
RNA Molecules and RNA Processing Functions and Modifications of RNA Molecules.

Structure and Dynamics of the Iron ResponsiveElement RNA: Implications for Binding of the RNA byIron Regulatory Binding Proteins
Kenneth J. Addess1, James P. Basilion2, Richard D. Klausner2
Tracey A. Rouault2 and Arthur Pardi1*
1Department of Chemistry andBiochemistry, University ofColorado, BoulderCO 80309-0215, USA2Cell Biology and MetabolismBranch, National Institute ofChild Health DevelopmentNational Institutes of HealthBethesda, MD 20892, USA
The iron responsive element (IRE) is a �30 nucleotide RNA hairpin thatis located in the 50 untranslated region of all ferritin mRNAs and in the 30untranslated region of all transferrin receptor mRNAs. The IREs arebound by two related IRE-binding proteins (IRPs) which help controlintracellular levels of iron by regulating the expression of both ferritinand transferrin receptor genes. Multi-dimensional NMR and compu-tational approaches were used to study the structure and dynamics ofthe IRE RNA in solution. The NMR data are consistent with formation ofA-form helical stem regions, a one-base internal bulge and a Watson-Crick C �G base-pair between the ®rst and ®fth nucleotides in the loop.A superposition of re®ned structures indicates that the conserved C inthe internal bulge, and three residues in the six-nucleotide hairpin loopare quite dynamic in this RNA. The structural roles of the stems, theloop and the bulge in the function of the IRE RNA and in possible inter-actions with the iron regulatory protein are discussed.
# 1997 Academic Press Limited
Keywords: iron regulation; RNA structure; hairpin loops and bulges; IRP;NMR structure*Corresponding author
Introduction
Iron homeostasis is maintained in mammaliancells through post-transcriptional regulation ofgenes coding for ferritin, a protein that sequestersiron from the cytoplasm of eukaryotic cells, andthe transferrin receptor (TfR), a protein that deli-vers iron to the cytoplasm by receptor-mediated
endocytosis (Klausner et al., 1993; Hentze & Kuhn,1996). A cis-acting component of post-transcrip-tional regulation of both genes is a 30-nucleotidehairpin loop RNA known as the iron responsiveelement (IRE; Aziz & Munro, 1987; Hentze et al.,1987). IREs are located within the 50 untranslatedregion of all ferritin mRNAs and the 30 untrans-lated region of all transferrin receptor mRNAs.They are bound with high af®nity by two relatedIRE binding proteins now known as iron regulat-ory protein 1 (IRP1) and iron regulatory protein 2(IRP2; Rouault et al., 1992; Samaniego et al., 1994;Guo et al., 1994). IRP1 is a bifunctional cytosolicprotein with its function determined by the pre-sence or absence of a [4Fe-4S] cluster. When aeukaryotic cell is depleted of iron, the iron-freeform of IRP1 binds with high af®nity to the IRE.When bound to the RNA, the protein represses theexpression of ferritin by preventing the binding oftranslational initiation factors to the 50 cap site ofthe ferritin mRNA (Gray & Hentze, 1994). It simul-taneously increases the expression of the TfR geneby protecting the TfR mRNA against degradationby cellular ribonucleases (Binder et al., 1994).
Phylogenetic comparison and sequence analysisof naturally occurring IREs were used to de®ne a
Present addresses: K. J. Addess, ComputationalBiology Branch, National Center for BiotechnologyInformation, National Library of Medicine, NationalInstitutes of Health, Bethesda, MD 20894, USA;J. P. Basilion, Variagenics, Inc., 1 Kendall Square,Building 400, Cambridge, MA 02139, USA.
Abbreviations used: IRE, iron responsive element;NTP, nucleotide 50-triphosphate; IRP, iron regulatoryprotein; TfR, transferrin receptor; mRNA, messengerRNA; DEAE, diethylaminoethyl; NOE, nuclearOverhauser effect; NOESY, nuclear Overhauser effectspectroscopy; COSY, correlation spectroscopy; TOCSY,total correlation spectroscopy; E.COSY, exclusivecorrelation spectroscopy; HSQC, heteronuclear singlequantum coherence; CT-HSQC, constant timeheteronuclear single quantum coherence; CPMG,Carr-Purcell-Meiboom-Gill sequence; RMSD, root-mean-square deviation.
J. Mol. Biol. (1997) 274, 72±83
0022±2836/97/460072±12 $25.00/0/mb971377 # 1997 Academic Press Limited

28-nucleotide minimal consensus sequence of theRNA, which consists of two stem regions separ-ated by a single bulged cytosine located ®ve base-pairs away from a six-nucleotide loop (Barton et al.,1990; Bettany et al., 1992; Jaffrey et al., 1993; Theil,1994). All known IREs contain a highly conservedsix-base loop of the sequence 50-CAGUGX-30,where X at position 6 can be either an A, C or Ubut never a G. The stem regions of the IRE mustform stable double helices but there is generally nosequence requirement for high-af®nity binding byIRP1. The lower stem varies in the number ofcomplementary base-pairs whereas there is a strictrequirement for ®ve base-pairs in the upper stemfor RNA function (Kikinis et al., 1995; Jaffrey et al.,1993). Chemical protection experiments and muta-tional analysis of the consensus sequence indicatethat the fold of the IRE conforms to its predictedsecondary structure (Bettany et al., 1992; Harrellet al., 1991; Wang et al., 1991, 1990).
Competition studies of the consensus IRE withRNAs containing single mutations in the loop orbulged residues demonstrated that deviations fromthe consensus sequence result in a decrease in thebinding af®nity of the RNA for the protein (Jaffreyet al., 1993). However, these studies did not rigor-ously de®ne the structural and functional roles ofeach of the seven conserved residues. In vitro selec-tion experiments were used to identify alternateRNA ligands for the iron regulatory protein.Initially, Henderson et al. (1996) discovered anotherIRE sequence that binds with high af®nity to IRP1but not IRP2; this IRP1-speci®c ligand containedthe loop sequence 50-UAGUAX-30 where X was a Uor C, but not a G or an A. In this sequence the C atposition 1 is changed to a U and the only otherchange in the sequence is a G to A mutation atposition 5. This co-variation led to the hypothesisthat residues C1 and G5 in the loop form a base-pair in the wild-type sequence which could be animportant structural feature of functional IREs(Henderson et al., 1994). By selecting for ligandsagainst two different IRPs, Butt et al. (1996) notedthat the RNA hairpins selected in a competitivebinding study by IRP1 and IRP2 always containeda C at the bulge position, a G at position 3 of theloop and almost always contained an A at position2 of the loop. Many of the selected sequences alsohad changes at position 4 of the loop, where theconserved U was either an A or a G, implying thatsequence variation at this position did not signi®-cantly interfere with high-af®nity binding. One ofthe goals of the present study is to complement themutagenesis and biochemical studies with directinformation on the conformation of the IRE RNA.
During the past decade, NMR spectroscopy hasdeveloped into an important tool for studying thethree-dimensional structure of RNA in solution(for reviews see Varani & Tinoco, 1991; Pardi,1995). NMR also represents an important probe ofthe dynamics of molecules in solution; for exampleNMR can be used to differentiate those parts of amolecule that are motionally disordered in solution
from those that adopt a precise conformation.Here, we have applied heteronuclear multidimen-sional NMR techniques to study the structure anddynamics of the IRE RNA to help understand thestructural basis for the consensus sequence of theRNA and how the RNA binds to the IRP1.
