![-Chaconine [20562-03-2] and -Solanine [20562-02-1] · -Chaconine [20562-03-2] and -Solanine ... Solanine exhibited a hyperglycemic effect in intact rats and a hypoglycemic effect](https://static.fdocuments.in/doc/165x107/5ad6f65c7f8b9a32618bb6e7/-chaconine-20562-03-2-and-solanine-20562-02-1-20562-03-2-and-solanine-.jpg)
Steroidal glycoalkaloids in Solanum chacoense - hort.vt.edu · been shown to accumulate...
9
Steroidal glycoalkaloids in Solanum chacoense Alice M. Mweetwa a,1 , Danielle Hunter a , Rebecca Poe a , Kim C. Harich b , Idit Ginzberg c , Richard E. Veilleux a , James G. Tokuhisa a,⇑ a Department of Horticulture, Virginia Tech, Blacksburg, VA 24061, USA b Department of Biochemistry, Virginia Tech, Blacksburg, VA 24061, USA c Institute of Plant Sciences, ARO, The Volcani Center, Bet Dagan 50250, Israel article info Article history: Received 19 September 2011 Received in revised form 28 November 2011 Available online 2 January 2012 Keywords: Solanum chacoense Solanaceae Chaco potato Developmental profile Steroidal glycoalkaloids Leptines hmg pss sgt abstract Potato (Solanum tuberosum L.), a domesticated species that is the fourth most important world agricul- tural commodity, requires significant management to minimize the effects of herbivore and pathogen damage on crop yield. A wild relative, Solanum chacoense Bitt., has been of interest to plant breeders because it produces an abundance of novel steroidal glycoalkaloid compounds, leptines and leptinines, which are particularly effective deterrents of herbivory by the Colorado potato beetle (Leptinotarsa decemlineata Say). Biochemical approaches were used in this study to investigate the formation and accu- mulation of SGAs in S. chacoense. SGA contents were determined in various organs at different stages of organ maturity during a time course of plant development. Leptines and leptinines were the main con- tributors to the increased levels in SGA concentration measured in the aerial versus the subterranean organs of S. chacoense accession 8380-1. Leptines were not detected in aboveground stolons until the stage where shoots had formed mature chlorophyllous leaves. To gain insights into SGA biosynthesis, the abundance of SGAs and steady-state transcripts of genes coding for enzymes of the central terpene and SGA-specific pathways in various plant organs at anthesis were compared. For two genes of primary terpene metabolism, transcript and SGA abundances were correlated, although with some discrepancies. For genes associated with SGA biosynthesis, transcripts were not detected in some tissues containing SGAs; however these transcripts were detected in the progenitor tissues, indicating the possibility that under our standard growth conditions, SGA biosynthesis is largely limited to highly proliferative tissues such as shoot, root and floral meristems. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Steroidal glycoalkaloids (SGAs) are a class of plant natural prod- ucts found in many species of the genus Solanum including potato and tomato (Milner et al., 2011). They are also found in the genus Veratrum of the monocotyledon family Melanthiaceae (Kaneko et al., 1977). SGAs have been shown to function as defense com- pounds in non-host resistance by Solanaceous plants against herbi- vores (Tingey, 1984) and eukaryotic microbial pathogens (Deahl et al., 1973; Fewell and Roddick, 1993). Their biological activity has been attributed to two modes of action: inhibition of (acetyl)cholinesterase activity (Roddick, 1989) and disruption of the eukaryotic cell membrane structure through binding with the sterol component of the membranes (Bouarab et al., 2002). The biological properties of SGAs can adversely affect humans and are the reason for limits on their abundance in the tubers of cul- tivated potato. Commercial potato breeders reject cultivars that have SGA concentrations in the tubers exceeding 0.2 lmol g À1 fr. wt. However, SGAs are present in the ‘‘skins’’ of potato tubers (Krits et al., 2007) and unripe tomato fruit (Moco et al., 2007) and give a characteristic bitterness that contributes to the organoleptic proper- ties of the cooked potato tuber and unripe tomato fruit. Typically the SGA levels found in supermarket potatoes are 0.03–0.06 lmol g À1 fr. wt. In other tissues of cultivated and wild potatoes, SGAs accumulate to various and typically greater levels (Nema et al., 2008). For in- stance, SGAs are present in the leaves of cultivated potato at concen- trations up to 1.7 lmol g À1 fr. wt. and in wild potato leaves to 8.5 lmol g À1 fr. wt. (Gregory et al., 1981; Friedman and Dao, 1992). Certain accessions of the diploid wild potato species Solanum chacoense have some of the highest contents of SGAs among wild potato species (Sinden et al., 1986b). Whereas a-chaconine (1) and a-solanine (2) are the SGAs found in cultivated potato (Fig. 1), S. chacoense produces leptinines (3,4) and leptines (5,6), un- ique classes of SGAs that are the hydroxylated and O-acetylated 0031-9422/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.phytochem.2011.12.003 ⇑ Corresponding author. Address: 503 Latham Hall, Department of Horticulture, Virginia Tech, Blacksburg, VA 24060, USA. Tel.: +1 540 231 5653; fax: +1 540 231 3347. E-mail address: [email protected] (J.G. Tokuhisa). 1 Present address: School of Agricultural Science, University of Zambia, Lusaka, Zambia. Phytochemistry 75 (2012) 32–40 Contents lists available at SciVerse ScienceDirect Phytochemistry journal homepage: www.elsevier.com/locate/phytochem
Transcript of Steroidal glycoalkaloids in Solanum chacoense - hort.vt.edu · been shown to accumulate...

Phytochemistry 75 (2012) 32–40
Contents lists available at SciVerse ScienceDirect
Phytochemistry
journal homepage: www.elsevier .com/locate /phytochem
Steroidal glycoalkaloids in Solanum chacoense
Alice M. Mweetwa a,1, Danielle Hunter a, Rebecca Poe a, Kim C. Harich b, Idit Ginzberg c,Richard E. Veilleux a, James G. Tokuhisa a,⇑a Department of Horticulture, Virginia Tech, Blacksburg, VA 24061, USAb Department of Biochemistry, Virginia Tech, Blacksburg, VA 24061, USAc Institute of Plant Sciences, ARO, The Volcani Center, Bet Dagan 50250, Israel
a r t i c l e i n f o a b s t r a c t
Article history:Received 19 September 2011Received in revised form 28 November 2011Available online 2 January 2012
Keywords:Solanum chacoenseSolanaceaeChaco potatoDevelopmental profileSteroidal glycoalkaloidsLeptineshmgpsssgt
0031-9422/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.phytochem.2011.12.003
⇑ Corresponding author. Address: 503 Latham HallVirginia Tech, Blacksburg, VA 24060, USA. Tel.: +1 543347.
E-mail address: [email protected] (J.G. Tokuhisa).1 Present address: School of Agricultural Science, U
Zambia.
