
Specificity of a Y-linked gene in the flea beetle Phyllotreta nemorum for defences in Barbarea...
10
Entomologia Experimentalis et Applicata 91: 359–368, 1999. © 1999 Kluwer Academic Publishers. Printed in the Netherlands. 359 Specificity of a Y-linked gene in the flea beetle Phyllotreta nemorum for defences in Barbarea vulgaris Jens Kvist Nielsen Chemistry Department, Royal Veterinary and Agricultural University, Thorvaldsensvej 40, DK-1871 Frederiks- berg C, Denmark Accepted: February 11, 1999 Key words: Barbarea vulgaris ssp. arcuata, Cruciferae, Phyllotreta nemorum, Chrysomelidae, Alticinae, flea beetle, plant defence, host plant range, near-isogenic, Y-linkage, evolution Abstract A Y-linked gene (R-gene) in the flea beetle Phyllotreta nemorum L. (Coleoptera: Chrysomelidae: Alticinae) confer the ability of larvae to survive on types of the plant Barbarea vulgaris R.Br. (Brassicaceae) which are immune to attack by susceptible conspecifics. Two near-isogenic flea beetle lines were developed. The YE-line contained the Y-linked R-gene, and male larvae from this line survived on B. vulgaris. The ST-line did not contain the gene and did not survive on the plant. The YE-line had been developed through 8–9 generations of backcrosses (YE-males with ST-females) and the two lines were considered to be isogenic except for genes located on the Y-chromosome. A single copy of the Y-linked gene is sufficient to transfer a susceptible genotype (ST) into a resistant genotype (YE) which is able to utilize a plant that is immune to attack by specimens without R-genes. The Y-linked gene had no effects on survival on other plant species tested. The gene did not have any effect on developmental times and weights of adult beetles reared on other plants than B. vulgaris. Developmental times of larvae with the Y-linked gene were longer on B. vulgaris than on normal host plants, R. sativus and S. arvensis, but the adults obtained the same size on these plant species. No trade-offs of the Y-linked gene were discovered. The results suggest that the occurrence of the Y-linked gene is a derived trait which has enabled the flea beetle to expand its host plant range. The evolution of a host shift to B. vulgaris seems not to be favoured by the presence of this single gene. Introduction A large number of phytophagous insect species are food specialists with a narrow host plant range (Thompson, 1994). Specialization may make host plant finding more critical, and many insects have developed refined sensory receptors and behavioural responses which enable them to find their particular host plants among other green plants in their habitat (van Loon, 1996). The selection pressures which have promoted the evolution of host plant specialization are poorly known (Bernays & Graham, 1988; Futuyma & Keese, 1992; Thompson, 1994). A narrow host plant range might provide (1) better adaptation to a par- ticular combination of nutrients in the host plant(s), (2) avoidance of many toxic or anti-nutritive com- pounds occurring in non-host plants, (3) escape from enemies and competitors (Schoonhoven et al., 1998). Evolutionary changes in host plant range require intraspecific genetic variation in herbivorous insects for traits influencing behaviour (preference) and per- formance on different plants. Such variation has been demonstrated in a number of cases (reviewed by Fu- tuyma & Keese, 1992; Thompson, 1994; Via, 1990). In a few cases the inheritance has been unravelled, and a polygenic basis has been demonstrated in most cases. Few attempts have been made to investigate the role of single genes influencing preference or performance on different plants, although such studies might provide a better insight in the evolutionary processes. It might be important to know whether the genes are specific for particular plant defences or whether they have more general effects. Studies of effects of single genes re-
-
Upload
jens-kvist-nielsen -
Category
Documents
-
view
214 -
download
2
Transcript of Specificity of a Y-linked gene in the flea beetle Phyllotreta nemorum for defences in Barbarea...

Entomologia Experimentalis et Applicata91: 359–368, 1999.© 1999Kluwer Academic Publishers. Printed in the Netherlands.
359
Specificity of a Y-linked gene in the flea beetlePhyllotreta nemorumfordefences inBarbarea vulgaris
Jens Kvist NielsenChemistry Department, Royal Veterinary and Agricultural University, Thorvaldsensvej 40, DK-1871 Frederiks-berg C, Denmark
Accepted: February 11, 1999
Key words: Barbarea vulgarisssp. arcuata, Cruciferae,Phyllotreta nemorum, Chrysomelidae, Alticinae, fleabeetle, plant defence, host plant range, near-isogenic, Y-linkage, evolution
Abstract
A Y-linked gene (R-gene) in the flea beetlePhyllotreta nemorumL. (Coleoptera: Chrysomelidae: Alticinae) conferthe ability of larvae to survive on types of the plantBarbarea vulgarisR.Br. (Brassicaceae) which are immune toattack by susceptible conspecifics. Two near-isogenic flea beetle lines were developed. The YE-line contained theY-linked R-gene, and male larvae from this line survived onB. vulgaris. The ST-line did not contain the gene anddid not survive on the plant. The YE-line had been developed through 8–9 generations of backcrosses (YE-maleswith ST-females) and the two lines were considered to be isogenic except for genes located on the Y-chromosome.
A single copy of the Y-linked gene is sufficient to transfer a susceptible genotype (ST) into a resistant genotype(YE) which is able to utilize a plant that is immune to attack by specimens without R-genes. The Y-linked gene hadno effects on survival on other plant species tested. The gene did not have any effect on developmental times andweights of adult beetles reared on other plants thanB. vulgaris. Developmental times of larvae with the Y-linkedgene were longer onB. vulgaristhan on normal host plants,R. sativusandS. arvensis, but the adults obtained thesame size on these plant species. No trade-offs of the Y-linked gene were discovered. The results suggest that theoccurrence of the Y-linked gene is a derived trait which has enabled the flea beetle to expand its host plant range.The evolution of a host shift toB. vulgarisseems not to be favoured by the presence of this single gene.
