
spatial learning cells that code for space synaptic plasticity in the hippocampus
40
spatial learning cells that code for space synaptic plasticity in the hippocampus experiments that are knockouts summary PART 4: BEHAVIORAL PLASTICITY #25: SPATIAL NAVIGATION IN RATS II
description
PART 4: BEHAVIORAL PLASTICITY #25: SPATIAL NAVIGATION IN RATS II. spatial learning cells that code for space synaptic plasticity in the hippocampus experiments that are knockouts summary. PART 4: BEHAVIORAL PLASTICITY #26: SPATIAL NAVIGATION IN RATS II. spatial learning - PowerPoint PPT Presentation
Transcript of spatial learning cells that code for space synaptic plasticity in the hippocampus

spatial learning cells that code for space synaptic plasticity in the hippocampus experiments that are knockouts summary
PART 4: BEHAVIORAL PLASTICITY#25: SPATIAL NAVIGATION IN RATS II

spatial learning cells that code for space synaptic plasticity in the hippocampus experiments that are knockouts summary
PART 4: BEHAVIORAL PLASTICITY#26: SPATIAL NAVIGATION IN RATS II

CODING SPACE – HIPPOCAMPAL PLACE CELLS
place cells encode more than simple space T-maze, trained (fruit loops) to alternate L & R turns subset of place cells showed interesting pattern e.g., activity (sector 3) anticipating right turns only
suggests hippocampal network represents episodic memories, cells are small segments of an episode link of cells with overlapping episodes memories
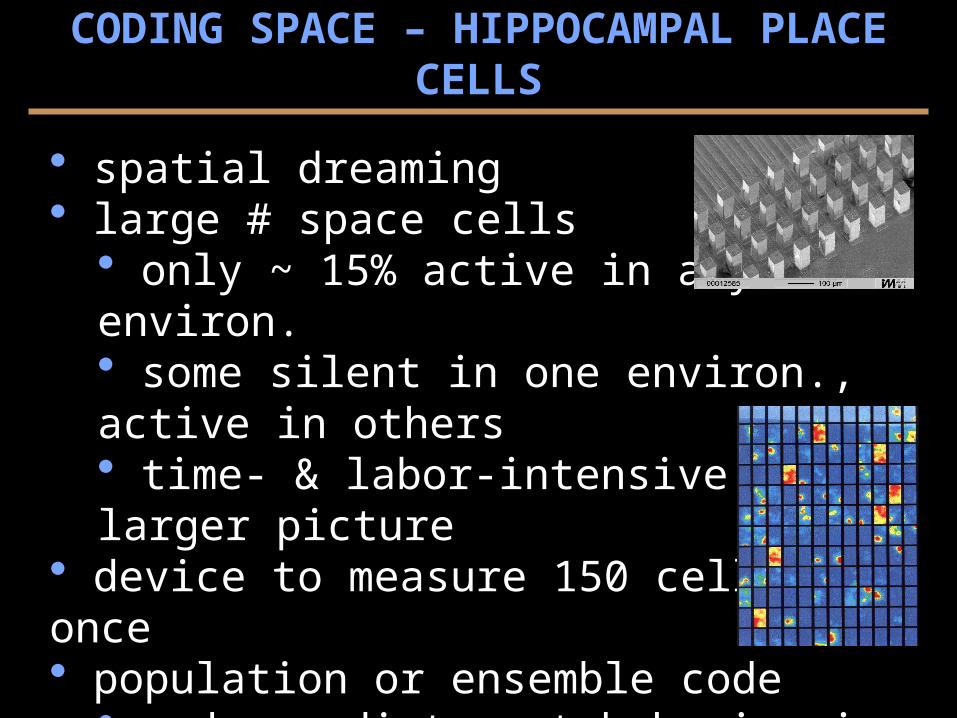
spatial dreaming large # space cells
only ~ 15% active in any 1 environ. some silent in one environ., active in others time- & labor-intensive to get larger picture
device to measure 150 cells at once population or ensemble code
code predicts rat behavior in maze many environments & codes overlapping, not interfering
used to study plasticity...
CODING SPACE – HIPPOCAMPAL PLACE CELLS
spatial dreaming plasticity
strengthening of code learning accompanied by reduced inhibitory activity
does code relate to consolidated (permanent) memory trained rats in spatial task measured code during
training sleeping before training sleeping after training
dreaming replay of events memory consolidation
CODING SPACE – HIPPOCAMPAL PLACE CELLS

