
Staging of middle and late embryonic development in the medicinal leech,Hirudo medicinalis
13
Staging of Middle and Late Embryonic Development in the Medicinal Leech, Hirudo medicinalis SHIRLEY A. REYNOLDS, KATHLEEN A. FRENCH,* ANDREAS BAADER, AND WILLIAM B. KRISTAN, JR. Department of Biology, University of California - San Diego, La Jolla, California 92093–0357 ABSTRACT We present a description of the last half of embryonic development in the European medicinal leech, Hirudo medicinalis, based entirely on externally visible morphological features, and establish reliably observable stages during that development. Embryogenesis, from the time fertilized eggs are deposited in an eggcase (called a cocoon) to the emergence of juveniles from the cocoon, takes approximately 4 weeks at room temperature. The stages described in this paper extend from the completion of segmentation to the appearance of the final bands of pigmentation. Developmental stages are expressed as percentages of total embryonic developmental time. This staging table was constructed for embryos kept at 20°C. In addition, the development of animals kept at 17°C or at 24°C was compared with those held at 20°C. Development proceeds more quickly at higher temperatures. Because development in embryos held at higher or lower temperatures was linearly related to the stages determined for embryos held at 20°C, the rate of development at any intermediate temperature can be predicted from the staging table at 20°C by simple multiplication. J. Comp. Neurol. 402:155–167, 1998. r 1998 Wiley-Liss, Inc. Indexing terms: embryogenesis; behavior; nervous system; cell fate; ontogeny The medicinal leech, an annelid worm, has been widely used to study the neuronal basis of behavior and develop- ment of the central nervous system (Muller et al., 1981; Loer and Kristan, 1989; French et al., 1992; Drapeau et al., 1995). Although early development of this leech has been previously described in a paper covering early embryonic organization and segmentation, which occur during ap- proximately the first half of embryogenesis (Fernandez and Stent, 1982), the last half of embryogenesis has remained less well categorized. In this paper, we present a staging table which covers the last 60% of embryogenesis, overlapping with the previous staging table just enough to bring them into register. In nature, fertilized leech eggs are deposited at the edge of a freshwater pond in a watertight, yolky-fluid-filled cocoon, from which they emerge as adult-like juveniles approximately 4 weeks later. Emerging juveniles behave very much like adults, except that they lack reproductive behavior (Reynolds et al., 1998). Much of a leech’s body form and behavior develops during the last 60% of embryo- genesis. However, until now there has been no staging table for this later developmental period, and the timing of events has typically been characterized in terms of the estimated number of days elapsed since the eggs were deposited (Glover and Mason, 1986; Loer, et al., 1987; Jellies and Kristan, 1991; French et al., 1992). To facilitate further study of events during the latter half of develop- ment, we have established specific criteria to define recog- nizable stages in the development of this species. We have used only morphological features that are visible in intact, undyed, freely moving embryos. Because the rate of devel- opment varies with ambient temperature, we have ex- pressed the stages in terms of percentage of developmental progress, as in grasshopper staging (Bentley et al., 1979), rather than in absolute number of days. MATERIALS AND METHODS Developmental observations Progenitor leeches were obtained from Leeches, USA, Westbury, NY. After multiple blood meals, they were maintained in groups of 10 in terraria containing sphag- num moss and some standing water. The terraria were Grant sponsor: NIH; Grant number: NS25916. *Correspondence to: Kathleen A. French, Dept. of Biology, 0357, U.C.S.D., 9500 Gilman Drive, La Jolla, CA 92093–0357. E-mail: [email protected] Received 28 October 1997; Revised 23 July 1998; Accepted 28 July 1998 THE JOURNAL OF COMPARATIVE NEUROLOGY 402:155–167 (1998) r 1998 WILEY-LISS, INC.
Transcript of Staging of middle and late embryonic development in the medicinal leech,Hirudo medicinalis

Staging of Middle and Late EmbryonicDevelopment in the Medicinal Leech,
Hirudo medicinalis
SHIRLEY A. REYNOLDS, KATHLEEN A. FRENCH,* ANDREAS BAADER,
AND WILLIAM B. KRISTAN, JR.
Department of Biology, University of California - San Diego, La Jolla, California 92093–0357
ABSTRACTWe present a description of the last half of embryonic development in the European
medicinal leech, Hirudo medicinalis, based entirely on externally visible morphologicalfeatures, and establish reliably observable stages during that development. Embryogenesis,from the time fertilized eggs are deposited in an eggcase (called a cocoon) to the emergence ofjuveniles from the cocoon, takes approximately 4 weeks at room temperature. The stagesdescribed in this paper extend from the completion of segmentation to the appearance of thefinal bands of pigmentation. Developmental stages are expressed as percentages of totalembryonic developmental time. This staging table was constructed for embryos kept at 20°C.In addition, the development of animals kept at 17°C or at 24°C was compared with those heldat 20°C. Development proceeds more quickly at higher temperatures. Because development inembryos held at higher or lower temperatures was linearly related to the stages determinedfor embryos held at 20°C, the rate of development at any intermediate temperature can bepredicted from the staging table at 20°C by simple multiplication. J. Comp. Neurol.402:155–167, 1998. r 1998 Wiley-Liss, Inc.
Indexing terms: embryogenesis; behavior; nervous system; cell fate; ontogeny
The medicinal leech, an annelid worm, has been widelyused to study the neuronal basis of behavior and develop-ment of the central nervous system (Muller et al., 1981;Loer and Kristan, 1989; French et al., 1992; Drapeau et al.,1995). Although early development of this leech has beenpreviously described in a paper covering early embryonicorganization and segmentation, which occur during ap-proximately the first half of embryogenesis (Fernandezand Stent, 1982), the last half of embryogenesis hasremained less well categorized. In this paper, we present astaging table which covers the last 60% of embryogenesis,overlapping with the previous staging table just enough tobring them into register.
In nature, fertilized leech eggs are deposited at the edgeof a freshwater pond in a watertight, yolky-fluid-filledcocoon, from which they emerge as adult-like juvenilesapproximately 4 weeks later. Emerging juveniles behavevery much like adults, except that they lack reproductivebehavior (Reynolds et al., 1998). Much of a leech’s bodyform and behavior develops during the last 60% of embryo-genesis. However, until now there has been no stagingtable for this later developmental period, and the timing ofevents has typically been characterized in terms of theestimated number of days elapsed since the eggs weredeposited (Glover and Mason, 1986; Loer, et al., 1987;
Jellies and Kristan, 1991; French et al., 1992). To facilitatefurther study of events during the latter half of develop-ment, we have established specific criteria to define recog-nizable stages in the development of this species. We haveused only morphological features that are visible in intact,undyed, freely moving embryos. Because the rate of devel-opment varies with ambient temperature, we have ex-pressed the stages in terms of percentage of developmentalprogress, as in grasshopper staging (Bentley et al., 1979),rather than in absolute number of days.
