SPATIAL OCCURRENCE OF A HABITAT-TRACKING SAPROXYLIC BEETLE INHABITING A MANAGED FOREST LANDSCAPE
10
Ecological Applications, 17(3), 2007, pp. 900–909 Ó 2007 by the Ecological Society of America SPATIAL OCCURRENCE OF A HABITAT-TRACKING SAPROXYLIC BEETLE INHABITING A MANAGED FOREST LANDSCAPE L. MARTIN SCHROEDER, 1 THOMAS RANIUS,BARBARA EKBOM, AND STIG LARSSON Swedish University of Agricultural Sciences, Department of Entomology, P.O. Box 7044, SE-750 07 Uppsala, Sweden Abstract. Because of the dynamic nature of many managed habitats, proper evaluation of conservation efforts calls for models that take into account both spatial and temporal habitat dynamics. We develop a metapopulation model for successional-type systems, in which habitat quality changes over time in a predictable fashion. The occupancy and recruitment of the predatory saproxylic (dependent on dead wood) beetle Harminius undulatus was studied in a managed boreal forest landscape, covering 24 449 ha, in central Sweden. In a first step, we analyzed the beetle’s occupancy pattern in relation to stand characteristics, and the amounts of present and past habitat in the surrounding landscape. Managed forest is suitable habitat when 60 years old, and immediately after cutting, but not between the ages of 10 and 60 years. The observed occupancy of H. undulatus was positively correlated with the stand’s age as habitat. We used a metapopulation model to predict the current probability of occurrence in each forest stand, given the spatiotemporal distribution of suitable forest stands during the last 50 years. Metapopulation parameters were estimated by matching predicted spatial distributions with observed spatial distributions. The model predicted observed spatial distributions better than a similar model that assumed constant habitat quality of each forest stand. Thus, metapopulation models for successional-type systems, such as dead wood dependent organisms in managed forest landscapes, should include habitat dynamics. An estimated 82% of the landscape-wide recruitment took place in managed stands, which covered 87% of the forest area, in comparison with 18% in unmanaged stands, which covered 13% of the forest area. Among the managed stand types, 60-year-old stands and 3–7-year- old clear-cuttings contributed to 79% of the total recruitment while 8–59-year-old stands only contributed 3%. The results suggest the following guidelines to improve conditions for H. undulatus and other species with similar habitat requirements: (1) the proportion of the landscape constituted by younger stands should not be allowed to grow too large, (2) the rotation period of managed stands should not be allowed to be too short, and (3) dead wood should be retained and created at final cutting. Key words: habitat-tracking metapopulation; Harminius undulatus; incidence function model; landscape-wide recruitment; modeling; occupancy pattern; saproxylic beetle. INTRODUCTION The metapopulation concept provides a theoretical basis for studies of population dynamics in spatially structured habitats (Hanski and Gaggiotti 2004). In classical metapopulation models, the spatial structure of the habitat is considered, but no temporal changes of the habitat are taken into account (e.g., Hanski 1994). It has been argued, however, that in many cases metapopula- tion dynamics are strongly affected by the process of destruction and creation of habitat patches (Thomas 1994). Such metapopulations are referred to as habitat tracking (Harrison and Taylor 1997) because the populations track the habitat when they colonize newly generated habitat patches and become extinct in patches that deteriorate. Incorporating habitat dynamics in metapopulation models may have a large impact on metapopulation persistence (Keymer et al. 2000, Akc¸a- kaya et al. 2004, Bergman and Kindvall 2004). Thus, models that take habitat dynamics into account are useful tools for proper evaluation of conservation efforts performed in dynamic landscapes (Stelter et al. 1997, Wahlberg et al. 2002, Biedermann 2004, Verheyen et al. 2004, Akc¸akaya et al. 2005, Sna¨ll et al. 2005). Boreal forest landscapes in Fennoscandia are typically managed by clear-cutting and consist of a mosaic of stands of different ages (Esseen et al. 1997). Within each managed stand, tree age is similar. When the stands are mature they are harvested, and a new stand is regenerated. Thus, each successional stage (e.g., clear- cut, young stand, and mature stand) persists for only a limited period of time. The availability of coarse woody debris (CWD) in managed stands is strongly linked to stand age (Fridman and Walheim 2000, Ranius et al. 2003). During the first years after clear-cutting, CWD is found in stands as remnants from the harvested mature stands as well as CWD deliberately left for biodiversity conservation. Gradually, CWD decays and disappears Manuscript received 18 January 2006; revised 7 September 2006; accepted 3 October 2006. Corresponding Editor: D. L. Peterson. 1 E-mail: [email protected] 900
Transcript of SPATIAL OCCURRENCE OF A HABITAT-TRACKING SAPROXYLIC BEETLE INHABITING A MANAGED FOREST LANDSCAPE

Ecological Applications, 17(3), 2007, pp. 900–909� 2007 by the Ecological Society of America
SPATIAL OCCURRENCE OF A HABITAT-TRACKING SAPROXYLICBEETLE INHABITING A MANAGED FOREST LANDSCAPE
L. MARTIN SCHROEDER,1 THOMAS RANIUS, BARBARA EKBOM, AND STIG LARSSON
Swedish University of Agricultural Sciences, Department of Entomology, P.O. Box 7044, SE-750 07 Uppsala, Sweden
Abstract. Because of the dynamic nature of many managed habitats, proper evaluation ofconservation efforts calls for models that take into account both spatial and temporal habitatdynamics. We develop a metapopulation model for successional-type systems, in whichhabitat quality changes over time in a predictable fashion. The occupancy and recruitment ofthe predatory saproxylic (dependent on dead wood) beetle Harminius undulatus was studied ina managed boreal forest landscape, covering 24 449 ha, in central Sweden. In a first step, weanalyzed the beetle’s occupancy pattern in relation to stand characteristics, and the amountsof present and past habitat in the surrounding landscape. Managed forest is suitable habitatwhen �60 years old, and immediately after cutting, but not between the ages of 10 and 60years. The observed occupancy of H. undulatus was positively correlated with the stand’s ageas habitat. We used a metapopulation model to predict the current probability of occurrencein each forest stand, given the spatiotemporal distribution of suitable forest stands during thelast 50 years. Metapopulation parameters were estimated by matching predicted spatialdistributions with observed spatial distributions. The model predicted observed spatialdistributions better than a similar model that assumed constant habitat quality of each foreststand. Thus, metapopulation models for successional-type systems, such as dead wooddependent organisms in managed forest landscapes, should include habitat dynamics. Anestimated 82% of the landscape-wide recruitment took place in managed stands, whichcovered 87% of the forest area, in comparison with 18% in unmanaged stands, which covered13% of the forest area. Among the managed stand types, �60-year-old stands and 3–7-year-old clear-cuttings contributed to 79% of the total recruitment while 8–59-year-old stands onlycontributed 3%. The results suggest the following guidelines to improve conditions for H.undulatus and other species with similar habitat requirements: (1) the proportion of thelandscape constituted by younger stands should not be allowed to grow too large, (2) therotation period of managed stands should not be allowed to be too short, and (3) dead woodshould be retained and created at final cutting.
