
Size of the Unwound Region of DNA in Escherichia … of the Unwound Region of DNA in Escherichia...
8
10.1101/SQB.1983.047.01.054 Access the most recent version at doi: 1983 47: 455-461 Cold Spring Harb Symp Quant Biol H.B. Gamper and J.E. Hearst Ternary Complexes RNA Polymerase and Calf Thymus RNA Polymerase II Escherichia coli Size of the Unwound Region of DNA in References http://symposium.cshlp.org/content/47/455.refs.html This article cites 26 articles, 8 of which can be accessed free at: service Email alerting click here the box at the top right corner of the article or Receive free email alerts when new articles cite this article - sign up in http://symposium.cshlp.org/subscriptions go to: Cold Spring Harbor Symposia on Quantitative Biology To subscribe to Copyright © 1983 Cold Spring Harbor Laboratory Press Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.org Downloaded from
Transcript of Size of the Unwound Region of DNA in Escherichia … of the Unwound Region of DNA in Escherichia...

10.1101/SQB.1983.047.01.054Access the most recent version at doi: 1983 47: 455-461Cold Spring Harb Symp Quant Biol
H.B. Gamper and J.E. Hearst Ternary ComplexesRNA Polymerase and Calf Thymus RNA Polymerase II
Escherichia coliSize of the Unwound Region of DNA in
References http://symposium.cshlp.org/content/47/455.refs.html
This article cites 26 articles, 8 of which can be accessed free at:
serviceEmail alerting
click herethe box at the top right corner of the article orReceive free email alerts when new articles cite this article - sign up in
http://symposium.cshlp.org/subscriptions go to: Cold Spring Harbor Symposia on Quantitative BiologyTo subscribe to
Copyright © 1983 Cold Spring Harbor Laboratory Press
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

Size of the Unwound Region of DNA in Escherichia coli RNA Polymerase and
Calf Thymus RNA Polymerase II Ternary Complexes
H.B. GAMPER AND J .E. HEARST Department of Chemistry, University of California, Berkeley, California 94720
Based almost entirely on studies with the bacterial en- zyme, a general model has emerged for the ternary com- plex between DNA, RNA, and RNA polymerase. A central feature of this model is the existence of an enzyme-stabilized, unwound region in the DNA double helix that permits the coding strand to specify the com- plementary RNA sequence through hydrogen bonding. Helix unwinding first occurs at the promoter site in con- junction with formation of the open-promoter complex. Strand separation in this complex has been directly observed by analysis of the unwinding angle (Saucier and Wang 1972; Wang et al. 1977) and DNA-sequenc- ing technology (Siebenlist 1979). Recent studies in- dicate that 12-15 bp of DNA are unwound between the Pribnow box and the site of initiation (Melnikova et al. 1978; Siebenlist et al. 1980). It is believed that the size of the unwound region does not change during elonga- tion and that the RNA-DNA hybrid associated with the coding strand is approximately 12 bp long (Kumar and Krakow 1975).
We are interested in refining the structure of the RNA polymerase ternary complex so as to understand better the mechanisms of pausing and termination. Our ap- proach has been to determine the size of the DNA un- wound region in Escherichia coli RNA polymerase and calf thymus RNA polymerase II (pol II) ternary com- plexes formed on superhelical SV40 DNA. Polymerase- induced unwinding has been quantified by relaxing the ternary complexes and electrophoresing the deprotein- ized DNA. Analysis shows that the unwound region is 17 • 1 bp (580 • 30 ~ long in the E. coli complex and at least 10-11 bp (360 ~ long in the calf thymus com- plex. The results with the bacterial enzyme allow us to propose a topological model for transcription. This model has been described in detail elsewhere (Gamper and Hearst 1982).
EXPERIMENTAL PROCEDURES
E. coli holoenzyme was obtained from M. Chamber- lin (University of California at Berkeley) and A.-Y.M. Woody (Colorado State University). Calf thymus pol II was purchased from Bethesda Research Laboratories and diluted before use with an equal volume of 50 mM Tris-HCl (pH 7.9) buffer containing 0.1 mM (NH4)2SO4, 0.1 mM EDTA, 1 mM dithiothreitol (DTT), and 50% glycerol. Nicking-closing enzyme consisted of the fraction-I supernatant of lysed HeLa cell nuclei described by Vosberg et al. (1975). Superhelical SV40
DNA was purified from lytically infected TC-7 African green monkey kidney cells as outlined by Hallick et al. (1978).