Introduction
A large number of phytophagous insect species arefood specialists with a narrow host plant range(Thompson, 1994). Specialization may make hostplant finding more critical, and many insects havedeveloped refined sensory receptors and behaviouralresponses which enable them to find their particularhost plants among other green plants in their habitat(van Loon, 1996). The selection pressures which havepromoted the evolution of host plant specialization arepoorly known (Bernays & Graham, 1988; Futuyma &Keese, 1992; Thompson, 1994). A narrow host plantrange might provide (1) better adaptation to a par-ticular combination of nutrients in the host plant(s),(2) avoidance of many toxic or anti-nutritive com-
pounds occurring in non-host plants, (3) escape fromenemies and competitors (Schoonhoven et al., 1998).
Evolutionary changes in host plant range requireintraspecific genetic variation in herbivorous insectsfor traits influencing behaviour (preference) and per-formance on different plants. Such variation has beendemonstrated in a number of cases (reviewed by Fu-tuyma & Keese, 1992; Thompson, 1994; Via, 1990).In a few cases the inheritance has been unravelled, anda polygenic basis has been demonstrated in most cases.Few attempts have been made to investigate the role ofsingle genes influencing preference or performance ondifferent plants, although such studies might provide abetter insight in the evolutionary processes. It might beimportant to know whether the genes are specific forparticular plant defences or whether they have moregeneral effects. Studies of effects of single genes re-

360
quire access to near-isogenic lines of insects whichdiffer only in the presence vs. absence of single geneswhich influence responses to different host plants. Apair of such near-isogenic lines has recently been de-veloped in the flea beetle,Phyllotreta nemorumL.(Nielsen, 1996).
Phyllotreta nemorum(Coleoptera: Chrysomelidae:Alticinae) live on a small number of glucosinolatecontaining plant species from the family Brassicaceae(Cruciferae) (Nielsen, 1977, 1989). Its interactionwith the plant species,Barbarea vulgarisR.Br. inDenmark is rather complex, since some plant ‘types’are suitable for all individual beetles, while others aredefended against ‘susceptible’, but not against ‘resis-tant’ beetles (Nielsen, 1996, 1997a,b). The ability ofresistant beetles to utilize defendedB. vulgarisplantsis controlled by major, dominant R-genes. Some ofthese genes seems to be located on sex chromosomes(X and Y) while others are located on autosomes(Nielsen, 1997b). The R-genes control adult behaviour(acceptance) as well as larval survival on defendedB. vulgarisplants (Nielsen, 1996 and unpubl.).
Near-isogenic lines of flea beetles with or withoutR-genes are currently being developed. A suscepti-ble line (ST) without R-genes is unable to surviveon defendedB. vulgaristypes. A resistant line (YE)contain the Y-linked R-gene, and males from this linecan survive on defendedB. vulgaris, while the fe-males cannot. The YE-line has been developed andis being maintained by repeated backcrossing of YE-males with ST-females (8–9 generations). The twolines are therefore considered to be isogenic apart fromgenes which are closely linked to the R-gene on theY-chromosome. The two lines are known to differ insurvivorship on defendedB. vulgaris, but nothing isknown about their performance on other plant species.The present study is the first attempt to compare per-formance of near-isogenic insect lines on a range ofactual and potential host plants. Particular attentionwas paid to the following questions: (1) Is the Y-linkedR-gene specific for defences ofB. vulgarisor does ithave more general effects on survival on other plantsas well? (2) does the gene confer any disadvantages(trade-offs) for the larvae when they are feeding onother actual or potential host plants? and (3) does thegene represent a derived or a primitive trait?
Materials and methods
Flea beetle lines. Two near-isogenic lines have beencompared. Ancestors of the ST-line were collectedon radish (Raphanus sativusL.) in Taastrup (T-population; Nielsen, 1997a,b). A small proportion(10–20%) of beetles from the susceptible T-populationproved to contain some R-genes (de Jong & Nielsen,1999). The ST-line (susceptible line fromTaastrup)was developed from the T-population by selecting onlythose pairs which did not produce progeny that wasable to survive in defended types ofBarbarea vulgarisR. Br. ssp.arcuata (Opiz.) Simkovics. The YE-line(Y-linked R-gene fromEjby) was developed by re-peated backcrossing of males from the E-population(collected originally on the G-type ofB. v. ssp.ar-cuata) with females from the T-population (first threegenerations) or from the ST-line (4th–9th generation).Results from the first three generations of these crossesare given by Nielsen, 1997b).
Plants. Seeds were obtained from different sources:R. sativusL. (radish), Cheiranthus cheiriL. (wall-flower), Iberis amaraL. (candituft), andLunaria an-nuawere purchased from Dæhnfeldt, Odense;Sinapisalba L. cv. Savor andBrassica napusL. var. oleiferacv. Star (oilseed rape) were gifts from Svalöf AB andDansk Planteforædling, Store Heddinge, respectively.Brassica nigraL. (Koch) (black mustard) andThlaspiarvenseL. (pennycress) were gifts from the Dept. ofAgricultural Science, RVAU, Taastrup,Hirschfeldiaincana (L.) Lagrèze-Fossat,Erysimum hieracifoliumL. and Reseda albaL. from the Botanic Garden ofthe University of Copenhagen,Matthiola fruticulosa(L.) Maire ssp.valesiaca(Gay) Ball from the botanicgarden in Champex, Switzerland.Barbarea vulgarisssp. arcuata (accession no. 3; G-type which is de-fended againstP. nemorum; Nielsen, 1997a),Capsellabursa-pastoris(L.) Med., Sinapis arvensisL. (char-lock), Hesperis matronalisL. (dame’s violet),Alliariapetiolata(Bieb.) Cavara & Grande, andReseda luteolaL were collected at natural growth places in Denmark,andErucastrum nasturtiifolium(Poiret) Schultz froma natural growth place in Wallis, Switzerland. Rootsof Armoracia rusticanaG., M. & Sch. were from theBotanic Garden of the Agricultural University. Theplants were grown in a mixture (5:1) of peat (Enhet-sjord - K-jord) and vermiculite (grade 3) in a growthchamber at 20±2 ◦C and a L18:D6 photo cycle. Lightwas supplied by 400 W HPI/T-lamps which supplied160–200µmol quanta m−2 s−1 at the level of the leaf

361
surface. Plants in the vegetative stage, i.e. before budformation, were used in the bioassays. Rosette leavesof Bunias orientalisL. and ofIsatis tinctoriaL. werecollected at natural growth sites in Denmark.
