
Structure formation in active networks
7
ARTICLES PUBLISHED ONLINE: 24 APRIL 2011 | DOI: 10.1038/NMAT3009 Structure formation in active networks Simone Köhler, Volker Schaller and Andreas R. Bausch * Structure formation and constant reorganization of the actin cytoskeleton are key requirements for the function of living cells. Here we show that a minimal reconstituted system consisting of actin filaments, crosslinking molecules and molecular-motor filaments exhibits a generic mechanism of structure formation, characterized by a broad distribution of cluster sizes. We demonstrate that the growth of the structures depends on the intricate balance between crosslinker-induced stabilization and simultaneous destabilization by molecular motors, a mechanism analogous to nucleation and growth in passive systems. We also show that the intricate interplay between force generation, coarsening and connectivity is responsible for the highly dynamic process of structure formation in this heterogeneous active gel, and that these competing mechanisms result in anomalous transport, reminiscent of intracellular dynamics. C ell adhesion, cytokinesis and cell migration are fascinating examples of the power of the cytoskeleton to self-organize locally into complex structures. Whereas such processes are tightly controlled by the biochemical activation of the par- ticipating proteins, structures need to be formed by a physical self-organization process. A stable structure formation mechanism requires the intricate interplay between locally produced forces and the stabilization of the filamentous network by crosslink- ing molecules. Despite the fundamental importance of self- organization processes in such active systems, the microscopic mechanisms and their consequences are poorly understood. To this end, the concept of active gels has been introduced 1–3 , with the structure formation in the cytoskeleton being a prime example 4–6 . Active gel systems rely on pattern-forming mechanisms very differ- ent from those in other soft materials: unlike structure formation in passive out-of-equilibrium systems dominated by nucleation and growth processes, the intriguing dynamical properties of active gels are the result of the interplay between active force generation and force dissipation in the (visco-)elastic environment 7 . A minimal in vitro system of a dynamic cytoskeletal network consists of the combination of only three components: actin filaments, myosin-II motor filaments and crosslinking proteins 8–10 . In a parameter regime where homogeneous networks are still observed, the effect of molecular motors can directly be detected by the bulk properties of the networks 11,12 . In another concentration regime, local and quasi-static structures evolve 13,14 . However, cells rely on highly heterogeneous dynamic structures. It is in this regime of active gels where cells are able to constantly reorganize their structure and mechanics to their local needs. Here we present a minimal model system of an active gel consisting of actin filaments, fascin as a crosslinking molecule and myosin-II filaments as molecular motors. We identify and quantify the key properties of the highly heterogeneous and dynamic system and connect them to the underlying microscopic mechanisms. To this end, we combine fluorescence microscopy, digital image analysis including recognition and tracking of actin structures, and phenomenological simulations. At sufficiently high myosin-II concentrations, we observe a coarsening process, which results in a dynamic steady state, which is characterized by a broad cluster-size distribution. Three distinct cluster sizes with their respective characteristic stability and dynamics can be identified. Lehrstuhl für Biophysik E27, Technische Universität München, James-Franck-Straße 1, 85748 Garching, Germany. *e-mail: [email protected]. Only clusters of actin filaments above a critical size of 10–20 μm in diameter are able to grow and coarsen. Their growth up to several hundred micrometres in diameter is limited by the concomitantly decreasing connectivity within the actin network and the subsequently decreased possibility of active transport. This dependence of the overall mobility on the growth and coarsening rate proves robust for all varied parameters, indicating that this is a generic feature of such active networks. The transport and thus growth dynamics are anomalous, reminiscent of the intracellular dynamics in living cells. Results In vitro, actin filaments in the presence of the crosslinking molecule fascin at a molar ratio of 1:1 assemble into a network of stiff and rigid bundles with a well-defined bundle thickness of about 20 filaments per bundle and lengths of up to several hundred micrometres 15 (Fig. 1a). Thermally excited motions of the bundles are barely visible, and the structure remains stable for hours although permanent unbinding and rebinding of the crosslinking molecules occur 16 . Once the system is switched from a passive to an active state by the presence of myosin-II filaments and ATP, the network’s structure and dynamics are markedly changed. Thereby, it is the concentration ratio between active components and static crosslinkers κ = c myosin :c fascin that determines the network properties (Fig. 1). The actin polymerization triggers a structure formation process that can last for up to 90 min, resulting in a dynamic steady state. This dynamic steady state shows a characteristic size distribution of the evolved patterns and distinct dynamical properties, both of which crucially depend on the chosen parameter set. Two different steady-state scenarios can be observed: a quasi-static regime and a highly dynamic regime. A prevalence of crosslinkers (κ / 1:50) leads to the quasi-static network regime with only minor reorganizations taking place (Fig. 1b,e and Supplementary Movie S1). In this regime, forces of the motor filaments are not sufficient to induce any large-scale dynamics in this crosslinker-dominated network. Consequently, a single percolated network with heterogeneous bundle thicknesses is formed. Only small patches, which are stabilized by a few crosslinkers, can be displaced by the relatively small number of motor filaments. 462 NATURE MATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials © 2011 Macmillan Publishers Limited. All rights reserved
Transcript of Structure formation in active networks

ARTICLESPUBLISHED ONLINE: 24 APRIL 2011 | DOI: 10.1038/NMAT3009
Structure formation in active networksSimone Köhler, Volker Schaller and Andreas R. Bausch*
Structure formation and constant reorganization of the actin cytoskeleton are key requirements for the function of living cells.Here we show that a minimal reconstituted system consisting of actin filaments, crosslinking molecules and molecular-motorfilaments exhibits a generic mechanism of structure formation, characterized by a broad distribution of cluster sizes. Wedemonstrate that the growth of the structures depends on the intricate balance between crosslinker-induced stabilizationand simultaneous destabilization by molecular motors, a mechanism analogous to nucleation and growth in passive systems.We also show that the intricate interplay between force generation, coarsening and connectivity is responsible for the highlydynamic process of structure formation in this heterogeneous active gel, and that these competing mechanisms result inanomalous transport, reminiscent of intracellular dynamics.