CODING SPACE – HEAD DIRECTION CELLS
navigation requires knowledge of place direction... another class of cells...
in another structure... postsubiculum cells fire ~ head position

basic features of head direction cells retain direction preference in novel environments ~ 90° arc around preferred direction populations of cells with different preferences not ~ rat position in environment ~ independent of rat’s own behavior
CODING SPACE – HEAD DIRECTION CELLS

common features of head direction cells & place cells influenced by salient external cues direction cells also fire after cues (light) removed capable of deduced reckoning using ideothetic cues informed by vestibular and visual input direction cells do not remap in a novel environments
CODING SPACE – HEAD DIRECTION CELLS
navigation involves computation by the brain temporal process (~ video vs photograph) memory of past events prediction of future events processed by sub-populations of head direction cells 2 areas measured in behaving rats
postsubicular cortex (PSC) anterodorsal nucleus (ADN) of thalamus
CODING SPACE – HEAD DIRECTION CELLS

navigation involves computation by the brain analyzed firing pattern relative to
momentary head direction both cell types have preferred direction
CODING SPACE – HEAD DIRECTION CELLS

navigation involves computation by the brain analyzed firing pattern relative to
angular velocity PSC retain preference ADN shift preference future position
CODING SPACE – HEAD DIRECTION CELLS
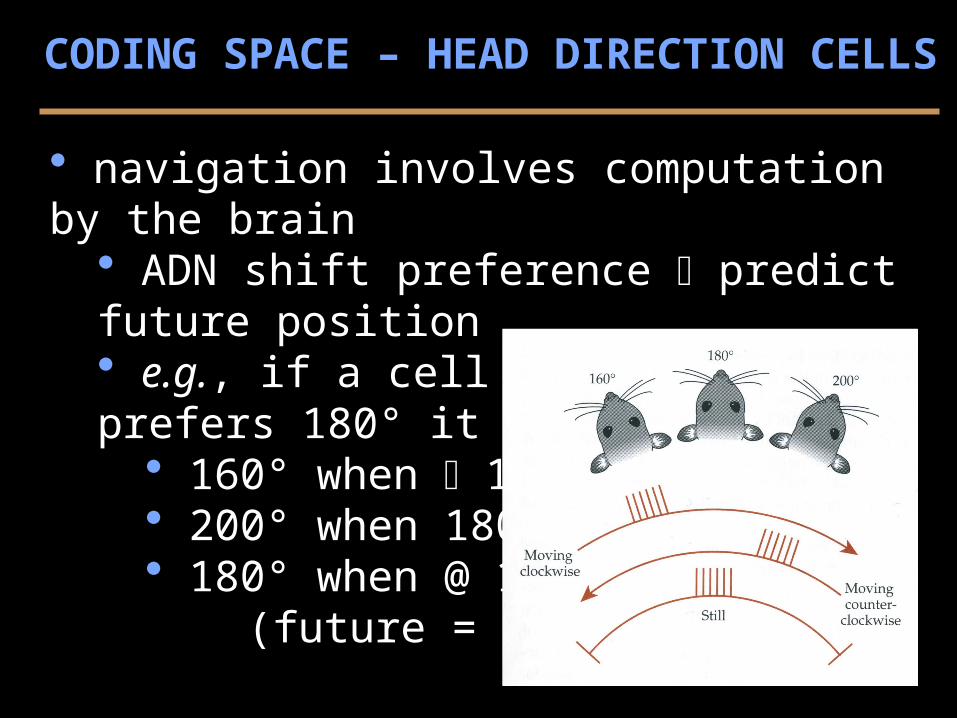
navigation involves computation by the brain ADN shift preference predict future position e.g., if a cell (of many) prefers 180° it may fire @
160° when 180° 200° when 180° 180° when @ 180° (future = present)
CODING SPACE – HEAD DIRECTION CELLS

why bother with all of this?... in theory... deductive reckoning circuit direction cells work by integrating internal cues ADN cells combine information about
current head direction head movement (turning)
proposed that PSC & ADN cells... constitute a looping circuit, compute direction by integrating motion/time but... how is “time” measured?
CODING SPACE – HEAD DIRECTION CELLS