Key words: habitat-tracking metapopulation; Harminius undulatus; incidence function model;landscape-wide recruitment; modeling; occupancy pattern; saproxylic beetle.
INTRODUCTION
The metapopulation concept provides a theoretical
basis for studies of population dynamics in spatially
structured habitats (Hanski and Gaggiotti 2004). In
classical metapopulation models, the spatial structure of
the habitat is considered, but no temporal changes of the
habitat are taken into account (e.g., Hanski 1994). It has
been argued, however, that in many cases metapopula-
tion dynamics are strongly affected by the process of
destruction and creation of habitat patches (Thomas
1994). Such metapopulations are referred to as habitat
tracking (Harrison and Taylor 1997) because the
populations track the habitat when they colonize newly
generated habitat patches and become extinct in patches
that deteriorate. Incorporating habitat dynamics in
metapopulation models may have a large impact on
metapopulation persistence (Keymer et al. 2000, Akca-
kaya et al. 2004, Bergman and Kindvall 2004). Thus,
models that take habitat dynamics into account are
useful tools for proper evaluation of conservation efforts
performed in dynamic landscapes (Stelter et al. 1997,
Wahlberg et al. 2002, Biedermann 2004, Verheyen et al.
2004, Akcakaya et al. 2005, Snall et al. 2005).
Boreal forest landscapes in Fennoscandia are typically
managed by clear-cutting and consist of a mosaic of
stands of different ages (Esseen et al. 1997). Within each
managed stand, tree age is similar. When the stands are
mature they are harvested, and a new stand is
regenerated. Thus, each successional stage (e.g., clear-
cut, young stand, and mature stand) persists for only a
limited period of time. The availability of coarse woody
debris (CWD) in managed stands is strongly linked to
stand age (Fridman and Walheim 2000, Ranius et al.
2003). During the first years after clear-cutting, CWD is
found in stands as remnants from the harvested mature
stands as well as CWD deliberately left for biodiversity
conservation. Gradually, CWD decays and disappears
Manuscript received 18 January 2006; revised 7 September2006; accepted 3 October 2006. Corresponding Editor: D. L.Peterson.
1 E-mail: [email protected]
900

causing a scarcity of CWD when the forest is 20–60
years old. Thereafter, the amount of CWD increases
continuously until final cutting (Fridman and Walheim
2000, Ranius et al. 2003). This pattern is even more
pronounced for CWD that is still covered by bark
(Ekbom et al. 2006). In addition to managed stands,
some unmanaged stands may also be present in
managed forest landscapes. The amount of CWD is
higher in these stands than in managed stands (Fridman
and Walheim 2000). In Fennoscandia, the proportion of
the forest land left unmanaged is generally very small
(National Board of Forestry 2004). Organisms depen-
dent on CWD will be compelled to either rely solely on
the small proportion of unmanaged stands or be able to
track new habitat patches (that might arise when the
forest stand is about 60 years old) to compensate for
extinctions in stands following final cutting. Conse-
quently, the concept of habitat-tracking metapopula-
tions may be useful to describe the dynamics of species
inhabiting CWD in managed forest landscapes.
The occurrence pattern of species inhabiting a
dynamic forest landscape is expected to be related not
only to the current spatial structure of their habitat, but
also to the history of spatial structure change because
populations do not respond immediately to landscape
changes (Tilman et al. 1994, Hanski 1999, Ovaskainen
and Hanski 2002). When studying the dynamics of a
habitat-tracking metapopulation, long-term large-scale
experiments would provide the most reliable results.
However, such experiments are very expensive, and the
delay in obtaining results is long. An alternative is to use
occupancy data from snapshot studies in combination
with information about the historical distribution of
habitat (e.g., Snall et al. 2004, Verheyen et al. 2004).
In this study, we assessed the occupancy and
recruitment patterns of the CWD-dependent beetle
Harminius undulatus (De Geer) (Coleoptera: Elateridae)
in one 24 449-ha managed forest landscape in central
Sweden. We related its occupancy to attributes of
individual forest stands and to the spatiotemporal
distribution of habitat in the landscape. The investigated
stand attributes included variables that reflect habitat
quality and stand history. The landscape variables
reflected current and past density and distribution of
habitat in the landscape. We developed a metapopula-
tion model for successional-type systems, in which
habitat quality changed over time in a predictable
fashion. We used this metapopulation model to predict
the current probability of occurrence in each forest
stand. Metapopulation parameters were estimated by
matching predicted spatial distributions with observed
spatial distributions. We compared the observed occur-
rence pattern with the outcome from this model, and
also with a model assuming constant habitat quality of
each forest stand. This was done to test whether a
habitat-tracking model is superior to a static model in
describing this system. Harminius undulatus was chosen
for the study because (1) it shares its unspecific habitat
requirements regarding tree species with many other
beetle species dependent on CWD (Dahlberg andStokland 2004), and (2) it is a species at risk of declining
in the future and therefore can serve as a model for otherthreatened CWD-dependent organisms.
The aims of this study were (1) to determine theimportance of stand and landscape attributes for the
stand occupancy of H. undulatus in a managed forestlandscape, (2) to test if a habitat-tracking metapopula-tion model can help explain the current pattern of H.
undulatus occurrence, and (3) to estimate the relativeimportance of different kinds of forest stands for the
recruitment of H. undulatus across large landscapes.
MATERIALS AND METHODS
Harminius undulatus
The larva of H. undulatus is predatory and lives under
bark of dead wood. No specific tree species seem to befavored (Nilsson and Baranowski 1996, Dahlberg and
Stokland 2004). Developmental time has been reportedto be four years (Laibner 2000), but no reference as to
the source of this statement is provided. In Sweden, thespecies pupates in June, and adults occur in June andJuly (Palm 1946). Harminius undulatus has a wide
geographical distribution in northern Europe, Asia,and North America (Laibner 2000). In Sweden, the
species has been found in only a few localities in thesouth, whereas it becomes more common toward the
north (Lundberg 1995, Nilsson and Baranowski 1996,Gardenfors 2000). It has been suggested that H.
undulatus has become rare in southern Sweden due tothe dramatic decrease in suitable habitats in the forest
landscape in the 18th and 19th century (Nilsson andBaranowski 1996). It is likely that the clear-cutting of
forests in central and northern Sweden will similarlyresult in decreased H. undulatus populations there.