Potato (Solanum tuberosum L.), a domesticated species that is the fourth most important world agricul-tural commodity, requires significant management to minimize the effects of herbivore and pathogendamage on crop yield. A wild relative, Solanum chacoense Bitt., has been of interest to plant breedersbecause it produces an abundance of novel steroidal glycoalkaloid compounds, leptines and leptinines,which are particularly effective deterrents of herbivory by the Colorado potato beetle (Leptinotarsadecemlineata Say). Biochemical approaches were used in this study to investigate the formation and accu-mulation of SGAs in S. chacoense. SGA contents were determined in various organs at different stages oforgan maturity during a time course of plant development. Leptines and leptinines were the main con-tributors to the increased levels in SGA concentration measured in the aerial versus the subterraneanorgans of S. chacoense accession 8380-1. Leptines were not detected in aboveground stolons until thestage where shoots had formed mature chlorophyllous leaves. To gain insights into SGA biosynthesis,the abundance of SGAs and steady-state transcripts of genes coding for enzymes of the central terpeneand SGA-specific pathways in various plant organs at anthesis were compared. For two genes of primaryterpene metabolism, transcript and SGA abundances were correlated, although with some discrepancies.For genes associated with SGA biosynthesis, transcripts were not detected in some tissues containingSGAs; however these transcripts were detected in the progenitor tissues, indicating the possibility thatunder our standard growth conditions, SGA biosynthesis is largely limited to highly proliferative tissuessuch as shoot, root and floral meristems.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Steroidal glycoalkaloids (SGAs) are a class of plant natural prod-ucts found in many species of the genus Solanum including potatoand tomato (Milner et al., 2011). They are also found in the genusVeratrum of the monocotyledon family Melanthiaceae (Kanekoet al., 1977). SGAs have been shown to function as defense com-pounds in non-host resistance by Solanaceous plants against herbi-vores (Tingey, 1984) and eukaryotic microbial pathogens (Deahlet al., 1973; Fewell and Roddick, 1993). Their biological activityhas been attributed to two modes of action: inhibition of(acetyl)cholinesterase activity (Roddick, 1989) and disruption ofthe eukaryotic cell membrane structure through binding with thesterol component of the membranes (Bouarab et al., 2002).
ll rights reserved.
, Department of Horticulture,0 231 5653; fax: +1 540 231
niversity of Zambia, Lusaka,
The biological properties of SGAs can adversely affect humansand are the reason for limits on their abundance in the tubers of cul-tivated potato. Commercial potato breeders reject cultivars thathave SGA concentrations in the tubers exceeding 0.2 lmol g�1 fr.wt. However, SGAs are present in the ‘‘skins’’ of potato tubers (Kritset al., 2007) and unripe tomato fruit (Moco et al., 2007) and give acharacteristic bitterness that contributes to the organoleptic proper-ties of the cooked potato tuber and unripe tomato fruit. Typically theSGA levels found in supermarket potatoes are 0.03–0.06 lmol g�1 fr.wt. In other tissues of cultivated and wild potatoes, SGAs accumulateto various and typically greater levels (Nema et al., 2008). For in-stance, SGAs are present in the leaves of cultivated potato at concen-trations up to 1.7 lmol g�1 fr. wt. and in wild potato leaves to8.5 lmol g�1 fr. wt. (Gregory et al., 1981; Friedman and Dao, 1992).
Certain accessions of the diploid wild potato species Solanumchacoense have some of the highest contents of SGAs among wildpotato species (Sinden et al., 1986b). Whereas a-chaconine (1)and a-solanine (2) are the SGAs found in cultivated potato(Fig. 1), S. chacoense produces leptinines (3,4) and leptines (5,6), un-ique classes of SGAs that are the hydroxylated and O-acetylated

α-L-Rha-(1-4)
α-L-Rha-(1-2)
β-D-Glc-O-
β-D-Glc-(1-3)
α-L-Rha-(1-2)
β-D-Gal-O-
β-D-Glc-(1-3)
β-D-Glc-(1-2)
β-D-Glc-(1-4) -- β-D-Gal-O-
SolatriosylChacotriosyl Commertetraosyl
Fig. 1. Structures and molecular weight of the major SGAs of Solanum chacoense accession 8380-1. The basic solanidine aglycone is modified by hydroxylation and O-acetylation at C23 (R2) and named as indicated. The glycosides are formed by the addition of a chacotriosyl, solatriosyl or commertetraosyl group at C3 (R1).
A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40 33
derivatives, respectively, of a-chaconine (1) and a-solanine (2;Kuhn and Löw, 1961). Leptines (5,6) and leptinines (3,4) are absentfrom cultivated potato varieties and even among wild species thisclass of SGAs has a limited distribution (Distl and Wink, 2009).These derivative compounds have been shown to be more effectivethan the parental SGAs as deterrents against the Colorado potatobeetle (Sinden et al., 1988). Furthermore, leptines (5,6) andleptinines (3,4) accumulate only in the aerial parts of the plant.
S. chacoense was targeted as an experimental system to explorethe role of SGAs in the natural chemical defense system of wild po-tato. In this study it was assumed that the reduction of SGA contentto ensure the safety of tubers for human consumption (Johns andAlonso, 1990) has also reduced the herbivore and pathogendefenses of cultivated potato. An understanding of the formationand function of SGAs in an entire plant whose genotype has beendefined by natural selection rather than domestication will allowus to more fully exploit the defense potential of SGAs in cultivatedpotato. As an initial investigation, described herein are: (i) a timecourse survey of SGA concentration and composition in various or-gans of the developing plant, and (ii) transcript levels of genes cod-ing for selected isoprenoid biosynthetic enzymes including somegenes directly involved with SGA biosynthesis.
2. Results and discussion
2.1. Qualitative differences in the SGA profile of S. chacoense
Different accessions of S. chacoense vary in SGA profile andcontent (Ronning et al., 2000). In this study, the high leptine accu-mulating line S. chacoense accession 8380-1 was chosen to investi-gate the chemical defense biochemistry because it has been usedpreviously for analytical and ecological studies (Sinden et al.,
1986a). In this study, the accession under controlled growing con-ditions was cultivated to analyze SGA accumulation at the organlevel and to monitor the dynamics of these plant defense com-pounds at different developmental stages.
SGAs were extracted from plant organs at various developmen-tal stages (Table 1) and fractionated by reversed phase HPLC withmonitoring at A202nm (Fig. 2). The chromatographic profiles dif-fered most significantly between the aerial (flowers and leaves)and subterranean (roots and tubers) organ extracts. However, allsamples had two peaks with retention times (RTs) of 7.9 and8.4 min that co-migrated with authentic standards of a-solanine(2) and a-chaconine (1), respectively. To confirm the identity ofthe additional HPLC peaks of the tissue extracts, leaf and root sam-ples as well as subfractions of the HPLC eluate of leaf samples wereanalyzed by LC–MS. For the major peaks of the leaf extract, the val-ues of the molecular [M + H]+ and fragment ions were consistentwith those observed for a-chaconine (1), a-solanine (2), leptines(5,6) and their closely related compounds, leptinines (3,4) (Table2; Distl and Wink, 2009). The specific 8380-1 accession used hasbeen maintained through clonal propagation and has been thesource material for analyses of leptine I (5) and II (6) and leptinineI (3) and II (4) by thin layer chromatography and HPLC (Sindenet al., 1986b; Kowalski et al., 2000), and for the characterizationof their respective aglycones by NMR and GC–MS (Lawson et al.,1992, 1997). Taken together, the identities in leaf sample extractsof leptinine II (4; RT 4.4 min), leptinine I (3; RT 4.7 min), a-solanine(2; RT 7.9 min), a-chaconine (1; RT 8.4 min), leptine II (6;RT10.6 min) and leptine I (5; RT 11.0 min) were confirmed.