SYNATPTIC PLASTICITY IN THE HIPPOCAMPUS
how do place cells and head directions cells learn to change their preferences? maintain their preferences over time?
clues from electrophysiology experiments... brief, high-frequency stimulation of trisynaptic circuit...
all 3 pathways
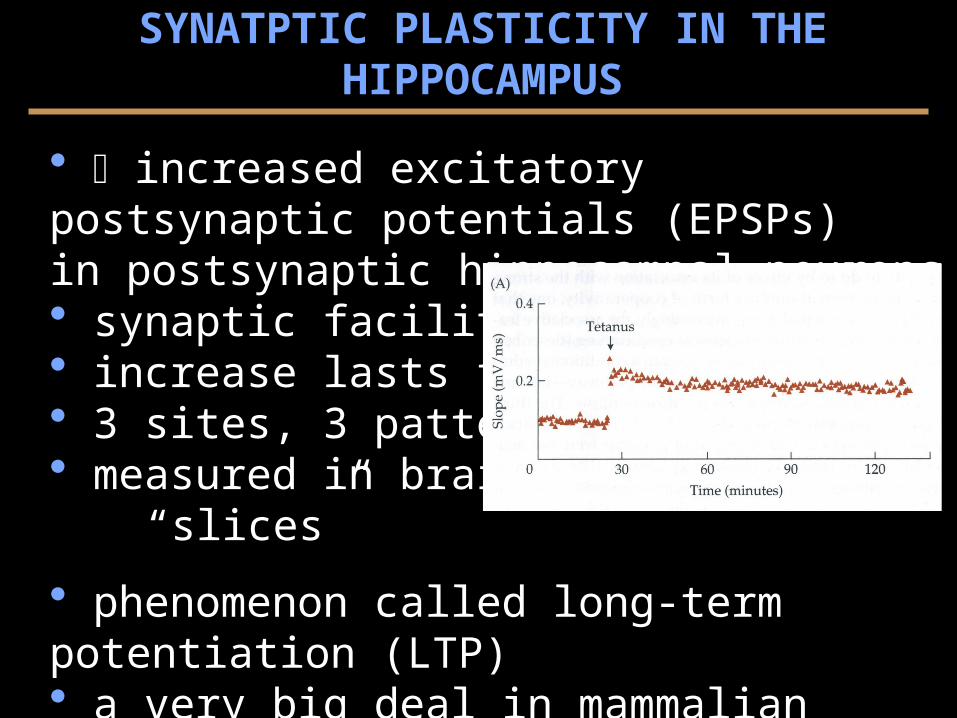
increased excitatory postsynaptic potentials (EPSPs) in postsynaptic hippocampal neurons
synaptic facilitation increase lasts for hours 3 sites, 3 patterns, CA1 measured in brain
“slices”
phenomenon called long-term potentiation (LTP) a very big deal in mammalian cell.-phys. of learning but... difficult to demonstrate relevance for behavior
SYNATPTIC PLASTICITY IN THE HIPPOCAMPUS
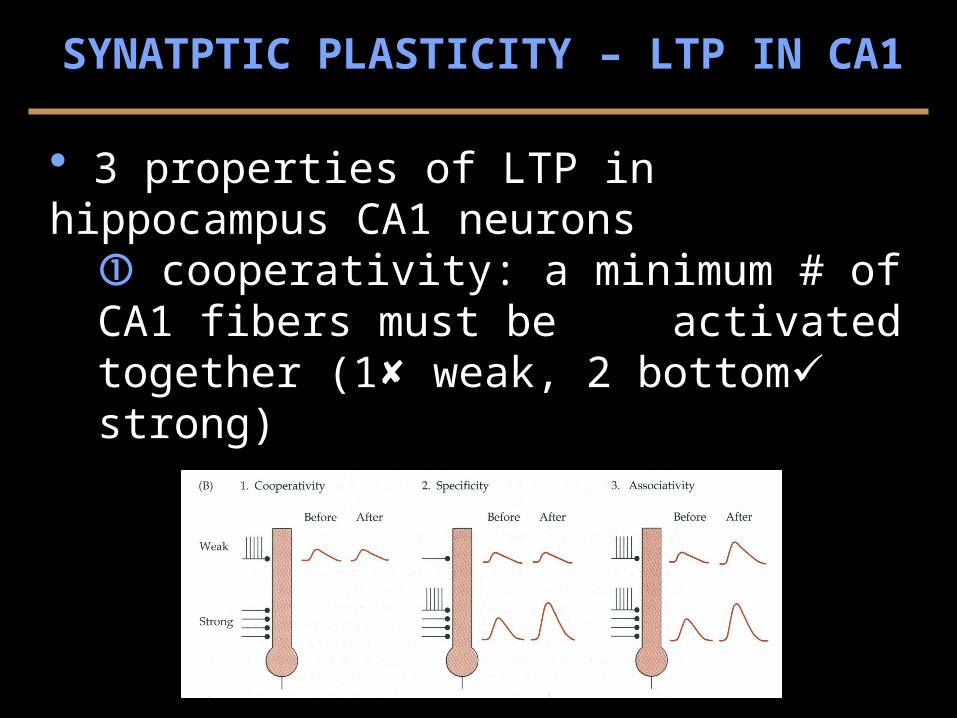
3 properties of LTP in hippocampus CA1 neurons cooperativity: a minimum # of CA1 fibers must be
activated together (1 weak, 2 bottom strong)
SYNATPTIC PLASTICITY – LTP IN CA1