Cell adhesion, cytokinesis and cell migration are fascinatingexamples of the power of the cytoskeleton to self-organizelocally into complex structures. Whereas such processes
are tightly controlled by the biochemical activation of the par-ticipating proteins, structures need to be formed by a physicalself-organization process. A stable structure formation mechanismrequires the intricate interplay between locally produced forcesand the stabilization of the filamentous network by crosslink-ing molecules. Despite the fundamental importance of self-organization processes in such active systems, the microscopicmechanisms and their consequences are poorly understood. To thisend, the concept of active gels has been introduced1–3, with thestructure formation in the cytoskeleton being a prime example4–6.Active gel systems rely on pattern-forming mechanisms very differ-ent from those in other soft materials: unlike structure formationin passive out-of-equilibrium systems dominated by nucleation andgrowth processes, the intriguing dynamical properties of active gelsare the result of the interplay between active force generation andforce dissipation in the (visco-)elastic environment7.
A minimal in vitro system of a dynamic cytoskeletal networkconsists of the combination of only three components: actinfilaments, myosin-II motor filaments and crosslinking proteins8–10.In a parameter regime where homogeneous networks are stillobserved, the effect of molecular motors can directly be detected bythe bulk properties of the networks11,12. In another concentrationregime, local and quasi-static structures evolve13,14. However, cellsrely on highly heterogeneous dynamic structures. It is in this regimeof active gels where cells are able to constantly reorganize theirstructure andmechanics to their local needs.
Here we present a minimal model system of an active gelconsisting of actin filaments, fascin as a crosslinking molecule andmyosin-II filaments as molecular motors. We identify and quantifythe key properties of the highly heterogeneous and dynamic systemand connect them to the underlying microscopic mechanisms.To this end, we combine fluorescence microscopy, digital imageanalysis including recognition and tracking of actin structures,and phenomenological simulations. At sufficiently high myosin-IIconcentrations, we observe a coarsening process, which resultsin a dynamic steady state, which is characterized by a broadcluster-size distribution. Three distinct cluster sizes with theirrespective characteristic stability and dynamics can be identified.
Lehrstuhl für Biophysik E27, Technische Universität München, James-Franck-Straße 1, 85748 Garching, Germany. *e-mail: [email protected].
Only clusters of actin filaments above a critical size of 10–20 µmin diameter are able to grow and coarsen. Their growth upto several hundred micrometres in diameter is limited by theconcomitantly decreasing connectivity within the actin networkand the subsequently decreased possibility of active transport. Thisdependence of the overall mobility on the growth and coarseningrate proves robust for all varied parameters, indicating that this isa generic feature of such active networks. The transport and thusgrowth dynamics are anomalous, reminiscent of the intracellulardynamics in living cells.
ResultsIn vitro, actin filaments in the presence of the crosslinkingmolecule fascin at a molar ratio of 1:1 assemble into a networkof stiff and rigid bundles with a well-defined bundle thicknessof about 20 filaments per bundle and lengths of up to severalhundred micrometres15 (Fig. 1a). Thermally excited motionsof the bundles are barely visible, and the structure remainsstable for hours although permanent unbinding and rebindingof the crosslinking molecules occur16. Once the system isswitched from a passive to an active state by the presenceof myosin-II filaments and ATP, the network’s structure anddynamics are markedly changed. Thereby, it is the concentrationratio between active components and static crosslinkers κ =cmyosin:cfascin that determines the network properties (Fig. 1). Theactin polymerization triggers a structure formation process thatcan last for up to 90min, resulting in a dynamic steadystate. This dynamic steady state shows a characteristic sizedistribution of the evolved patterns and distinct dynamicalproperties, both of which crucially depend on the chosen parameterset. Two different steady-state scenarios can be observed: aquasi-static regime and a highly dynamic regime. A prevalenceof crosslinkers (κ / 1:50) leads to the quasi-static networkregime with only minor reorganizations taking place (Fig. 1b,eand Supplementary Movie S1). In this regime, forces of themotor filaments are not sufficient to induce any large-scaledynamics in this crosslinker-dominated network. Consequently, asingle percolated network with heterogeneous bundle thicknessesis formed. Only small patches, which are stabilized by a fewcrosslinkers, can be displaced by the relatively small number ofmotor filaments.
462 NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials
© 2011 Macmillan Publishers Limited. All rights reserved

NATURE MATERIALS DOI: 10.1038/NMAT3009 ARTICLES
20 μm 200 μm
60 min 65 min
Passive Quasi-static Dynamic
a b c
d e f
κ
Figure 1 | Effect of κ on actin/fascin/myosin network structure. a–f, Fluorescence micrographs 90 min after initiation of polymerization (a–c) andcoloured time overlays for 60–65 min after initiation of polymerization (d–f) of 1 µM actin/1 µM fascin at different myosin/fascin ratios. Without myosin(a,d), a passive actin/fascin network with thin, uniform bundles is observed. The inset in a shows a larger magnification of these bundles; all other imageshave the same scale bar as indicated in c. Only small fluctuations occur, resulting in a homogeneously coloured time overlay. Low myosin concentrations(κ = 1:50; b,e) result in quasi-static huge clusters spanning the whole field of view (quasi-static regime). The overall network structure remains constant,and only minor movements of the whole network are observable in the time overlay. At high myosin concentrations (κ = 1:10; c,f), large, dense clusters andsmall clusters coexist in a dynamic steady state (highly dynamic regime). These dynamic reorganizations result in heterogeneous displacements in thetime overlay.