Nevertheless, at present the species still occurs in anumber of the forest stands in our study landscape,
which makes an analysis of its occurrence patternpossible.
Study landscape
The study was conducted in central Sweden during the
summers of 2001–2003. The study area is situated in thecentral boreal region in the province of Halsingland
close to Delsbo (628 N, 168 E) about 40 km southwest ofSundsvall. One large block of land (hereafter denoted
landscape) owned by the forest company Holmen SkogAB was used for the study. The size of the landscape is
24 449 ha of which 20 294 ha consists of productiveforest land (wood production � 1 m3�ha�1�yr�1). Theremaining area is covered by lakes, mires, agriculturalland, and unproductive forest land (wood production ,
1 m3�ha�1�yr�1, i.e., mainly wooded mires and rockyoutcrops). Unproductive forests were estimated to cover701 ha, i.e., 2.9% of the landscape. These were not
included in the sampling, because no forest data wereavailable. This should not have influenced the results
April 2007 901SPATIAL OCCURRENCE OF A BEETLE

because the amount of CWD is low in this type of
habitat (Gessler 1998). The landscape is typical of the
Swedish boreal region with Scots pine (Pinus sylvestris
L.) and Norway spruce (Picea abies (L.) Karsten) the
dominant tree species. Birch (Betula pendula Roth. and
B. pubescens Ehrh.) and aspen (Populus tremula L.) are
the most common deciduous tree species, but in
managed stands they rarely constitute more than a
small proportion of the standing volume. Because
Holmen Skog AB is certified since 1998 according to
FSC (Forest Stewardship Council) guidelines, the forest
is currently managed using methods developed to
preserve biodiversity (guidelines available online).2 These
practices include green-tree retention and creation of
high stumps at final cuttings. Using logging machinery,
3–5 m high stumps are left standing when live trees are
cut. At present, an average of 1.2 high stumps/ha are
created (Schroeder et al. 2006) and 40 living trees per ha
are retained at final cuttings in this landscape (O. Karen,
personal communication).
Selection of stands to be sampled
The forest land of the landscape was stratified into
seven stand categories based on stand age and manage-
ment (Table 1). The first two categories, 0–2 years and
3–7 years, represent clear-cuts on which creation of high
stumps and green-tree retention have been practiced at
final cutting. Older clear-cuts, 8–14 years, were cut
before these practices were adopted. In young stands,
15–59 years, recruitment of CWD has generally not yet
begun because tree mortality is low at these ages (Ranius
et al. 2003). Furthermore, in many of the young stands
tree diameter is ,10 cm. In stands �60 years,
recruitment of CWD has started. Set-asides are stands
that have been excluded from management but have no
legal protection. Reserves are legally protected forest
areas owned by the state.
In each category, except reserves, stands locatedthroughout the landscape were randomly sampled
(Table 1). No stands were sampled in the first category,
0–2 years, because CWD created at cutting was not oldenough to be colonized by H. undulatus. The largest
sampling effort was directed toward the 3–7 years old
clear-cuts, i.e., the stands where CWD had been createdas a result of adoption of FSC guidelines. The
probability for selection was proportional to the areaof the stand. All three nature reserves located in the
study landscape were sampled. Because the reserves were
larger than managed stands the reserves were dividedinto sub-areas (denoted stands). The two largest
(Stensjon och Lomtjarn, 427 ha; Hagasen, 242 ha) were
divided into six sub-areas of approximately equal sizeand the smallest (Flottholjan, 82 ha) was divided into
two sub-areas. The areas given for the reserves include
only forest land. In each chosen stand, amounts ofCWD and occurrence of H. undulatus were estimated.
Sampling of bark on CWD
Sampling of logs (downed CWD) was carried out
using the line intersect sampling method (Marshall et al.
2000). Four randomly selected transects of 100 m (twoeast–west and two north–south) were sampled in each
stand. The same transects were used to delimit sampling
plots for snags (standing CWD). The plot extended 10 mon each side of the transect forming a 20 3 100 m
rectangle (0.2 ha). Logs were included if the diameter atthe line intersect was at least 10 cm for conifers, and 7 cm
for deciduous trees. Snags were included if the diameter
at 1.3 m (dbh) was at least 10 cm and if the length was atleast 0.5 m. The following information was recorded for
every object: (1) classification as snag or log, (2) intact or
broken/cut snag, (3) diameter (at transect line for logsand at breast height for snags), (4) height of snag, (5) tree
species, (6) proportion of surface area covered by bark,
and (7) decay class. For standing objects shorter than 1.3m, the top diameter was measured. For a more detailed
description of CWD estimation and results see Ekbom et
al. (2006). The classification into decay classes was based
TABLE 1. Composition of the landscape at Delsbo, central Sweden, number of stands sampled forHarminius undulatus, and CWDbark area (decay class 2–3) for each of the different stand categories.
Stand category No. stands Area (ha) No. sampled stands Bark area (m2/ha)
Managed forest
0–2 years 20 343 � �3–7 years 47 666 20 88.808–14 years 82 1744 10 2.1515–59 years 796 9459 11 8.80�60 years 853 5411 28 85.60
Unmanaged forest
Set asides 375 1920 9 176.90Reserves 14 751 7 166.90
Sum of forest 2176 20 294 85Non-forest 4155Entire landscape 24 449
Note: Estimates of bark area are from Ekbom et al. (2006); CWD is coarse woody debris.� This stand category was not sampled.
2 hwww.fsc-sweden.orgi
L. MARTIN SCHROEDER ET AL.902 Ecological ApplicationsVol. 17, No. 3

on the hardness of the wood. We used the same
classification system as Siitonen and Saaristo (2000),
i.e., six categories where decay class one is fresh wood
and six is late decay.
The area of bark on single snags was calculated by
using the formula for area of a cone multiplied by the
proportion of remaining bark. Surface area of logs was
estimated by modifying the formula for ‘‘total projected
area’’ (Marshall et al. 2000) by replacing diameter with
circumference. Surface area for each object was multi-
plied by the proportion of remaining bark. For each
stand an average for each variable was calculated based
on the four plots or intersect lines.