3 properties of LTP in hippocampus CA1 neurons associativity: a weak tetanus paired with a strong
will gain - by association - value of strong measured in response after “training” (3 top) features ~ behavior, associative learning
SYNATPTIC PLASTICITY – LTP IN CA1
SYNATPTIC PLASTICITY – LTP IN CA1
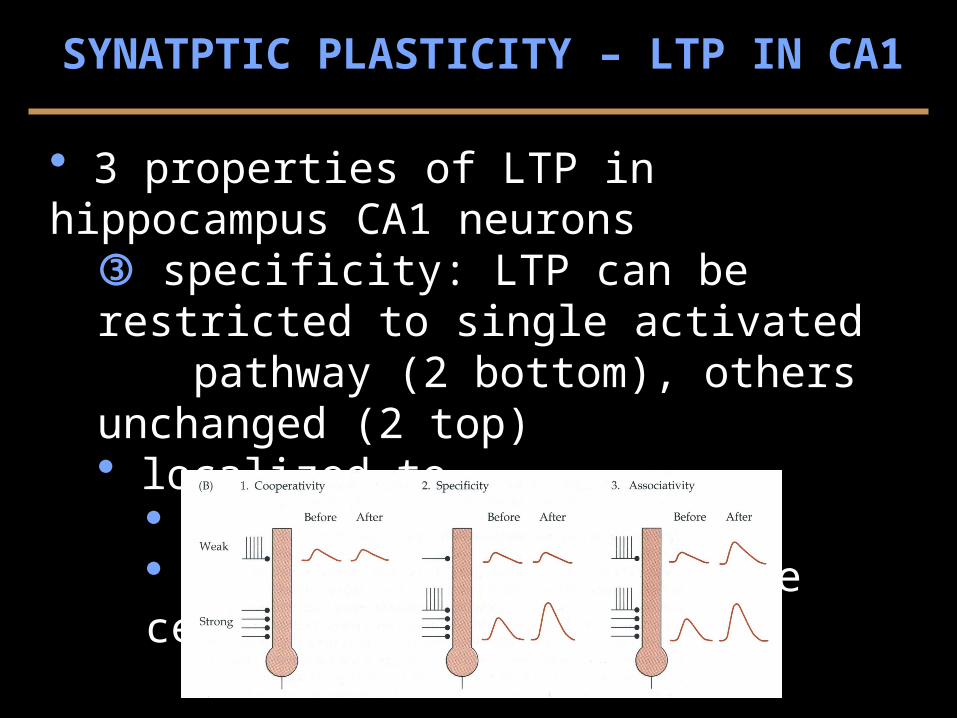
3 properties of LTP in hippocampus CA1 neurons specificity: LTP can be restricted to single activated
pathway (2 bottom), others unchanged (2 top) localized to
regions of hippocampus inputs regions on single cells (2)

