
Soil bacterial community and functional shifts in response ... · Soil bacterial community and...
16
SOIL, 2, 459–474, 2016 www.soil-journal.net/2/459/2016/ doi:10.5194/soil-2-459-2016 © Author(s) 2016. CC Attribution 3.0 License. SOIL Soil bacterial community and functional shifts in response to altered snowpack in moist acidic tundra of northern Alaska Michael P. Ricketts 1 , Rachel S. Poretsky 1 , Jeffrey M. Welker 2 , and Miquel A. Gonzalez-Meler 1 1 Biological Sciences Department, Ecology and Evolution, University of Illinois at Chicago, Chicago, IL, USA 2 Department of Biological Sciences, University of Alaska, Anchorage, AK, USA Correspondence to: Michael P. Ricketts ([email protected]) Received: 10 December 2015 – Published in SOIL Discuss.: 18 January 2016 Revised: 1 July 2016 – Accepted: 20 July 2016 – Published: 8 September 2016 Abstract. Soil microbial communities play a central role in the cycling of carbon (C) in Arctic tundra ecosys- tems, which contain a large portion of the global C pool. Climate change predictions for Arctic regions include increased temperature and precipitation (i.e. more snow), resulting in increased winter soil insulation, increased soil temperature and moisture, and shifting plant community composition. We utilized an 18-year snow fence study site designed to examine the effects of increased winter precipitation on Arctic tundra soil bacterial com- munities within the context of expected ecosystem response to climate change. Soil was collected from three pre- established treatment zones representing varying degrees of snow accumulation, where deep snow ∼ 100 % and intermediate snow ∼ 50 % increased snowpack relative to the control, and low snow ∼ 25 % decreased snowpack relative to the control. Soil physical properties (temperature, moisture, active layer thaw depth) were measured, and samples were analysed for C concentration, nitrogen (N) concentration, and pH. Soil microbial commu- nity DNA was extracted and the 16S rRNA gene was sequenced to reveal phylogenetic community differences between samples and determine how soil bacterial communities might respond (structurally and functionally) to changes in winter precipitation and soil chemistry. We analysed relative abundance changes of the six most abundant phyla (ranging from 82 to 96 % of total detected phyla per sample) and found four (Acidobacteria, Acti- nobacteria, Verrucomicrobia, and Chloroflexi) responded to deepened snow. All six phyla correlated with at least one of the soil chemical properties (% C, % N, C : N, pH); however, a single predictor was not identified, suggest- ing that each bacterial phylum responds differently to soil characteristics. Overall, bacterial community structure (beta diversity) was found to be associated with snow accumulation treatment and all soil chemical properties. Bacterial functional potential was inferred using ancestral state reconstruction to approximate functional gene abundance, revealing a decreased abundance of genes required for soil organic matter (SOM) decomposition in the organic layers of the deep snow accumulation zones. These results suggest that predicted climate change scenarios may result in altered soil bacterial community structure and function, and indicate a reduction in de- composition potential, alleviated temperature limitations on extracellular enzymatic efficiency, or both. The fate of stored C in Arctic soils ultimately depends on the balance between these mechanisms. Published by Copernicus Publications on behalf of the European Geosciences Union.
Transcript of Soil bacterial community and functional shifts in response ... · Soil bacterial community and...

SOIL 2 459ndash474 2016wwwsoil-journalnet24592016doi105194soil-2-459-2016copy Author(s) 2016 CC Attribution 30 License
SOIL
Soil bacterial community and functional shifts inresponse to altered snowpack in moist acidic tundra of
northern Alaska
Michael P Ricketts1 Rachel S Poretsky1 Jeffrey M Welker2 and Miquel A Gonzalez-Meler1
1Biological Sciences Department Ecology and Evolution University of Illinois at Chicago Chicago IL USA2Department of Biological Sciences University of Alaska Anchorage AK USA
Correspondence to Michael P Ricketts (rickett4uicedu)
Received 10 December 2015 ndash Published in SOIL Discuss 18 January 2016Revised 1 July 2016 ndash Accepted 20 July 2016 ndash Published 8 September 2016
Abstract Soil microbial communities play a central role in the cycling of carbon (C) in Arctic tundra ecosys-tems which contain a large portion of the global C pool Climate change predictions for Arctic regions includeincreased temperature and precipitation (ie more snow) resulting in increased winter soil insulation increasedsoil temperature and moisture and shifting plant community composition We utilized an 18-year snow fencestudy site designed to examine the effects of increased winter precipitation on Arctic tundra soil bacterial com-munities within the context of expected ecosystem response to climate change Soil was collected from three pre-established treatment zones representing varying degrees of snow accumulation where deep snow sim 100 andintermediate snowsim 50 increased snowpack relative to the control and low snowsim 25 decreased snowpackrelative to the control Soil physical properties (temperature moisture active layer thaw depth) were measuredand samples were analysed for C concentration nitrogen (N) concentration and pH Soil microbial commu-nity DNA was extracted and the 16S rRNA gene was sequenced to reveal phylogenetic community differencesbetween samples and determine how soil bacterial communities might respond (structurally and functionally)to changes in winter precipitation and soil chemistry We analysed relative abundance changes of the six mostabundant phyla (ranging from 82 to 96 of total detected phyla per sample) and found four (Acidobacteria Acti-nobacteria Verrucomicrobia and Chloroflexi) responded to deepened snow All six phyla correlated with at leastone of the soil chemical properties ( C N C N pH) however a single predictor was not identified suggest-ing that each bacterial phylum responds differently to soil characteristics Overall bacterial community structure(beta diversity) was found to be associated with snow accumulation treatment and all soil chemical propertiesBacterial functional potential was inferred using ancestral state reconstruction to approximate functional geneabundance revealing a decreased abundance of genes required for soil organic matter (SOM) decomposition inthe organic layers of the deep snow accumulation zones These results suggest that predicted climate changescenarios may result in altered soil bacterial community structure and function and indicate a reduction in de-composition potential alleviated temperature limitations on extracellular enzymatic efficiency or both The fateof stored C in Arctic soils ultimately depends on the balance between these mechanisms
Published by Copernicus Publications on behalf of the European Geosciences Union
460 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
1 Introduction
Broad and rapid environmental changes are driving bothabove- and belowground community shifts in the Arctic (El-mendorf et al 2012a b Tape et al 2006 2012 Wallen-stein et al 2007) It is well established that soil micro-bial communities may alter their composition in responseto changing environmental factors such as nutrient availabil-ity moisture pH temperature and aboveground vegetationshifts (Lauber et al 2009 Morgado et al 2015 Semenovaet al 2015) and ecological and climate-induced changes toArctic soil microbial community structure and function haveimportant effects on ecosystem carbon (C) cycling and nutri-ent availability for plant growth (Deslippe et al 2012 Gra-ham et al 2012 Waldrop et al 2010 Zak and Kling 2006)Because many of these environmental features are rapidlychanging in Arctic tussock tundra ecosystems (Anisimov etal 2007 Liston and Hiemstra 2011) and because of thelarge amounts of C stored in Arctic soils (Hugelius et al2013 Ping et al 2008 Schuur et al 2009 Tarnocai et al2009) it is imperative to examine microbial responses in thissystem
Soil microorganisms play a key role in the decompositionof soil organic matter (SOM) on a global scale releasing nu-trients into the soil and stored C into the atmosphere in theforms of CO2 and CH4 two major greenhouse gases thatcontribute to global warming (Anisimov et al 2007) De-composition of SOM by soil microorganisms amounts to atleast half of the 80ndash90 Gt C released each year by soil respi-ration the second largest terrestrial flux after gross primaryproductivity (GPP Davidson and Janssens 2006 Hopkinset al 2013 Raich et al 2002) Because global soils con-tain about 2000 Gt of Csim 1500 Gt of which is in the form ofSOM (Batjes 1996 IPCC 2000) large-scale changes in therate of microbial decomposition will have an impact on therate at which CO2 accumulates in the atmosphere (Schimeland Schaeffer 2012)
The decomposition rate of SOM resulting in heterotrophicrespiration from soils (Rh) has been shown to be sensitiveto temperature and moisture (Davidson and Janssens 2006Frey et al 2013 Hopkins et al 2012 2013 Xia et al 2014)As the Arctic climate warms increasing Rh may be capableof producing a positive feedback on the climate system as Cstored in soils over millennia is released back to the atmo-sphere (Czimczik and Welker 2010 Jonasson et al 1999Lupascu et al 2014b Mack et al 2004 Nowinski et al2010 Shaver and Chapin III 1980 1986)
Northern latitude permafrost soils may house over 50 ofthe worldrsquos soil organic C (SOC the C component of SOM)approximately twice the amount of C present in the atmo-sphere (Hugelius et al 2013 Ping et al 2008 Schuur et al2009 Tarnocai et al 2009) In addition Arctic ecosystemsare more susceptible to the effects of climate change warm-ing at approximately twice the rate as temperate zones andexhibiting increased winter precipitation patterns (Anisimov
et al 2007 Liston and Hiemstra 2011) Deeper snow hasa suite of cascading consequences in tundra ecosystems assnow acts to insulate soil from extreme winter air tempera-tures resulting in soil temperatures under deeper snowpackup to 10 C warmer than soils under ambient snow depths(Schimel et al 2004) Altered soil conditions under deepersnow may thus lead to increased SOM decomposition caus-ing changes in SOC stocks while also releasing nutrientsfor plant and microbial growth (Anisimov et al 2007 Lef-fler and Welker 2013 Rogers et al 2011 Welker et al2005) The predicted increase in soil temperature as a resultof deeper winter snow accumulation should enhance the rateof SOM decomposition by (1) a direct temperature effect onenzyme kinetics and (2) by increasing substrate availabilityto decomposers as the active layer deepens and permafrostthaws (Luumltzow and Koumlgel-Knabner 2009 Nowinski et al2010 Schuur et al 2008) Therefore warming and deepersnow in the Arctic are likely to expose C stored over millen-nia to decomposers resulting in a major source of C to theatmosphere
However ecosystem C loss may be offset by increasedsoil moisture causing hypoxic conditions and limiting Rh(Blanc-Betes et al 2016) Also microbial mineralization ofplant nutrients such as nitrogen (N) and phosphorus (P)from SOM decomposition are likely to contribute to in-creased net primary productivity (NPP Hinzman et al 2005Natali et al 2012 Pattison and Welker 2014) and causeshifts in vegetation from herbaceous species (cottongrass tus-sock ndash Eriophorum vaginatum) towards woody species (Arc-tic shrubs ndash Betula nana and Salix pulchra) that may producea larger amount of plant litter compounds that are more re-sistant to decomposition (Bret-Harte et al 2001 Pearson etal 2013 Sturm et al 2005 Wahren 2005) The balancebetween these processes will determine the extent to whichArctic tundra ecosystems feedback on the global climatemaking the fate of this stored C unclear (Sistla et al 2013)
This study examined changes in soil bacterial commu-nity composition due to increased winter snow accumula-tion and subsequent altered biotic and abiotic factors usinga long-term snow fence manipulation experiment that mim-ics changes in winter precipitation by creating a gradient ofsnow depths from much deeper than ambient to shallowerthan ambient levels (Jones et al 1998 Pattison and Welker2014 Welker et al 2000) We postulated that increased soilthermal insulation from deeper winter snow accumulationwould elicit bacterial community response via (1) altered soilphysical characteristics such as soil temperature moistureor O2 availability and (2) altered soil chemistry producedby increased microbial mineralization of SOM resulting inincreased nutrient availability and changes in plant speciescomposition and litter Here we evaluated phylum-level shiftsin bacterial community phylogeny using 16S rRNA geneanalysis and predicted bacterial functions using the programPICRUSt (Langille et al 2013) to test whether increasedsnow accumulation and associated changes in soil conditions
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 461
Figure 1 Modified from Walker et al (1999) Schematic of snow accumulation depth at moist acidic tundra site from snow fence manipu-lation Three soil cores were obtained from each treatment zone (labelled Deep Intermediate and Low) and a control zone located gt 30 moutside the effect of the snow fence
(warmer temperatures altered plant inputs and increased hy-poxia) would cause shifts in bacterial community structureand functional potential that reflect increased SOM decom-position and nutrient mineralization
2 Methods
21 Site description and sample collection
The study utilized a long-term snow depth manipulation ex-periment site (Jones et al 1998 Walker et al 1999) es-tablished in 1994 in a moist acidic tundra ecosystem lo-cated near Toolik Lake Field Station Alaska (6837prime N14932primeW) It consists of a strategically placed snow fencedesigned to simulate the increased precipitation patternsand continuous snow-cover episodes predicted under globalwarming scenarios resulting in a gradient of increasing snowaccumulation (and thus increasing soil thermal insulationsoil temperatures and active layer thaw depthpermafrostthaw) with proximity to the fence While snowfall variedfrom year to year the wind drift from the fence provided con-sistent relative snow accumulation at similar distances fromthe fence every winter (Fahnestock et al 2000 Welker et al2005) The soil is classified as Typic Aquiturbel exhibitingcharacteristics of cryoturbation and poor drainage (Ping etal 1998 Soil Survey 2015) Four experimental zones wereidentified according to their snow accumulation regime con-trol (ldquoControlrdquo taken gt 30 m outside the effects of the snowfence) deep snow (ldquoDeeprdquo sim 100 increase in snowpackrelative to the control) intermediate snow (ldquoIntrdquo sim 50 in-crease in snowpack relative to the control) and low snow(ldquoLowrdquo sim 25 decrease in snowpack relative to the con-trol Fig 1) The Deep snow zone is unique in that it is wa-terlogged during thaw periods and dominated not by cot-tongrass tussock or woody shrub species (eg Eriophorumvaginatum Betula nana or Salix pulchra) but by a sedgespecies Carex bigelowii However the vegetative history ofthis plot includes a transition from tussock cottongrass towoody shrub species and finally to wet-sedge species (Arftet al 1999 Walker and Wahren 2006)
Three replicate soil cores were taken approximately 15ndash20 m apart from each experimental snow zone (totalling12 soil cores) in August of 2012 and analysed separately
All soil coring equipment was cleaned and sterilized in thefield between each sample using water and 100 ethanolThe top 10ndash16 cm representing the organic layers was takenfirst using a sharpened steel pipe (55 cm diametertimes 20 cmlength) and serrated knife to cut through surface vegeta-tion and to minimize soil compaction A slide hammer with51 cmtimes 305 cm split soil core sampler (AMS Inc IDUSA) was used to obtain the remainder of the active layerdown to permafrost (sim 35ndash65 cm soil depth) including min-eral soil layers The soil cores were stored in sterile Whirl-pakreg bags immediately frozen on site and shipped to theStable Isotope Laboratory at the University of Illinois atChicago where they were sectioned horizontally into 2 cmdepth segments using a sterilized ice-core cutter providinga 2 cm resolution soil depth profile for each core A por-tion of each segment was ground into a fine powder usinga Spexmill mixermill 8000 (SPEX SamplePrep NJ USA)and analysed for C and N concentration and stable isotopesusing a Costech elemental analyser (Valencia CA USA) inline with a Finnigan DELTAplus XL IRMS (isotope ratiomass spectrometer Bremen Germany) Soil pH was mea-sured from portions of the same segments by creating a soilslurry mixture (2 mL H2O 1 g soil) and using an AccumetBasic AB15 pH meter with a calomel reference pH elec-trode (Thermo Fisher Scientific Inc MA USA) In addi-tion at the time of collection soil temperature soil mois-ture and active layer thaw depth were measured and recordedat four points around each soil core hole (n= 12 per treat-ment) to characterize the soil environment Soil tempera-tures (C) were measured using a 12 cm Taylor TruTemp dig-ital instant-read probe thermometer (Taylor Precision Prod-ucts Inc NM USA) surface (top 12 cm) volumetric watercontent () was measured using a HydroSense soil watercontent measurement system (Campbell Scientific Inc UTUSA) and active layer thaw depths (cm) were measured byinserting a metre stick attached to a metal rod into the grounduntil it hit ice