The formation and analysis of E. coil RNA polymer- ase transcriptional complexes, including the electropho- retie determination of mean superhelicity, have been described previously (Gamper and Hearst 1982). Binary complexes of pol II were formed by incubating 0.5 gg of SV40 DNA with pol II for 5 minutes at 37~ in 9 gl of 100 mM Tris-HC1 (pH 7.9), 25 mM (NH4)2SO4, 3 mM MnC12, and 0.5 mM DTT. Initiation complexes were formed by making the reaction buffer 200 #M each in ATP, GTP, and CTP and incubating it an additional 5 minutes at 37~ Finally, ternary complexes were formed by diluting the reaction mixture sixfold with 50 #1 of 100 mM Tris-HC1 (pH 7.9), 10 mM MgCI2, 150 mM NaC1, and 1 rnM spermidine containing 200 gM each ATP, GTP, and CTP, 12 gM UTP, and diluted nicking- closing extract. RNA synthesis and enzymatic relaxa- tion of the DNA was allowed to proceed for 15 minutes at 37~ Binary and initiation complexes were relaxed in the same buffer containing the appropriate supple- ment of NTPs.
When pol II transcriptional complexes were analyzed directly by gel electrophoresis, the enzymatic relaxation (and RNA synthesis) was terminated by making the reaction mixtures 1% in SDS. The DNA was then precipitated with ethanol, dissolved in electrophoresis buffer containing 3.5% sucrose and 0.0035% brom- phenol blue, and loaded onto a horizontal 1% agarose gel immersed in 40 mM Tris-HCl and 5 mM MgC12. The samples were electrophoresed at 4~ for 35 hours across a 1.2 V/cm voltage gradient. The gel was stained with ethidium bromide and photographed on Polaroid type-665 positive/negative film.
For isolation of relaxed pol II ternary complexes on nitrocellulose, the reaction volume was scaled up threefold. Enzymatic relaxation and RNA synthesis were terminated by adding a 2.5-fold volume of 10 mM NaHzPO4 (pH 7.4), 1.25 U NaCl, 50 m~ EDTA, and 250 gg/ml of heparin. After 10 minutes at 4~ each sample was diluted to 1.0 ml with 10 mM NaH2PO4 (pH 7.4), 0.6 M NaCI, 0.1 mu EDTA, 0.1 mM DTT, and 10 gg/ml of heparin, filtered through nitrocellulose at a flow rate of 4 ml/min, and washed with 3 ml of the same solution. The DNA was recovered by shaking each filter in 0.8 ml of 10 mM Tris-HC1 (pH 7.2), 1 mM EDTA, and 0.2% SDS at room temperature for 40 minutes. After adjusting the solution to 0.2 M NaC1 and adding 40
455
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

456 GAMPER AND HEARST
#g of tRNA carrier, the DNA was precipitated with ethanol and analyzed by gel electrophoresis.
RESULTS
E. coli RNA Polymerase
At low ratios of enzyme to DNA, E. coli RNA poly- merase initiates transcription from either of two adja- cent thymidine residues at positions 2556 and 2555 on the early strand of SV40 (Reisbig and Hearst 198 I). The open-promoter complex readily forms at this preferred promoter site and is converted into a ternary complex by the addition of all four NTPs. Since termination and reinitiation does not occur under the conditions of our assay (data not shown), the number of active polymer- ases per SV40 DNA molecule can be determined by [7-32p]ATP end-labeling of the transcript.
Both binary and ternary transcriptiorial complexes have been analyzed for their ability to unwind DNA. The methodology involves relaxing superhelical DNA containing one or more transcriptional complexes to covalently closed, relaxed circles, denaturing the reac- tion mixtures with SDS, and electrophoresing the DNA through agarose under conditions that permit resolution of the topoisomeric bands. Figure 1 shows the gel pat- terns for SV40 DNA relaxed at 37~ (A) or 5~ (B) in the presence of a range of ternary complex concentra- tions.
Covalently closed DNA is unique in that its topologi- cal winding number (c0 is a fixed integer. This dictates that any change in helicity (/3) of the molecule be bal- anced by an equal and opposite change in superhelicity (z). Since ternary complexes locally unwind the DNA, denaturation of the enzymatically relaxed SV40 com- plexes permits the helix to reform and generates a slight superhelicity in the DNA. The polymerase-dependent shift in mean superhelicity of the DNA samples in Figure 1 is obvious. The direction of shift is opposite in
the two gels because the corresponding DNAs have superhelicities of different handedness.
