental stability of Copera annulata (Odonata: Zygoptera) larvae. Biol. J. Linn. Soc. 96, 44–50. doi:10.1111/j.1095-8312.2008.01036.x
Clean Water Act, 1972. U.S.C. Cole, M., Lindeque, P., Fileman, E., Halsband, C., Galloway, T.S., 2015. The Im-
pact of Polystyrene Microplastics on Feeding, Function and Fecundity in the Marine Copepod Calanus helgolandicus. Environ. Sci. Technol. 49, 1130–37. doi:10.1021/es504525u
Coors, A., Frische, T., 2011. Predicting the aquatic toxicity of commercial pesticide mixtures. Environ. Sci. Eur. 23, 22. doi:10.1186/2190-4715-23-22
De Coen, W., Janssen, C.R., 2003. The missing biomarker link: relationships be-tween effects on the celluar energy allocaiton biomarker of toxicant-stressed Daphnia magna and corresponding population characteristics. Environ. Toxicol. 22, 1632–1641.
Debecker, S., Sanmartín-Villar, I., de Guinea-Luengo, M., Cordero-Rivera, A., Stoks, R., 2016. Integrating the pace-of-life syndrome across species, sexes and individuals: covariation of life history and personality under pesticide exposure. J. Anim. Ecol. 85, 726–738.
Deo, R.P., Halden, R.U., 2013. Pharmaceuticals in the built and natural water envi-ronment of the United States. Water (Switzerland). doi:10.3390/w5031346
Díaz, F., Salas, A., Denisse Re, A., Gonzalez, M., Reyes, I., 2006. Thermal prefer-ence and tolerance of green abalone Haliotis fulgens (Philippi, 1845) and pink abalone Haliotis corrugata (Gray, 1828). Aquac. Res. 37, 877–884.
Dris, R., Gasperi, J., Rocher, V., Saad, M., Renault, N., Tassin, B., 2015. Micro-plastic contamination in an urban area: a case study in Greater Paris. Environ. Chem. 12, 592–599. doi:10.1071/EN14167
Dris, R., Gasperi, J., Saad, M., Mirande, C., Tassin, B., 2016. Synthetic fibers in atmospheric fallout: A source of micrplastics in the environment? Mar. Pollut. Bull. 104, 290–293. doi:10.1016/j.marpolbul.2016.01.006
DrugBank, 2017. Diphenhydramine [WWW Document]. URL https://www.drugbank.ca/drugs/DB01075 (accessed 11.28.17).
Dudgeon, D., Arthington, A.H., Gessner, M.O., Kawabata, Z.I., Knowler, D.J., Lé-vêque, C., Naiman, R.J., Prieur-Richard, A.H., Soto, D., Stiassny, M.L.J., Sulli-van, C.A., 2006. Freshwater biodiversity: Importance, threats, status and con-servation challenges. Biol. Rev. Camb. Philos. Soc. doi:10.1017/S1464793105006950
E.G.Bligh, W.J.Dyer, 1959. A rapid method of total lipid extraction and purification. Natl. Res. Counc. CANADA 37, 911–917. doi:10.1139/o59-099
Eliason, E.J., Anttila, K., 2017. Temperature and the Cardiovascular System, in: Fish Physiology. Elsevier, pp. 235–297. doi:https://doi.org/10.1016/bs.fp.2017.09.003
Ensley, S., 2007a. Pyrethrins and pyrethroids, in: Gupta, R.C. (Ed.), Veterinary Toxicology Basic and Clinical Principles. Elsevier Inc., pp. 494–498.

61
Ensley, S., 2007b. Imidacloprid, in: Gupta, R. (Ed.), Veterinary Toxicology Basic and Clinical Principles. Elsevier Inc., pp. 505–507. doi:https://doi.org/10.1016/B978-012370467-2/50141-3
Eriksson, C., Burton, H., 2003. Origins and biological accumulation of small plastic particles in fur seals from Macquarie Island. AMBIO A J. Hum. Environ. 32, 380–384. doi:10.1579/0044-7447-32.6.380
European Commission, 2018a. Neonicotinoids [WWW Document]. Policies, infor-mation, Serv. URL https://ec.europa.eu/food/plant/pesticides/approval_active_substances/approval_renewal/neonicotinoids_en (accessed 5.15.18).
European Commission, 2018b. A European Strategy for Plastics in a Circular Econ-omy. Brussels.
European Commission, 2012. Communication from the Commission to the Council - The combination effects of chemicals. Chemical mixtures.
European Commission, 2000. Directive 2000/60/EC of the European Parliament and the Council of 23 October 2000 establishing a framework for Community action in the field of water. Off. J. Eur. Communities L, 1–72.
