Staurosporine induces apoptosis in human papillomavirus positive oral cancer cells at G2/M phase by...
6
Staurosporine induces apoptosis in human papillomavirus positive oral cancer cells at G2/M phase by disrupting mitochondrial membrane potential and modulation of cell cytoskeleton Jayanta Sarkar * , Neetu Singh, Sanjeev Meena, Sudhir Sinha Drug Target Discovery and Development Division, Central Drug Research Institute, Lucknow 226 001, India article info Article history: Received 2 April 2009 Received in revised form 23 April 2009 Accepted 23 April 2009 Available online 6 June 2009 Keywords: Oral cancer Staurosporine Apoptosis Cell cycle Mitochondrial membrane potential Cell cytoskeleton summary Our study demonstrates that staurosporine (STS), a protein kinase inhibitor from Streptomyces sp., induces apoptosis in human papillomavirus positive oral carcinoma cells (KB) in a dose dependent man- ner. Growth inhibition studies revealed an IC 50 value of approximately 100 nM. STS induced marked nuclear fragmentation and inter-nucleosomal cleavage compared to untreated cells. It also caused dose dependent disruption of mitochondrial membrane potential and activation of caspase-3, indicating involvement of mitochondria-mediated cell death signaling in KB cell apoptosis. We found time-depen- dent arrest of the KB cells at G2/M phase of cell cycle. Using fluorescence microscopy, we have further shown that STS treatment disrupts microtubules and reorganizes F-actin after 6 h exposure. Taken together, our results suggest that STS induces mitochondria-mediated KB cell apoptosis at G2/M phase by altering cell cytoskeletal network. Ó 2009 Elsevier Ltd. All rights reserved. Introduction Staurosporine (STS), an indolo (2,3-alpha) carbazole was discov- ered in the course of screening of extracts of the bacterium Strep- tomyces sp. for constituent alkaloids with protein kinase C (PKC) inhibitory activity, 1 and it has become the ‘lead’ compound among PKC inhibitors with therapeutic potential. Staurosporine has been used extensively to induce apoptosis in tumor cell lines, 2–4 lym- phocytes, 5,6 neurons 7,8 and other primary cells. 9,10 Two structural analogs of staurosporine, GCP41251 and UCN-01 are currently un- der clinical evaluation as potential anti cancer drugs. 11 Induction of apoptosis in STS treated mammalian cells involves multiple pathways, depending upon the cell type. In primary cul- tures of chick retinal cells, human colonic carcinoma cells SW620, human neuroblastoma cell line SK-N-MC and hepatoma cell lines, STS activates mitochondrial apoptotic pathway. 12–15 In mouse hepatocytes STS induces apoptosis by a mechanism inde- pendent of caspase-8, cytochrome c, caspase-9 and caspase-12. 9 In rat mesenchymal stem cells, STS-induced apoptosis is mediated by voltage-dependent potassium channel. 16 Moreover, a wide range of STS concentrations (100 nM–100 lM) have been used to induce apoptosis in different cell types and a variety of methods have been employed to quantify cell death. Oral cancer refers to a subgroup of head and neck cancers and is synonymous with squamous cell carcinoma (SSC) of oral mucosa origin. Human papillomavirus (HPV) is present in three times as many precancerous oral mucosa and almost five times as many oral cancers compared with normal oral mucosa, indicating its possible role in transforming the oral mucosa. 17 Oncogenic proteins HPV-E6 and HPV-E7 deregulate cell cycle by inactivating p53 and pRb, which may promote tumorogenesis. 18 On the other hand, in HPV immortalized human endocervical cells, an increased level of anti-apoptotic proteins of the mitochondrial pathway makes the cells relatively resistant to apoptosis induced by STS or cisplatin. 19 The present study was aimed at elucidation of the role of mito- chondria and cell cytoskeletoal components in STS-induced apop- tosis in a HPV positive oral cancer cell line KB. We show that mitochondria mediated intrinsic pathway and modulation of cell cytoskeleton play key roles in STS induced KB cell death. Materials and methods Materials Staurosporine (STS), trypsin–EDTA, minimum essential medium Eagle (MEM), gentamicin sulfate salt, amphotericin B, 3-(4,5-di- methyl-2-thiazyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), propidium iodide (PI), Hoechst 33258, caspase-3 apoptosis detection kit, ribonuclease A were purchased from Sigma Chemical 1368-8375/$ - see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.oraloncology.2009.04.009 * Corresponding author. Tel.: +91 522 2612411 4313; fax: +91 522 2623405. E-mail addresses: [email protected], [email protected] (J. Sarkar). Oral Oncology 45 (2009) 974–979 Contents lists available at ScienceDirect Oral Oncology journal homepage: www.elsevier.com/locate/oraloncology
-
Upload
jayanta-sarkar -
Category
Documents
-
view
214 -
download
1
Transcript of Staurosporine induces apoptosis in human papillomavirus positive oral cancer cells at G2/M phase by...