MATERIALS AND METHODS
Developmental observations
Progenitor leeches were obtained from Leeches, USA,Westbury, NY. After multiple blood meals, they weremaintained in groups of 10 in terraria containing sphag-num moss and some standing water. The terraria were
Grant sponsor: NIH; Grant number: NS25916.*Correspondence to: Kathleen A. French, Dept. of Biology, 0357, U.C.S.D.,
9500 Gilman Drive, La Jolla, CA 92093–0357. E-mail: [email protected] 28 October 1997; Revised 23 July 1998; Accepted 28 July 1998
THE JOURNAL OF COMPARATIVE NEUROLOGY 402:155–167 (1998)
r 1998 WILEY-LISS, INC.

observed daily, and the time at which cocoons were depos-ited was noted as precisely as possible. The moss wassearched 2–3 times per week, and any cocoons found in themoss were removed and labeled. Six embryos from each offive cocoons were allowed to develop in their cocoon atroom temperature (about 22°C) for the first 10 days, thenremoved from the cocoon and placed in individually num-bered Petri dishes at 20°C 6 0.5°C for an additional 20days. Two additional cocoons were used to determine thetiming of early features and the total time required fordevelopment at 20°C. These two cocoons were placed in the20°C incubator within the first 24 hours after they weredeposited and were held there, except during periods ofobservation, until the ventrolateral stripe appeared.Twenty embryos were observed from these cocoons. Deathof embryos reduced the number observed at some of thelater time points, but we used an average of 32 leeches tocalculate the time of onset for each trait. All cocoons weredeposited at least a month apart except for the final two,which were deposited 1 day apart and were observed intandem. The deposition of cocoons is not usually observed,so the timing of the first five cocoons was not knownexactly. The approximate stage at which each cocoon wasopened was judged by candling the intact cocoon, as well asby counting calendar days since the estimated time ofcocoon deposition. (Breeding tanks were observed everyday, so in most cases we knew the day and time when eggdeposition began. In addition, the color of a cocoon changesfrom white through pink to tan during the 2 days followingdeposition, providing further evidence of the chronologicalage of embryos.) In two cases, embryos were also observedwhile they were still in the open cocoon to confirm theirdevelopmental progress.
Morphological and behavioral observations were madethrough a Wild M7A dissection microscope (63–303)every 8 hours for 17 days, and then once every 24 hours for3 more days, or, in the case of the two cocoons heldcontinuously at 20°C, every 8 hours up until the embryosreached the endpoint. To avoid heating the embryos duringobservations, fiber-optic lighting was used, and many ofthe features were best seen by using side-lighting. Physi-cal features observed for developmental changes were thelarval mouth, the rear sucker, annuli, genital pores, larvalsac peristalsis, dorsal fusion, lateral heart tubes, gutpouches, eyes, and body pigmentation. The time of onsetlisted for each feature is the mean time when it firstappeared for all the embryos. Times are listed as percent ofthe total time of embryonic development, abbreviated aspercent embryonic development (% ED). Photomicro-graphs were made with the same microscope used for theserial observations, except Figure 4, which was takenthrough a Zeiss compound fluorescence microscope. Someembryos were anesthetized in 4–8% ethanol prior to beingphotographed. The behavioral changes observed will bediscussed in another paper (Reynolds et al., 1998).
Correlating data from individual cocoons
For the first five cocoons, the stage of the embryos at thetime the cocoons were opened was between 47% and 57% ofthe total time of embryonic development, judged post hocas follows. After observations were complete, the five setsof developmental data needed to be temporally alignedwith each other. Because the time at which the cocoon wasdeposited was known only approximately, the beginning ofembryogenesis was undefined for these cocoons. In addi-
tion, a defined endpoint was required, and the time atwhich embryos normally hatch from their cocoon turnedout to be unsatisfactory. First, from observing many co-coons other than our experimental subjects, we discoveredthat the time at which embryos emerge from their cocoonsvaries greatly. Second, because we removed the embryosfrom their cocoon we could not know when they would havehatched. We therefore needed a morphological criterionthat reliably appeared near the end of embryogenesis. Thedark longitudinal stripes on the ventral surface firstappear as faint pigment speckles in this area very late inembryogenesis, making them an appropriate identifiableendpoint for the time vs. temperature study as well as forour final two cocoons used in the staging study.
In addition, because five cocoons were initially raised atroom temperature and only later moved to 20°C, weexpected the rate of development to have changed as aresult of the temperature shift. To compare observations ofthese embryos with observations of embryos held at 20°Cthroughout development, it was necessary to determinethe time required for an embryo to shift from the develop-mental rate characteristic of room temperature to the ratecharacteristic of 20°C. Our time and temperature studyrevealed that the effects of changing temperature from20°C to 24°C were first detected no sooner than 15 hoursafter the temperature change. Based on this observation,when we aligned the data for these cocoons with those forthe constant-temperature cocoons, we omitted the first 16hours of data following the temperature shift.
The final two cocoons were opened slightly earlier thanany of the first five cocoons, and 10 embryos from eachcocoon were held at 20°C and observed until they reachedthe endpoint criterion. We observed these embryos throughthe initial appearance of pigment in the ventral stripesand then temporally aligned all seven cocoons beforecalculating the mean time at which each trait appeared.The temporal alignment was based on the time at whichpigment appeared on caudal sucker. This trait was chosenbecause it was the trait with the smallest temporalvariability, both within and between cocoons. (Variabilitybetween cocoons was calculated by determining the rela-tive timing of several readily discernible traits, each ofwhich had low temporal variability within a cocoon.) Thetime from cocoon deposition to the appearance of tailpigment in the cocoons raised entirely at 20°C was used asthe standard; the mean time at which the tail pigmentappeared in embryos from each of the other five cocoonswas set equal to this standard, and the time at which eachof the other features appeared was calculated back fromthis benchmark.
When embryos were removed from their cocoon, siblingswithin each cocoon varied by no more than 1.5% ED fromthe mean stage in their cocoon, and they remained within2% of the mean stage for 99.5% of the observationsthroughout embryogenesis.
Effects of temperature on the timing ofdevelopmental events
Two cocoons were used to determine quantitatively theeffect of temperature on developmental rate. Embryoswere raised in their cocoons at 20°C for the first 46% (forone cocoon) or 56% (for the other cocoon) of developmentand were then placed in individual Petri dishes and held atone of three temperatures: 17°C, 20°C, or 24°C. Eachexperimental group contained three to five embryos. These
156 S.A. REYNOLDS ET AL.

temperatures were chosen because preliminary ob-servations indicated that they are near the limits ofviability for the embryos. Observations were made asdescribed above throughout the remainder of embryogen-esis. The embryos held at 20°C in these experiments werenot included in the calculation of values for the stagingtable.