Sampling of Harminius undulatus
The presence and abundance of H. undulatus larvae in
individual stands were assessed by peeling bark from
CWD objects, sieving the material, and extracting the
larvae in Tullgren funnels. By sampling H. undulatus
larvae under bark we could be sure that reproduction
had taken place in the substrate and thereby obtained a
measure of the addition of new individuals to a stand. If
adults captured in flight-interception traps had been
tallied, we would not know if reproduction was actually
taking place in the stand (i.e., if the stand was
colonized). Ten pieces of CWD of decay class 2–3 were
sampled in each stand. The sampling was restricted to
these two decay classes because H. undulatus does not
occur in the first decay class and because decay classes
older than class 3 were assumed to be too old for the
species. This assumption was based on the fact that H.
undulatus requires CWD with bark (M. Jonsell and J. N.
Stokland, personal communication) and that relatively
small amounts of bark were present on decay classes
older than 3 (Ekbom et al. 2006). Both logs and snags
were sampled when they both occurred. Only spruce
objects were sampled during the first year, while pine
and birch were also included during the following two
years. In stands where 10 suitable objects were not
available, all appropriate objects were sampled. In 2001,
each sample consisted of 0.3–0.5 m2 bark. As a result of
the low density of H. undulatus per area of bark, sample
area was increased to 1 m2 per object in 2002 and 2003.
Occupancy of Harminius undulatus
The relationship between stand and landscape attri-
butes and occupancy of H. undulatus per stand was
analyzed by multiple logistic regression. Two stand
categories, managed stands 8–14 years old and 15–59
years old, were considered as non-habitat based on the
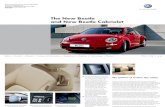
low number of individuals present per hectare (Fig. 1).
Nonforest land was assumed to host no CWD and was
classified as non-habitat. All other stand types, which
generally contain suitable substrate for H. undulatus
were considered as habitat. Thus, in the analysis we only
considered 56 sampled stands belonging to forest types
deemed to be habitat for H. undulatus.
The following variables were used: (1) stand bark area
(m2/ha), (2) stand size (ha), (3) stand altitude (m above
sea level), (4) age as habitat (i.e., how long ago the stand
became habitat given that stands become suitable when
they are 60 years), (5) number of sampled dead wood
units, and (6) the proportion of the landscape within a
radius of 500 m covered by habitat in 2002, 1992, 1982,
1972, 1962, and 1952. Age as habitat was set to 50 years
for nature reserves, forest stands .110 years, and clear-
cuts. The historical occurrence of habitat in the
landscape was recreated by counting backwards from
present stand ages, and assuming that all stands were
more than 110 years old at their latest final cutting. The
proportion of the surrounding area covered by habitat
was quantified using ARC/View 8.2 (ESRI, Redlands,
California, USA). During the last 50 years, the percent-
age of the landscape covered by forest greater than or
equal to 60 years decreased by approximately 50% (Fig.
2). Clear-cuts from final cuttings in 1994 or later were
classified as habitat because this was the approximate
starting time for creation of high stumps (Schroeder et al.
2006) and green-tree retention. Stand altitude was
included in the analysis because it is a factor that
influences the distribution of single species as well as
species diversity (Begon et al. 1990). In the regression,
FIG. 1. Number of Harminius undulatus recruited per ha in the different stand categories. The two stand categories 8–14 yearsand 15–59 years were considered as non-habitat in the analyses.
April 2007 903SPATIAL OCCURRENCE OF A BEETLE

nonsignificant variables (P . 0.10) were excluded from
the final model using backward elimination (log likeli-
hood ratio significance) (SPSS 10.05 Advanced Statistics
software; SPSS, Inc. Chicago, Illinois, USA). The
response variable (beetle occupancy) was tested for
spatial autocorrelation (PASSAGE 1.0; software avail-
able online).3 Moran’s I was highest at the shortest
distance tested (I¼ 0.094, P¼ 0.10, 202 pairs). However,
no significant (P , 0.05) correlation was found at any of
the 10 tested distance classes. Thus, the assumption of
independent observations for the logistic regression was
not violated (Lennon 2000).
The occurrence of H. undulatus was also analyzed in
relation to the expected probability of presence predict-
ed by two metapopulation models, one habitat static
and one habitat tracking. This was done with a multiple
logistic regression model where we also included altitude
(that was not considered in the metapopulation models)
and stand size (because the effect of stand size was
minimized in the metapopulation model by assuming the
local extinction risk to be independent of stand size) as
independent variables, but excluded variables that we
should expect to be correlated with the model outcome.
The simulation outcome may be spatially correlated, but
it has been shown that logistic regression is robust to
moderate levels of spatial correlation in metapopulation
occupancy patterns (Johnson 2005).
Using a metapopulation model to explain observed
occupancy patterns
We modeled the occupancy of H. undulatus in forest
stands using a habitat-tracking metapopulation model
and assumptions about when forest stands become
habitat or non-habitat. This was compared with a
habitat-static metapopulation model that was similar,
except for the assumption that the current distribution
of habitat per non-habitat had been the same over the
last 50 years. The result was the probability of current
occurrence in each stand, which was used as an
independent variable in the logistic regression model.
By choosing different parameter combinations in the
metapopulation model, different spatial and temporal
scales were considered. The main aims were (1) to
analyze whether a metapopulation model, taking into
account the spatial structure of the habitat patches,
could explain anything of the observed occupancy
pattern, and (2) to test whether the prediction from a
habitat-tracking model is superior to a simpler habitat-
static model. Because the aim was not to predict
minimum viable metapopulation sizes, or to compare
extinction risks using different management scenarios,
we could use a simple model (e.g., without stochastic
variability of any parameter over time), and solve the
problem of lack of empirical data for parameter
estimation by testing many different parameter combi-
nations. The approach applied was developed by Gu et
al. (2002) and is based on the incidence function model
(Hanski 1994, 1999). We modified the model slightly,
because in the present study the occupancy of H.
undulatus was sampled per stand instead of per grid cell.
Forest stands, differing in size, were thus considered as
habitat patches. We assumed that habitat patch size
(denoted by A) is equal to the size of the stand (in ha) if
it was a suitable habitat, otherwise it was set to zero.
For each year, the colonization probability for an
empty stand is a function of its connectivity (S) to
existing local populations. The connectivity of stand i is
defined as
Si ¼X
expð�adijÞpjAj ð1Þ
where p¼ 0 for empty and p¼ 1 for occupied stands, dijis the distance between stands i and j, and 1/a is the
average migration distance. This equation is the same as
that in Hanski (1994) except that the parameter b, which
scales the rate of emigration by area, was set to 1. The
colonization probability C is assumed to be a sigmoid
function of connectivity:
Ci ¼ S2i =ðS2
i þ y2Þ ð2Þ
where y is a parameter regulating the colonization rate.