Assays. A three-day bioassay was performed as de-scribed previously (Nielsen, 1997a). Single leaveswere presented to inexperienced, neonate larvae insmall plastic vials (20 or 160 ml depending on the sizeof the leaf; 1 larva per leaf). In a few cases leaf discs(d= 14 mm) were presented in smaller vials (12.5 ml;two discs/vial). After three–four days when survivinglarvae were about to perform their first moult, the sur-vival or death of each larva was recorded as well as thenumber of mines it had made. Leaves remained freshduring this period and were not replaced.
Survival until adult emergence was measured inother experiments using growing plants. The plantswere kept as described above until the experimentswere started and during the experiments at 22± 2 ◦Cand a L18:D6 photo cycle. Light was provided by twowhite Philips and two red (Osram Fluora) 36 W tubeswhich supplied ca. 40µmol quanta m−2 s−1 at thelevel of the leaf surface. Inexperienced, neonate larvaewere released on the upper leaf surface. Larval devel-opment was followed daily, and leaves with nearly fullgrown larvae were collected and transferred to plasticvials (V = 500 ml) containing a mixture of peat andvermiculite where the larvae could pupate. Each daya new vial was used so that the time of pupation wasknown within a 24 h period. Vials with larvae and pu-pae were kept at 22± 2 ◦C until the adults emergedfrom the peat. Within 24 h after emergence and beforeany feeding had occurred, the beetles were collected insmall vials used for sexing and weighing (on a MettlerH20 balance, accuracy: 0.01 mg; weight of vials: ca.3 g).
Statistical analysis. The likelihood ratio test (G-test)was made with the GENMOD procedure in SAS (SASInstitute Inc., 1993) as described by Nielsen (1997a).A significance level of P= 0.01 was used (ns: P>0.01). Analyses of variance (Type 3) and Duncan’sMulitiple Range Tests (significance level: P= 0.05)were performed on untransformed data using the GLMprocedure in SAS. The distribution of beetle freshweights showed no deviation from normality (UNI-VARIATE procedure in SAS) or equality of variances(Bartlett’s Test) (P> 0.05). Pairwise comparisonsof mean larval and pupal developmental times wereperformed with the Mann–Whitney U-test, since the
distribution of these values deviated from normality.A significance level of P= 0.01 was chosen in or-der to avoid type I errors when performing multiplecomparisons.
Results
The most pronounced difference between the twonear-isogenic flea beetle lines was found in survivalrates on the G-type ofBarbarea vulgarisssp.arcuata.No larvae from the ST-line survived on this plant while47.1% (0.691×0.682×100%)of the YE-line surviveduntil the first moult in the three-day bioassay (Ta-ble 1). These results demonstrate that the majority ofYE-males (>90%) survive on the G-type ofBarbareavulgaris ssp.arcuata, since the sex ratio of neonatelarvae is assumed to be close to 50% of each sex (seelater). A single Y-linked gene is therefore sufficientto transfer a susceptible genotype (ST) into a resistantgenotype (YE) and to enable the carrier to utilize aplant that is immune to attack by specimens withoutthe gene.
The Y-linked gene had no or less dramatic effectson survival rates on other plant species (Table 1).A statistical analysis including all 21 plant speciesshowed significant effects of plant species, insect lineas well as a significant interaction between these twovariables. WhenB. v. ssp.arcuatawas removed fromthe dataset, there was no longer a significant differ-ence between survival rates of the two insect linesand no significant interactions between plant speciesand insect line (Table 2). The variation between insectlines can therefore be fully explained by differences inresponses toB. v. ssp.arcuata. There were still signif-icant differences between plant species (Table 2), butthe ranking of plant species was more or less parallelfor the two insect lines (Table 1).
The YE-line had higher mine initiation rates thanthe ST-line onB. v. ssp.arcuataas well as on someof the other plant species (Tables 1 and 2). This maybe a more general effect of the Y-linked gene on larvalbehaviour, since there were significant differences be-tween flea beetle lines even whenB. v. ssp.arcuatawas excluded from the analysis (Table 2). In somecases, however, this possible effect of the gene hadvery little effect on survival, e.g., onReseda albainthe three-day bioassay (Table 1) or onThlaspi arvensein the test using intact plants (Table 3).
