
Sources and diagenetic status of organic matter in the ... Sikes et al OGC 07.pdf · Sources and...
18
Sources and diagenetic status of organic matter in the Hauraki Gulf, New Zealand: Evidence from the carbon isotopic composition of D- and L-amino acids Maria E. Uhle a , Elisabeth L. Sikes b, * , Scott D. Nodder c , Conrad A. Pilditch d a Department of Earth and Planetary Sciences, University of Tennessee, 1412 Circle Drive, Knoxville, TN 37996, USA b Institute of Marine and Coastal Sciences, Rutgers University, 71 Dudley Road, New Brunswick, NJ 08901, USA c National Institute of Water and Atmospheric Research, Private Bag 14901, Kilbirnie, Wellington, New Zealand d Department of Biological Sciences, University of Waikato, Private Bag 3105, Hamilton, New Zealand Received in revised form 22 December 2005; accepted 1 June 2006 Available online 14 August 2006 Abstract The origin of sedimentary organic matter and the influence of microbial reworking in the Hauraki Gulf, New Zealand, were determined using the stable carbon isotopic compositions of sedimentary D- and L-amino acids. Core top sediments from three offshore and five inner Gulf sites were collected using a multi-corer. After correction for microbial alteration of original source materials, source apportionment of L-amino acids indicated that the organic matter at the outer Gulf sites was predominantly derived from a marine source, whereas the inner Gulf sites reflected a terrestrial source. The isotopic compositions of D-amino acids were consistent with preferential microbial utilization of marine organic matter in the outer Gulf and preferential utilization of terrestrial material in the inner Gulf. Microbial reworking was most prevalent at the outer Gulf sites where up to 58% of the total hydrolyzable amino acids (THAA) were derived from microbial synthesis. Although the inner Gulf sites contained relatively less reworked material, the microbial contribution to the THAA pool ranged from 40 to 50%. This study showed that the sediment carbon dynamics in the Hauraki Gulf are complex as micro- bial reworking influences the isotopic compositions of the sedimentary organic matter throughout the Hauraki Gulf. Ó 2006 Elsevier Ltd. All rights reserved. 1. Introduction Early diagenesis in marine sediments is generally thought to entail the preferential loss of labile organic compounds such as amino acids, owing to a series of chemical and microbially mediated reac- tions. Amino acids are ubiquitous in continental margin sediments and their distribution in marine sediments can yield information about the degree to which the original organic matter has undergone decay (Dauwe et al., 1999; Lee et al., 2000). Whereas the loss of amino acids is consistent with early diagenesis, recent studies have shown that dia- genesis also entails synthesis of these labile com- pounds through the production of microbial biomass (Macko et al., 1994; Keil and Fogel, 2001). The microbial production of amino acids can alter the organic matter composition of marine 0146-6380/$ - see front matter Ó 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.orggeochem.2006.06.007 * Corresponding author. Tel.: +1 732 932 6555; fax: +1 732 932 8578. E-mail address: [email protected] (E.L. Sikes). Organic Geochemistry 38 (2007) 440–457 www.elsevier.com/locate/orggeochem Organic Geochemistry
Transcript of Sources and diagenetic status of organic matter in the ... Sikes et al OGC 07.pdf · Sources and...

Organic Geochemistry 38 (2007) 440–457
www.elsevier.com/locate/orggeochem
OrganicGeochemistry
Sources and diagenetic status of organic matter in theHauraki Gulf, New Zealand: Evidence from the carbon
isotopic composition of D- and L-amino acids
Maria E. Uhle a, Elisabeth L. Sikes b,*, Scott D. Nodder c, Conrad A. Pilditch d
a Department of Earth and Planetary Sciences, University of Tennessee, 1412 Circle Drive, Knoxville, TN 37996, USAb Institute of Marine and Coastal Sciences, Rutgers University, 71 Dudley Road, New Brunswick, NJ 08901, USAc National Institute of Water and Atmospheric Research, Private Bag 14901, Kilbirnie, Wellington, New Zealand
d Department of Biological Sciences, University of Waikato, Private Bag 3105, Hamilton, New Zealand
Received in revised form 22 December 2005; accepted 1 June 2006Available online 14 August 2006
Abstract
The origin of sedimentary organic matter and the influence of microbial reworking in the Hauraki Gulf, New Zealand,were determined using the stable carbon isotopic compositions of sedimentary D- and L-amino acids. Core top sedimentsfrom three offshore and five inner Gulf sites were collected using a multi-corer. After correction for microbial alteration oforiginal source materials, source apportionment of L-amino acids indicated that the organic matter at the outer Gulf siteswas predominantly derived from a marine source, whereas the inner Gulf sites reflected a terrestrial source. The isotopiccompositions of D-amino acids were consistent with preferential microbial utilization of marine organic matter in the outerGulf and preferential utilization of terrestrial material in the inner Gulf. Microbial reworking was most prevalent at theouter Gulf sites where up to 58% of the total hydrolyzable amino acids (THAA) were derived from microbial synthesis.Although the inner Gulf sites contained relatively less reworked material, the microbial contribution to the THAA poolranged from 40 to 50%. This study showed that the sediment carbon dynamics in the Hauraki Gulf are complex as micro-bial reworking influences the isotopic compositions of the sedimentary organic matter throughout the Hauraki Gulf.� 2006 Elsevier Ltd. All rights reserved.
1. Introduction
Early diagenesis in marine sediments is generallythought to entail the preferential loss of labileorganic compounds such as amino acids, owing toa series of chemical and microbially mediated reac-tions. Amino acids are ubiquitous in continental
0146-6380/$ - see front matter � 2006 Elsevier Ltd. All rights reserveddoi:10.1016/j.orggeochem.2006.06.007
* Corresponding author. Tel.: +1 732 932 6555; fax: +1 732 9328578.
E-mail address: [email protected] (E.L. Sikes).
margin sediments and their distribution in marinesediments can yield information about the degreeto which the original organic matter has undergonedecay (Dauwe et al., 1999; Lee et al., 2000).Whereas the loss of amino acids is consistent withearly diagenesis, recent studies have shown that dia-genesis also entails synthesis of these labile com-pounds through the production of microbialbiomass (Macko et al., 1994; Keil and Fogel,2001). The microbial production of amino acidscan alter the organic matter composition of marine
.

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 441
sediments masking other diagenetic changes in theorganic matter. To assess the influence of microbialnew production on the fate of organic matter inmarine sediments, it is necessary to distinguish com-pounds derived from bacterial production fromthose found in the original source materials.
Bacterial communities produce many of the samecompounds found in the original organic matter,making source identification and apportionment-based solely on chemical structure, difficult. The chi-ral nature of amino acids, however, can help assessmicrobial reworking of sedimentary organic matter.Living biomass consists primarily of L-amino acidsand the presence of D-stereoisomers in marine sedi-ments is attributed to either the racemization of theL-amino acids to their D-equivalents (e.g. Badaet al., 1970) or to the formation D-amino acids asfound in bacterial peptidoglycan (Walsh, 1989;Pedersen et al., 2001; Jorgensen et al., 2003).
Peptidoglycan is the primary building block ofeubacterial cell walls (Koch, 1990) and its structureconsists of b-1,4 linked N-acetylglucosamine and N-acetylmuramic acid strands cross-linked by peptidebridges (Schleifer and Kandler, 1972; Koch, 1990).Three of the five amino acids in the peptide bridgesare D-amino acids. Whereas it may be expected thatbacterial cells undergo diagenesis quite rapidly, thestructure of peptidoglycan appears to render thestructure more resistant to degradation. Empty bac-terial cells have been observed in marine sediments(Moriarty and Hayward, 1982) and peptidoglycanhas been associated with 4000–6000 year old, highmolecular weight, dissolved organic matter in oceanwaters indicating its refractory nature (McCarthyet al., 1998).
The presence of D-amino acids in marine sedi-ments may be linked to the peptidoglycan in the cellwalls of Gram-negative (G�) bacteria owing to theirpredominance in both oxic and anoxic marine envi-ronments (Moriarty and Hayward, 1982; Hagstromet al., 2000). The highest concentration of D-stereo-isomers in a sediment core from Aarhus Bay, Den-mark was observed at 10 cm depth (Pedersenet al., 2001), which was coincident with the maxi-mum bacterial density in these sediments (Jorgensenet al., 1990). While the D-alanine (D-Ala) and D-glu-tamic acid (D-Glu) dominate the peptide bridges ofpeptidoglycan derived from both Escherichia coli
(Gram-negative (G�) bacteria) and Bascillus subtilis
(Gram-positive (G+) bacteria), the principal aminoacid components of the peptide interbridges varyamong bacterial strains. The peptide bridges of
other bacterial strains may include D-aspartic acid(D-Asp), D-leucine (D-Leu), and D-serine (D-Ser)(Pedersen et al., 2001; Jorgensen et al., 2003).
Non-protein amino acids such as b-alanine (b-Ala) and c-aminobutyric acid (c-Aba) may beformed from microbial decarboxylation of proteinamino acids (Lee and Cronin, 1982) and indicatemicrobial reworking of organic matter in sediments.Increases in the percentage of non-protein aminoacids relative to their protein amino acid precursorshave been used as an indicator of microbial metab-olism of sedimentary organic matter in several mar-ine settings, including the Peru upwelling system,Saanich Inlet and the North Sea (Lee and Cronin,1982; Dauwe and Middelburg, 1998; Cowie andHedges, 1992).
In addition to concentrations of non-protein andD-amino acids, the isotopic composition of individ-ual amino acids may provide a way to distinguishnew microbial production of amino acids fromthose derived from marine and terrestrial sources.Compound specific isotope analysis (CSIA) can alsohelp determine the source of organic matter utilizedby microbial communities in the sediments. Recentwork monitoring the isotopic composition of aminoacids along a transect from the mouth of theColumbia River to the waters of the Washingtoncoast, showed that most of the terrigenous aminoacids were replaced by their marine equivalentsand that microbial reworking further altered thed13C values of amino acids in the offshore sediments(Keil and Fogel, 2001). The d13C values of certainamino acids (proline (Pro) and Leu) reflected asmuch as 80% replacement by marine derived aminoacids. The isotopic compositions of other aminoacids, such as Ala, isoleucine (Ile), Glu, Asp, andphenylalanine (Phe), were consistent with microbialresynthesis using marine substrates and/or preserva-tion of approximately 50% of the original terrige-nous material.
The focus of our present study was to identify thesedimentary organic matter sources and assess theinfluence of microbial reworking on the organicmatter in coastal sediments through the stable car-bon isotopic composition of D- and L-amino acids.We assumed that the carbon isotopic compositionof D-amino acids reflects the organic matter sourceutilized by bacterial communities. Keil and Fogel(2001) utilized the d13C values of amino acids toindicate the relative fates of terrigenous and mar-ine-derived organic matter in coastal environ-ments. That study, however, did not resolve the

