
Specificity of the Binding Interaction between Human Serum ...
7
THE JOURNAL UI BIOI.OUICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 269, No. 13, Issue of April 1, pp. 9760-9766, 1994 Printed in U.S.A. Specificity of the Binding Interaction between Human Serum Amyloid P-component and Immobilized Human C-reactive Protein* (Received for publication, September 14, 1993, and in revised form, December 17, 1993) Robert B. ChristnerS and Richard F. MortensenP From the Department of Microbiology, Ohio State Uniuersity, Columbus, Ohio 43210 C-reactive protein (CRP) and serum amyloid P-com- ponent (SAP) are two members of a group of plasma proteins termed pentraxins, which are composed of five identical noncovalently linked subunits that display Ca2’-dependent binding to a wide variety of substrates. Purified human SAP binds to CRP, only when the latter is immobilized, in a Ca2+-dependent manner under physiological conditions. Externally labeled SAP rap- idly binds to two distinct forms of immobilized CRP (di- rect and phosphorylcholine captured) with a relatively high affinity (K, = 5 MI) at a molar ratioof specifically bound SAP/CRP = 0.3. Studies of binding inhibition us- ing monoclonal antibodies to CRP or synthetic peptides of CRP revealed that residues 134-148 and the COOH- terminal region (residues 191-206) were recognized by SAP. A fragment of CRP consisting of the COOH-termi- nal60 residues within each subunit was also selectively bound by SAP. The ability of immobilized CRP to bind SAP was distinguished fromCRP’s lectin-like binding reactivity since deglycosylated SAP retained its binding reactivity forCRP and sugars that inhibit CRP’s lectin- like binding activity failed to inhibit binding. A peptide from trypsin digested SAP composed of residues 144-199 retained CRP binding activity, implicating the COOH- terminal region of SAP as the CRP recognition site. C-reactive protein (CRP)’ and serum amyloid-P component (SAP) are the two membersof a small group of conserved blood proteins termed pentraxins, proteins that possess a common structural feature of five identical non-covalently linked sub- units arranged in a flat planar disc (1-3). CRP is the arche- typical acute-phase reactant in most vertebrates, and its in- creased synthesis by hepatocytes in response to inflammatory cytokines may result in a severalthousandfoldincreasein blood levels within 24 h after tissue damage or infection, whereas SAP levels increaseby approximately 50% in humans and most other mammals (4). CRP and SAP share extensive * This investigation was supported by United States Public Health Service Grant CA 30015 and by Public Health Senice Grant 2 P30 CA 16058 awarded by the National Cancer Institute, Department of Health and Human Services to the James Cancer Hospital and Research In- stitute of The Ohio State University. The costs of publication of this article were defrayed in part by the payment of page charges. This with 18 U.S.C. Section 1734 solely to indicate this fact. article must therefore be hereby marked “advertisement” in accordance $ Recipient of an Ohio State University Graduate Student Alumni Research Award. 5 To whom correspondence should be addressed: Dept. of Microbiol- ogy, Ohio State University, 484 W. 12thAve.,Columbus, OH 43210. Tel.: 614-292-3360; Fax: 614-292-1538. rylcholine;Ab, antibody(ies1; mAb, monoclonal antibody(ies); KLH, Key- ’ The abbreviations used are: CRP, C-reactive protein; PC, phospho- hole Limpet hemocyanin; B, biotinylated; SAP, serum amyloid P-com- ponent; Pep, peptide; PNG-F, peptide-N-glycosidase-F; PAGE, polyacrylamide gel electrophoresis; HPLC, high performance liquid chromatography; ELISA, enzyme-linked immunosorbent assay. amino acid sequence similarity (-70%) and possess the same gene organization suggesting a common ancestral gene (5-7). CRP has 206 residueshbunit and is not glycosylated (5, 6), whereas SAP has 204 amino acidshbunit with 11% N-linked carbohydrate (7,8). CRP circulates as a single pentraxin, while human SAP is composed of two pentraxins bound face-to-face (9). The two pentraxin proteins share several molecular and bio- chemical properties. They both bind free Ca2+ ions which induce a conformational change that allows binding to a variety of substrates (10-12). CRP and SAP bind to chromatin (13, 14), histones H1 and H2A (151, and fibronectin (16-18). Both CRP and SAP activate the classical complement cascade by inter- acting with the collagen-like stem region of Clq (19-21). The two proteins also have unique binding reactivities. SAP ap- pears to bind to DNAitself (221, whereas CRP binds uniquely to the U1 small nuclear RNP (23). The defining characteristic for CRP from all species is its Caz+-dependent binding to the phos- phorylcholine (PC) moiety (1) on certain polysaccharides and phospholipids (24, 25). By contrast, SAP does not bind to PC, but does bind to agarose (261, zymosan (ll), the glycosamin- oglycans heparin and dermatan sulfate (27), a variety of man- nose-rich glycoproteins (281, and to C4b-binding protein (16). SAP was originally named on the basis of its identity to the P-component that is present in all amyloid deposits (29) regard- less of the composition of the amyloid fibrils (1). SAP binds to both isolated amyloid fibrils (30), as well as to amyloid in uiuo (31). Despite investigations of the molecular and binding prop- erties of these two proteins, a unique biological role for either pentraxin has yet to be established. While investigating functional regions within CRP we ob- served that SAP binds selectively to immobilized, but not soluble CRP, in a Ca2+-dependent manner in buffers of physi- ological ionic strength (32). The purpose of the experiments described here was to characterize this binding interaction for its relative affinity and its specificity in terms of the recognition site on the immobilized CRP and the binding site on SAP for CRP. The interaction between these pentraxins may underlie events in inflammation leading to localization of these pentrax- ins to sites of tissue damage and subsequent repair. EXPERIMENTAL PROCEDURES Purification and Labeling of CRP and SAP-CRP was purified from human ascites fluids by Ca2+-dependent affinity chromatography on PC-phenyl-Sepharose(Pierce Chemical Co.) exactly as described else- where (32,33). Human SAP was purified by affinity chromatographyon agarose beads as described elsewhere (11, 32). Protein concentration was estimated using the reported extinction coefficients at 280 nm of 1.82 for SAP and 1.95 for CRP (11, 32). The concentration of SAP was also determined by rocket immunoelectrophoresis using a monospecific rabbit anti-human SAP antibody (Ab) and a single sample of normal human (male) serum at 40 pg/ml of SAP. Biotin succinimide ester (A” hydroxy-succinimide-Biotin, Pierce Chemical Co.) was added at 250 pg of biotidmg of purified pentraxin protein in 0.1 M sodium borate buffer, pH = 8.8, containing 5 mM EDTA and mixed gently for 4 h. The bioti- nylated (B) proteins were dialyzed against Tris-buffered saline (TBS), 9760
Transcript of Specificity of the Binding Interaction between Human Serum ...