Feo, M.L., Ginebreda, A., Eljarrat, E., Barceló, D., 2010. Presence of pyrethroid pesticides in water and sediments of Ebro River Delta. J. Hydrol. 393, 156–162. doi:10.1016/j.jhydrol.2010.08.012
Ferrer, I., Heine, C.E., Thurman, E.M., 2004. Combination of LC/TOF-MS and LC/ion trap MS/MS for the identification of diphenhydramine in sediment sam-ples. Anal. Chem. 76, 1237–1444. doi:10.1021/ac034794m
Field, L.M., Davies, · T G Emyr, Andrias, ·, O ’reilly, O., Williamson, M.S., Wal-lace, · B A, 2017. Voltage‑gated sodium channels as targets for pyrethroid in-secticides. Eur Biophys J 46, 675–679. doi:10.1007/s00249-016-1195-1
Fleeger, J.W., Carman, K.R., Nisbet, R.M., 2003. Indirect effects of contaminants in aquatic ecosystems. Sci. Total Environ. 317, 207–233. doi:10.1016/S0048-9697(03)00141-4
Fonck, C., Easter, A., Pietras, M.R., Bialecki, R.A., 2015. CNS Adverse Effects: From Functional Observation Battery/Irwin Tests to Electrophysiology, in: Pugsley, M.K., Curtis, M.J. (Eds.), Principles of Safety Pharmacology. Spring-er-Verlag, Heidelberg New York Dordrecht London, pp. 83–114. doi:10.1007/978-3-662-46943-9
Fong, P.P., Bury, T.B.S., Donovan, E.E., Lambert, O.J., Palmucci, J.R., Adamczak, S.K., 2017. Exposure to SSRI-type antidepressants increases righting time in the marine snail Ilyanassa obsoleta. Environ. Sci. Pollut. Res. 24, 725–731. doi:10.1007/s11356-016-7855-y
Forbes, V.E., Cold, A., 2005. Effects of the pyrethroid esfenvalerate on life-cycle traits and population dynamics of Chironomus riparius--importance of exposure scenario. Environ. Toxicol. Chem. 24, 78–86. doi:10.1897/03-547.1
Ford, A.T., Fong, P.P., 2016. The effects of antidepressants appear to be rapid and at environmentally relevant concentrations. Environ. Toxicol. Chem. 35, 794–798. doi:10.1002/etc.3087
Froese, J.M.W., Smits, J.E.G., Forsyth, D.J., Wickstrom, M.L., 2009. Toxicity and immune system effects of dietary deltamethrin exposure in tiger salamanders (Ambystoma tigrinum). J. Toxicol. Environ. Health. A 72, 518–26. doi:10.1080/15287390802706363
Galeotti, N., Ghelardini, C., Bartolini, A., 1999. The role of potassium channels in antihistamine analgesia. Neuropharmacology 38, 1893–1901. doi:10.1016/S0028-3908(99)00068-4

62
Ganesh, K.N., Urmila, J., Vijayan, V.A., 2003. Pyrethroid susceptibility & enzyme activity in two malaria vectors, Anopheles stephensi (Liston) & A. culicifacies (Giles) from Mysore, India. Indian J. Med. Res. 117, 30–38.
Gerald, G.W., 2005. The influence of chemical cues and conspecific density on the temperature selection of a freshwater snail (Melanoides tuberculata). J. Therm. Biol. 30, 237–245. doi:10.1016/j.jtherbio.2004.12.002
Gilliom, R.J., 2007. Pesticides in U.S. Streams and Groundwater. Environ. Sci. Technol. 41, 3408–3414. doi:10.1021/es072531u
Goedkoop, W., Spann, N., Åkerblom, N., 2010. Sublethal and sex-specific cyperme-thrin effects in toxicity tests with the midge Chironomus riparius Meigen. Eco-toxicology 19, 1201–1208. doi:10.1007/s10646-010-0505-0
Gordon, C.J., 2005. Temperature and Toxicology: An Integrative, Comparative, and Environmental Approach. Taylor & Francis.
Gowland, B.T.G., Moffat, C.F., Stagg, R.M., Houlihan, D.F., Davies, I.M., 2002. Cypermethrin induces glutathione S-transferase activity in the shore crab, Car-cinus maenas. Mar. Environ. Res. 54, 169–177. doi:10.1016/S0141-1136(02)00105-8
Graaf, S., Adielsson, S., Kreuger, J., 2010. Resultat från miljöövervakningen av bekämpningsmedel (växtskyddsmedel) -- Årssammanställning 2010.
Graham, E.R., Thompson, J.T., 2009. Deposit- and suspension-feeding sea cucum-bers (Echinodermata) ingest plastic fragments. J. Exp. Mar. Bio. Ecol. 368, 22–29. doi:10.1016/j.jembe.2008.09.007
Green, M., 2008. Histamine in the central nervous system. Proc. West. Pharmacol. Soc. 21, 337–339. doi:10.1152/physrev.00043.2007.
Greenberg, A.E., Trussell, R.R., Clesceri, L.S., 1985. Standard Methods for the Examination of Water and Wastewater. Washington D.C.
Gunnarson, L., Jauhiainen, A., Kristiansson, E., Nerman, O., Larsson, D.G., 2008. Evolutionary conservation of human drug targets in organisms used for envi-ronmental risk assessments. Environ. Sci. Technol. 42, 5807–13. doi:10.1021/es8005173
Hardersen, S., Wratten, S.D., 1996. The sensitivity of the nymphs of two New Zea-land Damselfly species (Odonata: Zygoptera) to azinphos-methyl and carbaryl. Aust. J. Ecotoxicol.
Hasenbein, S., Lawler, S.P., Geist, J., Connon, R.E., 2015. The use of growth and behavioral endpoints to assess the effects of pesticide mixtures upon aquatic or-ganisms. Ecotoxicology 24, 746–759. doi:10.1007/s10646-015-1420-1
Hellou, J., Cook, A., Lalonde, B., Walker, P., Dunphy, K., Macleod, S., Cook, A., Lalonde, B., Walker, P., Dunphy, K., Macleod, S., Hellou, J., Cook, A., Lalonde, B., Walker, P., Dunphy, K., Macleod, S., 2009. Escape and survival of Corophium volutator and Ilyanassa obsoleta exposed to freshwater and chloro-thalonil. J. Environ. Sci. Heal. Part A 44, 778–790. doi:10.1080/10934520902928404
Hong, S., Bang, S., Paik, D., Kang, J., Hwang, S., Jeon, K., Chun, B., Hyun, S., Lee, Y., Kim, J., 2006. Histamine and Its Receptors Modulate Temperature-Preference Behaviors in Drosophila. Neuroscience 26, 7245–7256.