The difference between the two flea beetle lineswas apparent also when larvae were reared on grow-

362
Table 1. Suitability of plants for neonate larvae of two near-isogenic flea beetle lines in athree-day bioassay. Males of the YE-line carry a Y-linked R-gene, which confer resistanceto defences inBarbarea vulgaris, while the ST-line does not contain R-genes. N= numberof larvae transferred; I= number of larvae initiating a leaf mine (% of N); S/I= number oflarvae that survived for three days divided by the number that initiated a leaf mine. Species(other thanBarbarea v. ssp.arcuata) are ranked according to over-all survival (S× I) ofthe ST-line
Plant species YE-line ST-line
N I (%) S/I (%) N I (%) S/I (%)
Brassicaceae
Barbarea v. ssp.arcuataa 123 69.1 68.2 85 34.1 0.0
Raphanus sativus 45 93.3 95.2 40 95.0 97.4
Armoracia rusticanab 40 90.0 97.2 40 95.0 97.4
Isatis tinctoriab 40 100.0 97.5 40 95.0 97.4
Erucastrum nasturtiifolium 40 100.0 95.0 30 96.7 93.1
Brassica napusb 40 95.0 73.7 40 100.0 82.5
Sinapis alba 40 100.0 95.0 35 88.6 90.3
Capsella bursa-pastoris 40 77.5 96.8 40 82.5 84.8
Erysimum hieracifolium 50 74.0 89.2 30 66.7 95.0
Brassica nigra 60 90.0 70.4 40 77.5 67.7
Hirschfeldia incana 40 85.0 55.9 25 80.0 35.0
Thlaspi arvense 60 61.7 83.8 50 34.0 82.4
Cheiranthus cheiri 60 28.3 52.9 40 15.0 50.0
Iberis amara 60 93.3 23.2 50 88.0 6.8
Lunaria annua 40 2.5 0.0 42 2.4 (100.0)c
Alliaria petiolata 40 82.5 0.0 40 82.5 0.0
Bunias orientalisb 40 77.5 0.0 40 77.5 0.0
Hesperis matronalis 40 7.5 0.0 30 16.7 0.0
Matthiola fruticulosa 40 10.0 0.0 30 6.7 0.0
Resedaceae
Reseda luteola 60 35.0 85.7 40 30.0 100.0
Reseda alba 40 75.0 0.0 40 52.5 0.0
aG-type which is defended against susceptible flea beetles (Nielsen, 1997a)bLeaf discs were used from wax-covered species,B. napusandI. tinctoria, and leaf pieceswere used fromA. rusticanaandB. orientaliswith very large leavescEstimate of survival onL. annuais misleading since only one larva initiated a leaf mine.
ing plants of the G-type ofBarbarea vulgarisssp.arcuataand survival until adult emergence was mea-sured (Table 3). Only males survived on this plant,and they all came from the YE-line. Analyses of sexratios demonstrated significant differences betweenplant species whenB. v. ssp.arcuatawas included,but not when it was excluded from the analysis. Therewere no significant differences between insect linesand no interaction between plant species and insectline (Table 3). None of the sex ratios obtained on otherplants thanB. v. ssp.arcuatawere significantly differ-ent from 50% (G-test). These results suggest that thesex ratio of neonate larvae is close to 50% of each sex
in both flea beetle lines. This ratio is maintained on themajority of plant species regardless of whether the Y-linked gene is present or absent. However, onB. v. ssp.arcuataall the female larvae die and all the survivorsare males. Under this assumption, the survival of YEmales is approximately the same onB. v. ssp.arcuataand the normal host plants,R. sativusandS. arvensis(Table 3). The results demonstrate a high specificityof the Y-linked gene for defences inB. v. ssp.arcuata.The gene does not confer any adaptation to defencesin A. petiolata, I. amara and T. arvense, which areunsuitable for both lines (Table 3).

363
Table 2. Statistical analysis (G-test) on data on mine initiation and survivalof flea beetle larvae on different plants in the three-day bioassay (Table 1).The analysis was performed initially on the whole data set, and subsequentlyon all plants exceptBarbarea v.ssp.arcuata(G-type) in order to illustrate thecontribution of this plant to the total variation
Explanatory Mine initiation rates Survival rates
variable G df P G df P
Barbarea v.ssp.arcuata(G-type) included (whole data set):
Plant species 846.0 20 <0.0001 810.7 20 <0.0001
Insect line 25.9 1 <0.0001 15.0 1 <0.0001
Interaction 36.1 20 ns 55.3 20 <0.0001
Barbarea v.ssp.arcuata(G-type) excluded:
Plant species 835.1 19 <0.0001 802.8 19 <0.0001
Insect line 10.2 1 <0.01 1.6 1 ns
Interaction 26.6 19 ns 17.0 19 ns
Fresh weights of adultP. nemorumvaried depend-ing on the plant species on which it had been reared,but there were no significant differences between fe-males and males nor between different lines (Table 4).There were no significant differences in weights ofYE-males reared on the G-type ofB. v. ssp.arcuata,R. sativus, S. arvensis, and R. luteola, while malesreared onE. hieracifoliumwere significantly smaller.ST-males reared onR. luteola obtained the highestmean fresh weights although survival rates on thisplant were relatively low (Tables 3 and 4).
The duration of larval development also dependedon plant species, but there were no significant differ-ences between females and males nor between insectlines (Table 5). The fastest development occurred onR. sativusandS. arvensis, while development on theG-type ofB. v. ssp.arcuata, R. luteola, andE. hieraci-foliumwas significantly slower. The duration of pupaldevelopment depended on plant species although dif-ferences were small. Females developed faster thanmales, but there were no differences between the twoinsect lines (Table 6).
Discussion
Specificity of the Y-linked gene.The importance ofa single Y-linked gene for survival ofP. nemorumlarvae onB. v. ssp. arcuata was confirmed. It hadbeen suggested previously in genetic studies usingbeetles collected in the field (Nielsen, 1997b), but inthese studies there were other genes involved, too. In
the present study using near-isogenic lines, it couldbe demonstrated that survival of progeny from ST-females without any R-genes was determined by thekind of males with which they were mated. If a femalewas mated with a YE-male with the Y-linked gene, hermale progeny survived onB. v. ssp.arcuata. However,if she was mated with a male from the ST-line, herprogeny did not survive on this plant, but they didsurvive on the usual host plants,R. sativusandS. ar-vensis. A single gene is therefore sufficient to transfera susceptible genotype (ST) into a resistant genotype(YE) which is able to utilize a plant that is immune toattack by conspecifics without R-genes. The presentresults confirm that the gene is Y-linked since it isalways inherited from fathers to sons (Nielsen, 1997b).