442 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
amino acid enantiomers and assessment of micro-bial carbon sources was done indirectly. Here, webuild on previous work and identify organic mattersources utilized by microbial communities throughstereoisomer distributions and isotopic composi-tions of hydrolyzable sedimentary D- and L-aminoacids.
2. Methods
2.1. Study area and oceanographic setting
The Hauraki Gulf is a shallow, semi-enclosedshelf sea located on the northeast coast of the NorthIsland, New Zealand (Fig. 1). The continental shelfin this region is fairly narrow, typically <40 km, butthe width of the shelf increases to �80 km in theHauraki area (Manighetti and Carter, 1999). Sea-floor gradients in the central Gulf are as low as1:2000, steepening in the outer Gulf and reaching1:400 towards the shelf-break at approximately150 m water depth (Manighetti and Carter, 1999).Sediments form a heterogeneous mosaic of differenttypes across the shelf, ranging from sandy muds atthe shelf-break and outer shelf to sandy sediments,
Fig. 1. Study area. Locations of sampling stations are indicated by blHauraki Gulf are driven by residual tidal flows augmented by regionalgrey arrows. Whangerei is a sizeable regional city and river as well aAuckland city is just south of the Whangaparoa peninsula. Site 14 (FoT)rivers enter the Firth at the southernmost end.
containing varying proportions of calcareous sandand gravel and terrigenous mud, on the inner andmid-shelf (Carter and Eade, 1980; Manighetti andCarter, 1999). Located between the northern tip ofCoromandel Peninsula and Great Barrier Island,Colville Channel is floored by very coarse, calcare-ous sediments, as are Jellicoe and Craddock chan-nels to the north on the mid-shelf. The Firth ofThames is dominated by fine-grained terrigenousdeposition (Carter and Eade, 1980).
The Gulf is bounded to the south by the HaurakiPlains, to the east by the Coromandel Peninsula andGreat Barrier Island and to the west by the Auck-land metropolitan area (Fig. 1). The total riverineinflow to the inner Gulf is about 160 m3 s�1, whichis delivered primarily to the southeastern Gulf viathe Firth of Thames (100 m3 s�1) (Zeldis et al.,2004). The seafloor in the Firth of Thames regionshoals gradually from 35 m at the northern entranceof the Firth, where sediments are predominantlyshelly sandy silts, to swampy, estuarine mudflatsat a gradient of 1:1500 (Manighetti and Carter,1999). The Waihou and Piako rivers drain the dairyareas of the Hauraki Plains and smaller, steepercatchments are located in native forest regions of
ack dots and designated by number. The general currents of theflow of the East Auckland Current and are indicated by the lights the site of the country’s only refinery. The sewage outfall forsits at the mouth of the Firth of Thames. The Waiohau and Piako

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 443
the Coromandel Peninsula. These rivers carry dis-solved inorganic phosphorous and nitrogen derivedfrom the pastoral catchments to the Firth ofThames (Waikato, 1998a,b) and have importanteffects on the flow of water, salt and nutrientsthrough the Firth of Thames and the inner HaurakiGulf (Zeldis and Smith, 1999; Zeldis et al., 2004).
On the western side of the Hauraki Gulf, thereare significant discharges of treated sewage effluent.The major source is at North Shore, Auckland,while other smaller discharges occur along the westcoast of the Gulf. Although the water volume ofsewage is small relative to other freshwater inputs,dissolved P and N are highly concentrated in theeffluent. Consequently, nutrient fluxes from sewageoutfalls are significant relative to other freshwaternutrient sources (Waikato, 1998a,b). Zeldis andSmith (1999) estimated that �30% of the dissolvedinorganic nitrogen load in the gulf was due to river-ine input mainly via the Firth of Thames, while�10% was due to Auckland sewerage effluent andthe remaining �60% from ocean-derived nutrients.
Subtropical waters are delivered to the Gulf andits shelf-slope system by the southeastward flowingEast Auckland Current, which passes along theouter shelf and dominates the overall flow in theregion (Fig. 1). Net circulation in the Gulf is fromnorth to south driven by residual tidal current flow(Sharples, 1997; Zeldis et al., 2004). In the innerGulf, where weaker current flows diverge, finergrained sediments are deposited at higher rates,and the total organic content is twice that of outershelf sites (�8%). Strong flows exiting the Gulf nearthe southern mouth are evident from the gravellypavement in the Colville Channel between GreatBarrier Island and the Coromandel Peninsula(Fig. 1) (Manighetti and Carter, 1999).
Seasonal upwelling occurs in the late winter toearly spring when westerly winds predominate.Upwelling in the outer Hauraki Gulf is believed tofuel phytoplankton productivity by delivering nutri-ents during these seasonal upwelling events. Phyto-plankton blooms occur in late winter and spring inthe outer Gulf and along the outer shelf (Changet al., 2003b; Zeldis, 2004). Upwelled nutrient-replete water occurs at the surface in the northernGulf, but to the south, in the inner Gulf, thesewaters are only present below the surface mixedlayer. Consequently, in the southern, shallow por-tions of the Gulf phytoplankton assemblages aredominated by dinoflagellates, which make betteruse of nutrients delivered at depth (Chang et al.,
2003b; Zeldis, 2004). The seasonal upwelling thatdrives blooms in the spring to early summer arereplaced by downwelling in the summer when thewinter westerlies shift to easterlies at which timethe Gulf becomes strongly stratified (Sharples,1997; Zeldis et al., 2004). At this time the Gulfchanges from a net autotrophic system to one moreclosely balanced between autotrophy and heterotro-phy (Chang et al., 2003b) and nutrients appear to bederived from remineralization in the lower watercolumn and sediments (Zeldis, 2004). Seasonally,the inner gulf and Firth of Thames show greaterproportions of remineralized nitrogen than theouter area, particularly when stratification is mostintense (Zeldis, 2004). Importantly, on an annualbasis, both the Gulf and Firth of Thames act asnet sinks for dissolved inorganic nitrogen, suggest-ing that denitrification by sediment microbial andheterotrophic communities is an important processin regulating the nitrogen load in these two environ-ments (Giles, 2001; Zeldis, 2004).
2.2. Sample collection
Sediments were collected using an Ocean Instru-ments MC-800 multi-corer during a cruise on theRV Tangaroa in December 1999 (TAN9915). Aftercollection, the sediment cores were sub-sampledimmediately at 0.5 and 1 cm intervals. Samples forthe determination of bulk sediment properties weresealed in zip-lock plastic bags while those for aminoacid analysis were placed in pre-cleaned glass jarsand both were stored frozen until analyzed. Sites 2and 11 are located along the shelf-break and serveas marine stations (Fig. 1). The other six sites arefairly close to shore with most of them locatedwithin the inner gulf. Site 1 is situated in an areaof intense spring upwelling and related productivity(Chang et al., 2003b; Zeldis et al., 2004). Sites 4 and8 are located near Whangarei and Whangaparoaharbors, respectively, which may be affected by ter-restrial run-off and sewage outfalls. Site 14 is locatedat the entrance to the Firth of Thames where signif-icant terrestrial inputs and nutrients are delivered tothe inner gulf from the Waihou and Piako rivers.Site 10 is located within Colville Channel betweenGreat Barrier Island and the Coromandel Penin-sula, which is a suspected outflow channel for innergulf waters (Manighetti and Carter, 1999). Site 7 islocated north of Craddock Channel on the northside of Great Barrier Island, where extensive man-grove forests thrive.

444 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
2.3. Bulk sediment characteristics
Grain-size parameters for the top 0–1 cm at eachsite were established on oven-dried sediments usingstandard sieve (% sand: 500–63 lm) and Sedigraphtechniques (% silt and clay: <63 lm fraction). Car-bonate content was measured volumetrically witha ±1% accuracy and total organic matter (TOM)was estimated by loss-on-ignition (500 �C for 4 h).Following acidification with 8% sulfurous acid (Ver-ardo et al., 1990), percent organic carbon and nitro-gen were determined by a CHN analyzer (CEInstruments NC2500) with 2% machine precision.
2.4. Bulk carbon and nitrogen isotopic analyses
The stable carbon and nitrogen isotopic compo-sitions of bulk sedimentary organic matter analyseswere determined by a ThermoQuest FinniganDelta-Plus mass spectrometer. Acidified sedimentsamples were combusted in an NA 1500 elementalanalyzer (Fisons Instruments). The isotopic compo-sitions were reported relative to PDB (13C) andatmospheric air (15N) standards via secondary labo-ratory standards. Precision of both d13C and d15N isbetter than 0.25&.
2.5. Amino acid isolation and characterization
Amino acids were extracted from lyophilized andground sediments through acid hydrolysis. Sedi-ments were placed in culture tubes, 6 N HCl wasadded to excess. The samples were sealed underN2 and digested at 100 �C for 24 h. The extractswere dried under N2 at 25 �C and the amino acidswere derivatized to their TFA isopropyl ester formfollowing the procedure described by Silfer et al.(1991), to facilitate analysis by gas chromatogra-phy–mass spectrometry (GC/MS) and gas chroma-tography–isotope ratio mass spectrometry (GC/IRMS). Briefly, the hydrolyzates were esterifiedwith acidified 2-propanol for 1 h at 110 �C. Thereaction was quenched and the solvent was removedby evaporation under a gentle stream of N2 at 25 �Cafter which CH2Cl2 was added and subsequentlyevaporated to remove excess 2-propanol and water.The amino acid isopropyl esters were acylated usingtrifluoroacetic anhydride (TFAA) and CH2Cl2 for10 min at 110 �C. The vials were then chilled andexcess TFAA and CH2Cl2 were removed at 0 �Cby evaporation under N2 in an ice bath. Glutamicand aspartic acids represent an unknown contribu-
tion from glutamine and asparagine, which are con-verted to the dicarboxylic acid during acidhydrolysis. These amino acids are reported as Glxand Asx, respectively.
The TFA isopropyl esters of the amino acidswere analyzed by GC/MS using a Hewlett-Packard6890 gas chromatograph interfaced to a HewlettPackard 5973 GC/MSD with helium as the carriergas. A J&W Scientific Chirasil-Val fused silica cap-illary column (50 m · 0.25 mm i.d.) was used toresolve the TFA isopropyl esters of amino acidswith the oven temperature programmed from45 �C for 3 min to 70 �C at 45 �C/min. The temper-ature was held isothermally at 70 �C for 15 min,increased from 70 �C to 190 �C at 2�C/min andfinally held at 190 �C for 45 min.
2.6. Compound-specific stable carbon isotope
analyses
The TFA isopropyl esters of the amino acidswere directly analyzed for their stable carbon isoto-pic compositions using a ThermoFinnigan GC/IRMS system. This system consists of a Hewlett-Packard 6890 gas chromatograph interfaced to aThermoFinnigan Deltaplus XL isotope ratio massspectrometer via a combustion furnace/nafion watertrap. The isotopic compositions of the amino acidTFA isopropyl esters were determined under thetemperature conditions and capillary columndescribed for GC/MS analysis.
2.7. Source apportionment model
The relative contributions of marine and terres-trial organic inputs to the sedimentary pool forthe D- and L-amino acids at each of the samplingsites was estimated based on a two end-membermixing model such that:
fmarine ¼d13CSedAA � d13Cterrestrial
d13Cmarine � d13Cterrestrial
ð1Þ
fmarine ¼ 1� fterrestrial ð2Þ
where d13CSedAA, d13Cterrestrial, and d13Cmarine arethe carbon isotopic compositions of the sedimentaryamino acid (either D or L), terrestrial and marineend-members, respectively, and where fmarine andfterrestrial are the fractions of marine and terrestrialderived organic matter. Unfortunately, no corre-sponding ‘‘pure’’ end-member samples were takenat the time of coring. Owing to the lack of marine