Oral Oncology 45 (2009) 974–979
Contents lists available at ScienceDirect
Oral Oncology
journal homepage: www.elsevier .com/locate /ora loncology
Staurosporine induces apoptosis in human papillomavirus positive oral cancercells at G2/M phase by disrupting mitochondrial membrane potential andmodulation of cell cytoskeleton
Jayanta Sarkar *, Neetu Singh, Sanjeev Meena, Sudhir SinhaDrug Target Discovery and Development Division, Central Drug Research Institute, Lucknow 226 001, India
a r t i c l e i n f o
Article history:Received 2 April 2009Received in revised form 23 April 2009Accepted 23 April 2009Available online 6 June 2009
Keywords:Oral cancerStaurosporineApoptosisCell cycleMitochondrial membrane potentialCell cytoskeleton
1368-8375/$ - see front matter � 2009 Elsevier Ltd. Adoi:10.1016/j.oraloncology.2009.04.009
* Corresponding author. Tel.: +91 522 2612411 431E-mail addresses: [email protected], j_sarka
s u m m a r y
Our study demonstrates that staurosporine (STS), a protein kinase inhibitor from Streptomyces sp.,induces apoptosis in human papillomavirus positive oral carcinoma cells (KB) in a dose dependent man-ner. Growth inhibition studies revealed an IC50 value of approximately 100 nM. STS induced markednuclear fragmentation and inter-nucleosomal cleavage compared to untreated cells. It also caused dosedependent disruption of mitochondrial membrane potential and activation of caspase-3, indicatinginvolvement of mitochondria-mediated cell death signaling in KB cell apoptosis. We found time-depen-dent arrest of the KB cells at G2/M phase of cell cycle. Using fluorescence microscopy, we have furthershown that STS treatment disrupts microtubules and reorganizes F-actin after 6 h exposure. Takentogether, our results suggest that STS induces mitochondria-mediated KB cell apoptosis at G2/M phaseby altering cell cytoskeletal network.
� 2009 Elsevier Ltd. All rights reserved.
Introduction
Staurosporine (STS), an indolo (2,3-alpha) carbazole was discov-ered in the course of screening of extracts of the bacterium Strep-tomyces sp. for constituent alkaloids with protein kinase C (PKC)inhibitory activity,1 and it has become the ‘lead’ compound amongPKC inhibitors with therapeutic potential. Staurosporine has beenused extensively to induce apoptosis in tumor cell lines,2–4 lym-phocytes,5,6 neurons7,8 and other primary cells.9,10 Two structuralanalogs of staurosporine, GCP41251 and UCN-01 are currently un-der clinical evaluation as potential anti cancer drugs.11
Induction of apoptosis in STS treated mammalian cells involvesmultiple pathways, depending upon the cell type. In primary cul-tures of chick retinal cells, human colonic carcinoma cellsSW620, human neuroblastoma cell line SK-N-MC and hepatomacell lines, STS activates mitochondrial apoptotic pathway.12–15 Inmouse hepatocytes STS induces apoptosis by a mechanism inde-pendent of caspase-8, cytochrome c, caspase-9 and caspase-12.9
In rat mesenchymal stem cells, STS-induced apoptosis is mediatedby voltage-dependent potassium channel.16 Moreover, a widerange of STS concentrations (100 nM–100 lM) have been used toinduce apoptosis in different cell types and a variety of methodshave been employed to quantify cell death.
ll rights reserved.
3; fax: +91 522 [email protected] (J. Sarkar).
Oral cancer refers to a subgroup of head and neck cancers and issynonymous with squamous cell carcinoma (SSC) of oral mucosaorigin. Human papillomavirus (HPV) is present in three times asmany precancerous oral mucosa and almost five times as many oralcancers compared with normal oral mucosa, indicating its possiblerole in transforming the oral mucosa.17 Oncogenic proteins HPV-E6and HPV-E7 deregulate cell cycle by inactivating p53 and pRb,which may promote tumorogenesis.18 On the other hand, in HPVimmortalized human endocervical cells, an increased level ofanti-apoptotic proteins of the mitochondrial pathway makes thecells relatively resistant to apoptosis induced by STS or cisplatin.19
The present study was aimed at elucidation of the role of mito-chondria and cell cytoskeletoal components in STS-induced apop-tosis in a HPV positive oral cancer cell line KB. We show thatmitochondria mediated intrinsic pathway and modulation of cellcytoskeleton play key roles in STS induced KB cell death.