For each year, the probability of extinction (E ) in an
unsuitable patch is 1, and the probability of extinction in
a suitable patch is
Ei ¼ min 1; uð1� CiÞf g ð3Þ
where u is a parameter regulating the extinction risk and
(1 � C) represents the rescue effect. When no suitable
habitat is available, the larvae of H. undulatus never
survive, and therefore we assumed that the local
population immediately goes extinct when a habitat
patch becomes unsuitable. In the model by Gu et al.
(2002), extinction probability was assumed to scale by
the area, but this was removed from the equation.
Moreover, in contrast to Gu et al. (2002), we did not
FIG. 2. Historical and present percentages of the studylandscape covered by forest stand categories defined as habitatfor Harminius undulatus. These stand categories includemanaged stands 3–7 years old (only in 2002) and �60 yearsold, and the unmanaged stand types set asides and reserves. Thehistorical percentages were recreated by counting backwardfrom present stand ages.
3 hhttp://www.passagesoftware.neti
L. MARTIN SCHROEDER ET AL.904 Ecological ApplicationsVol. 17, No. 3

implement any regional stochasticity. The modeled
landscape was 28.6 3 28.1 km, which was the smallest
rectangle possible where all sampled stands were .4 km
from the nearest landscape boundary. We considered all
forest stands in the landscape. In the habitat-tracking
model, we estimated when the stands switched between
the two possible states, being suitable or unsuitable for
H. undulatus, over 50 years. Simulations were started
with an initial condition of 60% occupancy of randomly
selected suitable stands in 1952. This means that we
assumed the occupancy per suitable stand to be higher at
that time than today, because the density of suitable
stands was higher. The landscape structure was updated
every year. Populations occupying the suitable stands
that turned into unsuitable stands were lost with their
habitat. In the habitat-static model we assumed that
each stand has been either suitable or unsuitable
constantly over the last 50 years, according to the
current situation. The initial occupancy was 30%, which
is close to the occupancy in field data.
We did not have the required information in order to
estimate model parameters (a, u, y) for H. undulatus.
Therefore, we first applied a very wide range of
parameter combinations. We found that a rather low
extinction risk (low value of u) and long dispersal
distances (low value of a) were required in order to
obtain occupancy patterns that approximated field data.
We then systematically searched for parameter combi-
nations that generated outcomes significantly correlated
with field data. We used three different values of u: 0.06,
0.12, and 0.18. Five different mean dispersal distances
(1/a) were used: 500 m, 1000 m, 1500 m, 2000 m, and
2500 m. To obtain reasonable rates of occupancy, we
adjusted y in order to obtain occupancy of about 30% in
the surveyed forest stands. This was close to the average
proportion of occupied stands observed (which was
27%, see Results). This means that when the local
extinction risk (u) increased it had to be compensated for
with increased colonization (y decreased), which implies
that we compared different turnover rates. The connec-
tivity values (S in Eq. 1) were high in our study because
the habitat patches are large and numerous. Therefore,
to obtain a colonization rate appropriate for obtaining
the desired occupancy rate (30%), the y values (in Eq. 2)
became very large in comparison to some other studies
using the incidence function model (Moilanen et al.
1998, Hanski 1999). When the mean dispersal distance
increased (a decreased), it was compensated for by a
lower colonization probability (y increased). One
hundred simulations were run for each of the parameter
combinations, which was enough for obtaining stable
outcomes.
Recruitment of Harminius undulatus
The number of H. undulatus larvae developing in each
stand category (‘‘recruitment’’) was estimated based on
proportion of occupied stands, mean density (no./m2
bark) for occupied stands, and bark area. In the
calculations, the average proportion of occupied stands
for two groups of stand types that were significantly
different (see Results) was used. The first consisted of 3–
7- and �60-year-old stands, and the second group
contained the remaining stand types. The mean density
in occupied stands was calculated across stand catego-
ries, because the number of observations for each stand
category was low. The calculations were based on all H.
undulatus larvae irrespective of size.
RESULTS
Occupancy of Harminius undulatus
Occupancy increased with age as habitat, whereas it
decreased with altitude (Table 2) for habitat stands (i.e.,
those 56 stands sampled belonging to unmanaged stand
types and the managed stand types 3–7 years old and
�60 years old). Neither bark area per ha nor proportion
of habitat in the surroundings (within 500 m) in the
present or the past were significantly related to the
occupancy of H. undulatus.
The relationship between probability of occurrence
predicted from the metapopulation model and observed
occupancy was stronger (the P value was generally
lower) for the habitat-tracking model than for the
habitat-static model (Table 3). For both models,
strongly significant relationships were more common
for parameter combinations with fairly large dispersal
range (1/a � 1500), and a low extinction risk per stand
(u ¼ 0.06 or 0.12).
Recruitment of Harminius undulatus
Based on the proportion of stands occupied by H.
undulatus, the stand types could be grouped into three
categories: (1) 3–7-year-old clear-cuts and managed
stands �60 years old in which about 30% of the stands
were occupied, (2) managed stands 15–59 years old, set
asides and reserves in which 9–14% of the stands were
TABLE 2. Results from multiple logistic regression withpresence (1) or absence (0) of Harminius undulatus per standas the dependent variable, and variables describing the standand the surroundings, currently and historically, as indepen-dent variables.
Variable Coefficient P
Included in the model
Altitude �0.029 0.004Age as habitat 0.078 0.018Stand area �0.039 0.092Intercept 6.06
Excluded from the model
Proportion of habitat 1952 0.743Proportion of habitat 1962 0.439Proportion of habitat 1972 0.707Proportion of habitat 1982 0.926Proportion of habitat 1992 0.865Proportion of habitat 2002 0.833Bark area per hectare 0.998No. sampled trees 0.565
Note: The model was obtained by backward eliminationbased on log likelihood ratio significance; N¼ 56 stands.
April 2007 905SPATIAL OCCURRENCE OF A BEETLE

occupied, and (3) managed stands 8–14 years old in
which no occupied stands were found (Fig. 3). The
occupancy was significantly higher in stands of the first
category than in the other two categories (category 1
compared with category 2, v2¼ 3.84, P¼ 0.05; category
1 compared with category 3, v2¼ 4.21, P¼ 0.04), while
the second category was not significantly different from
the third category (Fisher’s exact test). The average
proportion of occupied stands was 27% for stands
classified as habitat. The mean density of H. undulatus in
occupied stands was 1.5 individuals/m2 bark (SD¼ 0.9).