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 445
and terrestrial d13C end-member values for the indi-vidual amino acids derived from the organic mattersources in the Hauraki Gulf, we base our mixingmodel calculations on the isotopic values for themarine and terrestrial end-members reported byKeil and Fogel (2001). We believe this approach iswarranted owing to the similarities between thetwo marine environments. Firstly, the bulk carbonand nitrogen isotopic compositions of organic mat-ter for the marine and slope sites of the Washingtoncoast (d13C = �22.1&; d15N = 5–7&) are similar tothe isotopic compositions of the Hauraki Gulf sed-iments (Table 1). Secondly, the terrestrial organicmatter source in both environments is dominatedby C3 vegetation and the C:N ratios for both sitesare similar (9.4–10 – Keil and Fogel (2001) vs.9.2–16.0 for this study). The D- and L-enantiomersof the source materials were not resolved by Keiland Fogel (2001); we assume here that their re-ported d13C values essentially represent the carbonisotopic composition of the L-stereoisomer of thesource materials.
The isotopic compositions of the terrestrial andmarine end-members in our model have been mod-ified to include the potential reworking of the origi-nal organic matter source. Keil and Fogel (2001)showed that all the marine sediments sampled alongthe Washington coast showed extensive reworkingand that many of the amino acids in the sedimentswere derived from local bacterial populations. Otherstudies have shown that amino acids are systemati-cally depleted by up to 5& relative to the bulk iso-topic signal of the growth substrate (Macko andEstep, 1984; Macko et al., 1987). To account forthe influence of microbial reworking, the isotopiccompositions of the marine and terrestrial end-
Table 1Bulk sediment characteristics and carbon and nitrogen isotopic values
Site Waterdepth(m)
Bulk sediments >63 lm
TOM(%)
CaCO3
(%)POC(%)
PON(%)
C:N %C d1
Site l 82 4.97 26.4 1.1 0.1 9.2 2.0 �Site 2 153 5.00 27.3 0.6 0.1 10.9 1.0 �Site 6a 54 2.65 22.0 0.4 0.1 14.0 2.1 �Site 7 85 7.87 28.7 1.3 0.1 11.4 1.6 �Site 8 28 5.73 8.5 1.6 0.2 9.7 1.8 �Site 10 41 1.90 30.5 – – – 2.9 �Site 11 335 5.98 24.4 0.6 0.1 11.5 0.8 �Site 14 32 2.35 3.3 0.6 0.1 16.0 1.8 �
a Bulk sediment characteristics for site 6 are reported and should be ctwo sites. Bulk data for site 4 was not determined.
members are corrected to reflect this 5& depletionin 13C.
2.8. Estimates of microbial reworking
The contributions of microbially derived L-aminoacids were determined using a two end-member mix-ing model similar to that used for amino acid sourceapportionment (Eqs. (1) and (2)). The reworkingmodel partitions the d13C values of the L-amino acidsbetween a microbial end-member represented by theD-enantiomers of certain amino acids, and either themarine or the terrestrial source. It is assumed that thesedimentary D-amino acid enantiomers are directlyderived from the microbial populations at each siteand therefore, represent the microbial end-memberfor each stereoisomer. The core top sediments areessentially un-racemized and are not a substantialsource of D-amino acids to the Hauraki Gulf sedi-ments (see later discussion). The d13C values of theseD-enantiomers are taken to reflect the source oforganic matter utilized by the local microbial popu-lations. If one assumes that little isotopic differenceoccurs during the synthesis of D- vs. L-amino acidsfrom an organism, then the d13C values of microbi-ally produced amino acids should be roughly equiv-alent (D- or L- : see later discussion). Employing thisassumption, the D-stereoisomers of Ala, Asx, andGlx can be employed with the d13C values of the ter-restrial or marine end-members to estimate the per-cent contribution of microbial derived amino acidsto the sediments such that:
fmicrobial ¼d13Cl-AA � d13Cterr=mar
d13Cd-AA � d13Cterr=mar
ð3Þ
fmicrobial ¼ 1� fterr=mar ð4Þ
<63 lm
3C %N d15N C:N %C d13C %N d15N C:N
20.1 0.3 +7.1 7.0 2.3 �20.2 0.4 +6.7 6.421.4 0.1 +7.6 9.0 2.4 �20.5 0.3 +6.2 8.319.4 0.3 +7.4 7.4 2.5 �20.2 0.3 +7.3 7.619.7 0.2 +7.7 7.4 2.0 �20.6 0.3 +7. 1 7.720.3 0.3 +9.8 7.1 1.9 �20.2 0.3 +8.5 7.020.2 0.4 +8.3 6.7 2.9 �20.3 0.5 +8.3 6.620.7 0.1 +9.2 7.4 1.1 �20.7 0.2 +6.9 7.220.8 0.2 +8.9 7.6 2.5 �20.4 0.3 +8.6 7.1
onsidered representative of site 4 owing to the proximity of these
446 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
where d13CL-AA, d13Cterr/mar, and d13CD-AA are thecarbon isotopic compositions of the sedimentaryL-amino acid, the terrestrial/marine end-member,and the sedimentary D-amino acid, respectively,and where fmicrobial is the fraction of the microbialinput and fterr/mar is the fraction of the marine/ter-restrial derived organic matter, respectively.
3. Results
3.1. Bulk characteristics
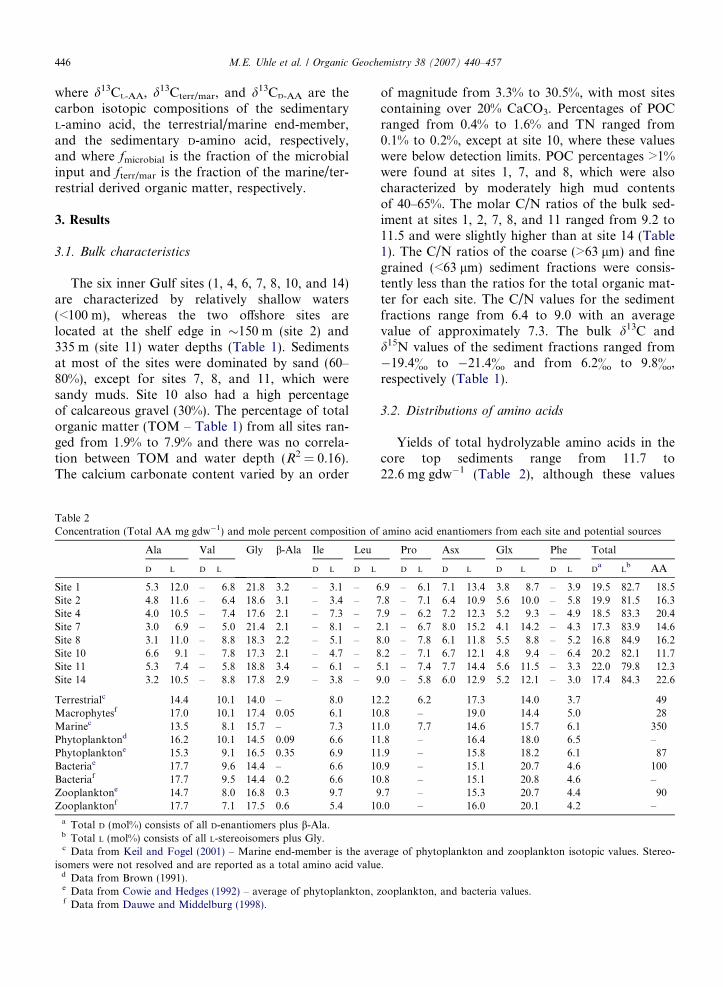
The six inner Gulf sites (1, 4, 6, 7, 8, 10, and 14)are characterized by relatively shallow waters(<100 m), whereas the two offshore sites arelocated at the shelf edge in �150 m (site 2) and335 m (site 11) water depths (Table 1). Sedimentsat most of the sites were dominated by sand (60–80%), except for sites 7, 8, and 11, which weresandy muds. Site 10 also had a high percentageof calcareous gravel (30%). The percentage of totalorganic matter (TOM – Table 1) from all sites ran-ged from 1.9% to 7.9% and there was no correla-tion between TOM and water depth (R2 = 0.16).The calcium carbonate content varied by an order
Table 2Concentration (Total AA mg gdw�1) and mole percent composition of
Ala Val Gly b-Ala Ile Leu
D L D L D L D L
Site 1 5.3 12.0 – 6.8 21.8 3.2 – 3.1 – 6Site 2 4.8 11.6 – 6.4 18.6 3.1 – 3.4 – 7Site 4 4.0 10.5 – 7.4 17.6 2.1 – 7.3 – 7Site 7 3.0 6.9 – 5.0 21.4 2.1 – 8.1 – 2Site 8 3.1 11.0 – 8.8 18.3 2.2 – 5.1 – 8Site 10 6.6 9.1 – 7.8 17.3 2.1 – 4.7 – 8Site 11 5.3 7.4 – 5.8 18.8 3.4 – 6.1 – 5Site 14 3.2 10.5 – 8.8 17.8 2.9 – 3.8 – 9
Terrestrialc 14.4 10.1 14.0 – 8.0 12Macrophytesf 17.0 10.1 17.4 0.05 6.1 10Marinec 13.5 8.1 15.7 – 7.3 11Phytoplanktond 16.2 10.1 14.5 0.09 6.6 11Phytoplanktone 15.3 9.1 16.5 0.35 6.9 11Bacteriae 17.7 9.6 14.4 – 6.6 10Bacteriaf 17.7 9.5 14.4 0.2 6.6 10Zooplanktone 14.7 8.0 16.8 0.3 9.7 9Zooplanktonf 17.7 7.1 17.5 0.6 5.4 10
a Total D (mol%) consists of all D-enantiomers plus b-Ala.b Total L (mol%) consists of all L-stereoisomers plus Gly.c Data from Keil and Fogel (2001) – Marine end-member is the ave
isomers were not resolved and are reported as a total amino acid valued Data from Brown (1991).e Data from Cowie and Hedges (1992) – average of phytoplankton,f Data from Dauwe and Middelburg (1998).
of magnitude from 3.3% to 30.5%, with most sitescontaining over 20% CaCO3. Percentages of POCranged from 0.4% to 1.6% and TN ranged from0.1% to 0.2%, except at site 10, where these valueswere below detection limits. POC percentages >1%were found at sites 1, 7, and 8, which were alsocharacterized by moderately high mud contentsof 40–65%. The molar C/N ratios of the bulk sed-iment at sites 1, 2, 7, 8, and 11 ranged from 9.2 to11.5 and were slightly higher than at site 14 (Table1). The C/N ratios of the coarse (>63 lm) and finegrained (<63 lm) sediment fractions were consis-tently less than the ratios for the total organic mat-ter for each site. The C/N values for the sedimentfractions range from 6.4 to 9.0 with an averagevalue of approximately 7.3. The bulk d13C andd15N values of the sediment fractions ranged from�19.4& to �21.4& and from 6.2& to 9.8&,respectively (Table 1).
3.2. Distributions of amino acids
Yields of total hydrolyzable amino acids in thecore top sediments range from 11.7 to22.6 mg gdw�1 (Table 2), although these values
amino acid enantiomers from each site and potential sources
Pro Asx Glx Phe Total
D L D L D L D L Da
Lb AA
.9 – 6.1 7.1 13.4 3.8 8.7 – 3.9 19.5 82.7 18.5
.8 – 7.1 6.4 10.9 5.6 10.0 – 5.8 19.9 81.5 16.3
.9 – 6.2 7.2 12.3 5.2 9.3 – 4.9 18.5 83.3 20.4
.1 – 6.7 8.0 15.2 4.1 14.2 – 4.3 17.3 83.9 14.6
.0 – 7.8 6.1 11.8 5.5 8.8 – 5.2 16.8 84.9 16.2
.2 – 7.1 6.7 12.1 4.8 9.4 – 6.4 20.2 82.1 11.7
.1 – 7.4 7.7 14.4 5.6 11.5 – 3.3 22.0 79.8 12.3
.0 – 5.8 6.0 12.9 5.2 12.1 – 3.0 17.4 84.3 22.6
.2 6.2 17.3 14.0 3.7 49
.8 – 19.0 14.4 5.0 28
.0 7.7 14.6 15.7 6.1 350
.8 – 16.4 18.0 6.5 –
.9 – 15.8 18.2 6.1 87
.9 – 15.1 20.7 4.6 100
.8 – 15.1 20.8 4.6 –
.7 – 15.3 20.7 4.4 90
.0 – 16.0 20.1 4.2 –
rage of phytoplankton and zooplankton isotopic values. Stereo-.
zooplankton, and bacteria values.
M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 447
are relatively high, they are consistent with othersurface sediments from modern coastal marineenvironments (Cowie and Hedges, 1992; Keil andFogel, 2001; Pantoja and Lee, 2003). Between16% and 26% of the organic carbon and 35–50%of the nitrogen is contained in the sediments asamino acids with the exception of site 10, wherethe amino acids account for only 10% and 13%of the carbon and nitrogen in these sediments,respectively. Gly is the most abundant of the indi-vidual amino acids accounting for between 17 and20 mol% of the total hydrolyzable amino acid(THAA) pool. There is a strong L- over D-aminoacid predominance at all sites, with the L-stereoiso-mers constituting 80 and 85 mol% of the THAApool and the D-enantiomers and b-Ala rangingfrom 17 to 22 mol%. L-Ala, L-Glx, and L-Asx arethe most abundant of the L-amino acids with acombined presence of 30–36 mol% of the L-aminoacids with their D-enantiomers contributing halfas much: 14–18 mol% of the THAA (Table 2). L-Valine (Val), L-Ile, L-Leu, L-Pro and the non-pro-tein amino acid, b-Ala constitute the remainderof the amino acids detected in the sediments. Com-pared to the amino acid distributions of potentialsource materials determined in previous studies(Cowie and Hedges, 1992; Dauwe and Middelburg,1998; Keil and Fogel, 2001), Hauraki Gulf sedi-mentary distributions are depleted in L-Ala, L-Leu, L-Ile (except sites 4, 7, and 11), L-Asx (exceptsite 7), and L-Glx (except site 7). Conversely, thesediments are enriched in Gly and they containup to �70 times more b-Ala. There is no data onthe D-enantiomers of Ala, Asx, and Glx, for thepotential sources (Table 2).
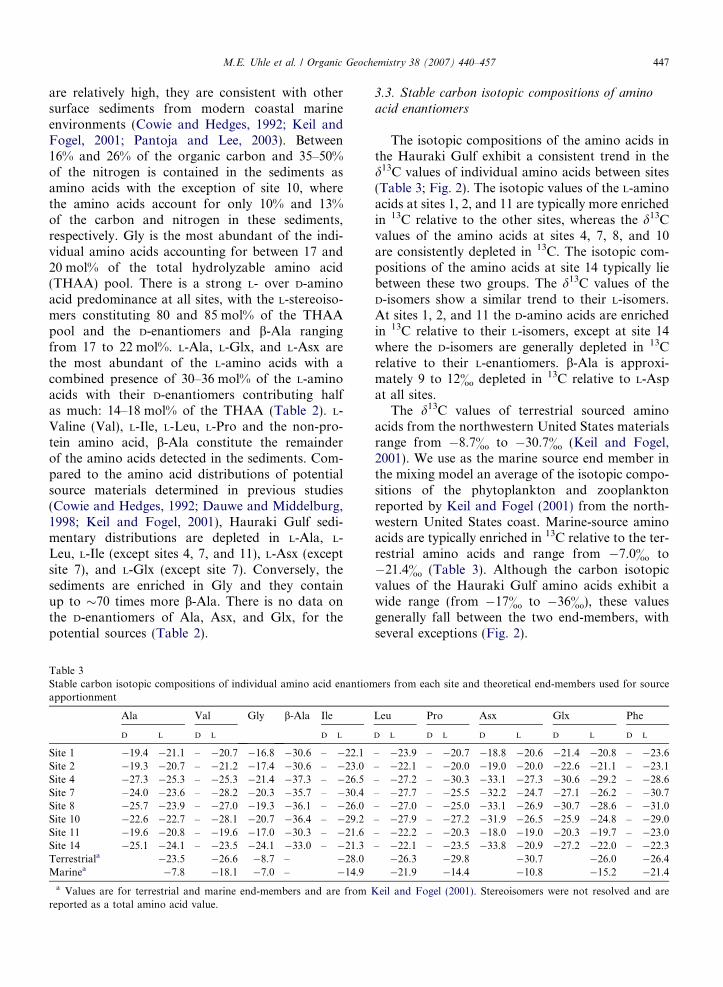
Table 3Stable carbon isotopic compositions of individual amino acid enantiomapportionment
Ala Val Gly b-Ala Ile
D L D L D L
Site 1 �19.4 �21.1 – �20.7 �16.8 �30.6 – �22.1Site 2 �19.3 �20.7 – �21.2 �17.4 �30.6 – �23.0Site 4 �27.3 �25.3 – �25.3 �21.4 �37.3 – �26.5Site 7 �24.0 �23.6 – �28.2 �20.3 �35.7 – �30.4Site 8 �25.7 �23.9 – �27.0 �19.3 �36.1 – �26.0Site 10 �22.6 �22.7 – �28.1 �20.7 �36.4 – �29.2Site 11 �19.6 �20.8 – �19.6 �17.0 �30.3 – �21.6Site 14 �25.1 �24.1 – �23.5 �24.1 �33.0 – �21.3Terrestriala �23.5 �26.6 �8.7 – �28.0Marinea �7.8 �18.1 �7.0 – �14.9
a Values are for terrestrial and marine end-members and are from Kreported as a total amino acid value.
3.3. Stable carbon isotopic compositions of amino
acid enantiomers
The isotopic compositions of the amino acids inthe Hauraki Gulf exhibit a consistent trend in thed13C values of individual amino acids between sites(Table 3; Fig. 2). The isotopic values of the L-aminoacids at sites 1, 2, and 11 are typically more enrichedin 13C relative to the other sites, whereas the d13Cvalues of the amino acids at sites 4, 7, 8, and 10are consistently depleted in 13C. The isotopic com-positions of the amino acids at site 14 typically liebetween these two groups. The d13C values of theD-isomers show a similar trend to their L-isomers.At sites 1, 2, and 11 the D-amino acids are enrichedin 13C relative to their L-isomers, except at site 14where the D-isomers are generally depleted in 13Crelative to their L-enantiomers. b-Ala is approxi-mately 9 to 12& depleted in 13C relative to L-Aspat all sites.
The d13C values of terrestrial sourced aminoacids from the northwestern United States materialsrange from �8.7& to �30.7& (Keil and Fogel,2001). We use as the marine source end member inthe mixing model an average of the isotopic compo-sitions of the phytoplankton and zooplanktonreported by Keil and Fogel (2001) from the north-western United States coast. Marine-source aminoacids are typically enriched in 13C relative to the ter-restrial amino acids and range from �7.0& to�21.4& (Table 3). Although the carbon isotopicvalues of the Hauraki Gulf amino acids exhibit awide range (from �17& to �36&), these valuesgenerally fall between the two end-members, withseveral exceptions (Fig. 2).
ers from each site and theoretical end-members used for source
Leu Pro Asx Glx Phe
D L D L D L D L D L
– �23.9 – �20.7 �18.8 �20.6 �21.4 �20.8 – �23.6– �22.1 – �20.0 �19.0 �20.0 �22.6 �21.1 – �23.1– �27.2 – �30.3 �33.1 �27.3 �30.6 �29.2 – �28.6– �27.7 – �25.5 �32.2 �24.7 �27.1 �26.2 – �30.7– �27.0 – �25.0 �33.1 �26.9 �30.7 �28.6 – �31.0– �27.9 – �27.2 �31.9 �26.5 �25.9 �24.8 – �29.0– �22.2 – �20.3 �18.0 �19.0 �20.3 �19.7 – �23.0– �22.1 – �23.5 �33.8 �20.9 �27.2 �22.0 – �22.3�26.3 �29.8 �30.7 �26.0 �26.4�21.9 �14.4 �10.8 �15.2 �21.4
eil and Fogel (2001). Stereoisomers were not resolved and are

