
Severe Preeclampsia-Related Changes in Gene Expression at...
11
Severe Preeclampsia-Related Changes in Gene Expression at the Maternal-Fetal Interface Include Sialic Acid-Binding Immunoglobulin-Like Lectin-6 and Pappalysin-2 Virginia D. Winn, Matthew Gormley, Agnes C. Paquet, Kasper Kjaer-Sorensen, Anita Kramer, Kristen K. Rumer, Ronit Haimov-Kochman, Ru-Fang Yeh, Michael T. Overgaard, Ajit Varki, Claus Oxvig, and Susan J. Fisher Departments of Obstetrics, Gynecology, and Reproductive Sciences (V.D.W., M.G., R.H.-K.), Cell and Tissue Biology (R.H.-K., S.J.F.), Medicine Lung Biology Center (A.C.P., R.-F.Y.), and Anatomy and Pharmaceutical Chemistry (S.J.F.), University of California, San Francisco, San Francisco, California 94143; Department of Obstetrics and Gynecology (V.D.W., A.K., K.K.R.), University of Colorado Denver, Aurora, Colorado 80045; Department of Molecular and Cellular Medicine (A.V.), University of California, San Diego, San Diego, California 92093; and Department of Molecular Biology (K.K.-S., M.T.O., C.O.), University of Aarhus, DK-8000 Aarhus C, Denmark Preeclampsia (PE), which affects 4 – 8% of human pregnancies, causes significant maternal and neonatal morbidity and mortality. Within the basal plate, placental cytotrophoblasts (CTBs) of fetal origin invade the uterus and extensively remodel the maternal vasculature. In PE, CTB invasion is often shallow, and vascular remodeling is rudimentary. To better understand possible causes, we conducted a global analysis of gene expression at the maternal-fetal interface in placental samples from women with PE (n 12; 24 –36 wk) vs. samples from women who delivered due to preterm labor with no evidence of infection (n 11; 24 –36 wk), a condition that our previous work showed is associated with normal CTB invasion. Using the HG-U133A&B Affymetrix GeneChip platform, and statistical significance set at log odds-ratio of B 0, 55 genes were differentially expressed in PE. They encoded proteins previously associated with PE [e.g. Flt-1 (vascular endothelial growth factor receptor-1), leptin, CRH, and inhibin] and novel molecules [e.g. sialic acid binding Ig-like lectin 6 (Siglec-6), a potential leptin receptor, and pappalysin-2 (PAPP-A2), a protease that cleaves IGF- binding proteins]. We used quantitative PCR to validate the expression patterns of a subset of the genes. At the protein level, we confirmed PE-related changes in the expression of Siglec-6 and PAPP-A2, which localized to invasive CTBs and syncytiotrophoblasts. Notably, Siglec-6 placental expression is uniquely human, as is spontaneous PE. The functional significance of these novel observations may provide new insights into the pathogenesis of PE, and assaying the circulating levels of these proteins could have clinical utility for predicting and/or diagnosing PE. (Endocrinology 150: 452– 462, 2009) P reeclampsia (PE), a pregnancy complication, is manifested by the onset of hypertension and proteinuria in the second half of pregnancy. PE is relatively common (4 – 8% of pregnan- cies), with potentially deadly consequences for the mother and/or her offspring. Currently, the only definitive treatment for this condition is delivery of the placenta, and therefore the infant, accounting for 15% of all preterm births in the United States. Despite decades of research, a full understanding of the patho- genesis of PE remains elusive, hampering the development of better predictive tests, methods of diagnosis, and therapeutic interventions. Nevertheless, it is clear that the placenta plays a central role; the signs of PE can occur in molar pregnancies, which lack a fetus, and the disease resolves once the placenta is delivered. Interestingly, spontaneous PE does not occur in other animals, including nonhuman primates. Formation of the human placenta entails differentiation of its ISSN Print 0013-7227 ISSN Online 1945-7170 Printed in U.S.A. Copyright © 2009 by The Endocrine Society doi: 10.1210/en.2008-0990 Received July 2, 2008. Accepted September 15, 2008. First Published Online September 25, 2008 Abbreviations: CTB, Cytotrophoblast; IGFBP, IGF-binding protein; PAPP-A2, pappalysin-2; PE, preeclampsia; PTL, preterm labor; PVDF, polyvinylidene difluoride; Q-PCR, quantitative PCR; Siglec-6, sialic acid binding Ig-like lectin 6; SOCS, suppressor of cytokine signaling; STB, syncytiotrophoblast; VEGFR, vascular endothelial growth factor receptor. REPRODUCTION-DEVELOPMENT 452 endo.endojournals.org Endocrinology, January 2009, 150(1):452– 462 at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.org Downloaded from
Transcript of Severe Preeclampsia-Related Changes in Gene Expression at...