In the study landscape overall, managed stands
contributed more to H. undulatus recruitment than
unmanaged stands. An estimated 82% of the total
recruitment took place in managed stands (covering 87%
of forest area) compared with 18% in unmanaged stands
(covering 13%; Fig. 4). Stands �60 years old contributed
70% of the total recruitment while 8–59-year-old stands
only contributed 3%. About 9% of the total recruitment
of H. undulatus in the landscape took place in 3–7-year-
old clear-cuts.
Most H. undulatus larvae were small. Of the largest
larvae (head capsule width � 2.0 mm), there was never
more than one individual per 1-m2 sample. In contrast,
up to seven individuals of the smallest larvae (head
capsule width � 0.4 mm) were found per 1-m2 sample.
Larvae of the largest and smallest size were often found
in the same sample, suggesting thatH. undulatus uses the
same CWD object as oviposition substrate for several
years.
DISCUSSION
In this study, we demonstrated that metapopulation
models can be extended to deal with successional-type
systems in which habitat suitability changes over time in
a predictable pattern. The cycle of succession is well
known in many managed systems, and this knowledge
can be used to classify habitat quality. Both spatial and
temporal aspects can thus be examined using metapop-
TABLE 3. Significance of the simulation model predictions in a multiple logistic regression model with forest stand area, altitude,and simulation model prediction as independent variables, and observed occupancy of Harminius undulatus per stand as thedependent variable.
1/a (m)
P y
u ¼ 0.06 u ¼ 0.12 u ¼ 0.18 u ¼ 0.06 u ¼ 0.12 u ¼ 0.18
Habitat-tracking model
500 0.046 0.081 0.095 170 110 901000 0.012 0.054 0.067 700 455 3451500 0.004 0.020 0.018 1500 950 7502000 0.010 0.025 0.013 2500 1600 12502500 0.004 0.001 0.007 3800 2400 1880
Habitat-static model
500 0.441 0.400 0.437 130 85 701000 0.406 0.382 0.571 445 300 2551500 0.379 0.253 0.177 970 640 5052000 0.045 0.048 0.090 1600 1040 8302500 0.153 0.055 0.053 2250 1520 1200
Notes: P values (likelihood ratio significance) are from a regression model obtained with backward elimination; boldfaceindicates P , 0.05. The predictions were from metapopulation models with different parameter combinations: one model assumeda dynamic habitat, and the other a static habitat. Definitions: 1/a is the mean dispersal distance, and u is a parameter reflecting theextinction risk per stand. Values of y were changed to compensate for the different values of 1/a and u in order to obtain observedoccupancy.
FIG. 3. Percentage of stands occupied by Harminius undulatus in the different stand categories: 3–7 yr, N¼ 20; 8–14 yr, N¼ 10;15–59 yr, N ¼ 11; �60 yr, N ¼ 28; set asides, N ¼ 9; and reserves, N ¼ 7. N is number of stands per category.
L. MARTIN SCHROEDER ET AL.906 Ecological ApplicationsVol. 17, No. 3

ulation models. Our results strongly suggest that, in a
managed forest landscape, the CWD dependent beetle
H. undulatus can be described as a habitat-tracking
metapopulation (sensu Thomas 1994). Three lines of
evidence lend support to this suggestion. First, there was
a strong relationship between habitat quality (area of
bark cover) and stand age. This means that managed
forests shift between the states of habitat and non-
habitat in a predictable manner. Managed forest stands
change from habitat to non-habitat for H. undulatus
immediately or after a delay of several years after clear-
cutting depending on whether or not suitable CWD is
retained at final cutting. The shift from non-habitat to
habitat occurs about 60 years after final cutting (Fig. 1).
Second, there was a significant relationship between H.
undulatus occurrence and the stand’s age as habitat.
After a stand becomes suitable it often seems to take
some time before it becomes colonized. Thus dispersal
between forest stands may be so restricted that each
forest stand can be regarded as a habitat patch, with a
local population only somewhat connected with other
populations. Third, there was a significant relationship
between H. undulatus occurrence and the probability of
occurrence per stand predicted from the habitat-tracking
metapopulation model, which was stronger than the
relationship using the prediction from the habitat-static
metapopulation. We assumed that local extinction risk
was the same in all suitable stands as long as they were
not affected by any rescue effect. Thus, the significant
relationship obtained must be the result of the fact that
the colonization rate or rescue effect (which both result
from dispersal) is constrained where dispersal sources
are, or have been, small and distant.
We also show that it is possible to roughly estimate
metapopulation model parameters by matching predict-
ed and observed spatial distributions. This provides an
approach to estimating individual parameters in the
incidence function model even if equilibrium between
local colonizations and extinctions could not be
assumed. Results from the metapopulation model
indicate that H. undulatus has a large dispersal range;
the relationship between observed occupancy and the
prediction from the metapopulation model was stronger
when the mean dispersal distance was set to �1500 m in
comparison with shorter distances (Table 3). This may
explain the lack of a relationship between H. undulatus
occupancy and the proportion of present habitat within
a radius of 500 m. Although the dispersal range might be
wide, two sets of data suggest that the occurrence
pattern may also be influenced by a restricted ability to
colonize suitable stands. First, H. undulatus did not
occur in all suitable stands, but its occupancy pattern
was significantly correlated with the outcome from the
metapopulation model when we assumed a low turnover
rate (i.e., low extinction risk and colonization rate) per
stand. Second, occupancy was positively correlated with
the time that the stands had been a suitable habitat.
Stands that had recently become habitat had lower
occupancy, probably because of a shorter time for
colonization in comparison with older stands.
The present study demonstrates that managed stands
are important for the landscape-wide recruitment of H.
undulatus; the main part (82%) of the estimated
landscape-wide recruitment took place in managed
stands. Especially important among the managed stand
types were stands � 60 years, which accounted for 70%
of the landscape-wide recruitment. In contrast to
managed stands � 60 years, the contribution to
recruitment of H. undulatus by managed stands 8–59
years was small, despite the fact that they covered more
than 50% of the forest land of the landscape. This is a
consequence of the low amounts of CWD with bark
FIG. 4. Percentage of the landscape covered by each stand category (percentage of forest area) and the percentage of the totalrecruitment (percentage of landscape-wide recruitment) of Harminius undulatus occurring in each stand category. Managed stands0–2 years old were not included in the analyses because they were not sampled.
April 2007 907SPATIAL OCCURRENCE OF A BEETLE

occurring in these stand types (Ekbom et al. 2006). Thus,
the age distribution of stands within managed land-
scapes is an important factor for saproxylic organisms
dependent on bark.