Fig. 2. Isotopic compositions of individual amino acids from Hauraki Gulf sediments and potential source materials used in the sourceapportionment model. The heavy dashed lines represent the terrestrial end-member value for each amino acid and the lighter stippled linecorresponds to the marine end-member value. The shaded boxes represent a 5& depletion in 13C owing to potential microbial reworkingof original source materials. Terrestrial and marine end-member values are from Keil and Fogel (2001).
448 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 449
The isotopic compositions of Gly are depletedsignificantly in 13C relative to both end-membersand the d13C values of D- and L-Ala at sites 4, 7,8, and 14 fall below the terrestrial boundary. Simi-larly, the isotopic values of L-Val and L-Ile at sites7 and 10 are depleted in 13C relative to the end-members. Relatively depleted values of D-Asx andD-Glx also occur at the inner Gulf sites (4, 7, 8,10, and 14 – Fig. 2). The values that lie outsidethe boundaries defined by the terrestrial and marineend-members are technically not amenable to mix-ing of these two sources. However, the modelaccounts for microbial reworking of the originalsource materials as shown by the shaded box, whichrepresents up to a 5& depletion of the originalsource values. When this reworking is accountedfor, the d13C values of all the amino acids exceptGly can be explained by mixing between the marineand terrestrial sources. The d13C values of Gly in theHauraki Gulf are depleted substantially in 13C rela-tive to the reported values of marine and terrestrialsources (Keil and Fogel, 2001). Because these valuesare not amenable for use in the numerical mixingmodel Gly can only be qualitatively considered inour attempt to apportion sources.
3.4. Source apportionment of amino acid
stereoisomers
Owing to the lack of marine and terrestrial d13Cend-member values for the individual amino acidsderived from the organic matter sources in the Hau-raki Gulf, we have based our mixing model on theisotopic values for the marine and terrestrial end-members reported by Keil and Fogel (2001). Weattempted to estimate a putative ‘‘pure’’ terrestrialand marine end-member number (with valuesapproaching those by reported by Keil and Fogel,2001). With the exception of Gly, the isotopic com-position of the L-amino acids at sites 4 and 8, ourmost terrestrial sites, agree well with the Washingtoncoast terrestrial end-member values. Alternately,none of our sites approaches previously reportedmarine end-member values (Table 3; Keil and Fogel,2001). Thus, we conclude that even our most ‘‘mar-ine’’ sites have a substantial terrestrial influence, aconclusion that is supported by the alkane distribu-tions and their 13C signatures in matching samplesfrom the same cores (Sikes et al., 2004). We believethe use of literature values for our calculations iswarranted owing to the similarities in 13C signaturesbetween the two marine environments.
Calculations from the mixing model (Table 4)indicate that the d13C values of the D- and L-aminoacids from the inner Gulf sites (sites 4, 7, 8, and 10)have a predominantly terrestrial source, averagingapproximately 70% terrestrial organic matter input.The isotopic compositions of the D- and L-isomersfrom the northern Gulf and shelf sites (sites 1, 2,and 11) have a stronger marine signal (�72%), withthe exception of the Ala enantiomers. Both stereo-isomers of Ala show a strong terrestrial signal(>70%) at the inner Gulf sites (4, 7, 8, 10, and 14)and a roughly 60:40 mixture of terrestrial and mar-ine sources at the outer Gulf sites (1, 2, and 11). Adominantly terrestrial signal for Ala is consistentwith the findings of Keil and Fogel (2001), whoreported that Ala retained a terrestrial signal alongthe river-coastal ocean gradient. At site 14 in theFirth of Thames (FoT; Fig. 1), the L-stereoisomersindicate a marine source (with the exception of L-Ala, Table 4), whereas the D-isomers of Ala, Asxand Glx reflect a strong terrestrial signal. The d13Cvalues of these bacterial derived D-amino acidsreflect between 76% and 92% input of terrestrialorganic matter, which is in contrast to their L-isomers.
3.5. Extent of microbial reworking
Proportional estimates of the L-amino acidsproduced by microbial populations were calcu-lated for L-Ala, L-Asx, and L-Glx (Table 5) asthese were the only amino acids that had D-iso-mers detected in sufficient concentrations to per-mit isotopic analysis. Microbial contribution tothe L-stereoisomer pools of Ala, Asx, and Glxranges widely, from �20% to �90% (Table 5).In general, Ala is the least influenced by microbialreworking, whereas the microbial contribution tothe L-Asx and L-Glx pools averages �78%. At site10 however, up to �90% of the L-Ala is derivedfrom microbial synthesis. L-Asx at sites 1, 2, and11 shows a slight terrestrial influence as does L-Glx at sites 4 and 8. The strongest influence ofmicrobial production for all three L-isomers isfound at sites 10 and 11, where microbial synthe-sis accounts for an average of �85% of the L-Ala,L-Asx, and L-Glx amino acid pools. Sites 8 and 14show the least influence of microbial reworking,although, on average, between 45% and 50% ofL-Ala, L-Asx, and L-Glx are derived from in situproduction by the microbial populations in thesediments.
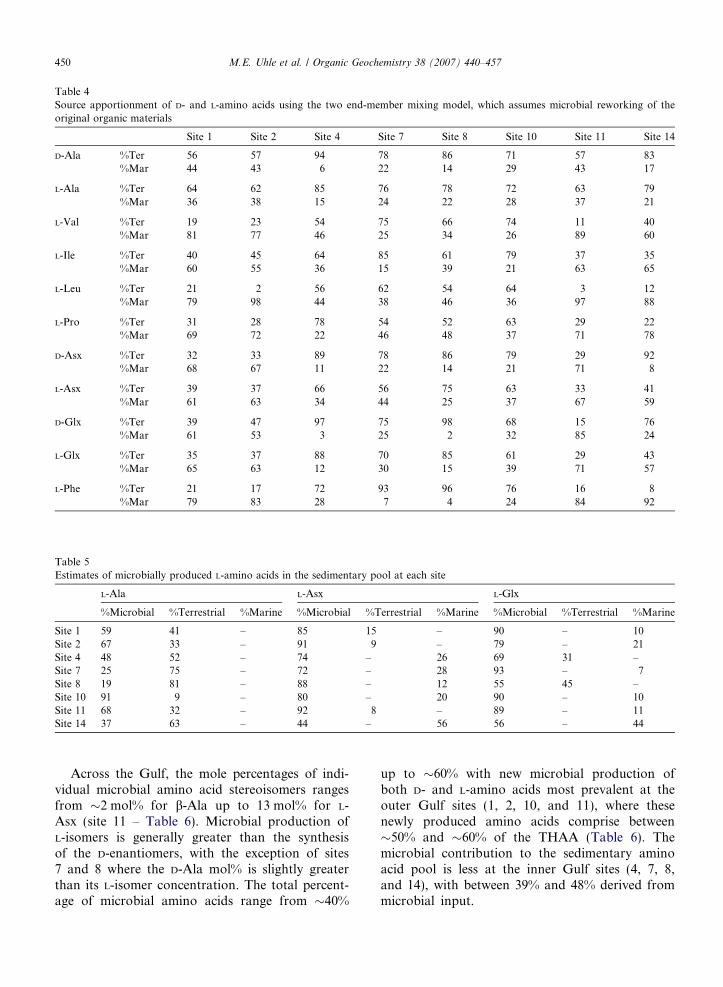
Table 4Source apportionment of D- and L-amino acids using the two end-member mixing model, which assumes microbial reworking of theoriginal organic materials
Site 1 Site 2 Site 4 Site 7 Site 8 Site 10 Site 11 Site 14
D-Ala %Ter 56 57 94 78 86 71 57 83%Mar 44 43 6 22 14 29 43 17
L-Ala %Ter 64 62 85 76 78 72 63 79%Mar 36 38 15 24 22 28 37 21
L-Val %Ter 19 23 54 75 66 74 11 40%Mar 81 77 46 25 34 26 89 60
L-Ile %Ter 40 45 64 85 61 79 37 35%Mar 60 55 36 15 39 21 63 65
L-Leu %Ter 21 2 56 62 54 64 3 12%Mar 79 98 44 38 46 36 97 88
L-Pro %Ter 31 28 78 54 52 63 29 22%Mar 69 72 22 46 48 37 71 78
D-Asx %Ter 32 33 89 78 86 79 29 92%Mar 68 67 11 22 14 21 71 8
L-Asx %Ter 39 37 66 56 75 63 33 41%Mar 61 63 34 44 25 37 67 59
D-Glx %Ter 39 47 97 75 98 68 15 76%Mar 61 53 3 25 2 32 85 24
L-Glx %Ter 35 37 88 70 85 61 29 43%Mar 65 63 12 30 15 39 71 57
L-Phe %Ter 21 17 72 93 96 76 16 8%Mar 79 83 28 7 4 24 84 92
Table 5Estimates of microbially produced L-amino acids in the sedimentary pool at each site
L-Ala L-Asx L-Glx
%Microbial %Terrestrial %Marine %Microbial %Terrestrial %Marine %Microbial %Terrestrial %Marine
Site 1 59 41 – 85 15 – 90 – 10Site 2 67 33 – 91 9 – 79 – 21Site 4 48 52 – 74 – 26 69 31 –Site 7 25 75 – 72 – 28 93 – 7Site 8 19 81 – 88 – 12 55 45 –Site 10 91 9 – 80 – 20 90 – 10Site 11 68 32 – 92 8 – 89 – 11Site 14 37 63 – 44 – 56 56 – 44
450 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
Across the Gulf, the mole percentages of indi-vidual microbial amino acid stereoisomers rangesfrom �2 mol% for b-Ala up to 13 mol% for L-Asx (site 11 – Table 6). Microbial production ofL-isomers is generally greater than the synthesisof the D-enantiomers, with the exception of sites7 and 8 where the D-Ala mol% is slightly greaterthan its L-isomer concentration. The total percent-age of microbial amino acids range from �40%
up to �60% with new microbial production ofboth D- and L-amino acids most prevalent at theouter Gulf sites (1, 2, 10, and 11), where thesenewly produced amino acids comprise between�50% and �60% of the THAA (Table 6). Themicrobial contribution to the sedimentary aminoacid pool is less at the inner Gulf sites (4, 7, 8,and 14), with between 39% and 48% derived frommicrobial input.