Results
Sequence design of the IREs used in thestructural study
The sequence and secondary structure of the IREfound in the 50 untranslated region of the humanferritin H-chain mRNA is shown in Figure 1(a)(Klausner et al., 1993). Residues 5 through 25 in thewild-type IRE sequence (Figure 1(b)) being studiedhere, are identical to residues 9 through 29 in thehuman ferritin H-chain IRE. The bottom four base-pairs in the lower stem were chosen to helpimprove the transcriptional ef®ciency with T7RNA polymerase. Though formation of these fourbase-pairs is required to conform to the IRE con-sensus sequence, there is no sequence-speci®crequirement for these base-pairs (Bettany et al.,1992). Thus changes in the sequence of these basepairs have little or no effect on the binding of bothIREs by human IRP1. The only difference betweenthe wild-type and U1A5 IRE RNA hairpins shownin Figure 1 is in the hairpin loop where the wild-type has a 50-CAGUGC-30 loop sequence, whereasthe U1A5 mutant IRE contains substitutions of Ufor C at residue 13 and A for G at residue 17
Figure 1. Nucleotide sequence and secondary structureof (a) the human ferritin H-chain IRE, (b) wild-type and(c) U1A5 mutant iron responsive element (IRE) RNAs.Residues 13 and 17, enclosed in ovals, form a base-pairin the NMR structures (see the text). The binding con-stants of these sequences for the human IRP1 are in therange of 20 to 40 pM (see the text)
Structure of the Iron Responsive Element 73

(Figure 1(c)). Competition assays were used tomeasure the binding af®nity of the IREs, depictedin Figure 1(b) and 1(c), for human IRP1, asdescribed (Haile et al., 1989; Jaffrey et al., 1993).Based on measured IC50 values, the Kd values forbinding of these RNAs to human IRP1 are 20 to 40pM, which is not signi®cantly different from the Kd
for binding of the human ferritin H-chain IRE tohuman IRP1. Thus the sequences used here rep-resent a biologically relevant form of the IRE RNA.
Assignments of exchangeableproton resonances
The one-dimensional (1D) proton NMR spec-trum in H2O of the wild-type IRE RNA is shownin Figure 2(a). The assignments were made on thebasis of sequential NOE connectivities as describedbelow. All of the imino proton resonances pre-dicted by the secondary structure of the IRE RNAare observed in the 1D spectrum except for U19H3which only appears as a broad peak at 14.5 ppm inthe spectrum collected at pH 5.5 (Figure 2(b)).Temperature studies of both the wild-type and theU1A5 IRE show that each RNA is stable because
imino proton resonances are observed up to 55�C(data not shown). A resonance at 10.7 ppm isobserved in the imino proton spectrum of thewild-type IRE but not in the U1A5 IRE RNA (notshown). This resonance sharpens at pH 5.5 andwas assigned as the imino proton of G17 in theloop, based on the guanine-speci®c 15N chemicalshift observed in the 2D (15N,1H) HSQC.
The exchangeable imino and amino protons ofboth the wild-type and U1A5 mutant IRE RNAswere assigned from (15N,1H) HSQC and NOESYspectra in H2O at 15�C. The NOESY spectrum ofthe wild-type IRE is shown in Figure 3. The iminoproton assignments were made from imino protonto imino proton sequential connectivities as illus-trated in Figure 3. Sequential assignments of thelower helix were made starting with the G5 �U25
wobble base-pair which has an extremely strongintra-base-pair NOE (WuÈ thrich, 1986). In the upperstem, only the imino proton to imino proton NOEcross-peak between G21 and U20 is observed; how-
Figure 2. One-dimensional imino proton NMR spectraof the wild-type IRE RNA at (a) pH 6.5 and (b) at pH5.5. The spectra were acquired at 10�C under conditionsdescribed in the text.
Figure 3. (a) Expanded region of the imino, amino andaromatic proton region of the H2O NOESY spectrum ofthe wild-type IRE RNA at 10�C. Resonance assignmentof the imino protons and the A3H2 and A11H2 areshown. (b) Imino proton to imino proton sequentialNOE connectivities are illustrated. The spectrum wasacquired with a sweep width of 12,000 Hz in bothdimensions, 300 complex t1 points of 80 scans and 2048complex t2 points.
74 Structure of the Iron Responsive Element

ever, sequential NOE connectivities between theC10 �G21 and the U9 �G22 base pairs were observedin a (15N, 1H) CPMG-NOESY spectrum (data notshown). The G17 imino proton showed a 1D NOEto a C amino proton at 8.25 ppm indicating thatG17 forms a Watson-Crick base-pair with C13. Thisbase-pair has been observed in a previous NMRstudy of the upper stem and loop of the IRE (Laing& Hall, 1996).
Assignment of the non-exchangeableproton resonances
Recently developed triple resonance experimentsthat correlate exchangeable and non-exchangeablebase protons in nucleic acids were next used toextend assignment of the exchangeable base pro-tons to the H6 and H8 resonances for most of theresidues in the upper and lower stems of the IRE(Simorre et al., 1996a,b, 1995). Pyrimidine H6 pro-ton resonances were identi®ed from the HNCCCHexperiments and purine H8 resonances were ident-i®ed from the HNC-TOCSY-CH experiments. TheAH2 resonance assignments were also used to helpidentify the AH8 resonances. Two of these, A3H2and A11H2, were identi®ed by strong NOEs to theU27H3 and U20H3 protons, respectively, both ofwhich are indicated in the H2O NOESY shown inthe top half of Figure 3. A23H2 was identi®ed fromits NOE cross-peak to the U8H3 in the H2ONOESY spectrum at 1�C (data not shown). TheA12H2 and A14H2 resonances were assigned fromthe (1H,13C) HSQC spectrum and from the 3DNOESY-HSQC spectrum.
Assignment of the ribose proton resonancesbegan with H10-C10 correlations in a C20 decoupled(1H,13C) HSQC spectrum. Once all 29 H10-C10 cor-relations were identi®ed, the other ribose protonswere assigned from a combination of (13C,13C,1H)HCCH-TOCSY, 3D (1H,13C,1H) HCCH-TOCSY and3D (1H,13C,1H) HCCH-COSY experiments (Pardi,1995). Aromatic to H10 and aromatic to H20sequential assignments were made from the 2Dand 3D NOESY spectra using previously describedmethods (Nikonowicz & Pardi, 1993; WuÈ thrich,1986). The intensity of the G15H8-G15H10 cross-peak is much stronger than all the other aromaticto H10 cross-peaks and comparable to the H5-H6cross-peaks of U5 and C26 (data not shown). In ashort mixing time NOESY spectrum, the volume ofthis cross-peak integrates to a distance of approxi-mately 2.5 AÊ and is twice the volume of the G15H8to H20 cross-peak, indicating the glycosidic torsionangle for G15 is in the syn conformation.
Conformation of the sugars
The homonuclear three bond 3JH10,H20 couplingconstants in Table 1 were measured directly from a3D HCCH-E.COSY spectrum (Schwalbe et al.,1994). G15 and U16 have coupling constants greaterthan 10 Hz and therefore the ribose sugars of thesetwo residues adopt primarily S-type puckers(Altona, 1982). G1, G6, C7, G17 and C18 have valuesless than 10 Hz but greater than 2.0 Hz and aretherefore interconverting between S-type and N-type sugar puckers. The ribose sugars of the other22 residues have 3JH10,H20 coupling constants42.0 Hz, which is characteristic of an N-type orC30-endo conformation.
Structure calculations
A family of 15 re®ned, ®nal structures of theIRE-1 were generated from 50 starting structures,as described in Materials and Methods. The three-dimensional structure of the RNA was calculatedwith 314 experimental NOE restraints, 153 torsionangle restraints and 54 base-pair constraints (seeTable 2). No other restraints were included in themolecular dynamics calculations, and loose upperand lower bounds were applied to all NOErestraints to re¯ect the dynamic behavior of theRNA, as discussed below.