Materials and methods
Materials
Staurosporine (STS), trypsin–EDTA, minimum essential mediumEagle (MEM), gentamicin sulfate salt, amphotericin B, 3-(4,5-di-methyl-2-thiazyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT),propidium iodide (PI), Hoechst 33258, caspase-3 apoptosisdetection kit, ribonuclease A were purchased from Sigma Chemical

J. Sarkar et al. / Oral Oncology 45 (2009) 974–979 975
Corporation, USA. MitoCaptureTM apoptosis detection kit wassourced from Merck-Calbiochem, Germany, fetal bovine serum(FBS) was obtained from GIBCO BRL, USA and DNA ladder kit waspurchased from Invitrogen Corporation, USA. Cytoskeletal Rear-rangement kit was purchased from Cellomics Inc., USA.
Cell culture
KB (Human Oral Carcinoma) was procured from ATCC, USA (cat-alog number CCL-17). The cells were cultured in MEM containinggentamicin (50 lg/ml) and amphotericin B (2.5 lg/ml) supple-mented with 10% FBS. Cells up to 15 passages were used for thisstudy.
MTT assay
A colorimetric assay20 using MTT was adopted for a quantitativemeasurement of cell growth and viability. Briefly, 104 cells/well (in96-well culture plates) were exposed to various concentrations ofSTS for 48 h. Untreated cells served as control. Later, 25 ll ofMTT reagent was added to each well and incubated for 2 h. TheMTT solution was removed and formazan crystals dissolved in150 ll of DMSO. After vortexing, plates were read at 540 nm. Cyto-toxicity was expressed as percent cell viability of STS treated cells,relative to untreated controls.
Nuclear morphology study by Hoechst 33258
For monitoring nuclear morphology, KB cells were exposed tovarious concentrations of STS for 24 h, fixed with 4% para formalde-hyde and stained with Hoechst dye (10 lg/ml) for 30 min. Afterwashing with PBS, the cells were analyzed under a fluorescencemicroscope (Leica, Germany) and photographed. Cells with con-densed or fragmented nuclei were considered apoptotic. For eachtreatment minimum 200 cells were counted and the apoptotic indexwas calculated as the percentage of apoptotic cells in the population.
DNA ladder detection
Inter nucleosomal DNA fragmentation (ladder) was determinedusing DNA ladder kit. DNA was extracted from untreated control aswell as STS treated cell as per manufacturer’s protocol. Equal vol-umes of DNA samples were subjected to electrophoresis at 80 Vin 1.5% agarose gel containing 1 lg/ml ethidium bromide.
Colorimetric caspase-3 assay
The caspase-3 activity was measured with a caspase-3 apopto-sis detection kit according to manufacturer’s protocol. Briefly, KBcells were exposed to varying concentrations of STS for 24 h, har-vested and lysed in lysis buffer (4 �C for 30 min). Equal amountsof protein extracts (100 lg) were loaded onto a 96 well microplateand incubated (37 �C for overnight) with reaction buffer, caspasesubstrate, with or without the caspase inhibitors. The liberatedp-nitroaniline (pNA) was read at 405 nm on a micro-plate reader.
DNA cell cycle analysis by flow cytometry
Briefly, 4 � 105 cells/well were seeded in a 6-well plate andtreated with 100 nM STS for varying time periods. Later, the cellswere trypsinized, washed twice with cold PBS, and centrifuged.The cells were then fixed with chilled 75% ethanol (30 min at4 �C), centrifuged, washed with cold PBS and re-suspended in500 ll PBS containing PI (40 lg/ml), RNAse (100 lg/ml) and0.05% Triton X100. After incubation (37 �C, 40 min) in dark, thecells were finally centrifuged, washed with PBS and re-suspended
in 500 ll of PBS. Flow cytometry was performed on fluorescenceactivated cell sorter (FACS, Becton-Dickinson), employing CellQuest software.
Mitochondrial membrane potential analysis (Dwm)
Modulation of mitochondrial membrane potential (Dwm) wasmeasured by fluorescence based MitoCaptureTM apoptosis detectionkit. Cells were seeded into 6-well plates and each well received4 � 105 cells. After overnight growth, cells were exposed to varyingconcentrations of STS for 24 h and harvested in PBS. The cells werethen centrifuged at 500g for 5 min and the pellet was re-suspendedin 1 ml of diluted MitoCaptureTM reagent (1:1000 in incubation buf-fer). Samples were incubated in dark at 37 �C for 15–20 min andcentrifuged. Finally the pellet was re-suspended in 1 ml of pre-warmed incubation buffer and analyzed by flow cytometry.