Table 6Mole percent composition and total percentage of microbially produced amino acids in sediments at each site
D-Ala L-Ala b-Ala D-Asx L-Asx D-Glx L-Glx Total D-isomer Total L-isomer Total THAA %
Site 1 5 7 3 7 11 4 8 19 26 55Site 2 5 8 3 6 10 6 8 20 26 54Site 4 4 5 2 7 9 5 6 19 21 46Site 7 3 2 2 8 11 4 13 17 26 48Site 8 3 2 2 6 10 5 5 17 17 40Site 10 3 8 2 7 10 5 8 17 26 51Site 11 5 5 3 8 13 6 10 22 29 58Site 14 3 4 3 6 6 5 7 17 17 39
M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 451
4. Discussion
Terrigenous and marine organic materials aretypically considered the principal sources of organiccarbon to coastal marine sediments. If the bulkcharacteristics are taken at face value, the HaurakiGulf sediments are typical coastal margin sediments(Table 1). The C/N ratios of terrestrial organic mat-ter sources exhibit a wide range (15–400) anddecrease during diagenesis, whereas the C/N ratiosof phytoplankton (6–10) are almost constant(Meyers, 1997). The C/N ratios of the bulk sedi-ments at most sites in the Haruaki Gulf (�9 to�11) are consistent with a marine source (Meyers,1997), whereas at site 14 the higher C/N ratio (16)reflects greater input from terrestrial sources(Meyers, 1997). The C/N ratios for the coarse andfine-grained sediment fractions, however, exhibitslightly lower values and a more narrow range thanthe bulk ratios and are more strongly indicative oforganic matter input from phytoplankton. This issupported by the bulk carbon and nitrogen isotopicdata for the sediment size fractions. This mayreflect some preferential loss of terrestrial organicmatter during processing of the sediments for size,but we cannot propose a firm mechanism for this.Mineral-associated organic matter has been sug-gested to reflect sorption of dissolved marineorganic matter more strongly than terrestrial mat-ter. Keil and Fogel (2001) found that terrigenousorganic matter was replaced by marine organicmaterial sorbed to clay isolates along the Washing-ton coast.
It is interesting that the bulk isotopic values ofthe sediment fractions are indicative of a marinesource at sites 8 and 6 (as a proxy for site 4) consid-ering that these sites are closest to terrestrial sourcesand sewage outfall areas (Fig. 1). The influence ofthe sewage outfall flux is inconsequential (Zeldis,2004), however, the effluent is rich in nutrients,
which may help support marine phytoplanktonblooms (Manighetti and Carter, 1999).
Whereas the bulk data (C/N and bulk d13C val-ues) suggest that terrigenous and marine materialsare the principal sources of organic matter to theHauraki Gulf sediments, these bulk parameters arean integrated signal from both allochthonous andautochthonous sources. The bulk data record theisotopic history of all components and cannot pro-vide a detailed picture of individual organic carbonsources, or diagenetic pathways, such as microbialreworking, which can alter the organic matter com-position in the sediments.
Microbial reworking includes not only the degra-dation of labile organic matter but also to the addi-tion of labile organic matter to the sedimentsthrough microbial synthesis. Consequently, micro-bial reworking can alter the isotopic compositionof labile compounds, such as amino acids, throughproduction of microbial biomass, which containsthe same compounds as the original source materi-als, as well as those unique to the bacterial commu-nities in the sediments (Macko et al., 1994; Keil andFogel, 2001). The isotopic compositions and chem-ical distributions of individual amino acids suggestthat microbial reworking of organic matter is signif-icant in the Hauraki Gulf sediments. Three lines ofevidence support this observation.
First, bacteria (and other diagenetic processes)produce non-protein amino acids, (e.g. b-Ala) dur-ing degradation of protein amino acids and the rel-ative enrichment of these non-protein amino acidsin sediments is indicative of early diagenesis. b-Alais predominantly formed from the decarboxylationof aspartic acid (Lee and Cronin, 1982). It is gener-ally accepted that the loss of one of the carboxylgroups in aspartic acid can yield b-Ala, but, thereis some evidence that degradation of the pyrimidineuracil can also form b-Ala (Lee and Cronin, 1982).Concentrations of b-Ala in vascular plants, bacteria