The relative orientation of the upper and lowerstems is not well-de®ned because superposition offull structures led to an average pairwise RMSD of4.0 AÊ ; however, the local pairwise RMSDs revealthat the upper and lower stems form well-de®nedstructural units. The average pairwise RMSDs foreither the upper or lower stem increases when the
Table 1. 3JH10 ,H20 coupling constants measured in the wild-type IRE RNA
Residue G1 G2 A3 G4 U5 G6 C7 U8 U9 C10 A11 A12 C13 A14 G15
3JH10 ,H20 (Hz) 5.7 <2.0 <2.0 <2.0 <2.0 5.8 6.0 <2.0 N.O. <2.0 N.O. N.O. <2.0 <2.0 11.6Residue U16 G17 C18 U19 U20 G21 G22 A23 C24 G25 C26 U27 C28 C293JH10 ,H20 (Hz) 10.5 5.7 5.6 <2.0 <2.0 <2.0 <2.0 <2.0 <2.0 <2.0 <2.0 <2.0 N.O. N.O.
N.O. indicates that these coupling constants could not be observed.
Table 2. Summary of NMR derived-distance and torsionangle restraints and root-mean-squared deviations forthe ®nal structures of the wild-type IRE RNA
NOE restraints 314Interresidue 133Intraresidue 181Torsion angle restraints 153Base-pairing restraints for upper and lowerstems 54
RMSDa (AÊ )Lower stem (residues 1±6, 24±29) 1.71 � 0.44Lower stem and bulge (residues 1±7, 24±29) 2.14 � 0.50Upper stem (residues 8±12, 19±23) 1.63 � 0.37Upper stem and bulge (residues 7±12, 19±23) 2.19 � 0.46Loop residues 13, 14, 17 1.09 � 0.29Loop residues 13±18 2.44 � 0.92
a These are the average pairwise RMSDs of the 15 re®ned struc-tures.
Structure of the Iron Responsive Element 75
bulged C is included in the calculation (Table 2).This indicates that the conformation of the bulgedC is not well de®ned. For the loop, the averagepairwise RMSD for C13, A14 and G17 is low butincreases signi®cantly when G15, U16 and C18 areincluded in the calculation. This indicates that theconformation of loop residues C13, A14 and G17 arewell de®ned, whereas G15, U16 and C18 are disor-dered in the structure.
Rotating spin-lattice relaxation times for C10sugar resonances
The rotating frame spin-lattice relaxation times,T1r, were measured using a series of 1H,13C HSQC
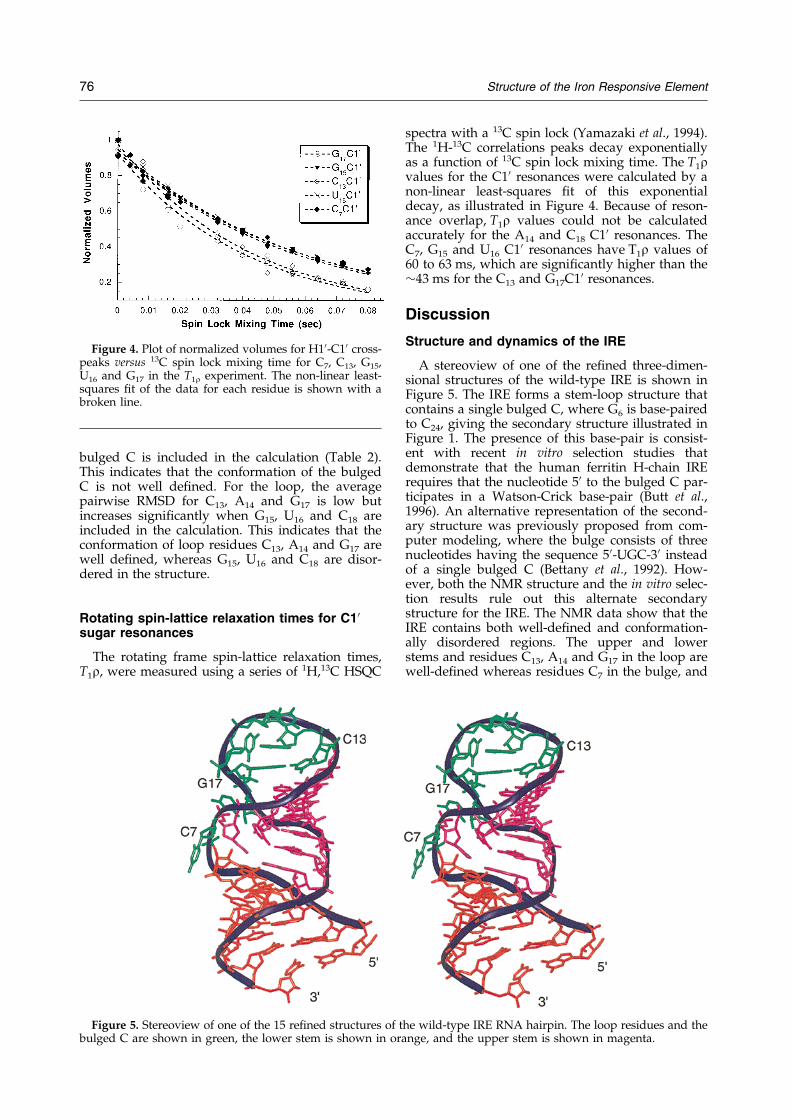
spectra with a 13C spin lock (Yamazaki et al., 1994).The 1H-13C correlations peaks decay exponentiallyas a function of 13C spin lock mixing time. The T1rvalues for the C10 resonances were calculated by anon-linear least-squares ®t of this exponentialdecay, as illustrated in Figure 4. Because of reson-ance overlap, T1r values could not be calculatedaccurately for the A14 and C18 C10 resonances. TheC7, G15 and U16 C10 resonances have T1r values of60 to 63 ms, which are signi®cantly higher than the�43 ms for the C13 and G17C10 resonances.
Discussion
Structure and dynamics of the IRE
A stereoview of one of the re®ned three-dimen-sional structures of the wild-type IRE is shown inFigure 5. The IRE forms a stem-loop structure thatcontains a single bulged C, where G6 is base-pairedto C24, giving the secondary structure illustrated inFigure 1. The presence of this base-pair is consist-ent with recent in vitro selection studies thatdemonstrate that the human ferritin H-chain IRErequires that the nucleotide 50 to the bulged C par-ticipates in a Watson-Crick base-pair (Butt et al.,1996). An alternative representation of the second-ary structure was previously proposed from com-puter modeling, where the bulge consists of threenucleotides having the sequence 50-UGC-30 insteadof a single bulged C (Bettany et al., 1992). How-ever, both the NMR structure and the in vitro selec-tion results rule out this alternate secondarystructure for the IRE. The NMR data show that theIRE contains both well-de®ned and conformation-ally disordered regions. The upper and lowerstems and residues C13, A14 and G17 in the loop arewell-de®ned whereas residues C7 in the bulge, and
Figure 4. Plot of normalized volumes for H10-C10 cross-peaks versus 13C spin lock mixing time for C7, C13, G15,U16 and G17 in the T1r experiment. The non-linear least-squares ®t of the data for each residue is shown with abroken line.
Figure 5. Stereoview of one of the 15 re®ned structures of the wild-type IRE RNA hairpin. The loop residues and thebulged C are shown in green, the lower stem is shown in orange, and the upper stem is shown in magenta.
76 Structure of the Iron Responsive Element

G15, U16 and C18 in the loop are not well-de®ned inthe structure.