Cell cytoskeleton assay
The cytoskeletal components of the STS treated KB cells wereanalyzed using Cellomics cytoskeletal rearrangement high contentscreening (HCS) reagent kit. The kit contains fluorescent dyes to la-bel and monitor cell morphology, nuclear morphology, microfila-ment-F actin and microtubule–tubulin. KB cells were treatedwith STS at different concentrations for 6 h and labeled with fluo-rescent dyes as per manufacturer’s protocol. Fluorescent imageswere acquired using 20� objective and analyzed using CellomicsArrayscan� VTI Reader (Cellomics Inc., USA) using Morphology Bio-application software module.
Statistical analysis
All data represents at least three independent experiments eachperformed in duplicate and are expressed as mean ± SE. The signif-icance of statistical data was determined by two tailed Student’s t-test with P < 0.05.
Results
Effect of STS on KB cell viability
The cytotoxicity of STS on KB cells was investigated after anincubation period of 48 h, using colorimetric MTT assay. KB cellproliferation was inhibited in a dose dependent manner in re-sponse to increasing concentration of STS. Based on the dose re-sponse curve, the obtained IC50 value of STS on KB cells wasapproximately 100 nM.
Induction of apoptosis by STS in KB cells
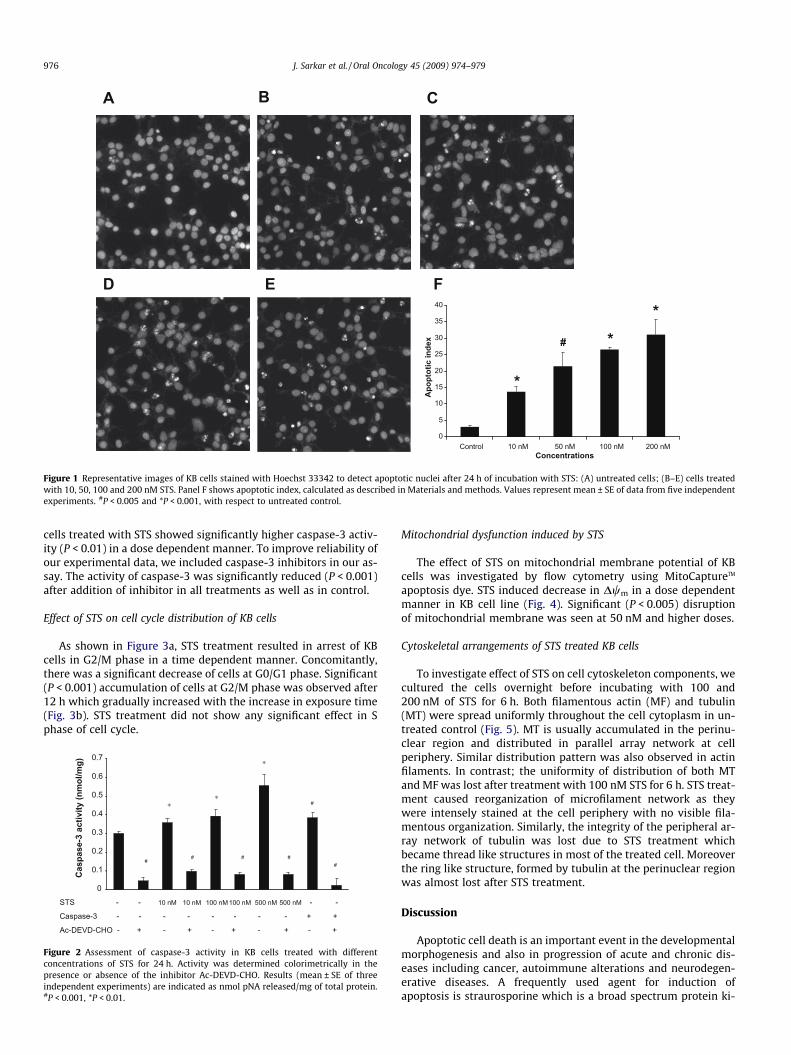
To assess cellular apoptosis induced by STS, KB cells were ex-posed to STS for 24 h, stained with Hoechst dye and examined byfluorescence microscopy. As shown in Figure 1a–e, STS inducedmarked morphologic changes (fragmented nuclei) which are thetypical feature of apoptosis. The level of apoptosis gradually in-creased in a dose dependent manner (Fig. 1f). Consistent withthe morphologic changes seen by Hoechst staining, a DNA ladderpattern of inter-nucleosomal fragmentation – another characteris-tic feature of apoptosis – was distinctly seen for STS treated cells(data not shown).