Hoppe, P.D., Rosi-Marshall, E.J., Bechtold, H.A., 2012. The antihistamine cimetid-ine alters invertebrate growth and population dynamics in artificial streams. Freshw. Sci. 31, 379–388.
Horton, A.A., Walton, A., Spurgeon, D.J., Lahive, E., Svendsen, C., 2017. Micro-plastics in freshwater and terrestrial environments: Evaluating the current under-standing to identify the knowledge gaps and future research priorities. Sci. Total Environ. doi:10.1016/j.scitotenv.2017.01.190

63
Huang, Y., Zhang, J., Han, X., Huang, T., 2014. The use of zebrafish (Danio rerio) behavioral responses in identifying sublethal exposures to deltamethrin. Int. J. Environ. Res. Public Health 11, 3650–3660. doi:10.3390/ijerph110403650
Hyne, R. V, Maher, W.A., 2003. Invertebrate biomarkers: links to toxicosis that predict population decline. Ecotoxicol. Environ. Saf. 54, 366–374. doi:10.1016/S0147-6513(02)00119-7
Ingram, E.M., Augustin, J., Ellis, M.D., Siegfried, B.D., 2015. Evaluating sub-lethal effects of orchard-applied pyrethroids using video-tracking software to quantify honey bee behaviors. Chemosphere 135, 272–277. doi:10.1016/j.chemosphere.2015.04.022
Ishaaya, I., Elsner, A., Ascher, K.R.S., Casida, J.E., 1983. Synthetic pyrethroids: Toxicity and synergism on dietary exposure of Tribolium casaneum (Herbst) larvae. Pest Manag. Sci. 14, 367–372.
Janssens, L., Stoks, R., 2014. Reinforcing effects of non-pathogenic bacteria and predation risk: from physiology to life history. Oecologia 176, 323–332. doi:10.1007/s00442-014-3030-7
Janssens, L., Stoks, R., 2013. Fitness Effects of Chlorpyrifos in the Damselfly Enal-lagma cyathigerum Strongly Depend upon Temperature and Food Level and Can Bridge Metamorphosis. PLoS One 8. doi:10.1371/journal.pone.0068107
Janssens, L., Stoks, R., 2012. How does a pesticide pulse increase vulnerability to predation? Combined effects on behavioral antipredator traits and escape swimming. Aquat. Toxicol. 110–111, 91–98. doi:10.1016/j.aquatox.2011.12.019
Janssens, L., Van Dievel, M., Stoks, R., 2015. Warming reinforces nonconsumptive predator effects on prey growth, physiology, and body stoichiometry. Ecology 96, 3270–3280. doi:10.1890/15-0030.1
Jeschke, P., Nauen, R., Schindler, M., Elbert, A., 2010. Overview of the Status and Global Strategy for Neonicotinoids (dagger). J. Agric. Food Chem. 1–7. doi:10.1021/jf101303g
Johansson, M., 2015. Adjusting to the extreme : thermal adaptation in a freshwater gastropod.
Johnson, N., Revenga, C., Echeverria, J., 2001. Managing water for people and nature. Science (80-. ). 292, 1071.
Jonsson, M., Fick, J., Klaminder, J., Brodin, T., 2014. Antihistamines and aquatic insects: Bioconcentration and impacts on behavior in damselfly larvae (Zy-goptera). Sci. Total Environ. 472, 108–111. doi:10.1016/j.scitotenv.2013.10.104
Kahl, M.D., Makynen, E. a, Kosian, P. a, Ankley, G.T., 1997. Toxicity of 4-nonylphenol in a life-cycle test with the midge Chironomus tentans. Ecotoxicol. Environ. Saf. 38, 155–60. doi:10.1006/eesa.1997.1572
Katzenberger, M., Hammond, J., Duarte, H., Tejedo, M., Calabuig, C., Relyea, R.A., 2014. Swimming with predators and pesticides: How environmental stressors affect the thermal physiology of tadpoles. PLoS One 9, 1–11. doi:10.1371/journal.pone.0098265
Kay, P., Hiscoe, R., Moberley, I., Bajic, L., Mckenna, N., Kay, P., 2018. Wastewater treatment plants as a source of microplastics in river catchments. Environ. Sci. Pollut. Res. 2014–2017. doi:10.1007/s11356-018-2070-7
Kepner, J., 2004. Synergy: The big unknowns of pesticide exposure. Pestic. You 23, 17–20.