The Y-linked gene did not have such a dramaticeffect on performance ofP. nemorumlarvae on otherplant species. There were significant differences be-tween the two insect lines in mine initiation rates onsome less suitable plant species, e.g.,Thlaspi arvense.However, it remains to be established whether theseeffects are caused by the Y-linked gene. It was notpossible to measure the sex ratio of larvae that initiateda mine inT. arvense, since larvae could not be reareduntil adulthood on this plant (Table 3).
No significant effects of the Y-linked gene on sur-vival on other plants thanB. v. ssp.arcuatacould bedemonstrated in the present study (Tables 1–3). Thegene did not confer any ability to survive onA. peti-olata, B. orientalis, andR. albaalthough these plantsseem to be defended by a mechanism similar to the
364
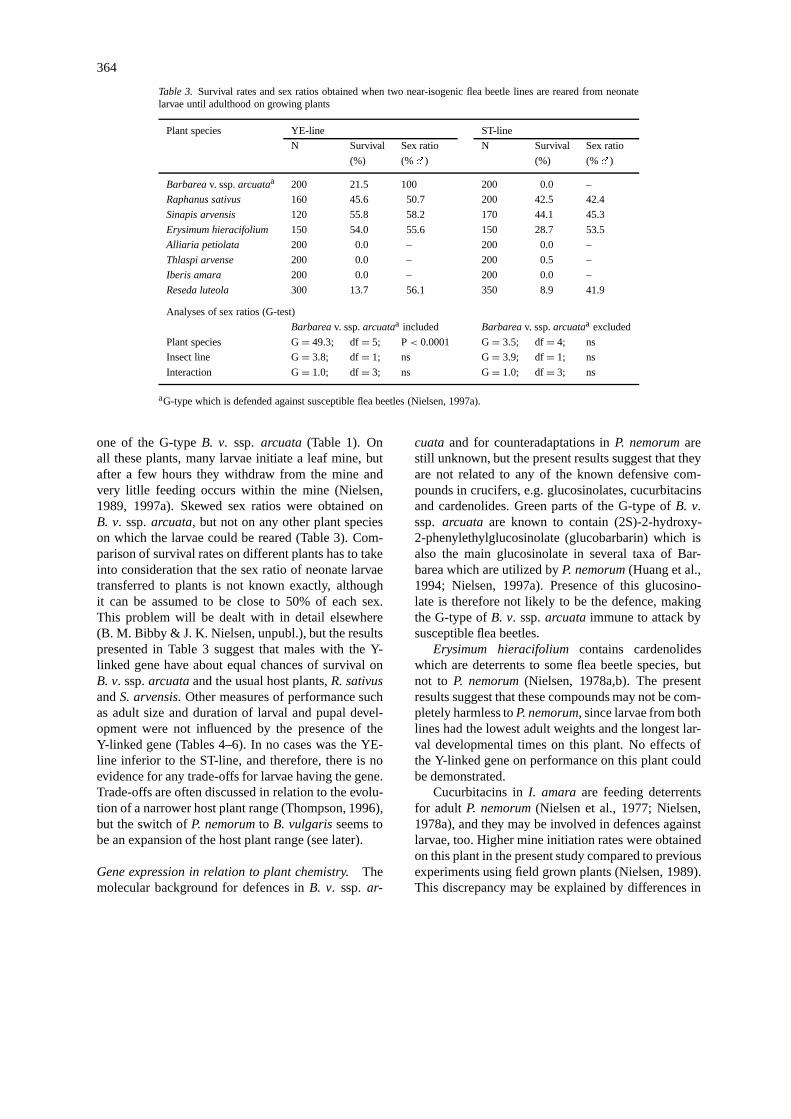
Table 3. Survival rates and sex ratios obtained when two near-isogenic flea beetle lines are reared from neonatelarvae until adulthood on growing plants
Plant species YE-line ST-line
N Survival Sex ratio N Survival Sex ratio
(%) (%�) (%) (%�)
Barbareav. ssp.arcuataa 200 21.5 100 200 0.0 –
Raphanus sativus 160 45.6 50.7 200 42.5 42.4
Sinapis arvensis 120 55.8 58.2 170 44.1 45.3
Erysimum hieracifolium 150 54.0 55.6 150 28.7 53.5
Alliaria petiolata 200 0.0 – 200 0.0 –
Thlaspi arvense 200 0.0 – 200 0.5 –
Iberis amara 200 0.0 – 200 0.0 –
Reseda luteola 300 13.7 56.1 350 8.9 41.9
Analyses of sex ratios (G-test)
Barbareav. ssp.arcuataa included Barbareav. ssp.arcuataa excluded
Plant species G= 49.3; df= 5; P< 0.0001 G= 3.5; df= 4; ns
Insect line G= 3.8; df= 1; ns G= 3.9; df= 1; ns
Interaction G= 1.0; df= 3; ns G= 1.0; df= 3; ns
aG-type which is defended against susceptible flea beetles (Nielsen, 1997a).
one of the G-typeB. v. ssp. arcuata (Table 1). Onall these plants, many larvae initiate a leaf mine, butafter a few hours they withdraw from the mine andvery litlle feeding occurs within the mine (Nielsen,1989, 1997a). Skewed sex ratios were obtained onB. v. ssp.arcuata, but not on any other plant specieson which the larvae could be reared (Table 3). Com-parison of survival rates on different plants has to takeinto consideration that the sex ratio of neonate larvaetransferred to plants is not known exactly, althoughit can be assumed to be close to 50% of each sex.This problem will be dealt with in detail elsewhere(B. M. Bibby & J. K. Nielsen, unpubl.), but the resultspresented in Table 3 suggest that males with the Y-linked gene have about equal chances of survival onB. v. ssp.arcuataand the usual host plants,R. sativusandS. arvensis. Other measures of performance suchas adult size and duration of larval and pupal devel-opment were not influenced by the presence of theY-linked gene (Tables 4–6). In no cases was the YE-line inferior to the ST-line, and therefore, there is noevidence for any trade-offs for larvae having the gene.Trade-offs are often discussed in relation to the evolu-tion of a narrower host plant range (Thompson, 1996),but the switch ofP. nemorumto B. vulgarisseems tobe an expansion of the host plant range (see later).