STS induced caspase-3 mediated apoptosis
Caspase-3 activity was assayed colorimetrically to determine itsrole in STS mediated apoptosis. As can be seen from Figure 2, KB
A
0
5
10
15
20
25
30
35
40
Control 10 nM 50 nM 100 nM 200 nMConcentrations
Apo
ptot
ic in
dex
*
**
#
B C
D E F
Figure 1 Representative images of KB cells stained with Hoechst 33342 to detect apoptotic nuclei after 24 h of incubation with STS: (A) untreated cells; (B–E) cells treatedwith 10, 50, 100 and 200 nM STS. Panel F shows apoptotic index, calculated as described in Materials and methods. Values represent mean ± SE of data from five independentexperiments. #P < 0.005 and *P < 0.001, with respect to untreated control.
976 J. Sarkar et al. / Oral Oncology 45 (2009) 974–979
cells treated with STS showed significantly higher caspase-3 activ-ity (P < 0.01) in a dose dependent manner. To improve reliability ofour experimental data, we included caspase-3 inhibitors in our as-say. The activity of caspase-3 was significantly reduced (P < 0.001)after addition of inhibitor in all treatments as well as in control.
Effect of STS on cell cycle distribution of KB cells
As shown in Figure 3a, STS treatment resulted in arrest of KBcells in G2/M phase in a time dependent manner. Concomitantly,there was a significant decrease of cells at G0/G1 phase. Significant(P < 0.001) accumulation of cells at G2/M phase was observed after12 h which gradually increased with the increase in exposure time(Fig. 3b). STS treatment did not show any significant effect in Sphase of cell cycle.
## # #
#
#
∗∗
∗
STS - - 10 nM 10 nM 100 nM 100 nM 500 nM 500 nM - -
Caspase-3 - - - - - - - - + +
Ac-DEVD-CHO - + - + - + - + - +
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Cas
pase
-3 a
ctiv
ity (n
mol
/mg)
Figure 2 Assessment of caspase-3 activity in KB cells treated with differentconcentrations of STS for 24 h. Activity was determined colorimetrically in thepresence or absence of the inhibitor Ac-DEVD-CHO. Results (mean ± SE of threeindependent experiments) are indicated as nmol pNA released/mg of total protein.#P < 0.001, *P < 0.01.
Mitochondrial dysfunction induced by STS
The effect of STS on mitochondrial membrane potential of KBcells was investigated by flow cytometry using MitoCaptureTM
apoptosis dye. STS induced decrease in Dwm in a dose dependentmanner in KB cell line (Fig. 4). Significant (P < 0.005) disruptionof mitochondrial membrane was seen at 50 nM and higher doses.
Cytoskeletal arrangements of STS treated KB cells
To investigate effect of STS on cell cytoskeleton components, wecultured the cells overnight before incubating with 100 and200 nM of STS for 6 h. Both filamentous actin (MF) and tubulin(MT) were spread uniformly throughout the cell cytoplasm in un-treated control (Fig. 5). MT is usually accumulated in the perinu-clear region and distributed in parallel array network at cellperiphery. Similar distribution pattern was also observed in actinfilaments. In contrast; the uniformity of distribution of both MTand MF was lost after treatment with 100 nM STS for 6 h. STS treat-ment caused reorganization of microfilament network as theywere intensely stained at the cell periphery with no visible fila-mentous organization. Similarly, the integrity of the peripheral ar-ray network of tubulin was lost due to STS treatment whichbecame thread like structures in most of the treated cell. Moreoverthe ring like structure, formed by tubulin at the perinuclear regionwas almost lost after STS treatment.
Discussion
Apoptotic cell death is an important event in the developmentalmorphogenesis and also in progression of acute and chronic dis-eases including cancer, autoimmune alterations and neurodegen-erative diseases. A frequently used agent for induction ofapoptosis is straurosporine which is a broad spectrum protein ki-

010203040
50607080
Control 12 h 15 h 18 h 21 h 24 hTime intervals
% o
f ce
lls
G0/G1 phase G2/M Phase
* * * **
*
*
*
*
*
Figure 3 STS induced cell cycle arrest in KB cells. Cells were treated with 100 nM STS and evaluated by flow cytometry at indicated time intervals. Panels A–E show cell cyclephase distribution. Panel F shows quantification (mean ± SE of three experiments) of cells in G0/G1 and G2/M phases. *P < 0.001.