THE JOURNAL UI BIOI.OUICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 13, Issue of April 1, pp. 9760-9766, 1994 Printed in U.S.A.
Specificity of the Binding Interaction between Human Serum Amyloid P-component and Immobilized Human C-reactive Protein*
(Received for publication, September 14, 1993, and in revised form, December 17, 1993)
Robert B. ChristnerS and Richard F. MortensenP From the Department of Microbiology, Ohio State Uniuersity, Columbus, Ohio 43210
C-reactive protein (CRP) and serum amyloid P-com- ponent (SAP) are two members of a group of plasma proteins termed pentraxins, which are composed of five identical noncovalently linked subunits that display Ca2’-dependent binding to a wide variety of substrates. Purified human S A P binds to CRP, only when the latter is immobilized, in a Ca2+-dependent manner under physiological conditions. Externally labeled S A P rap- idly binds to two distinct forms of immobilized CRP (di- rect and phosphorylcholine captured) with a relatively high affinity (K, = 5 MI) at a molar ratio of specifically bound SAP/CRP = 0.3. Studies of binding inhibition us- ing monoclonal antibodies to CRP or synthetic peptides of CRP revealed that residues 134-148 and the COOH- terminal region (residues 191-206) were recognized by SAP. A fragment of CRP consisting of the COOH-termi- nal60 residues within each subunit was also selectively bound by SAP. The ability of immobilized CRP to bind S A P was distinguished from CRP’s lectin-like binding reactivity since deglycosylated SAP retained its binding reactivity for CRP and sugars that inhibit CRP’s lectin- like binding activity failed to inhibit binding. A peptide from trypsin digested SAP composed of residues 144-199 retained CRP binding activity, implicating the COOH- terminal region of SAP as the CRP recognition site.
C-reactive protein (CRP)’ and serum amyloid-P component (SAP) are the two members of a small group of conserved blood proteins termed pentraxins, proteins that possess a common structural feature of five identical non-covalently linked sub- units arranged in a flat planar disc (1-3). CRP is the arche- typical acute-phase reactant in most vertebrates, and its in- creased synthesis by hepatocytes in response to inflammatory cytokines may result in a several thousandfold increase in blood levels within 24 h after tissue damage or infection, whereas SAP levels increase by approximately 50% in humans and most other mammals (4). CRP and SAP share extensive
* This investigation was supported by United States Public Health Service Grant CA 30015 and by Public Health Senice Grant 2 P30 CA 16058 awarded by the National Cancer Institute, Department of Health and Human Services to the James Cancer Hospital and Research In- stitute of The Ohio State University. The costs of publication of this article were defrayed in part by the payment of page charges. This
with 18 U.S.C. Section 1734 solely to indicate this fact. article must therefore be hereby marked “advertisement” in accordance
$ Recipient of an Ohio State University Graduate Student Alumni Research Award.
5 To whom correspondence should be addressed: Dept. of Microbiol- ogy, Ohio State University, 484 W. 12thAve., Columbus, OH 43210. Tel.: 614-292-3360; Fax: 614-292-1538.
rylcholine;Ab, antibody(ies1; mAb, monoclonal antibody(ies); KLH, Key- ’ The abbreviations used are: CRP, C-reactive protein; PC, phospho-
hole Limpet hemocyanin; B, biotinylated; SAP, serum amyloid P-com- ponent; Pep, peptide; PNG-F, peptide-N-glycosidase-F; PAGE, polyacrylamide gel electrophoresis; HPLC, high performance liquid chromatography; ELISA, enzyme-linked immunosorbent assay.
amino acid sequence similarity (-70%) and possess the same gene organization suggesting a common ancestral gene (5-7). CRP has 206 res idueshbuni t and is not glycosylated (5, 6), whereas SAP has 204 amino acidshbuni t wi th 11% N-linked carbohydrate (7,8). CRP circulates as a single pentraxin, while human SAP is composed of two pentraxins bound face-to-face (9).
The two pentraxin proteins share several molecular and bio- chemical properties. They both bind free Ca2+ ions which induce a conformational change that allows binding to a variety of substrates (10-12). CRP and SAP bind to chromatin (13, 14), histones H1 and H2A (151, and fibronectin (16-18). Both CRP and SAP activate the classical complement cascade by inter- acting with the collagen-like stem region of C l q (19-21). The two proteins also have unique binding reactivities. SAP ap- pears to bind to DNAitself (221, whereas CRP binds uniquely to the U1 small nuclear RNP (23). The defining characteristic for CRP from all species is its Caz+-dependent binding to the phos- phorylcholine (PC) moiety (1) on certain polysaccharides and phospholipids (24, 25). By contrast, SAP does not bind to PC, but does bind to agarose (261, zymosan ( l l ) , t he glycosamin- oglycans heparin and dermatan sulfate (27), a variety of man- nose-rich glycoproteins (281, and to C4b-binding protein (16). SAP was originally named on the basis of its identity to the P-component that is present in all amyloid deposits (29) regard- less of the composition of the amyloid fibrils (1). SAP binds to both isolated amyloid fibrils (30), as well as to amyloid in uiuo (31). Despite investigations of the molecular and binding prop- erties of these two proteins, a unique biological role for either pentraxin has yet to be established.
While investigating functional regions within CRP we ob- served that SAP binds selectively to immobilized, but not soluble CRP, in a Ca2+-dependent manner in buffers of physi- ological ionic strength (32). The purpose of the experiments described here was to characterize this binding interaction for its relative affinity and its specificity in terms of the recognition site on the immobilized CRP and the binding site on SAP for CRP. The interaction between these pentraxins may underlie events in inflammation leading to localization of these pentrax- i n s to sites of tissue damage and subsequent repair.
EXPERIMENTAL PROCEDURES Purification and Labeling of CRP and SAP-CRP was purified from
human ascites fluids by Ca2+-dependent affinity chromatography on PC-phenyl-Sepharose (Pierce Chemical Co.) exactly as described else- where (32,33). Human SAP was purified by affinity chromatography on agarose beads as described elsewhere (11, 32). Protein concentration was estimated using the reported extinction coefficients at 280 nm of 1.82 for SAP and 1.95 for CRP (11, 32). The concentration of SAP was also determined by rocket immunoelectrophoresis using a monospecific rabbit anti-human SAP antibody (Ab) and a single sample of normal human (male) serum at 40 pg/ml of SAP. Biotin succinimide ester ( A ” hydroxy-succinimide-Biotin, Pierce Chemical Co.) was added at 250 pg of biotidmg of purified pentraxin protein in 0.1 M sodium borate buffer, pH = 8.8, containing 5 mM EDTA and mixed gently for 4 h. The bioti- nylated (B) proteins were dialyzed against Tris-buffered saline (TBS),
9760

Binding of SAP to CRP 9761
pH 7.6, containing 10 lll~ EDTA to remove excess biotin. The biotinyl- ated SAP (B-SAP) migrated in SDS-PAGE as a single band of 31 kDa, corresponding to the 204-amino-acid subunit with 11% (wiw) carbohy- drate (8). B-CRP migrated as a 25-kDa protein by SDS-PAGE and retained its PC binding activity.