Gene expression in relation to plant chemistry.Themolecular background for defences inB. v. ssp.ar-
cuata and for counteradaptations inP. nemorumarestill unknown, but the present results suggest that theyare not related to any of the known defensive com-pounds in crucifers, e.g. glucosinolates, cucurbitacinsand cardenolides. Green parts of the G-type ofB. v.ssp. arcuata are known to contain (2S)-2-hydroxy-2-phenylethylglucosinolate (glucobarbarin) which isalso the main glucosinolate in several taxa of Bar-barea which are utilized byP. nemorum(Huang et al.,1994; Nielsen, 1997a). Presence of this glucosino-late is therefore not likely to be the defence, makingthe G-type ofB. v. ssp.arcuataimmune to attack bysusceptible flea beetles.
Erysimum hieracifolium contains cardenolideswhich are deterrents to some flea beetle species, butnot to P. nemorum(Nielsen, 1978a,b). The presentresults suggest that these compounds may not be com-pletely harmless toP. nemorum, since larvae from bothlines had the lowest adult weights and the longest lar-val developmental times on this plant. No effects ofthe Y-linked gene on performance on this plant couldbe demonstrated.
Cucurbitacins inI. amara are feeding deterrentsfor adult P. nemorum(Nielsen et al., 1977; Nielsen,1978a), and they may be involved in defences againstlarvae, too. Higher mine initiation rates were obtainedon this plant in the present study compared to previousexperiments using field grown plants (Nielsen, 1989).This discrepancy may be explained by differences in

365
Table 4. Fresh weights (mg) of newly emerged adultPhyllotreta nemorumreared on fivedifferent plant species
Plant species Insect Females Males
line Mean± s.e. N Mean± s.e. N
Barbarea v.ssp.arcuata YE – 0 1.71± 0.035b 31
(G-type) ST – 0 – 0
Raphanus sativus YE 1.68± 0.036abc 35 1.66± 0.029b 34
ST 1.62± 0.034bcd 49 1.69± 0.036b 36
Sinapis arvensis YE 1.75± 0.044ab 28 1.63± 0.033bc 38
ST 1.66± 0.032abc 41 1.64± 0.042bc 32
Erysimum hieracifolium YE 1.55± 0.034cd 29 1.54± 0.025cd 33
ST 1.50± 0.056d 17 1.50± 0.024d 19
Reseda luteola YE 1.68± 0.046abc 18 1.67± 0.043b 21
ST 1.78± 0.051a 18 1.87± 0.049a 13
Results from overall analysis of variance: Plant species: F= 13.1; df= 4; P< 0.0001; Insectline: F= 0.1; df= 1; ns; Sex: F= 0.2; df= 1; ns.Numbers with same letter within each column are not significantly different (Duncan’sMultiple Range Test; P> 0.05).
the content of cucurbitacins or in other secondarycompounds, since the content of these compoundsmay well be influenced by environmental factors (Wa-terman & Mole, 1989). The failure of both lines todevelop until adulthood on this plant demonstrate thatthe Y-linked gene does not confer any adaptation todefences inI. amara.
Gene expression and evolution of host plant range.The phylogeny of host plant utilization ofP. nemo-rum is not known. However, the knowledge of thepresent host plant range suggests that for a consider-able period of time, the species has been oligophagous,feeding on the crucifers that it can attack. The contem-porary host plants are not confined to any particulargrouping of the crucifers (Hedge, 1976). The tribeBrassiceae is well represented (Raphanus sativus(cul-tivated radish),R. raphanistrum(wild radish),Sinapisarvensis(charlock) andErucastrum nasturtiifolium),but species from Hesperideae (Erysimumspp.), Le-pidieae (Cardaria draba) and Arabideae (Barbareaspp.) are important host plants, too. All these tribesalso contain species which are not or rarely attacked byP. nemorum(Nielsen, 1977, 1989). With the presentknowledge of host plant selection of the species, thebest definition of the host range seems to be: anyglucosinolate containing plant that does not containparticular defences (Nielsen, 1978b, 1989).
A number of different defensive mechanismsagainstP. nemorumand other insects have been identi-fied in the crucifers. Some of these defences are clearly
derived e.g. cucurbitacins inIberis species and carde-nolides inErysimumandCheiranthusspecies and theiremergence may have restricted the host plant rangeof several insects, includingP. nemorum(Nielsen,1978a,b; Sachdev-Gupta et al., 1993a,b). The uti-lization of plant species belonging to several tribesof Cruciferae suggests that host range expansion hasalso been possible although it has been limited toplants containing glucosinolates. The interaction be-tweenP. nemorumand glucosinolate containing plantstherefore seems to be dynamic and expansion as wellas narrowing of the host plant range is possible.
The defensive mechanism of the G-type ofBar-berea vulgarisssp. arcuata is not known, but itsrestricted occurrence suggest that it is a derived char-acter. It is present in the subspeciesB. v.ssp.vulgarisand the G-type ofB. v.spp.arcuata, but absent in theP-type ofB. v.spp.arcuataas well as inB. intermediaandB. stricta (Nielsen, 1997a). Similar mechanismshave been found in other crucifers, e.g.,Alliaria peti-olata, Buniasspecies,Matthiola species andResedaalba (Table 1 and Nielsen, 1989), but the defences ofthese plants seem not to be identical to those ofB. vul-garis since the Y-linked R-gene does not confer anycounteradaptations to them as it does to defences inB. vulgaris.