Root extracts analyzed by HPLC exhibited a distinct peak at RT7.7 min. LC–MS analyses indicated m/z values of 1046.4 for themolecular ion [M + H]+, and 398, 560, and 722 for the fragment ions.This pattern was identical to the LC–MS characterization of dehyd-rocommersonine (7; Distl and Wink, 2009). Accession 8380-1 has
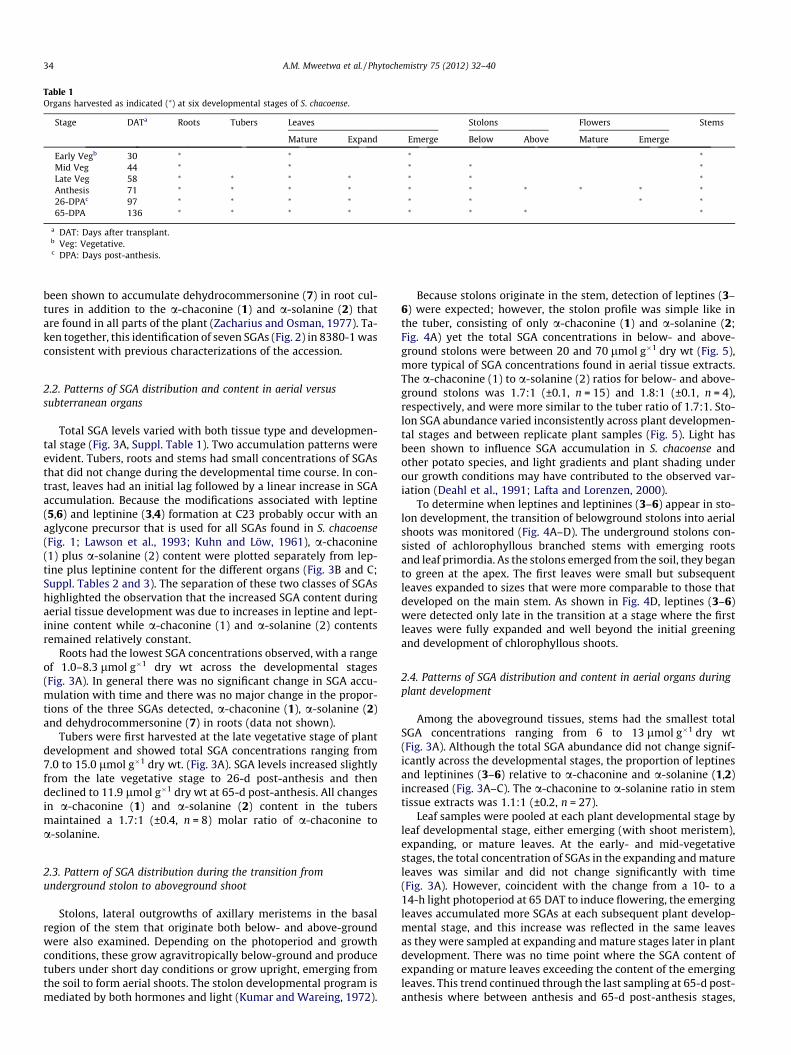
Table 1Organs harvested as indicated (⁄) at six developmental stages of S. chacoense.
Stage DATa Roots Tubers Leaves Stolons Flowers Stems
Mature Expand Emerge Below Above Mature Emerge
Early Vegb 30 ⁄ ⁄ ⁄ ⁄
Mid Veg 44 ⁄ ⁄ ⁄ ⁄ ⁄
Late Veg 58 ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄
Anthesis 71 ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄
26-DPAc 97 ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄
65-DPA 136 ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄ ⁄
a DAT: Days after transplant.b Veg: Vegetative.c DPA: Days post-anthesis.
34 A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40
been shown to accumulate dehydrocommersonine (7) in root cul-tures in addition to the a-chaconine (1) and a-solanine (2) thatare found in all parts of the plant (Zacharius and Osman, 1977). Ta-ken together, this identification of seven SGAs (Fig. 2) in 8380-1 wasconsistent with previous characterizations of the accession.
2.2. Patterns of SGA distribution and content in aerial versussubterranean organs
Total SGA levels varied with both tissue type and developmen-tal stage (Fig. 3A, Suppl. Table 1). Two accumulation patterns wereevident. Tubers, roots and stems had small concentrations of SGAsthat did not change during the developmental time course. In con-trast, leaves had an initial lag followed by a linear increase in SGAaccumulation. Because the modifications associated with leptine(5,6) and leptinine (3,4) formation at C23 probably occur with anaglycone precursor that is used for all SGAs found in S. chacoense(Fig. 1; Lawson et al., 1993; Kuhn and Löw, 1961), a-chaconine(1) plus a-solanine (2) content were plotted separately from lep-tine plus leptinine content for the different organs (Fig. 3B and C;Suppl. Tables 2 and 3). The separation of these two classes of SGAshighlighted the observation that the increased SGA content duringaerial tissue development was due to increases in leptine and lept-inine content while a-chaconine (1) and a-solanine (2) contentsremained relatively constant.
Roots had the lowest SGA concentrations observed, with a rangeof 1.0–8.3 lmol g�1 dry wt across the developmental stages(Fig. 3A). In general there was no significant change in SGA accu-mulation with time and there was no major change in the propor-tions of the three SGAs detected, a-chaconine (1), a-solanine (2)and dehydrocommersonine (7) in roots (data not shown).
Tubers were first harvested at the late vegetative stage of plantdevelopment and showed total SGA concentrations ranging from7.0 to 15.0 lmol g�1 dry wt. (Fig. 3A). SGA levels increased slightlyfrom the late vegetative stage to 26-d post-anthesis and thendeclined to 11.9 lmol g�1 dry wt at 65-d post-anthesis. All changesin a-chaconine (1) and a-solanine (2) content in the tubersmaintained a 1.7:1 (±0.4, n = 8) molar ratio of a-chaconine toa-solanine.
2.3. Pattern of SGA distribution during the transition fromunderground stolon to aboveground shoot
Stolons, lateral outgrowths of axillary meristems in the basalregion of the stem that originate both below- and above-groundwere also examined. Depending on the photoperiod and growthconditions, these grow agravitropically below-ground and producetubers under short day conditions or grow upright, emerging fromthe soil to form aerial shoots. The stolon developmental program ismediated by both hormones and light (Kumar and Wareing, 1972).