To investigate more thoroughly the motion ofthe residues in the bulge and loop regions of theIRE, the T1r relaxation times were measured in thewild-type IRE. The T1r relaxation times for the C10resonances on residues C7, G15 and U16 are signi®-cantly longer than the T1r values for C10 reson-ances on other residues (�60 versus �43 ms). Thisindicates that the conformations of the bulged C7
residue, and loop G15, U16 residues are dynamic inthe IRE, whereas the other loop residues haverelaxation times similar to resonances in the stemregions. The difference in T1r values between ¯ex-ible residues and well-de®ned residues correlateswell with the local RMSDs reported in Table 2.
Structure and function of the lower and upperstems of the IRE-1
The re®ned average structures of both lower andupper stems are shown in Figures 6 and 7 andreveal that these regions of the IRE have an A-formconformation (Saenger, 1984). For the upper andlower stems, interresidue NOEs from the H20 pro-ton of residue n to the H6, H8 protons of residuen � 1 are stronger than the intraresidue NOEsfrom pyrimidine H5 proton to H6 proton, charac-teristic of the A-form conformation. All the basesin the stem regions have anti glycosidic torsionangles and all the sugar puckers, with the excep-tion of terminal G1 and G6, are N-type (near C30-endo). These upper and lower stems are locallywell de®ned but do not have a ®xed orientationwith respect to each other. This suggests that thejunction between the two stems functions as a ¯ex-ible hinge that could allow the RNA to undergo aglobal conformational change upon binding by theIRP. The U imino proton resonance of the U8 �A23
base-pair is not observed in a 2D NOESY spectrum
above 1�C; the intensity imino to imino NOE cross-peak of the U9 �G22 base-pair in the upper stem isweaker than that of the U5 �G25 base-pair in thelower stem. This suggests that the imino protons ofthe A �U and the G �U base-pairs above the bulgedC have faster solvent exchange rates than the otherinternal base-pairs in the stem regions of the RNA.This supports the notion that the junction betweenthe two stems is ¯exible.
The variation in length and lack of sequencespeci®city of the lower stem suggests that bases inthe lower stem do not interact directly with theprotein in the complex. However, in vitro selectionexperiments showed that formation of the G6 �C24
base-pair adjacent to the bulged C in the lowerstem favors high-af®nity binding by the IRP1 andIRP2 (Butt et al., 1996). Positioned between thebulge and the loop, the upper stem plays a moredirect role in binding and regulation where a ®ve-base-pair stem is required for iron-dependent regu-lation (Kikinis et al., 1995). However, there is nosequence conservation for these base-pairs. Thislength requirement of the upper stem combinedwith the lack of sequence speci®city for these base-pairs suggest that it functions primarily as a ``mol-ecular ruler''. The upper stem would then providethe correct distance and spatial orientation betweenthe bulged C7 and G15 in the loop. In this modelwe propose that the bases of both C7 and G15 makesequence-speci®c interactions with the IRP.
Conformation and function of the bulged Cand hairpin loop
Figure 6 shows that the conformation of thebulged C7 is not well-de®ned in the three-dimen-sional structures of wild-type IRE RNA. Deletionof this residue from RNA results in a �400-folddecrease in the binding af®nity as compared to thewild-type RNA (Jaffrey et al., 1993). All RNAs in
Figure 6. The relative positions of the bulged C7 residue from the 15 low-energy structures of the wild-type IREwhen superimposed on the average structure of the lower stem.
Structure of the Iron Responsive Element 77

the in vitro selection experiments with IRP1 andIRP2 contain a cytosine at the bulge position (Buttet al., 1996). This strongly suggests that the bulgedC interacts directly with the protein in a sequence-speci®c manner; for example, the bulged C mayform a hydrogen bond with a side-chain residue inthe protein.
A superposition of the loop residues from thefamily of ®nal converged structures overlaid withthe average structure of the upper stem is dis-played in Figure 7. Three residues in the loop, C13,A14 and G17, have well-de®ned conformationswhereas these for G15, U16 and C18 are not well-de®ned. Of the three ¯exible residues in the loop,only G15 is required for high-af®nity binding of theRNA by the protein. The intensity of the G15H8 toG15H10 NOE cross-peak is much stronger than theother intraresidue H8, H6 to H10 NOE cross-peaksin this RNA. Although different relaxation proper-ties of G15 could contribute to the strong intensityof this H8 to H10 NOE cross-peak, the intensity ofthis cross-peak is twice that of the G15H8 to G15H20NOE cross-peak. Furthermore, the sugar pucker ofG15 is S-type (near C20-endo). Taken together, theNMR results indicate that G15 adopts a syn-typeglycosidic angle within the range of motion of thisresidue. This syn conformation for G15 might berequired for sequence-speci®c recognition of theRNA by the protein.
In the solution structure, C13 and G17 are basepaired to each other. A14, the other nucleotide inthe loop that is well-de®ned, stacks directly aboveG17 (Figure 8). In almost all cases, the A is requiredat this position for high-af®nity binding to the pro-tein. A single point mutation of an A to a G in the
IRE of the L-ferritin mRNA results in hyperferriti-nemia and bilateral cataract formation in humans(Beaumont et al., 1995). The observation that the Ato G mutation leads to development of a humandisease clearly indicates that A14 is important forIRE function, though phylogenetic and biochemicalevidence have not clearly de®ned its structuralrole. Similarly a mutation of G15 to C also resultsin severe cataract formation and hyperferritinemiain patients that carry this mutation (Girelli et al.,1995). There are several indications that loop resi-dues 1 (C13) and 5 (G17) do not interact directlywith the protein but instead are important for thestructure and stability of the loop. If C13 or G17
make base-speci®c contacts with the protein, it isunlikely that the U1A5 mutant IRE would havebeen selected as a high-af®nity RNA ligand for theIRP1. UV melting studies of an IRE containing the
Figure 7. Stereoview illustrating the variation in the conformations of the loop residues of the re®ned structures ofthe wild-type IRE. The structures were superimposed on the average structure of the upper stem. Residues in theaverage structure are shown in magenta, residues C13, A14 and G17 are in black, G15 in orange, U16 in green and C18
in cyan.
Figure 8. Conformation of C13, A14 and G17 vieweddown the helical axis, which shows how A14 stacksdirectly over G17. This Figure was generated with MOL-SCRIPT (Kraulis, 1991).
78 Structure of the Iron Responsive Element

loop sequence 50-CAGUAC-30 lowers the Tm of theRNA (Laing & Hall, 1996; Sierzputowska-Graczet al., 1995). This mutant is not bound by the IRP1with high af®nity (Jaffrey et al., 1993). This indi-cates that presence of this base-pair stabilizes thelocal conformation of the loop which is necessaryfor high-af®nity binding and iron-dependent regu-lation. Formation of the C13 �G17 base-pair couldexplain why residue 18 cannot be a G. A G at pos-ition 18 would likely base-pair with C13 and there-fore disrupt the base-pair between G17 and C13.This would lead to a different conformation of theloop which could then result in loss of function.
Comparison with the U1A5 mutant
The loop regions of both wild-type and U1A5
mutant IRE RNAs are shown in Figure 9. Overall,the structural characteristics of the loop regions ofthese two IREs are nearly identical. Analogous tothe wild-type sequence, the conformation of thebulged C in the U1A5 mutant IRE is not wellde®ned. This suggests that the conformation of thebulged C is the same in a complex of either thewild-type or U1A5 mutant IRE with the IRP. Forthe loop region, G15, U16 and C18 are not well-de®ned and have increased dynamics from C10 T1rmeasurements, whereas U13, A14 and A17 are well-de®ned. Identical with the wild-type sequence, G15
in the U1A5 RNA adopts a syn conformation.In the NOESY spectrum of the U1A5 mutant IRE,
A17H2 has NOEs to A14H10 and to U19H10. This isconsistent with a U13 �A17 base-pair in the loop,which is analogous to the C13 �G17 base pair of thewild-type IRE. However, no imino proton reson-ance was observed for U13, therefore this iminoproton is exchanging rapidly with solvent. Withoutdirect evidence for base-pairing in the loop, twoseparate structure calculations were performed. Inthe ®rst set, the base-pair between residue U13 andresidue A17 was restrained. As expected, this base-pair was formed in all the ®nal structures andthese restraints did not introduce any signi®cantdistance violations. In the second set of calculationsno restraints were included for this base-pair, and30% of the ®nal structures contained the base-pairwhereas in the other structures the U13 and A17
were not well-de®ned. The T1r data are consistentwith formation of the U13 �A17 base-pair and
together these results indicate that a U13 �A17 base-pair occurs in the structure of the U1A5 mutantIRE, as suggested by the in vitro selection exper-iments (Henderson et al., 1994).