Unwinding can be quantified by tracing the photo- graphic negative and determining a ~ for each DNA sample in the gel from peak-area measurements. The shift in ~ relative to that for a DNA control relaxed in the absence of polymerase is equivalent to the helical unwinding induced by a binary or ternary complex. An unwinding angle can be calculated for the complexes when these data are combined with end-labeling data on the number of active polymerases bound per SV40 genome. The results of such an analysis for binary and ternary complexes relaxed at 37~ (A) and at 50C (B) are shown in Figure 2. At the two temperatures, the ter- nary complex possesses an unwinding angle of 17.0 bp (580 ~ and 18.0 bp (6100), respectively. The binary complex exhibits a temperature-dependent unwinding characteristic of the conversion between open- and closed-promoter complexes (Wang et al. 1977). At 37~ the open-promoter form unwinds 17.0 bp of DNA (580~ whereas at 5~ no unwinding is observed with the equivalent of the closed-promoter form.
The unwinding angles determined with DNA end- labeling data have been verified by a semiindependent approach based on nitrocellulose filter trapping. This technique takes advantage of the stability of RNA polymerase ternary complexes at low temperature (Rhodes and Chamberlin 1974) and in the presence ot high salt (Richardson 1966). When ternary complexes are filtered through nitrocellulose in the presence of 0.6 M NaCI and 100/zg/ml of heparin, they are selectively retained with an efficiency of approximately 60% (Table 1). By filtering relaxed complexes formed at a low ratio of enzyme to DNA, the retained DNA has one ternary complex per genome, and the shift in ~ of that DNA relative to a control relaxed in the absence of polymerase should equal the unwinding angle of the complex. When DNA from such an experiment is recovered from the filter and electrophoresed, the
Figure 1. DNA unwinding induced by E. coil RNA polymerase ternary complexes at 37~ (A) and 5~ (B). Ternary complexes were formed on SV40 DNA at 18~ employing enzyme-to-DNA ratios of 0 (lane 1), 0.75 (lane 2), 1.50 (lane 3), 2.25 (lane 4), 3.00 (lane 5), and 3.75 (lane 6). The complexes were then enzymatically relaxed to equilibrium at 37~ (A) or 5~ (B), and the deproteinized DNA was electrophoresed in 1% agarose.
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

DNA UNWINDING DURING TRANSCRIPTION 457
Figure 2. Shift in mean superhelicity of SV40 DNA by E. coli RNA polymemse binary and ter- nary complexes at 37"C (A) and 5"C (B). Binary and ternary complexes were formed at 18"C and then enzymatically relaxed at either 37"C or 5"C. The DNA from these complexes was analyzed for polymerase-induced unwinding by gel elec- trophoresis. The number of active polymerases per SV40 genome was determined separately by measuring the uptake of [3,-~zP]ATP into acid- insoluble RNA. At 37"C the two complexes exhibit identical unwinding, whereas at 5"C only the ter- nary complex (O) induces unwinding.
2.0
1.5 I&~l
I.O
0.5
i i L O0 0.5 1.0 1.5
A c t i v e po lymer per qenome
2.5
3.0
2.5
2 .0
I&~'l 1.5
1.0
0.5
0 0
B
_" ' L i & i �9
0.5 I.O 1.5 2.0
A c t i v e p o l y m e r o s e s per genome
observed unwinding angle is 19 bp (645*) at 37"C and 17 bp (580*) at 5"C. Furthermore, the recovered DNA forms a tight Gaussian distribution in the gel, indicating that the size of the unwound region in the ternary com- plex is homogeneous. This conclusion is supported by the observation that the unwinding angle of the ternary complex is independent of its position on the SV40 genome (data not shown).
Calf Thymus RNA Polymerase H
In the presence of MgCI2, purified pol II from both plant and animal sources initiates nonspecific RNA syn- thesis with low efficiency (Mandel and Chambon 1974a,b; Dynan and Burgess 1981). The purified en- zyme lacks one or more factors required for promoter recognition and efficient formation of an open complex with DNA. In the absence of these as yet unidentified factors, initiation occurs preferentially at nicks (Lewis and Burgess 1980) and at single- to double-strand junc- tions (Lilley and Houghton 1979). Supercoiled DNA is also a good substrate for pol 11 (Lescure et al. 1978b). Torsional strain in the molecule is believed to facilitate open-complex formation by the enzyme. Nonspecific initiation off the form-I substrate can be further en-
Table 1. Selective Retention of E. coli and Calf Thymus RNA Polymerase Ternary Complexes on Nitrocellulose
Percent binding
Sample E. coli Calf thymus
Superhelical substrate DNA 1.1 0.7 binary complex 2.9 2.2 ternary complex 100 100
Enzymatically relaxed substrate DNA 3.3 14 binary complex 3.6 14 ternary complex 100 100
Free SV40 3H-labeled DNA or the same DNA containing approximately 0.7 E. coil polymerase or 2-3 calf thymus pol II transcriptional complexes per genome was filtered through nitrocellulose in the presence of 0.6 M NaCI and 10 teg/ml of heparin. Approximately 40% of the DNA in the E. coli ter- nary complex sample and 45% of the DNA in the corresponding pol II sam- ple was retained by the filter. Different nicking-closing extracts were used to relax the two DNA reaction series, and this accounts for the high background obtained with the relaxed calf-thymus-treated substrates.