Because stolons originate in the stem, detection of leptines (3–6) were expected; however, the stolon profile was simple like inthe tuber, consisting of only a-chaconine (1) and a-solanine (2;Fig. 4A) yet the total SGA concentrations in below- and above-ground stolons were between 20 and 70 lmol g�1 dry wt (Fig. 5),more typical of SGA concentrations found in aerial tissue extracts.The a-chaconine (1) to a-solanine (2) ratios for below- and above-ground stolons was 1.7:1 (±0.1, n = 15) and 1.8:1 (±0.1, n = 4),respectively, and were more similar to the tuber ratio of 1.7:1. Sto-lon SGA abundance varied inconsistently across plant developmen-tal stages and between replicate plant samples (Fig. 5). Light hasbeen shown to influence SGA accumulation in S. chacoense andother potato species, and light gradients and plant shading underour growth conditions may have contributed to the observed var-iation (Deahl et al., 1991; Lafta and Lorenzen, 2000).
To determine when leptines and leptinines (3–6) appear in sto-lon development, the transition of belowground stolons into aerialshoots was monitored (Fig. 4A–D). The underground stolons con-sisted of achlorophyllous branched stems with emerging rootsand leaf primordia. As the stolons emerged from the soil, they beganto green at the apex. The first leaves were small but subsequentleaves expanded to sizes that were more comparable to those thatdeveloped on the main stem. As shown in Fig. 4D, leptines (3–6)were detected only late in the transition at a stage where the firstleaves were fully expanded and well beyond the initial greeningand development of chlorophyllous shoots.
2.4. Patterns of SGA distribution and content in aerial organs duringplant development
Among the aboveground tissues, stems had the smallest totalSGA concentrations ranging from 6 to 13 lmol g�1 dry wt(Fig. 3A). Although the total SGA abundance did not change signif-icantly across the developmental stages, the proportion of leptinesand leptinines (3–6) relative to a-chaconine and a-solanine (1,2)increased (Fig. 3A–C). The a-chaconine to a-solanine ratio in stemtissue extracts was 1.1:1 (±0.2, n = 27).
Leaf samples were pooled at each plant developmental stage byleaf developmental stage, either emerging (with shoot meristem),expanding, or mature leaves. At the early- and mid-vegetativestages, the total concentration of SGAs in the expanding and matureleaves was similar and did not change significantly with time(Fig. 3A). However, coincident with the change from a 10- to a14-h light photoperiod at 65 DAT to induce flowering, the emergingleaves accumulated more SGAs at each subsequent plant develop-mental stage, and this increase was reflected in the same leavesas they were sampled at expanding and mature stages later in plantdevelopment. There was no time point where the SGA content ofexpanding or mature leaves exceeding the content of the emergingleaves. This trend continued through the last sampling at 65-d post-anthesis where between anthesis and 65-d post-anthesis stages,
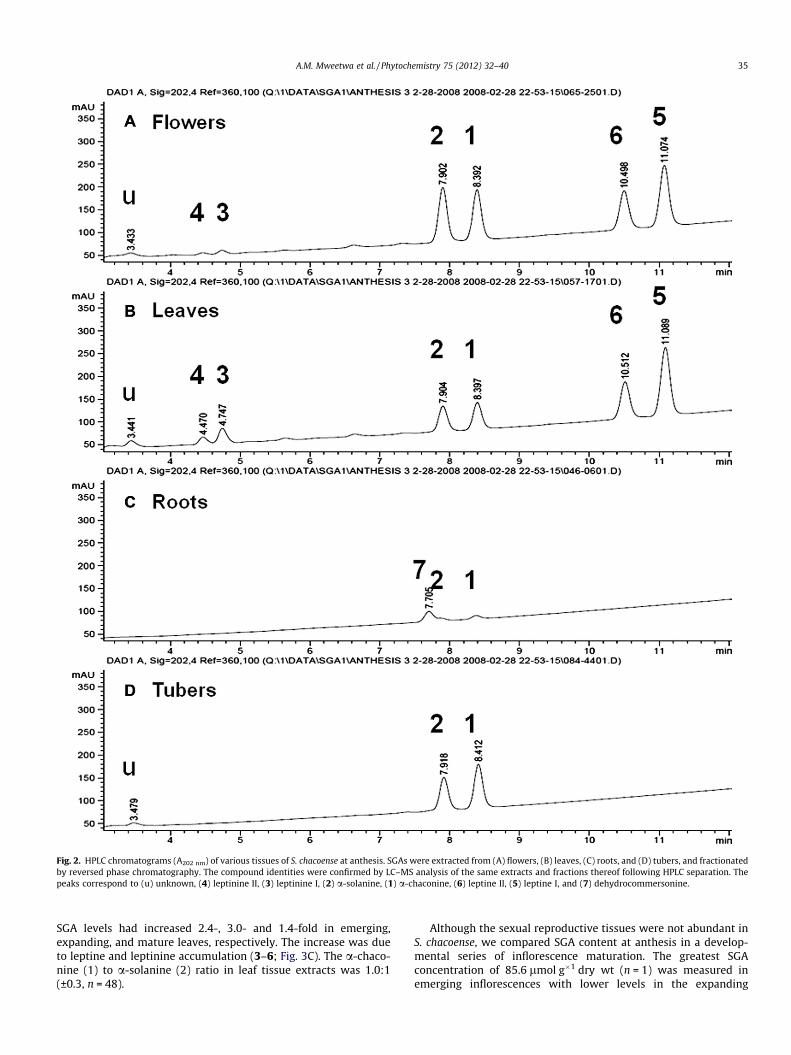
Fig. 2. HPLC chromatograms (A202 nm) of various tissues of S. chacoense at anthesis. SGAs were extracted from (A) flowers, (B) leaves, (C) roots, and (D) tubers, and fractionatedby reversed phase chromatography. The compound identities were confirmed by LC–MS analysis of the same extracts and fractions thereof following HPLC separation. Thepeaks correspond to (u) unknown, (4) leptinine II, (3) leptinine I, (2) a-solanine, (1) a-chaconine, (6) leptine II, (5) leptine I, and (7) dehydrocommersonine.
A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40 35
SGA levels had increased 2.4-, 3.0- and 1.4-fold in emerging,expanding, and mature leaves, respectively. The increase was dueto leptine and leptinine accumulation (3–6; Fig. 3C). The a-chaco-nine (1) to a-solanine (2) ratio in leaf tissue extracts was 1.0:1(±0.3, n = 48).