In the Delsbo landscape larval density in occupied
stands was not higher in set-asides or reserves than in the
managed stands � 60 years and new forestry clear-cuts
(Fig. 1). The proportion of occupied stands was lower in
set-asides or reserves than in the other stand types. This
may, to some extent, be due to the fact that the reserve
stands generally were larger than other stands, and H.
undulatus would probably have been found in most
reserve stands if the number of sampled CWD objects
had been higher. On the other hand, this would imply a
lower larval density of the species in occupied reserve
stands compared with the other stand types. Thus, it is
unlikely that a sampling regime based on amount of
available resource in each stand would dramatically
change the results regarding the importance of managed
vs. unmanaged stands for the landscape-wide recruit-
ment.
Management implications for species preservation
Stands 8–59 years old contributed little to the
landscape-wide recruitment because of the low avail-
ability of bark on CWD. The persistence probability for
a species is related to the proportion of habitat. It is
therefore important that the proportion of the landscape
comprised by older stands is not allowed to become too
small. The proportion of the landscape covered by forest
older than 60 years has decreased in the last 50 years
(Fig. 2). The reduction of old and mature forest areas in
recent years is a general pattern in large parts of boreal
Fennoscandia (Kouki et al. 2001 and references therein).
This situation could be improved by modifying the age
distribution of the forest stands. However, such an
improvement would require a considerable amount of
time and most likely be expensive. An alternative
strategy would be to transform younger stands into
habitats for H. undulatus by increasing the amounts of
CWD with bark. This could be done by increasing the
number of living trees retained at final harvest. Some of
these trees in the young stands will die from natural
causes or could be cut to increase the recruitment of
CWD at a later date (Ranius et al. 2003). It is also
important that the habitat quality of older stands is
maintained. Management practices, such as intensive
thinning, that result in lower quantities of CWD should
be avoided.
The size and persistence of a habitat-tracking
metapopulation can be strongly affected by the duration
of individual habitat patches (Fahrig 1992, Keymer et al.
2000). In a managed forest landscape like Delsbo,
rotation length determines the period of time a stand is
suitable habitat. A shortened rotation period of
managed stands would have a detrimental effect on H.
undulatus. The situation after harvest will also influence
the length of time the habitat patches remain suitable; if
CWD is retained and created at final harvest, the beetle
species is expected to persist for some years while if no
care is taken at harvest it may go extinct shortly after
cutting.
This study focused on a beetle species dependent on
bark-covered CWD. This species was common enough
to enable quantitative analyses and rare enough to be
considered a model for other saproxylic organisms
threatened by modern forestry. About 60% of all
saproxylic beetles inhabiting spruce are directly depen-
dent on bark and of these 20% are included in the
Swedish red-list (Gardenfors 2005; M. Jonsell, personal
communication). Among the species-rich group of
saproxylic beetles (1260 species in Sweden [Dahlberg
and Stokland 2004]) bark dependency is also common
among species not on the Red List (Saalas 1917, Palm
1951, 1959). We should expect a highly variable set of
species responses to low levels of dead wood. Some
species may persist even when the amount of CWD is
very low or have such a high colonization ability that
they can occur in almost any managed forest where there
is suitable habitat. Such species do not have to be
considered in evaluating biodiversity-oriented forestry.
Other species (such as red-listed polypores [Penttila et al.
2004]) may be absent in stands with low amounts of
CWD such as those where H. undulatus were found.
Such species are not relevant for evaluation of biodi-
versity-oriented management, because they obviously
need unmanaged forest land for persistence. Intermedi-
ate to these two extremes are species like H. undulatus,
which will be able to persist in managed forests if
management is adapted to their needs. Assuring a
supply of bark-covered CWD over long periods of time
in individual stands may contribute appreciably to the
persistence of these species.
ACKNOWLEDGMENTS
Support for this project came from the project ‘‘Conserva-tion of Biodiversity in Managed Forest’’ financed by theFaculty of Forestry at the Swedish University of AgriculturalSciences and Thomas Ranius’ project ‘‘Predicting extinctionrisks for threatened wood-living insects in dynamic landscapes’’financed by The Swedish Research Council for Environment,Agricultural Sciences and Spatial Planning. Holmen Skog ABgenerously hosted our field study, and Per-Gunnar Jacobssonand Ola Karen were always ready to provide information. Weare indebted to Matilda Apelqvist, Bjorn Forsberg, MarkusFranzen, David Isaksson, Niklas Jonsson, Mats Larsson, PerLarsson, Carola Orrmalm, Erik Sahlin, Mans Svensson, andJan ten Hoopen for assistance in the field and laboratory. Wethank Tord Snall and Melodie McGeoch for comments on themanuscript.
LITERATURE CITED
Akcakaya, H. R., J. Franklin, A. D. Syphard, and J. R.Stephensson. 2005. Viability of Bell’s sage sparrow (Amphis-piza belli ssp. belli): altered fire regimes. Ecological Applica-tions 15:521–531.
Akcakaya, H. R., V. C. Radeloff, D. J. Mladenoff, and H. S.He. 2004. Integrating landscape and metapopulation model-ing approaches: viability of the sharp-tailed grouse in adynamic landscape. Conservation Biology 18:526–537.
L. MARTIN SCHROEDER ET AL.908 Ecological ApplicationsVol. 17, No. 3

Begon, M., J. L. Harper, and C. R. Townsend, editors. 1990.Ecology: individuals, populations and communities. Black-well Scientific Publications, London, UK.
Bergman, K.-O., and O. Kindvall. 2004. Population viabilityanalysis of the butterfly Lopinga achine in a changinglandscape in Sweden. Ecography 27:49–58.
Biedermann, R. 2004. Modelling the spatial dynamics andpersistence of the leaf beetle Goniactena olivacea in dynamichabitats. Oikos 107:645–653.
Dahlberg, A., and J. N. Stokland. 2004. Vedlevande arters kravpa substrat: sammanstallning och analys av 3600 arter.Skogsstyrelsen, Rapport 7. Jonkoping, Sweden. [In Swedishwith English summary.]
Ekbom, B., M. Schroeder, and S. Larsson. 2006. Stand specificoccurrence of coarse woody debris in a managed boreal forestlandscape in central Sweden. Forest Ecology and Manage-ment 221:2–12.
Esseen, P.-A., B. Ehnstrom, L. Ericson, and K. Sjoberg. 1997.Boreal forests. Ecological Bulletins 46:16–47.
Fahrig, L. 1992. Relative importance of spatial and temporalscales in a patchy environment. Theoretical PopulationBiology 41:300–314.
Fridman, J., and M. Walheim. 2000. Amount, structure, anddynamics of dead wood on managed forestland in Sweden.Forest Ecology and Management 131:23–36.