SYNATPTIC PLASTICITY – LTP IN POSTSYNAPTIC CELLS
CA1 pyramidal neurons LTP in CA1 is dependent on pyramidal neurons (PNs)
inhibition of PN activity blocks LTP in CA1 hyperpolarize PN membrane blocks LTP in CA1 blocked inhibition of PN facilitates LTP in CA1 depolarize PN membrane
facilitates LTP in CA1 during weak tetanus not on its own (i.e., effect is associative)
the postsynaptic cell must be depolarized for LTP to occur in the presynaptic cell

SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS
glutamate (GLU), main excitatory transmitter (brain) N-methyl-D-aspartate (NMDA) 1 (of many) receptors LTP requires depolarization to open NMDA channel
doubly gated channel, by.. GLU (receptor) & voltage (sensor)

evidence for NMDA involvement in LTP NMDA blockers, e.g. aminophosphnovalerate (APV)
blocks NMDA activity blocks LTP
cooperativity: GLU from weak input depolarize postsynaptic cell strong input depolarizes postsynaptic cell
associativity: GLU from strong input depolarizes postsynaptic cell weak input (paired) opens NMDA channels*
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

evidence for NMDA involvement in LTP Hebb’s Rule: synapses are strengthened if a presynaptic cell repeatedly participates in driving spikes in a postsynaptic cell GLU & NMDA receptor satisfies the rule have coincident activity of cells
presynaptic release of GLU receptors postsynaptic depolarization by non-NMDA receptors
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

Ca++ influx into the postsynaptic cell is required for LTP block calcium (buffer)
blocks LTP calcium influx through NMDA receptor/channel
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS
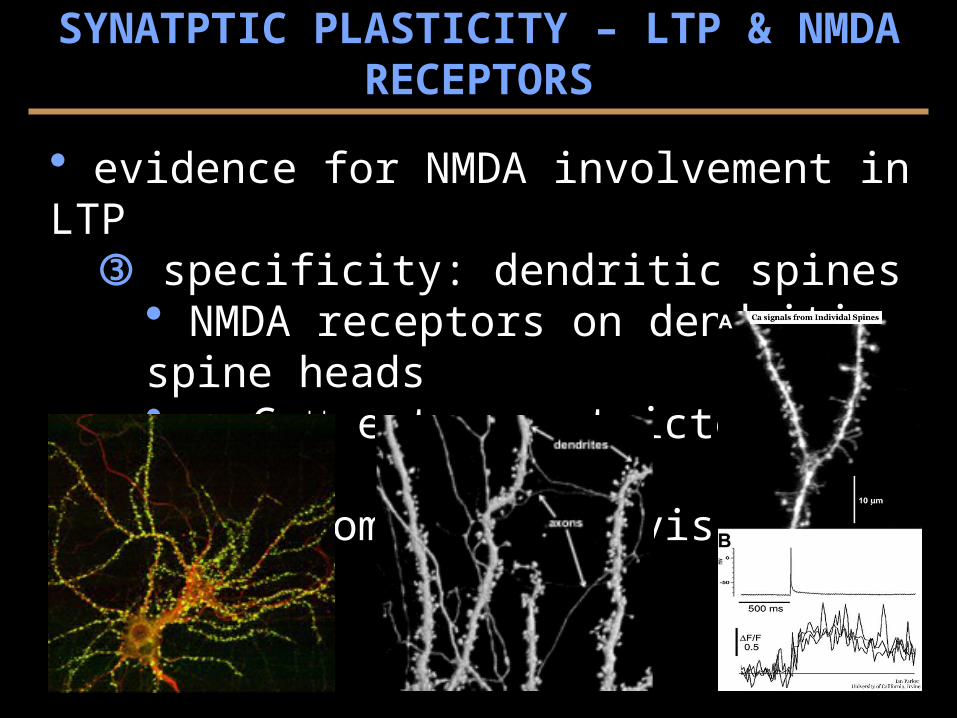
evidence for NMDA involvement in LTP specificity: dendritic spines
NMDA receptors on dendritic spine heads Ca++ entry restricted by necks anatomical subdivisions
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