Although the sexual reproductive tissues were not abundant inS. chacoense, we compared SGA content at anthesis in a develop-mental series of inflorescence maturation. The greatest SGAconcentration of 85.6 lmol g�1 dry wt (n = 1) was measured inemerging inflorescences with lower levels in the expanding
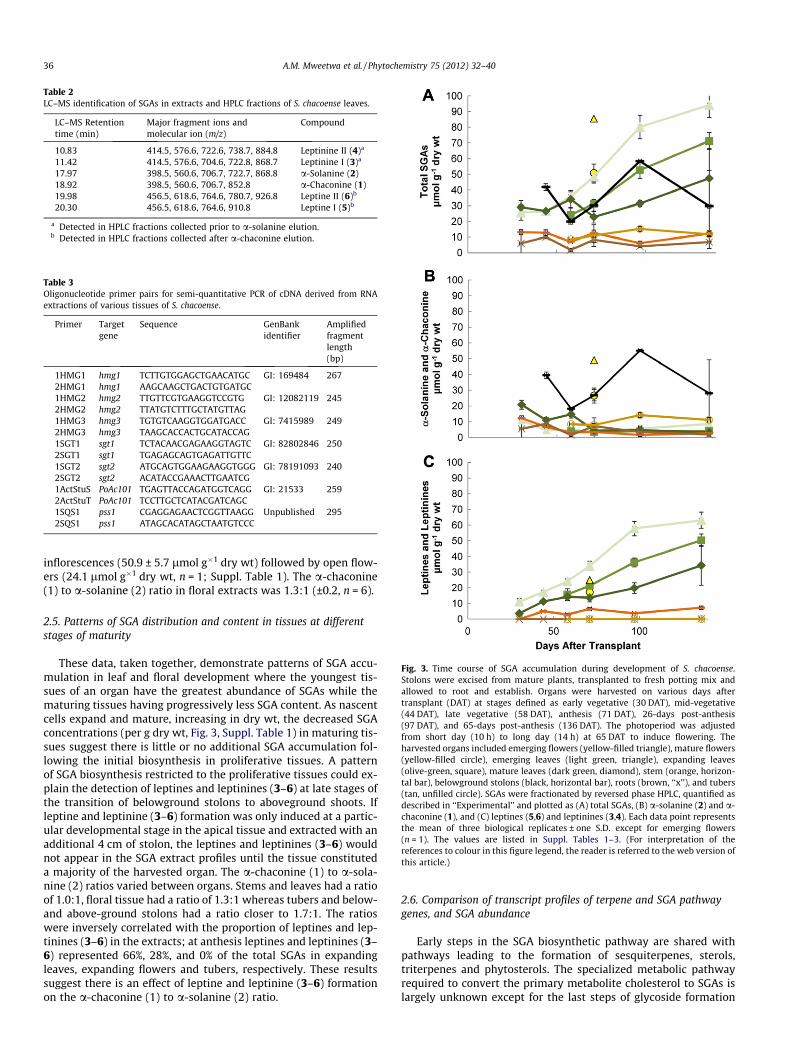
Table 2LC–MS identification of SGAs in extracts and HPLC fractions of S. chacoense leaves.
LC–MS Retentiontime (min)
Major fragment ions andmolecular ion (m/z)
Compound
10.83 414.5, 576.6, 722.6, 738.7, 884.8 Leptinine II (4)a
11.42 414.5, 576.6, 704.6, 722.8, 868.7 Leptinine I (3)a
17.97 398.5, 560.6, 706.7, 722.7, 868.8 a-Solanine (2)18.92 398.5, 560.6, 706.7, 852.8 a-Chaconine (1)19.98 456.5, 618.6, 764.6, 780.7, 926.8 Leptine II (6)b
20.30 456.5, 618.6, 764.6, 910.8 Leptine I (5)b
a Detected in HPLC fractions collected prior to a-solanine elution.b Detected in HPLC fractions collected after a-chaconine elution.
Table 3Oligonucleotide primer pairs for semi-quantitative PCR of cDNA derived from RNAextractions of various tissues of S. chacoense.
Primer Targetgene
Sequence GenBankidentifier
Amplifiedfragmentlength(bp)
1HMG1 hmg1 TCTTGTGGAGCTGAACATGC GI: 169484 2672HMG1 hmg1 AAGCAAGCTGACTGTGATGC1HMG2 hmg2 TTGTTCGTGAAGGTCCGTG GI: 12082119 2452HMG2 hmg2 TTATGTCTTTGCTATGTTAG1HMG3 hmg3 TGTGTCAAGGTGGATGACC GI: 7415989 2492HMG3 hmg3 TAAGCACCACTGCATACCAG1SGT1 sgt1 TCTACAACGAGAAGGTAGTC GI: 82802846 2502SGT1 sgt1 TGAGAGCAGTGAGATTGTTC1SGT2 sgt2 ATGCAGTGGAAGAAGGTGGG GI: 78191093 2402SGT2 sgt2 ACATACCGAAACTTGAATCG1ActStuS PoAc101 TGAGTTACCAGATGGTCAGG GI: 21533 2592ActStuT PoAc101 TCCTTGCTCATACGATCAGC1SQS1 pss1 CGAGGAGAACTCGGTTAAGG Unpublished 2952SQS1 pss1 ATAGCACATAGCTAATGTCCC
Fig. 3. Time course of SGA accumulation during development of S. chacoense.Stolons were excised from mature plants, transplanted to fresh potting mix andallowed to root and establish. Organs were harvested on various days aftertransplant (DAT) at stages defined as early vegetative (30 DAT), mid-vegetative(44 DAT), late vegetative (58 DAT), anthesis (71 DAT), 26-days post-anthesis(97 DAT), and 65-days post-anthesis (136 DAT). The photoperiod was adjustedfrom short day (10 h) to long day (14 h) at 65 DAT to induce flowering. Theharvested organs included emerging flowers (yellow-filled triangle), mature flowers(yellow-filled circle), emerging leaves (light green, triangle), expanding leaves(olive-green, square), mature leaves (dark green, diamond), stem (orange, horizon-tal bar), belowground stolons (black, horizontal bar), roots (brown, ‘‘x’’), and tubers(tan, unfilled circle). SGAs were fractionated by reversed phase HPLC, quantified asdescribed in ‘‘Experimental’’ and plotted as (A) total SGAs, (B) a-solanine (2) and a-chaconine (1), and (C) leptines (5,6) and leptinines (3,4). Each data point representsthe mean of three biological replicates ± one S.D. except for emerging flowers(n = 1). The values are listed in Suppl. Tables 1–3. (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version ofthis article.)
36 A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40
inflorescences (50.9 ± 5.7 lmol g�1 dry wt) followed by open flow-ers (24.1 lmol g�1 dry wt, n = 1; Suppl. Table 1). The a-chaconine(1) to a-solanine (2) ratio in floral extracts was 1.3:1 (±0.2, n = 6).
2.5. Patterns of SGA distribution and content in tissues at differentstages of maturity
These data, taken together, demonstrate patterns of SGA accu-mulation in leaf and floral development where the youngest tis-sues of an organ have the greatest abundance of SGAs while thematuring tissues having progressively less SGA content. As nascentcells expand and mature, increasing in dry wt, the decreased SGAconcentrations (per g dry wt, Fig. 3, Suppl. Table 1) in maturing tis-sues suggest there is little or no additional SGA accumulation fol-lowing the initial biosynthesis in proliferative tissues. A patternof SGA biosynthesis restricted to the proliferative tissues could ex-plain the detection of leptines and leptinines (3–6) at late stages ofthe transition of belowground stolons to aboveground shoots. Ifleptine and leptinine (3–6) formation was only induced at a partic-ular developmental stage in the apical tissue and extracted with anadditional 4 cm of stolon, the leptines and leptinines (3–6) wouldnot appear in the SGA extract profiles until the tissue constituteda majority of the harvested organ. The a-chaconine (1) to a-sola-nine (2) ratios varied between organs. Stems and leaves had a ratioof 1.0:1, floral tissue had a ratio of 1.3:1 whereas tubers and below-and above-ground stolons had a ratio closer to 1.7:1. The ratioswere inversely correlated with the proportion of leptines and lep-tinines (3–6) in the extracts; at anthesis leptines and leptinines (3–6) represented 66%, 28%, and 0% of the total SGAs in expandingleaves, expanding flowers and tubers, respectively. These resultssuggest there is an effect of leptine and leptinine (3–6) formationon the a-chaconine (1) to a-solanine (2) ratio.