Purified SAP was also labeled externally with "'1 by mixing 0.25 mCi of Na"'1 (Amersham Corp. 1 with 0.5 mg of protein in 0.5 ml of TBS plus 10 mM EDTAin a glass tube coated with Iodogen (Pierce Chemical Co.). The specific activity of the labeled SAP after reisolation by agarose affinity chromatography was 0.3-0.4 pCUpg protein. '"I-Labeled CRP was prepared as described previously and was reisolated by PC affinity chromatography (33).
CRP Synthetic Peptides-The following peptides were synthesized and correspond to the designated residues within the 206-amino-acid subunit of mature human CRP: Pep 134-148 (1-134) ILGQEQDSFGGN- FEG (r148); Pep 191-206 (r191) KYEVQGEVFTKF'QLWP (r206); Pep 47-63 (r47) RGYSIFSYATKRQDNEI (1-63); Pep 1-15 (rl) QTDM- SRKAFVFPKES (r15); Pep 27-38 (r27) TKPLKAFTVCLH (r38); Pep 152-178 (1-152) LVGIGNVNMWDFVLSPDEINTIRL (1-178).
The Pep 47-63 corresponds to the highly conserved PC-binding re- gion of CRP (34) that by itself binds PC (35). The Pep 134-148 corre- sponds to a highly conserved region of both SAP and CRP (34) that contributes to the Ca2+ binding activity of intact CRP (36,37). The Pep 27-38 corresponds to a conserved region of CRP that mediates cell attachment (38). The peptides were synthesized by The Ohio State University Biochemical Instrument Center using t-Boc synthesis on a model 9500 Peptide Synthesizer (Milligefliosearch, Millipore, Burl- ington, M A ) . The peptides were purified by HPLC, lyophilized, and stored desiccated a t -20 "C. The peptides were dissolved in 5% acetic acid or in phosphate-buffered saline a t 5 mg/ml just before use. The peptides were >97% pure based on their HPLC profile. Peptide integrity was confirmed by analysis of amino acid composition.
Antibodies to Human SAP and CRP-Polyclonal rabbit Abs were generated by injecting 250 pg of purified SAP dissolved in complete Freund's adjuvant a t multiple subcutaneous sites into New Zealand White rabbits and challenging 2 months later with 200 pg of SAP intravenously. The antiserum was monospecific for SAP in an immuno- electrophoretic comparison of normal human serum with 40 pg/ml SAP uersus SAP-depleted serum. The anti-Pep 134-148 mAb CaC2 (IgG1) was described previously by us and shown to bind to a surface epitope on CRP (37). Rabbit polyclonal Ab to Pep 27-38 of CRP (CB02) was generated by immunization with the peptide-KLH conjugate in com- plete Freund's adjuvant, followed by intravenous injection of free pep- tide at 3-week intervals until an ELISA titer of >1500 was obtained. The mAb 4B10 (specific for NH,-terminal residues 1-15) and 3H12 (specific for COOH-terminal residues 191-206) were kindly provided by Drs. Joan Siege1 and Henry Gewurz, Rush Medical Center, Chicago, IL (39). I g G fractions of all Ab and mAb were prepared by affinity chromatog- raphy on staphylococcal protein A-Sepharose beads (Pierce Chemical Co.).
Pronase Digestion and Blotting of CRP-Purified CRP a t 1 mg/ml in TBS was exposed to Pronase-Sepharose beads (Pierce Chemical Co.) (20% v/v) for 48 h at 37 "C in the presence of 10 mM EDTA with con- tinuous mixing. The partially digested CRP was analyzed by SDS- PAGE (15% gel) under reducing conditions to reveal 16- and 7-kDa fragments corresponding to residues 1-146 and 147-206, respectively (36). The separated proteins were blotted onto a polyvinylidene difluo- ride membrane for analysis of binding of B-SAP to the CRP fragments.
SAP Deglycosylation-B-SAP was deglycosylated with neuramini- dase as described elsewhere (40) or recombinant peptide-N-glycosi- dase-F (PNGaseF) (New England Biolabs) according to the supplier's specifications with some modifications. Briefly, 0.5 units of neuramini- dase were added to 400 pg of SAP in 0.5 M sodium acetate with 10 mM EDTA, pH 5.0, and incubated overnight a t 37 "C (40). Prior to PNGaseF treatment, 20 pg og SAP was denatured by boiling for 10 min in the presence of 0.5% SDS and 1% 2-mercaptoethanol. SAP was deglycosy- lated with 15,000 units of PNGaseF in 1% Nonidet P-40 overnight at 37 "C. Deglycosylation was confirmed by a mobility shift of the SAP subunit on SDS-PAGE (15% gel) (8).
ELISA Measure of SAP Binding-CRP, SAP, and their ligands were coated overnight a t 4 "C onto Immulon I1 plates (Dynatech, Inc.) using a carbonate-bicarbonate buffer, pH 9.6: CRP, 200 ng/well; SAP, 100 ng/well; PC-KLH, 50 ng/well; and agarose or zymosan a t 1 pg/well. The plates were washed and blocked with 10 mg/ml bovine serum albumin in TBS containing 2 mM Ca" and 0.05% Tween-20. Human CRP (2 pg/ml) and SAP (1 pg/ml) at equimolar concentrations were allowed to bind to their ligands for 1 h at 37 "C. B-SAP in TBS plus 1 mM CaZ+ at 2-fold dilutions over the range of 5 to 1000 ng/well (200 pl/well) was
allowed to bind to immobilized CRP and other ligands for 1 h at 37 "C. The bound B-SAP was detected with horseradish peroxidase-streptavi- din (1:2000 dilution) followed by the 3,3',5,5'-tetramethylbenzidine sub- strate for horseradish peroxidase (Kirkegaard & Perry Laboratories, Gaithersburg, MD) and reading the absorbance a t 450 nm on an ELISA plate reader (Dynatech). Inhibition of SAP binding to CRP was deter- mined by allowing 200 ng of B-SAP to bind to the immobilized CRP in the presence of CRP peptides, monosaccharides, disaccharides, or after reacting the CRP with region-specific Ab.