Only in a parameter range where the influence of bothcomponents, passive and active crosslinkers, is balanced (κ ' 1:50),a highly dynamic regime of structure formation and constantreorganization can arise. Here the concentration of molecularmotors is high enough to readily disrupt the network (Fig. 1c).Instead of well-defined bundles, condensed and interconnectedactin–fascin clusters with diameters of up to hundreds ofmicrometres and variable shapes emerge (see Methods). Onceformed, the clusters are constantly subjected to the action ofmolecular motors (Fig. 1f and Supplementary Fig. S1a,b). As aconsequence, they are highly mobile and move in a successionof persistent runs with a mean velocity of 0.5 µms−1 (Fig. 2and Supplementary Movie S2). The actin structures stabilized bythe crosslinking proteins ensure a certain connectivity betweenindividual clusters, which is mandatory for force percolation andthe emergence of highlymobile and dynamic structures.
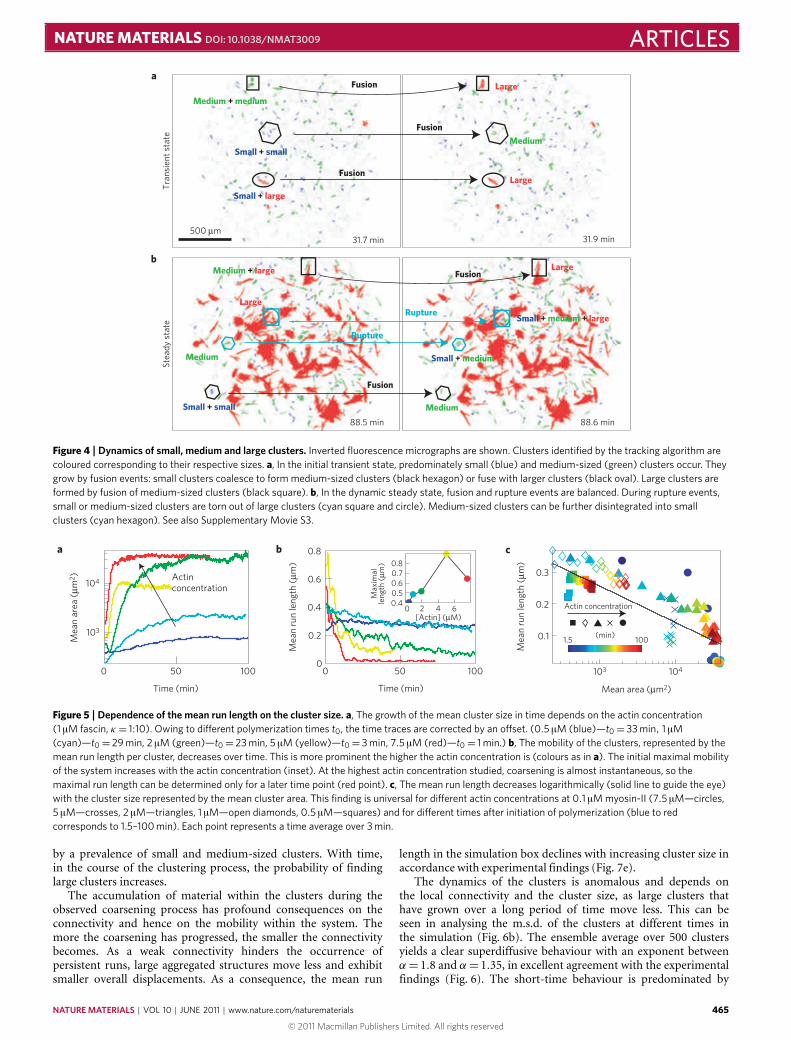
In this regime, the cluster sizes in the steady state are broadlydistributed (Fig. 3a, red).We define three characteristic sizes: small,bundle-like structures (average diameter d ≈ 6 µm), medium-sizedclusters (d ≈ 10 µm) and large clusters (d ≈ 34 µm). The timeevolution of the cluster-size distribution shows that, initially,predominantly small clusters consisting of several actin/fascinbundles are formed spontaneously (Figs 3 and 4a, SupplementaryMovie S3). Once formed, these small clusters fuse to formmedium-sized clusters, yet disruption events of medium- and large-sizedclusters result in their reformation (Fig. 3b). At the same time,small clusters are also stable as they are unlikely to be disruptedowing to their small contact area. The resulting low probabilityof motor filaments binding in an appropriate manner to furtherdisrupt the small clusters limits their disintegration. Consequently,a large number of small clusters can be found in the active systemat all times. Medium-sized clusters are formed continuously byfusion of small clusters or disruption of large clusters. At the sametime, they can be annihilated either by their adsorption to largerclusters or by disruption into small clusters. All of these effects
44 min 42 min
100 µm4 µm s¬1
46 min
a b
c d
Figure 2 |Dynamics of actin/fascin/myosin networks. a–d, Fluorescencemicrographs (a–c) and the resulting coloured time overlay (d) of activeactin networks at the indicated times after initiation of polymerization showa marked structural rearrangement within the network (1 µM actin, 1 µMfascin, 0.1 µM myosin, 1 mM ATP). Arrows indicate the cluster velocities.The overlay (d) represents 42–46 min after polymerization in blue to red.
contribute to the fact that medium-sized clusters predominate innumber (Fig. 3c), although individual medium-sized clusters areintrinsically unstable (Fig. 4b). The large clusters are stable overtime and increase their size by fusion events with clusters of all sizes.Small parts of them are torn out frequently without compromisingtheir structural integrity. Thus, over time a dynamic steady stateof cluster-size distributions evolves with a frequent exchange ofmaterial between the different cluster sizes (Figs 3c and 4b).