2.6. Comparison of transcript profiles of terpene and SGA pathwaygenes, and SGA abundance
Early steps in the SGA biosynthetic pathway are shared withpathways leading to the formation of sesquiterpenes, sterols,triterpenes and phytosterols. The specialized metabolic pathwayrequired to convert the primary metabolite cholesterol to SGAs islargely unknown except for the last steps of glycoside formation

Fig. 4. SGA profiles during the transition from belowground stolon to aerial shoot. Organs were harvested as belowground branching stolons (A), stolons as emerging shoots(B), shoots with expanding leaves (C), and shoots with mature leaves (D). The HPLC eluate was monitored at A202 nm and the numbered peaks correspond to (4) leptinine II, (3)leptinine I, (2) a-solanine, (1) a-chaconine, (6) leptine II, and (5) leptine I.
A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40 37
(McCue et al., 2007). We compared SGA abundance with the tran-script levels of genes either shared by terpene/sterol pathways orthose associated only with SGA biosynthesis. We used semiquanti-tative RT-PCR to estimate steady-state transcript levels of thegenes coding for 3-hydroxymethylglutaryl-CoA reductase (hmg1,hmg2, hmg3), squalene synthase (pss), solanidine glycosyltransfer-ase (sgt1, sgt2) and actin (PoAc101) in the various organs (Fig. 6).
Transcripts of hmg2 and hmg3 were not detected in any organ,as expected, as the plants were grown under minimal stress condi-tions (Choi et al., 1992; Yang et al., 1991). The transcript abun-dance profiles for hmg1 and pss correlated approximately withthe SGA levels across the various organs (Fig. 6). Similar observa-tions and conclusions were reported in a survey of wild and culti-vated potato genotypes limited to leaf and tuber organs (Krits et al.,2007). For the genes directly associated with SGA biosynthesis, sgttranscript was not detected in several organs where SGAs weredetected including tubers (sgt1 and sgt2), roots (sgt2), mature and
expanding leaves (sgt1), and mature flowers (sgt1 and sgt2). Asimilar result was observed by Belknap and coworkers (Moehset al., 1997) in the initial characterization of sgt1. Thus, the tran-script levels for sgt1 and sgt2, genes most closely associated withSGA biosynthesis, did not correlate with SGA abundance whereasgenes associated with central terpene metabolism showed acorrelation.
Transcripts of sgt genes were detected in the emerging leaf pri-mordia and floral buds, tissues that develop into the leaf and floralorgans which lack sgt transcripts but contain SGAs. These resultssuggest that uncharacterized sgt gene family members encodesolanidine glycosyltransferase activity in those organs lackingdetectable sgt1 and sgt2 transcript or that SGT1 and SGT2 enzymeactivity persists in mature tissues although the transcripts of theencoding genes were not detected. Alternatively, SGA biosynthesismay occur primarily in proliferating tissues such as the root apicalmeristems, emerging floral buds and leaf primordia and the SGAs

Fig. 5. Total SGA concentration of stolons at different developmental stages. SGAextracts were prepared from belowground (black bar) and aboveground (white bar)stolons. Each data point represents the mean of three biological replicates ± one S.D.except aboveground stolons at anthesis (n = 1). The absence of a bar indicates nosample analyzed. Abbreviations: dPA, days post-anthesis.
38 A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40
detected in other tissues represent the initial biosynthesis or short-range transport. Long-range transport of SGAs is unlikely asreciprocal rootstock and scion grafts of potato and tomato havedemonstrated that species-specific SGAs are not transported acrossthe graft junction into the tissues of the other species (Roddick,1982).
Fig. 6. Comparison of SGA accumulation and steady-state transcript levels for genes ofabundance (lmol g�1 dry wt) in extracts of tubers (Tbr), belowground stolons (BG Stl), rostems (Stm), open flowers (Mat Flr), and emerging flowers (Emr Flr). Values for tottranscriptase-PCR of cDNA prepared from RNA extracts of the same tissues. Oligonuclemethylglutaryl-CoA-reductase (hmg1, hmg2, hmg3), squalene synthase (pss), solanidiabbreviation, oligonucleotide primer sequences, GenBank identifiers and expected DNA
3. Concluding remarks
Leptines and leptinines (3–6) are the dominant SGAs in the leaf,stem and floral organs of S. chacoense. It was found that their accu-mulation accounts for the large increase in SGA content after floralinduction. Although leptines and leptinines (3–6) were present inmost aerial tissues, their absence in developing stolons indicatedthat factors other than organ specificity and light control theiraccumulation. Correlations were also found for the various plantorgans between SGA concentrations and the transcript levels fortwo major genes in central terpene metabolism, but not betweenSGA concentrations and the transcript levels for genes closely asso-ciated with SGA biosynthesis. These results could indicate that SGAbiosynthesis occurs in most tissues but they also do not precludethe possibility of SGA biosynthesis localized to proliferative tissueswith little or no additional biosynthesis in the expanding and ma-ture tissues of the organ under normal growth conditions.
4. Experimental
4.1. Plant material and reagents
Solanum chacoense accession 8380-1 (derived from PI 458310,Potato Germplasm Introduction Station, Sturgeon Bay, WI; (Sindenet al., 1986b) was the original source of plant material which hasbeen maintained in tissue culture for several years. a-Solanine(1; Cat. # 045K7350) and a-chaconine standards (2; Cat. #
terpene/sterol and SGA biosyntheses in various organs at anthesis. (A) Total SGAots (Rt), mature leaves (Mat Lf), expanding leaves (Exp Lf), emerging leaves (Emr Lf),al SGA concentrations are listed in Suppl. Table 1. (B) Semiquantitative reverseotide primer pairs were specific for gene family members coding for 3-hydroxy-3-ne glycosyltransferase (sgt1, sgt2) and actin (PoAc101). The primer name, geneproduct sizes for the genes are listed in Table 3.

A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40 39
089H7032; 99% purity, from potato sprouts) were obtained fromSigma–Aldrich (St. Louis, MO). Oligonucleotide primers were fromIntegrated DNA Technologies (Coralville, IA, Table 2).