Comparison with other RNAs
The simultaneous presence of well-de®ned and¯exible residues in the three-dimensional structureof the IRE has also been observed in the NMRstructural studies of other RNAs, including theRev response element (Battiste et al., 1995, 1994;Peterson et al., 1994), the 30 untranslated region ofthe U1A pre-RNA (Allain et al., 1996), the TARRNA (Puglisi et al., 1993), an ATP-binding aptamerRNA (Dieckmann et al., 1996; Jiang et al., 1996),and a theophylline-binding RNA aptamer(Zimmermann et al., 1997). For these RNAs, theresidues critical to binding often appear disorderedin the free RNA and become well-ordered uponcomplex formation. Thus we expect that the disor-dered residues C7 and G15 in the IRE become wellde®ned upon complex formation. However, wewould not expect that the well-de®ned residues,C13, A14 and G17, will undergo a major confor-mational change upon complex formation.
Interaction of the IRE RNA with the IRP
The 4Fe-4S cluster form of IRP1 is a cytosolicaconitase (Kennedy et al., 1992), which catalyzesthe isomerization of citrate to isocitrate, but cannotbind IREs with high af®nity (Haile et al., 1989).Restoration of RNA-binding activity and destruc-tion of aconitase activity occurs only when the 4Fe-4S cluster dissociates from the IRP1 in iron-depleted cells. The X-ray crystal structure of thepig heart mitochondrial aconitase has been solved(Robbins & Stout, 1989) and this protein has 30%sequence identity with human IRP1 (Rouault et al.,1991). The high sequence identity between IRP andmitochondrial aconitase suggests similar structuresfor both proteins. Unfortunately, there is no X-raycrystal structure of the apoprotein and it is notpossible to dock the IRE RNA into the structure ofthe aconitase with the 4Fe-4S cluster in a way thatis consistent with the NMR structure observedhere and the mutagenesis data on the IRP1. Never-theless, the NMR data combined with the bio-chemical results provide insights into how the IRPcould interact with the IRE. UV-cross-linking stu-dies (Basilion et al., 1994; Swenson & Walden,1994) demonstrated that the IRE hairpin interactswith residues 121 to 130 in human IRP1. In mito-chondrial aconitase, four active site arginine resi-dues are involved in substrate binding andspeci®city (Zheng et al., 1992; Lauble et al., 1992).All four active site arginine residues are found inIRP1, and site-directed mutagenesis reveals thatthree of these arginine residues are important inIRE binding (Butt et al., 1996; Philpott et al., 1993).In IRP1, mutagenesis of arginine 541 or 780 to glu-tamine (residues 452 and 644 in pig heart mito-
Figure 9. Structures of the loop region from (a) thewild-type IRE and (b) U1A5 IRE RNA hairpins. Residues13 and 17 in the loop are shown in gray and form aWatson-Crick base pair in both of these sequences (seethe text).
Structure of the Iron Responsive Element 79

chondrial aconitase) results in a 103 to 104-folddecrease in binding af®nity (Philpott et al., 1994).Arginine to guanine hydrogen bonds have beenobserved in the three-dimensional structure of sev-eral protein-DNA complexes, including zif268(Pavletich & Pabo, 1991), the glucocorticoid recep-tor (Luisi et al., 1991), and Escherichia coli Trprepressor (Otwinowski et al., 1988), and it is poss-ible that G15 in the IRE forms a speci®c interactionwith an active site arginine in the IRP. When the4Fe-4S cluster is bound to the protein, the bindingcleft, situated between domains 1 to 3 and domain4, is in a ``closed'' state, which permits arginine541 or 780 to bind isocitrate (Lauble et al., 1992)but prevents these amino acid residues from inter-action with residues in the hairpin loop of the IRE.This may be one way the presence of the 4Fe-4Scluster inhibits high af®nity binding of the RNA. Itis likely that these arginine residues become acces-sible to the RNA when the cleft is opened up by aconformational change in the hinge linker peptidethat connects domain 3 and domain 4 (Lauble et al.,1992; Basilion et al., 1994). Recently, Schalinske et al.(1997) demonstrated an increase in the rate of pro-teolysis of sites near the proposed binding cleftand hinge linker regions of the protein when the4Fe-4S cluster was removed. This indicates that thecleft cannot exist in this ``open'' state without dis-sociation of the 4Fe-4S cluster.
The crystal structure of a complex of the coatprotein from MS2 bacteriophage with its operatorRNA has been determined (Valegard et al., 1994)and the RNA has a hairpin-single base bulge motifthat has a qualitatively similar structure to the IRERNA (Borer et al., 1995). For the MS2 complex, theloop and bulged residues of the operator RNA thatare essential for binding form hydrogen bondand/or stacking interactions with conserved aminoacid residues (Valegard et al., 1994). In addition,the protein makes speci®c contacts with the phos-phate backbone of the RNA. We expect that similartypes of interactions will be observed in the IRP1-IRE complex. We are currently testing this hypoth-esis by photo-cross-linking studies of the protein-RNA complex and modi®cations of the criticalnucleotides in the IRE RNA hairpin.
Materials and Methods
Sample preparation
The wild-type and U1A5 IRE RNAs shown in Figure 1were synthesized by in vitro transcription on a single-stranded DNA template using phage T7 RNA polymer-ase. The transcription reactions were performed usingstandard conditions except each reaction contained atotal NTP concentration of 8 mM (Milligan et al., 1987).The 13C/15N NTPs were prepared as described(Nikonowicz et al., 1992; Batey et al., 1992). The crudeRNA from each transcription reaction was puri®ed bypreparative gel electrophoresis with a 20% (w/v) dena-turing gel containing 7 M urea. For each gel, the productband was visualized with UV shadowing and thenexcised from the gel. The RNA was recovered from the
excised product band by electroelution and ethanol pre-cipitation of the electroeluant. The RNA was further pur-i®ed by ion-exchange DEAE Sephacel chromatography.Fractions from the ion-exchange column containing theRNA were then dialyzed extensively against an NMRbuffer solution of 10 mM sodium phosphate (pH 6.5),10 mM NaCl, 0.2 mM EDTA. A 5 mg sample of puri®ed13C/15N-labeled wild-type IRE was synthesized from a50 ml transcription which yielded a 1.5 mM sample in350 ml. A 4.5 mg sample of the puri®ed, uniformly13C/15N-labeled, mutant U1A5 IRE was obtained from a40 ml transcription reaction yielding a 1.2 mM sample in350 ml. Before acquisition of NMR data, the sampleswere heated to 85�C for two minutes and then snap-cooled on ice to induce formation of the RNA hairpin.