Staining used to determine cell fates
Additional embryos (n 5 6) were used to determine thefate of cells within the larval mouth. Portions of the larvalmouth were labeled by applying the lipophilic fluorescentmarker Di-I (1,1-dioctadecyl-3,3,38,38-tetramethyl-indocar-bocyanine perchlorate, Molecular Probes, Eugene, OR).Di-I spreads throughout the membrane of the cells towhich it is applied, but does not spread to other cells(Honig and Hume, 1986). Embryos were labeled between44% and 46% ED, and the location of the marker wasobserved at 71–95% ED by using a fluorescence microscopewith a rhodamine filter, both in the whole embryo andagain after dissection.
RESULTS
The relative order of developmental events observed inthis study (Tables 1 and 2) was invariant, although theprecise timing with which features appeared varied amongindividual embryos. Figures 1 and 2 show the progressionthrough developmental time, at intervals of 5% ED. Spe-cific features will be discussed below. Embryonic sizecannot be used as a staging indicator because individualembryos normally vary greatly in size.
Larval shape
At 40% ED (Fig. 1), the germinal plate of each embryoappeared as a thin strip of opaque white tissue on a muchlarger balloon of clearer tissue, the larval sac, whichcontains yolk granules and a yolky fluid. At this stage, the
overall shape of the embryo was very irregular, lookingroughly like a bubbly triangle (Fig. 3). Over the next fewdays, the embryo gradually assumed a smoother oblateform, like a round-ended cigar.
Peristalsis of the larval sac
The larval sac contains larval muscles, which are de-rived from larval mesenchyme (Sawyer, 1986). Prior toabout 40% ED, these muscles caused apparently randomlyoccurring contractions of the yolk sac. However, byabout 45% ED when the larval sac was becoming oblate,the larval muscles contracted in peristaltic waves thattraveled longitudinally along the sac. Strong peristalticcontractions continued from this time until about58% ED, when they became noticeably weaker. The larvalsac ceased contracting altogether by 62.5% ED, whenthe germinal plate had overgrown most of the larval sactissue. The cessation of peristalsis also provided ad-vance notice that fusion along the dorsal midline wouldstart in another 3% ED (approximately 1 day at 20°C).Premature weakening or cessation of larval sac peristalsiswas often the first indication that an embryo was sick ordying.
Larval mouth
The early embryonic leech (‘‘larva’’) ingests yolky fluidvia a larval mouth located at the anterior end of thegerminal plate; the yolky fluid surrounding embryos insidea cocoon is ingested as the larval mouth opens and closes,filling the larval sac (Fernandez and Stent, 1982). At 40%ED (the earliest time cocoons were opened in this study),
TABLE 1. Appearance of Hirudo medicinalis Embryos at Intervals of 5%of the Time Between Cocoon Deposition (0% Embryonic Development, ED)
and the Appearance of the Ventrolateral Stripe (100% ED)1
0–40% ED: Refer to Fernandez and Stent (1982).40% ED: Larval mouth actively opens and closes;1 germinal plate is small and fan-
shaped (widest at the anterior end, tapering to a point at the posterior end);nephridia have not yet formed; annuli have not yet appeared; overall larvalshape is irregular.
45% ED: Larval shape tends towards oblate, but is still somewhat irregular;nephridia and annuli have not yet appeared; rear sucker primordium is still con-tinuous with the rest of the germinal plate.
50% ED: Developing rear sucker primordium is distinct and slightly raised; annuli,nephridiopores, and genital pores are present; larval shape is oblate; larvalmouth has been internalized; germinal plate is ovoid in shape.
55% ED: Large medial eyes are present, but unpigmented; a short region of dorsalfusion has occurred at anterior; rudimentary heart tubes may be visible, but arenot contractile; rear sucker is puckered or bowl-shaped; rudimentary jaws canbe seen in mouth.
60% ED: Heart tubes are contractile and contain blood; large anterior eyes arefaintly pigmented; larval nephridia are collapsing into the fluid space under thelarval membrane; gut pouches are clearly visible; larval sac peristalsis is weak.
65% ED: Secondary blood vessels are visible; larval sac peristalsis has ceased;larval nephridia are gone; posteriormost pair of gut pouches is much longer thanneighboring gut pouches; there is a short region of dorsal fusion at the posteriorend of the germinal plate.
70% ED: Dorsal fusion is complete; posterior pair of gut pouches have reached fulllength.
75% ED: Dorsal fusion line has decreased in prominence.80% ED: Primary body pigment stripes are visible.85% ED: Dorsal fusion line is visible subcutaneously only.90% ED: Secondary body pigmentation is visible.95% ED: Tail sucker has pigment stripes; dorsal fusion line is completely gone.100% ED: Dark ventrolateral stripe is barely visible.
1Italics indicate the most unequivocal feature.
TABLE 2. Development of Morphological Features of Hirudo medicinalisEmbryos in Chronological Order1
Age(%ED) Developments
45.5 Larval mouth inactive and receding46.0 Nephridiopores appear46.5 Larval shape oblate47.5 Rear sucker primordium distinct from the rest of the germinal plate;
annuli appear48.0 Germinal plate ovoid, rather than fan-shaped48.5 Larval mouth internalized49.0 Genital pores appear50.0 Rear sucker primordium raised above surface of larval sac51.0 Rudimentary jaws appear51.5 Rear sucker puckered or bowl-shaped52.0 Slight dorsal fusion at anterior end of germinal plate54.0 Eyes appear, but are unpigmented; rudimentary heart tubes appear55.5 Ridges begin to appear on surface of yolk membrane in the region of the
future gut pouches56.0 Larval nephridia loosening from inside surface of the larval sac58.0 Larval sac peristalsis weak58.5 Gut pouches clearly visible59.5 Heart tubes contain blood and are contractile60.0 Eyes faintly pigmented; larval nephridia are collapsing into the fluid
space under the larval sac membrane60.5 Slight dorsal fusion at posterior end of germinal plate61.0 Posterior gut pouches longer62.5 Larval nephridia gone; larval sac peristalsis ceases63.0 Secondary blood vessels visible65.5 Posterior gut pouches full length68.5 Dorsal fusion complete79.0 Primary body pigment84.0 Dorsal fusion line visible subcutaneously86.0 Secondary body pigment93.5 Dorsal fusion line gone94.0 Tail pigment
100 Dark ventrolateral stripe
1Age is given as a percent of the time between cocoon deposition and the appearance ofthe ventrolateral stripe (% embryonic development, ED), rounded to the nearest 0.5%.Further description of morphological features appears in the text.
HIRUDO STAGING 157

the larval mouth was a large and prominent sphere (Fig.1), whose aperture opened and closed actively. By 45.5%ED (Fig. 1), the larval mouth was no longer active and hadstarted to recede in prominence, apparently because otherhead structures were growing over it. Within about a day,the larval mouth was completely overgrown by the defini-tive mouth.