hanced by carrying out the reaction in the presence of MnCl2 and (NH4)2SO 4 (Mandel and Chambon 1974a). These conditions further destabilize the helix and favor its unwinding by the enzyme.
RNA synthesis off nicked or superhelical substrates can lead to aberrant transcription by pol II. With nicked DNA, the 3 ' side of the nick can actually prime RNA synthesis, resulting in the generation of a covalent linkage between the noncoding DNA strand and the RNA transcript (Lewis and Burgess 1980). With superhelical DNA it is reported that pol II is unable to displace the transcript off the coding strand, leading to extensive R-loop formation (Lescure et al. 1978b; Lilley and Houghton 1979). We have utilized a three- step reaction scheme to obtain high levels of initiation off superhelical DNA without subsequent R-loop forma- tion. Purified pol II from calf thymus is incubated with SV40 DNA at 37~ in the presence of MnCI2 and (NH4)2SO4. After 5 minutes, ATP, GTP, and CTP are added to the binary reaction mixture to form initiation complexes. Elongation is then started by the simultane- ous addition of the fourth nucleotide, UTP, together with HeLa nicking-closing enzyme in a buffer contain- ing MgCl2 and NaCI. Elongation thus proceeds off a relaxed DNA substrate, and R-loop formation is avoid- ed. When RNA synthesis is monitored by the uptake of [ct-32p]UTP into acid-insoluble product, over 99% of the transcriptional activity is inhibited by 1.0 #g/ml of ct-amanitin. This level of inhibition is diagnostic for a type-II RNA polymerase.
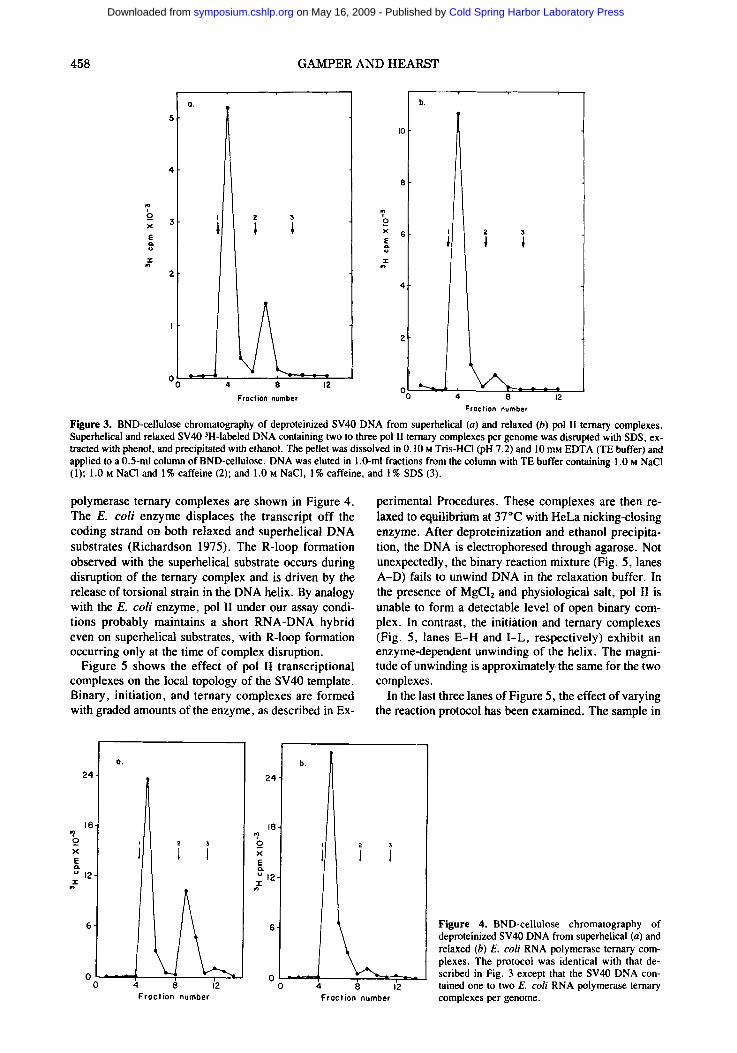
Benzoylated, naphthoylated DEAE (BND)-cellulose column chromatography has been used to verify an absence of R loops on the DNA transcribed by pol 11. This column matrix, which retains nucleic acids in 0.3 i NaCI, releases double-stranded DNA in 1.0 i NaCl and single-stranded DNA and RNA in 1.0 i NaCl and 1% caffeine. Figure 3 shows that 96% of the relaxed DNA and 78% of the superhelical DNA from SDS-disrupted ternary complexes elute with 1.0 i NaC1. This indicates that nearly all of the relaxed and most of the superhelical ternary complexes release RNA when pol II is de- natured.