If the defences inB. vulgarisare derived, the Y-linked gene is most likely a derived trait, too, sinceit is very specific for defences in this plant. If it hadbeen a primitive (plesiomorphic) trait, it is unlikelythat it would be so specific for a single plant species

366
Table 5. Duration (days) of larval feeding period ofPhyllotreta nemorumreared ondifferent plant species
Plant species Insect Females Males
line Mean± s.e. N Mean± s.e. N
Barbarea v. ssp.arcuata YE – 0 13.3± 0.17b 41
(G-type) ST – 0 – 0
Raphanus sativus YE 11.0± 0.19cd 36 11.2± 0.19de 37
ST 10.7± 0.17d 49 10.8± 0.17e 36
Sinapis arvensis YE 11.3± 0.12c 28 11.3± 0.11d 39
ST 11.3± 0.10c 41 11.5± 0.18d 34
Erysimum hieracifolium YE 14.0± 0.24a 36 14.3± 0.26a 45
ST 14.1± 0.42ab 20 13.9± 0.37ab 23
Reseda luteola YE 12.7± 0.39b 18 12.6± 0.25c 21
ST 12.9± 0.16b 18 13.0± 0.28bc 13
Results from overall analysis of variance: Plant species: F= 142.1; df = 4; P< 0.0001;Insect line: F= 0.5; df= 1; ns; Sex: F= 0.8; df= 1; ns.Numbers with the same letter within each column are not significantly different (pairwisecomparisons with the Mann–Whitney U-test; P> 0.01).
and even for certain types of this species (Nielsen,1997a). This conclusion is exaggerated by the failureto detect any disadvantages (trade-offs) of the geneand to unravel any selection pressures that could havereduced the frequency of the gene in flea beetle popu-lations. The gene therefore seems to represent a novelopportunity forP. nemorumto expand its host plantrange by incorporating certain types and subspeciesof B. vulgaris. This opportunity may have importantecological implications since the new host plants arecommon in many parts of Europe (Hegi, 1986).
The Y-linked gene may have originated as a ran-dom mutation on the Y-chromosome or on a sectionof another chromosome which has later been translo-cated to the Y-chromosome. The fitness advantagesof having the gene on the Y-chromosome rather thanon an autosome or X-chromosome is not at all clear.Since the gene is only found in males, it cannot conferhost range expansion of an entire flea beetle popula-tion. The fitness advantages of having the Y-linkedgene is therefore likely to depend on whether auto-somal and/or X-linked resistance genes are present infemales of the same population. This assumption issupported by the rarity of the Y-linked gene in Danishflea beetle populations, where indeed it seems to be re-stricted to populations where autosomal and X-linkedresistance genes are abundant, too (de Jong & Nielsen,1999).
Host range expansion could be the first step inthe evolution of specificity for the novel host plant(Thompson, 1996). Since evolution of host specificity
has occurred many times in phytophagous insects, thefactors favouring it have been discussed extensively(Bernays & Graham, 1988; Futuyma & Keese, 1992;Schoonhoven et al., 1998; Thompson, 1994; Via,1990). Central for this discussion is the idea that thereare negative genetic correlations between performanceon host and non-host plants, but such correlations haverarely been found. It has therefore been hypothesizedthat trade-offs favouring selection for specificity arenot inherent to single genes but to coadaptation be-tween several genes (Thompson, 1994, 1996). Thepresent results demonstrate that the Y-linked gene doesnot cause any trade-offs on any plant species, andtherefore it is unlikely that the presence of this genewould lead to specialization on defended types ofB. vulgaris. Faster larval development on the old hostplants, radish and charlock, may also counteract theevolution of specificity forB. vulgaris. Speed of larvaldevelopment is probably a measure of nutritive valueand/or content of toxic or antinutritional compounds,and the results suggest that the new host plant is stillless suitable than the old host plants. Longer develop-mental times mean longer exposures to parasitoids andpredators as well as adverse weather conditions, andmay be a significant disadvantage in the field (Bernays& Graham, 1988). This disadvantage may then becounterbalanced by lower intraspecific competition onthe new host plant, but the outcome of these opposingfactors need further studies under field conditions.
The occurrence of the Y-linked R-gene only inmales may present another constraint on its involve-

367
Table 6. Duration (days) of pupal development ofPhyllotreta nemorumreared on differentplant species
Plant species Insect Females Males
line Mean± s.e. N Mean± s.e. N
Barbarea v. ssp.arcuata YE – 0 14.0± 0.14ab 41
(G-type) ST – 0 – 0
Raphanus sativus YE 13.3± 0.10a 36 13.6± 0.11bcd 37
ST 13.2± 0.09ab 49 13.4± 0.10cd 36
Sinapis arvensis YE 13.1± 0.12ab 28 13.5± 0.12bcd 39
ST 12.9± 0.08b 41 13.3± 0.11d 34
Erysimum hieracifolium YE 13.5± 0.11a 36 13.8± 0.10b 45
ST 12.9± 0.21ab 20 13.7± 0.15bc 23
Reseda luteola YE 13.1± 0.21ab 18 13.5± 0.27bcd 21
ST 13.6± 0.15a 18 14.3± 0.18a 13
Results from overall analysis of variance: Plant species: F= 5.7; df= 4; P< 0.001; Insectline: F= 1.9; df= 1; ns; Sex: F= 32.1; df= 1; P< 0.0001.Numbers with same letter within each column are not significantly different (pairwisecomparisons with the Mann–Whitney U-test; P> 0.01).
ment in the evolution of specialization onB. vulgaris,since it might reduce their chances to find mates ifthey tended to aggregate on a plant which the majorityof females could not feed on. It has been shown pre-viously that beetles with the Y-linked R-gene acceptB. v.spp.arcuatawhile beetles without R-genes do not(Nielsen, 1996 and unpubl.). The chances for maleswith the Y-linked R-gene to find a mate and repro-duce are therefore assumed to be higher if they adopta wider host plant range. It seems to be more likelythat genes located on autosomes and X-chromosomesmight be involved in evolution of host plant special-ization since they can occur in both sexes. Flea beetlelines with autosomal or X-linked R-genes are currentlybeing developed, and comparison of a larger selec-tion of near-isogenic lines may yield further insightin (1) the origin of the R-genes, (2) the mechanismof gene expression, and (3) the implications of thelocation of R-genes on different chromosomes for theevolution of host plant range.