22 DNA extraction sequencing and analysis
Samples from organic and mineral layers of each soil coreas well as the transition between the two were selectedfor DNA extraction initially based on visual examination
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
462 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
of each individual core section and further classified by C in saturated soils as per the Soil Survey DivisionStaff (1993) (organic ge 12 SOC mineral lt 12 SOC)Organic samples were collected just below where plant tis-sue transitioned into dark brownblack soil (mean soil depthplusmn standard error [SE]= 56plusmn 13 cm Control n= 4 Deepn= 4 Int n= 3 Low n= 4) transitional samples weretaken from the visual border between organic and minerallayers based on change in soil colour (mean soil depthplusmnSE= 148plusmn 18 cm Control n= 3 Deep n= 3 Int n= 4Low n= 3) and mineral samples were collected 10 cm be-low this transition (mean soil depth plusmnSE= 251plusmn 17 cmControl n= 3 Deep n= 4 Int n= 3 Low n= 3) totalling41 samples To maintain consistency only these sampleswere used to analyse C N and pH relationships Sam-ples were sent to Argonne National Laboratory for DNA ex-traction amplification and sequencing as per standards usedby the Earth Microbiome Project (Gilbert et al 2014) DNAextractions were performed using MoBiorsquos PowerSoilreg-htp96-well soil DNA isolation kit as per protocol the V4 re-gion of the 16S rRNA gene was amplified using PCRprimers 515F806R (Caporaso et al 2012) DNA quantifica-tion was performed using PicoGreen and 2times 150 bp paired-end sequencing was performed using an Illumina MiSeq in-strument
Samples were barcoded prior to sequencing for down-stream sample identification and paired-end assembly de-multiplexing quality filtering and operational taxonomicunit (OTU) picking and preliminary diversity analyses wereperformed using the QIIME software package version 180(Caporaso et al 2010) Forward and reverse reads were as-sembled using fastq-join (Aronesty 2011) with 15 bp over-lap at 15 maximum difference Quality filtering includedremoval of reads that did not have at least 75 consecutivehigh-quality (phredgtq20) base calls and truncation of readswith more than three consecutive low-quality (phredltq20)base calls This resulted in an assembled-read median se-quence length of 253 bp
To reveal phylogenetic abundance and relationships se-quence reads were assigned taxonomic identities usingclosed-reference OTU picking that clusters and matches eachread to a reference database Any read that did not match asequence in the reference database was discarded All defaultQIIME parameters were used (reference database Green-genes (13_8) OTU picking method uclust and sequencesimilarity threshold 97 ) Because many organisms areknown to possess multiple copies of the 16S rRNA genein their genome the abundance assignments were correctedbased on known copy numbers using PICRUStrsquos normal-ize_by_copy_numberpy script The relative abundances ofthe six most abundant phyla comprising 82ndash96 of total de-tected phyla per sample were analysed for treatment effectsand alpha and beta diversities were examined using the Shan-non diversity index to estimate within sample diversity and
BrayndashCurtis dissimilarity matrices to determine communitystructure differences
The genetic functional potential of bacterial communitieswas determined using the software package PICRUSt ver-sion 100 (Langille et al 2013) which predicts functionalgene copy numbers in a community based on 16S rRNA se-quencing results Recent advances in sequencing technolo-gies and bioinformatics have greatly enhanced our currentknowledge of the genetic potential of soil microorganismsallowing us to determine what genes a group of organismsis likely to possess based on ancestral state reconstructionof metagenome assemblies from current genomic databases(Langille et al 2013 Martiny et al 2013) PICRUSt utilizesthis knowledge revealing functional potential in the formof gene abundance associated with phylogenetic communitystructure For this study we targeted Kyoto Encyclopedia ofGene and Genomes (KEGG) ortholog assignments for enzy-matic genes commonly associated with SOM decompositionnutrient (nitrogen and phosphate) mobilization and environ-mental stress responses (Sinsabaugh et al 2008 Waldropet al 2010 full list in Table S1 in the Supplement) Thesegenes were then grouped according to functional role result-ing in the following nine gene groups (1) lignin degradation(2) chitin degradation (3) cellulose degradation (4) pectindegradation (5) xylan degradation (6) arabinoside degrada-tion (7) nitrogen mobilization (8) phosphate mobilizationand (9) superoxide dismutation
23 Statistical analyses
Differences between soil layers (organic transition min-eral) and snow accumulation treatments (Control Deep IntLow) including abiotic measurements and relative abun-dance of bacterial 16S rRNA and functional genes were de-termined using the KruskalndashWallis test in the R statisticalsoftware package with a significance threshold of plt 005Due to significant differences between soil layers (Table S2)each layer was analysed separately Only organic and min-eral layers are reported All abiotic factors phyla relativeabundances and relative abundances of functional genes wereanalysed individually to elucidate the treatment effects foreach group and pairwise comparisons were made to deter-mine significant differences between treatments using theNemenyi post hoc test In addition linear regressions wereperformed to determine relationships between soil chemicalproperties ( C N C N and pH) and bacterial abundanceat the phylum level as well as the gene abundances of SOM-degrading enzymes (Figs S1ndashS15 in the Supplement) Toensure accurate comparisons soil chemical properties weremeasured from the same samples that DNA was extractedfrom Only R2 valuesgt 030 are discussed
Bacterial diversity statistics were calculated using QI-IME (Caporaso et al 2010) specifically the com-pare_alpha_diversitypy compare_categoriespy and com-pare_distance_matricespy scripts Pairwise comparisons of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 463
Table 1 Abiotic characteristics of soil from snow accumulation treatments (Low=sim 25 less snowpack than the Control Int=sim 50 more snowpack than the Control Deep=sim 100 more snowpack than the Control) Values are means plusmn standard errors Soil chemicalproperties were obtained from samples used for DNA extraction while temperature and thaw depth were measured in situ (n= 12) Organicand mineral samples were analysed separately using the Nemenyi post hoc test Results are indicated by subscripts ldquoardquo ldquobrdquo or ldquocrdquo onlywhere plt 005
Treatment Soil layers Sample C N C N pH Temp at Thaw depthdepths (cm) 12 cm (C) (cm)
ControlOrganic (n= 4) 675plusmn 312 4521plusmn 109ab 101plusmn 020 5004plusmn 944 459plusmn 009
432plusmn 027b 5917plusmn 123bcMineral (n= 3) 2600plusmn 551 257plusmn 039 015plusmn 003 1767plusmn 134 515plusmn 005ab
LowOrganic (n= 4) 550plusmn 189 4663plusmn 073a 106plusmn 007 4459plusmn 254 444plusmn 008
292plusmn 024b 5092plusmn 320cMineral (n= 3) 2700plusmn 115 418plusmn 192 022plusmn 011 1942plusmn 065 516plusmn 020ab
IntOrganic (n= 3) 367plusmn 067 4059plusmn 243ab 117plusmn 025 3838plusmn 885 469plusmn 041
408plusmn 025b 6188plusmn 119abMineral (n= 3) 2367plusmn 203 258plusmn 049 014plusmn 002 1858plusmn 145 501plusmn 004a
DeepOrganic (n= 4) 600plusmn 370 3651plusmn 427b 140plusmn 007 2627plusmn 341 561plusmn 021
649plusmn 020a 6542plusmn 149aMineral (n= 4) 2400plusmn 442 165plusmn 019 010plusmn 001 1641plusmn 056 583plusmn 017b
the Shannon alpha diversity metrics from soil layer andeach treatment group were made using non-parametric two-sample t tests with 999 Monte Carlo permutations Beta di-versity was analysed by comparing BrayndashCurtis dissimilaritymatrices of bacterial abundance data from each sample to soilchemical properties as well as between soil layers and snowaccumulation treatments using adonis tests with 999 permu-tations Organic and mineral layers were also analysed sep-arately when comparing snow accumulation treatments andsoil chemical properties (Table 2) Analyses of soil chemicalproperties were further substantiated by Mantel tests againusing 999 permutations These data were visualized by cre-ating a non-metric multidimensional scaling (NMDS) plot(stress= 0090 Shepard plot non-metric R2
= 0992) in theR package phyloseq (McMurdie and Holmes 2013) usingthe same BrayndashCurtis dissimilarity matrices (Fig 2)
3 Results
31 Environmental changes
Significant differences in soil temperature (n= 12H = 3329 df= 3 plt 0001) active layer thaw depth(n= 12 H = 2135 df= 3 plt 0001) and organic layers C (n= 4 H = 974 df= 3 p= 0021) were associatedwith the four different snow zones Post hoc tests revealedhigher temperatures in the Deep snow zone relative to theControl (p= 0009) the Int (p= 0001) and the Low snowzone (plt 0001 Table 1) Active layer depth data revealedsimilar results increasing in the Deep snow accumulationzone and decreasing as snowpack was experimentallyreduced Only in the Deep zone was the active layer thawdepth significantly (p= 0020) deeper than the Control zoneHowever along the snow accumulation gradient thaw depthsignificantly increased from Low to Deep plots (LowInt ndashp= 0021 LowDeep ndash plt 0001 Table 1) Soil moisturewas not correlated with snow accumulation possibly the
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
minus10
minus05
00
05
minus1 0 1 2NMDS1
NM
DS
2
Treatment Cont
Deep
Int
Low
N
05
10
15
10
20
30
40
C
Figure 2 Non-metric multidimensional scaling (NMDS) plot us-ing BrayndashCurtis dissimilarity matrices (stress= 0090 Shepard plotnon-metric R2
= 0992) Each point represents the bacterial com-munity structure within one of the 41 total samples used for DNAextraction from all soil depths (organic transition and mineral)Colours indicate C ranging from 14 (light blue) to 486 (dark blue) bubble size indicates N ranging from 009 (small)to 195 (large) and shapes indicate snow accumulation treatments(Control Deep Int Low) Ellipse centroids represent treatmentgroup means while the shape is defined by the covariance withineach group
result of surface hydrology at the site which was largelysaturated throughout the growing season In the organicsoil layers the C concentration of soil declined withincreased snow accumulation (LowDeep ndash p= 003) whilethe N concentration only slightly increased (LowDeep ndashp= 032) This resulted in a decreasing trend in C N ratiosacross snow accumulation treatment zones and relative tothe control (ControlDeep ndash p= 014 Table 1) Soil pH
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

460 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
1 Introduction
Broad and rapid environmental changes are driving bothabove- and belowground community shifts in the Arctic (El-mendorf et al 2012a b Tape et al 2006 2012 Wallen-stein et al 2007) It is well established that soil micro-bial communities may alter their composition in responseto changing environmental factors such as nutrient availabil-ity moisture pH temperature and aboveground vegetationshifts (Lauber et al 2009 Morgado et al 2015 Semenovaet al 2015) and ecological and climate-induced changes toArctic soil microbial community structure and function haveimportant effects on ecosystem carbon (C) cycling and nutri-ent availability for plant growth (Deslippe et al 2012 Gra-ham et al 2012 Waldrop et al 2010 Zak and Kling 2006)Because many of these environmental features are rapidlychanging in Arctic tussock tundra ecosystems (Anisimov etal 2007 Liston and Hiemstra 2011) and because of thelarge amounts of C stored in Arctic soils (Hugelius et al2013 Ping et al 2008 Schuur et al 2009 Tarnocai et al2009) it is imperative to examine microbial responses in thissystem
Soil microorganisms play a key role in the decompositionof soil organic matter (SOM) on a global scale releasing nu-trients into the soil and stored C into the atmosphere in theforms of CO2 and CH4 two major greenhouse gases thatcontribute to global warming (Anisimov et al 2007) De-composition of SOM by soil microorganisms amounts to atleast half of the 80ndash90 Gt C released each year by soil respi-ration the second largest terrestrial flux after gross primaryproductivity (GPP Davidson and Janssens 2006 Hopkinset al 2013 Raich et al 2002) Because global soils con-tain about 2000 Gt of Csim 1500 Gt of which is in the form ofSOM (Batjes 1996 IPCC 2000) large-scale changes in therate of microbial decomposition will have an impact on therate at which CO2 accumulates in the atmosphere (Schimeland Schaeffer 2012)
The decomposition rate of SOM resulting in heterotrophicrespiration from soils (Rh) has been shown to be sensitiveto temperature and moisture (Davidson and Janssens 2006Frey et al 2013 Hopkins et al 2012 2013 Xia et al 2014)As the Arctic climate warms increasing Rh may be capableof producing a positive feedback on the climate system as Cstored in soils over millennia is released back to the atmo-sphere (Czimczik and Welker 2010 Jonasson et al 1999Lupascu et al 2014b Mack et al 2004 Nowinski et al2010 Shaver and Chapin III 1980 1986)
Northern latitude permafrost soils may house over 50 ofthe worldrsquos soil organic C (SOC the C component of SOM)approximately twice the amount of C present in the atmo-sphere (Hugelius et al 2013 Ping et al 2008 Schuur et al2009 Tarnocai et al 2009) In addition Arctic ecosystemsare more susceptible to the effects of climate change warm-ing at approximately twice the rate as temperate zones andexhibiting increased winter precipitation patterns (Anisimov
et al 2007 Liston and Hiemstra 2011) Deeper snow hasa suite of cascading consequences in tundra ecosystems assnow acts to insulate soil from extreme winter air tempera-tures resulting in soil temperatures under deeper snowpackup to 10 C warmer than soils under ambient snow depths(Schimel et al 2004) Altered soil conditions under deepersnow may thus lead to increased SOM decomposition caus-ing changes in SOC stocks while also releasing nutrientsfor plant and microbial growth (Anisimov et al 2007 Lef-fler and Welker 2013 Rogers et al 2011 Welker et al2005) The predicted increase in soil temperature as a resultof deeper winter snow accumulation should enhance the rateof SOM decomposition by (1) a direct temperature effect onenzyme kinetics and (2) by increasing substrate availabilityto decomposers as the active layer deepens and permafrostthaws (Luumltzow and Koumlgel-Knabner 2009 Nowinski et al2010 Schuur et al 2008) Therefore warming and deepersnow in the Arctic are likely to expose C stored over millen-nia to decomposers resulting in a major source of C to theatmosphere
However ecosystem C loss may be offset by increasedsoil moisture causing hypoxic conditions and limiting Rh(Blanc-Betes et al 2016) Also microbial mineralization ofplant nutrients such as nitrogen (N) and phosphorus (P)from SOM decomposition are likely to contribute to in-creased net primary productivity (NPP Hinzman et al 2005Natali et al 2012 Pattison and Welker 2014) and causeshifts in vegetation from herbaceous species (cottongrass tus-sock ndash Eriophorum vaginatum) towards woody species (Arc-tic shrubs ndash Betula nana and Salix pulchra) that may producea larger amount of plant litter compounds that are more re-sistant to decomposition (Bret-Harte et al 2001 Pearson etal 2013 Sturm et al 2005 Wahren 2005) The balancebetween these processes will determine the extent to whichArctic tundra ecosystems feedback on the global climatemaking the fate of this stored C unclear (Sistla et al 2013)
This study examined changes in soil bacterial commu-nity composition due to increased winter snow accumula-tion and subsequent altered biotic and abiotic factors usinga long-term snow fence manipulation experiment that mim-ics changes in winter precipitation by creating a gradient ofsnow depths from much deeper than ambient to shallowerthan ambient levels (Jones et al 1998 Pattison and Welker2014 Welker et al 2000) We postulated that increased soilthermal insulation from deeper winter snow accumulationwould elicit bacterial community response via (1) altered soilphysical characteristics such as soil temperature moistureor O2 availability and (2) altered soil chemistry producedby increased microbial mineralization of SOM resulting inincreased nutrient availability and changes in plant speciescomposition and litter Here we evaluated phylum-level shiftsin bacterial community phylogeny using 16S rRNA geneanalysis and predicted bacterial functions using the programPICRUSt (Langille et al 2013) to test whether increasedsnow accumulation and associated changes in soil conditions