NMR Spectroscopy
All NMR experiments were performed at 500 MHz oneither Varian VXR or UnityPlus instruments equippedwith pulsed-®eld gradients. All spectra were trans-formed with the program FELIX version 2.30 or 95.0(Biosym/ MSI, San Diego, CA). A variety of homo- andheteronuclear magnetic resonance experiments wereused to make resonance assignments and to generatestructural constraints for the IRE RNAs including: 2D(1H,1H) NOESY (Kumar et al., 1980) and (1H,1H) DQF-COSY (Rance et al., 1983); 2D (15N,1H) HSQC, 2D(13C,1H) HSQC, 2D (15N,1H) HNCCCH (Simorre et al.,1995), 2D (15N,1H) HNC-TOCSY-CH (Simorre et al.,1996a,b), 2D (15N,1H) CPMG-NOESY (Mueller et al.,1995), 3D (1H,13C,1H) HCCH-E.COSY (Schwalbe et al.,1994), 3D (13C,13C,1H) HCCH-TOCSY, 3D (1H,13C,1H)HCCH-TOCSY, 3D (1H,13C,1H) HCCH-COSY, and 3D(1H,13C,1H) NOESY-HSQC (Pardi, 1995) experiments. Forall two-dimensional and three-dimensional heteronuclearexperiments, 13C or 15N WALTZ or GARP broadbanddecoupling was applied during the acquisition period.A DIPSI-2 mixing scheme was applied during the 25 ms13C spin lock period in the HCCH-TOCSY experiments.In the NOESY and HSQC spectra acquired in 90% H2O/10% 2H2O, water suppression was performed with either11 echo (SklenaÂr & Bax, 1987) or ¯ip-back WATERGATEpulses (Piotto et al., 1992).
Structure determination
For each RNA molecule, interproton distancerestraints were obtained from three 3D (1H,13C,1H)NOESY-HSQC spectra at mixing times of 75, 180 and300 ms. NOE cross-peak intensities were classi®ed asstrong if their intensities were greater than or equal tothose of the pyrimidine H5 to H6 cross-peaks in the75 ms 3D NOESY spectrum, medium if they wereobserved in the 75 ms and the 180 ms mixing time 3DNOESY spectra and their intensities were weaker thanthe pyrimidine H5 to H6 cross-peak, and weak if theywere observed only in the 300 ms mixing time 3DNOESY spectrum. The lower distance bounds for allinterproton distance pairs were set to 1.8 AÊ . Upper dis-tance bounds of 3.5, 4.5 and 5.5 AÊ were applied forstrong, medium and weak NOE cross-peaks, respect-ively. An additional upper bound of 1.0 AÊ was appliedto NOE cross-peaks involving exchangeable protons inthe NOESY spectrum in H2O at 10�C. Hydrogen-bond-ing constraints were included for the 11 base-pairs of theupper and lower stems, as described (Jucker et al., 1996).Sugar pucker conformations were classi®ed as N-type(near C30-endo), S-type (near C20-endo) or mixed confor-
80 Structure of the Iron Responsive Element

mation based on the JH10 ,H20 coupling constants (Altona,1982) measured in a 3D (1H,13C,1H) HCCH-E.COSYspectrum. For sugars falling into either N-type or S-typeconformations the n0 to n4 endocyclic torsion angles wererestrained to published values and no restraints wereincluded for sugars showing mixed conformation(Saenger, 1984). For residues with resolved H20-C20 cor-relation peaks in a spin echo difference CT-HSQC spec-trum, the e torsion angles were calculated frommeasured 3JC20P coupling constants (Legault et al., 1995).Most of these coupling constants were obtained for resi-dues in the loop and the bulge.
All calculations were performed on a Silicon GraphicsIndigo2 workstation using X-PLOR version 3.1 (BruÈ nger,1992). A total of 50 initial structures were generated byrandomizing the backbone torsion angles of the linear-ized 29mer RNA molecule. Re®nement of these startingstructures occurred in three stages. In the ®rst stage, themolecules were subjected to a simulated annealing pro-tocol, where each structure was heated to 3000 K andthen cooled to 300 K over 15 ps of restrained moleculardynamics. This allowed the RNA to fold into its globalhairpin conformation. During this stage, only experimen-tal distance and hydrogen bonding restraints were used.In the second stage, experimental dihedral anglerestraints were included in the re®nement procedure,which consisted of 2.5 ps of simulated annealing at3000 K after which the bath was cooled to 300 K over6.25 ps. In the ®nal step, the re®ned structures under-went 1000 steps of restrained energy minimization. Ofthe 50 starting structures, the 15 lowest energy structuresthat had no violations greater than 0.3 AÊ were selectedfor analysis and all these structures had a total potentialenergy of less than ÿ100 kcal/mol. Coordinates of the®nal structures have been deposited in the Protein DataBank under accession number 1AQO.
Acknowledgments
This work was supported in part by NIH grantAI33098 to A. P. and NIH NRSA Fellowship GM16577to K. J. A. We thank the Colorado RNA Center for theirsupport of RNA science on the Boulder campus.
References
Allain, F. H., Gubser, C. C., Howe, P. W., Nagai, K.,Neuhaus, D. & Varani, G. (1996). Speci®city of ribo-nucleoprotein interaction determined by RNA fold-ing during complex formulation. Nature, 380, 646±650.
Altona, C. (1982). Conformational analysis of nucleicacids. Determination of backbone geometry ofsingle-helical RNA and DNA in aqueous solution.Rec. Trav. Chim. Pays-Bas. 101, 413±432.
Aziz, N. & Munro, H. N. (1987). Iron regulates ferritinmRNA translation through a segment of its 50untranslated region. Proc. Natl Acad. Sci. USA, 84,8478±82.
Barton, H. A., Eisenstein, R. S., Bomford, A. & Munro,H. N. (1990). Determinants of the interactionbetween the iron-responsive element-binding pro-tein and its binding site in rat L-ferritin mRNA.J. Biol. Chem. 265, 7000±7008.
Basilion, J. P., Rouault, T. A., Massinople, C. M.,Klausner, R. D. & Burgess, W. H. (1994). The iron-
responsive element-binding protein: localization ofthe RNA-binding site to the aconitase active-sitecleft. Proc. Natl Acad. Sci. USA, 91, 574±8.
Batey, R. T., Inada, M., Kujawinski, E., Puglisi, J. D. &Williamson, J. R. (1992). Preparation of isotopicallylabeled ribonucleotides for multidimensional NMRspectroscopy of RNA. Nucl. Acids Res. 20, 4515±4523.
Battiste, J. L., Tan, R., Frankel, A. D. & Williamson, J. R.(1994). Binding of an HIV Rev peptide to Revresponsive element RNA induces formation of pur-ine-purine base pairs. Biochemistry, 33, 2741±2747.
Battiste, J. L., Tan, R., Frankel, A. D. & Williamson, J. R.(1995). Assignment and modeling of the Revresponse element RNA bound to a Rev peptideusing 13C-heteronuclear NMR. J. Biomol. NMR, 6,375±389.
Beaumont, C., Leneuve, P., Devaux, I., Scoazec, J. Y.,Berthier, M., Loiseau, M. N., Grandchamp, B. &Bonneau, D. (1995). Mutation in the iron responsiveelement of the L ferritin mRNA in a family withdominant hyperferritinaemia and cataract. NatureGenet. 11, 444±446.
Bettany, A. J., Eisenstein, R. S. & Munro, H. N. (1992).Mutagenesis of the iron-regulatory element furtherde®nes a role for RNA secondary structure in theregulation of ferritin and transferrin receptorexpression. J. Biol. Chem. 267, 16531±16537.
Binder, R., Horowitz, J. A., Basilion, J. P., Koeller, D. M.,Klausner, R. D. & Harford, J. B. (1994). Evidencethat the pathway of transferrin receptor messenger-RNA degradation involves an endonucleolytic clea-vage within the 30 UTR and does not involvepoly(A) tail shortening. EMBO J. 13, 1969±1980.
Borer, P. N., Lin, Y., Wang, S., Roggenbuck, M. W.,Gott, J. M., Uhlenbeck, O. C. & Pelczer, I. (1995).Proton NMR and structural features of a 24±nucleo-tide RNA hairpin. Biochemistry, 34, 6488±6503.