Western Blotting-Purified SAP, CRP, and Pronase-digested CRP were resolved by SDS-PAGE (15% gel) under reducing conditions and transferred to a polyvinylidene difluoride membrane. The membranes were blocked with 2% nonfat dry milk in TBS with 2 mM Ca2+ for 1 h followed by washing in TBS/Ca"'. The transferred proteins were incu- bated with B-CRP or B-SAP at 5 pg/ml in a TBS/Ca" buffer with 1%' nonfat dry milk overnight. Binding of the biotinylated proteins was detected with streptavidin-horseradish peroxidase and the substrate 3,3'5,5'-tetramethylbenzidine.
Radioimmunoassay of Specific Binding-Human CRP, at 200 ng/well was coated at 4 "C overnight in carbonate-bicarbonate buffer onto Im- mulon I1 Removawells. The amount of the immobilized CRP was deter- mined using trace amounts of '"I-labeled CRP; approximately 40% of the 200 ng of directly immobilized CRP was boundwell (0.64 pmol) and 33%' of the 200 ng of the CRP (0.48 pmol) was bound to the PC-KLH. Wells were blocked and radiolabeled SAP was allowed to bind in 200 pl for 1 h a t 37 "C. After washing, the bound SAP was measured by count- ing the entire well in a Beckman-300s gamma counter. Specific binding of SAP was >90% of the total binding at 0.5 saturation based on the amount of bound '"I-SAP inhibitable by a 100-fold excess of unlabeled SAP.
7kypsin Digestion of B-SAP-Purified B-SAP was dialyzed into wa- ter, lyophilized, and denatured by incubation for 2 h in 8 M urea, 100 mM EDTA, 100 mM NH,HCO,. I.-1-Tosyl-amido-2-phenylethyl chloromethyl ketone treated-trypsin-Sepharose beads were washed in 100 mM NH,HCO, and added a t 100 pl of beadd200 pg of SAP in 0.4 ml and incubated for 18 h at 37 "C. The SAP was recovered by centrifugation, frozen a t -20 "C for sequencing, or used immediately for binding. The digested SAP was analyzed by SDS-PAGE (12.58 gel) run under reduc- ing conditions.
HPLC Analysis of Protease-generated Peptides from SAP-Analysis of the peptides generated from S A P was done by reversed-phase HPLC on a Waters model 600E chromatograph with a model 486 detector. Initial characterization of the peptides was done on a Regis Val-U-Pak HP analytical C18 column using a 40-min gradient of 100% Buffer A (0.1'2 trifluoroacetic acid in water) to 100% Buffer B (100% acetonitrile with 0.1% trifluoroacetic acid). Scaled up purification of the peptides was accomplished on a Bio-Rad Hi-Pore RP-318 semipreparative C18 column. Peptide fractions from the trypsin digest were analyzed by SDS-PAGE (20% gel) and by isoelectric focusing on a Phastsystem (Pharmacia LKB Biotechnology Inc.). The peptide fractions were dia- lyzed into water and tested for the presence of biotin by direct ELISA. The peptide fractions were tested for their ability to bind to immobilized CRP by diluting into TBSlcalciumibovine serum albumidbeen-20 to 2 pg/ml.
Amino Acid Analysis and Peptide Sequencing-Amino acid analysis was performed to determine the molar concentration of the isolated peptides using the Pica-Tag Amino Acid Analysis System (Waters). Pep- tide sequencing was performed by automated Edman degradation using an Applied Biosystems model 470A sequencer, a model 120A phenyl- thiohydantoin analyzer, and a model 900A data module.
RESULTS
Binding of Labeled SAP to Immobilized CRP-To obtain quantitative data on the specific binding interaction between SAP and immobilized CRP, purified human SAP was externally labeled with and its binding to directly coated CRP or to CRP captured on microplate wells via binding to PC as PC- KLH was measured in buffers at physiological ionic strength. The binding approached saturation when 1000 ng (4.3 pmol) of '251-SAP was added to 80 ng (0.6 pmol) of directly immobilized CRP, or a molar ratio of soluble SAF'hmmobilized CRP of ap- proximately 7, based on a molecular mass of 230,000 for SAP and 125,000 for CRP (Fig. lA 1. Scatchard analysis of the bind- ing data yielded a KD = 5 x lo-' M (5 nM) and a molar ratio of specifically bound SAPKRP = 0.28 at saturation (Fig. I A , in-
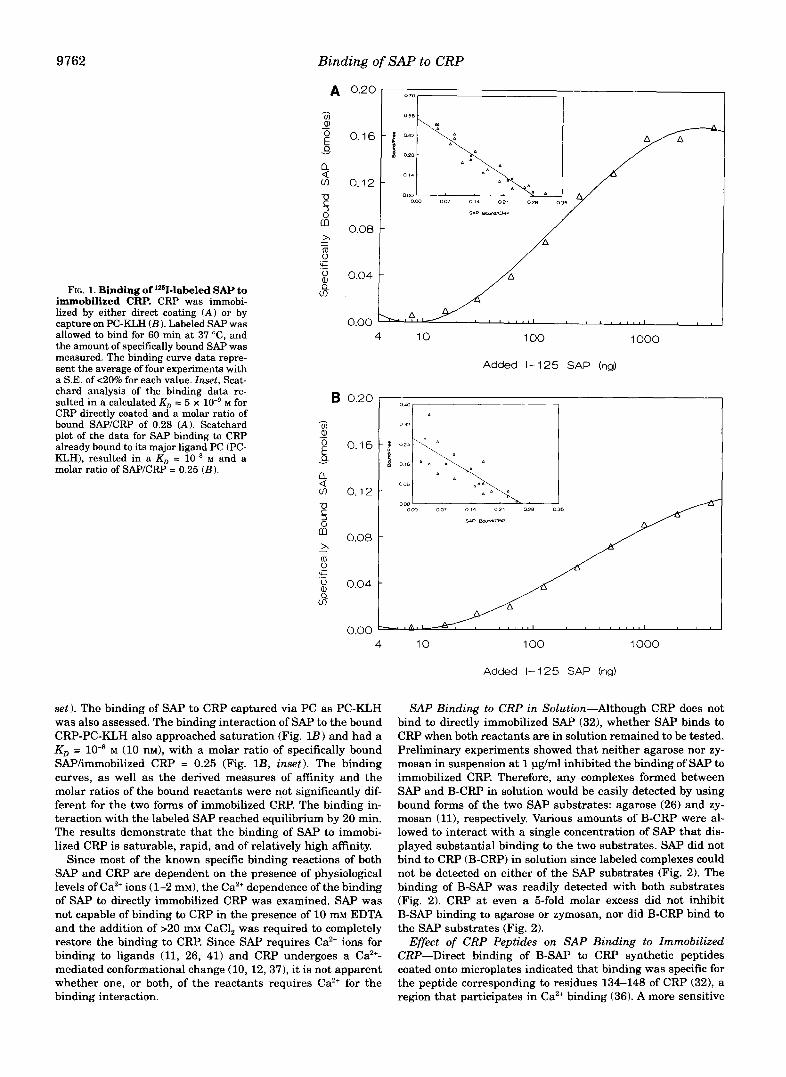
9762 Binding of SAP to CRP
FIG. 1. Binding of laaI-labeled SAP to immobilized CRP. CRP was immobi- lized by either direct coating (A) or by capture on PC-KLH ( B ) . Labeled SAP was allowed to bind for 60 min at 37 "C, and the amount of specifically bound SAP was measured. The binding curve data repre- sent the average of four experiments with a S.E. of <20% for each value. Inset, Scat- chard analysis of the binding data re- sulted in a calculated KD = 5 x M for
bound SAP/CRP of 0.28 (A). Scatchard CRP directly coated and a molar ratio of
plot of the data for SAP binding to CRP already bound to its major ligand PC (PC- KLH), resulted in a KD = lo-@ M and a molar ratio of SAP/CRP = 0.25 ( B ) .