0
5
10
15
20
25
Ctrl 10nM 50nM 100nM 200nM 500nM
Concentrations
Mito
Cap
ture
TM F
luor
esce
ne in
tens
ity
(Red
/Gre
en ra
tio)
* * **
Figure 4 Loss of mitochondrial membrane potential (Dwm) in KB cells induced bySTS. Cells treated with STS were harvested and stained with MitoCaptureTM cationicdye and analyzed by flow cytometry. Dwm value (mean ± SE of three independentexperiments) was based on red/green fluorescence ratio. *P < 0.001.
J. Sarkar et al. / Oral Oncology 45 (2009) 974–979 977
nase inhibitor isolated from Streptomyces. Although there arenumerous reports on induction of apoptosis by staurosporine andits structural analogs in a variety of cell types, the molecular mech-anism(s) involved is still not clear and it is considered more com-plex than any other anticancer drugs.21
To evaluate cytotoxic activity, we had exposed KB cells to strau-rosporine for 48 h so as to get optimum effect of the molecule atlower concentrations. On the other hand, apoptosis was deter-mined at an earlier time point (24 h) because in cultured mamma-lian cells apoptosis precedes secondary necrosis.22 Further, sinceapoptosis occurs in a complex cascade of signaling pathways andmany features of apoptosis and necrosis can overlap, it is impor-tant to employ two or more assays to confirm that cell death isoccurring via apoptosis. In this study, we employed two methodsto define apoptosis – (a) fluorescence microscopy of Hoechst dyestained KB cells and (b) inter nucleosomal DNA fragmentation(DNA ladder pattern).
Our results show that staurosporine arrests KB cell cycle at G2/M phase with a concomitant decrease in G0/G1 population. Similareffects of staurosporine have earlier been reported for squamouscell carcinoma and oral cancer cells,23,24 though its structural ana-log UCN-01 (7-hydroxystaurosporine) arrested the cells at G1phase.25 We have also observed that the cell cycle arrest as wellas apoptosis in KB cells occurs at lower concentrations than that
(20 lM) reported earlier.23 It has been shown that HPV-E6 proteindecreases the vigilance of mitotic checkpoints by targeting p53degradation.26 Therefore, down regulation of HPV-E6 by stauro-sporine27 and the resultant elevation in p53 activity could also con-tribute to KB cell cycle arrest at G2/M phase.
Straurosporine caused significant modulation of mitochondrialmembrane potential (Dwm) in a dose dependent manner. Concom-itantly, we also observed a boost in caspase-3 activity in KB cellsafter exposure to straurosporine. These results, supported withthe DNA fragmentation analysis, suggest cytochrome c mediatedactivation of ‘apoptosome’ upon loss of mitochondrial membranepotential.28 Apoptosome subsequently cleaves and activates down-stream caspase-3 which in turn degrades of cellular DNA.29
We found that straurosporine disrupts tubulin network in intactKB cells. Microtubules play an important role in maintaining a vari-ety of fundamental cellular functions including sustained shape,transportation of vesicles and protein complexes, and regulationof motility and cell division.30 In addition, disruption of microtu-bules by straurosporine may lead to altered mitochondrial distri-bution and interference in mitochondrial activity by modulatingits shape.31 Survivin – an antiapoptotic protein abundantly ex-pressed in oral squamous cell carcinoma32,33 – controls apoptosisduring the G2/M phase of cell cycle through its association withmitotic spindle microtubules. Disruption of survivin–microtubuleinteraction results in loss of antiapoptotic function and increasedcaspase-3 activity.34 It can thus be assumed that straurosporine-in-duced early phase (6 h) disruption of microtubules (thereby dis-ruption of the survivin–microtubule complex) is an importantmediator of late phase (12 h onwards) cell cycle arrest and apopto-sis in the G2/M phase.
In the present study we also compared the configuration of ac-tin filaments in healthy and apoptotic cells. Our results clearlydemonstrate the reorganization of filamentous network of F-actinwhich stains intensely towards cell periphery in the apoptoticcells. Actin cytoskeleton has already been reported as a critical ele-ment in apoptotic cellular remodeling. The two main morphologi-cal features mediated by actin in process of apoptosis areapoptosis-blebbing of the plasma membrane, and cell disintegra-tion into apoptotic bodies.35,36
In conclusion, our results clearly demonstrate that straurospo-rine induces apoptosis in KB cancer cells by arresting them inG2/M phase which may be due to the disorganization of cytoskel-

Figure 5 Cytoskeletal configuration of KB cells before (upper panels, with 20� objective) and after (middle and lower panels) STS treatment. Images were acquired usingCellomics ArrayScan HCS Reader.