evidence for NMDA involvement in LTP specificity: dendritic spines
NMDA receptors on dendritic spine heads Ca++ entry restricted by necks anatomical subdivisions
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

Ca++ influx into the postsynaptic cell is required for LTP Ca++ LTP mediated by 2nd messenger signaling
Ca++/calmodulin kinase (CaMKII) protein kinase C (PKC)
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

2 types of LTP described in CA1 neurons early-phase LTP (E-LTP)
1 3 h cAMP & protein synthesis-independent
late-phase LTP (L-LTP) 10 h + cAMP & protein synthesis-dependent
LTP in rats ~ long-term synaptic facilitation in Aplysia long-term memory in Drosophila
SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS

SYNATPTIC PLASTICITY – LTP & NMDA RECEPTORS
2 types of LTP described in CA1 neurons early-phase LTP (E-LTP)
1 3 h cAMP & protein synthesis-independent
late-phase LTP (L-LTP) 10 h + cAMP & protein synthesis-dependent
LTP in rats ~ long-term synaptic facilitation in Aplysia long-term memory in Drosophila

SYNATPTIC PLASTICITY – LTP & SPATIAL LEARNING
does LTP have anything to do with learning?... difficult spatial learning & memory in the water maze
block LTP with AP5 block memory ask the 3 Qs...
correlation? necessity? sufficiency?
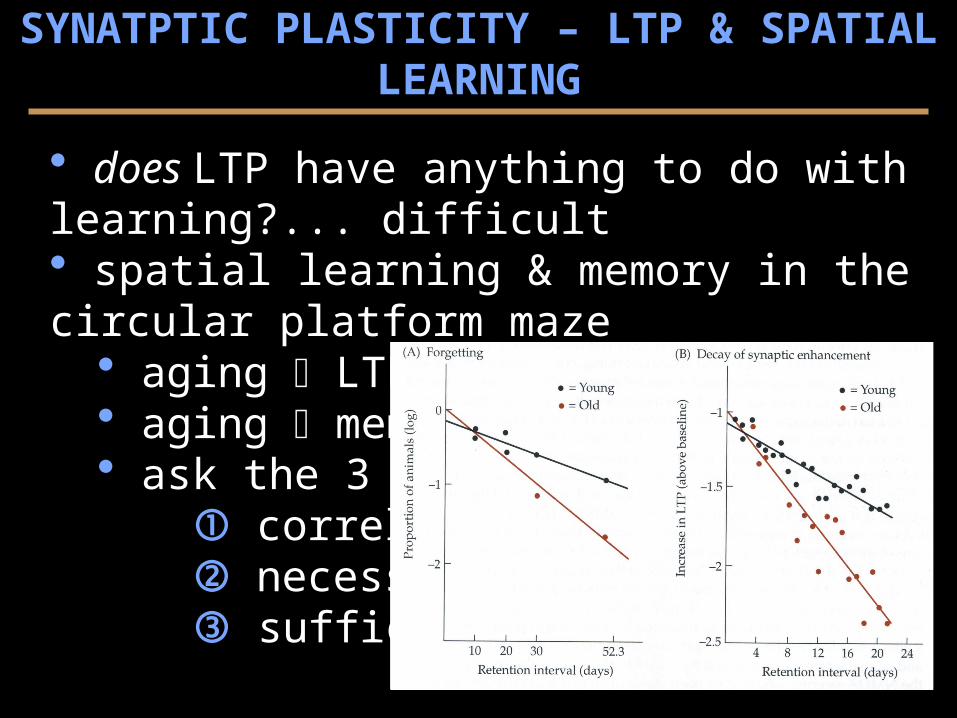
does LTP have anything to do with learning?... difficult spatial learning & memory in the circular platform maze
aging LTP ~ aging memory ask the 3 Qs...
correlation? necessity? sufficiency?
SYNATPTIC PLASTICITY – LTP & SPATIAL LEARNING