0.16
0.12
f
0.08 -
0.04 -
0.00 - , A s ' I
4 10 100 1000
Added I- 125 SAP (ng)
B 0.20 040 I
I l a I
0.12
0.08
0.04
0. 00 - 4
set). The binding of SAP to CRP captured via PC as PC-KLH was also assessed. The binding interaction of SAP to the bound CRP-PC-KLH also approached saturation (Fig. lB) and had a KO = lo-' M (10 nM), with a molar ratio of specifically bound SAP/immobilized CRP = 0.25 (Fig. l B , inset). The binding curves, as well as the derived measures of affinity and the molar ratios of the bound reactants were not significantly dif- ferent for the two forms of immobilized CRP. The binding in- teraction with the labeled SAP reached equilibrium by 20 min. The results demonstrate that the binding of SAP to immobi- lized CRP is saturable, rapid, and of relatively high affinity.
Since most of the known specific binding reactions of both SAP and CRP are dependent on the presence of physiological levels of Ca2+ ions (1-2 mM), the Ca2+ dependence of the binding of SAP to directly immobilized CRP was examined. SAP was not capable of binding to CRP in the presence of 10 mM EDTA and the addition of >20 mM CaCl, was required to completely restore the binding to CRP. Since SAP requires Ca2+ ions for binding to ligands (11, 26, 41) and CRP undergoes a CaZ+- mediated conformational change (10, 12,37), it is not apparent whether one, or both, of the reactants requires Ca2+ for the binding interaction.
I I
10 100 1000
Added 1-125 SAP (nQ)
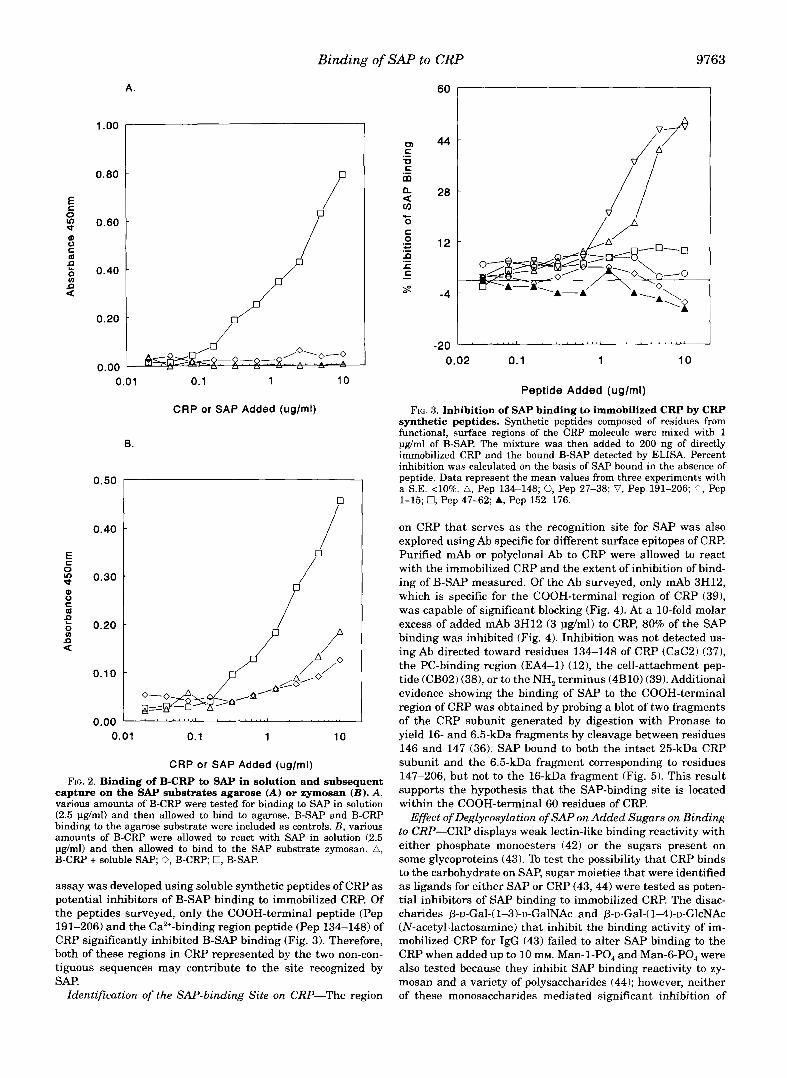
SAP Binding to CRP in Solution-Although CRP does not bind to directly immobilized SAP (32), whether SAP binds to CRP when both reactants are in solution remained to be tested. Preliminary experiments showed that neither agarose nor zy- mosan in suspension a t 1 pg/ml inhibited the binding of SAP to immobilized CRP. Therefore, any complexes formed between SAP and B-CRP in solution would be easily detected by using bound forms of the two SAP substrates: agarose (26) and zy- mosan ( l l ) , respectively. Various amounts of B-CRP were al- lowed to interact with a single concentration of SAP that dis- played substantial binding to the two substrates. SAP did not bind to CRP (B-CRP) in solution since labeled complexes could not be detected on either of the SAP substrates (Fig. 2). The binding of B-SAP was readily detected with both substrates (Fig. 2). CRP at even a 5-fold molar excess did not inhibit B-SAP binding to agarose or zymosan, nor did B-CRP bind to the SAP substrates (Fig. 2).
Effect of CRP Peptides on SAP Binding to Immobilized CRP-Direct binding of B-SAP to CRP synthetic peptides coated onto microplates indicated that binding was specific for the peptide corresponding to residues 134-148 of CRP (32), a region that participates in Ca2+ binding (36). A more sensitive
Binding of SAP to CRP 9763
A.
l.OO ~
0.80
0 E $ 0.60
2 C (0 n ;j 0.40 n a a
0.20
0.00 0.01 0.1 1 10
CRP or SAP Added (uglml)
B.