452 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
and plankton species are typically below 1 mol%(Cowie and Hedges, 1992), whereas the concentra-tions of b-Ala in sedimentary marine environmentscan reach values �10 mol% (Cowie et al., 1992,1995; Cowie and Hedges, 1994; Suthhof et al.,2000; Keil and Fogel, 2001). The molar concentra-tions of non-protein amino acids typically increasewith diagenetic alteration of organic matter as thelabile compounds are utilized preferentially by thebiota in favor of the non-protein amino acids(Cowie et al., 1992, 1995; Cowie and Hedges,1994; Dauwe and Middelburg, 1998). In highlydegraded abyssal sediments, non-protein aminoacids can constitute up to 70% of the total sedimen-tary amino acids and their concentrations routinelyincrease down core relative to their parent aminoacids precursors (Cowie and Hedges, 1994; Dauweand Middelburg, 1998). The concentrations of b-Ala in the Hauraki Gulf sediments range from 2.4to 3.7 mol% and these sediments are highly enrichedin b-Ala relative to values that have been measuredin potential source materials (Table 2). The concen-trations of non-protein amino acids in the HaurakiGulf sediments are indicative of microbial rework-ing of the organic matter and these concentrationsare consistent with the concentrations generallyencountered in coastal marine environments whichhave undergone appreciable but not extreme dia-genesis (Cowie and Hedges, 1994; Dauwe and Mid-delburg, 1998; Suthhof et al., 2000; Keil and Fogel,2001).
Second, the presence of D-enantiomers in coretop sediments substantiates microbial reworking ofthe sedimentary organic matter. Whereas D-aminoacids can form through racemization of L-aminoacids after cell death, the time scale for this conver-sion is on the order of �104–106 yr at 0 �C (Bada,1982). Owing to the slow rate of this reaction, anexclusively bacterial origin for D-amino acids inmodern marine environments has been suggestedin a number of studies (Lee and Bada, 1977; Pollockand Kvenvolden, 1978; Bada, 1982; Jorgensen et al.,1990; McCarthy et al., 1998; Pedersen et al., 2001;Perez et al., 2003). The Hauraki Gulf sedimentssampled in this study encompass the top 1 cm ofsediments and are therefore, essentially ‘‘zero-age’’such that the D-enantiomers in these surface sedi-ments have probably originated from the peptido-glycan in bacterial cell walls rather than throughracemization.
The distributions of the three D-enantiomersrecovered from the Hauraki Gulf sediments are con-
sistent with the distribution reported for AarhusBay, Denmark surface sediments (Pedersen et al.,2001), where D-Asp (�14%), D-Ala (�9%), and D-Glu (�8%) dominated the THAA in surface sedi-ments. The mole percentages in those sedimentshowever, were slightly greater than the concentra-tions in the Hauraki Gulf sediments, which maysuggest that the Hauraki Gulf sediments are lessdegraded. Slightly lower concentrations of D-Ala(�8 mol%), and D-Asx (�7 mol%) and D-Glx(�2 mol%) were reported for southern California,surface sediments (Pollock and Kvenvolden, 1978)and are more consistent with the percentage foundin the Hauraki Gulf sediments. The THAA profilefrom southern California sediments, however, wasdominated by D-Ala, which contrasts with boththe Aarhus Bay and Hauraki Gulf sediments.Although D-Ala and D-Glu are reported to be theprincipal D-amino acids in peptidoglycan (Schleiferand Kandler, 1972; Koch, 1990), peptidoglycanstructures between bacterial species has not beenclearly defined (McCarthy et al., 1998; Pedersenet al., 2001). Relatively high concentrations of D-Asp found in some bacterial strains suggests thatthe structure of the peptidoglycan varies betweenbacterial strains. Thus, the presence of D-aminoacids in modern sediments of the Hauraki Gulfare most likely of bacterial origin, which impliessubstantial microbial reworking of organic matterin this Southern Hemisphere temperateenvironment.
The third line of evidence to support microbialreworking of the Hauraki Gulf sediments is basedon the results of the mixing models. The isotopiccompositions of several amino acids lie outside theboundaries defined by the d13C values of the marineand terrestrial end-members (Fig. 2). The presenceof outliers is indicative of amino acid synthesis fromcarbon sources other than those used in the mixingmodel. Amino acids synthesized by heterotrophicbacteria are typically depleted in 13C relative tothe growth substrate utilized by the bacteria (Mackoand Estep, 1984; Macko et al., 1987). Only when theisotopic effects of reworking are accounted for, arethe d13C values of all the amino acids (excludingGly) amenable to the mixing model (Fig. 2). Thisobservation suggests that microbial resynthesis ofboth the marine and terrestrial end-members is animportant process affecting the fate of organic mat-ter throughout the Hauraki Gulf.
Despite correction for microbial resynthesis oforiginal organic matter sources, Gly cannot be mod-

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 453
eled by the mixing equations, which suggests thatthis amino acid is derived from resynthesis of cellu-lar compounds other than amino acids. Highlydepleted values of Gly may be indicative of resyn-thesis from a source depleted in 13C relative toamino acids such as carbohydrates (Degens, 1969;Keil and Fogel, 2001), although we can only specu-late on what the might be the actual substrate mol-ecule(s). This resynthesis is plausible as Gly is anintermediary in many biosynthetic pathways associ-ated with fermentation and its isotopic compositionmay reflect its synthesis from other compounds(Degens, 1969; Keil and Fogel, 2001). Interestingly,the isotopic compositions of Gly are depleted signif-icantly in 13C relative to our chosen end-membervalues, but the overall pattern in which the innerGulf sites are more depleted relative to the outerGulf sites is still observed. This may indicate thatthe Gly at sites 1, 2, and 11 reflect the contributionof Gly from marine phytoplankton which are typi-cally more enriched in amino acids than terrestrialplant materials which are highly carbohydrate-rich(Keil and Fogel, 2001). Keil and Fogel (2001)reported a similar depletion in 13C for Gly in sedi-ments relative to their source materials, which theyattributed to complete resynthesis of Gly from anisotopically lighter source than the amino acids.Complete resynthesis of Gly from carbohydratesmay explain these depleted d13C values observedin the sediments.
There is sufficient evidence to suggest that theorganic matter in the Hauraki Gulf sediments hassignificant amounts of microbial reworking. In lightof this evidence, two questions arise. What are thedominant organic matter sources to the sedimentsat each sampling site? What organic carbon sourcesare the microbial communities utilizing at thesesites? The mixing model calculations show that thed13C values of the D- and L-amino acids from mostof the inner Gulf sites (4, 7, 8, and 10) are domi-nated by a terrestrial source (Table 4), a signal thatis not reflected in the bulk isotope data (Table 1).These sites are located close to the mainland andto potential sewage outfall areas (Fig. 2), wherethe nutrient (as opposed to carbon) flux associatedwith the sewage outfall is suspected to be significantrelative to other freshwater nutrient sources. Zeldisand Smith (1999) estimated that sewage effluentcontributed �40% of the total dissolved inorganicnitrogen flux delivered to the gulf. The influx of sew-age-derived nutrients may support local phyto-plankton blooms in these areas, but autotrophic
production in the gulf is driven ultimately by oce-anic sources of nutrients (Zeldis, 2004). The differ-ences between the bulk and compound specificisotope data suggest that the sedimentary organicmatter at these inner Gulf sites is derived fromtwo sources; marine organic matter which accumu-lates in the sediments and terrestrial materials thatare utilized preferentially by bacterial populationsas a carbon source.
The carbon dynamics at site 14 are consistentwith input of both marine and terrestrial organicmatter. Unlike the other inner Gulf sites, whichrecord a terrestrial signal in the d13C values of boththe D- and L-amino acids, the isotopic compositionsof the D- and L-isomers at site 14 reflect two separatesources. The d13C values of the bulk organic matterand the L-amino acids, with the exception of Ala,are consistent with a marine source, whereas the iso-topic values of the D-enantiomers and b-Ala reflect aterrestrial source. The Firth of Thames receives themajority of the freshwater delivered to the HaurakiGulf via the Waihou River (Zeldis and Smith, 1999;Zeldis, 2004). This river transports nutrient-richwaters and large volumes of sediment from sur-rounding agricultural land. The marine signalrecorded by the d13C values of the bulk organic mat-ter may reflect organic matter from phytoplanktonblooms associated with the delivery of nutrientsvia the Firth of Thames. The terrestrial signal asso-ciated with the D-enantiomers may be a result ofpreferential bacterial utilization of labile terrestrialmaterials deposited in the firth by the WaihouRiver, consistent with the findings of from thenorthern Baltic Sea (Sandberg et al., 2004). Intensemicrobial activity is also inferred from nutrient bud-get and modeling studies in the Gulf and Firth,which suggest that denitrification processes are adominant mechanism in controlling the fate of dis-solved inorganic nitrogen on the inner Hauraki Gulfshelf and Firth of Thames (Giles, 2001; Zeldis,2004).
Rapid loss of terrestrial material and replacementby marine organic matter at site 14 is further sub-stantiated by the source apportionment of L-Leu,L-Pro, and L-Phe. These amino acids have essen-tially a completely marine signal whereas the otherL-isomers at this site reflect a 60:40 marine:terres-trial mixture. The d13C values of these amino acidsreflect substantial loss of the terrestrial signal whichhas been replaced by marine-derived L-Leu, L-Pro,and L-Phe. A similar trend was observed in coastalWashington sediments where 60–70% of the riverine