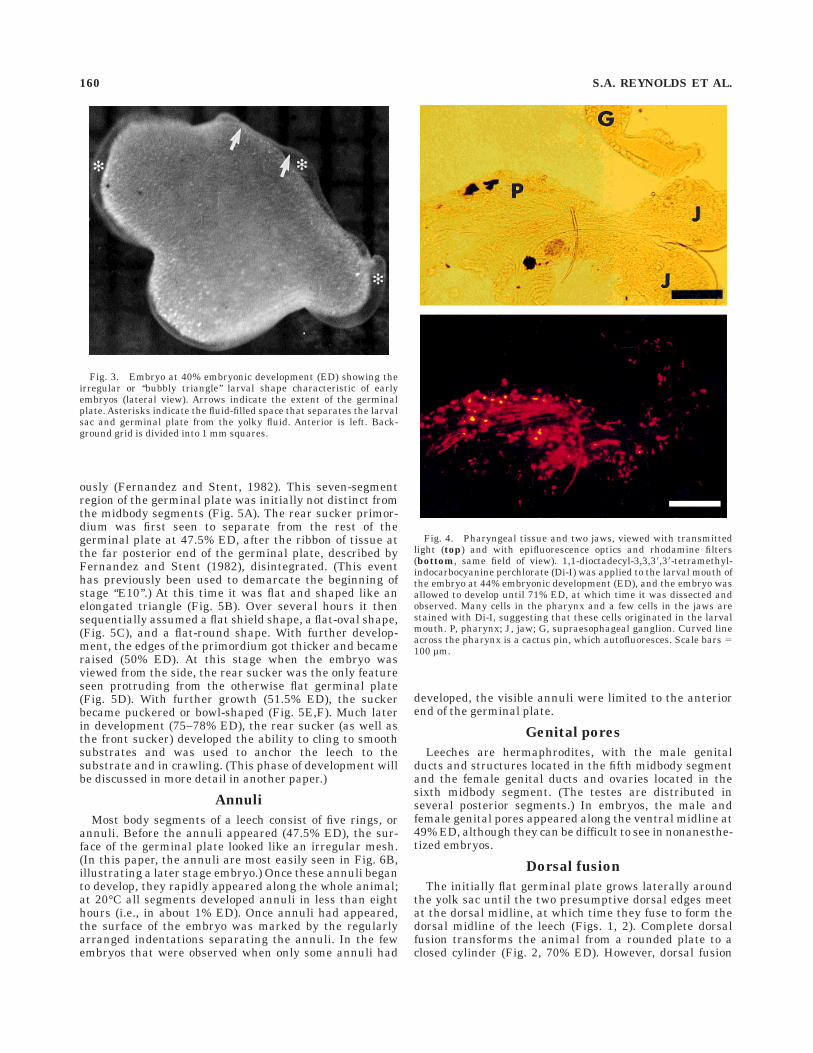
Di-I labeling of different parts of the larval mouthproduced staining in the pharynx, oral epithelium, and
jaws of later embryos (Fig. 4). It was impossible to stainevery cell in the larval mouth of any one animal, but ourresults suggest that after it is overgrown by other struc-tures the larval mouth may become the adult pharynx.(The pharynx is a muscular structure located in theanterior segments; it creates a suction that draws bloodinto the anterior intestinal tract [Sawyer, 1986], much asthe larval mouth draws cocoon fluid into the yolk sac of theearly embryo.)
Fig. 1. Development of Hirudo embryos from 40% embryonicdevelopment (ED) through 65% ED. Solid arrows indicate the presump-tive dorsal edges of the germinal plate; open white arrows indicate thedeveloping rear sucker; 40% ED: curved white arrow indicates theactive larval mouth; 50% ED: small white arrows indicate paired
nephridiopores near the posterior end of the embryo; 40–45%: ventro-lateral view; 50–60%: lateral view; 65%: dorsal view. Age is given as apercentage of ED. Anterior is to the left. Scale bars and units in thebackground grid 5 1 mm.
158 S.A. REYNOLDS ET AL.

Nephridiopores
The surface of the germinal plate was very smooth until46% ED, at which time the nephridiopores appeared(small white arrows in Fig.1, 50% ED). When viewed inoblique lighting they appear bright white in two longitudi-nal rows. These pores connect the interior of the nephridia,the leech’s excretory organs, to the external environment(Sawyer, 1986).
Jaws
Hirudo medicinalis has three jaws, each jaw havingmany small teeth on its rim (Sawyer, 1986). The develop-
ing jaws were first apparent in 51% ED embryos as threenubs that did not yet meet in the center of the mouth; thenas development proceeded the lips grew over them andthey were no longer visible from the outside. Developmentof the jaws may have started slightly earlier, but we wereunable to see them in unanesthetized embryos.
Rear sucker
Because the development of the germinal plate occurs ina rostrocaudal sequence, the seven segments making upthe rear sucker are the last to form, as described previ-
Fig. 2. Development of Hirudo embryos from 70% embryonic development (ED) through 110% ED;75% ED: dorsolateral view; all others: dorsal view. Age is given in percentage of ED. Anterior is to the left.Background grid 5 1 mm.
HIRUDO STAGING 159
ously (Fernandez and Stent, 1982). This seven-segmentregion of the germinal plate was initially not distinct fromthe midbody segments (Fig. 5A). The rear sucker primor-dium was first seen to separate from the rest of thegerminal plate at 47.5% ED, after the ribbon of tissue atthe far posterior end of the germinal plate, described byFernandez and Stent (1982), disintegrated. (This eventhas previously been used to demarcate the beginning ofstage ‘‘E10’’.) At this time it was flat and shaped like anelongated triangle (Fig. 5B). Over several hours it thensequentially assumed a flat shield shape, a flat-oval shape,(Fig. 5C), and a flat-round shape. With further develop-ment, the edges of the primordium got thicker and becameraised (50% ED). At this stage when the embryo wasviewed from the side, the rear sucker was the only featureseen protruding from the otherwise flat germinal plate(Fig. 5D). With further growth (51.5% ED), the suckerbecame puckered or bowl-shaped (Fig. 5E,F). Much laterin development (75–78% ED), the rear sucker (as well asthe front sucker) developed the ability to cling to smoothsubstrates and was used to anchor the leech to thesubstrate and in crawling. (This phase of development willbe discussed in more detail in another paper.)
Annuli
Most body segments of a leech consist of five rings, orannuli. Before the annuli appeared (47.5% ED), the sur-face of the germinal plate looked like an irregular mesh.(In this paper, the annuli are most easily seen in Fig. 6B,illustrating a later stage embryo.) Once these annuli beganto develop, they rapidly appeared along the whole animal;at 20°C all segments developed annuli in less than eighthours (i.e., in about 1% ED). Once annuli had appeared,the surface of the embryo was marked by the regularlyarranged indentations separating the annuli. In the fewembryos that were observed when only some annuli had
developed, the visible annuli were limited to the anteriorend of the germinal plate.