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 461
Figure 1 Modified from Walker et al (1999) Schematic of snow accumulation depth at moist acidic tundra site from snow fence manipu-lation Three soil cores were obtained from each treatment zone (labelled Deep Intermediate and Low) and a control zone located gt 30 moutside the effect of the snow fence
(warmer temperatures altered plant inputs and increased hy-poxia) would cause shifts in bacterial community structureand functional potential that reflect increased SOM decom-position and nutrient mineralization
2 Methods
21 Site description and sample collection
The study utilized a long-term snow depth manipulation ex-periment site (Jones et al 1998 Walker et al 1999) es-tablished in 1994 in a moist acidic tundra ecosystem lo-cated near Toolik Lake Field Station Alaska (6837prime N14932primeW) It consists of a strategically placed snow fencedesigned to simulate the increased precipitation patternsand continuous snow-cover episodes predicted under globalwarming scenarios resulting in a gradient of increasing snowaccumulation (and thus increasing soil thermal insulationsoil temperatures and active layer thaw depthpermafrostthaw) with proximity to the fence While snowfall variedfrom year to year the wind drift from the fence provided con-sistent relative snow accumulation at similar distances fromthe fence every winter (Fahnestock et al 2000 Welker et al2005) The soil is classified as Typic Aquiturbel exhibitingcharacteristics of cryoturbation and poor drainage (Ping etal 1998 Soil Survey 2015) Four experimental zones wereidentified according to their snow accumulation regime con-trol (ldquoControlrdquo taken gt 30 m outside the effects of the snowfence) deep snow (ldquoDeeprdquo sim 100 increase in snowpackrelative to the control) intermediate snow (ldquoIntrdquo sim 50 in-crease in snowpack relative to the control) and low snow(ldquoLowrdquo sim 25 decrease in snowpack relative to the con-trol Fig 1) The Deep snow zone is unique in that it is wa-terlogged during thaw periods and dominated not by cot-tongrass tussock or woody shrub species (eg Eriophorumvaginatum Betula nana or Salix pulchra) but by a sedgespecies Carex bigelowii However the vegetative history ofthis plot includes a transition from tussock cottongrass towoody shrub species and finally to wet-sedge species (Arftet al 1999 Walker and Wahren 2006)
Three replicate soil cores were taken approximately 15ndash20 m apart from each experimental snow zone (totalling12 soil cores) in August of 2012 and analysed separately
All soil coring equipment was cleaned and sterilized in thefield between each sample using water and 100 ethanolThe top 10ndash16 cm representing the organic layers was takenfirst using a sharpened steel pipe (55 cm diametertimes 20 cmlength) and serrated knife to cut through surface vegeta-tion and to minimize soil compaction A slide hammer with51 cmtimes 305 cm split soil core sampler (AMS Inc IDUSA) was used to obtain the remainder of the active layerdown to permafrost (sim 35ndash65 cm soil depth) including min-eral soil layers The soil cores were stored in sterile Whirl-pakreg bags immediately frozen on site and shipped to theStable Isotope Laboratory at the University of Illinois atChicago where they were sectioned horizontally into 2 cmdepth segments using a sterilized ice-core cutter providinga 2 cm resolution soil depth profile for each core A por-tion of each segment was ground into a fine powder usinga Spexmill mixermill 8000 (SPEX SamplePrep NJ USA)and analysed for C and N concentration and stable isotopesusing a Costech elemental analyser (Valencia CA USA) inline with a Finnigan DELTAplus XL IRMS (isotope ratiomass spectrometer Bremen Germany) Soil pH was mea-sured from portions of the same segments by creating a soilslurry mixture (2 mL H2O 1 g soil) and using an AccumetBasic AB15 pH meter with a calomel reference pH elec-trode (Thermo Fisher Scientific Inc MA USA) In addi-tion at the time of collection soil temperature soil mois-ture and active layer thaw depth were measured and recordedat four points around each soil core hole (n= 12 per treat-ment) to characterize the soil environment Soil tempera-tures (C) were measured using a 12 cm Taylor TruTemp dig-ital instant-read probe thermometer (Taylor Precision Prod-ucts Inc NM USA) surface (top 12 cm) volumetric watercontent () was measured using a HydroSense soil watercontent measurement system (Campbell Scientific Inc UTUSA) and active layer thaw depths (cm) were measured byinserting a metre stick attached to a metal rod into the grounduntil it hit ice
22 DNA extraction sequencing and analysis
Samples from organic and mineral layers of each soil coreas well as the transition between the two were selectedfor DNA extraction initially based on visual examination
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
462 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
of each individual core section and further classified by C in saturated soils as per the Soil Survey DivisionStaff (1993) (organic ge 12 SOC mineral lt 12 SOC)Organic samples were collected just below where plant tis-sue transitioned into dark brownblack soil (mean soil depthplusmn standard error [SE]= 56plusmn 13 cm Control n= 4 Deepn= 4 Int n= 3 Low n= 4) transitional samples weretaken from the visual border between organic and minerallayers based on change in soil colour (mean soil depthplusmnSE= 148plusmn 18 cm Control n= 3 Deep n= 3 Int n= 4Low n= 3) and mineral samples were collected 10 cm be-low this transition (mean soil depth plusmnSE= 251plusmn 17 cmControl n= 3 Deep n= 4 Int n= 3 Low n= 3) totalling41 samples To maintain consistency only these sampleswere used to analyse C N and pH relationships Sam-ples were sent to Argonne National Laboratory for DNA ex-traction amplification and sequencing as per standards usedby the Earth Microbiome Project (Gilbert et al 2014) DNAextractions were performed using MoBiorsquos PowerSoilreg-htp96-well soil DNA isolation kit as per protocol the V4 re-gion of the 16S rRNA gene was amplified using PCRprimers 515F806R (Caporaso et al 2012) DNA quantifica-tion was performed using PicoGreen and 2times 150 bp paired-end sequencing was performed using an Illumina MiSeq in-strument
Samples were barcoded prior to sequencing for down-stream sample identification and paired-end assembly de-multiplexing quality filtering and operational taxonomicunit (OTU) picking and preliminary diversity analyses wereperformed using the QIIME software package version 180(Caporaso et al 2010) Forward and reverse reads were as-sembled using fastq-join (Aronesty 2011) with 15 bp over-lap at 15 maximum difference Quality filtering includedremoval of reads that did not have at least 75 consecutivehigh-quality (phredgtq20) base calls and truncation of readswith more than three consecutive low-quality (phredltq20)base calls This resulted in an assembled-read median se-quence length of 253 bp
To reveal phylogenetic abundance and relationships se-quence reads were assigned taxonomic identities usingclosed-reference OTU picking that clusters and matches eachread to a reference database Any read that did not match asequence in the reference database was discarded All defaultQIIME parameters were used (reference database Green-genes (13_8) OTU picking method uclust and sequencesimilarity threshold 97 ) Because many organisms areknown to possess multiple copies of the 16S rRNA genein their genome the abundance assignments were correctedbased on known copy numbers using PICRUStrsquos normal-ize_by_copy_numberpy script The relative abundances ofthe six most abundant phyla comprising 82ndash96 of total de-tected phyla per sample were analysed for treatment effectsand alpha and beta diversities were examined using the Shan-non diversity index to estimate within sample diversity and
BrayndashCurtis dissimilarity matrices to determine communitystructure differences
The genetic functional potential of bacterial communitieswas determined using the software package PICRUSt ver-sion 100 (Langille et al 2013) which predicts functionalgene copy numbers in a community based on 16S rRNA se-quencing results Recent advances in sequencing technolo-gies and bioinformatics have greatly enhanced our currentknowledge of the genetic potential of soil microorganismsallowing us to determine what genes a group of organismsis likely to possess based on ancestral state reconstructionof metagenome assemblies from current genomic databases(Langille et al 2013 Martiny et al 2013) PICRUSt utilizesthis knowledge revealing functional potential in the formof gene abundance associated with phylogenetic communitystructure For this study we targeted Kyoto Encyclopedia ofGene and Genomes (KEGG) ortholog assignments for enzy-matic genes commonly associated with SOM decompositionnutrient (nitrogen and phosphate) mobilization and environ-mental stress responses (Sinsabaugh et al 2008 Waldropet al 2010 full list in Table S1 in the Supplement) Thesegenes were then grouped according to functional role result-ing in the following nine gene groups (1) lignin degradation(2) chitin degradation (3) cellulose degradation (4) pectindegradation (5) xylan degradation (6) arabinoside degrada-tion (7) nitrogen mobilization (8) phosphate mobilizationand (9) superoxide dismutation
23 Statistical analyses
Differences between soil layers (organic transition min-eral) and snow accumulation treatments (Control Deep IntLow) including abiotic measurements and relative abun-dance of bacterial 16S rRNA and functional genes were de-termined using the KruskalndashWallis test in the R statisticalsoftware package with a significance threshold of plt 005Due to significant differences between soil layers (Table S2)each layer was analysed separately Only organic and min-eral layers are reported All abiotic factors phyla relativeabundances and relative abundances of functional genes wereanalysed individually to elucidate the treatment effects foreach group and pairwise comparisons were made to deter-mine significant differences between treatments using theNemenyi post hoc test In addition linear regressions wereperformed to determine relationships between soil chemicalproperties ( C N C N and pH) and bacterial abundanceat the phylum level as well as the gene abundances of SOM-degrading enzymes (Figs S1ndashS15 in the Supplement) Toensure accurate comparisons soil chemical properties weremeasured from the same samples that DNA was extractedfrom Only R2 valuesgt 030 are discussed
Bacterial diversity statistics were calculated using QI-IME (Caporaso et al 2010) specifically the com-pare_alpha_diversitypy compare_categoriespy and com-pare_distance_matricespy scripts Pairwise comparisons of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 463
Table 1 Abiotic characteristics of soil from snow accumulation treatments (Low=sim 25 less snowpack than the Control Int=sim 50 more snowpack than the Control Deep=sim 100 more snowpack than the Control) Values are means plusmn standard errors Soil chemicalproperties were obtained from samples used for DNA extraction while temperature and thaw depth were measured in situ (n= 12) Organicand mineral samples were analysed separately using the Nemenyi post hoc test Results are indicated by subscripts ldquoardquo ldquobrdquo or ldquocrdquo onlywhere plt 005
Treatment Soil layers Sample C N C N pH Temp at Thaw depthdepths (cm) 12 cm (C) (cm)
ControlOrganic (n= 4) 675plusmn 312 4521plusmn 109ab 101plusmn 020 5004plusmn 944 459plusmn 009
432plusmn 027b 5917plusmn 123bcMineral (n= 3) 2600plusmn 551 257plusmn 039 015plusmn 003 1767plusmn 134 515plusmn 005ab
LowOrganic (n= 4) 550plusmn 189 4663plusmn 073a 106plusmn 007 4459plusmn 254 444plusmn 008
292plusmn 024b 5092plusmn 320cMineral (n= 3) 2700plusmn 115 418plusmn 192 022plusmn 011 1942plusmn 065 516plusmn 020ab
IntOrganic (n= 3) 367plusmn 067 4059plusmn 243ab 117plusmn 025 3838plusmn 885 469plusmn 041
408plusmn 025b 6188plusmn 119abMineral (n= 3) 2367plusmn 203 258plusmn 049 014plusmn 002 1858plusmn 145 501plusmn 004a
DeepOrganic (n= 4) 600plusmn 370 3651plusmn 427b 140plusmn 007 2627plusmn 341 561plusmn 021
649plusmn 020a 6542plusmn 149aMineral (n= 4) 2400plusmn 442 165plusmn 019 010plusmn 001 1641plusmn 056 583plusmn 017b
the Shannon alpha diversity metrics from soil layer andeach treatment group were made using non-parametric two-sample t tests with 999 Monte Carlo permutations Beta di-versity was analysed by comparing BrayndashCurtis dissimilaritymatrices of bacterial abundance data from each sample to soilchemical properties as well as between soil layers and snowaccumulation treatments using adonis tests with 999 permu-tations Organic and mineral layers were also analysed sep-arately when comparing snow accumulation treatments andsoil chemical properties (Table 2) Analyses of soil chemicalproperties were further substantiated by Mantel tests againusing 999 permutations These data were visualized by cre-ating a non-metric multidimensional scaling (NMDS) plot(stress= 0090 Shepard plot non-metric R2
= 0992) in theR package phyloseq (McMurdie and Holmes 2013) usingthe same BrayndashCurtis dissimilarity matrices (Fig 2)
3 Results
31 Environmental changes
Significant differences in soil temperature (n= 12H = 3329 df= 3 plt 0001) active layer thaw depth(n= 12 H = 2135 df= 3 plt 0001) and organic layers C (n= 4 H = 974 df= 3 p= 0021) were associatedwith the four different snow zones Post hoc tests revealedhigher temperatures in the Deep snow zone relative to theControl (p= 0009) the Int (p= 0001) and the Low snowzone (plt 0001 Table 1) Active layer depth data revealedsimilar results increasing in the Deep snow accumulationzone and decreasing as snowpack was experimentallyreduced Only in the Deep zone was the active layer thawdepth significantly (p= 0020) deeper than the Control zoneHowever along the snow accumulation gradient thaw depthsignificantly increased from Low to Deep plots (LowInt ndashp= 0021 LowDeep ndash plt 0001 Table 1) Soil moisturewas not correlated with snow accumulation possibly the
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
minus10
minus05
00
05
minus1 0 1 2NMDS1
NM
DS
2
Treatment Cont
Deep
Int
Low
N
05
10
15
10
20
30
40
C
Figure 2 Non-metric multidimensional scaling (NMDS) plot us-ing BrayndashCurtis dissimilarity matrices (stress= 0090 Shepard plotnon-metric R2
= 0992) Each point represents the bacterial com-munity structure within one of the 41 total samples used for DNAextraction from all soil depths (organic transition and mineral)Colours indicate C ranging from 14 (light blue) to 486 (dark blue) bubble size indicates N ranging from 009 (small)to 195 (large) and shapes indicate snow accumulation treatments(Control Deep Int Low) Ellipse centroids represent treatmentgroup means while the shape is defined by the covariance withineach group
result of surface hydrology at the site which was largelysaturated throughout the growing season In the organicsoil layers the C concentration of soil declined withincreased snow accumulation (LowDeep ndash p= 003) whilethe N concentration only slightly increased (LowDeep ndashp= 032) This resulted in a decreasing trend in C N ratiosacross snow accumulation treatment zones and relative tothe control (ControlDeep ndash p= 014 Table 1) Soil pH
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 461
Figure 1 Modified from Walker et al (1999) Schematic of snow accumulation depth at moist acidic tundra site from snow fence manipu-lation Three soil cores were obtained from each treatment zone (labelled Deep Intermediate and Low) and a control zone located gt 30 moutside the effect of the snow fence
(warmer temperatures altered plant inputs and increased hy-poxia) would cause shifts in bacterial community structureand functional potential that reflect increased SOM decom-position and nutrient mineralization
2 Methods
21 Site description and sample collection