The characteristic temporal evolution of the cluster-size dis-tribution shown in Fig. 3 is very robust: it is observed for allstudied actin or myosin concentrations. Only at the very low actin
NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials 463
© 2011 Macmillan Publishers Limited. All rights reserved

ARTICLES NATURE MATERIALS DOI: 10.1038/NMAT3009
50
1002 3 4Log(area (µm2)) Tim
e (m
in)
Small
Large
Medium
Small
Medium
Large
Prop
ortio
n
Time (min)
0
5,000
10,000
Num
ber
15,000
0.1
0.2
0.3
0.4
40 60 80 100
a
b c
Figure 3 | Time evolution of the cluster-size distribution. a, Afterpolymerization, three types of clusters with distinct sizes emerge. Thecluster sizes are determined by their respective areas. The distribution ofthe cluster areas is calculated for 18.4 min time intervals and represents theintegrated total number of clusters found in the field of view in this timeinterval. The positions of the peaks for small, medium and large clusters donot change with time. The temporal evolution is robust for all studied actinconcentrations (see Supplementary Fig. S2). b, Material exchange occursbetween clusters of all sizes. c, With time, the population of the differentcluster sizes changes, as seen in their number proportion: small,homogeneously sized clusters (blue, with area smaller than 17.8 µm2) firstfuse, forming medium-sized clusters (green, area larger than 17.8 µm2 andsmaller than 63.1 µm2). These medium-sized clusters grow further, forminglarge and stable clusters (red, area larger than 63.1 µm2). A dynamic steadystate evolves because medium-sized clusters are constantly destroyed andreformed from a few small clusters, and large clusters are stable.
concentration of 0.5 µM, large clusters hardly form and the meancluster size barely increases (Fig. 5a and Supplementary Fig. S2a,b).At higher actin concentrations, large clusters grow to span thewholefield of view, decreasing the statistics of the cluster-size distribution(Supplementary Fig. S2c,d). Furthermore, with increasing actinconcentration (above 2 µM), the formation of huge clusters resultsin a heterogeneous network organization and only a part of thenetwork structure can be observed in the field of view. This maylead to only fragmentary determination of the mean cluster sizes asobserved for 5 µM actin in Fig. 5a. For all parameter variations, thesystem reaches a dynamic steady state, where the cluster-size dis-tribution and consequently the mean cluster size remain constant(Fig. 5a and Supplementary Fig. S1c).
The plateau value of the mean cluster size and the time toreach the plateau crucially depend on the actin concentration.Conceptually, a high actin concentration enhances the networkconnectivity and thus facilitates an effective self-organizationprocess. Subsequently, the steady state is reached faster, asan increased actin concentration boosts the initial mobility inthe system, as measured by the mean run length per cluster(Fig. 5b, inset). A rapid nucleation of small clusters is alsoobservable. For all actin concentrations, the subsequent formationof medium and large clusters results in a decline of the network’sconnectivity that in turn yields a decreased cluster mobility
(Fig. 5b): the more material is accumulated in the large clusters,the less material is available to connect the different clusterswith each other. Thus, if the tracks necessary for myosin-II toeffectively transport actin structures are lacking, the mobility ofclusters is impared. Importantly, at high actin concentrations,higher average cluster sizes are observed in the steady state(Fig. 5a). These clusters are less connected, yielding a lowermobility (Fig. 5b).
Indeed, we observe a generic dependence of the mean runlength on the cluster size and thus on the connectivity of thenetwork. A logarithmic decrease of the mean run length per clusterwith increasing cluster size is found (Fig. 5c), independent ofthe specific parameter set or the time the system has coarsened(for details see Methods). At the highest actin concentrationused (7.5 µM), a macroscopic contraction into a single, non-mobile cluster is observed, resulting in a freezing of the activenetwork with only minor reorganizations taking place (Fig. 5c,circles and Supplementary Fig. S2c–d). However, at very low actinconcentrations (0.5 µM), the cluster sizes remain predominantlysmall or medium sized. Consequently, the clusters remain in ahighly active state even after 100min.
The dynamics of the contraction is set by the transportmechanism within the network and is anomalous in nature.The dynamics of the individual structures in the network isbest described by the mean square displacement (m.s.d.), whichis computed for over 4,000 individual clusters (for details seeMethods). For Brownian diffusion, the m.s.d. of a trajectory r(t ),〈|r(t + τ ) − r(t )|2〉 ∝ τ α , is expected to increase with time τwith an exponent α = 1, whereas 0 ≤ α < 1 and 1 < α ≤ 2 areindicative of sub- or superdiffusion, respectively17,18. For all actinconcentrations studied, a clear superdiffusive behaviour is found atall times (Fig. 6a and Supplementary Fig. S3). This superdiffusionwith a power-law exponent larger than 1 can be traced backto the complex alternation of runs and stalls in the individualtrajectories (Fig. 6a, inset). Whereas this dynamics with α ≈ 1.7persists during the transient state where the structures evolve,a gradual decrease of the mobility is observed at longer times(α≈ 1.3) with increasing degree of clustering of the structures. Athigh actin concentrations, the long-time behaviour at late timesbecomes subdiffusive, corresponding to immobile clusters showingonly local rearrangements (Supplementary Fig. S3b).
To relate the mesoscopic dynamics and the observed structure-formation processes to the underlying microscopic mechanisms,the microscopic interactions are implemented in a numericalsimulation: in a phenomenological approach, fascin bundlesare modelled as monodisperse and polar rigid rods in a two-dimensional geometry. The bundles are propelled and crosslinked,reflecting transport and binding processes. Both active and passivebinding processes are subjected to rupture events based on aforce-dependent unbinding kinetics (for details see Methods andSupplementary Information).