454 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
Phe and Leu was replaced by their marine-derivedcounterparts and significant inputs of marine Prowere observed at the continental slope (Keil andFogel, 2001). The microbial populations in the sed-iments at site 14 appear to utilize available labile ter-restrial material over marine organic matter at thissite. Dissolved organic carbon from terrestrialinputs can constitute an important and labile nutri-ent and energy source for bacterioplankton and maybe important in controlling the balance between netheterotrophic and autotrophic production in somecoastal environments (Sandberg et al., 2004).
Preferential bacterial utilization of terrestrialmaterials at the inner Gulf sites is further substanti-ated by the isotopic compositions of b-Ala. The car-boxyl groups of aspartic acid isolated from marinephotoautotrophs are typically enriched in 13C rela-tive to the total amino acid by �10& (Abelsonand Hoering, 1961; Savidge and Blair, 2004). Micro-bial decarboxylation of aspartic acid could yield b-Ala that is �10& depleted in 13C relative to theaspartic acid precursor. The d13C values of b-Alaare between 10& and 12& depleted in 13C relativeto their presumed parent precursor, L-Asx (Table3; Fig. 2), which is consistent with formation of b-Ala through microbial decarboxylation of L-Asx.The d13C values of b-Ala at the inner Gulf sitesreflect microbial utilization of predominantly terres-trial material.
The isotopic compositions of both the bulkorganic matter and the D- and L-isomers at sites1, 2, and 11 reflect a predominantly marine source.The exceptions to this trend are the d13C values ofboth Ala stereoisomers and b-Ala, which reflectterrestrial organic matter input to the sedimentsat all Hauraki Gulf sites (Table 3; Fig. 2). Thed13C values of Ala in Washington coastal sedi-ments were also consistent with a terrestrial origin.This suggests that terrestrial Ala does not undergomineralization in marine environments as quicklyas other amino acids particularly Leu and Phe(Keil and Fogel, 2001) making Ala a potentialindicator of terrestrial organic matter input tomarine sediments. These outer Gulf sites are dom-inated by marine organic deposition and the micro-bial communities appear to be utilizing this marineorganic material as a carbon source. This is consis-tent with the proximity of these sites to the shelfedge where nutrient replete waters upwell and phy-toplankton blooms occur especially during the latewinter and early spring (Chang et al., 2003b; Zel-dis, 2004).
Whereas the d13C values of the L-amino acid ster-eoisomers at most sites can be explained as a mix-ture of marine and terrestrial sources, these valuesmay also reflect input from the microbial popula-tions (Macko et al., 1994; Keil and Fogel, 2001).Bacteria produce both D-amino acids and L-isomers,which contribute to the THAA pool. Estimates ofbacterial production of L-amino acids were calcu-lated using mass balance Eqs. (3) and (4) thatassume no difference between the d13C values of D-and L-amino acids produced by heterotrophic bacte-ria. At present, there is no data to determine if anisotopic fractionation occurs during biologicallymediated racemization. For the purpose of thisstudy, however, we hypothesize that the D- and L-amino acids produced by bacteria are isotopicallyequivalent. To our knowledge, no studies havereported fractionation in the isotopic compositionsof D- and L-amino acids from a bacteria populationto test our assumption (Macko et al., 1994). Enzy-matic processes, however, typically exhibit isotopicfractionation. Racemization of L-Ala to D-Ala isthe first step in microbial synthesis of peptidogly-can. This reaction involves racemase enzymes,which convert L-amino acids to their D-stereoiso-mers (Walsh, 1989). If there is an isotopic depletionduring enzymatic racemization, then estimates ofthe microbially produced L-amino acids producedby our model are maximum values for those L-amino acids which are depleted in 13C relative totheir D-enantiomers. If the microbially produced L-amino acid was actually depleted relative to the D-isomers, then the mixing equation will overestimatethe fraction derived from the microbial pool. Con-versely, if the L-amino acids are enriched in 13C rel-ative to their D-enantiomers, then the estimates areminimum values. This effect is not seen in the isoto-pic compositions of D- and L-enantiomers in fossilshell materials where racemization occurs abioti-cally (Macko and Engel, 1991).
The influence of microbial reworking on theorganic matter in the Hauraki Gulf sediments issubstantial. Our data indicate that reworking ismost prevalent at the outer Gulf sites (1, 2, and11), where roughly 55–60% of the total sedimentaryamino acids are derived from local microbial popu-lations. The microbial contributions of L-Ala and L-Asx are considered maximum estimates. Thus,microbial production at these sites estimated bythe model should be considered maxima (Table 6).Marine organic sources constitute the remainderof the amino acids in the outer Gulf sediments

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 455
owing to the importance of seasonal phytoplanktonbiomass accumulation at these sites (Chang et al.,2003b). At the inner Gulf sites 8 and 14, the contri-bution of microbially reworked amino acids is esti-mated at 40%. While this value is substantial, itrepresents a minimum estimate and represents alower percentage of re-worked material than theouter Gulf sites. The microbial contribution at sites4, 7, and 10 are estimated at 50% of the THAA poolwith the other half derived predominantly from ter-restrial material. Each of these sites has proximity toterrestrial sources delivered either via surface runoffor current transport in the Gulf (Fig. 1). Site 4 islocated near Whangarei Harbor where several smallstreams empty into the Hauraki Gulf. Sites 7 and 10sit down current of the wider Gulf. Additionally,site 10 is down current of the Firth of Thames (site14). Waters, which flow through the Colville Chan-nel, will eventually be transported to the shelf at site11. The progressive loss of fresh terrestrial material(replacement by marine material) and the increasein microbial derived organics from sites 14 to 11via site 10 reinforces this idea.
Although our calculations of the microbial con-tribution to the L-amino acid pool in the HaurakiGulf sediments may be considered rough estimates,the impact of microbial reworking of the sedimen-tary organic matter is significant at all sites. Theconcentrations of D-amino acids in the core top sed-iments alone constitute between 17 and 22 mol% ofthe THAA and the outer shelf sites exhibit moremicrobially reworked sediments relative to the innerGulf sites. Including estimates of the microbiallyproduced L-stereoisomers does influence the overallextent of microbial reworking but, these values donot change the spatial pattern of microbial rework-ing throughout the Hauraki Gulf.
5. Conclusions
The organic matter in the Hauraki Gulf sedi-ments is derived from terrestrial, marine and bacte-rial sources. The isotopic data and results of themixing models strongly suggest that microbialreworking of the sediments in the Hauraki Gulf isextensive. At a minimum, 20–40% of the THAAappear to be produced in situ by local microbialpopulations. The carbon sources utilized by bacte-rial populations are site specific and influenced bythe proximity of the site to terrestrial or marineinfluences. The presence of Gly, the D-stereoisomersand b-Ala are good indicators of microbial rework-
ing and their isotopic compositions yield informa-tion about the carbon source utilized by the localpopulations. The d13C values of Gly in the HaurakiGulf suggest that this amino acid is not synthesizedfrom other amino acids, but rather some isotopi-cally depleted source relative to the amino acids. Itis likely that this source is terrestrially derived car-bohydrates at all sites, even those located on theshelf. The presence of terrestrial organic matterthroughout the Hauraki Gulf is supported by thed13C values of D- and L-Ala, which reflect a terres-trial organic matter source. Our data agree with pre-vious work suggesting that Ala is a good indicatorof terrestrial input to marine sediments (Keil andFogel, 2001). Results here indicate higher terrestrialloading in the inner Hauraki Gulf and Firth ofThames which supports previous observations ofhigh levels of nutrient and organic matter remineral-ization, net heterotrophy, and denitrification, whichact to reduce dissolved inorganic nitrogen loads incoastal regions (Giles, 2001; Chang et al., 2003b;Zeldis, 2004).
To our knowledge, this is the first study to utilizethe D- and L-stereoisomers of amino acids to deter-mine the source and extent of microbial alterationof organic matter in marine sediments. Our resultssuggest that microbial reworking of marine sedi-mentary amino acids can be significant in agreementwith the data of Keil and Fogel (2001). In additionto amino acids, microbial populations also produceother labile organic compounds such as carbohy-drates, which may be susceptible to microbial alter-ation and reworking. Early diagenesis does not onlyencompass degradation of organic matter but alsoproduction of new compounds that potentially havedifferent isotopic compositions than the originalsource materials. Microbial ‘‘new production’’ oflabile compounds can therefore markedly alter theisotopic composition of organic matter in marinesediments during early diagenesis.
Acknowledgements
We thank the crew of the RV Tangaroa for theirassistance in sample collection and an all-around-great cruise. The assistance of Mike Costa (Univer-sity of Auckland) in sample recovery and processingis greatly appreciated. Natalie Goodkin and Mela-nie Newell (University of Auckland) provided helpin sample processing and shipping. Melissa Hageand Dr. Kaycie Billmark (UTK) assisted in aminoacid sample extraction and isotopic analysis, and