Gardenfors, U. 2000. The 2000 Red List of Swedish species.ArtDatabanken, Sveriges Lantbruksuniversitet, Uppsala,Sweden.
Gardenfors, U. 2005. The 2005 Red List of Swedish species.ArtDatabanken, Sveriges Lantbruksuniversitet, Uppsala,Sweden.
Gessler, C. 1998. Impedimentens potentiella betydelse forbiologisk mangfald. En studie av myr- och bergimpedimenti ett skogslandskap i Vasterbotten. [In Swedish with Englishsummary.] Arbetsrapport 45, Institutionen for skogligresurshushallning och geometik, Sveriges Lantbruksuniversi-tet, Umea, Sweden.
Gu, W., R. Heikkila, and I. Hanski. 2002. Estimating theconsequences of habitat fragmentation on extinction risk indynamic landscapes. Landscape Ecology 17:699–710.
Hanski, I. 1994. A practical model of metapopulationdynamics. Journal of Animal Ecology 63:151–162.
Hanski, I. 1999. Metapopulation ecology. Oxford UniversityPress, Oxford, UK.
Hanski, I., and O. E. Gaggiotti, editors. 2004. Ecology,genetics, and evolution of metapopulations. Elsevier Aca-demic Press, Amsterdam, The Netherlands.
Harrison, S., and A. D. Taylor. 1997. Empirical evidence formetapopulation dynamics. Pages 27–42 in I. A. Hanski andM. E. Gilpin, editors. Metapopulation biology. Ecology,genetics, and evolution. Academic Press, San Diego, Cal-ifornia, USA.
Johnson, D. M. 2005. Metapopulation models: an empiricaltest of model assumptions and evaluation methods. Ecology86:3088–3098.
Keymer, J. E., P. A. Marquet, J. X. Velasco-Hernandez, andS. A. Levin. 2000. Extinction thresholds and metapopulationpersistence in dynamic landscapes. American Naturalist 156:478–494.
Kouki, J., S. Lofman, P. Martikainen, S. Rouvinen, and A.Uotila. 2001. Forest fragmentation in Fennoscandia: linkinghabitat requirements of wood-associated threatened speciesto landscape and habitat changes. Scandinavian Journal ofForest Research Supplement 3:27–37.
Laibner, S. 2000. Elateridae of the Czech and Slovac Republics.Kabourek, Zlin, Czech Republic.
Lennon, J. J. 2000. Red-shifts and red herrings in geographicalecology. Ecography 23:101–113.
Lundberg, S. 1995. Catalogus Coleopterum Sueciae. [InSwedish.] Naturhistoriska riksmuseet, Stockholm, Sweden.
Marshall, P. L., G. Davis, and V. M. LeMay. 2000. Using lineintersect sampling for coarse woody debris. Forest ReserchTechnical Report, Vancouver Forest Region, Canada, TR-003, Ecology. Forest Service, British Columbia, Canada.
Moilanen, A., A. T. Smith, and I. Hanski. 1998. Long-termdynamics in a metapopulation of the American pika.American Naturalist 152:530–542.
National Board of Forestry. 2004. Statistical yearbook offorestry 2004: official statistics of Sweden. National Board ofForestry, Jonkoping, Sweden.
Nilsson, S. G., and R. Baranowski. 1996. Changes in theSwedish distribution of click beetles (Elateridae) occurring inthe boreal forest. [In Swedish with English summary.]Entomologisk Tidskrift 117:87–101.
Ovaskainen, O., and I. Hanski. 2002. Transient dynamics inmetapopulation response to perturbation. Theoretical Pop-ulation Biology 61:285–295.
Palm, T. 1946. Coleopterfaunan i en jamtlandsk lavgranskog:traden och svampfaunan. [In Swedish.] Entomologisk Tid-skrift 67:109–139.
Palm, T. 1951. Die Holz- und Rinden-Kafer der nordschwe-dishen Laubbaume. [In German.] Meddelanden fran StatensSkogsforskningsinstitut 40:1–242.
Palm, T. 1959. Die Holz und Rinden-kafer der sud undmittelschwedischen Laubbaume. [In German.] OpusculaEntomologica Supplementum XVI:1–347.
Penttila, R., J. Siitonen, and M. Kuusinen. 2004. Polyporediversity in managed and old-growth boreal Picea abiesforests in southern Finland. Biological Conservation 117:271–283.
Ranius, T., O. Kindvall, N. Kruys, and B. G. Jonsson. 2003.Modelling dead wood in Norway spruce stands subject todifferent management regimes. Forest Ecology and Manage-ment 182:13–29.
Saalas, U. 1917. Die Fichtenkafer Finnlands I. SuomalaisenTiedekatemian Kustantama, Helsinki, Finland. [In German.]
Schroeder, L. M., T. Ranius, B. Ekbom, and S. Larsson. 2006.Recruitment of saproxylic beetles in high stumps created formaintaining biodiversity in a boreal forest landscape.Canadian Journal of Forest Research 36:2168–2178.
Siitonen, J., and L. Saaristo. 2000. Habitat requirements andconservation of Pytho kolwensis, a beetle species of old-growth boreal forest. Biological Conservation 94:211–220.
Snall, T., A. Hagstrom, J. Rudolphi, and H. Rydin. 2004.Distribution pattern of the epiphyte Neckera pennata onthree spatial scales: importance of past landscape structure,connectivity and local conditions. Ecography 27:757–766.
Snall, T., J. Pennanen, L. Kivisto, and I. Hanski. 2005.Modelling epiphyte metapopulation dynamics in a dynamicforest landscape. Oikos 109:209–222.
Stelter, C., M. Reich, V. Grimm, and C. Wissel. 1997.Modelling persistence in dynamic landscapes: lessons froma metapopulation of the grasshopper Bryodema tuberculata.Journal of Animal Ecology 66:508–518.
Thomas, C. D. 1994. Extinction, colonisation and metapopu-lations: environmental tracking by rare species. ConservationBiology 8:373–378.
Tilman, D., R. M. May, C. L. Lehman, and M. A. Nowak.1994. Habitat destruction and the extinction debt. Nature371:65–66.
Verheyen, K., M. Vellend, H. van Calster, G. Peterken, and M.Hermy. 2004. Metapopulation dynamics in changing land-scapes: a new spatially realistic model for forest plants.Ecology 85:3302–3312.
Wahlberg, N., T. Klemetti, and I. Hanski. 2002. Dynamicpopulations in a dynamic landscape: the metapopulationstructure of the marsh fritillary butterfly. Ecography 25:224–232.
April 2007 909SPATIAL OCCURRENCE OF A BEETLE