Acknowledgements
I thank Bo M. Bibby and Peter W. de Jong for crit-ical comments. I am grateful for seed supplies fromthe Dept. of Agricultural Science, RVAU, Taastrup,The Botanic Garden of the University of Copen-hagen, The Botanic Garden in Champex, Switzerland,Dansk Planteforædling, and Svalöf AB. The work wassupported by the Danish Agricultural and VeterinaryResearch Council.
References
Bernays, E.A. & M. Graham, 1988. On the evolution of hostspecificity in phytophagous arthropods. Ecology 69: 886–892.
Futuyma, D. J. & M. C. Keese, 1992. Evolution and coevolutionof plants and phytophagous arthropods. In: G. A. Rosenthal &M. R. Berenbaum (eds), Herbivores: Their Interaction with Sec-ondary Plant Metabolites, vol. II. Ecological and EvolutionaryProcesses. Academic Press, San Diego, pp. 439–475.
Hedge, I. E., 1976. A systematic and geographical survey of the oldworld Cruciferae. In: J. G. Vaughan, A. J. MacLeod & B. M. G.Jones (eds.), The Biology and Chemistry of the Cruciferae,Academic Press, London, pp. 1–45.
Hegi, G., 1986. Illustrierte Flora von Mitteleuropa, Band IV, Teil 1,Verlag Paul Parey, Berlin.
Huang, X., J. A. A. Renwick & K. Sachdev-Gupta, 1994. Ovipo-sition stimulants inBarbarea vulgaris for Pieris rapae andP. napi oleracea: Isolation, identification and differential activity.Journal of Chemical Ecology 20: 423–438.
Jong, P. W. de & J. K. Nielsen, 1999. Polymorphism in a flea beetlefor the ability to utilize an atypical host plant. Proceedings of theRoyal Society of London B266: 103–111.
Loon, J. J. A. van, 1996. Chemosensory basis of feeding and ovipo-sition behaviour in herbivorous insects: a glance at the periphery.Entomologia Experimentalis et Applicata 80: 7–13.
Nielsen, J. K., 1977. Host plant relationships ofPhyllotreta nemo-rum L. (Coleoptera: Chrysomelidae). I. Field studies. Zeitschriftfür Angewandte Entomologie 84: 396–407.
Nielsen, J. K., 1978a. Host plant discrimination within Cruciferae:Feeding responses of four leaf beetles (Coleoptera: Chrysomel-idae) to glucosinolates, cucurbitacins and cardenolides. Ento-mologia Experimentalis et Applicata 24: 41–54.
Nielsen, J. K., 1978b. Host plant selection in monophagous andoligophagous flea beetles feeding on crucifers. EntomologiaExperimentalis et Applicata 24: 562–569.
Nielsen, J. K., 1989. Host plant relations ofPhyllotreta nemorumL.(Coleoptera: Chrysomelidae). II. Various defensive mechanismsin plants and their role in defining the host plant range. Journalof Applied Entomology 107: 193–202.

368
Nielsen, J. K., 1996. Intraspecific variation in adult flea beetlebehaviour and larval performance on an atypical host plant.Entomologia Experimentalis et Applicata 80: 160–162.
Nielsen, J. K., 1997a. Variation in defences of the plantBarbareavulgarisand in counteradaptations by the flea beetlePhyllotretanemorum. Entomologia Experimentalis et Applicata 82: 25–35.
Nielsen, J. K., 1997b. Genetics of the ability ofPhyllotreta nemorumlarvae to survive in an atypical host plant,Barbarea vulgarisssp.arcuata. Entomologia Experimentalis et Applicata 82: 37–44.
Nielsen, J. K., L. M. Larsen & H. Sørensen, 1977. CucurbitacinE and I in Iberis amara: Feeding inhibitors forPhyllotretanemorum. Phytochemistry 16: 1519–1522.
Sachdev-Gupta, K., C. D. Radke & J. A. A. Renwick, 1993a.Antifeedant activity of cucurbitacins fromIberis amaraagainstlarvae ofPieris rapae. Phytochemistry 33: 1385–1388.
Sachdev-Gupta, K., C. D. Radke, J. A. A. Renwick & M. B.Dimock, 1993b. Cardenolides fromErysimum cheiranthoides:Feeding deterrents toPieris rapaelarvae. Journal of ChemicalEcology 19: 1355–1369.
SAS Institute Inc., 1993. SASr Technical Report P-243,SAS/STATr Software: The GENMOD Procedure, Release6.09, Cary.
Schoonhoven, L. M., T. Jermy & J. J. A. van Loon, 1998. Insect –Plant Biology. From Physiology to Evolution. Chapman & Hall,London.
Thompson, J. N., 1994. The Coevolutionary Process. The Universityof Chicago Press, Chicago.
Thompson, J. N., 1996. Trade-offs in larval performance on normaland novel hosts. Entomologia Experimentalis et Applicata 80:133–139.
Via, S., 1990. Ecological genetics and host adaptation in herbiv-orous insects: The experimental study of evolution in naturaland agricultural systems. Annual Review of Entomology 35:421–446.
Waterman, P. G. & S. Mole, 1989. Extrinsic factors influenc-ing production of secondary metabolites in plants. In: E. A.Bernays (ed), Insect-Plant Interactions I. CRC Press, BocaRaton, pp. 107–133.


