Khalil, A.M., 2015. Toxicological effects and oxidative stress responses in freshwa-ter snail, Lanistes carinatus, following exposure to chlorpyrifos. Ecotoxicol. Environ. Saf. 116, 137–142. doi:10.1016/j.ecoenv.2015.03.010

64
Kleinteich, J., Seidensticker, S., Marggrander, N., Zarf, C., 2018. Microplastics reduce short-term effects of environmental contaminants. part II: Polyethylene particles decrease the effect of polycyclic aromatic hydrocarbons on microor-ganisms. Int. J. Environ. Res. Public Health 15. doi:10.3390/ijerph15020287
Koelmans, A.A., Besseling, E., Wegner, A., Foekema, E.M., 2013. Plastic as a Car-rier of POPs to Aquatic Organisms: A Model Analysis. Environ. Sci. Technol. 47, 7812–7820. doi:10.1021/es401169n
Kortenkamp, A., 2007. Ten years of mixing cocktails: A review of combination effects of endocrine-disrupting chemicals. Environ. Health Perspect. 115, 98–105. doi:10.1289/ehp.9357
Kosonen, J., Kronberg, L., 2009. The occurrence of antihistamines in sewage waters and in recipient rivers. Environ. Sci. Pollut. Res. 16, 555–564. doi:10.1007/s11356-009-0144-2
Kostaropoulos, I., Papadopoulos, A.I., Metaxakis, A., Boukouvala, E., Papadopou-lou-Mourkidou, E., 2001. Glutathione S–transferase in the defence against pyre-throids in insects. Insect Biochem. Mol. Biol. 31, 313–319. doi:http://dx.doi.org/10.1016/S0965-1748(00)00123-5
Kreuger, J., 1998. Pesticides in stream water within an agricultural catchment in southern Sweden, 1990-1996. Sci. Total Environ. 216, 227–251. doi:10.1016/S0048-9697(98)00155-7
Kreutzweiser, D.P., Good, K.P., Chartrand, D.T., Scarr, T.A., Thompson, D.G., 2008. Toxicity of the systemic insecticide, imidacloprid, to forest stream insects and microbial communities. Bull. Environ. Contam. Toxicol. 80, 211–214. doi:10.1007/s00128-007-9347-8
Kristofco, L.A., Brooks, B.W., 2017. Global scanning of antihistamines in the envi-ronment: Analysis of occurrence and hazards in aquatic systems. Sci. Total En-viron. 592, 477–487. doi:10.1016/j.scitotenv.2017.03.120
Kumar, S., Thomas, A., Pillai, M.K.K., 2011. Deltamethrin: Promising mosquito control agent against adult stage of Aedes aegypti L. Asian Pac. J. Trop. Med. 4, 430–435. doi:10.1016/S1995-7645(11)60120-X
Landrum, P.F., Gedeon, M.L., Burton, G.A., Greenberg, M.S., Rowland, C.D., 2002. Biological Responses of Lumbriculus variegatus Exposed to Fluoran-thene-Spiked Sediment. Arch. Environ. Contam. Toxicol. 42, 292–302. doi:10.1007/s00244-001-0032-3
Liess, M., Schulz, R., Liess, M.H.D., Rother, B., Kreuzig, R., 1999. Determination of insecticide contamination in agricultural headwater streams. Water Res. 33, 239–247. doi:10.1016/S0043-1354(98)00174-2
Liess, M., Von Der Ohe, P.C., 2005. Analyzing effects of pesticides on invertebrate communities in streams. Environ. Toxicol. Chem. 24, 954–965. doi:10.1897/03-652.1
Lindström, B., Kreuger, J., 2015. Resultat från miljöövervakningen av bekämp-ningsmedel ( växtskyddsmedel ) - Årssammanställning 2013.
Lusher, A.L., McHugh, M., Thompson, R.C., 2013. Occurrence of microplastics in the gastrointestinal tract of pelagic and demersal fish from the English Channel. Mar. Pollut. Bull. 67, 94–99. doi:10.1016/j.marpolbul.2012.11.028
Lydy, M., Belden, J., Wheelock, C., Hammock, B., Denton, D., 2004. Challenges in regulating pesticide mixtures. Ecol. Soc. 9. doi:1

65
Macherey, A., Dansette, P.M., 2015. Biotransformations Leading to Toxic Metabo-lites: Chemical Aspect, in: Wermuth, C.G., Aldous, D., Raboisson, P., Rognan, D. (Eds.), The Practice of Medicinal Chemistry. Elsevier Ltd, Tokyo, pp. 585–614. doi:10.1016/B978-0-12-417205-0.00025-0
Malaj, E., Von Der Ohe, P.C., Grote, M., Kühne, R., Mondy, C.P., Usseglio-Polatera, P., Brack, W., Schäfer, R.B., 2014. Organic chemicals jeopardize the health of freshwater ecosystems on the continental scale. Proc. Natl. Acad. Sci. U. S. A. 111, 9549–54. doi:10.1073/pnas.1321082111
Maltby, L., 1999. Studying stress: The importance of organism-level responses. Ecol. Appl. 9, 431–440.