4.2. Plant growth conditions
Plants were grown in an environmental growth chamber(Conviron, Winnipeg, Manitoba) under a 10 h photoperiod at24 �C and night temperature of 19 �C. A combination of fluorescentand incandescent light sources provided photosynthetically activeradiation of approximately 275 lmol photons m�2 s�1. The relativehumidity was maintained at 60%. Plants were fertilized every 14 dwith All Purpose Miracle-Gro (NPK of 24-8-16, Scotts, Marysville,OH) at a concentration of 1 g l�1. Stolon cuttings were planted incell packs containing Sunshine Mix # 1 (Sun Gro Horticulture,Bellevue, WA) and then transferred to 10-cm pots after 28 d. Flow-ering was induced by lengthening the photoperiod to 14 h at65 days after transplanting.
4.3. Plant developmental stages and organs for SGA analysis
Organs were harvested at different developmental stages repre-senting the entire life cycle of the plant as summarized in Table 1.Plants were harvested at the same time of day and separated intovarious organs. Organ fresh weights were determined after whichthe samples were lyophilized (Labconco, Kansas City, MO) for72 h (Sagredo et al., 2006). The number of plants representing eachbiological replicate varied with developmental stage but therewere at least three biological replicates for each sample.
4.4. SGA extraction
Freeze-dried samples were ground by mortar and pestle withliq. N2. Extraction and profile analyses using high performance li-quid chromatography (HPLC) followed the procedure of Edwardsand Cobb (1996). In brief, SGAs were extracted with 0.02 M hep-tanesulfonic acid in 1% aq. CH3COOH (v/v), then concentratedand enriched by solid phase extraction using C18 Classic cartridgecolumns (Waters, Milford, MA).
4.5. SGA identification and quantification I
SGAs were fractionated by HPLC (Agilent HP 1200 Series, SantaClara, CA) on a C18 reversed-phase column (Agilent Eclipse XDB-C18, 5 lm pore size and 4.6 � 150 mm). SGAs were eluted usinga gradient of Solvent A, CH3CN/0.01 M Tris–HCl (30:70, v/v pH8.0), and Solvent B, CH3CN/0.01 M Tris–HCl (80:20, v/v, pH 8.0)at a flow rate of 1 ml min�1 at 25 �C. Gradient elution was0–10 min, 0–40% B; 10–13 min, 40–100% B; 13–15 min, 100% B;15–15.5 min, 100–0% B; 15.5–20 min, 0% B. The eluent was moni-tored by diode array detection from 190 to 240 nm. Quantificationof SGAs was based on peak absorbance area at A202 nm, which wasconverted to molar concentrations using relative response factorsgenerated from calibration curves for a-solanine and a-chaconinemade with purified standards. The plots were linear within therange of 125–2000 ng and generated response factors of3.0 ng mAU�1 and 3.4 ng mAU�1, respectively. For quantificationof other SGAs, which all contained the aglycone solanidine withside modifications at C3 and C23 (Fig. 2), the response factor wasused for a-chaconine (1). For LC–MS, extracted SGAs (2 ll) were in-jected directly onto the analytical column described above with aflow rate of 0.5 ml min�1 at 50 �C and a gradient consisting of0.1% aq. HCO2H (Solvent A) and 0.1% aq. HCO2H in CH3CN (SolventB). The run consisted of 0.0–2.00 min, 30% solvent B; 2.00–10.00 min, 30–60% solvent B; 10–15 min, 60% solvent B; 15–15.01 min, 60–30% solvent B; 15.01–17 min, 30% solvent B. Eluant
was analyzed directly using the 3200 Q TRAP Linear Ion Trap Quad-rupole LC/MS/MS System (AB Sciex, Carlsbad, CA), equipped with aTurboSpray ion source.
4.6. SGA identification II: HPLC and LC/MS
Additional HPLC analyses were conducted with a smaller formatC18 reversed-phase column (Agilent Eclipse XDB-C18, 2.1 �100 mm, 3.5 lm pore size) and MS-compatible Solvent A, 10 mM(NH4)CO3 pH 8.0, and Solvent B, CH3CN/80 mM NH4CO3 (7:1) witha gradient elution of 0–0.2 min, 0% B; 0.2–1.0 min, 0–36% B; 1.0–16.0 min, 36–48% B; 16.0–22.0 min, 48–100% B; 22.0–25.0 min,100% B; 25.0–25.5 min, 100–0% B; and 25.5–32.0 min, 0% B. Thesesame column and conditions were used for LC–MS with the eluentanalyzed by mass spectrometry as described above (Section 4.5).
4.7. RNA extraction and cDNA synthesis
Tubers, stolons, roots, leaves, stems, flowers and floral buds of S.chacoense were harvested at anthesis, chilled with liquid nitrogenand stored at �80 �C. Total RNA was extracted from the tissuesusing TRIzol reagent (Invitrogen, Carlsbad, CA) according to instruc-tions. RNA was quantified by spectrophotometry and assessed byseparation on formaldehyde-denaturing agarose gels. First-strandcDNA was prepared from 2 lg of total RNA using 200 U of M-MLVreverse transcriptase SuperScript II (Invitrogen), and oligo (dT)20
as the oligonucleotide primer in a reaction volume of 50 ll.
4.8. Semi-quantitative reverse transcription polymerase chain reaction(RT-PCR)
Relative steady-state transcript levels of selected genes weredetermined from first-strand cDNA product from 40 ng of RNA.The oligonucleotide primers and corresponding genes are listedin Table 2. The cycle conditions were an initial denaturation at95 �C for 3 min, and 27 cycles at 94 �C for 30 s, 58 �C for 30 s,72 �C for 1 min, followed by 5 min incubation at 72 �C. Shorterand longer cycles (25, 30, 35 and 40) were conducted to ensurethat the PCR was in a linear response range at 27 cycles. The PCRproducts were stained with ethidium bromide following 1% aga-rose gel electrophoresis. Restriction enzyme digests confirmedDNA product identity. The gene PoAc101 (Drouin and Dover,1990) coding for actin was used for normalization.
4.9. Statistical analysis
Analysis of variance for SGA content was conducted with SASsoftware (Version 9.1, SAS Institute Inc., Cary, NC) with mean sep-aration using the Ryan-Einot-Gabriel-Welsch multiple range test(REGWQ) at P 6 0.05.
Acknowledgements
Generous support was provided by the Thomas F. and Kate Mill-er Jeffress Memorial Trust (J-871) and the Binational AgriculturalResearch and Development Fund (IS-4134-08).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.phytochem.2011.12.003.

40 A.M. Mweetwa et al. / Phytochemistry 75 (2012) 32–40
References
Bouarab, K., Melton, R., Peart, J., Baulcombe, D., Osbourn, A., 2002. A saponin-detoxifying enzyme mediates suppression of plant defences. Nature 418, 889–892.
Choi, D., Ward, B.L., Bostock, R.M., 1992. Differential induction and suppression ofpotato 3-hydroxy-3-methylglutaryl Coenzyme A reductase genes in response toPhytophthora infestans and to its elicitor arachidonic acid. Plant Cell 4, 1333–1344.
Deahl, K.L., Cantelo, W.W., Sinden, S.L., Sanford, L.L., 1991. The effect of light-intensity on Colorado potato beetle resistance and foliar glycoalkaloidconcentration of four Solanum chacoense clones. Am. Potato J. 68, 659–666.