EXPERIMENTS THAT ARE KNOCKOUTS (MOUSE)
genetic engineering - e.g. already with Drosophila transgenic “knockouts” (also “knockins”) single gene manipulations LTP & spatial learning fyn gene knockout are tyrosine kinase– and... knockouts of CaMKII–
LTP in CA1 cells spatial learning ask the 3 Qs...
correlation? necessity? sufficiency?
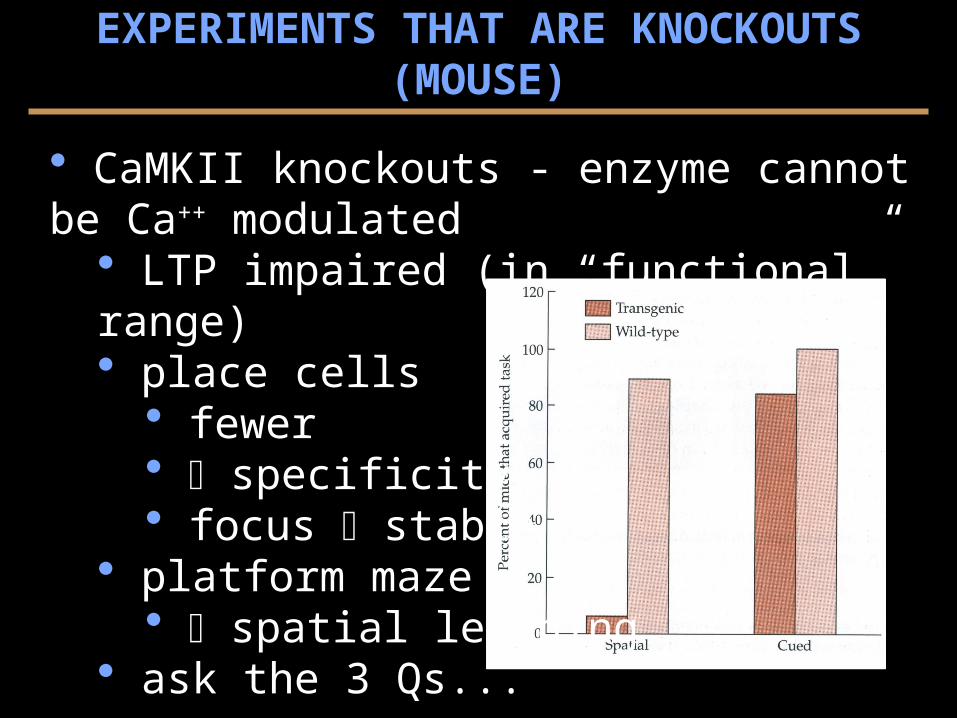
CaMKII knockouts - enzyme cannot be Ca++ modulated LTP impaired (in “functional” range) place cells
fewer specificity focus stable
platform maze spatial learning
ask the 3 Qs...
EXPERIMENTS THAT ARE KNOCKOUTS (MOUSE)
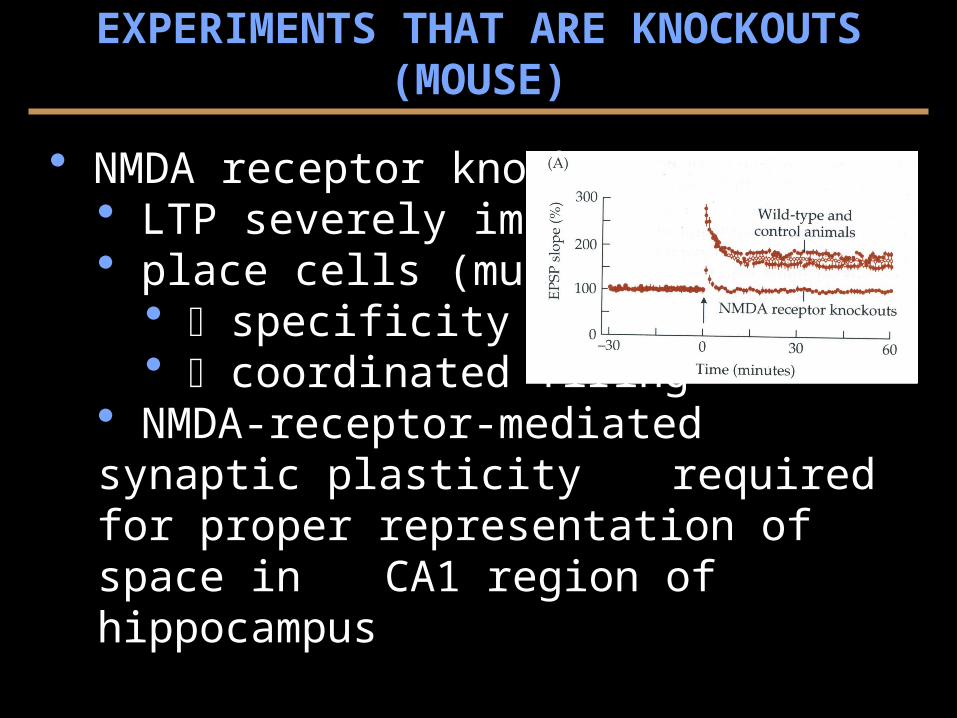
NMDA receptor knockouts LTP severely impaired place cells (multi-elect.)
specificity coordinated firing
NMDA-receptor-mediated synaptic plasticity required for proper representation of space in CA1 region of hippocampus
EXPERIMENTS THAT ARE KNOCKOUTS (MOUSE)