0.50
0.40
0.30
0.20
/
d /
0.01 0.1 1 10
CRP or SAP Added (uglml) FIG. 2. Binding of B-CRP to SAP in solution and subsequent
capture on the SAP substrates agarose (A) or zymosan ( B ) . A, various amounts of B-CRP were tested for binding to SAP in solution (2.5 pg/ml) and then allowed to bind to agarose. B-SAP and B-CRP binding to the agarose substrate were included as controls. E , various amounts of B-CRP were allowed to react with SAP in solution (2.5 pg/ml) and then allowed to bind to the SAP substrate zymosan. A, B-CRP + soluble SAP; 0, B-CRP; 0, B-SAP.
assay was developed using soluble synthetic peptides of CRP as potential inhibitors of B-SAP binding to immobilized CRP. Of the peptides surveyed, only the COOH-terminal peptide (Pep 191-206) and the Ca2+-binding region peptide (Pep 134-148) of CRP significantly inhibited B-SAP binding (Fig. 3). Therefore, both of these regions in CRP represented by the two non-con- tiguous sequences may contribute to the site recognized by SAP.
Identification of the SAP-binding Site on CRP-The region
-20 ' ' ' ' " " '
0.02 0.1 1 10
Peptide Added (uglml)
FIG. 3. Inhibition of S A P binding to immobilized CRP by CRP synthetic peptides. Synthetic peptides composed of residues from functional, surface regions of the CRP molecule were mixed with 1 pg/ml of B-SAP. The mixture was then added to 200 ng of directly immobilized CRP and the bound B-SAP detected by ELISA. Percent inhibition was calculated on the basis of SAP bound in the absence of peptide. Data represent the mean values from three experiments with a S.E. <lo%. A, Pep 134-148; 0, Pep 27-38; V, Pep 191-206; C, Pep 1-15; 0, Pep 47-62; A, Pep 152-176.
on CRP that serves as the recognition site for SAP was also explored using Ab specific for different surface epitopes of CRP. Purified mAb or polyclonal Ab to CRP were allowed to react with the immobilized CRP and the extent of inhibition of bind- ing of B-SAP measured. Of the Ab surveyed, only mAb 3H12, which is specific for the COOH-terminal region of CRP (39), was capable of significant blocking (Fig. 4). At a 10-fold molar excess of added mAb 3H12 (3 pg/ml) to CRP, 80% of the SAP binding was inhibited (Fig. 4). Inhibition was not detected us- ing Ab directed toward residues 134-148 of CRP (CaC2) (371, the PC-binding region (EA4-1) (121, the cell-attachment pep- tide (CB02) (38), or to the NH, terminus (4B10) (39). Additional evidence showing the binding of SAP to the COOH-terminal region of CRP was obtained by probing a blot of two fragments of the CRP subunit generated by digestion with Pronase to yield 16- and 6.5-kDa fragments by cleavage between residues 146 and 147 (36). SAP bound to both the intact 25-kDa CRP subunit and the 6.5-kDa fragment corresponding to residues 147-206, but not to the 16-kDa fragment (Fig. 5 ) . This result supports the hypothesis that the SAP-binding site is located within the COOH-terminal 60 residues of CRP.
Effect of Deglycosylation of SAP on Added Sugars on Binding to CRP-CRP displays weak lectin-like binding reactivity with either phosphate monoesters (42) or the sugars present on some glycoproteins (43). To test the possibility that CRP binds to the carbohydrate on SAP, sugar moieties that were identified as ligands for either SAP or CRP (43,441 were tested as poten- tial inhibitors of SAP binding to immobilized CRP. The disac- charides P-o-Gal-(1-3)-~-GalNAc and p-D-Gal-(l-4)-D-GlcNAc (N-acetyl-lactosamine) that inhibit the binding activity of im- mobilized CRP for IgG (43) failed to alter SAP binding to the CRP when added up to 10 mM. Man-1-PO, and Man-6-P04 were also tested because they inhibit SAP binding reactivity to zy- mosan and a variety of polysaccharides (44); however, neither of these monosaccharides mediated significant inhibition of

9764 Binding of SAP to CRP
loo I
0.05 0.1 1 10
Antibody Added (uglml)
FIG. 4. Blocking of SAP binding to immobilized CRP by anti- CRPAb. Purified mAb and polyclonal Ab to different surface regions of CRP were allowed to bind to 200 ng of immobilized CRP. The unbound Ab were removed and 100 ng of B-SAP was allowed to bind. Bound SAP was detected by ELISA, and the percentage inhibition of binding was calculated on the basis of SAP bound in the absence of antibody. Data are mean values from two experiments. A, 3H12; 0, EA4-1; 0, CaC2; 0, 4B10; 0, CB02.
25K-1-
1 6 K -
6 . 5 K -
1 2 3 4 FIG. 5. Binding of SAP to fragments of CRP. Purified CRP was
partially digested with Pronase-Sepharose beads to yield 16- and 6.5- kDa fragments by selective cleavage between residues 146 and 147. Undigested CRP (lanes 1 and 3 ) and cleaved CRP (lanes 2 and 4 ) were separated by SDS-PAGE on a 15% gel run under reducing conditions and transferred to polyvinylidene difluoride membranes. The trans- ferred protein was stained with Amido Black (lanes 1 and 2) or probed with 5 pg/ml of B-SAP (lanes 3 and 4 ).
SAP binding to CRP. The results suggest that the sugar moi- eties on SAP and the lectin-binding site of CRP are not involved in the SAP to CRP interaction.
CRP binding to serum glycoproteins was shown to be in- creased by removal of terminal sialic acids and abrogated by deglycosylation of the glycoproteins with PNGaseF (43). The role of the N-linked carbohydrate of SAP in the binding of SAP to CRP was tested by deglycosylating SAP with either neura- minidase or PNGaseF. The two forms of deglycosylated SAP,
1 .oo
0.80
0 E
0.60 m C
$ 0.40 n < in
0.20
0.00 0.01 0.1 1 10
SAP Added (uglml)
FIG. 6. Effect of deglycosylation of SAP on its ability to bind to immobilized CRP. Purified SAP was biotinylated and then treated with either neuraminidase or PNGaseF. The desialylated (N-SAP) and deglycosylated (G-SAP) preparations of SAP were then allowed to bind to directly immobilized CRP (200 ng/well). The binding of the SAP preparations was measured by ELISA. Data are from a representative experiment. 0, SAP; A, N-SAP, 0, G-SAP.
which displayed the predicted increases in mobility on SDS- PAGE (8), bound as efficiently to the immobilized CRP as did the unmodified SAP (Fig. 6). The two deglycosylated prepara- tions of SAP had equivalent amounts of biotin when compared with unmodified SAP as determined by a direct ELISA. There- fore, the carbohydrate moieties present on SAP are not likely to participate in the binding interaction with CRP.