Manjon, C., Troczka, B.J., Zaworra, M., Beadle, K., Randall, E., Hertlein, G., Singh, K.S., Zimmer, C.T., Homem, R.A., Lueke, B., Reid, R., Kor, L., Kohler, M., Benting, J., Williamson, M.S., Davies, T.G.E., Field, L.M., Bass, C., Nauen, R., 2018. Unravelling the Molecular Determinants of Bee Sensitivity to Neonico-tinoid Insecticides. Curr. Biol. 28, 1137–1143.e5. doi:10.1016/j.cub.2018.02.045
Manríquez, P.H., Jara, M.E., Mardones, M.L., Navarro, J.M., Torres, R., Lardies, M.A., Vargas, C.A., Duarte, C., Widdicombe, S., Salisbury, J., Lagos, N.A., 2013. Ocean Acidification Disrupts Prey Responses to Predator Cues but Not Net Prey Shell Growth in Concholepas concholepas (loco). PLoS One 8, 1–10. doi:10.1371/journal.pone.0068643
Martin, T.L., Huey, R.B., 2008. Why “suboptimal” is optimal: Jensen’s inequality and ectotherm thermal preferences. Am. Nat. 171, E102–E118. doi:10.1086/527502
Matsuo, R., Fukata, R., Kumagai, M., Kobayashi, A., Kobayashi, S., Matsuo, Y., 2016. Distribution of histaminergic neurons and their modulatory effects on os-cillatory activity in the olfactory center of the terrestrial slug Limax. J. Comp. Neurol. 524, 119–135. doi:10.1002/cne.23829
Maul, J.D., Brennan, A.A., Harwood, A.D., Lydy, M.J., 2008. Effect of sediment- associated pyrethroids, fipronil, and metabolites on Chironomus tentans growth rate, body mass, condition index, immobilization, and survival. Environ. Toxi-col. Chem. 27, 2582–2590. doi:10.1897/08-185.1
Maund, S.J., Campbell, P.J., Giddings, J.M., Hamer, M.J., Henry, K., Pilling, E.D., Warinton, J.S., Wheeler, J.R., 2012. Ecotoxicology of synthetic pyrethroids. Top Curr. Chem. 314, 137–65. doi:10.1007/128_2011_260
McLoughlin, N., IDaqiang, Y., Maltby, L., Wood, R.M., Hongxia, Y., 2000. Evalua-tion of sesensitivity and specificity of two crustacean biochemical biomarkers. Environ. Toxicol. Chem. 19, 2085–2092.
Meinertz, J.R., Schreier, T.M., Bernardy, J.A., Franz, J.L., 2010. Chronic toxicity of diphenhydramine hydrochloride and erythromycin thiocyanate to Daphnia, Daphnia magna, in a continuous exposure test system. Bull. Environ. Contam. Toxicol. 85, 447–451. doi:10.1007/s00128-010-0117-7
Mintenig, S.M., Int-Veen, I., Löder, M.G.., Primpke, S., Gerdts, G., 2017. Identifica-tion of microplastic effluents of waste water treatment plants using focal plane array-based micro-Fourier-transform infrared imaging. Water Res. 108, 365–372.
Murray, F., Cowie, P.R., 2011. Plastic contamination in the decapod crustacean Nephrops norvegicus Linnaeus. Mar. Pollut. Bull. 62, 1207–17. doi:10.1016/j.marpolbul.2011.03.032

66
Nicholls, R., Fowler, R., Niven, J.E., Gilbert, J.D., Goulson, D., 2017. Larval expo-sure to field-realistic concentrations of clothianidin has no effect on develop-ment rate, over-winter survival or adult metabolic rate in a solitary bee, Osmia bicornis . PeerJ 5. doi:10.7717/peerj.3417
Nyman, A.M., Hintermeister, A., Schirmer, K., Ashauer, R., 2013. The Insecticide Imidacloprid Causes Mortality of the Freshwater Amphipod Gammarus pulex by Interfering with Feeding Behavior. PLoS One 8. doi:10.1371/journal.pone.0062472
OECD, 2000. Guidance document on aquatic toxicity testing of difficult substances and mixtures. Environ. Heal. Saf. Publ. Ser. Test. Assess. - N°23 53. doi:ENV/JM/MONO(2007)10
Oliveira, C., Almeida, J., Guilhermino, L., Soares, A.M.V.M., Gravato, C., 2012. Acute effects of deltamethrin on swimming velocity and biomarkers of the common prawn Palaemon serratus. Aquat. Toxicol. 124–125, 209–216. doi:10.1016/j.aquatox.2012.08.010
Oliveira, M., Ribeiro, A., Hylland, K., Guilhermino, L., 2013. Single and combined effects of microplastics and pyrene on juveniles (0+ group) of the common goby Pomatoschistus microps (Teleostei, Gobiidae). Ecol. Indic. 34.