By varying the ratio of motors and passive crosslinkers,the experimentally observed phase behaviour can be retrieved:whereas an excess amount of passive crosslinkers leads toquasi-static structures with only minor reorganizations, dynamicstructure formation processes occur only when motors and passivecrosslinkers are relatively balanced. Here, the interplay of activetransport and crosslinking leads to the formation of clusters ofaggregated actin/fascin bundles. Starting from a homogeneousinitial state, the simulation shows a coarsening behaviour in thecourse of which small structures such as individual bundles or smallclusters coalesce and gradually form larger structures (Fig. 7a–d,fand Supplementary Movie S4). This is reflected in the temporalevolution of the cluster-size distributions (Supplementary Fig. S4),which is in good agreement with the experimental findings. As inthe experiment, the initial cluster-size distribution is characterized
464 NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials
© 2011 Macmillan Publishers Limited. All rights reserved
NATURE MATERIALS DOI: 10.1038/NMAT3009 ARTICLES
500 µm
Fusion
Fusion
Medium + medium
Small + smallMedium
Small + large
Large
Large
FusionMedium + large
Small + medium
Medium
Small + medium + large
Large
Small + small
Medium
Rupture
Fusion
Stea
dy s
tate
Tra
nsie
nt s
tate
Rupture
Fusion
31.9 min
88.6 min
Large
31.7 min
88.5 min
a
b
Figure 4 |Dynamics of small, medium and large clusters. Inverted fluorescence micrographs are shown. Clusters identified by the tracking algorithm arecoloured corresponding to their respective sizes. a, In the initial transient state, predominately small (blue) and medium-sized (green) clusters occur. Theygrow by fusion events: small clusters coalesce to form medium-sized clusters (black hexagon) or fuse with larger clusters (black oval). Large clusters areformed by fusion of medium-sized clusters (black square). b, In the dynamic steady state, fusion and rupture events are balanced. During rupture events,small or medium-sized clusters are torn out of large clusters (cyan square and circle). Medium-sized clusters can be further disintegrated into smallclusters (cyan hexagon). See also Supplementary Movie S3.
103
104
Time (min)
Mea
n ar
ea (
µm2 ) Actin
concentration
Mea
n ru
n le
ngth
(µm
)
4 60.40.50.60.70.8
[Actin] (µM)
Max
imal
le
ngth
(µm
)
0 50 1000
0.2
0.4
0.6
0.8
0 2
Time (min)
0 50 100
a b
Mea
n ru
n le
ngth
(µm
)
Mean area (µm2)
103 104
0.1
0.2
0.3
Actin concentration
1.5(min)
100
c
Figure 5 |Dependence of the mean run length on the cluster size. a, The growth of the mean cluster size in time depends on the actin concentration(1 µM fascin, κ = 1:10). Owing to different polymerization times t0, the time traces are corrected by an offset. (0.5 µM (blue)—t0= 33 min, 1 µM(cyan)—t0= 29 min, 2 µM (green)—t0= 23 min, 5 µM (yellow)—t0= 3 min, 7.5 µM (red)—t0= 1 min.) b, The mobility of the clusters, represented by themean run length per cluster, decreases over time. This is more prominent the higher the actin concentration is (colours as in a). The initial maximal mobilityof the system increases with the actin concentration (inset). At the highest actin concentration studied, coarsening is almost instantaneous, so themaximal run length can be determined only for a later time point (red point). c, The mean run length decreases logarithmically (solid line to guide the eye)with the cluster size represented by the mean cluster area. This finding is universal for different actin concentrations at 0.1 µM myosin-II (7.5 µM—circles,5 µM—crosses, 2 µM—triangles, 1 µM—open diamonds, 0.5 µM—squares) and for different times after initiation of polymerization (blue to redcorresponds to 1.5–100 min). Each point represents a time average over 3 min.
by a prevalence of small and medium-sized clusters. With time,in the course of the clustering process, the probability of findinglarge clusters increases.
The accumulation of material within the clusters during theobserved coarsening process has profound consequences on theconnectivity and hence on the mobility within the system. Themore the coarsening has progressed, the smaller the connectivitybecomes. As a weak connectivity hinders the occurrence ofpersistent runs, large aggregated structures move less and exhibitsmaller overall displacements. As a consequence, the mean run
length in the simulation box declines with increasing cluster size inaccordance with experimental findings (Fig. 7e).
The dynamics of the clusters is anomalous and depends onthe local connectivity and the cluster size, as large clusters thathave grown over a long period of time move less. This can beseen in analysing the m.s.d. of the clusters at different times inthe simulation (Fig. 6b). The ensemble average over 500 clustersyields a clear superdiffusive behaviour with an exponent betweenα= 1.8 and α= 1.35, in excellent agreement with the experimentalfindings (Fig. 6). The short-time behaviour is predominated by
NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials 465
© 2011 Macmillan Publishers Limited. All rights reserved

ARTICLES NATURE MATERIALS DOI: 10.1038/NMAT3009
102 103
10¬2
10¬3
10¬4
Time (s)
m.s
.d. (µ
m2 )
Time
1.71.3
m.s
.d. (L2
)
Experiment Simulation1.8
1.35
20 µm
Simulation time
L101
102
100 101
a b
Figure 6 |Mean square displacement. a, The ensemble-averaged m.s.d. ofthe clusters at different time intervals shows superdiffusive behaviour witha power-law exponent larger than 1. Initially, in the transient state(17–35 min, blue circles) a high superdiffusivity persists for long times.With increasing time (36–54 min, cyan crosses; 55–72 min; green triangles;73–90, yellow diamonds; 91–109 min, orange squares; 110–127 min; redstars), stalling events of the clusters for long times increase, resulting in abiphasic behaviour of the m.s.d.: a strong superdiffusivity due to localrearrangements at short times and only a slight superdiffusivity on largertime scales. Solid black lines are power laws with indicated exponents toguide the eyes. b, As in the experiment, the transport in the simulations ischaracterized by a clear superdiffusive behaviour. With time, individualclusters grow larger and move less, visible in the sample trajectory shownin the inset. This results in a successive decline in the m.s.d. exponent αand the experimentally observed biphasic behaviour. Insets showexemplary trajectories for the experiment (a) and the simulation (b).
short runs, whereas the long-time behaviour of the system at latetimes becomes increasingly dominated by the dynamic arrest ofthe clusters. This explains the observed biphasic behaviour of them.s.d. in the steady state.