Identification of the CRP-binding Sequence on SAP-To lo- calize the CRP-binding site within SAP, polypeptide fragments from SAP digested with trypsin were evaluated for their bind- ing activity toward immobilized CRP. Trypsin digestion pro- duced four large fragments of 16,20,29, and 4.5 kDa that were resolved by reversed-phase HPLC and recovered as B-polypep- tides. One fragment with a relative molecular mass of 4.5 kDa displayed substantially more binding reactivity for immobi- lized CRP than the 16- and 20-kDa fragments when assayed at 2 pg/ml. This CRP-binding fragment was analyzed by SDS- PAGE and isoelectric focusing and found to be sufficiently ho- mogeneous for sequencing. Amino acid analysis was performed on the isolated 4.5-kDa fragment and then it was sequenced. The NH,-terminal 11 residues were an exact match for residues 144-154 of the derived sequence for SAP (7) (Table I). The entire fragment most likely spans residues 144-199 since Lys- 199 is the next trypsin cleavage and biotinylation site.
DISCUSSION
The experiments described here were done to quantitate and to characterize the specificity of the unidirectional binding in- teraction between the two human pentraxins SAP and CRP. Our initial report of this interaction showed that the CRP must be immobilized and occurred in a Ca2+-dependent manner even in the presence of serum (32). The new findings described herein show that binding of soluble SAP to two distinct forms of immobilized CRP is saturable, of relative high affinity with a KD in the 5-10 nM range of SAP, and specific for residues within the COOH-terminal third of the 206-amino-acid CRP subunit. These properties taken together clearly distinguish the SAP to CRP interaction from the autoaggregation of SAP which is not saturable and readily prevented in the presence of carrier pro- teins, such as the 10 mg/ml of bovine serum albumin present in the binding studies described herein (41). Whether the directly immobilized CRP recognized by SAP is the equivalent of the

Binding of SAP to CRP 9765
TABLE I NH2-terminal sequence of the CRP-binding fragment from SAP
The most active fragment from trypsin-digested SAP was sequenced. Picomole ratio corresponds to the value obtained by dividing the ad- justed picomole for each residue by the S.D. of the background curve for the residue.
Cycle pmol no.
Amino acid ratio identified
1 297 Phe 2 145 3 188
ASP -4%
4 43 5 235
Ser
6 28 Gln
7 Ser
76 8 82
Phe Val
9 24 G ~ Y 10 74 11 66
Glu Ile
conformation represented by neo-CRP is uncertain (45); none- theless, the native CRP conformation “captured” by immobi- lized PC-KLH was still recognized by SAP suggesting that binding is not dependent on any single conformation of CRP. The calculated valence of 0.25-0.3 SAP molecules boundCRP molecule suggests that SAP may bridge or cross-link several CRP molecules to form a matrix. Although the biological sig- nificance of this matrix formation remains to be elucidated, it is possible that the binding event itself may lead to the develop- ment of a nucleating site for the localization of both pentraxins to a single focus, especially during tissue repair (46-48).
The results clearly show that S A P recognizes a site within the COOH-terminal region of each of the CRP subunits on the basis of three different observations. The first is that SAP bind- ing to CRP was selectively inhibited by a mAb (3H12) specific for residues 199-206 in this region of CRP (49). The second is that S A P binds to a 6.5-kDa fragment of CRP composed of residues 147-206 (36). The third is that SAP binding to CRP is inhibited by either one of two synthetic peptides composed of residues 134-148 or 191-206 of CRP, and therefore the site recognized may be composed of non-contiguous amino acids. Since mAb 3H12 recognizes an epitope on CRP that is not altered by the unfolding of CRP it appears that SAP may also recognize a site composed of contiguous amino acids. Recogni- tion of linear epitopes on CRP by SAP seems likely since rela- tively short synthetic peptides inhibit the SAP binding, and denatured CRP subunits, as well as fragments thereof, are recognized by SAP. Since binding of S A P appears to not require an intact CRP molecule and there is considerable evidence that CRP is degraded to biological active peptides by both human neutrophils (49-51) and monocytes (52), SAP has the potential to bind certain CRP peptides generated at inflamed sites with leukocyte infiltrates. It is noteworthy that recovered neutrophil digestion fragments of CRP have been shown to include amino acids 134-148 and 199-206 (49).
Because denatured SAP had the same binding characteris- tics for CRP as the native protein, peptides generated from a trypsin digest of SAP were examined in an attempt to map the CRP-binding site within SAP. SAP digested with trypsin yielded relatively few large fragments that were easily sepa- rated and isolated. However, only one of these retained binding reactivity for immobilized CRP. The NH,-terminal sequence of the reactive peptide corresponded to residues 144-154 of SAP. Although the COOH terminus of this 4.5-kDa fragment was not determined, the next predicted trypsin cleavage site is at Lys- 199. The precise location of the CRP-binding site on the SAP subunits is being pursued with mAb specific for human SAP. The NH, terminus of the SAP fragment with CRP binding activity partially overlaps with residues 134-148 of CRP that
corresponds to at least one of the sites on CRP recognized in the SAP to CRP interaction. Because of the extensive structural and functional similarities between these two pentraxins, this region is one of the two Ca2+-binding sites of CRP and may correspond to a Ca” binding region within SAP. A comparison of the sequence from this conserved region of both pentraxins reveals that SAP has only two non-conservative neutral to charged substitutions at residues 143 and 146 of SAP: CRP, (134)I-L-G-Q-E-Q-D-S-F-G-G-N-F-E-G-S-Q-S-L-V-G-D( 155); SAP, (132)V-*-*-*-*-*-*-*-y-*-K-*-K-*-D-R-*-*-*-F-*-*-E(153),
The specific binding interaction between soluble SAP and CRP in a bound conformation may also occur as a normal event during either tissue repair or development (or both) with SAP eventually serving as a structural component of tissues (48). Although this notion is very speculative, the overall structure of SAP of two flat pentameric discs with 10 identical glycosy- lated subunits is compatible with a process requiring cross- linking of protein ligands.
Acknowledgments-We thank Dr. John Lowbridge and Jane Tolley of the Ohio State University Biochemical Instrument Center for assis- tance with protein sequencing and peptide synthesis and Don Ordaz and Gary Kleman of the Ohio State Microbiology Department Fermen- tation Facility for help with HPLC.
REFERENCES
2. Ballou, S., and Kushner, I. (1992)Adu. Znt. Med. 37, 313-336 1. Pepys, M., and Baltz, M. (1983)Adu. Zmmunol. 34, 141-211
3. Kilpatrick, J. , and Volanakis, J. (1991) Zmmunol. Res. 10, 43-53 4. Pepys, M., Dash, A., Markham, R., Thomas, H., Williams, B., and Petrie, A.