The study utilized a long-term snow depth manipulation ex-periment site (Jones et al 1998 Walker et al 1999) es-tablished in 1994 in a moist acidic tundra ecosystem lo-cated near Toolik Lake Field Station Alaska (6837prime N14932primeW) It consists of a strategically placed snow fencedesigned to simulate the increased precipitation patternsand continuous snow-cover episodes predicted under globalwarming scenarios resulting in a gradient of increasing snowaccumulation (and thus increasing soil thermal insulationsoil temperatures and active layer thaw depthpermafrostthaw) with proximity to the fence While snowfall variedfrom year to year the wind drift from the fence provided con-sistent relative snow accumulation at similar distances fromthe fence every winter (Fahnestock et al 2000 Welker et al2005) The soil is classified as Typic Aquiturbel exhibitingcharacteristics of cryoturbation and poor drainage (Ping etal 1998 Soil Survey 2015) Four experimental zones wereidentified according to their snow accumulation regime con-trol (ldquoControlrdquo taken gt 30 m outside the effects of the snowfence) deep snow (ldquoDeeprdquo sim 100 increase in snowpackrelative to the control) intermediate snow (ldquoIntrdquo sim 50 in-crease in snowpack relative to the control) and low snow(ldquoLowrdquo sim 25 decrease in snowpack relative to the con-trol Fig 1) The Deep snow zone is unique in that it is wa-terlogged during thaw periods and dominated not by cot-tongrass tussock or woody shrub species (eg Eriophorumvaginatum Betula nana or Salix pulchra) but by a sedgespecies Carex bigelowii However the vegetative history ofthis plot includes a transition from tussock cottongrass towoody shrub species and finally to wet-sedge species (Arftet al 1999 Walker and Wahren 2006)
Three replicate soil cores were taken approximately 15ndash20 m apart from each experimental snow zone (totalling12 soil cores) in August of 2012 and analysed separately
All soil coring equipment was cleaned and sterilized in thefield between each sample using water and 100 ethanolThe top 10ndash16 cm representing the organic layers was takenfirst using a sharpened steel pipe (55 cm diametertimes 20 cmlength) and serrated knife to cut through surface vegeta-tion and to minimize soil compaction A slide hammer with51 cmtimes 305 cm split soil core sampler (AMS Inc IDUSA) was used to obtain the remainder of the active layerdown to permafrost (sim 35ndash65 cm soil depth) including min-eral soil layers The soil cores were stored in sterile Whirl-pakreg bags immediately frozen on site and shipped to theStable Isotope Laboratory at the University of Illinois atChicago where they were sectioned horizontally into 2 cmdepth segments using a sterilized ice-core cutter providinga 2 cm resolution soil depth profile for each core A por-tion of each segment was ground into a fine powder usinga Spexmill mixermill 8000 (SPEX SamplePrep NJ USA)and analysed for C and N concentration and stable isotopesusing a Costech elemental analyser (Valencia CA USA) inline with a Finnigan DELTAplus XL IRMS (isotope ratiomass spectrometer Bremen Germany) Soil pH was mea-sured from portions of the same segments by creating a soilslurry mixture (2 mL H2O 1 g soil) and using an AccumetBasic AB15 pH meter with a calomel reference pH elec-trode (Thermo Fisher Scientific Inc MA USA) In addi-tion at the time of collection soil temperature soil mois-ture and active layer thaw depth were measured and recordedat four points around each soil core hole (n= 12 per treat-ment) to characterize the soil environment Soil tempera-tures (C) were measured using a 12 cm Taylor TruTemp dig-ital instant-read probe thermometer (Taylor Precision Prod-ucts Inc NM USA) surface (top 12 cm) volumetric watercontent () was measured using a HydroSense soil watercontent measurement system (Campbell Scientific Inc UTUSA) and active layer thaw depths (cm) were measured byinserting a metre stick attached to a metal rod into the grounduntil it hit ice
22 DNA extraction sequencing and analysis
Samples from organic and mineral layers of each soil coreas well as the transition between the two were selectedfor DNA extraction initially based on visual examination
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
462 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
of each individual core section and further classified by C in saturated soils as per the Soil Survey DivisionStaff (1993) (organic ge 12 SOC mineral lt 12 SOC)Organic samples were collected just below where plant tis-sue transitioned into dark brownblack soil (mean soil depthplusmn standard error [SE]= 56plusmn 13 cm Control n= 4 Deepn= 4 Int n= 3 Low n= 4) transitional samples weretaken from the visual border between organic and minerallayers based on change in soil colour (mean soil depthplusmnSE= 148plusmn 18 cm Control n= 3 Deep n= 3 Int n= 4Low n= 3) and mineral samples were collected 10 cm be-low this transition (mean soil depth plusmnSE= 251plusmn 17 cmControl n= 3 Deep n= 4 Int n= 3 Low n= 3) totalling41 samples To maintain consistency only these sampleswere used to analyse C N and pH relationships Sam-ples were sent to Argonne National Laboratory for DNA ex-traction amplification and sequencing as per standards usedby the Earth Microbiome Project (Gilbert et al 2014) DNAextractions were performed using MoBiorsquos PowerSoilreg-htp96-well soil DNA isolation kit as per protocol the V4 re-gion of the 16S rRNA gene was amplified using PCRprimers 515F806R (Caporaso et al 2012) DNA quantifica-tion was performed using PicoGreen and 2times 150 bp paired-end sequencing was performed using an Illumina MiSeq in-strument
Samples were barcoded prior to sequencing for down-stream sample identification and paired-end assembly de-multiplexing quality filtering and operational taxonomicunit (OTU) picking and preliminary diversity analyses wereperformed using the QIIME software package version 180(Caporaso et al 2010) Forward and reverse reads were as-sembled using fastq-join (Aronesty 2011) with 15 bp over-lap at 15 maximum difference Quality filtering includedremoval of reads that did not have at least 75 consecutivehigh-quality (phredgtq20) base calls and truncation of readswith more than three consecutive low-quality (phredltq20)base calls This resulted in an assembled-read median se-quence length of 253 bp
To reveal phylogenetic abundance and relationships se-quence reads were assigned taxonomic identities usingclosed-reference OTU picking that clusters and matches eachread to a reference database Any read that did not match asequence in the reference database was discarded All defaultQIIME parameters were used (reference database Green-genes (13_8) OTU picking method uclust and sequencesimilarity threshold 97 ) Because many organisms areknown to possess multiple copies of the 16S rRNA genein their genome the abundance assignments were correctedbased on known copy numbers using PICRUStrsquos normal-ize_by_copy_numberpy script The relative abundances ofthe six most abundant phyla comprising 82ndash96 of total de-tected phyla per sample were analysed for treatment effectsand alpha and beta diversities were examined using the Shan-non diversity index to estimate within sample diversity and
BrayndashCurtis dissimilarity matrices to determine communitystructure differences
The genetic functional potential of bacterial communitieswas determined using the software package PICRUSt ver-sion 100 (Langille et al 2013) which predicts functionalgene copy numbers in a community based on 16S rRNA se-quencing results Recent advances in sequencing technolo-gies and bioinformatics have greatly enhanced our currentknowledge of the genetic potential of soil microorganismsallowing us to determine what genes a group of organismsis likely to possess based on ancestral state reconstructionof metagenome assemblies from current genomic databases(Langille et al 2013 Martiny et al 2013) PICRUSt utilizesthis knowledge revealing functional potential in the formof gene abundance associated with phylogenetic communitystructure For this study we targeted Kyoto Encyclopedia ofGene and Genomes (KEGG) ortholog assignments for enzy-matic genes commonly associated with SOM decompositionnutrient (nitrogen and phosphate) mobilization and environ-mental stress responses (Sinsabaugh et al 2008 Waldropet al 2010 full list in Table S1 in the Supplement) Thesegenes were then grouped according to functional role result-ing in the following nine gene groups (1) lignin degradation(2) chitin degradation (3) cellulose degradation (4) pectindegradation (5) xylan degradation (6) arabinoside degrada-tion (7) nitrogen mobilization (8) phosphate mobilizationand (9) superoxide dismutation
23 Statistical analyses
Differences between soil layers (organic transition min-eral) and snow accumulation treatments (Control Deep IntLow) including abiotic measurements and relative abun-dance of bacterial 16S rRNA and functional genes were de-termined using the KruskalndashWallis test in the R statisticalsoftware package with a significance threshold of plt 005Due to significant differences between soil layers (Table S2)each layer was analysed separately Only organic and min-eral layers are reported All abiotic factors phyla relativeabundances and relative abundances of functional genes wereanalysed individually to elucidate the treatment effects foreach group and pairwise comparisons were made to deter-mine significant differences between treatments using theNemenyi post hoc test In addition linear regressions wereperformed to determine relationships between soil chemicalproperties ( C N C N and pH) and bacterial abundanceat the phylum level as well as the gene abundances of SOM-degrading enzymes (Figs S1ndashS15 in the Supplement) Toensure accurate comparisons soil chemical properties weremeasured from the same samples that DNA was extractedfrom Only R2 valuesgt 030 are discussed
Bacterial diversity statistics were calculated using QI-IME (Caporaso et al 2010) specifically the com-pare_alpha_diversitypy compare_categoriespy and com-pare_distance_matricespy scripts Pairwise comparisons of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 463
Table 1 Abiotic characteristics of soil from snow accumulation treatments (Low=sim 25 less snowpack than the Control Int=sim 50 more snowpack than the Control Deep=sim 100 more snowpack than the Control) Values are means plusmn standard errors Soil chemicalproperties were obtained from samples used for DNA extraction while temperature and thaw depth were measured in situ (n= 12) Organicand mineral samples were analysed separately using the Nemenyi post hoc test Results are indicated by subscripts ldquoardquo ldquobrdquo or ldquocrdquo onlywhere plt 005
Treatment Soil layers Sample C N C N pH Temp at Thaw depthdepths (cm) 12 cm (C) (cm)
ControlOrganic (n= 4) 675plusmn 312 4521plusmn 109ab 101plusmn 020 5004plusmn 944 459plusmn 009
432plusmn 027b 5917plusmn 123bcMineral (n= 3) 2600plusmn 551 257plusmn 039 015plusmn 003 1767plusmn 134 515plusmn 005ab
LowOrganic (n= 4) 550plusmn 189 4663plusmn 073a 106plusmn 007 4459plusmn 254 444plusmn 008
292plusmn 024b 5092plusmn 320cMineral (n= 3) 2700plusmn 115 418plusmn 192 022plusmn 011 1942plusmn 065 516plusmn 020ab
IntOrganic (n= 3) 367plusmn 067 4059plusmn 243ab 117plusmn 025 3838plusmn 885 469plusmn 041
408plusmn 025b 6188plusmn 119abMineral (n= 3) 2367plusmn 203 258plusmn 049 014plusmn 002 1858plusmn 145 501plusmn 004a
DeepOrganic (n= 4) 600plusmn 370 3651plusmn 427b 140plusmn 007 2627plusmn 341 561plusmn 021
649plusmn 020a 6542plusmn 149aMineral (n= 4) 2400plusmn 442 165plusmn 019 010plusmn 001 1641plusmn 056 583plusmn 017b
the Shannon alpha diversity metrics from soil layer andeach treatment group were made using non-parametric two-sample t tests with 999 Monte Carlo permutations Beta di-versity was analysed by comparing BrayndashCurtis dissimilaritymatrices of bacterial abundance data from each sample to soilchemical properties as well as between soil layers and snowaccumulation treatments using adonis tests with 999 permu-tations Organic and mineral layers were also analysed sep-arately when comparing snow accumulation treatments andsoil chemical properties (Table 2) Analyses of soil chemicalproperties were further substantiated by Mantel tests againusing 999 permutations These data were visualized by cre-ating a non-metric multidimensional scaling (NMDS) plot(stress= 0090 Shepard plot non-metric R2
= 0992) in theR package phyloseq (McMurdie and Holmes 2013) usingthe same BrayndashCurtis dissimilarity matrices (Fig 2)
3 Results
31 Environmental changes
Significant differences in soil temperature (n= 12H = 3329 df= 3 plt 0001) active layer thaw depth(n= 12 H = 2135 df= 3 plt 0001) and organic layers C (n= 4 H = 974 df= 3 p= 0021) were associatedwith the four different snow zones Post hoc tests revealedhigher temperatures in the Deep snow zone relative to theControl (p= 0009) the Int (p= 0001) and the Low snowzone (plt 0001 Table 1) Active layer depth data revealedsimilar results increasing in the Deep snow accumulationzone and decreasing as snowpack was experimentallyreduced Only in the Deep zone was the active layer thawdepth significantly (p= 0020) deeper than the Control zoneHowever along the snow accumulation gradient thaw depthsignificantly increased from Low to Deep plots (LowInt ndashp= 0021 LowDeep ndash plt 0001 Table 1) Soil moisturewas not correlated with snow accumulation possibly the
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
minus10
minus05
00
05
minus1 0 1 2NMDS1
NM
DS
2
Treatment Cont
Deep
Int
Low
N
05
10
15
10
20
30
40
C
Figure 2 Non-metric multidimensional scaling (NMDS) plot us-ing BrayndashCurtis dissimilarity matrices (stress= 0090 Shepard plotnon-metric R2
= 0992) Each point represents the bacterial com-munity structure within one of the 41 total samples used for DNAextraction from all soil depths (organic transition and mineral)Colours indicate C ranging from 14 (light blue) to 486 (dark blue) bubble size indicates N ranging from 009 (small)to 195 (large) and shapes indicate snow accumulation treatments(Control Deep Int Low) Ellipse centroids represent treatmentgroup means while the shape is defined by the covariance withineach group
result of surface hydrology at the site which was largelysaturated throughout the growing season In the organicsoil layers the C concentration of soil declined withincreased snow accumulation (LowDeep ndash p= 003) whilethe N concentration only slightly increased (LowDeep ndashp= 032) This resulted in a decreasing trend in C N ratiosacross snow accumulation treatment zones and relative tothe control (ControlDeep ndash p= 014 Table 1) Soil pH
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

462 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
of each individual core section and further classified by C in saturated soils as per the Soil Survey DivisionStaff (1993) (organic ge 12 SOC mineral lt 12 SOC)Organic samples were collected just below where plant tis-sue transitioned into dark brownblack soil (mean soil depthplusmn standard error [SE]= 56plusmn 13 cm Control n= 4 Deepn= 4 Int n= 3 Low n= 4) transitional samples weretaken from the visual border between organic and minerallayers based on change in soil colour (mean soil depthplusmnSE= 148plusmn 18 cm Control n= 3 Deep n= 3 Int n= 4Low n= 3) and mineral samples were collected 10 cm be-low this transition (mean soil depth plusmnSE= 251plusmn 17 cmControl n= 3 Deep n= 4 Int n= 3 Low n= 3) totalling41 samples To maintain consistency only these sampleswere used to analyse C N and pH relationships Sam-ples were sent to Argonne National Laboratory for DNA ex-traction amplification and sequencing as per standards usedby the Earth Microbiome Project (Gilbert et al 2014) DNAextractions were performed using MoBiorsquos PowerSoilreg-htp96-well soil DNA isolation kit as per protocol the V4 re-gion of the 16S rRNA gene was amplified using PCRprimers 515F806R (Caporaso et al 2012) DNA quantifica-tion was performed using PicoGreen and 2times 150 bp paired-end sequencing was performed using an Illumina MiSeq in-strument
Samples were barcoded prior to sequencing for down-stream sample identification and paired-end assembly de-multiplexing quality filtering and operational taxonomicunit (OTU) picking and preliminary diversity analyses wereperformed using the QIIME software package version 180(Caporaso et al 2010) Forward and reverse reads were as-sembled using fastq-join (Aronesty 2011) with 15 bp over-lap at 15 maximum difference Quality filtering includedremoval of reads that did not have at least 75 consecutivehigh-quality (phredgtq20) base calls and truncation of readswith more than three consecutive low-quality (phredltq20)base calls This resulted in an assembled-read median se-quence length of 253 bp
To reveal phylogenetic abundance and relationships se-quence reads were assigned taxonomic identities usingclosed-reference OTU picking that clusters and matches eachread to a reference database Any read that did not match asequence in the reference database was discarded All defaultQIIME parameters were used (reference database Green-genes (13_8) OTU picking method uclust and sequencesimilarity threshold 97 ) Because many organisms areknown to possess multiple copies of the 16S rRNA genein their genome the abundance assignments were correctedbased on known copy numbers using PICRUStrsquos normal-ize_by_copy_numberpy script The relative abundances ofthe six most abundant phyla comprising 82ndash96 of total de-tected phyla per sample were analysed for treatment effectsand alpha and beta diversities were examined using the Shan-non diversity index to estimate within sample diversity and