Orr, M. V, El-Bekai, M., Lui, M., Watson, K., Lukowiak, K., 2007. Predator detec-tion in Lymnaea stagnalis. J. Exp. Biol. 210, 4150–4158. doi:10.1242/jeb.010173
Orton, F., Rosivatz, E., Scholze, M., Kortenkamp, A., 2012. Competitive androgen receptor antagonism as a factor determining the predictability of cumulative antiandrogenic effects of widely used pesticides. Environ. Health Perspect. 120, 1578–1584. doi:10.1289/ehp.1205391
Osborne, N.N., Patel, S., 1984. Localisation of histamine-immunoreactivity in spe-cific neurones of the gastropod CNS including a cell which has been identified as histaminergic. Histochemistry 81, 207–208. doi:10.1007/BF00490120
Osterberg, J.S., Darnell, K.M., Blickley, T.M., Romano, J.A., Rittschof, D., 2012. Acute toxicity and sub-lethal effects of common pesticides in post-larval and ju-venile blue crabs, Callinectes sapidus. J. Exp. Mar. Bio. Ecol. 424–425, 5–14. doi:10.1016/j.jembe.2012.05.004
Palmquist, K., Salatas, J., Fairbrother, A., 2012. Pyrethroid Insecticides: Use, Envi-ronmental Fate, and Ecotoxicology, in: Perveen, F. (Ed.), Insecticides -- Ad-vances in Integrated Pest Management. Intech, pp. 251–279. doi:10.5772/29495
Palmquist, K.R., Jepson, P.C., Jenkins, J.J., 2008. Impact of aquatic insect life stage and emergence strategy on sensitivity to esfenvalerate exposure. Environ. Toxi-col. Chem. 27, 1728–34. doi:10.1897/07-499.1
Piasečná, K., Pončová, A., Tejedo, M., Gvoždík, L., 2015. Thermoregulatory strate-gies in an aquatic ectotherm from thermally-constrained habitats: An evaluation of current approaches. J. Therm. Biol. 52, 97–107. doi:10.1016/j.jtherbio.2015.06.007
Population Council, 2005. Environmental Degradation and Human Well-Being: Report of the Millennium Ecosystem Assessment. Popul. Dev. Rev. 31, 389–398. doi:10.1111/j.1728-4457.2005.00073.x
R Core Team, 2016. R: A language and environment for statistical computing. Vi-enna, Austria, R Foundation for Statistical Computing
Rasmussen, J.J., Nørum, U., Jerris, M.R., Wiberg-Larsen, P., Kristensen, E.A., Friberg, N., 2013. Pesticide impacts on predator-prey interactions across two levels of organisation. Aquat. Toxicol. 140–141, 340–345. doi:10.1016/j.aquatox.2013.06.019

67
Rehse, S., Kloas, W., Zarfl, C., 2016. Short-term exposure with high concentrations of pristine microplastic particles leads to immobilisation of Daphnia magna. Chemosphere 153, 91–99. doi:10.1016/j.chemosphere.2016.02.133
Relyea, R., Hoverman, J., 2006. Assessing the ecology in ecotoxicology: A review and synthesis in freshwater systems. Ecol. Lett. 9, 1157–1171. doi:10.1111/j.1461-0248.2006.00966.x
Relyea, R.A., 2009. A cocktail of contaminants: How mixtures of pesticides at low concentrations affect aquatic communities. Oecologia 159, 363–376. doi:10.1007/s00442-008-1213-9
Reynaldi, S., Duquesne, S., Jung, K., Liess, M., 2006. Linking feeding activity and maturation of Daphnia magna following short-term exposure to fenvalerate. Environ. Toxicol. Chem. 25, 1826–1830. doi:10.1897/05-469R.1
Reynaldi, S., Liess, M., 2005. Influence of duration of exposure to the pyrethroid fenvalerate on sublethal responses and recovery of Daphnia magna. Environ. Toxicol. Chem. 24, 1160–1164. doi:10.1897/04-218R.1
Ricciardi, A., Rasmussen, J.B., 1999. Extinction rates of North American freshwater fauna. Conserv. Biol. 13, 1220–1222. doi:10.1046/j.1523-1739.1999.98380.x
Ristola, T., Pellinen, J., Ruokolainen, M., Kostamo, A., Kukkonen, J., 2009. Effect of sediment type, feeding level, and larval density on growth and development of a midge (Chironomous riparius). Environ. Toxicol. Chem. 18, 756–764.
Rochman, C.M., Browne, M.A., Halpern, B.S., Hentschel, B.T., Hoh, E., Karap-anagioti, H.K., Rios-Mendoza, L.M., Takada, H., Teh, S., Thompson, R.C., 2013. Policy: Classify plastic waste as hazardous. Nature 494, 169–71. doi:10.1038/494169a
Rodrigues, A.C.M., Gravato, C., Quintaneiro, C., Barata, C., Soares, A.M.V.M., Pestana, J.L.T., 2015a. Sub-lethal toxicity of environmentally relevant concen-trations of esfenvalerate to Chironomus riparius. Environ. Pollut. 207, 273–279. doi:10.1016/j.envpol.2015.09.035
Rodrigues, A.C.M., Gravato, C., Quintaneiro, C., Golovko, O., Zlábek, V., Barata, C., Soares, A.M.V.M., Pestana, J.L.T., 2015b. Life history and biochemical ef-fects of chlorantraniliprole on Chironomus riparius. Sci. Total Environ. 508, 506–513. doi:10.1016/j.scitotenv.2014.12.021
Romero, A., Ares, I., Ramos, E., Castellano, V., Martínez, M., Martínez-Larrañaga, M.R., Anadón, A., Martínez, M.A., 2015. Evidence for dose-additive effects of a type II pyrethroid mixture. In vitro assessment. Environ. Res. 138, 58–66. doi:10.1016/j.envres.2015.02.008
Rosi-marshall, E.J., Royer, T. V, 2012. Pharmaceutical Compounds and Ecosystem Function : An Emerging Research Challenge for Aquatic Ecologists. Ecosys-tems 15, 867–880. doi:10.1007/s10021-012-9553-z
Rumpf, S., Hetzel, F., Frampton, C., 1997. Lacewings (Neuroptera: Hemerobiidae and chrysopidae) and integrated pest management: Enzyme activity as bi-omarker of sublethal insecticide exposure. J. Econ. Entomol. 90, 102–108. doi:10.1093/jee/90.1.102
Samsøe-Petersen, L., Gustavson, K., Madsen, T., Mogensen, B.B., Lassen, P., Skjernov, K., Christoffersen, K., Jørgensen, E., 2001. Fate and effects of esfen-valerate in agricultural ponds. Environ. Toxicol. Chem. 20, 1570–1578. doi:10.1002/etc.5620200722
Sayeed, I., Parvez, S., Pandey, S., Bin-Hafeez, B., Haque, R., Raisuddin, S., 2003. Oxidative stress biomarkers of exposure to deltamethrin in freshwater fish, Channa punctatus Bloch. Ecotoxicol. Environ. Saf. 56, 295–301. doi:10.1016/S0147-6513(03)00009-5

68
Scherer, C., Brennholt, N., Reifferscheid, G., Wagner, M., 2017. Feeding type and development drive the ingestion of microplastics by freshwater invertebrates. Sci. Rep. 7. doi:10.1038/s41598-017-17191-7
Schram, J.B., Schoenrock, K.M., McClintock, J.B., Amsler, C.D., Angus, R.A., 2014. Multiple stressor effects of near-future elevated seawater temperature and decreased pH on righting and escape behaviors of two common Antarctic gas-tropods. J. Exp. Mar. Bio. Ecol. 457, 90–96. doi:10.1016/j.jembe.2014.04.005
Schulz, R., 2004. Field studies on exposure, effects, and risk mitigation of aquatic nonpoint-source insecticide pollution: A review. J. Environ. Qual. 33, 419–48.