5. Lei, K., Liu, T., Zon, G., Soravia, A,, Liu, T., and Goldman, N. (1985) J. Bid.
6. Woo, P., Korenberg, J., and Whitehead, A. (1985) J. Biol. Chem. 260, 13384-
7. Mantzouranis, E., Dowton, S., Whitehead, A., Edge, M., Bruns, G., and Colten,
8. Hamazaki, H. (1990) Biochem. Biophys. Acta 1037,435438 9. Osmand,A., Friedenson, B., Gewun, H., Painter, R., Hoffman, T., and Shelton,
E. (1977) Proc. Natl. Acad. Sci. U. S. A. 54, 739-743
(1978) Clin. Exp. Zmmunol. 32, 119-124
Chem. 260, 13377-13383
13388
H. (1985) J. Biol. Chem. 260, 7752-7’756
10. Young, N., and Williams, R. (1978) J. Zmmunol. 121, 1893-1898 11. Potempa, L., Kubak, B., and Gewurz, H. (1985) J. Bid. Chem. 260, 12142-
12. Kilpatrick, J., Kearney, J. , and Volanakis, J. (1982) Mol. Zmmunol. 19, 1159-
13. Robey, F., Jones, K., Tanaka, T., and Liu, T. (1984) J. Bid. Chem. 259, 7311-
14. Butler, P., Tennent, G., Pepys, M., Cook, R., and Caspi, D. (1990) J. Exp. Med.
15. DuClos, T., Zlock, L., and Rubin, R. (1988) J. Immunol. 141, 42664270 16. deBeer, F., Baltz, M., Holford, S., Feinstein, A,, and Pepys, M. (1981) J. Exp.
12147
1165
7316
172, 13-18
Med. 154, 1134-1139 17. Salonen, E., Vartio, T., Hedman, K., and Vaheri, A. (1984) J. Bid . Chem. 259,
18. Tseng, J., and Mortensen, R. (1988) Mol. Immunol. 25, 679486
20. Hicks, P., Saunero-Nava, L., DuClos, T., and Mold, C. (1992) J. Zmmunol. 149, 19. Jiang, H., Siegel, J. , and Gewurz, H. (1991) J. Zmmunol. 146, 2324-2330
21. Ying, S., Gewurz, A,, Jiang, H., and Gewurz, H. (1993) J. Zmmunol. 150,
22. Pepys, M., and Butler, P. (1987) Biochem. Biophys. Res. Commun. 148,308-
23. DuClos, T. (1989) J. Zmmunol. 143, 2553-2559 24. Volanakis, J., and Kaplan, M. (1971) Proc. Soc. Exp. Bid. Med. 236, 612-614 25. Volanakis, J., and Narkates, A. (1981) J. Zmmunol. 126, 1820-1825 26. Pepys, M., Dash, A., Munn, E., Feinstein, A., Skinner, M., Cohen, A,, Gewurz,
27. Hamazaki, H. (1987) J. Bid. Chem. 262, 1456-1460 28. Kubak, B., Potempa, L., Anderson, B., Mahklouf, S., Venegas, M., Gewurz, H.,
29. Cathcart, E., Comerford, I . , and Cohen, A. (1965) N. Engl. J. Med. 273, 143-
30. Pepys, M., Dyck, R., deBeer, F., Skinner, M., and Cohen, A. (1979) Clin. Exp.
31. Baltz, M., Dyck, R., and Pepys, M. (1980) Immunology 4 1 , 5 9 4 6 32. Swanson, S., Christner, R., and Mortensen, R. (1992) Biochim. Biophys. Acta
33. Tebo, J., and Mortensen, R. (1990) J. Zmmunol. 144,231-238 34. Liu, T., Syin, C., Nguyen, N., Suzuki, A,, Boykins, R., Lei , K., and Goldman, N.
35. Swanson, S., and Mortensen, R. (1990) Mol. Zmmunol. 27, 679487 36. Kinoshita C. M., Ying, S., Hugli, T., Siegel, J., Potempa, L., Jiang, H.,
Houghten, R., and Gewurz, H. (1989) Biochemistry 28, 9840-9848 37. Swanson, S., Mullenix, M., and Mortensen, R. (1991) J. Zmmunol. 147, 2248-
2252
1496-1501
3689-3694
169-176
313
H., Osmand, A., and Painter, R. (1977) Lancet 1, 1029-1031
and Gewurz, A. (1988) Mol. Zmmunol. 25, 851-858
145
Zmmunol. 38,284-293
1160,309-316
(1985) J. Prot. Chem. 6, 263-2’72

9766 Binding of SAP to CRP 38. Fernandez, M., Mullenix, M., Christner, R., and Modensen, R. (1992) J. Cell.
39. Ying, S., Gewurz, H., Kinoshita, C., Potempa, L., and Siegel, J. (1989) J.
40. Moyle, W., Bahl, O., and Marz, L. (1975) J. Biol. Chem. 250, 9163-9169 41. Baltz, M., deBeer, F., Feinstein, A., and Pepys, M. (1982) Biochem. Biophys.
42. Soelter, J., and Uhlenbruck, G . (1986) Immunology 58, 139-144 43. Kottgen, E., Hell, E., Kage, A., and Tauber, R. (1992) J. Immunol. 149,
Biochem. 60,83-92
Immunol. 143,221-228
Acta 701,229-236
445453
EMBO J. 11, 813-819 44. Loveless, R., Floyd-OSullivan, G., R a p e s , J., Yuen, C., and Feizj, T. (1992)
45. Shields, M. (1993) Immunol. Res. 12,3747 46. Kushner, I., Rakita, L., and Kaplan, M. (1963) J. Clin. Inuest. 42, 286-295 47. Rees, R., Gewurz, H., Siegel, J., Coon, J., and Poternpa, L. (1988) Clin. Immu-
48. Inoue, S. (1991) Cell. Tissue Res. 263, 431438 49. Ying, S., Shephard, E., deBeer, F., Siegel, J., Hams, D., Gewurz, B., Fridkin,
50. Robey, F., Ohura, K., Futaki, S., Fujii, N., Yajimi, H.. Goldrnan, N., Jones, K.
51. Shephard, E.,Anderson, R., Rosen, O., Myer, M., Fridkin, M., Strachan,A., and
52. Teho, J. and Mortensen, R. (1991) Bioehem. Biophys. Acta 1096,210-216
nol. Immunopathol. 48, 95-107
M., and Gewurz, H. (1992) Mol. Immunol. 29,677-687
D., and Wahl, S. (1987) J. B i d . Chem. 262, 7053-7057
deBeer, F. (1990) J . Immunol. 145, 1469-1476


