Genital pores
Leeches are hermaphrodites, with the male genitalducts and structures located in the fifth midbody segmentand the female genital ducts and ovaries located in thesixth midbody segment. (The testes are distributed inseveral posterior segments.) In embryos, the male andfemale genital pores appeared along the ventral midline at49% ED, although they can be difficult to see in nonanesthe-tized embryos.
Dorsal fusion
The initially flat germinal plate grows laterally aroundthe yolk sac until the two presumptive dorsal edges meetat the dorsal midline, at which time they fuse to form thedorsal midline of the leech (Figs. 1, 2). Complete dorsalfusion transforms the animal from a rounded plate to aclosed cylinder (Fig. 2, 70% ED). However, dorsal fusion
Fig. 3. Embryo at 40% embryonic development (ED) showing theirregular or ‘‘bubbly triangle’’ larval shape characteristic of earlyembryos (lateral view). Arrows indicate the extent of the germinalplate. Asterisks indicate the fluid-filled space that separates the larvalsac and germinal plate from the yolky fluid. Anterior is left. Back-ground grid is divided into 1 mm squares.
Fig. 4. Pharyngeal tissue and two jaws, viewed with transmittedlight (top) and with epifluorescence optics and rhodamine filters(bottom, same field of view). 1,1-dioctadecyl-3,3,38,38-tetramethyl-indocarbocyanine perchlorate (Di-I) was applied to the larval mouth ofthe embryo at 44% embryonic development (ED), and the embryo wasallowed to develop until 71% ED, at which time it was dissected andobserved. Many cells in the pharynx and a few cells in the jaws arestained with Di-I, suggesting that these cells originated in the larvalmouth. P, pharynx; J, jaw; G, supraesophageal ganglion. Curved lineacross the pharynx is a cactus pin, which autofluoresces. Scale bars 5100 µm.
160 S.A. REYNOLDS ET AL.

occurs in stages. Dorsal fusion began in a small region ofthe head at 52% ED (Fig. 6A). As the germinal plate grewlarger, the extreme tail end of the embryo also fusedanterior to the rear sucker (60.5% ED). At first, the headand tail fusion lines extended slowly. At approximately65.5 % ED, extension accelerated, and approximately 70%
of the dorsal midline fused during an interval equal to onlyabout 3% ED. In this final rapid phase of fusion, the larvalsac tissue located between the anterior and posteriorfusion lines took on a filmy whitish appearance (Fig. 1,65% ED) Dorsal fusion was complete at 68.5% ED, leavinga raised white line along the dorsal midline (Fig. 2, 70%
Fig. 5. Development of rear sucker. A: Rear sucker primordium isindistinct from germinal plate; approximately 44% embryonic develop-ment (ED). B: Rear sucker primordium recently separated fromgerminal plate (arrow) and still triangular in shape; 47.5% ED. C:Rear sucker primordium distinct from germinal plate (arrow) and
somewhat oval in shape; 48.5% ED. D: Rear sucker (arrowhead)rounded and seen from the side to be raised; 50.5% ED. E: Rear suckerbowl-shaped; 52% ED. F: Bowl-shaped rear sucker seen from the side;53% ED. Anterior is to the left. Scale bars and/or units in backgroundgrid 5 1 mm.
HIRUDO STAGING 161

ED). This line disappeared over the next several days (Fig.2) with a somewhat variable time course: it was initiallyvisible on the surface, later visible only subcutaneously(84% ED), and then disappeared altogether (93.5% ED).
Gut pouches
The leech germinal plate and the yolky fluid are sepa-rated from one another by a clear fluid (Fig. 3), and boththe germinal plate (French et al., 1992) and the yolky fluidappear to be bounded by membranes that separate themfrom the fluid. Prior to gut pouch formation, the yolkmembrane underlying the germinal plate was smooth andcontinuous (Fig. 7A). The 10–11 pairs of gut pouches in amature leech first appeared in the anterior two-thirds ofthe embryo as a series of ridges on the surface of the yolkmembrane, particularly when the embryo shortened dur-ing peristalsis (55.5% ED, seen in Fig. 7B). Previousauthors have suggested that the gut pouches are formedwhen growing segmental septa encroach on the yolk sac(Fernandez and Stent, 1982; Sawyer, 1986). We cannotconfirm this hypothesis, because the segmental septa areinvisible in an unstained living embryo. By 58.5% ED, the
gut pouches could be seen even when the embryonic platewas fully lengthened (Fig. 7C). The posteriormost gutpouches then began to grow faster than their more ante-rior neighbors (Fig. 7D). This growth was continuous, sofor the purpose of defining a developmental stage, wenoted when at least one of the two posteriormost gutpouches had reached at least twice the average length ofthe pouches anterior to it (61% ED). These posterior gutpouches continued growing until they reached the end ofthe body cavity. Although dorsal fusion was not completeby this time, a body cavity was formed by local fusion of thepresumptive dorsal edges at the posterior end of theembryo (see ‘‘Dorsal fusion’’ above). The two posterior gutpouches did not always grow at the same rate, and the gutwas scored as full length when at least one of the posterior-most pair of gut pouches reached the end of the body cavity(65.5% ED).
Heart tubes
Rudimentary heart tubes were first visible as thin whitelines located inside the embryo and just lateral to thenephridiopores (54% ED). They ran the full length of thebody, although intersegmental continuity was not alwaysseen in the earliest stages. Red blood appeared in the hearttubes about when they began to contract (59.5% ED). Theblood in the heart tubes makes them easy to recognize atthis point (Fig. 8A). The next readily observable event invascular development was the appearance of secondaryblood vessels (63% ED). The first ones visible were branchesemanating from the heart tube at right angles (Fig. 8B). Asthe heart tube contracted, it forced blood into these smallblood vessels, making them at least temporarily visible.
Eyes
Hirudo medicinalis has five bilateral pairs of eyeslocated on the dorsal margin of the head (Fig. 6B), one pairin each of the first five segments (Kretz et al., 1976). Thefirst eyes detectable were the two largest and most ante-rior pairs (54% ED), then later the three smaller, moreposterior pairs of eyes appeared. At first the eyes wereunpigmented, and only later, a faint pigmentation could beseen, beginning in the anterior eyes (60% ED). The eyeswere initially a faint golden color, then medium brown,and finally very dark brown or black.