For comparison, the elution profiles of SV40 DNA from SDS-disrupted, relaxed and superhelical, E. coli
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from
458 GAMPER AND HEARST
a.
o = z x 3
o
O0 4 8 I~'
Fraction number
b.
I0
8
4
2
. . . . . .__,
0
x 6 E o. o
2
2
1 t
,i 8 Frncti0n number
12
Figure 3. BND-cellulose chromatography of deproteinized SV40 DNA from superhelical (a) and relaxed (b) pol I1 ternary complexes. Superhelical and relaxed SV40 3H-labeled DNA containing two to three pol It ternary complexes per genome was disrupted with SDS, ex- tracted with phenol, and precipitated with ethanol. The pellet was dissolved in 0.10 M Tris-HC1 (pH 7.2) and 10 mM EDTA (TE buffer) and applied to a 0.5-ml column of BND-cellulose. DNA was eluted in 1.0-ml fractions from the column with TE buffer containing 1.0 m NaCI (1); 1.0 M NaCI and 1% caffeine (2); and 1.0 m NaCI, 1% caffeine, and 1% SDS (3).
polymerase ternary complexes are shown in Figure 4. The E. coli enzyme displaces the transcript off the coding strand on both relaxed and superhelical DNA substrates (Richardson 1975). The R-loop formation observed with the superhelical substrate occurs during disruption of the ternary complex and is driven by the release of torsional strain in the DNA helix. By analogy with the E. coli enzyme, pol II under our assay condi- tions probably maintains a short RNA-DNA hybrid even on superhelical substrates, with R-loop formation occurring only at the time of complex disruption.
Figure 5 shows the effect of pol II transcriptional complexes on the local topology of the SV40 template. Binary, initiation, and ternary complexes are formed with graded amounts of the enzyme, as described in Ex-
perimental Procedures. These complexes are then re- laxed to equilibrium at 37~ with HeLa nicking-closing enzyme. After deproteinization and ethanol precipita- tion, the DNA is electrophoresed through agarose. Not unexpectedly, the binary reaction mixture (Fig. 5, lanes A-D) fails to unwind DNA in the relaxation buffer. In the presence of MgCI2 and physiological salt, pol II is unable to form a detectable level of open binary com- plex. In contrast, the initiation and ternary complexes (Fig. 5, lanes E-H and~I-L, respectively) exhibit an enzyme-dependent unwinding of the helix. The magni- tude of unwinding is approximately the same for the two complexes.
In the last three lanes of Figure 5, the effect of varying the reaction protocol has been examined. The sample in
2 4 -
1 8 - ?
x E
" 12 3"
m
6 �84
0 0
0~
4
i i
8 12 F r a c t i o n number
o
x
u
_~4-
184
0 0 4
7 i
Frac t i on number
Figure 4. BND-cellulose chromatography of deproteinized SV40 DNA from superhelical (a) and relaxed (b) E. coli RNA polymerase ternary com- plexes. The protocol was identical with that de- scribed in Fig. 3 except that the SV40 DNA con- tained one to two E. coli RNA polymerase ternary complexes per genome.