NMDA receptor knockouts water maze
spatial learning
ask the 3 Qs... arguments more compelling with each experiment spatial & temporal targeting of knockout, correlation of lesion, LTP, behavior remains
EXPERIMENTS THAT ARE KNOCKOUTS (MOUSE)

SUMMARY spatial navigation uses 2 types of cues
external (landmarks) internal (ideothetic) deductive reckoning (memory)
spatial navigation studied in rats using radial arm maze T-maze water maze circular platform maze

tasks are designated as spatial (using distal cues) cued (or non-spatial, using proximal cues)
lesion studies, hippocampus for spatial learning if lesions precede learning
working & reference memory tasks are impaired cued tasks are not impaired
if learning precedes lesions time between events important usually older memories are less affected
SUMMARY

two classes of neurons encode space place cells, CA1 hippocampus
firing field stability ~ weeks, memory influenced by
external cues (landmarks) internal cues (vestibular, visual ~ motion)
field in dark ~ active can be event-related, predictive (e.g., turning) work together ensemble code replay in sleep... consolidation?... dreaming?
SUMMARY

two classes of neurons encode space head direction cells, CA1 hippocampus
fire ~ head direction similarly influenced by
external cues (landmarks) internal cues (vestibular, visual ~ motion)
2 types of cells PSC cells encode current direction ADN cells encode future direction
SUMMARY

LTP is a prominent form of hippocampal synaptic plasticity, with the following properties:
cooperativity associativity specificity
LTP in CA1 neurons ~ NMDA receptor, 2 requirements: depolarization of the postsynaptic cell binding of glutamate with the NMDA receptor allows channel opening, Na+ & Ca++ influx Ca++ influx is required for induction of LTP
SUMMARY

NMDA receptor mechanism for Hebb’s Rule
Evidence that LTP underlies (or is involved with) mechanisms for learning
drugs blocking LTP also block spatial learning aging affects LTP and spatial learning mice knockouts for “LTP genes” show deficits in
LTP place cell properties spatial learning
SUMMARY


