Body pigmentation
Embryonic leeches were unpigmented until well afterthe completion of dorsal fusion, and when it first appeared,pigment could be seen only as tiny spots visible when theembryo was examined through a microscope. Pigmenta-tion arose in stages (Fig. 9). The first pigment to appear(the ‘‘primary pigmentation,’’ 79% ED) consisted of twodorsolateral stripes of dark pigment which ran the entirelength of the body, but not onto the tail sucker. The stripeswere usually patchy, rather than continuous. The pigmentwas faint when it first appeared and was most readilydetected by looking at the anterior half of the dorsum whenthe animal shortened. The secondary body pigmentation(86% ED) consisted of an iridescent pigment arranged in acheckerboard pattern around each of the primary stripes.At the same time, a solid stripe of similar iridescentpigment appeared along the lateral edge of the body, whichbecame the yellowish longitudinal stripes at the lateraledge of an adult leech. Next, the tail pigmentation (94%
Fig. 6. Development of the head. A: The expanding edges of thegerminal plate (arrows) have met and fused on the dorsal surface ofthe embryonic head; 52% embryonic development (ED). B: The eyesappear at 54% ED, become faintly pigmented at 60% ED, thengradually become darkly pigmented, as seen in this animal at 78% ED.Anterior is to the left. Units of the background grid 5 1 mm.
162 S.A. REYNOLDS ET AL.

ED) appeared as a pattern of dark stripes on the posteriorsucker, which looked like a continuation of the two pri-mary stripes on the midbody. An additional dark stripealong the dorsal midline of the sucker appeared at thesame time. Finally, the dark ventrolateral stripe (100%ED) appeared just ventral to the yellowish longitudinalstripe along the lateral edge. The ventrolateral stripecould best be seen by inspecting the ventral surface of thebody as the animal crawled along the surface of a clearPetri dish, or by turning the animal over with a probe orforceps as it elongated while crawling. These stripes werefaint and usually patchy or speckled, rather than solid,when they first appeared and darkened as the juvenileleech got older, although in some adults the stripes remainspeckled.
After the ventrolateral stripe appeared, the pigmentover the entire body of the leech gradually darkened, butthe rate at which the pigment became darker was notreadily quantifiable and the time course of this phasevaried among individuals. Because the appearance of theventrolateral stripe was the last quantifiable event inembryogenesis, it was taken as 100% ED.
Effect of temperature on the rateof embryogenesis
To determine how strongly temperature affects the rateof embryogenesis, sibling embryos from single cocoonswere raised at different temperatures (17°C, 20°C, or24°C) for the last 45–55% of embryogenesis. At the time ofseparation, the siblings from each cocoon were at the samedevelopmental stage. Embryos raised in each temperaturedeveloped nearly synchronously with their siblings raisedin the same condition, but embryos raised at 17°C devel-oped substantially slower, and those raised at 24°C devel-oped faster, than those at 20°C . The mean times requiredby embryos from a single cocoon, but held at differenttemperatures, to reach a series of morphological stagingcriteria are graphed in Figure 10.
A regression line was fitted to the data for each group.The dashed line shows the expected rate of development at20°C, based on the data used to establish staging criteria.The 24°C group developed in 79% of the time taken by the20°C group, and the 17°C animals developed in 150% ofthe time taken by the 20°C animals. Morphological fea-
Fig. 7. Development of gut pouches. A: Lateral view of embryo,dorsal side up, at 48% embryonic development (ED). Gut pouches havenot yet begun to form; the surface of the yolk membrane is smooth.B–D: Ventrolateral views, looking through the germinal plate. B: Aseries of small outward puckers in the yolk membrane (arrowheads)indicate the beginning of gut pouch formation (57% ED). C: Thebilaterally paired gut pouches are approximately equally sized alongthe embryo (59% ED). D: The posteriormost pair of gut pouches has
grown to be more than twice as long as the neighboring pair of gutpouches (62% ED). The posteriormost pair of gut pouches will continueto grow until they reach the posterior end of the body cavity (notshown). Open arrows indicate position of the rear sucker. Large solidarrows in C and D indicate the two most posterior pairs of gut pouches.Small white arrows indicate gut pouch typical of each stage. Anterioris to the left. Scale bar and/or background grid 5 1 mm.
HIRUDO STAGING 163

tures appeared in the same order at all three tempera-tures, and development of these features was linear whenplotted against the staging percentages derived at 20°C.The developmental rates for Hirudo embryos raised at17°C, 20°C, and 24°C were 0.48, 0.32, and 0.25 days/percent ED, respectively, and because development waslinear at all three temperatures, we assume it is linear atintermediate temperatures as well. Results for the secondcocoon were similar (data not shown), but data for the twococoons could not be graphed together because the em-bryos were placed into the three different temperatures ata different developmental stage.
The regression lines on Figure 10 cross shortly after theleeches were placed into different temperatures, and thepoint where lines cross indicates the time at which thedifference in temperature started to affect the rate ofdevelopment. The lag time was as long as 15 hours.Although the time of onset for different developmentalevents was linear within each temperature condition, thechange in rate with temperature is not linear over therange of viable temperatures. For example, a 3°C decreasefrom 20°C to 17°C slowed the developmental rate much
more than the 4°C increase from 20°C to 24°C increasedthe rate.
DISCUSSION
This study characterizes the morphological developmentof Hirudo medicinalis starting just after the cessation ofsegment formation, and together with Fernandez andStent (1982) it provides a complete description of embryo-genesis. The current staging scheme is based entirely onobservation of intact living embryos, so all criteria can beused to stage embryos which then can be used in develop-mental or physiological experiments. Because the cuticleof the leech is transparent through most of embryogenesis,many changes in internal organs are externally visible andmay be charted along with the development of externalfeatures. We have found that many traits arise gradually,and the changes occur in an orderly and predictablemanner. These developmental features can be expressedcompactly as a line of developmental progress (Fig. 11). Infuture experiments, physiological or histological investiga-tions can be carried out on individuals that have beenstaged according to external criteria, and the developmen-tal progress of embryos exposed to experimental manipula-tions can be evaluated, allowing information at the cellu-lar and molecular levels to be added to an understanding ofHirudo development.
Staging schemes
Descriptions of the progression of developmental eventsin various animals have generally relied on one of twotypes of schemes: event-staging and time-staging. Theearliest schemes, for chicks (Hamburger and Hamilton,1951) and frogs (Nieuwkoop and Faber, 1967), were basedon event-staging in which major, readily recognizablefeatures were used to define the progression of stages.Event-staging has also been used for invertebrates, suchas the desert locust, Schistocerca gregaria Forskal (Shulovand Pener, 1963) and the leech, Theromyzon rude (Fernan-dez, 1980). The latter has been adapted for other glossipho-niid leeches (Weisblat et al., 1980; Kuwada and Kramer,1983) as well. The alternative scheme, time-staging, hasbeen used more recently for the grasshopper Schistocercanitens (Bentley et al., 1979) and in an earlier description ofdevelopment in Hirudo medicinalis (Fernandez and Stent,1982). Bentley et al. (1979) relied on relative timing,dividing all of embryogenesis into 5% bins, whereas theFernandez and Stent (1982) opted for a strict timingscheme at a fixed temperature (24°C 6 1°C). All of theseschemes are ultimately arbitrary, and each has itsstrengths. We wanted a scheme that would be both easy touse and flexible, and we have chosen to use a scheme basedon the percentage of embryonic time, for the followingreasons:
1. The temporal relationship between developmentalevents (rather than merely the order of events) is immedi-ately apparent from staging designations. In event-staging schemes, the duration of stages varies, so it is moredifficult to remember how long it will take an embryo to gofrom the current stage to five stages later.