As in the experiment, the stability of the clusters dependson their size. Large clusters are stable because they have grownsufficiently large and comprise a sufficiently large number ofpassive crosslinkers. Motors continuously reorganize even largeclusters internally by rupturing individual bonds. They are alsoable to tear out material from the clusters, yet they are notsufficiently strong and cooperative enough to entirely disintegratethese large structures. Medium-sized clusters, on the contrary,can readily be disintegrated by the action of molecular motors.Here, they find a large number of possible binding sites and thuscan induce unbinding events of the relatively small number ofpassive crosslinks within the cluster. For the smallest structurescomprising only two to five individual bundles, motor-inducedunbinding events play only a minor role: as small clustersdo not offer enough binding sites for motors, the clusterdisintegration predominantly occurs by stochastic unbindingevents not involving motor proteins.
DiscussionCompared with other phase-separation processes in soft materials,such as nucleation and growth of crystals19, phase separationin binary mixtures20 and diffusion-limited aggregation21, activegels in the parameter regime studied here show a remarkabledynamic behaviour.
The incessant local input of mechanical energy at the smallestscales by myosin-II filaments drives a constant reorganization ofthe actin/fascin network through forced unbinding and rebindingevents. In the course of this constant network reorganization,a highly dynamic steady state of aggregated clusters emerges inwhich the characteristic properties, such as the mean clustersize and the activity, crucially depend on the system’s keyparameters: the actin and motor filament concentration. Theactivity of the highly heterogeneous system depends on itsconnectivity, as molecular motors need tracks on which forces canbe exerted. This dependence of the activity on the connectivity
of the networks opens up the possibility that even smallperturbations can result in a large structural response, mediatedby the identified self-organization mechanisms of the active gel.Biochemical signals or mechanical stimuli could cause a shiftin the phase diagram, enabling a rapid, local and highly robustmechanosensing mechanism as observed in cytokinesis or onmechanical stimulation22.
Importantly, the transport dynamics recovered in the reconsti-tuted system is in excellent agreement with the dynamics insidethe cytoplasm of cells. Here too, a superdiffusive behaviour isobservable with exponents between 1.5 and 1.7 determined bydifferent means4,23,24. Whereas in cells the mechanism can be linkedonly to the energy consumption, presumably through the activityof molecular motors, the model system presented here allows theidentification of the underlying microscopic mechanism, which isthe competition between binding and rupturing events evoked bycrosslinkingmolecules andmolecular motors.
Although individual patterns, such as asters of cytoskeletalfilaments, have been successfully described both by mesoscopicmodels in the dilute regime25,26 and macroscopic models2,27, theexplicit modelling of the nonlinear dynamics of extended patternsand coarsening processes proves difficult. This is attributed to theinherent heterogeneity in many model systems that comprise apolymorphism of many coexisting patterns. Especially for genericapproaches that are based on linear irreversible thermodynamicswith geometric nonlinearities, it is therefore challenging to narrowdown the parameter space and to decide which additionalnonlinearities are important27.
Other physiological crosslinkers, such as α-actinin or filamin,already in the passive state result in a much more complicatedand even kinetically trapped network structure28. It remains aformidable challenge to address the effect of active molecularmotors on the local structure and dynamics in such networks.It is the excellent accessibility of the self-organization principlesand dynamics on all levels of description—from the molecularmechanisms to the formation of large-scalemacroscopic patterns—that makes the presented system based on fascin, myosin-II andactin a versatile benchmark for the exploration of this broadmaterial class of active gels. Itmay be the starting point for an in vitroreconstitution of cellular dynamics by a bottom-up approach.
MethodsProtein purification. Myosin29 and G-actin30,31 are extracted from rabbit skeletalmuscle. Actin is fluorescently labelled with Alexa Fluor 555 succinimidylester(Invitrogen) with a 25% degree of labelling. Recombinant human fascin is purifiedfrom Escherichia coli BL21-CodonPlus-RP and stored at−80 ◦C in 2mM Tris-HCl(pH 7.4) and 150mMKCl at 64 µM (ref. 32).
Fluorescence imaging. Actin in the presence of the indicated concentrations ofmyosin-II and fascin is polymerized by adding one-tenth of the sample volumeof 100mM imidazole, 2mM CaCl2 and 30mMMgCl2. The ATP concentration iskept constant at 1mM ATP by adding 20mM creatine phosphate and 0.1mgml−1creatine phospho kinase (Sigma). Casein (3mgml−1) is added to prevent anysurface interactions. In this assay buffer, myosin readily polymerizes into filamentswith amean length of 0.6 µm. Samples are enclosed in hermetically sealed chambersto eliminate any drift in the network. All data are acquired on a Zeiss Axiovert200 inverted microscope with either a ×10 (NA 0.2) long-distance objective or a×40 (NA 1.3) oil immersion objective. Images are captured at 0.84 frames s−1 witha CCD (charge-coupled device) camera (Orca ER, Hamamatsu) attached to themicroscope by a×0.4 camera mount.