BrayndashCurtis dissimilarity matrices to determine communitystructure differences
The genetic functional potential of bacterial communitieswas determined using the software package PICRUSt ver-sion 100 (Langille et al 2013) which predicts functionalgene copy numbers in a community based on 16S rRNA se-quencing results Recent advances in sequencing technolo-gies and bioinformatics have greatly enhanced our currentknowledge of the genetic potential of soil microorganismsallowing us to determine what genes a group of organismsis likely to possess based on ancestral state reconstructionof metagenome assemblies from current genomic databases(Langille et al 2013 Martiny et al 2013) PICRUSt utilizesthis knowledge revealing functional potential in the formof gene abundance associated with phylogenetic communitystructure For this study we targeted Kyoto Encyclopedia ofGene and Genomes (KEGG) ortholog assignments for enzy-matic genes commonly associated with SOM decompositionnutrient (nitrogen and phosphate) mobilization and environ-mental stress responses (Sinsabaugh et al 2008 Waldropet al 2010 full list in Table S1 in the Supplement) Thesegenes were then grouped according to functional role result-ing in the following nine gene groups (1) lignin degradation(2) chitin degradation (3) cellulose degradation (4) pectindegradation (5) xylan degradation (6) arabinoside degrada-tion (7) nitrogen mobilization (8) phosphate mobilizationand (9) superoxide dismutation
23 Statistical analyses
Differences between soil layers (organic transition min-eral) and snow accumulation treatments (Control Deep IntLow) including abiotic measurements and relative abun-dance of bacterial 16S rRNA and functional genes were de-termined using the KruskalndashWallis test in the R statisticalsoftware package with a significance threshold of plt 005Due to significant differences between soil layers (Table S2)each layer was analysed separately Only organic and min-eral layers are reported All abiotic factors phyla relativeabundances and relative abundances of functional genes wereanalysed individually to elucidate the treatment effects foreach group and pairwise comparisons were made to deter-mine significant differences between treatments using theNemenyi post hoc test In addition linear regressions wereperformed to determine relationships between soil chemicalproperties ( C N C N and pH) and bacterial abundanceat the phylum level as well as the gene abundances of SOM-degrading enzymes (Figs S1ndashS15 in the Supplement) Toensure accurate comparisons soil chemical properties weremeasured from the same samples that DNA was extractedfrom Only R2 valuesgt 030 are discussed
Bacterial diversity statistics were calculated using QI-IME (Caporaso et al 2010) specifically the com-pare_alpha_diversitypy compare_categoriespy and com-pare_distance_matricespy scripts Pairwise comparisons of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 463
Table 1 Abiotic characteristics of soil from snow accumulation treatments (Low=sim 25 less snowpack than the Control Int=sim 50 more snowpack than the Control Deep=sim 100 more snowpack than the Control) Values are means plusmn standard errors Soil chemicalproperties were obtained from samples used for DNA extraction while temperature and thaw depth were measured in situ (n= 12) Organicand mineral samples were analysed separately using the Nemenyi post hoc test Results are indicated by subscripts ldquoardquo ldquobrdquo or ldquocrdquo onlywhere plt 005
Treatment Soil layers Sample C N C N pH Temp at Thaw depthdepths (cm) 12 cm (C) (cm)
ControlOrganic (n= 4) 675plusmn 312 4521plusmn 109ab 101plusmn 020 5004plusmn 944 459plusmn 009
432plusmn 027b 5917plusmn 123bcMineral (n= 3) 2600plusmn 551 257plusmn 039 015plusmn 003 1767plusmn 134 515plusmn 005ab
LowOrganic (n= 4) 550plusmn 189 4663plusmn 073a 106plusmn 007 4459plusmn 254 444plusmn 008
292plusmn 024b 5092plusmn 320cMineral (n= 3) 2700plusmn 115 418plusmn 192 022plusmn 011 1942plusmn 065 516plusmn 020ab
IntOrganic (n= 3) 367plusmn 067 4059plusmn 243ab 117plusmn 025 3838plusmn 885 469plusmn 041
408plusmn 025b 6188plusmn 119abMineral (n= 3) 2367plusmn 203 258plusmn 049 014plusmn 002 1858plusmn 145 501plusmn 004a
DeepOrganic (n= 4) 600plusmn 370 3651plusmn 427b 140plusmn 007 2627plusmn 341 561plusmn 021
649plusmn 020a 6542plusmn 149aMineral (n= 4) 2400plusmn 442 165plusmn 019 010plusmn 001 1641plusmn 056 583plusmn 017b
the Shannon alpha diversity metrics from soil layer andeach treatment group were made using non-parametric two-sample t tests with 999 Monte Carlo permutations Beta di-versity was analysed by comparing BrayndashCurtis dissimilaritymatrices of bacterial abundance data from each sample to soilchemical properties as well as between soil layers and snowaccumulation treatments using adonis tests with 999 permu-tations Organic and mineral layers were also analysed sep-arately when comparing snow accumulation treatments andsoil chemical properties (Table 2) Analyses of soil chemicalproperties were further substantiated by Mantel tests againusing 999 permutations These data were visualized by cre-ating a non-metric multidimensional scaling (NMDS) plot(stress= 0090 Shepard plot non-metric R2
= 0992) in theR package phyloseq (McMurdie and Holmes 2013) usingthe same BrayndashCurtis dissimilarity matrices (Fig 2)
3 Results
31 Environmental changes
Significant differences in soil temperature (n= 12H = 3329 df= 3 plt 0001) active layer thaw depth(n= 12 H = 2135 df= 3 plt 0001) and organic layers C (n= 4 H = 974 df= 3 p= 0021) were associatedwith the four different snow zones Post hoc tests revealedhigher temperatures in the Deep snow zone relative to theControl (p= 0009) the Int (p= 0001) and the Low snowzone (plt 0001 Table 1) Active layer depth data revealedsimilar results increasing in the Deep snow accumulationzone and decreasing as snowpack was experimentallyreduced Only in the Deep zone was the active layer thawdepth significantly (p= 0020) deeper than the Control zoneHowever along the snow accumulation gradient thaw depthsignificantly increased from Low to Deep plots (LowInt ndashp= 0021 LowDeep ndash plt 0001 Table 1) Soil moisturewas not correlated with snow accumulation possibly the
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
minus10
minus05
00
05
minus1 0 1 2NMDS1
NM
DS
2
Treatment Cont
Deep
Int
Low
N
05
10
15
10
20
30
40
C
Figure 2 Non-metric multidimensional scaling (NMDS) plot us-ing BrayndashCurtis dissimilarity matrices (stress= 0090 Shepard plotnon-metric R2
= 0992) Each point represents the bacterial com-munity structure within one of the 41 total samples used for DNAextraction from all soil depths (organic transition and mineral)Colours indicate C ranging from 14 (light blue) to 486 (dark blue) bubble size indicates N ranging from 009 (small)to 195 (large) and shapes indicate snow accumulation treatments(Control Deep Int Low) Ellipse centroids represent treatmentgroup means while the shape is defined by the covariance withineach group
result of surface hydrology at the site which was largelysaturated throughout the growing season In the organicsoil layers the C concentration of soil declined withincreased snow accumulation (LowDeep ndash p= 003) whilethe N concentration only slightly increased (LowDeep ndashp= 032) This resulted in a decreasing trend in C N ratiosacross snow accumulation treatment zones and relative tothe control (ControlDeep ndash p= 014 Table 1) Soil pH
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 463
Table 1 Abiotic characteristics of soil from snow accumulation treatments (Low=sim 25 less snowpack than the Control Int=sim 50 more snowpack than the Control Deep=sim 100 more snowpack than the Control) Values are means plusmn standard errors Soil chemicalproperties were obtained from samples used for DNA extraction while temperature and thaw depth were measured in situ (n= 12) Organicand mineral samples were analysed separately using the Nemenyi post hoc test Results are indicated by subscripts ldquoardquo ldquobrdquo or ldquocrdquo onlywhere plt 005
Treatment Soil layers Sample C N C N pH Temp at Thaw depthdepths (cm) 12 cm (C) (cm)
ControlOrganic (n= 4) 675plusmn 312 4521plusmn 109ab 101plusmn 020 5004plusmn 944 459plusmn 009
432plusmn 027b 5917plusmn 123bcMineral (n= 3) 2600plusmn 551 257plusmn 039 015plusmn 003 1767plusmn 134 515plusmn 005ab
LowOrganic (n= 4) 550plusmn 189 4663plusmn 073a 106plusmn 007 4459plusmn 254 444plusmn 008
292plusmn 024b 5092plusmn 320cMineral (n= 3) 2700plusmn 115 418plusmn 192 022plusmn 011 1942plusmn 065 516plusmn 020ab
IntOrganic (n= 3) 367plusmn 067 4059plusmn 243ab 117plusmn 025 3838plusmn 885 469plusmn 041
408plusmn 025b 6188plusmn 119abMineral (n= 3) 2367plusmn 203 258plusmn 049 014plusmn 002 1858plusmn 145 501plusmn 004a
DeepOrganic (n= 4) 600plusmn 370 3651plusmn 427b 140plusmn 007 2627plusmn 341 561plusmn 021
649plusmn 020a 6542plusmn 149aMineral (n= 4) 2400plusmn 442 165plusmn 019 010plusmn 001 1641plusmn 056 583plusmn 017b
the Shannon alpha diversity metrics from soil layer andeach treatment group were made using non-parametric two-sample t tests with 999 Monte Carlo permutations Beta di-versity was analysed by comparing BrayndashCurtis dissimilaritymatrices of bacterial abundance data from each sample to soilchemical properties as well as between soil layers and snowaccumulation treatments using adonis tests with 999 permu-tations Organic and mineral layers were also analysed sep-arately when comparing snow accumulation treatments andsoil chemical properties (Table 2) Analyses of soil chemicalproperties were further substantiated by Mantel tests againusing 999 permutations These data were visualized by cre-ating a non-metric multidimensional scaling (NMDS) plot(stress= 0090 Shepard plot non-metric R2
= 0992) in theR package phyloseq (McMurdie and Holmes 2013) usingthe same BrayndashCurtis dissimilarity matrices (Fig 2)
3 Results
31 Environmental changes
Significant differences in soil temperature (n= 12H = 3329 df= 3 plt 0001) active layer thaw depth(n= 12 H = 2135 df= 3 plt 0001) and organic layers C (n= 4 H = 974 df= 3 p= 0021) were associatedwith the four different snow zones Post hoc tests revealedhigher temperatures in the Deep snow zone relative to theControl (p= 0009) the Int (p= 0001) and the Low snowzone (plt 0001 Table 1) Active layer depth data revealedsimilar results increasing in the Deep snow accumulationzone and decreasing as snowpack was experimentallyreduced Only in the Deep zone was the active layer thawdepth significantly (p= 0020) deeper than the Control zoneHowever along the snow accumulation gradient thaw depthsignificantly increased from Low to Deep plots (LowInt ndashp= 0021 LowDeep ndash plt 0001 Table 1) Soil moisturewas not correlated with snow accumulation possibly the
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
Cont
Deep
Int
Low
minus10
minus05
00
05
minus1 0 1 2NMDS1
NM
DS
2
Treatment Cont
Deep
Int
Low
N
05
10
15
10
20
30
40
C
Figure 2 Non-metric multidimensional scaling (NMDS) plot us-ing BrayndashCurtis dissimilarity matrices (stress= 0090 Shepard plotnon-metric R2
= 0992) Each point represents the bacterial com-munity structure within one of the 41 total samples used for DNAextraction from all soil depths (organic transition and mineral)Colours indicate C ranging from 14 (light blue) to 486 (dark blue) bubble size indicates N ranging from 009 (small)to 195 (large) and shapes indicate snow accumulation treatments(Control Deep Int Low) Ellipse centroids represent treatmentgroup means while the shape is defined by the covariance withineach group
result of surface hydrology at the site which was largelysaturated throughout the growing season In the organicsoil layers the C concentration of soil declined withincreased snow accumulation (LowDeep ndash p= 003) whilethe N concentration only slightly increased (LowDeep ndashp= 032) This resulted in a decreasing trend in C N ratiosacross snow accumulation treatment zones and relative tothe control (ControlDeep ndash p= 014 Table 1) Soil pH
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

464 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tended to increase (became more neutral) with increasedsnow accumulation (LowDeep ndash p= 006) The changesin the mineral soil layers were less pronounced than in theorganic layers C N ratios again showed a decreasing trendas snow accumulation increased while soil pH increased inthe Deep zone but did not show a trend along the treatmentgradient (Table 1)
32 Bacterial community shifts
Some bacteria exhibited shifting trends in response to snowdepth both across treatments and relative to the controlwhile other community shifts were either not significant ordid not appear to be the result of the snow depth treat-ments (Figs 3 and S16ndashS20) Noticeable trends at the phy-lum level included a 16-fold increased abundance in Ver-rucomicrobia (p= 0068) a 21-fold increase in Actinobac-teria (p= 0083) and a 3290-fold increase in Chloroflexi(p= 0010) in the organic layers from the Low to Deepsnow zones Acidobacteria showed decreased abundancein all treatments relative to the Control with the Deepzone exhibiting the largest difference with a 198-fold de-crease (p= 0055 Fig 3) In the mineral layers signifi-cant increases in the phylum Chloroflexi (718-fold increasep= 0011) occurred from the Control to Deep zones whilesignificant decreases (284-fold decrease p= 0019) wereobserved from Control to Deep zones in the phylum Verru-comicrobia (Fig 3)
Bacterial abundance in each phylum correlated withat least one of the soil chemical properties we mea-sured ( C N C N or pH) The best overall pre-dictor was C correlating with four out of the sixphyla It showed negative relationships with Actinobacteria(R2= 038 plt 0001 Fig S4) and Chloroflexi (R2
= 034plt 0001 Fig S6) and positive relationships with Bac-teroidetes (R2
= 033 plt 0001 Fig S5) and Proteobac-teria (R2
= 032 plt 0001 Fig S2) Actinobacteria wasalso negatively correlated with N (R2
= 034 plt 0001Fig S4) and Chloroflexi was positively correlated with soilpH (R2
= 034 plt 0001 Fig S6) The best and only pre-dictor for Acidobacteria abundance was soil pH which corre-lated negatively (R2
= 046 plt 0001 Fig S1) Verrucomi-crobia abundance correlated positively with N (R2
= 036plt 0001 Fig S3)
Soil depth significantly affected bacterial relative abun-dance in all phyla except for Acidobacteria (Table S2) Theorganic layers were more abundant in Proteobacteria (159-fold difference plt 0001) Verrucomicrobia (148-fold dif-ference plt 0001) and Bacteroidetes (227-fold differencep= 0001) Phyla that were more abundant in the minerallayers were Actinobacteria (448-fold difference plt 0001)and Chloroflexi (1421-fold difference plt 0001)
Alpha diversity measured using the Shannon index wasfound to differ between soil layers (organicmineral ndashp= 0003) but not between snow accumulation treatments
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus04
minus02
00
02
04
00
02
04
06
minus06
minus04
minus02
00
00
05
10
15
20
25
minus05
00
05
10
0
2
4
6
8
Treatment
Deep
Int
Low
Mineral
Acidobacteria
Proteobacteria
Verrucomicrobia
Actinobacteria
Bacteroidetes
Chloroflexi
minus06
minus04
minus02
00
00
01
02
03
04
00
02
04
minus025
000
025
050
075
000
025
050
075
100
0
20
40
Re
lativ
e a
bu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Bacteria
ab
b
a
a
ab
b
a
ab
b
Figure 3 Averaged relative abundance of the six most abundantbacterial phylum relative to the control separated by snow accumu-lation treatment and in order of greatest abundance (top to bottompanels) Error bars represent standard error (standard error of con-trols ranged from 12929 in Chloroflexi to 0026 in Verrucomicro-bia) Significance determined by KruskalndashWallis tests is indicatedby asterisks (lowast=plt 01 lowastlowast=plt 005) while post hoc Nemenyitest results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except where therewere significant differences to the control
(Table S3) However beta diversity of bacterial communitiesvisualized by a NMDS plot of BrayndashCurtis dissimilarity in-dices constructed from community matrices (stress= 0090Shepard plot non-metric R2
= 0992 Fig 2) revealed signif-icant differences in community structure between all sam-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-
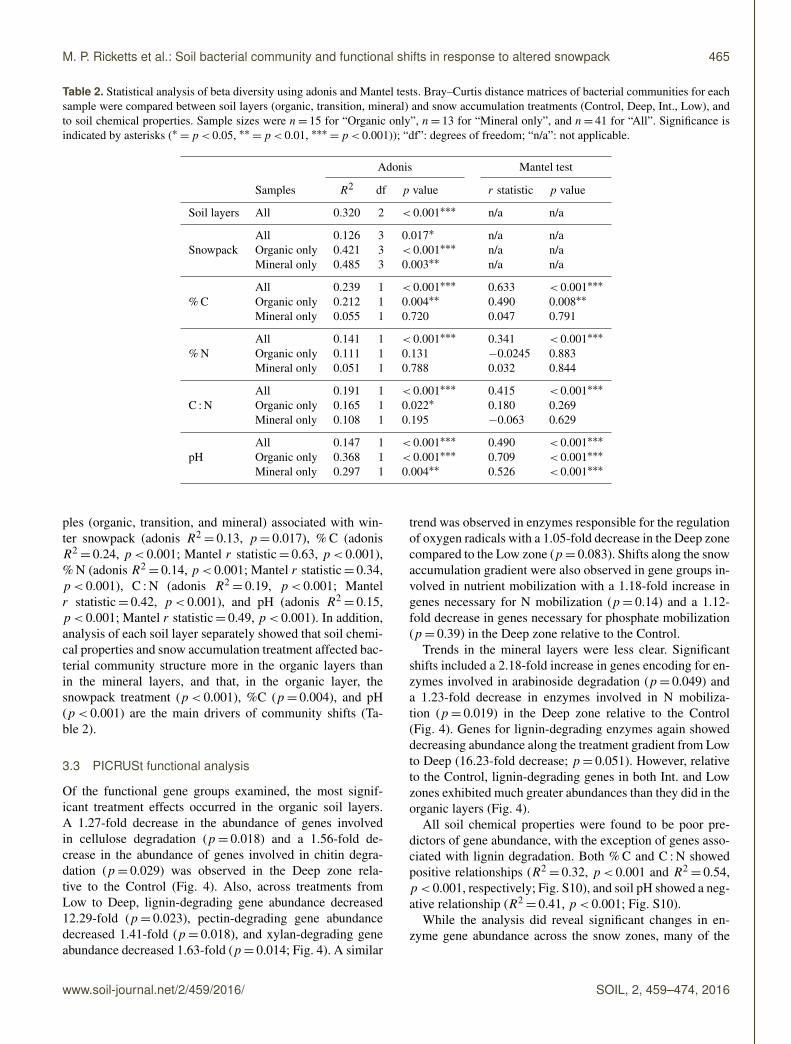
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 465
Table 2 Statistical analysis of beta diversity using adonis and Mantel tests BrayndashCurtis distance matrices of bacterial communities for eachsample were compared between soil layers (organic transition mineral) and snow accumulation treatments (Control Deep Int Low) andto soil chemical properties Sample sizes were n= 15 for ldquoOrganic onlyrdquo n= 13 for ldquoMineral onlyrdquo and n= 41 for ldquoAllrdquo Significance isindicated by asterisks (lowast=plt 005 lowastlowast=plt 001 lowastlowastlowast=plt 0001)) ldquodfrdquo degrees of freedom ldquonardquo not applicable