2. Establishing nomenclature for smaller subdivisions ofdevelopmental time, or for the timing of new developmen-tal events as they are discovered, will be convenient andconsistent with the existing nomenclature. If a neuron firststarts to grow its axon half-way between stages 45 and 50,
Fig. 8. Heart tube development. A: Heart tube in a 95% embryonicdevelopment (ED) embryo (arrowheads). Blood can be seen through anunpigmented region of cuticle, just ventral to the lateral secondarypigment stripe. B: Higher magnification of a 69% ED embryo revealssecondary blood vessels (arrows) branching from a heart tube (arrow-head). Anterior is to the left. Scale bar and units of the backgroundgrid 5 1 mm.
164 S.A. REYNOLDS ET AL.

for instance, it can conveniently be said to occur at 47.5%ED, not at the less precise and more cumbersome ‘‘mid-stage 12’’ or ‘‘stage 17 (day 2 of 4).’’
3. We use percentage time (% ED), rather than absolutetime, because the rate of development depends on theambient temperature. In a percentage scheme, two em-bryos that look alike have the same designation. In atiming scheme, two embryos that look exactly alike wouldhave different designations if they were raised at differenttemperatures.
4. A percentage of developmental time can be inter-preted by the nonspecialist more readily than can a stagenumber (Bentley et al., 1979).
In their study of the development of desert locusts,Shulov and Pener (1963) gave several reasons for using‘‘arbitrarily chosen stages’’ based on morphological changes,rather than using number of days, including: the definitionof embryonic stages is independent of temperature, someembryonic stages last more or less than a day, andcomparative developmental studies of related species isbest based on morphological stages. These argumentsapply equally well to the use of a percent staging system.In the same study, it was noted that the standard deviationfrom the mean stage number at any chosen time reflectedthe differences in duration of the stages: the standarddeviation from the mean value of a stage was greatest forthe shortest stages, and it was smallest for the longeststages, providing yet another argument in favor of percent-staging. In a later staging table for the grasshopper,Schistocerca nitens, the length of time between eachdescribed stage is an equal 5% of the total time ofembryonic development (Bentley et al., 1979). This sys-tem, however, does not indicate the time at which eachindividual feature or change in appearance arises, nordoes it give the relative order of developmental eventswithin each 5% period. We have addressed this problem bygiving descriptions in increments of 5% of the total develop-mental time (Table 1) as well as noting the timing ofindividual events in Hirudo embryogenesis (Table 2).
In Xenopus staging it has been noted that there wererelatively lengthy periods of development during whichchanges in external features suitable for staging criteriawere lacking, and this problem was solved by combiningand reducing the number of stages (Nieuwkoop and Faber,
Fig. 9. Lateral view of a 110% embryonic development (ED)juvenile Hirudo, showing pigmentation. Arrows, primary pigmentstripe (discontinuous stripe); stars and open arrowhead, secondarypigment around dorsolateral stripe and along lateral margin; solid
arrowhead, dark ventrolateral stripe. Some pigment is visible on therear sucker at the far right of the photo. Anterior is to the left.Background grid 5 1 mm.
Fig. 10. Developmental rates at different temperatures. Siblingswere raised together at 20°C until 46% ED, then were separated andraised to 100% ED at 17°C, 20°C, or 24°C. Arrow indicates time ofseparation. Dashed line indicates the time course of development thatwould have been expected based on staging described in this paper forembryos held at 20°C. The slope of each line gives the developmentalrate at that temperature.
HIRUDO STAGING 165

1967). In Hirudo the same problem exists, but we proposea different solution. The staging system presented herehas the advantage of temporal linearity even in theabsence of observable changes in external features, so nochanging of the numbering system is needed.
In the chick staging system, embryos exhibiting featureswhich fall between two stages are indicated by a stagenumber and a 1 or – sign (Hamburger and Hamilton,1951). This convention could potentially be followed forHirudo using the staging system presented here, but amore useful convention, which this system allows, is forfurther quantitative distinctions in development to receivetheir own numerical designation, indicating the degree ofdevelopmental advancement without changing the al-ready existing staging terminology or numerical stagingdesignations.
Our observations on the temperature dependence ofHirudo embryos were similar to those in the desert locust(Shulov and Pener, 1963), in which it was shown that theduration of every stage of development was very similar inembryos raised at 20°C and 27°C, when stage durationwas expressed as a percentage of the time required for thecompletion of embryonic development at each tempera-ture.
The absolute size of Hirudo embryos proved to beinadequate as a distinguishing morphological feature;individual embryos varied greatly in size, independent ofdevelopmental stage. The size of individuals varies be-tween different cocoons, and sometimes between siblingsfrom the same cocoon. Cocoons vary in size, as do thenumber of embryos per cocoon, and among cocoons of thesame size, a cocoon containing fewer embryos often yieldslarger embryos than does a cocoon containing a greaternumber of embryos.
Correlation with previousleech staging system
A previous study characterized the early stages ofdevelopment of Hirudo medicinalis (Fernandez and Stent,1982). It summarized the previous literature, which gavean overview of very early leech development, then investi-gated more fully the origin of the germinal plate and themechanisms of segmentation. The end of developmentaltime covered by their study overlaps the time at which ourstudy begins. We have compared their descriptions andphotographs with our own observations, allowing us tobring the two studies into register and to relate the twosystems of nomenclature.
The earliest embryos in the Fernandez and Stent (1982)study, up to and including their 6-day-old embryo, appearto be younger than any in this study. Their late 7-day-oldanimal (Fig. 5 in Fernandez and Stent, 1982) correspondsroughly to our 40% ED embryos. The ‘‘9-day-old’’ and ‘‘late9-day-old’’ (Figs. 6, 29, 30 of Fernandez and Stent, 1982)correspond to our 45–46% ED. Their two 11-day-old em-bryos (Figs. 31 and 7, respectively of Fernandez and Stent,1982) correspond to our 48.5% ED and 51% ED. Theslightly different ages in % ED does not preclude themboth from occurring on the same calendar day, as one dayof development at 20°C covers a range of approximately3% ED.
Predicting the timing ofdevelopmental events
The timing between any two events in Hirudo develop-ment can be calculated by the equation: T 5 SR, where:T 5 developmental time interval (days); S 5 developmen-tal stage (% ED); R 5 developmental rate (days/% ED). Todetermine the time between two developmental events (T),
Fig. 11. Time line for development of morphological features in Hirudo embryos. Time is given as apercentage of the total time from egg deposition to the appearance of the ventrolateral stripe (%embryonic development, ED). Morphological features are described more fully in the text. G.P., germinalplate; L., larval; L.S., larval sac; Post., posterior; R.S., rear sucker.