Image processing. Images are background-subtracted in ImageJ. To identifyindividual clusters, an intensity threshold value is applied in ImageJ to generatea binary image. The threshold value is set such that the noise level remainsconstant for all studied samples. The cluster size is determined by their areas.Diameters are square roots of their respective area. The clusters are traced overtime using the IDL tracking algorithm33 for the intensity-weighted centroid clusterpositions in Matlab R2008b (Mathworks). To minimize tracking artefacts, thetrajectories are subjected to a gliding average over 4 frames. For each parameterset, clusters were automatically identified and traced for 1–2 h with a time
466 NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials
© 2011 Macmillan Publishers Limited. All rights reserved

NATURE MATERIALS DOI: 10.1038/NMAT3009 ARTICLES
c
0.1
0.2
0.3
0.4 Simulation time
0 5,000
p on
Average cluster size100 101 102 103
Mea
n ru
n le
ngth
Ave
rage
clu
ster
siz
e
Simulation time T
b
a
Simulation time T = 100 Simulation time T = 250 Simulation time T = 1,000
Simulation time T = 1,000
0 200 800600400 1,000
300
100
0
200
c/d
Active crosslink
Passive crosslink
Free filament
Crosslinked filament
a
d e
f
b c
ρ
Figure 7 |Dynamics and coarsening behaviour in the simulations. a–c, Simulation snapshots for a density ρ of 14 rods per L2 (with L being the rod length),a crosslinker unbinding rate poff=0.02, a crosslinker binding rate pon=0.01, a motor unbinding rate roff=0.01 and a motor binding rate ron=0.1 (fordetails, see Methods). As in the experiment, the simulation shows a coarsening process. d, In the steady state, large clusters occur in the entire simulationbox. The black square depicts the region shown in c. e, The more material is accumulated in the clusters, the lower is the connectivity and the motility inthe system. This directly becomes manifest in the mean cluster run length, which decreases with increasing cluster size. The mean run length is defined asthe mean displacement of all clusters within simulation time intervals of1T= 10. As in the experiments, the concomitant decrease of the mean run lengthwith increasing cluster size is robust for a wide range of parameters (e). The actin density was varied from ρ=8 to ρ= 16 and the crosslinker on-rate frompon=0.007 to pon=0.013. The inset shows a schematic map of the used parameters. f, In accordance with the experiment, the simulation shows a rapidcoarsening process that leads to a steady state with a stable average cluster size. All lengths are measured in units of the rod length L= 1. With a rod lengthof 10 µm and a motor velocity of 1 µm s−1, the unit of the simulation time is 1 s.
resolution of 1.19 s. As clusters merge frequently or are annihilated (as describedin the text), or move out of or into the field of view, up to 10,000 clusters perexperiment were identified, although per frame not more than about 100 clustersare observed simultaneously.
To characterize the activity in the system, we calculate the mean run lengthper cluster. To this end, the trajectories are divided into ‘runs’ and ‘stalls’: runs areidentified as movements between two frames with velocities larger than 0.36 µms−1and a change in direction smaller than 30◦. The run length is defined as the lengthof a continuous run over at least 2 successive frames. The mean run length percluster is calculated in 3min time intervals, which is significantly longer thantypical run times (10 s average).
The m.s.d. is calculated for individual traces as 〈r2(τ )〉 = 〈(r(t +τ )− r(t ))2〉and ensemble-averaged subsequently for 18min time slots.
Simulation. In a minimal approach, fascin bundles are modelled as monodisperseand polar rigid rods in a quasi two-dimensional geometry. Despite beingtwo-dimensional, no excluded volume effects are taken into account. The bundlesare actively propelled by motors and crosslinked by passive crosslinkers. Bothactive and passive bindings are subjected to forced unbinding events. The explicitdynamics of motor proteins and crosslinkers is not taken into account; if twofilaments overlap, active or passive binding events occur based on probabilisticinteraction rules. In a similar manner unbinding processes are calculated.
The displacements that arise owing to the action of molecular motors arecalculated using generic velocity models similar to that of ref. 25. For details, seeSupplementary Information.
Received 22 December 2010; accepted 14 March 2011;published online 24 April 2011
References1. LeGoff, L., Amblard, F. & Furst, E. Motor-driven dynamics in actin-myosin
networks. Phys. Rev. Lett. 88, 18101 (2002).2. Kruse, K., Joanny, J-F., Juelicher, F., Prost, J. & Sekimoto, K. Asters, vortices,
and rotating spirals in active gels of polar filaments. Phys. Rev. Lett. 92,78101 (2004).
3. Joanny, J-F., Juelicher, F., Kruse, K. & Prost, J. Hydrodynamic theory formulti-component active polar gels. New. J. Phys. 9, 422 (2007).
4. Lau, A., Hoffman, B., Davies, A., Crocker, J. & Lubensky, T. Microrheology,stress fluctuations, and active behaviour of living cells. Phys. Rev. Lett. 91,198101 (2003).
5. Joanny, J-F. & Prost, J. Active gels as a description of the actin-myosincytoskeleton. HFSP J. 3, 94–104 (2009).
6. Fletcher, D. A. & Geissler, P. L. Active biological materials. Annu. Rev. Phys.Chem. 60, 469–486 (2009).
NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials 467
© 2011 Macmillan Publishers Limited. All rights reserved

ARTICLES NATURE MATERIALS DOI: 10.1038/NMAT3009
7. Guerin, T., Prost, J., Martin, P. & Joanny, J-F. Coordination and collectiveproperties ofmolecularmotors: Theory.Curr.Opin. Cell. Biol. 22, 14–20 (2010).
8. Kane, R. E. Interconversion of structural and contractile actin gels by insertionof myosin during assembly. J. Cell. Biol. 97, 1745–1752 (1983).
9. Janson, L. W., Kolega, J. & Taylor, D. L. Modulation of contractionby gelation/solation in a reconstituted motile model. J. Cell. Biol. 114,1005–1015 (1991).
10. Liverpool, T. B., Marchetti, M. C., Joanny, J-F. & Prost, J. Mechanical responseof active gels. Epl-Europhys. Lett. 85, 18007 (2009).