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

DNA UNWINDING DURING TRANSCRIPTION 459
Figure 5. Effect of pol II binary, initiation, and ternary complexes on the local topology of DNA. Pol II binary (lanes A-D), initiation (lanes E-H), and ternary (lanes l-L) complexes were formed as described in Experimental Procedures. The complexes were relaxed to equilibrium at 37"C, and the deproteinized DNA was electrophoresed in 1% agarose. For each of the three complexes, from left to right, 0.5 #g of SV40 DNA was treated with 0, 0.6, 1.2, and 1.8 #1 ofpol II extract. The last four samples in the gel show the effect of variations in experimental protocol as described in the text. Except for the sample in lane M, which lacked pol II, the DNA in lanes N-P was treated with 2.0 #1 of pol II extract.
lane N comes from a ternary complex reaction sequence carried out entirely in relaxation buffer. Since pol II does not initiate efficiently in this buffer, there is no detectable unwinding. The samples in lanes O and P come from ternary complexes allowed to elongate prior to the addition of or in the complete absence of nicking- closing enzyme. There is no indication from the banding pattern in the gel that the DNA contains an R loop.
The ternary complex unwinding angle has been deter- mined by using the nitrocellulose filter trapping tech- nique. Like the bacterial enzyme, pol II forms a very stable ternary complex resistant to polyanions and high salt (Leseure et al. 1978a,b; Dynan and Burgess 1981). When filtered through nitrocellulose in the presence of heparin and 0.6 M NaCI, DNA that contains a pol II ter- nary complex is selectively retained on the filter (Table 1). Since the number of active enzymes per microliter of extract is not known for the pol II preparation, several ternary complex reaction mixtures are formed with graded amounts of the enzyme. After relaxation at 37~ each is passed through nitrocellulose, and the of the retained DNA is determined by gel elec- trophoresis. In Figure 6 the AF of the DNA samples relative to a DNA control relaxed in the absence of pol II is plotted versus the amount of pol II extract in the reaction mixture. As the amount of pol II decreases, AT decreases to a plateau value. In the plateau region, there is less than one ternary complex per SV40 genome in the reaction mixture and Au is assumed to represent the ter- nary complex unwinding angle. On the basis of a helical repeat of 10.6 bp per turn and assuming that 10% of the filter-retained DNA does not possess a ternary complex (see Table 1), an unwinding angle of 10-11 bp (360 ~ is estimated for the pol II ternary complex at 37~
DISCUSSION
The DNA unwound region in the E. coli RNA poly- merase open-promoter and ternary complexes is 17 -4- 1
bp at 37~ This compares favorably with the estimate of 15 bp made by Melnikova et al. (1978) but is one- third larger than the value of 12 bp, arrived at by
1.6
1.2
A7
0 .8
I
0 .4
I I I I
I 2 ,3 4 ,5
FI RNA Polymerose TI"
Figure 6. Shift induced by pol II in the mean superhelicity of SV40 DNA recovered from nitrocellulose filters. Pol II ternary complexes on SV40 DNA were formed with the indicated volumes of extract and filtered through nitrocellulose as described in Ex- perimental Procedures. The shift in mean superhelicity of the recovered DNA was measured relative to a DNA sample relaxed in the absence of polymerase.
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

460 GAMPER AND HEARST
Figure 7. A topological model for the E. coli RNA polymerase ternary complex. The two oval regions represent the binding domains in the enzyme for the RNA-DNA hybrid and the noncoding DNA strand. (l~) Windase centers; (<]) catalytic site.
Siebenlist (1979) for the open-promoter complex. In agreement with earlier observations (Saucier and Wang 1972; Melnikova et al. 1978), both complexes have un- wound regions of identical size. Furthermore, the size of the unwound region in the ternary complex is in- dependent of its position on the SV40 genome. To- gether, these data indicate that the DNA unwound region is rigidly controlled by the polymerase.
A topological model of the ternary complex consistent with the unwinding angle data is shown in Figure 7. Two windase centers bracket the catalytic site and in- duce a rotation in the DNA helix on either side of the en- zyme, as well as in the RNA-DNA hybrid. The rotation is concerted and results in a separation of the DNA helix at the leading unwindase site and its formation again at
IIII llll
IIII j j l l l l l
l l l l ~ l l l l
III 11
Figure 8. Alternative secondary structures for the expression of a hyphenated dyad symmetry element during pausing and termina- tion. The palindromic regions are represented by a bar on the RNA.
the lagging rewindase site after displacement of the RNA transcript. In the unwound region, the noncoding DNA strand and the RNA-DNA hybrid interact with two separate binding domains on the polymerase, which converge at the windase centers. A central feature of this model is the rotation of the DNA double helix and not the RNA polymerase. As a result, the model pre- dicts that simultaneous convergent or divergent transcription requires a swivel in the intervening DNA such as could be provided by a topoisomerase. It further predicts that hindrance of DNA rotation, such as through the covalent addition of a large solid support to the helix, will inhibit transcription. These predictions are being tested.