166 S.A. REYNOLDS ET AL.

therefore, one needs to know the two stages (from Tables 1and 2) and the developmental rate, as determined from theslopes of the regression lines in Figure 10.
This rate is temperature dependent: it was 0.48, 0.32,and 0.25 days/percent ED at 17°C, 20°C, and 24°C, respec-tively. Hence, the total developmental time from cocoondeposition to the appearance of the ventral longitudinalstripe under these same conditions would have been 25days at 24°C, 32 days at 20°C, and 48 days at 17°C. Thiscalculation can also be used to manipulate the rate ofdevelopment for experimental purposes. For instance, ittakes 2.4 days (0.48 3 5) to advance from one stage to thenext in Table 1 (i.e., a change of 5% ED) if the embryos arekept at 17°C, but only 1.25 days (0.25 3 5) if they are keptat 24°C, so the dwell time within any 5% stage can beprolonged by roughly a day by changing the temperaturefrom 24°C to 17°C without, as far as we can tell, in any waychanging the sequence of events in the developmentalprogram.
In addition, the results from the temperature-shiftexperiments indicate that it is not necessary to know thetemperature at which embryos have been raised in orderto predict the future rate of development. When embryosare moved from one temperature to another, there is ashort period of adjustment, after which they develop at therate typical of the new temperature. Without additionalstudies, the effect of intermediate temperatures wouldhave to be interpolated, and this interpolation is notlinear: cooling embryos from 20°C has a larger effect ondevelopmental rate than does warming them from 20°C(Fig. 10).
Relationship to other eventsin leech development
Ultimately, the purpose of a scheme for staging embry-onic leeches is to facilitate further studies of embryogen-esis. We are interested in the development of neurons, andas a result, it will be convenient to predict when particularneuronal properties, such as neuronal outgrowth, electro-genesis (Kuwada and Kramer, 1983), and the synthesis ofneurotransmitters (Glover et al., 1987), will first appear.This staging scheme should allow the correlation of inter-nal events that are difficult or impossible to observe witheasily recognizable morphological features. In addition, itwill allow an observer to predict, based on an embryo’sappearance, how long it will take for a particular feature toappear and to modify the developmental rate by heating orcooling the embryos. Further, the scheme will make itpossible to determine if manipulations have changed thetiming of developmental events. We have used this stagingscheme to describe the onset and development of behaviorduring embryogenesis (Reynolds et al., 1998), and it willfacilitate future exploration of the developing neuronalcircuits responsible for these behaviors.
ACKNOWLEDGMENTS
We thank all the members of our laboratory for manylively discussions. This research was supported by an NIHresearch grant (NS25916) to W.B.K.
LITERATURE CITED
Bentley, D., H. Keshishian, M. Shankland, and A. Toroian-Raymond (1979)Quantitative staging of embryonic development of the grasshopper,Schistocerca nitens. J. Embryol. Exp. Morph. 54:47–74.
Drapeau, P., W.B. Kristan, Jr., and M.S. Shankland, eds. (1995) Neurobiol-ogy and development of the leech. J. Neurobiol. 27:267–445.
Fernandez, J. (1980) Embryonic development of the glossiphoniid leechTheromyzon rude: Structure and development of the germ bands. Dev.Biol. 76:245–262.
Fernandez, J. and G.S. Stent (1982) Embryonic development of thehirudinid leech Hirudo medicinalis: Structure, development and seg-mentation of the germinal plate. J. Embryol. Exp. Morphol. 72:71–96.
French, K.A., S.M. Jordan, C.M. Loer, and W.B. Kristan, Jr. (1992)Mesenchyme of embryonic reproductive ducts directs process out-growth of Retzius neurons in the medicinal leech. Dev. Biol. 153:122–140.
Glover, J. and A. Mason (1986) Morphogenesis of an identified leech neuron:Segmental specification of axonal outgrowth. Dev. Biol. 115:256–260.
Glover, J.C., D.K. Stuart, H.T. Cline, R.E. McCaman, C. Magill, and G.S.Stent (1987) Development of neurotransmitter metabolism in embryosof the leech Haementeria ghilianii. J. Neurosci. 7:581–594.
Hamburger, V. and H.L. Hamilton (1951) A series of normal stages in thedevelopment of the chick embryo. J. Morphol. 88:49–92.
Honig, M.G. and R.I. Hume (1986) Fluorescent carbocyanine dyes allowliving neurons of identified origin to be studied in long-term cultures. J.Cell Biol. 103:171–187.
Jellies, J. and W.B. Kristan, Jr. (1991) The oblique muscle organizer inHirudo medicinalis, an identified embryonic cell projecting multipleparallel growth cones in an orderly array. Dev. Biol. 148:334–354.
Kretz, J.R., G.S. Stent, and W.B. Kristan, Jr. (1976) Photosensory inputpathways in the medicinal leech. J. Comp. Physiol. 106:1–37.
Kuwada, J.Y. and A.P. Kramer (1983) Embryonic development of the leechnervous system: Primary axon outgrowth of identified neurons. J.Neurosci. 3:2098–2111.
Loer, C.M., J. Jellies, and W.B. Kristan, Jr. (1987) Morphological changes inleech Retzius neurons after target contact during embryogenesis. J.Neurosci. 7:2618–2629.
Loer, C.M. and W.B. Kristan, Jr. (1989) Peripheral target choice byhomologous neurons during embryogenesis of the medicinal leech. II.Innervation of ectopic reproductive tissue by non-reproductive Retziuscells. J. Neurosci. 9:528–538.
Muller, K.J., J.G. Nicholls, and G.S. Stent (1981) Neurobiology of the Leech.Cold Spring Harbor, NY: Cold Spring Harbor Laboratory.
Nieuwkoop, P.D. and J. Faber, eds. (1967) Normal Table of Xenopus laevis(Daudin). 2nd ed. Amsterdam: North Holland Publishing Co.
Reynolds, S.A., K.A. French, A. Baader, and W. B. Kristan, Jr. (1998)Development of spontaneous and evoked behaviors in the medicinalleech. J. Comp. Neurol. 402:168–180.
Sawyer, R.T. (1986) Leech Biology and Behaviour. Oxford: Oxford SciencePublications, Clarendon Press.
Shulov, A. and M.P. Pener (1963) Studies on the development of eggs of thedesert locust (Schistocerca gregaria Forskal) and its interruption underparticular conditions of humidity. Anti-Locust Bull. 41:1–59.
Weisblat, D.A., G. Harper, G.S. Stent, and R.T. Sawyer (1980) Embryoniccell lineages in the nervous system of the glossiphoniid leech Helobdellatriserialis. Dev. Biol. 76:58–78.
HIRUDO STAGING 167


