BruÈ nger, A. T. (1992). X-PLOR, Version 3.1: A System forX-ray Crystallography and NMR, Yale UniversityPress, New Haven, CT.
Butt, J., Kim, H. Y., Basilion, J. P., Cohen, S., Iwai, K.,Philpott, C. C., Altschul, S., Klausner, R. D. &Rouault, T. A. (1996). Differences in the RNA bind-ing sites of iron regulatory proteins and potentialtarget diversity. Proc. Natl Acad. Sci. USA, 93, 4345±4349.
Dieckmann, T., Suzuki, E., Nakamura, G. K. & Feigon, J.(1996). Solution structure of an ATP-binding RNAaptamer reveals a novel fold. RNA, 2, 628±640.
Girelli, D., Corrocher, R., Bisceglia, L., Olivieri, O.,Defranceschi, L., Zelante, L. & Gasparini, P.(1995). Molecular-basis for the recently describedhereditary hyperferritinemia cataract syndrome: amutation in the iron-responsive element of ferritinL-subunit gene (the Verona Mutation). Blood, 86,4050±4053.
Gray, N. K. & Hentze, M. W. (1994). Iron regulatoryprotein prevents binding of the 43 S translation pre-initiation complex to ferritin and eALAS messenger-RNAs. EMBO J. 13, 3882±3891.
Guo, B., Yu, Y. & Leibold, E. A. (1994). Iron regulatescytoplasmic levels of a novel iron-responsiveelement-binding protein without aconitase activity.J. Biol. Chem. 269, 24252±24260.
Haile, D. J., Hentze, M. W., Rouault, T. A., Harford,J. B. & Klausner, R. D. (1989). Regulation of inter-action of the iron-responsive element binding pro-
Structure of the Iron Responsive Element 81

tein with iron-responsive RNA elements. Mol. Cell.Biol. 9, 5055±5061.
Harrell, C. M., McKenzie, A. R., Patino, M. M., Walden,W. E. & Theil, E. C. (1991). Ferritin mRNA: inter-actions of iron regulatory element with translationalregulator protein P-90 and the effect on base-paired¯anking regions. Proc. Natl Acad. Sci. USA, 88,4166±4170.
Henderson, B. R., Menotti, E., Bonnard, C. & Kuhn, L. C.(1994). Optimal sequence and structure of iron-responsive elements. Selection of RNA stem-loopswith high af®nity for iron regulatory factor. J. Biol.Chem. 269, 17481±17489.
Henderson, B. R., Menotti, E. & Kuhn, L. C. (1996). Ironregulatory proteins 1 and 2 bind distinct sets ofRNA target sequences. J. Biol. Chem. 271, 4900±4908.
Hentze, M. W. & Kuhn, L. C. (1996). Molecular controlof vertebrate iron metabolism: mRNA-based regu-latory circuits operated by iron, nitric oxide, andoxidative stress. Proc. Natl Acad. Sci. USA, 93, 8175±8182.
Hentze, M. W., Caughman, S. W., Rouault, T. A.,Barriocanal, J. G., Dancis, A., Harford, J. B. &Klausner, R. D. (1987). Identi®cation of the iron-responsive element for the translational regulationof human ferritin mRNA. Science, 238, 1570±1573.
Jaffrey, S. R., Haile, D. J., Klausner, R. D. & Harford, J. B.(1993). The interaction between the iron-responsiveelement binding protein and its cognate RNA ishighly dependent upon both RNA sequence andstructure. Nucl. Acids Res. 21, 4627±4631.
Jiang, F., Kumar, R. A., Jones, R. A. & Patel, D. J. (1996).Structural basis of RNA folding and recognition inan AMP-RNA aptamer complex. Nature, 382, 183±186.
Jucker, F. M., Heus, H. A., Yip, P. F., Moors, E. H. M. &Pardi, A. (1996). A network of heterogeneoushydrogen bonds in GNRA tetraloops. J. Mol. Biol.264, 968±980.
Kennedy, M. C., Mendemueller, L., Blondin, G. A. &Beinert, H. (1992). Puri®cation and characterizationof cytosolic aconitase from beef-liver and itsrelationship to the iron-responsive element binding-protein. Proc. Natl Acad. Sci. USA, 89, 11730±11734.
Kikinis, Z., Eisenstein, R. S., Bettany, A. J. & Munro,H. N. (1995). Role of RNA secondary structure ofthe iron-responsive element in translational regu-lation of ferritin synthesis. Nucl. Acids Res. 23,4190±4195.
Klausner, R. D., Rouault, T. A. & Harford, J. B. (1993).Regulating the fate of mRNA: the control of cellulariron metabolism. Cell, 72, 19±28.
Kraulis, P. J. (1991). MOLSCRIPT: a program to produceboth detailed and schematic plots of proteinstructures. J. Appl. Crystallog. 24, 946±950.
Kumar, A., Ernst, R. R. & WuÈ thrich, K. (1980). A two-dimensional nuclear Overhauser enhancement (2DNOE) experiment for the elucidation of completeproton-proton cross-relaxation networks in biologi-cal macromolecules. Biochem. Biophys. Res. Commun.95, 1±6.
Laing, L. G. & Hall, K. B. (1996). A model of the ironresponsive element RNA hairpin loop structuredetermined from NMR and thermodynamic data.Biochemistry, 35, 13586±13596.
Lauble, H., Kennedy, M. C., Beinert, H. & Stout, C. D.(1992). Crystal-structures of aconitase with isocitrate
and nitroisocitrate bound. Biochemistry, 31, 2735±2748.
Legault, P., Jucker, F. M. & Pardi, A. (1995). Improvedmeasurement of 13C-31P coupling constants in isoto-pically labeled RNA. FEBS Letters, 362, 156±160.
Luisi, B. F., Xu, W. X., Otwinowski, Z., Freedman, L. P.,Yamamoto, K. R. & Sigler, P. B. (1991). Crystallo-graphic analysis of the interaction of the glucocorti-coid receptor with DNA. Nature, 352, 497±505.
Milligan, J. F., Groebe, D. R., Witherell, G. W. &Uhlenbeck, O. C. (1987). Oligoribonucleotide syn-thesis using T7 RNA polymerase and syntheticDNA templates. Nucl. Acids Res. 15, 8783±8789.
Mueller, L., Legault, P. & Pardi, A. (1995). ImprovedRNA structure determination by detection of NOEcontacts to exchange-broadened amino protons.J. Am. Chem. Soc. 117, 11043±11048.
Nikonowicz, E. P. & Pardi, A. (1993). An ef®cient pro-cedure for assignment of the proton, carbon andnitrogen resonances in 13C/15N labeled nucleicacids. J. Mol. Biol. 232, 1141±1156.
Nikonowicz, E. P., Sirr, A., Legault, P., Jucker, F. M.,Baer, L. M. & Pardi, A. (1992). Preparation of 13Cand 15N labelled RNAs for heteronuclear multi-dimensional NMR studies. Nucl. Acids Res. 20,4507±4513.
Otwinowski, Z., Schevitz, R. W., Zhang, R. G., Lawson,C. L., Joachimiak, A., Marmorstein, R. Q., Luisi,B. F. & Sigler, P. B. (1988). Crystal structure of trprepressor/operator complex at atomic resolution.Nature, 335, 321±329.
Pardi, A. (1995). Multidimensional heteronuclear NMRexperiments for structure determination of isotopi-cally labeled RNA. Methods Enzymol. 261, 350±380.
Pavletich, N. P. & Pabo, C. O. (1991). Zinc ®nger-DNArecognition: crystal structure of a Zif268-DNA com-plex at 2.1 AÊ . Science, 252, 809±817.