During in vitro transcription, E. coli RNA poly- merase moves discontinuously along the helix, pausing for long periods of time at certain distinct positions. One causative factor appears to be the presence in the DNA of a hyphenated dyad symmetry element with a center of symmetry 16 to 20 nucleotides and a start 5 to 10 nucle- otides upstream from the pause or termination site (Rosenberg and Court 1979; Farnham and Platt 1981). The three alternative secondary structures that could arise from such a symmetry element are shown in Figure 8. Only the RNA hairpin (Fig. 8b) (Adhya and Gottesman 1978; Farnham and Platt 1980) and the bridged RNA-DNA intermediate (Fig. 8c) (Gamper and Hearst 1982) are consistent with an unwound region of 17 bp. Formation of a DNA hairpin on the noncoding strand (Fig. 8d) (Hobom et al. 1979) would lead to a much greater unwinding of the DNA helix and so is con- sidered unlikely.
Due to the inability of purified pol II to form stable open-promoter complexes under the relaxation condi- tions, we have been unable to determine an unwinding angle for this complex. In contrast, the pol II ternary complex is a remarkably stable species, similar in many respects to the corresponding E. coli complex. It clearly is able to displace the nascent RNA transcript off the DNA coding strand and maintains an unwound region equivalent to one turn of helix. The length of the un- wound region is surprisingly small considering the size and complexity of the pol II enzyme. Nonetheless, it is reasonable to assume that the topological model outlined in Figure 7 also applies to pol II. One observation that suggests this is the case involves an examination of the termination site for transcription of the eukaryotic
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from

DNA UNWINDING DURING TRANSCRIPTION 461
17
b. AAu 12 u G C A UU
C-G U C G-C U - A C -G C-G C-G C ~ U-A ~ G C-G C-G G -C G -C
5 'A -UUUUUUU 3' 5' G -CACCA 3'
Figure 9. Potential RNA hairpin structure at the tryptophan at- tenuator site (a) and the histone gene termination site (b). The number of nucleotides between the dyad center and the stop site is given above the arrows.
histone gene. This site is highly conserved and contains a hyphenated dyad symmetry element upstream from the stop point (Birchmeier et al. 1982). The sequence of this palindromic region and the corresponding region of the tryptophan attenuator are shown in Figure 9. In each case the distance between the dyad center and the stop site for RNA synthesis is approximately equal to the measured length of the loop opened in each of the re- spective ternary complexes. This is what would be pre- dicted if pausing at the dyad symmetry element is re- lated to the formation of a bridged RNA-DNA inter- mediate.
ACKNOWLEDGMENTS
We thank E. Daniell for providing us with HeLa cells and J. Bartholomew and M. Botchan for generously donating SV40-infected TC-7 cultures. We also thank C.-C. Huang and J. Thompson for their interest and ad- vice. This work was supported, in part, by grants from the American Cancer Society (NP185) and the National Institutes of Health (GM-11180 and T32-ES07075) and by a California Division-American Cancer Society Junior Fellowship (J-21-82) to H.B.G.
REFERENCES
ADHYA, S. and M. GOTTESMAN. 1978. Control of transcrip- tion termination. Annu. Rev. Biochem. 47: 967.
BIRCHMEIER, C., R. GROSSCHEDL, and M.L. BIRNSTIEL. 1982. Generation of authentic 3' termini of an H2A mRNA in vivo is dependent on a short inverted DNA repeat and on spacer sequences. Cell 28: 739.
DYNAN, W.S. and R.R. BURGESS. 1981. In vitro transcription by wheat germ RNA polymerase II. Initiation of RNA syn- thesis on relaxed, closed circular template. J. BioL Chem. 256: 5866.
FARNHAM, P.J. and T. PLATT. 1980. A model for transcription termination suggested by studies on the trp attenuator in vitro using base analogs. Cell 20: 739.
- - . 1981. Rho-independent termination: Dyad symmetry in DNA causes RNA polymerase to pause during transcrip- tion in vitro. Nucleic Acids Res. 9: 563.
GAMPER, H.B. and J.E. HEARST. 1982. A topological model for transcription based on unwinding angle analysis of E. coil RNA polymerase binary, initiation, and ternary com- plexes. Cell 29: 81.