Adonis Mantel test
Samples R2 df p value r statistic p value
Soil layers All 0320 2 lt 0001lowastlowastlowast na na
SnowpackAll 0126 3 0017lowast na naOrganic only 0421 3 lt 0001lowastlowastlowast na naMineral only 0485 3 0003lowastlowast na na
CAll 0239 1 lt 0001lowastlowastlowast 0633 lt 0001lowastlowastlowast
Organic only 0212 1 0004lowastlowast 0490 0008lowastlowast
Mineral only 0055 1 0720 0047 0791
NAll 0141 1 lt 0001lowastlowastlowast 0341 lt 0001lowastlowastlowast
Organic only 0111 1 0131 minus00245 0883Mineral only 0051 1 0788 0032 0844
C NAll 0191 1 lt 0001lowastlowastlowast 0415 lt 0001lowastlowastlowast
Organic only 0165 1 0022lowast 0180 0269Mineral only 0108 1 0195 minus0063 0629
pHAll 0147 1 lt 0001lowastlowastlowast 0490 lt 0001lowastlowastlowast
Organic only 0368 1 lt 0001lowastlowastlowast 0709 lt 0001lowastlowastlowast
Mineral only 0297 1 0004lowastlowast 0526 lt 0001lowastlowastlowast
ples (organic transition and mineral) associated with win-ter snowpack (adonis R2
= 013 p= 0017) C (adonisR2= 024 plt 0001 Mantel r statistic= 063 plt 0001)
N (adonis R2= 014 plt 0001 Mantel r statistic= 034
plt 0001) C N (adonis R2= 019 plt 0001 Mantel
r statistic= 042 plt 0001) and pH (adonis R2= 015
plt 0001 Mantel r statistic= 049 plt 0001) In additionanalysis of each soil layer separately showed that soil chemi-cal properties and snow accumulation treatment affected bac-terial community structure more in the organic layers thanin the mineral layers and that in the organic layer thesnowpack treatment (plt 0001) C (p= 0004) and pH(plt 0001) are the main drivers of community shifts (Ta-ble 2)
33 PICRUSt functional analysis
Of the functional gene groups examined the most signif-icant treatment effects occurred in the organic soil layersA 127-fold decrease in the abundance of genes involvedin cellulose degradation (p= 0018) and a 156-fold de-crease in the abundance of genes involved in chitin degra-dation (p= 0029) was observed in the Deep zone rela-tive to the Control (Fig 4) Also across treatments fromLow to Deep lignin-degrading gene abundance decreased1229-fold (p= 0023) pectin-degrading gene abundancedecreased 141-fold (p= 0018) and xylan-degrading geneabundance decreased 163-fold (p= 0014 Fig 4) A similar
trend was observed in enzymes responsible for the regulationof oxygen radicals with a 105-fold decrease in the Deep zonecompared to the Low zone (p= 0083) Shifts along the snowaccumulation gradient were also observed in gene groups in-volved in nutrient mobilization with a 118-fold increase ingenes necessary for N mobilization (p= 014) and a 112-fold decrease in genes necessary for phosphate mobilization(p= 039) in the Deep zone relative to the Control
Trends in the mineral layers were less clear Significantshifts included a 218-fold increase in genes encoding for en-zymes involved in arabinoside degradation (p= 0049) anda 123-fold decrease in enzymes involved in N mobiliza-tion (p= 0019) in the Deep zone relative to the Control(Fig 4) Genes for lignin-degrading enzymes again showeddecreasing abundance along the treatment gradient from Lowto Deep (1623-fold decrease p= 0051) However relativeto the Control lignin-degrading genes in both Int and Lowzones exhibited much greater abundances than they did in theorganic layers (Fig 4)
All soil chemical properties were found to be poor pre-dictors of gene abundance with the exception of genes asso-ciated with lignin degradation Both C and C N showedpositive relationships (R2
= 032 plt 0001 and R2= 054
plt 0001 respectively Fig S10) and soil pH showed a neg-ative relationship (R2
= 041 plt 0001 Fig S10)While the analysis did reveal significant changes in en-
zyme gene abundance across the snow zones many of the
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

466 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus10
minus05
00
minus04
minus03
minus02
minus01
00
minus020
minus015
minus010
minus005
000
minus03
minus02
minus01
00
minus04
minus03
minus02
minus01
00
01
minus01
00
01
02
03
000
005
010
015
020
minus010
minus005
000
minus004
minus002
000
002
Re
l ge
ne
abu
nd
an
ce
ch
an
ge
c
on
tro
l
Organic
Lignin
Chitin
Cellulose
Pectins
Xylan
Arabinosides
N mobilization
P mobilization
Superoxides
minus1
0
1
2
000
025
000
005
010
015
020
minus01
00
01
02
minus03
minus02
minus01
00
01
02
03
00
05
10
15
minus020
minus015
minus010
minus005
000
minus02
minus01
00
minus010
minus005
000
Treatment
Deep
Int
Low
Mineral
Enzymes
aba
b
aabb
a
ab
b
ba
ab
a
abb
aab b
Figure 4 Averaged relative abundance of genes for enzyme func-tional groups relative to the control and separated by snow accumu-lation treatment Functional groups involved in soil organic matterdecomposition are ordered from recalcitrant to labile substrates (topto bottom) Error bars represent standard error (standard error ofcontrols ranged from 1220 in the lignin group to 0008 in the super-oxides group) Significance determined by KruskalndashWallis tests isindicated by asterisks (lowast=plt 01 lowastlowast=plt 005) while post hocNemenyi test results are indicated by ldquoardquo ldquobrdquo or ldquoabrdquo except wherethere were significant differences to the control
KEGG ortholog groups of enzymes targeted in this studywere either not found in any of the samples or were found invery low quantities including phenol oxidases peroxidasesand laccases (Table S1)
4 Discussion
This study documents changes in soil bacterial communitystructure in the active layer of moist acidic tundra in responseto long-term (18-year) experimental changes in winter pre-cipitation We examined how changes in bacterial commu-nity functional potential as a result of climate forcing fac-tors might affect SOM degradation and alter the C balanceof this Arctic tundra ecosystem Low temperatures in Arc-tic ecosystems limit soil C availability and decomposability(Conant et al 2011 Davidson and Janssens 2006) How-ever global-warming-induced permafrost thaw may partiallyalleviate this temperature limitation potentially releasinglarge amounts of C into the atmosphere via SOM decom-position and further increasing the rate of global warming(Lupascu et al 2013 2014a Luumltzow and Koumlgel-Knabner2009 Schuur et al 2008)
After 18 years of experimental winter snow addition bac-terial community structure and functional potential in Arc-tic moist acidic tundra changed under deeper winter snowaccumulation Our results indicate that increased snowpackreduced the abundance of genes associated with SOM de-composition in the organic soil layers suggesting a reducedSOM decomposition potential Possible explanations for thisfunctional shift may include (1) altered bacterial C substratepreferences towards more labile sources under lowered O2availability that would result in a decreased abundance ofgenes associated with SOM decomposition as well as (2) areduced amount of enzymatic machinery (and fewer genecopies Rocca et al 2014) necessary to accomplish simi-lar metabolic results as increased soil temperatures undersnow accumulation may alleviate kinetic limitations of en-zyme functioning (German et al 2012 Sinsabaugh et al2008)
41 Bacterial community shifts
Our results indicate that altered snow accumulation has a sig-nificant effect on soil bacterial community structure in Arcticmoist acidic tussock tundra ecosystems While large differ-ences in relative abundances were found between soil layers(Table S2) the most notable effects of snow accumulationoccurred in the organic layers For instance we observedshifts in the relative abundance in many of the most abun-dant phyla including Verrucomicrobia Acidobacteria andActinobacteria particularly in the Deep snow zone (Fig 3)Shifts in Verrucomicrobia were primarily driven by increasesin the order Chthoniobacterales in the Deep snow zones rel-ative to the Low snow zones This order contains facultativeaerobic heterotrophs able to utilize saccharide components of
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 467
plant biomass but unable to use amino acids or organic acidsother than pyruvate (Sangwan et al 2004) Shifts in Acti-nobacteria were dominated by the order ActinomycetalesGram-positive facultative bacteria that have been shown tostimulate the growth of ectomycorrhizal fungi a group of or-ganisms known to degrade recalcitrant C compounds (Good-fellow and Williams 1983 Maier et al 2004 Pridham andGottlieb 1948) While not as abundant the phylum Chlo-roflexi also responded to snowpack treatments increasing inabundance from Low to Deep snow zones (Fig 3) Shifts inChloroflexi were the result of increasing abundance of theclass Anaerolineae in the Deep zone Anaerolineae includegreen non-sulfur bacteria able to thrive in anaerobic environ-ments and have previously been found in similar cold water-saturated soils (Costello and Schmidt 2006) These resultsappear consistent with the increased soil moisture and de-creased partial pressure of O2 documented under increasedsnowpack at the study site (Blanc-Betes et al 2016)
These shifts in bacterial phyla indicate that even at thecoarsest level of phylogeny and a high degree of variancebetween samples deeper snow in winter and associatedchanges in soil conditions may be driving changes in the be-lowground community Bacterial community shifts may beresulting in potentially altered substrate use preference bydecomposers as well as different genetic functional activityThis is supported by other studies from Arctic soil and per-mafrost ecosystems that provide evidence of altered micro-bial community composition and rapid functional responseto temperature manipulations thawing soils or fertilizationtreatments (Deslippe et al 2012 Koyama et al 2014 Mack-elprang et al 2011) For example Actinobacteria abundancewas found to increase in response to both increased tempera-ture (Deslippe et al 2012) and in freshly thawed permafrostsoils (Mackelprang et al 2011) similar to the response weobserved in the Deep zone (Fig 3) Mackelprang et al (2011)also reported varying shifts in a wide array of functionalgenes in response to permafrost thaw In addition Koyamaet al (2014) documented a decrease in the oligotrophic Aci-dobacteria phylum in response to fertilizer soil inputs whichthey attributed to be a direct result of competition with co-piotrophic α- β- and γ -Proteobacteria which increased inabundance with fertilizer treatment While oligotrophic or-ganisms such as Acidobacteria are adapted to survive in lownutrient environments they are often outcompeted in morefertile soils by generalist copiotrophs (such as Proteobacte-ria) which are better equipped to harvest available nutrientsOur results did not show a clear pattern for Proteobacteriabut they do show that Acidobacteria abundance shifts asso-ciate negatively with Proteobacteria shifts in the Deep zonewhere C N soil values are lowest (most fertile Table 1 andFig 3)
Correlations between soil chemical characteristics ( C N C N and pH) and bacterial phylum abundance par-tially support findings reported in Fierer et al (2007) Theyidentified C mineralization rates (a proxy for C availabil-
ity) to be the best predictor of bacterial abundance in thedominant phyla including positive relationships with Bac-teroidetes and β-Proteobacteria and a negative relationshipwith Acidobacteria (Fierer et al 2007) We acknowledgethat C mineralization and availability differ from C in thatregardless of carbon concentration physical and chemicalfactors in the Arctic such as temperature limitations and hightannin concentrations may limit C mineralization (Davidsonand Janssens 2006 Schimel et al 1996) Physical protec-tion of SOM by soil aggregates and associations with organo-minerals also known to limit C mineralization does not playas large of a role in Arctic soils compared to other soil types(Houmlfle et al 2013 Ping et al 2015) Regardless of these dif-ference in C measurement our study did find weak positiverelationships between C and Proteobacteria (Fig S2) aswell as Bacteroidetes (Fig S5) similar to Fierer et al (2007)Interestingly although N can be a limiting factor for micro-bial growth N only correlated with two phyla positivelywith Verrucomicrobia (Fig S3) and negatively with Acti-nobacteria (Fig S4) While identifying individual abiotic fac-tors that may predict bacterial abundance at the phylum levelis informative it is important to recognize that often a varietyof interacting factors determine microbial community com-position and effects at the phylum scale may be too coarsefor adequate interpretation Our results suggest that whileC N (a proxy for SOM quality) is a poor indicator of individ-ual bacterial phylum abundance C and N (and in somecases soil pH) alone may be more relevant in these acidictundra soils More detailed studies that address the relation-ships between soil chemicalabiotic characteristics and mi-crobial community composition at finer phylogenetic scalesare needed to adequately identify dependable predictors
While the alpha diversity of soil bacterial communities viathe Shannon index did differ between soil layer it did notdiffer between snowpack treatment zones Also it does notelucidate community structural or functional differences be-tween samples and it fails to distinguish shifts in geneticpotential among treatments In contrast beta diversity anal-yses better revealed soil bacterial community responses tosnow accumulation Bacterial community structure signifi-cantly shifted between snowpack treatment zones at all soildepthslayers (Table 2) The NMDS plot (Fig 2) shows bac-terial community structures to be associated with the snowaccumulation treatment as soil chemical properties changed( C N C N and pH) indicating that bacterial beta di-versity may respond to indirect changes in soil chemistry inresponse to winter snow accumulation The initial effects ofincreased snowpack result in altered physical factors (greateractive layer thaw depth and increased soil temperatures andmoisture Blanc-Betes et al 2016) which may lead to in-creased SOM availability and faster enzyme activities withthe potential to enhance SOM decomposition Higher SOMmineralization may promote the documented shifts in above-ground plant communities and increased NPP (Natali et al2012 Sturm et al 2005 Anderson-Smith 2013) and vege-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

468 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
tation shifts to more shrubby species may alter the chemistryand quality of new litter inputs ultimately affecting decom-poser communities Moreover soil moisture and compactioncan reduce O2 diffusion into the soil inhibiting aerobicSOM decomposition (Blanc-Betes et al 2016 OrsquoBrien etal 2010) and altering bacterial community composition byselecting for microorganisms that utilize simple C substratesleaving behind complex organic compounds and plant poly-mers In addition tannins produced by expanding woodyshrubs may act to inhibit microbial activity (Schimel et al1996) further slowing decomposition This is supported bythe lower relative abundance of genes required for SOM de-composition in the Deep snow accumulation zone wherewe observed the most significant shifts in bacterial commu-nity composition (Figs 3 and 4) The balance between thesetwo competing processes and the functional shifts associ-ated with them will ultimately influence the C balance of thesystem
42 Functional shifts
To examine the influence of shifting bacterial abundanceson soil community functioning and the C balance of Arcticecosystems we focused on the genetic potential of the bac-terial community to produce enzymes required for the degra-dation of various forms of SOM We did this by using PI-CRUSt software to estimate functional gene abundance viaancestral state reconstruction (Langille et al 2013) Whilethis method does not provide direct measurements of geneabundance (eg does not account for horizontal gene trans-fer or unknown functionaltaxonomic linkages that may existin the sampled tundra soils) it does offer valuable insightsinto the functional capacities of bacterial communities us-ing 16S rRNA data (Langille et al 2013) Furthermore geneabundance in itself is not a direct measurement of gene ex-pression or enzyme activity (Wood et al 2015) However itdoes provide a measure of genetic potential and may be pos-itively correlated with enzyme activity and gene expression(Morris et al 2014 Neufeld et al 2001 Rocca et al 2014)Accurate measurement of enzymatic functional potential orgene expression would require a targeted metagenomic andmetatranscriptomic approach