978 J. Sarkar et al. / Oral Oncology 45 (2009) 974–979
etal network, especially microtubules. We have also shown thatmitochondrial intrinsic apoptotic pathway is the key mediator ofstraurosporine induced KB cell apoptosis. However, additionalanalysis of intrinsic pathway is needed. Further, the role of extrin-sic pathway in straurosporine mediated KB cell death is yet to beexplored.
Conflict of Interest Statement
None declared.
Acknowledgements
The work has been financially supported by CSIR NetworkProject IAP-0001 and MoES Project ‘Drugs from the Sea’. Weare thankful to Mr. A.L. Vishwakarma for experiments withFlow Cytometry. This manuscript is CDRI communication no.7699.
References
1. Omura S, Iwai Y, Hirano A, Nakagawa A, Awaya J, Tsuchya H, et al. A newalkaloid AM-2282 OF streptomyces origin. Taxonomy, fermentation, isolationand preliminary characterization. J Antibiot 1977;30(4):275–82.
2. Zhang H, Vollmer M, De Geyter M, Durrenberger M, De Geyter C. Apoptosis anddifferentiation induced by staurosporine in granulosa tumor cells is coupledwith activation of JNK and suppression of p38 MAPK. Int J Oncol2005;26(6):1575–80.
3. Takehara T, Liu X, Fujimoto J, Friedman SL, Takahashi H. Expression and role ofBcl-xL in human hepatocellular carcinomas. Hepatology 2001;34(1):55–61.
4. Giuliano M, Bellavia G, Lauricella M, D’Anneo A, Vassallo B, Vento R, et al.Staurosporine-induced apoptosis in Chang liver cells is associated with down-regulation of Bcl-2 and Bcl-XL. Int J Mol Med 2004;13(4):565–71.
5. Knox KA, Finney M, Milner AE, Gregory CD, Wakelam MJ, Michell RH, et al.Second-messenger pathways involved in the regulation of survival in germinal-centre B cells and in Burkitt lymphoma lines. Int J Cancer 1992;52(6):959–66.
6. Suzuki K, Azuma Y, Onishi Y, Kizaki H, Ishimura Y. Biphasic effect ofstaurosporine on thymocyte apoptosis. Biochem Mol Biol Int1995;35(5):1085–92.
7. McCarthy MJ, Rubin LL, Philpott KL. Involvement of caspases in sympatheticneuron apoptosis. J Cell Sci 1997;110(Pt 18):2165–73.
8. MacManus JP, Rasquinha I, Black MA, Laferriere NB, Monette R, Walker T, et al.Glutamate-treated rat cortical neuronal cultures die in a way different from theclassical apoptosis induced by staurosporine. Exp Cell Res 1997;233(2):310–20.
9. Feng G, Kaplowitz N. Mechanism of staurosporine-induced apoptosis in murinehepatocytes. Am J Physiol Gastrointest Liver Physiol 2002;282(5):G825–34.
10. Weil M, Jacobson MD, Coles HS, Davies TJ, Gardner RL, Raff KD, et al.Constitutive expression of the machinery for programmed cell death. J CellBiol 1996;133(5):1053–9.
11. Gescher A. Staurosporine analogues – pharmacological toys or usefulantitumour agents? Crit Rev Oncol Hematol 2000;34(2):127–35.
12. Gil J, Almeida S, Oliveira CR, Rego AC. Cytosolic and mitochondrial ROS instaurosporine-induced retinal cell apoptosis. Free Radic Biol Med2003;35(11):1500–14.
13. Heerdt BG, Houston MA, Mariadason JM, Augenlicht LH. Dissociation ofstaurosporine-induced apoptosis from G2-M arrest in SW620 human coloniccarcinoma cells: initiation of the apoptotic cascade is associated with elevationof the mitochondrial membrane potential (deltapsim). Cancer Res2000;60(23):6704–13.
14. Chandele A, Prasad V, Jagtap JC, Shukla R, Shastry PR. Upregulation of survivinin G2/M cells and inhibition of caspase 9 activity enhances resistance instaurosporine-induced apoptosis. Neoplasia 2004;6(1):29–40.
15. Wang Y, Zhang B, Peng X, Perpetua M, Harbrecht BG. Bcl-xL preventsstaurosporine-induced hepatocyte apoptosis by restoring protein kinase B/

J. Sarkar et al. / Oral Oncology 45 (2009) 974–979 979
mitogen-activated protein kinase activity and mitochondria integrity. J CellPhysiol 2008;215(3):676–83.