HALLICK, L.M., H.A. YOKOTA, J.C. BARTHOLOMEW, and J.E. HEARST. 1978. Photochemical addition of the cross- linking reagent 4,5',8-trimethylpsoralen (trioxsalen) to in- tracellular and viral simian virus 40 DNA-histone com- plexes../. Virol. 27: 127.
HOROM, G., K. GROSSCHEDL, M. LusKY, G. SCHERER, E. SCHWARZ, and H. K6SSEL. 1979. Functional analysis of the replicator structure of lamboid bacteriophage DNAs. Cold Spring Harbor Syrup. Quant. Biol. 43: 165.
KUMAR, S.A. and J.S. KRAKOW. 1975. Studies on the product binding site of the Azotobacter vinelandii ribonucleic acid polymerase. Jr. Biol. Chem. 250: 2878.
LEscu~, B., C. DAUGUET, and M. YANW. 1978a. Transcrip- tion of polyoma virus DNA in vitro. Localization of calf thymus RNA polymerase II binding sites. Jr. Mol. Biol. 124: 87.
LESCURE, B., A. CHESTIER, and M. YANW. 1978b. Transcrip- tion of polyoma virus DNA in vitro. Transcription of superhelical and linear polyoma DNA by RNA polymerase II. Jr. Biol. Chem. 124: 73.
LEWIS, M.K. and R.R. BURGESS. 1980. Transcription of si- mian virus 40 DNA by wheat germ RNA polymerase II. Priming of RNA synthesis by the 3'-hydroxyl of DNA at single strand nicks, jr. Biol. Chem. 255: 4928.
LILLEY, D.M.L and M. HOUGHTON. 1979. The interaction of RNA polymerase II from wheat with supercoiled and linear plasmid templates. Nucleic Acids Res. 6: 507.
MANDEL, J.L. and P. CHAMBON. 1974a. Animal DNA- dependent RNA polymerases. Studies on the reaction parameters of transcription in vitro of simian virus 40 DNA by mammalian RNA polymerases AI and B. Eur. jr. Biochem. 41: 367.
- - . 1974b. Animal DNA-dependent RNA polymerases. Analysis of the RNAs synthesized on simian virus 40 superhelical DNA by mammalian RNA polymerases AI and B. Eur. J. Biochem. 41: 379.
MELNIKOVA, A.F., R. BEABEALASHVILLI, and A.D. Mm- ZABEKOV. 1978. A study of unwinding of DNA and shielding of the DNA grooves by RNA polymerase by us- ing methylation with dimethylsulphate. Eur. J. Biochem. 84: 301.
REISBIG, R.R. and J.E. HEARST. 1981. Escherichia coli deoxy- ribonucleic acid dependent ribonucleic acid polymerase transcriptional pause sites on SV40 DNA F 1. Biochemistry 20: 1907.
RHODES, G. and M.J. CHAMBERUN. 1974. Ribonucleic acid chain elongation by Escherichia coli ribonucleic acid polymerase. Isolation of ternary complexes and the kinetics of elongation. J. Biol. Chem. 249: 6675.
RICHARDSON, J.P. 1966. Enzymic synthesis of RNA from T7 DNA. J. Mol. Biol. 21: 115.
- - . 1975. Attachment of nascent RNA molecules to superhelical DNA. J. Mol. Biol. 98: 565.
ROSENBERG, M. and D. COURT. 1979. Regulatory sequences involved in the promotion and termination of RNA transcription. Annu. Rev. Genet. 13: 319.
SAUCIER, J.-M. and J.C. WANG. 1972. Angular alteration of the DNA helix by E. coil RNA polymerase. Nat. New Biol. 239: 167.
S]EBENLIST, U. 1979. RNA polymerase unwinds an I l-base pair segment of a phage T7 promoter. Nature 279:651.
SmBENLlST, U., R.B. SIMPSON, and W. GILBERT. 1980. E. coli RNA polymerase interacts homologously with two dif- ferent promoters. Cell 20: 269.
VOSBERG, H.-P., LI. GROSSMAN, and J. VINOORAD. 1975. Isolation and partial characterization of the relaxation pro- tein from nuclei of cultured mouse and human ceils. Eur. J. Biochem. 55: 79.
WANG, J.C., J.H. JACOBSEN, and J.-M. SAUCIER. 1977. Physicochemical studies on interactions between DNA and RNA polymerase. Unwinding of the DNA helix by Escherichia coli RNA polymerase. Nucleic Acids Res. 4: 1225.
Cold Spring Harbor Laboratory Press on May 16, 2009 - Published by symposium.cshlp.orgDownloaded from


