Many bacterial genes encoding for enzymes associatedwith the degradation of lignin and other complex plant com-pounds (such as peroxides phenol oxidases and laccases)were not detected in this study This suggests that bacterialcommunities preferentially degrade microbial biomass andpolysaccharide polymers and that the decomposition of morerecalcitrant forms of C in Arctic soils is performed by othermicroorganisms such as fungi Fungi typically play a key rolein the degradation of recalcitrant organic matter by special-izing in the production of oxidative enzymes (Deslippe etal 2012 Morgado et al 2015) The absence of bacterialgenes that encode for peroxides phenol oxidases and lac-cases could also be due to the presence of tannins in the soil
which are common in the Alaskan floodplain and are pro-duced by encroaching shrub species (DeMarco et al 2014Schimel et al 1996) Tannic compounds have been shownto inhibit microbial activity and decrease decomposition bybinding to vital enzymes (Schimel et al 1996) If produc-tion of phenol oxidases and peroxides yields little to no ben-efit for bacteria in this ecosystem due to competition withfungi and interference from tannins and other phenolic com-pounds then genes encoding for these enzymes may be re-duced (Rocca et al 2014)
The PICRUSt-predicted copies of genes for enzymes re-sponsible for SOM decomposition while generally moreabundant in the organic layers (Table S2) were less abun-dant in the organic layers of the Deep snow zone than inthe Control and Low snow accumulation zones (Fig 4) Thegenes that are most affected encode enzymes required forthe breakdown of plant derived litter such as cellulose xy-lan or pectin all major constituents of plant cell walls Xy-lans in particular are common in woody plant tissues (Timell1967) The observed decrease in these genes in Deep snow-pack suggests bacterial preference of readily available sub-strates such as microbial biomass or root exudates (Sulli-van and Welker 2005 Sullivan et al 2007 2008) Produc-tion of these substrates may have been stimulated by in-creased soil temperatures and NPP predicted under a climatechange scenario and may require less energetic investmentin exo-enzyme production (Schimel and Weintraub 2003)The production of enzymes for the degradation of complexpolysaccharides is energetically demanding Therefore in anenergy- and nutrient-limited ecosystem such as the Arctictundra (Hobbie et al 2002 Jonasson et al 1999 Mack etal 2004 Shaver and Chapin III 1980 1986 Sistla et al2012) more labile substrates are likely preferable whichmay lead to accumulation of SOM and thus SOC (Lupascuet al 2013 2014a)
Our results indicating reduced decomposition potentialunder deeper snowpack are consistent with other long-termwarming and snow fence studies from Arctic tundra ecosys-tems that report zero net C loss (or even C gain) duringthe growing season (Natali et al 2012 2014 Sistla et al2013) We speculate that initial soil conditions likely favourdecomposer activity and decomposition rates increase in re-sponse to increased temperatures resulting in C loss Overtime changing soil conditions (eg increased moisture de-creased O2 availability changes in chemistry of litter inputs)may select for microorganisms that use anaerobic metabolicpathways such as methanogenesis (Blanc-Betes et al 2016)These hypoxic soil conditions would limit aerobic decom-position As bacterial communities increase the abundanceof genes encoding for enzymes involved in N mobilizationnewly available N would enhance microbial biomass produc-tion plant NPP and leaf litter N content and would induceplant community shifts (Pattison and Welker 2014 Schimeland Weintraub 2003 Welker et al 2005) A decrease inSOM decomposition is possibly supported by data from this
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 469
study which shows a decreased abundance of genes involvedin SOM decomposition in conjunction with trends suggestingincreased abundance of N mobilization genes in the organiclayers as snowpack increases (Fig 4)
Increased temperature may provide an alternate explana-tion to the decreased PICRUSt-predicted abundance of genesassociated with SOM decomposition in the organic layers ofthe Deep snow accumulation zone (Fig 4) Enzyme activ-ity is partially regulated by the rate of gene expression aswell as by post-transcriptional regulating factors which in-clude environmental factors (Gross et al 1989) MichaelisndashMenten enzyme kinetics are sensitive to temperature (Ger-man et al 2012) increasing the maximum rate of enzymeactivity (Vmax) by increasing the catalytic constant of the re-action (Razavi et al 2015) Increased Vmax may representan excess potential enzyme activity for the given substrate orgrowth conditions resulting in a down-regulation of genesrequired for the enzyme (eg Gonzalez-Meler et al 19992001) because fewer enzymes are needed to achieve simi-lar Vmax at higher temperatures Therefore increases in soiltemperature under deeper snow may partially explain the de-crease in PICRUSt-predicted abundance of genes requiredfor SOM decomposition (Table 1 and Fig 4)
43 Ecosystem response to snow accumulation
Whether bacterial communities are responding to changingplant inputs and corresponding altered SOM quality (de-creased C N Table 1) or whether they are directly alter-ing SOM chemistry through selective decomposition remainsunclear From the results of our study it is clear that in-creased snow accumulation may lead to changes in both bac-terial community composition and SOM chemistry in the or-ganic soil layers (Table 1 and Fig 3) Unlike other ecosys-tems where plants are the first responders to abiotic climatechange factors in the Arctic microbes are likely the firstresponders to changes in temperature by initially increas-ing nutrient mineralization These released nutrients facil-itate plant community shifts and increase ecosystem NPP(Chapin III et al 1995) Over time the combination of in-creased snow accumulation and soil compaction may leadto hypoxicanaerobic soil conditions (eg Blanc-Betes etal 2016) and further vegetative shifts to wet-sedge (Carex)species limiting SOM decomposition This in combinationwith a recent history of more recalcitrant plant litter inputscould result in re-accrual of SOC (eg Sistla et al 2012) ul-timately mitigating the positive feedback loop hypothesizedin the current literature (Davidson and Janssens 2006 Nataliet al 2014 Schuur et al 2009 Sturm et al 2005)
5 Conclusions
The results presented here support the hypothesis that bacte-rial community structure and function shift as a result of con-sistently deepened snowpack Increases in soil hypoxia un-
der deepened snow may have resulted in an increased abun-dance of anaerobic or facultative bacteria slowing decom-position Decreases in PICRUSt-predicted gene copies sug-gest that SOM decomposition may be slowed under accu-mulated snow and bacterial community substrate preferencemay shift to more labile compounds Concentrations of Cand N as opposed to C N better explained bacterial com-munity responses to snowpack treatments Together these re-sults strongly suggest that soil decomposers of moist acidictundra are key in determining the direction and magnitude ofpermafrost C feedbacks on the climate system
6 Code availability
All computer script text files used in QIIME and R pack-ages are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
7 Data availability
All data used in this report are publicly accessible throughtwo separate data repositories The 16S rRNA gene se-quences derived from Illumina Mi-Seq sequencing have beendeposited in the NCBI Sequence Read Archive (SRA) underaccession number SRP068302 All raw data and pertinent in-termediate files are available via the NSF Arctic Data Centerdoi1018739A2DP96 (Ricketts et al 2016)
The Supplement related to this article is available onlineat doi105194soil-2-459-2016-supplement
Author contributions Michael P Ricketts Jeffrey M Welkerand Miquel A Gonzalez-Meler designed the experimentRachel S Poretsky provided expertise and insight into the bioin-formatics and data analyses Michael P Ricketts performed allsample collections lab work and data analyses Michael P Rickettsprepared the manuscript with contributions from all co-authors
Acknowledgements This research was supported by fundingfrom the Department of Energy Terrestrial Ecosystem ScienceProgram to Miquel A Gonzalez-Meler (DE-SC 0006607) Jef-frey M Welker initiated and maintained the snow fence experimentwith NSF funds beginning in 1994 (9412782 0632184 06125340856728) Michael P Ricketts received additional support fromthe Elmer Hadley Graduate Research Award the WC and MayPreble Deiss Award for Graduate Research and the BodmerInternational Travel Award We would like to thank membersof the Stable Isotope Laboratory at the University of Illinois atChicago (UIC) particularly Jessica Rucks and Gerard Aubanellfor assistance with field worksample collection and processingWe thank Elena Blanc-Betes Douglas Johnston Douglas Lynchand Charlie Flower for advice on experimental design analysesand comments on the manuscript We would also like to thank
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

470 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
DrsquoArcy Meyer Dombard (Department of Earth and Environ-mental Sciences UIC) for use of her lab and soil expertiseand Imrose Kauser (Microbial Ecology lab UIC) for help withthe bioinformatics Finally we thank Olivia L Miller (PurdueUniversity) Niccole Van Hoey (University of Alaska Anchorage)all of the staff at Toolik Field Station and the field and lab UICundergraduate assistants Ben Thurnhoffer Andres Davila andBriana Certa
Edited by K KalbitzReviewed by three anonymous referees
References
Anderson-Smith M Remotely-sensed spectral data linked to in-creasing shrub abundance and greater growing season car-bon uptake in Alaskan Arctic tundra UMI 154257 Mas-terrsquos Thesis available at httpsearchproquestcomdocview1426631801 (last access 6 September 2016) 2013
Anisimov O Vaughan D Callaghan T Furgal C MarchantH Prowse T Vilhjalmsson H and Walsh J Polar regions(Arctic and Antarctic) in Climate Change 2007 Impacts adap-tation and vulnerability Contribution of working group II to thefourth assessment report of the Intergovernmental Panel on Cli-mate Change edited by Parry M L Canziani O F PalutikofJ P van der Linden P J and Hanson C E Cambridge Uni-versity Press Cambridge 653ndash685 2007
Arft A Walker M Gurevitch J Alatalo J M Bret-Harte MDale M Diemer M Gugerli F Henry G H R Jones M HHollister R D Jonsdottir I S Laine K Levesque E Mar-ion G M Molau U Molgaard P Nordenhall U RaszhivinV Robinson C H Starr G Stenstrom A Stenstrom M Tot-land O Turner P L Walker L J Webber P J Welker J Mand Wookey P A Responses of tundra plants to experimentalwarming meta-analysis of the international tundra experimentEcol Monogr 69 491ndash511 1999
Aronesty E ea-utils Command-line tools for processing bio-logical sequencing data httpsgithubcomExpressionAnalysisea-utils (last access 6 September 2016) 2011
Batjes N Total carbon and nitrogen in soils of the world Eur JSoil Sci 47 151ndash163 1996
Blanc-Betes E Welker J M Sturchio N C Chanton J Pand Gonzalez-Meler M A Winter precipitation and snowaccumulation drive the methane sink or source strength ofArctic tussock tundra Global Change Biol 22 2818ndash2833doi101111gcb13242 2016
Bret-Harte M S Shaver G R Zoerner J P Johnstone J FWagner J L Chavez A S Gunkelman IV R F Lippert S Cand Laundre J A Developmental plasticity allows Betula nanato dominate tundra subjected to an altered environment Ecology82 18ndash32 2001
Caporaso J G Kuczynski J Stombaugh J Bittinger K Bush-man F D Costello E K Fierer N Gonzalez A GoodrichJ K Gordon J I Huttley G A Kelley S T KnightsD Koenig J E Ley R E Lozupone C A McDonaldD Muegge B D Pirrung M Reeder J Sevinsky J RTurnbaugh P J Walters W A Widmann J Yatsunenko TZaneveld J and Knight R QIIME allows analysis of high-
throughput community sequencing data Nat Meth 7 335ndash336doi101038nmethf303 2010
Caporaso J G Lauber C L Walters W A Berg-Lyons DHuntley J Fierer N Owens S M Betley J Fraser LBauer M Gormley N Gilbert J A Smith G and KnightR Ultra-high-throughput microbial community analysis on theIllumina HiSeq and MiSeq platforms ISME J 6 1621ndash1624doi101038ismej20128 2012
Chapin III F S Shaver G R Giblin A E Nadelhoffer KJ and Laundre J A Responses of Arctic tundra to experi-mental and observed changes in climate Ecology 76 694ndash711doi1023071939337 1995
Conant R T Ryan M G Aringgren G I Birge H E DavidsonE A Eliasson P E Evans S E Frey S D Giardina C PHopkins F M Hyvoumlnen R Kirschbaum M U F LavalleeJ M Leifeld J Parton W J Steinweg J M Wallenstein MD Wetterstedt J Aring M and Bradford M A Temperature andsoil organic matter decomposition rates ndash synthesis of currentknowledge and a way forward Global Change Biol 17 3392ndash3404 doi101111j1365-2486201102496x 2011
Costello E K and Schmidt S K Microbial diversity in alpinetundra wet meadow soil novel Chloroflexi from a cold water-saturated environment Environ Microbiol 8 1471ndash1486doi101111j1462-2920200601041x 2006
Czimczik C I and Welker J M Radiocarbon content of CO2respired from high Arctic tundra in Northwest GreenlandArct Antarct Alp Res 42 342ndash350 doi1016571938-4246-423342 2010
Davidson E A and Janssens I A Temperature sensitivity of soilcarbon decomposition and feedbacks to climate change Nature440 165ndash173 doi101038nature04514 2006
DeMarco J Mack M and Bret-Harte M Effects of arctic shrubexpansion on biophysical versus biogeochemical drivers of litterdecomposition Ecology 95 1861ndash1875 2014
Deslippe J R Hartmann M Simard S W and Mohn WW Long-term warming alters the composition of Arctic soilmicrobial communities FEMS Microbiol Ecol 82 303ndash315doi101111j1574-6941201201350x 2012
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBjorkman A D Callaghan T V Collier L S Cooper EJ Cornelissen J H C Day T A Fosaa A M Gould WA Greacutetarsdoacutettir J Harte J Hermanutz L Hik D S Hof-gaard A Jarrad F Joacutensdoacutettir I S Keuper F Klanderud KKlein J A Koh S Kudo G Lang S I Loewen V May JL Mercado J Michelsen A Molau U Myers-Smith I HOberbauer S F Pieper S Post E Rixen C Robinson CH Schmidt N M Shaver G R Stenstroumlm A Tolvanen ATotland Oslash Troxler T Wahren C H Webber P J WelkerJ M and Wookey P A Global assessment of experimen-tal climate warming on tundra vegetation Heterogeneity overspace and time Ecol Lett 15 164ndash175 doi101111j1461-0248201101716x 2012a
Elmendorf S C Henry G H R Hollister R D Bjoumlrk R GBoulanger-Lapointe N Cooper E J Cornelissen J H C DayT A Dorrepaal E Elumeeva T G Gill M Gould W AHarte J Hik D S Hofgaard A Johnson D R Johnstone JF Joacutensdoacutettir I S Jorgenson J C Klanderud K Klein J AKoh S Kudo G Lara M Leacutevesque E Magnuacutesson B MayJ L Mercado-Diacuteaz J A Michelsen A Molau U Myers-
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 471
Smith I H Oberbauer S F Onipchenko V G Rixen CMartin Schmidt N Shaver G R Spasojevic M J THORNoacuterhalls-doacutettir THORN E Tolvanen A Troxler T Tweedie C E VillarealS Wahren C-H Walker X Webber P J Welker J M andWipf S Plot-scale evidence of tundra vegetation change andlinks to recent summer warming Nat Clim Change 2 453ndash457doi101038nclimate1465 2012b
Fahnestock J T Povirk K A and Welker J M Abi-otic and biotic effects of increased litter accumulation inarctic tundra Ecography 23 623ndash631 doi101111j1600-05872000tb00181x 2000
Fierer N Bradford M A and Jackson R B Toward an eco-logical classification of soil bacteria Ecology 88 1354ndash1364doi10189005-1839 2007
Frey S D Lee J Melillo J M and Six J The temperature re-sponse of soil microbial efficiency and its feedback to climateNat Clim Change 3 395ndash398 doi101038nclimate17962013
German D P Marcelo K R B Stone M M and Allison S DThe MichaelisndashMenten kinetics of soil extracellular enzymes inresponse to temperature a cross-latitudinal study Global ChangeBiol 18 1468ndash1479 doi101111j1365-2486201102615x2012
Gilbert J A Jansson J K and Knight R The Earth Micro-biome project successes and aspirations BMC Biol 12 69doi101186s12915-014-0069-1 2014
Gonzagravelez-Meler M A Ribas-Carbo M Giles L and Siedow JN The effect of growth and measurement temperature on the ac-tivity of the alternative respiratory pathway Plant Physiol 120765ndash772 doi101104pp1203765 1999
Gonzagravelez-Meler M A Giles L Thomas R B and Siedow J NMetabolic regulation of leaf respiration and alternative pathwayactivity in response to phosphate supply Plant Cell Environ 24205ndash215 doi101046j1365-3040200100674x 2001
Goodfellow M and Williams S T Ecology of acti-nomycetes Annu Rev Microbiol 37 189ndash216doi101146annurevmi37100183001201 1983
Graham D E Wallenstein M D Vishnivetskaya T A WaldropM P Phelps T J Pfiffner S M Onstott T C Whyte L GRivkina E M Gilichinsky D A Elias D A Mackelprang RVerBerkmoes N C Hettich R L Wagner D WullschlegerS D and Jansson J K Microbes in thawing permafrost theunknown variable in the climate change equation ISME J 6709ndash712 doi101038ismej2011163 2012
Gross R Arico B and Rappuoli R Families of bacterialsignal-transducing proteins Mol Microbiol 3 1661ndash1667doi101111j1365-29581989tb00152x 1989