Schulz, R., Liess, M., 2001. Acute and Chronic effects of particles-associated fen-valerate on stream macroinvertebrates: a runoff simulation study using outdoor microcosms. Arch. Environ. Contam. Toxicol. 40, 481–488. doi:10.1007/s002440010200
Setälä, O., Fleming-Lehtinen, V., Lehtiniemi, M., 2014. Ingestion and transfer of microplastics in the planktonic food web. Environ. Pollut. 185, 77–83. doi:10.1016/j.envpol.2013.10.013
Shi, X., Jiang, L., Wang, H., Qiano, K., Wang, D., Wang, K., 2011. Toxicities and sublethal effects of seven neonicotinoid insecticides on survival, growth and re-production of imidacloprid‑resistant cotton aphid, Aphis gossypii. Pest Manag. Sci. 67, 1528–1533. doi:10.1002/ps.2207
Simon-Delso, N., Amaral-Rogers, V., Belzunces, L.P., Bonmatin, J.M., Chagnon, M., Downs, C., Furlan, L., Gibbons, D.W., Giorio, C., Girolami, V., Goulson, D., Kreutzweiser, D.P., Krupke, C.H., Liess, M., Long, E., Mcfield, M., Mineau, P., Mitchell, E.A., Morrissey, C.A., Noome, D.A., Pisa, L., Settele, J., Stark, J.D., Tapparo, A., Van Dyck, H., Van Praagh, J., Van Der Sluijs, J.P., White-horn, P.R., Wiemers, M., 2015. Systemic insecticides (Neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 22, 5–34. doi:10.1007/s11356-014-3470-y
Simons, F.E.R., Simons, K.J., 2011. Histamine and H1-antihistamines: celebrating a century of progress. J. Allergy Clincal Immunol. 128, 11394–50.
Soderlund, D.M., 2012. Molecular Mechanisms of Pyrethroid Insecticide Neurotox-icity: Recent Advances. Arch. Toxicol. 86, 165–181. doi:doi:10.1007/s00204-011-0726-x
Stoks, R., de Block, M., Van de Meutter, F., Johansson, F., 2005. Predation cost of rapid growth: behavioral coupling and physiological decoupling. J. Anim. Ecol. 74, 708–715.
Stoughton, S.J., Liber, K., Culp, J., Cessna, A., 2008. Acute and chronic toxicity of imidacloprid to the aquatic invertebrates Chironomus tentans and Hyalella az-teca under constant- and pulse-exposure conditions. Arch. Environ. Contam. Toxicol. 54, 662–673. doi:10.1007/s00244-007-9073-6
Straub, S., Hirsch, P.E., Bukhardt-Holm, P., 2017. Biodegradable and Petroleum-Based Microplastics Do Not Differ in Their Ingestion and Excretion but in Their Biological Effects in a Freshwater Invertebrate Gammarus fossarum. Int. J. En-viron. Res. Public Health 14, 774.
Su, L., Xue, Y., Li, L., Yang, D., Kolandhasamy, P., Li, D., Shi, H., 2016. Micro-plastics in Taihu Lake, China. Environ. Pollut. 216, 711–719.
Sundbäck, K., Petersen, D.G., Dahllöf, I., Larson, F., 2007. Combined nutrient-toxicant effects on a shallow-water marine sediment system: sensitivity and re-silience of ecosystem function. Mar. Ecol. Prog. Ser. 330, 13–30. doi:10.3354/meps330013

69
Sussarellu, R., Suquet, M., Thomas, Y., Lambert, C., Fabioux, C., Pernet, M.E.J., Le Goïc, N., Quillien, V., Mingant, C., Epelboin, Y., Corporeau, C., Guyomarch, J., Robbens, J., Paul-Pont, I., Soudant, P., Huvet, A., 2016. Oyster reproduction is affected by exposure to polystyrene microplastics. Proc. Natl. Acad. Sci. 113, 2430–2435. doi:10.1073/pnas.1519019113
Syberg, K., Khan, F.R., Selck, H., Palmqvist, A., Banta, G.T., Daley, J., Sano, L., Duhaime, M.B., 2015. Microplastics: Addressing ecological risk through les-sons learned. Environ. Toxicol. Chem. 34, 945–953. doi:10.1002/etc.2914
Tennekes, H., 2010. The systemic insecticides : a disaster in the making. Weevers Walburg Communicatie, Zutphen, The Netherlands.