16. Wang S, Wang JA, Li J, Zhou J, Wang H. Voltage-dependent potassium channelsare involved in staurosporine-induced apoptosis of rat mesenchymal stemcells. Cell Biol Int 2008;32(2):312–9.
17. Reddout N, Christensen T, Bunnel A, Jensen D, Johnson D, O’Malley S, et al. Highrisk HPV types 18, 16 are potent modulators of oral squamous cell carcinomaphenotypes in vitro. Infect Agent Cancer 2007;2:21.
18. zur Hausen H. Papillomavirus infections – a major cause of human cancers.Biochim Biophys Acta 1996;1288(2):F55–78.
19. Yang X, Hao Y, Pater MM, Tang SC, Pater A. Enhanced expression of anti-apoptotic proteins in human papillomavirus immortalized and cigarette smokecondensate transformed human endocervical cells: correlation with resistanceto apoptosis induced by DNA damage. Mol Carcinog 1998;22(2):95–101.
20. Mosmann T. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J Immunol Methods1983;65(1–2):55–63.
21. Stepczynska A, Lauber K, Engels IH, Janssen O, Kabelitz D, Wesselborg S, et al.Staurosporine and conventional anticancer drugs induce overlapping, yetdistinct pathways of apoptosis and caspase activation. Oncogene2001;20(10):1193–202.
22. Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol2007;35(4):495–516.
23. Swe M, Bay BH, Sit KH. Interphase and M-phase oral KB carcinoma cells aretargetted in staurosporine-induced apoptosis. Cancer Lett 1996;104(2):145–52.
24. Zhang YX, Yu SB, Ou-Yang JP, Xia D, Wang M, Li JR. Effect of protein kinase Calpha, caspase-3, and survivin on apoptosis of oral cancer cells induced bystaurosporine. Acta Pharmacol Sin 2005;26(11):1365–72.
25. Otsubo A, Bhawal UK, Nomura Y, Mitani Y, Ozawa K, Kuniyasu H, et al. UCN-01(7-hydroxystaurosporine) induces apoptosis and G1 arrest of both primary and
metastatic oral cancer cell lines in vitro. Oral Surg Oral Med Oral Pathol OralRadiol Endod 2007;103(3):391–7.
26. Thompson DA, Belinsky G, Chang TH, Jones DL, Schlegel R, Munger K. Thehuman papillomavirus-16 E6 oncoprotein decreases the vigilance of mitoticcheckpoints. Oncogene 1997;15(25):3025–35.
27. Bernard B, Pretet JL, Charlot JF, Mougin C. Human papillomaviruses type 16+and 18+ cervical carcinoma cells are sensitive to staurosporine-mediatedapoptosis. Biol Cell 2003;95(1):17–26.
28. Arnoult D, Gaume B, Karbowski M, Sharpe JC, Cecconi F, Youle RJ. Mitochondrialrelease of AIF and EndoG requires caspase activation downstream of Bax/Bak-mediated permeabilization. Embo J 2003;22(17):4385–99.
29. Sakahira H, Enari M, Nagata S. Cleavage of CAD inhibitor in CAD activation andDNA degradation during apoptosis. Nature 1998;391(6662):96–9.
30. Margolis RL, Wilson L. Microtubule treadmilling: what goes around comesaround. Bioessays 1998;20(10):830–6.
31. Heggeness MH, Simon M, Singer SJ. Association of mitochondria withmicrotubules in cultured cells. Proc Natl Acad Sci USA 1978;75(8):3863–6.
32. Lo Muzio L, Pannone G, Staibano S, Mignogna MD, Rubini C, Mariggio MA, et al.Survivin expression in oral squamous cell carcinoma. Br J Cancer2003;89(12):2244–8.
33. Tanaka C, Uzawa K, Shibahara T, Yokoe H, Noma H, Tanzawa H. Expression of aninhibitor of apoptosis, surviving, in oral carcinogenesis. J Dent Res2003;82(8):607–11.
34. Li F, Ambrosini G, Chu EY, Plescia J, Tognin S, Marchisio PC, et al. Control ofapoptosis and mitotic spindle checkpoint by survivin. Nature1998;396(6711):580–4.
35. Cotter TG, Lennon SV, Glynn JM, Green DR. Microfilament-disrupting agentsprevent the formation of apoptotic bodies in tumor cells undergoing apoptosis.Cancer Res 1992;52(4):997–1005.
36. Cervinka M, Cerman J, Rudolf E. Apoptosis in Hep2 cells treated with etoposideand colchicine. Cancer Detect Prev 2004;28(3):214–26.