456 M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457
Lisa Northcote (NIWA) undertook the bulk sedi-ment analyses. This work was supported by Foun-dation of Research, Science and Technology (NZ)research grants to Scott Nodder and John Zeldis(NIWA) (C01822, C01X0027) and the Jones Organ-ic Geochemistry Endowment to Maria Uhle at theUniversity of Tennessee.
References
Abelson, P.H., Hoering, T.C., 1961. Carbon isotopic fraction-ation in formation of amino acids by photosynthetic organ-isms. Proceedings of the National Academy of Science U.S.A.47, 623–632.
Bada, J.L., 1982. Racemization of amino-acids in nature.Interdisciplinary Science Reviews 7 (1), 30–46.
Bada, J.L., Maynard, J.B., Luyendyk, B.P., 1970. Marinesediments – dating by racemization of amino acids. Science170 (NN395), 730.
Brown, M.R., 1991. The amino-acid and sugar composition of 16species of microalgae used in mariculture. Journal of Exper-imental Marine Biology and Ecology 145, 79–99.
Carter, L., Eade, J.V., 1980. Hauraki sediments. New ZealandOceanographic Institute Chart, Coastal Series. Scale 1:200000.
Chang, F.H., Zeldis, J., Gall, M., Hall, J., 2003b. Seasonal andspatial variation of phytoplankton assemblages, biomass, andcell size from spring to summer across the north-eastern NewZealand continental shelf. Journal of Plankton Research 25(7), 737–758.
Cowie, G.L., Hedges, J.I., 1992. Sources and reactivities ofamino-acids in a coastal marine-environment. Limnology andOceanography 37 (4), 703–724.
Cowie, G.L., Hedges, J.I., 1994. Biochemical indicators ofdiagenetic alteration in natural organic-matter mixtures.Nature 369 (6478), 304–307.
Cowie, G.L., Hedges, J.I., Calvert, S.E., 1992. Sources andrelative reactivities of amino-acids, neutral sugars, and ligninin an intermittently anoxic marine- environment. Geochimicaet Cosmochimica Acta 56 (5), 1963–1978.
Cowie, G.L., Hedges, J.I., Prahl, F.G., Delange, G.J., 1995.Elemental and major biochemical-changes across an oxida-tion front in a relict turbidite - an oxygen effect. Geochimicaet Cosmochimica Acta 59 (1), 33–46.
Dauwe, B., Middelburg, J.J., 1998. Amino acids and hexosaminesas indicators of organic matter degradation state in North Seasediments. Limnology and Oceanography 43 (5), 782–798.
Dauwe, B., Middelburg, J.J., Herman, P.M.J., Heip, C.H.R.,1999. Linking diagenetic alteration of amino acids and bulkorganic matter reactivity. Limnology and Oceanograph 44(7), 1809–1814.
Degens, E.T., 1969. Biogeochemistry of stable carbon isotopes.In: Eglinton, G., Murphy, M.T.J. (Eds.), Organic Geochem-istry. Springer-Verlag, pp. 305–329.
Giles, H., 2001. Modelling denitrification in continental shelfsediments. In: Geology. University of Waikato, Hamilton,New Zealand, p. 180.
Hagstrom, A., Pinhassi, J., Zweifel, U.L., 2000. Biogeographicaldiversity among marine bacterioplankton. Aquatic MicrobialEcology 21, 231–244.
Jorgensen, B.B., Bang, M., Blackburn, T.H., 1990. Anaerobicmineralization in marine-sediments from the Baltic-Sea-North-Sea transition. Marine Ecology-Progress Series 59 (1–2), 39–54.
Jorgensen, N.O.G., Stepanaukas, R., Pedersen, A.G.U., Hansen,M., Nybroe, O., 2003. Occurrence and degradation ofpeptidoglycan in aquatic environments. Fems MicrobiologyEcology 46 (3), 269–280.
Keil, R.G., Fogel, M.L., 2001. Reworking of amino acid inmarine sediments: stable carbon isotopic composition ofamino acids in sediments along the Washington coast.Limnology and Oceanography 46 (1), 14–23.
Koch, A.L., 1990. Growth and form of the bacterial cell wall.American Scientist 78, 327–341.
Lee, C., Bada, J.L., 1977. Dissolved amino-acids in equatorialPacific, Sargasso Sea, and Biscayne Bay. Limnology andOceanography 22 (3), 502–510.
Lee, C., Cronin, C., 1982. The vertical flux of particulate organicnitrogen in the sea: decomposition of amino acids in the Peruupwelling area and the equatorial Atlantic. Journal of MarineResearch 40, 227–251.
Lee, C., Wakeham, S.G., Hedges, J.I., 2000. Composition andflux of particulate amino acids and chloropigments inequatorial Pacific seawater and sediments. Deep-Sea ResearchPart I-Oceanographic Research Papers 47 (8), 1535–1568.
Macko, S., Engel, M., Qian, Y., 1994. Early diagenesis andorganic matter preservation – a molecular stable carbonisotope perspective. Chemical Geology 114, 365–379.
Macko, S.A., Engel, M.H., 1991. Assessment of indigeneity infossil organic matter: amino acids and stable isotopes.Philosophical Transactions Royal Society of London B 333,364–367.
Macko, S.A., Estep, M.L., 1984. Microbial alteration of stablenitrogen and carbon isotopic compositions of organic matter.Organic Geochemistry 6, 787–790.
Macko, S.A., Fogel (Estep), M.A., Hare, P.E., Hoering, T.C.,1987. Isotopic fractionation of nitrogen and carbon in thesynthesis of amino acids by microorganisms. ChemicalGeology (Isotope Geoscience Section) 65, 79–92.
Manighetti, B., Carter, L., 1999. Across-shelf sediment dis-persal, Hauraki Gulf, New Zealand. Marine Geology 160,271–300.
McCarthy, M.D., Hedges, J.I., Benner, R., 1998. Major bacterialcontribution to marine dissolved organic nitrogen. Science281 (5374), 231–234.
Meyers, P.A., 1997. Organic geochemical proxies of paleoceano-graphic, paleolimnologic, and paleoclimatic processes.Organic Geochemistry 27 (5–6), 213–250.
Moriarty, D.J.W., Hayward, A.C., 1982. Ultrastructure ofbacteria and the proportion of Gram-negative bacteria inmarine sediments. Microbial Ecology 8, 1–14.
Pantoja, S., Lee, C., 2003. Amino acid remineralization andorganic matter lability in Chilean coastal sediments. OrganicGeochemistry 34, 1047–1056.
Pedersen, A.G.U., Thomsen, T.R., Lomstein, B.A., Jorgensen,N.O.G., 2001. Bacterial influence on amino acid enantiomer-ization in a coastal marine sediment. Limnology and Ocean-ography 46 (6), 1358–1369.
Perez, M.T., Pausz, C., Herndl, G.J., 2003. Major shift inbacterioplankton utilization of enantiomeric amino acidsbetween surface waters and the ocean’s interior. Limnologyand Oceanography 48 (2), 755–763.

M.E. Uhle et al. / Organic Geochemistry 38 (2007) 440–457 457
Pollock, G.E., Kvenvolden, K.A., 1978. Stereochemistry ofamino acids in surface samples of a marine sediment.Geochimica et Cosmochimica Acta 42, 1903–1905.
Sandberg, J., Andersson, A., Johansson, S., Wikner, J., 2004.Pelagic food web structure and carbon budget in the northernBaltic Sea: potential importance of terrigenous carbon.Marine Ecology Progress Series 268, 13–29.
Savidge, W.B., Blair, N.E., 2004. Patterns of intramolecularcarbon isotopic heterogeneity within amino acids of auto-trophs and heterotrophs. Oecologia 139, 178–189.
Schleifer, K.H., Kandler, O., 1972. Peptidoglycan types ofbacterial-cell walls and their taxonomic implications. Bacte-riology Reviews 36, 407–477.
Sharples, J., 1997. Cross-shelf intrusion of subtropical water intothe coastal zone of northeast New Zealand. Continental ShelfResearch 17 (7), 835–857.
Sikes, E.L., Uhle, M.E., Nodder, S.D., Hage, M.M., Howard,M.E., 2004. Assessing the distribution and sources of organicmatter in a coastal marine environment using compoundspecific carbon isotopic analysis of sedimentary lipids:Evidence from the Hauraki Gulf, New Zealand. EOS,Transactions AGU 85 (17), Jt. Assem. Suppl. AbstractB43B-01.
Silfer, J.A., Engel, M.H., Macko, S.A., Jumeau, E.J., 1991. Stablecarbon isotope analysis of amino acid enantiomers byconventional isotope ratio mass spectrometry and combinedgas chromatography/isotope ratio mass spectrometry. Ana-lytical Chemistry 63 (4), 370–374.
Suthhof, A., Jennerhjahn, T.C., Schafer, P., Ittekkot, V., 2000.Nature of organic matter in surface sediments from thePakistan continental margin and the deep Arabian sea: aminoacids. Deep-Sea Research II 47, 329–351.
Verardo, D.J., Froelich, P.N., McIntyre, A., 1990. Determinationof organic carbon and nitrogen in marine sediments using theCarlo Erba NA-1500 analyzer. Deep-Sea Research 37, 157–165.
Waikato, E., 1998a. Environmental Monitoring 1997 DataReport. Environment Waikato Technical Report 1998/05.Environment Waikato.
Waikato, E., 1998b. Regional Rivers Water Quality MonitoringProgramme Data Report 1997. Environment Waikato Tech-nical Report 1998/12. Environment Waikato.
Walsh, C.T., 1989. Enzymes in the D-alanine branch of bacterialcell wall peptidoglycan assembly. The Journal of BiologicalChemistry 264 (5), 2393–2396.
Zeldis, J.R., 2004. New and remineralised nutrient supply andecosystem metabolism on the northeastern New Zealandcontinental shelf. Continental Shelf Research 24, 563–581.
Zeldis, J.R., Smith, S.V., 1999. Water, salt and nutrient budgetsfor Hauraki Gulf, New Zealand. In: Smith, S.V., Crossland,C.J. (Eds.), Australiasian Estutarine Systems: Carbon, nitro-gen and phosphorus fluxes/LOICZ Reports and Studies 121.LOICZ IPO, Texel, The Netherlands, pp. 182.
Zeldis, J.R., Walters, R.A., Greig, M.J.N., Image, K., 2004.Circulation over the northeastern New Zealand continentalslope, shelf and adjacent Hauraki Gulf, during spring andsummer. Continental Shelf Research 24, 543–561.







![Organic Portals Theory - Sources - Library AHlibrary.abundanthope.org/index_htm_files/opsources.pdf · Organic Portals Theory: Sources John Baines The Stellar Man (1985). [p 41] ...](https://static.fdocuments.in/doc/165x107/5bb81a3209d3f2930f8b7ec1/organic-portals-theory-sources-library-organic-portals-theory-sources-john.jpg)