Hinzman L D Bettez N D Bolton W R Chapin F S Dyurg-erov M B Fastie C L Griffith B Hollister R D HopeA Huntington H P Jensen A M Jia G J Jorgenson TKane D L Klein D R Kofinas G Lynch A H Lloyd AH McGuire A D Nelson F E Oechel W C OsterkampT E Racine C H Romanovsky V E Stone R S StowD A Sturm M Tweedie C E Vourlitis G L Walker MD Walker D A Webber P J Welker J M Winker K Sand Yoshikawa K Evidence and implications of recent climatechange in Northern Alaska and other Arctic regions ClimaticChange 72 251ndash298 doi101007s10584-005-5352-2 2005
Hobbie S E Nadelhoffer K J and Houmlgberg P A syn-thesis The role of nutrients as constraints on carbon bal-ances in boreal and arctic regions Plant Soil 242 163ndash170doi101023A1019670731128 2002
Houmlfle S Rethemeyer J Mueller C W and John S Organicmatter composition and stabilization in a polygonal tundra soil ofthe Lena Delta Biogeosciences 10 3145ndash3158 doi105194bg-10-3145-2013 2013
Hopkins F M Torn M S and Trumbore S E Warm-ing accelerates decomposition of decades-old carbon in for-est soils P Natl Acad Sci USA 109 E1753ndashE1761doi101073pnas1120603109 2012
Hopkins F M Gonzalez-Meler M A Flower C E Lynch DJ Czimczik C Tang J and Subke J-A Ecosystem-level con-trols on root-rhizosphere respiration New Phytol 199 339ndash3512013
Hugelius G Tarnocai C Broll G Canadell J G Kuhry Pand Swanson D K The Northern Circumpolar Soil CarbonDatabase spatially distributed datasets of soil coverage and soilcarbon storage in the northern permafrost regions Earth SystSci Data 5 3ndash13 doi105194essd-5-3-2013 2013
IPCC IPCC special report on land use land-use change andforestry edited by Watson R T Noble I R Bolin B Ravin-dranath N H Verardo D J and Dokken D J CambridgeUniversity Press Cambridge 3ndash24 2000
Jonasson S Michelsen A and Schmidt I K Coupling of nu-trient cycling and carbon dynamics in the Arctic integration ofsoil microbial and plant processes Appl Soil Ecol 11 135ndash146doi101016S0929-1393(98)00145-0 1999
Jones M H Fahnestock J T Walker D A Walker M D andWelker J M Carbon dioxide fluxes in moist and dry Arctic tun-dra during the snow-free season Responses to increases in sum-mer temperature and winter snow accumulation Arctic AntarctAlp Res 30 373ndash380 1998
Koyama A Wallenstein M D Simpson R T and Moore J CSoil bacterial community composition altered by increased nutri-ent availability in Arctic tundra soils Front Microbiol 5 1ndash16doi103389fmicb201400516 2014
Langille M G I Zaneveld J Caporaso J G McDonald DKnights D Reyes J A Clemente J C Burkepile D E VegaThurber R L Knight R Beiko R G and Huttenhower CPredictive functional profiling of microbial communities using16S rRNA marker gene sequences Nat Biotechnol 31 814ndash821 doi101038nbt2676 2013
Lauber C L Hamady M Knight R and Fierer NPyrosequencing-based assessment of soil pH as a predictor ofsoil bacterial community structure at the continental scale ApplEnviron Microb 75 5111ndash5120 doi101128AEM00335-092009
Leffler A J and Welker J M Long-term increases in snow packelevate leaf N and photosynthesis in Salix arctica responses toa snow fence experiment in the High Arctic of NW GreenlandEnviron Res Lett 8 1ndash10 doi1010881748-9326820250232013
Liston G E and Hiemstra C A The Changing Cryosphere Pan-Arctic Snow Trends (1979ndash2009) J Climate 24 5691ndash5712doi101175JCLI-D-11-000811 2011
Lupascu M Welker J M Seibt U Maseyk K Xu X andCzimczik C I High Arctic wetting reduces permafrost car-
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

472 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
bon feedbacks to climate warming Nat Clim Change 4 51ndash56doi101038NCLIMATE2058 2013
Lupascu M Welker J M and Czimczik C I Rates and radio-carbon content of summer ecosystem respiration to long-termdeeper snow in the High Arctic of NW Greenland J GeophysRes-Biogeo 119 1180ndash1194 doi1010022013JG0024332014a
Lupascu M Welker J M Seibt U Xu X Velicogna I Lind-sey D S and Czimczik C I The amount and timing of pre-cipitation control the magnitude seasonality and sources (14C)of ecosystem respiration in a polar semi-desert northwesternGreenland Biogeosciences 11 4289ndash4304 doi105194bg-11-4289-2014 2014b
Luumltzow M and Koumlgel-Knabner I Temperature sensitivity of soilorganic matter decomposition ndash what do we know Biol FertSoils 46 1ndash15 doi101007s00374-009-0413-8 2009
Mack M C Schuur E A G Bret-Harte M S Shaver G R andChapin III F S Ecosystem carbon storage in arctic tundra re-duced by long-term nutrient fertilization Nature 431 440ndash443doi101038nature02887 2004
Mackelprang R Waldrop M P DeAngelis K M David MM Chavarria K L Blazewicz S J Rubin E M and Jans-son J K Metagenomic analysis of a permafrost microbial com-munity reveals a rapid response to thaw Nature 480 368ndash371doi101038nature10576 2011
Maier A Riedlinger J Fiedler H-P and Hampp R Actinomyc-etales bacteria from a spruce stand characterization and effectson growth of root symbiotic and plant parasitic soil fungi in dualculture Mycol Prog 3 129ndash136 2004
Martiny A C Treseder K and Pusch G Phylogenetic conser-vatism of functional traits in microorganisms ISME J 7 830ndash838 doi101038ismej2012160 2013
McMurdie P J and Holmes S phyloseq An R Pack-age for Reproducible Interactive Analysis and Graph-ics of Microbiome Census Data PLoS ONE 8 e61217doi101371journalpone0061217 2013
Morgado L N Semenova T A Welker J M Walker M DSmets E and Geml J Summer temperature increase has dis-tinct effects on the ectomycorrhizal fungal communities of moisttussock and dry tundra in Arctic Alaska Global Change Biol21 959ndash972 doi101111gcb12716 2015
Morris R Schauer-Gimenez A Bhattad U Kearney C Stru-ble C A Zitomer D and Maki J S Methyl coenzyme M re-ductase (mcrA) gene abundance correlates with activity measure-ments of methanogenic H2CO2-enriched anaerobic biomassMicrob Biotechnol 7 77ndash84 doi1011111751-7915120942014
Natali S M Schuur E A G and Rubin R L Increasedplant productivity in Alaskan tundra as a result of experimen-tal warming of soil and permafrost J Ecol 100 488ndash498doi101111j1365-2745201101925x 2012
Natali S M Schuur E A G Webb E E Hicks Pries C Eand Crummer K G Permafrost degradation stimulates carbonloss from experimentally warmed tundra Ecology 95 602ndash6082014
Neufeld J D Driscoll B T Knowles R and Archibald F SQuantifying functional gene populations comparing gene abun-dance and corresponding enzymatic activity using denitrificationand nitrogen fixation in pulp and paper mill effluent treatment
systems Can J Microbiol 47 925ndash934 doi101139w01-0922001
Nowinski N S Taneva L Trumbore S E and Welker J MDecomposition of old organic matter as a result of deeper activelayers in a snow depth manipulation experiment Oecologia 163785ndash792 doi101007s00442-009-1556-x 2010
OrsquoBrien S L Jastrow J D Grimley D A and Gonzalez-MelerM A Moisture and vegetation controls on decadal-scale ac-crual of soil organic carbon and total nitrogen in restored grass-lands Global Change Biol 16 2573ndash2588 doi101111j1365-2486200902114x 2010
Pattison R R and Welker J M Differential ecophysiological re-sponse of deciduous shrubs and a graminoid to long-term ex-perimental snow reductions and additions in moist acidic tundraNorthern Alaska Oecologia 174 339ndash350 doi101007s00442-013-2777-6 2014
Pearson R G Phillips S J Loranty M M Beck P S ADamoulas T Knight S J and Goetz S J Shifts in Arcticvegetation and associated feedbacks under climate change NatClim Change 3 673ndash677 doi101038nclimate1858 2013
Ping C L Bockheim J G Kimble J M Michaelson G J andWalker D A Characteristics of cryogenic soils along a latitu-dinal transect in arctic Alaska J Geophys Res 103 28917ndash28928 doi10102998JD02024 1998
Ping C L Michaelson G J Jorgenson M T Kimble J MEpstein H Romanovsky V E and Walker D A High stocksof soil organic carbon in the North American Arctic region NatGeosci 1 615ndash619 doi101038ngeo284 2008
Ping C L Jastrow J D Jorgenson M T Michaelson G J andShur Y L Permafrost soils and carbon cycling SOIL 1 147ndash171 doi105194soil-1-147-2015 2015
Pridham T G and Gottlieb D The utilization of carbon com-pounds by actinomycetales as an aid for species determinationJ Bacteriol 56 107ndash114 1948
Raich J Potter C and Bhagawati D Interannual variability inglobal soil respiration 1980ndash94 Global Change Biol 8 800ndash812 2002
Razavi B S Blagodatskaya E Kuzyakov Y Nonlinear temper-ature sensitivity of enzyme kinetics explains canceling effect ndash acase study on loamy haplic Luvisol Front Microbiol 6 1ndash13doi103389fmicb201501126 2015
Ricketts M P Poretsky R S Welker J M and Gonzalez-MelerM A Soil bacterial community and functional shifts in responseto altered snow pack in moist acidic tundra of Northern AlaskaNSF Arctic Data Center doi1018739A2DP96 2016
Rocca J D Hall E K Lennon J T Evans S E Waldrop M PCotner J B Nemergut D R Graham E B and WallensteinM D Relationships between protein-encoding gene abundanceand corresponding process are commonly assumed yet rarelyobserved ISME J 9 1693ndash1699 doi101038ismej20142522014
Rogers M C Sullivan P F and Welker J M Exchange toincreasing levels of winter snow depth in the high Arctic ofNorthwest Greenland Arctic Antarct Alp Res 43 95ndash106doi1016571938-4246-43195 2011
Sangwan P Chen X Hugenholtz P and Janssen P H Chtho-niobacter flavus gen nov sp nov the First Pure-Culture Repre-sentative of Subdivision Two Spartobacteria classis nov of the
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack 473
Phylum Verrucomicrobia Appl Environ Microbiol 70 5875ndash5881 doi101128AEM70105875-58812004 2004
Schimel J P and Weintraub M N The implications of exoen-zyme activity on microbial carbon and nitrogen limitation insoil a theoretical model Soil Biol Biochem 35 549ndash563doi101016S0038-0717(03)00015-4 2003
Schimel J P and Schaeffer S M Microbial controlover carbon cycling in soil Front Microbiol 3 1ndash11doi103389fmicb201200348 2012
Schimel J P Cleve K Van Cates R G Clausen T P and Re-ichardt P B Effects of balsam poplar (Populus balsamifera)tannins and low molecular weight phenolics on microbial activ-ity in taiga floodplain soil implications for changes in N cyclingduring succession Can J Bot 74 84ndash90 doi101139b96-0121996
Schimel J P Bilbrough C and Welker J M Increased snowdepth affects microbial activity and nitrogen mineralization intwo Arctic tundra communities Soil Biol Biochem 36 217ndash227 doi101016jsoilbio200309008 2004
Schuur E A G Bockheim J Canadell J G Euskirchen EField C B Goryachkin S V Hagemann S Kuhry P LafleurP M Lee H Mazhitova G Nelson F E Rinke A Ro-manovsky V E Shiklomanov N Tarnocai C Venevsky SVogel J G and Zimov S A Vulnerability of permafrost car-bon to climate change Implications for the global carbon cycleBioscience 58 701ndash714 doi101641B580807 2008
Schuur E A G Vogel J G Crummer K G Lee H SickmanJ O and Osterkamp T E The effect of permafrost thaw onold carbon release and net carbon exchange from tundra Nature459 556ndash559 doi101038nature08031 2009
Semenova T A Morgado L N Welker J M WalkerM D Smets E and Geml J Long-term experimentalwarming alters community composition of ascomycetes inAlaskan moist and dry arctic tundra Mol Ecol 24 424ndash437doi101111mec13045 2015
Shaver G R and Chapin III F S Response to fertilization by var-ious plant growth forms in an Alaskan tundra Nutrient accumu-lation and growth Ecology 61 662ndash675 doi10230719374321980
Shaver G R and Chapin III F S Effect of fertilizer on productionand biomass of the tussock tundra Alaska USA Arct AlpRes 18 261ndash268 doi1023071550883 1986
Sinsabaugh R L Lauber C L Weintraub M N Ahmed B Al-lison S D Crenshaw C Contosta A R Cusack D Frey SGallo M E Gartner T B Hobbie S E Holland K KeelerB L Powers J S Stursova M Takacs-Vesbach C WaldropM P Wallenstein M D Zak D R and Zeglin L H Stoi-chiometry of soil enzyme activity at global scale Ecol Lett 111252ndash1264 doi101111j1461-0248200801245x 2008
Sistla S A Asao S and Schimel J P Detecting micro-bial N-limitation in tussock tundra soil Implications for Arc-tic soil organic carbon cycling Soil Biol Biochem 55 78ndash84doi101016jsoilbio201206010 2012
Sistla S A Moore J C Simpson R T Gough L Shaver G Rand Schimel J P Long-term warming restructures Arctic tundrawithout changing net soil carbon storage Nature 497 615ndash618doi101038nature12129 2013
Soil Survey Division Staff Soil survey manual Handbook SoilConservation Service US Department of Agriculture 18 pp
httpwwwnrcsusdagovwpsportalnrcsdetailsoilsrefcid=nrcs142p2_054262 (last access 6 September 2016) 1993
Soil Survey Staff Natural Resources Conservation Service UnitedStates Department of Agriculture Web Soil Survey availableat httpwebsoilsurveynrcsusdagov last access 17 Novem-ber 2015
Sturm M Schimel J Michaelson G Welker J M Ober-bauer S F Liston G E Fahnestock J and RomanovskyV E Winter biological processes could help convert Arctictundra to shrubland Bioscience 55 17ndash26 doi1016410006-3568(2005)055[0017WBPCHC]20CO2 2005
Sullivan P F and Welker J M Warming chambers stimulate earlyseason growth of an arctic sedge Results of a minirhizotron fieldstudy Oecologia 142 616ndash626 doi101007s00442-004-1764-3 2005
Sullivan P F Sommerkorn M Rueth H M Nadelhoffer K JShaver G R and Welker J M Climate and species affectfine root production with long-term fertilization in acidic tus-sock tundra near Toolik Lake Alaska Oecologia 153 643ndash652doi101007s00442-007-0753-8 2007
Sullivan P F Arens S J T Chimner R A and WelkerJ M Temperature and microtopography interact to controlcarbon cycling in a high Arctic fen Ecosystems 11 61ndash76doi101007s10021-007-9107-y 2008
Tape K Sturm M and Racine C The evidence for shrub ex-pansion in Northern Alaska and the Pan-Arctic Global ChangeBiol 12 686ndash702 doi101111j1365-2486200601128x2006
Tape K D Hallinger M Welker J M and Ruess R W Land-scape heterogeneity of shrub expansion in Arctic Alaska Ecosys-tems 15 711ndash724 doi101007s10021-012-9540-4 2012
Tarnocai C Canadell J G Schuur E A G Kuhry P Mazhi-tova G and Zimov S Soil organic carbon pools in the north-ern circumpolar permafrost region Global Biogeochem Cy 231ndash11 doi1010292008GB003327 2009
Timell T E Recent progress in the chemistry of wood hemicellu-loses Wood Sci Technol 1 45ndash70 doi101007BF005922551967
Wahren C Vegetation responses in Alaskan arctic tundra af-ter 8 years of a summer warming and winter snow ma-nipulation experiment Global Change Biol 11 537ndash552doi101111j1365-2486200500927x 2005
Waldrop M P Wickland K P White R Berhe A A HardenJ W and Romanovsky V E Molecular investigations intoa globally important carbon pool Permafrost-protected car-bon in Alaskan soils Global Change Biol 16 2543ndash2554doi101111j1365-2486200902141x 2010
Walker M and Wahren C Plant community responses to exper-imental warming across the tundra biome P Natl Acad SciUSA 103 1342ndash1346 doi101073pnas0503198103 2006
Walker M Walker D Welker J Arft A Bardsley T BrooksP Fahnestock J T Jones M Losleben M Parsons A Seast-edt T and Turner P Long-term experimental manipulation ofwinter snow regime and summer temperature in arctic and alpinetundra Hydrol Process 13 2315ndash2330 1999
Wallenstein M D McMahon S and Schimel J Bacterial andfungal community structure in Arctic tundra tussock and shrubsoils FEMS Microbiol Ecol 59 428ndash435 doi101111j1574-6941200600260x 2007
wwwsoil-journalnet24592016 SOIL 2 459ndash474 2016
474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-

474 M P Ricketts et al Soil bacterial community and functional shifts in response to altered snowpack
Welker J M Fahnestock J T and Jones M H Annual CO2 fluxin dry and moist arctic tundra Field responses to increases insummer temperatures and winter snow depth Climatic Change44 139ndash150 doi101023a1005555012742 2000
Welker J M Fahnestock J T Sullivan P F and Chimner RA Leaf mineral nutrition of Arctic plants in response to warm-ing and deeper snow in northern Alaska Oikos 109 167ndash177doi101111j0030-1299200513264x 2005
Wood S A Almaraz M Bradford M A McGuire K LNaeem S Neill C Palm C A Tully K L and Zhou JFarm management not soil microbial diversity controls nutrientloss from smallholder tropical agriculture Front Microbiol 61ndash10 doi103389fmicb201500090 2015
Xia J Chen J Piao S Ciais P Luo Y and Wan S Terres-trial carbon cycle affected by non-uniform climate warming NatGeosci 7 173ndash180 doi101038ngeo2093 2014
Zak D R and Kling G W Microbial community composition andfunction across an arctic tundra landscape Ecology 87 1659ndash1670 2006
SOIL 2 459ndash474 2016 wwwsoil-journalnet24592016
- Abstract
- Introduction
- Methods
-
- Site description and sample collection
- DNA extraction sequencing and analysis
- Statistical analyses
-
- Results
-
- Environmental changes
- Bacterial community shifts
- PICRUSt functional analysis
-
- Discussion
-
- Bacterial community shifts
- Functional shifts
- Ecosystem response to snow accumulation
-
- Conclusions
- Code availability
- Data availability
- Author contributions
- Acknowledgements
- References
-


