11. Koenderink, G. H. et al. Liquids and structural glasses special feature: An activebiopolymer network controlled by molecular motors. Proc. Natl Acad. Sci. USA106, 15192–15197 (2009).
12. Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletalprotein networks. Biophys. J. 94, 3126–3136 (2008).
13. Backouche, F., Haviv, L., Groswasser, D. & Bernheim-Groswasser, A.Active gels: Dynamics of patterning and self-organization. Phys. Biol. 3,264–273 (2006).
14. Smith, D. et al. Molecular motor-induced instabilities and cross linkersdetermine biopolymer organization. Biophys. J. 93, 4445–4452 (2007).
15. Claessens, M. M. A. E., Semmrich, C., Ramos, L. & Bausch, A. R. Helicaltwist controls the thickness of F-actin bundles. Proc. Natl Acad. Sci. USA 105,8819–8822 (2008).
16. Lieleg, O., Claessens, M. M. A. E., Luan, Y. & Bausch, A. R. Transientbinding and dissipation in cross-linked actin networks. Phys. Rev. Lett. 101,108101 (2008).
17. Metzler, R. & Klafter, J. The random walk’s guide to anomalous diffusion: Afractional dynamics approach. Phys. Rep. 339, 1–77 (2000).
18. Metzler, R. & Klafter, J. Accelerating Brownian motion: A fractional dynamicsapproach to fast diffusion. Europhys. Lett. 51, 492–498 (2000).
19. Weitz, D. A. & Oliveria, M. Fractal structures formed by kinetic aggregation ofaqueous gold colloids. Phys. Rev. Lett. 52, 1433–1436 (1984).
20. Langer, J. S. Theory of spinodal decomposition in alloys. Ann. Phys. 65,53–86 (1971).
21. Viscek, T. & Family, F. Dynamic scaling for aggregation of clusters. Phys. Rev.Lett. 52, 1669–1672 (1984).
22. Effler, J. C. et al. Mitosis-specific mechanosensing and contractile-proteinredistribution control cell shape. Curr. Biol. 16, 1962–1967 (2006).
23. Hoffman, B. D., Massiera, G., Citters, K. M. V. & Crocker, J. C. The consensusmechanics of cultured mammalian cells. Proc. Natl Acad. Sci. USA 103,10259–10264 (2006).
24. Gallet, F., Arcizet, D., Bohec, P. & Richert, A. Power spectrum ofout-of-equilibrium forces in living cells: Amplitude and frequency dependence.Soft Matter 5, 2947–2953 (2009).
25. Liverpool, T. B. & Marchetti, M. C. Instabilities of isotropic solutions of activepolar filaments. Phys. Rev. Lett. 90, 138102 (2003).
26. Aranson, I. S. & Tsimring, L. S. Pattern formation of microtubules and motors:Inelastic interaction of polar rods. Phys. Rev. E 71, 50901 (2005).
27. Juelicher, F., Kruse, K., Prost, J. & Joanny, J-F. Active behaviour of thecytoskeleton. Phys. Rep. 449, 3–28 (2007).
28. Schmoller, K. M., Lieleg, O. & Bausch, A. R. Structural and viscoelasticproperties of actin/filamin networks: Cross-linked versus bundled networks.Biophys. J. 97, 83–89 (2009).
29. Margossian, S. S. & Lowey, S. Preparation of myosin and its subfragments fromrabbit skeletal muscle.Meth. Enzymol. 85, 55–71 (1982).
30. Spudich, J. A. & Watt, S. The regulation of rabbit skeletal muscle contraction.I. Biochemical studies of the interaction of the tropomyosin-troponin complexwith actin and the proteolytic fragments of myosin. J. Biol. Chem. 246,4866–4871 (1971).
31. MacLean-Fletcher, S. & Pollard, T. D. Identification of a factor in conventionalmuscle actin preparations which inhibits actin filament self-association.Biochem. Biophys. Res. Commun. 96, 18–27 (1980).
32. Vignjevic, D. et al. Formation of filopodia-like bundles in vitro from a dendriticnetwork. J. Cell. Biol. 160, 951–962 (2003).
33. Crocker, J. C. & Grier, D. G. Methods of digital video microscopy for colloidalstudies. J. Colloid. Interface Sci. 179, 298–310 (1996).
AcknowledgementsWe gratefully acknowledge technical support by M. Rusp, G. Chmel and K. Vogt.We gratefully acknowledge the financial support of the DFG in the framework of theSFB 863, and partial support in the framework of the German Excellence Initiativeby the ‘Nanosystems Initiative Munich’ and the ‘Institute of Advanced Studies’(TUM-IAS). S.K. and V.S. thank the ‘International Graduate School for Science andEngineering’. V.S. acknowledges support from the Elite Network of Bavaria by thegraduate programme CompInt.
Author contributionsS.K. and A.R.B. designed experiments, carried out and analysed experiments. V.S., S.K.andA.R.B. conceived, carried out and analysed the simulations andwrote the paper.
Additional informationThe authors declare no competing financial interests. Supplementary informationaccompanies this paper on www.nature.com/naturematerials. Reprints and permissionsinformation is available online at http://www.nature.com/reprints. Correspondence andrequests for materials should be addressed to A.R.B.
468 NATUREMATERIALS | VOL 10 | JUNE 2011 | www.nature.com/naturematerials
© 2011 Macmillan Publishers Limited. All rights reserved
















![Active Perception based Formation Control for Multiple ...[11], active perception based formation control is addressed using a decentralized non-linear model predictive controller.](https://static.fdocuments.in/doc/165x107/5f085c617e708231d421a0af/active-perception-based-formation-control-for-multiple-11-active-perception.jpg)