Deahl, K.L., Young, R.J., Sinden, S.L., 1973. A study of the relationship of late blightresistance to glycoalkaloid content in fifteen potato clones. Am. Potato J. 50,248–253.
Distl, M., Wink, M., 2009. Identification and quantification of steroidal alkaloidsfrom wild tuber-bearing Solanum species by HPLC and LC-ESI-MS. Potato Res.52, 79–104.
Drouin, G., Dover, G.A., 1990. Independent gene evolution in the potato actin genefamily demonstrated by phylogenetic procedures for resolving geneconversions and the phylogeny of angiosperm actin genes. J. Mol. Evol. 31,132–150.
Edwards, E.J., Cobb, A.H., 1996. Improved high-performance liquid chromatographicmethod for the analysis of potato (Solanum tuberosum) glycoalkaloids. J. Agric.Food Chem. 44, 2705–2709.
Fewell, A.M., Roddick, J.G., 1993. Interactive antifungal activity of the glycoalkaloidsa-solanine and a-chaconine. Phytochemistry 33, 323–328.
Friedman, M., Dao, L., 1992. Distribution of glycoalkaloids in potato plants andcommercial potato products. J. Agric. Food Chem. 40, 419–423.
Gregory, P., Sinden, S.L., Osman, S.F., Tingey, W.M., Chessin, D.A., 1981.Glycoalkaloids of wild, tuber-bearing Solanum species. J. Agric. Food Chem.29, 1212–1215.
Johns, T., Alonso, J.G., 1990. Glycoalkaloid change during the domestication of thepotato, Solanum section Petota. Euphytica 50, 203–210.
Kaneko, K., Tanaka, M.W., Takahashi, E., Mitsuhashi, H., 1977. Teinemine andisoteinemine. Two new alkaloids from Veratrum grandiflorum. Phytochemistry16, 1620–1622.
Kowalski, S.P., Perez, F.G., Sanford, L.L., Deahl, K.L., 2000. Partial preparativepurification of leptine I from foliage of the wild potato, Solanum chacoense(Bitt.). Prep. Biochem. Biotechnol. 30, 133–144.
Krits, P., Fogelman, E., Ginzberg, I., 2007. Potato steroidal glycoalkaloid levels andthe expression of key isoprenoid metabolic genes. Planta 227, 143–150.
Kuhn, R., Löw, I., 1961. Zur Konstitution der Leptine. Chem. Ber. 94, 1088–1095.Kumar, D., Wareing, P.F., 1972. Factors controlling stolon development in the potato
plant. New Phytol. 71, 639–648.Lafta, A.M., Lorenzen, J.H., 2000. Influence of high temperature and reduced
irradiance on glycoalkaloid levels in potato leaves. J. Am. Soc. Hort. Sci. 125,563–566.
Lawson, D.R., Erb, W.A., Miller, A.R., 1992. Analysis of Solanum alkaloids usinginternal standardization and capillary gas-chromatography. J. Agric. Food Chem.40, 2186–2191.
Lawson, D.R., Green, T.P., Haynes, L.W., Miller, A.R., 1997. Nuclear magneticresonance spectroscopy and mass spectrometry of solanidine, leptinidine, andacetylleptinidine. Steroidal alkaloids from Solanum chacoense Bitter. J. Agric.Food Chem. 45, 4122–4126.
Lawson, D.R., Veilleux, R.E., Miller, A., 1993. Biochemistry and genetics of Solanumchacoense steroidal alkaloids: Natural resistance factors to the Colorado potatobeetle. Curr. Topics Bot. Res. 1, 335–352.
McCue, K.F., Allen, P.V., Shepherd, L.V.T., Blake, A., MacCree, M.M., Rockhold, D.R.,Novy, R.G., Stewart, D., Davies, H.V., Belknap, W.R., 2007. Potato glycosterolrhamnosyltransferase, the terminal step in triose side-chain biosynthesis.Phytochemistry 68, 327–334.
Milner, S.E., Brunton, N.P., Jones, P.W., O’ Brien, N.M., Collins, S.G., Maguire, A.R.,2011. Bioactivities of glycoalkaloids and their aglycones from Solanum species. J.Agric. Food Chem. 59, 3454–3484.
Moco, S., Capanoglu, E., Tikunov, Y., Bino, R.J., Boyacioglu, D., Hall, R.D., Vervoort, J.,De Vos, R.C.H., 2007. Tissue specialization at the metabolite level is perceivedduring the development of tomato fruit. J. Exp. Bot. 58, 4131–4146.
Moehs, C.P., Allen, P.V., Friedman, M., Belknap, W.R., 1997. Cloning and expressionof solanidine UDP-glucose glucosyltransferase from potato. Plant J. 11, 227–236.
Nema, P.K., Ramayya, N., Duncan, E., Niranjan, K., 2008. Potato glycoalkaloids:formation and strategies for mitigation. J. Sci. Food Agric. 88, 1869–1881.
Roddick, J.G., 1982. Distribution of steroidal glycoalkaloids in reciprocal grafts ofSolanum tuberosum L. and Lycopersicon esculentum Mill. Experientia 38, 460–462.
Roddick, J.G., 1989. The acetylcholinesterase-inhibitory activity of steroidalglycoalkaloids and their aglycones. Phytochemistry 28, 2631–2634.
Ronning, C.M., Kowalski, S.P., Sanford, L.L., Stommel, J.R., 2000. Geographicalvariation of solanidane aglycone glycoalkaloids in the wild potato speciesSolanum chacoense Bitter. Genet. Resourc. Crop Evol. 47, 359–369.
Sagredo, B., Lafta, A., Casper, H., Lorenzen, J., 2006. Mapping of genes associatedwith leptine content of tetraploid potato. Theor. Appl. Genet. 114, 131–142.
Sinden, S.L., Sanford, L.L., Cantelo, W.W., Deahl, K., 1988. Bioassays of segregatingplants: a strategy for studying chemical defenses. J. Chem. Ecol. 14, 1941–1950.
Sinden, S.L., Sanford, L.L., Cantelo, W.W., Deahl, K.L., 1986a. Leptine glycoalkaloidsand resistance to the Colorado potato beetle (Coleoptera, Chrysomelidae) inSolanum chacoense. Environ. Entomol. 15, 1057–1062.
Sinden, S.L., Sanford, L.L., Deahl, K.L., 1986b. Segregation of leptine glycoalkaloids inSolanum chacoense Bitter. J. Agric. Food Chem. 34, 372–377.
Tingey, W.M., 1984. Glycoalkaloids as pest resistance factors. Am. Potato J. 61, 157–167.
Yang, Z.B., Park, H.S., Lacy, G.H., Cramer, C.L., 1991. Differential activation of potato3-hydroxy-3-methylglutaryl coenzyme A reductase genes by wounding andpathogen challenge. Plant Cell 3, 397–405.
Zacharius, R.M., Osman, S.F., 1977. Glycoalkaloids in tissue culture of Solanumspecies. Dehydrocommersonine from cultured roots of Solanum chacoense. PlantSci. Lett. 10, 283–287.


