Teuten, E.L., Saquing, J.M., Knappe, D.R.U., Barlaz, M.A., Jonsson, S., Björn, A., Rowland, S.J., Thompson, R.C., Galloway, T.S., Yamashita, R., Ochi, D., Wa-tanuki, Y., Moore, C., Viet, P.H., Tana, T.S., Prudente, M., Boonyatumanond, R., Zakaria, M.P., Akkhavong, K., Ogata, Y., Hirai, H., Iwasa, S., Mizukawa, K., Hagino, Y., Imamura, A., Saha, M., Takada, H., 2009. Transport and release of chemicals from plastics to the environment and to wildlife. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 364, 2027–45. doi:10.1098/rstb.2008.0284
Thompson, R.C., Olsen, Y.S., Mitchell, R.P., Davis, A., Rowland, S.J., John, A.W.G., McGonigle, D., Russell, A.E., 2004. Lost at Sea: Where Is All the Plas-tic? Science (80-. ). 304, 838. doi:10.1126/science.1094559
Tomizawa, M., Casida, J.E., 2005. NEONICOTINOID INSECTICIDE TOXICOL-OGY: Mechanisms of Selective Action. Annu. Rev. Pharmacol. Toxicol. 45, 247–268. doi:10.1146/annurev.pharmtox.45.120403.095930
Topp, E., Sumarah, M.W., Sabourin, L., 2012. The antihistamine diphenhydramine is extremely persistent in agricultural soil. Sci. Total Environ. 439, 136–140. doi:10.1016/j.scitotenv.2012.09.033
Toumi, H., Boumaiza, M., Millet, M., Radetski, C.M., Felten, V., Fouque, C., Férard, J.F., 2013. Effects of deltamethrin (pyrethroid insecticide) on growth, reproduction, embryonic development and sex differentiation in two strains of Daphnia magna (Crustacea, Cladocera). Sci. Total Environ. 458–460, 47–53. doi:10.1016/j.scitotenv.2013.03.085
Trekels, H., Van de Meutter, F., Stoks, R., 2011. Effects of species-specific interac-tions with predation risk on the relative species sensitivities to a pesticide in wa-ter boatmen (Corixidae). Oikos 120, 897–905. doi:10.1111/j.1600-0706.2010.18852.x
UNEP, 2016. A Snapshot of the World’s Water Quality: Towards a Global Assess-ment. United Nations Environment Programme, Nairobi, Kenya. doi:978-92-807-3555-0
Vaughn, C.C., 2010. Biodiversity Losses and Ecosystem Function in Freshwaters: Emerging Conclusions and Research Directions. Bioscience 60, 25–35. doi:10.1525/bio.2010.60.1.7
Viran, R., Erkoç, F.Ü., Polat, H., Koçak, O., 2003. Investigation of acute toxicity of deltamethrin on guppies (Poecilia reticulata). Ecotoxicol. Environ. Saf. 55, 82–85. doi:10.1016/S0147-6513(02)00096-9
von Moos, N., Burkhardt-Holm, P., Koehler, A., 2012. Uptake and Effects of Mi-croplastics on Cells and Tissue of the Blue Mussel Mytilus edulis L. after an Experimental Exposure. Environ. Sci. Technol. 46, 327–335. doi:10.1021/es302332w

70
Wagner, M., Scherer, C., Alvarez-muñoz, D., Brennholt, N., Bourrain, X., Buching-er, S., Fries, E., Grosbois, C., Klasmeier, J., Marti, T., Rodriguez-mozaz, S., Ur-batzka, R., Vethaak, A.D., Winther-nielsen, M., Reifferscheid, G., 2014. Micro-plastics in freshwater ecosystems : what we know and what we need to know. Environ. Sci. Eur. 26. doi:10.1186/s12302-014-0012-7
Walters, E., Mcclellan, K., Halden, R.U., 2010. Occurrence and loss over three years of 72 pharmaceuticals and personal care products from biosolids -- soil mixtures in outdoor mesocosms. Water Res. 44, 6011–6020. doi:10.1016/j.watres.2010.07.051
Weithmann, N., Möller, J.N., Löder, M.G.J., Piehl, S., Laforsch, C., Freitag, R., 2018. Organic fertilizer as a vehicle for the entry of microplastic into the envi-ronment. Sci. Adv. 4. doi:10.1126/sciadv.aap8060
Weston, D.P., Ramil, H.L., Lydy, M.J., 2013. Pyrethroid insecticides in municipal wastewater. Environ. Toxicol. Chem. 32, 2460–2468. doi:10.1002/etc.2338
Widenfalk, A., Bertilsson, S., Sundh, I., Goedkoop, W., 2008. Effects of pesticides on community composition and activity of sediment microbes—responses at various levels of microbial community organization. Environ. Pollut. 152, 576–584. doi:10.1016/j.envpol.2007.07.003


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology 1692
Editor: The Dean of the Faculty of Science and Technology
A doctoral dissertation from the Faculty of Science andTechnology, Uppsala University, is usually a summary of anumber of papers. A few copies of the complete dissertationare kept at major Swedish research libraries, while thesummary alone is distributed internationally throughthe series Digital Comprehensive Summaries of UppsalaDissertations from the Faculty of Science and Technology.(Prior to January, 2005, the series was published under thetitle “Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Science and Technology”.)
Distribution: publications.uu.seurn:nbn:se:uu:diva-355236
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2018


