Peterson, R. D., Bartel, D. P., Szostak, J. W., Horvath,S. J. & Feigon, J. (1994). 1H-NMR studies of thehigh-af®nity rev binding-site of the rev responsiveelement of HIV-1 messenger-RNA: base pairing inthe core binding-element. Biochemistry, 33 (18),5357±5366.
Philpott, C. C., Haile, D., Rouault, T. A. & Klausner,R. D. (1993). Modi®cation of a free Fe-S clustercysteine residue in the active iron-responsiveelement-binding protein prevents RNA binding.J. Biol. Chem. 268, 17655±17658.
Philpott, C. C., Klausner, R. D. & Rouault, T. A. (1994).The bifunctional iron-responsive element bindingprotein/cytosolic aconitase: the role of active-siteresidues in ligand binding and regulation. Proc.Natl Acad. Sci. USA, 91, 7321±7325.
Piotto, M., Saudek, V. & Sklenar, V. (1992). Gradient-tai-lored excitation for single-quantum NMR-spec-troscopy of aqueous-solutions. J. Biomol. NMR, 2,661±665.
Puglisi, J. D., Chen, L., Frankel, A. D. & Williamson, J. R.(1993). Role of RNA structure in arginine recog-nition of TAR RNA. Proc. Natl Acad. Sci. USA, 90,3680±3684.
Rance, M., Sùrensen, O. W., Bodenhausen, G., Wagner,G., Ernst, R. R. & WuÈ thrich, K. (1983). Improvedspectral resolution in COSY 1H NMR spectra ofproteins via double quantum ®ltering. Biochem. Bio-phys. Res. Commun. 117, 479±485.
Robbins, A. H. & Stout, C. D. (1989). Structure of acti-vated aconitase: formation of the [4Fe-4S] cluster in
82 Structure of the Iron Responsive Element

the crystal. Proc. Natl Acad. Sci. USA, 86, 3639±3643.
Rouault, T. A., Stout, C. D., Kaptain, S., Harford, J. B. &Klausner, R. D. (1991). Structural relationshipbetween an iron-regulated RNA-binding protein(IRE-BP) and aconitase: functional implications. Cell,64, 881±883.
Rouault, T. A., Haile, D. J., Downey, W. E., Philpott,C. C., Tang, C., Samaniego, F., Chin, J., Paul, I.,Orloff, D. & Harford, J. B., et al. (1992). An iron-sul-fur cluster plays a novel regulatory role in the iron-responsive element binding protein. Biometals, 5(3),131±140.
Saenger, W. (1984). Principles of Nucleic Acid Structure,Springer-Verlag, New York.
Samaniego, F., Chin, J., Iwai, K., Rouault, T. A. &Klausner, R. D. (1994). Molecular characterization ofa second iron-responsive element binding protein,iron regulatory protein 2. Structure, function, andpost-translational regulation. J. Biol. Chem. 269,30904±30910.
Schalinkse, K. L., Anderson, S. L., Tuazon, P. T., Chen,O. S., Kennedy, M. C. & Eisenstein, R. S. (1997).The iron-sulfur cluster of iron regulatory protein 1modulates the accessibility of RNA binding andphosphorylation sites. Biochemistry, 36, 3950±3958.
Schwalbe, H., Marino, J. P., King, G. C., Wechselberger,R., Bermel, W. & Griesinger, C. (1994). Determi-nation of a complete set of coupling-constants in13C-labeled oligonucleotides. J. Biomol. NMR, 4,631±644.
Sierzputowska-Gracz, H., McKenzie, R. A. & Theil, E. C.(1995). The importance of a single G in the hairpinloop of the iron responsive element (IRE) in ferritinmRNA for structure: an NMR spectroscopy study.Nucl. Acids Res. 23, 146±153.
Simorre, J.-P., Zimmermann, G. R., Pardi, A., Farmer,B. T. I. I. & Mueller, L. (1995). Triple resonanceHNNCCH experiments for correlating exchangeableand nonexchangeable cytidine and uridine baseprotons in RNA. J. Biomol. NMR, 6, 427±432.
Simorre, J.-P., Zimmermann, G. R., Mueller, L. & Pardi,A. (1996a). Triple-resonance experiments for assign-ment of adenine base resonances in 13C/15N-labeledRNA. J. Am. Chem. Soc. 118, 5316±5317.
Simorre, J.-P., Zimmermann, G. R., Mueller, L. & Pardi,A. (1996b). Correlation of the guanosine exchange-able and nonexchangeable base protons in13C-/15N-labeled RNA with an HNC-TOCSY-CHexperiment. J. Biomol. NMR, 7, 153±156.
SklenaÂr, V. & Bax, A. (1987). Spin-echo water suppres-sion for the generation of pure-phase two-dimen-sional NMR spectra. J. Magn. Reson. 74, 469±479.
Swenson, G. R. & Walden, W. E. (1994). Localization ofan RNA-binding element of the iron-responsiveelement-binding protein within a proteolytic frag-
ment containing iron coordination ligands. Nucl.Acids Res. 22, 2627±2633.
Theil, E. C. (1994). Iron regulatory elements (IREs): afamily of mRNA non-coding sequences. Biochem. J.304, 1±11.
Varani & Tinoco (1991). RNA structure and NMRspectroscopy. Quart. Rev. Biophys. 24, 479±532.
Valegard, K., Murray, J. B., Stockley, P. G., Stonehouse,N. J. & Liljas, L. (1994). Crystal structure of a bac-teriophage-RNA coat protein-operator complex.Nature, 371, 623±626.
Wang, Y. H., Sczekan, S. R. & Theil, E. C. (1990). Struc-ture of the 50 untranslated regulatory region of ferri-tin mRNA studied in solution. Nucl. Acids Res. 18,4463±8.
Wang, Y. H., Lin, P. N., Sczekan, S. R., McKenzie,R. A. & Theil, E. C. (1991). Ferritin mRNA probed,near the iron regulatory region, with protein andchemical (1,10-phenanthroline-Cu) nucleases. Apossible role for base-paired ¯anking regions. Biol.Met. 4, 56±61.
WuÈ thrich, K. (1986). NMR of Proteins and Nucleic Acids,John Wiley & Sons, New York.
Yamazaki, T., Muhandiram, R. & Kay, L. E. (1994).NMR experiments for the measurement of carbonrelaxation properties in highly enriched, uniformly13C,15N-labeled proteins: application to 13Ca
carbons. J. Am. Chem. Soc. 116, 8266±8278.Zheng, L., Kennedy, M. C., Beinrt, H. & Zalkin, H.
(1992). Mutational analysis of active-site residues inpig-heart aconitase. J. Biol. Chem. 267, 7895±7903.
Zimmermann, G. R., Jenison, R. D., Wick, C. L.,Simorre, J.-P. & Pardi, A. (1997). Interlocking struc-tural motifs mediate molecular discrimination by atheophylline-binding RNA. Nature-Struct. Biol. 4,644±649.
Edited by I. Tinoco
(Received 12 June 1997; received in revised form18 August 1997; accepted 18 August 1997)
http://www.hbuk.co.uk/jmb
Supplementary material for this paper comprisingtwo Tables and one Figure is available from JMBOnline.
Structure of the Iron Responsive Element 83

![$PQZSJHIU …ousar.lib.okayama-u.ac.jp/files/public/5/57946/...(IRP2), an iron-responsive protein that is a master reg-ulator of intracellular iron homeostasis [7,27]. LIP is strongly](https://static.fdocuments.in/doc/165x107/5e72c50a70ae81741e1c6d38/pqzsjhiu-ousarlibokayama-uacjpfilespublic557946-irp2-an-iron-responsive.jpg)
