Severe Preeclampsia-Related Changes in GeneExpression at the Maternal-Fetal Interface IncludeSialic Acid-Binding Immunoglobulin-Like Lectin-6and Pappalysin-2
Virginia D. Winn, Matthew Gormley, Agnes C. Paquet, Kasper Kjaer-Sorensen,Anita Kramer, Kristen K. Rumer, Ronit Haimov-Kochman, Ru-Fang Yeh,Michael T. Overgaard, Ajit Varki, Claus Oxvig, and Susan J. Fisher
Departments of Obstetrics, Gynecology, and Reproductive Sciences (V.D.W., M.G., R.H.-K.), Cell and Tissue Biology(R.H.-K., S.J.F.), Medicine Lung Biology Center (A.C.P., R.-F.Y.), and Anatomy and Pharmaceutical Chemistry (S.J.F.),University of California, San Francisco, San Francisco, California 94143; Department of Obstetrics and Gynecology(V.D.W., A.K., K.K.R.), University of Colorado Denver, Aurora, Colorado 80045; Department of Molecular and CellularMedicine (A.V.), University of California, San Diego, San Diego, California 92093; and Department of MolecularBiology (K.K.-S., M.T.O., C.O.), University of Aarhus, DK-8000 Aarhus C, Denmark
Preeclampsia (PE), which affects 4–8% of human pregnancies, causes significant maternal andneonatal morbidity and mortality. Within the basal plate, placental cytotrophoblasts (CTBs) of fetalorigin invade the uterus and extensively remodel the maternal vasculature. In PE, CTB invasion isoften shallow, and vascular remodeling is rudimentary. To better understand possible causes, weconducted a global analysis of gene expression at the maternal-fetal interface in placental samplesfrom women with PE (n � 12; 24–36 wk) vs. samples from women who delivered due to pretermlabor with no evidence of infection (n � 11; 24–36 wk), a condition that our previous work showedis associated with normal CTB invasion. Using the HG-U133A&B Affymetrix GeneChip platform, andstatistical significance set at log odds-ratio of B �0, 55 genes were differentially expressed in PE.They encoded proteins previously associated with PE [e.g. Flt-1 (vascular endothelial growth factorreceptor-1), leptin, CRH, and inhibin] and novel molecules [e.g. sialic acid binding Ig-like lectin 6(Siglec-6), a potential leptin receptor, and pappalysin-2 (PAPP-A2), a protease that cleaves IGF-binding proteins]. We used quantitative PCR to validate the expression patterns of a subset of thegenes. At the protein level, we confirmed PE-related changes in the expression of Siglec-6 andPAPP-A2, which localized to invasive CTBs and syncytiotrophoblasts. Notably, Siglec-6 placentalexpression is uniquely human, as is spontaneous PE. The functional significance of these novelobservations may provide new insights into the pathogenesis of PE, and assaying the circulatinglevels of these proteins could have clinical utility for predicting and/or diagnosing PE.(Endocrinology 150: 452–462, 2009)
Preeclampsia (PE), a pregnancy complication, is manifestedby the onset of hypertension and proteinuria in the second
half of pregnancy. PE is relatively common (4–8% of pregnan-cies),withpotentiallydeadly consequences for themotherand/orher offspring. Currently, the only definitive treatment for thiscondition is delivery of the placenta, and therefore the infant,accounting for 15% of all preterm births in the United States.Despite decades of research, a full understanding of the patho-
genesis of PE remains elusive, hampering the development ofbetter predictive tests, methods of diagnosis, and therapeuticinterventions. Nevertheless, it is clear that the placenta plays acentral role; the signs of PE can occur in molar pregnancies,which lack a fetus, and the disease resolves once the placenta isdelivered. Interestingly, spontaneous PE does not occur in otheranimals, including nonhuman primates.
Formation of the human placenta entails differentiation of its
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2009 by The Endocrine Societydoi: 10.1210/en.2008-0990 Received July 2, 2008. Accepted September 15, 2008.First Published Online September 25, 2008
Abbreviations: CTB, Cytotrophoblast; IGFBP, IGF-binding protein; PAPP-A2, pappalysin-2;PE, preeclampsia; PTL, preterm labor; PVDF, polyvinylidene difluoride; Q-PCR, quantitativePCR; Siglec-6, sialic acid binding Ig-like lectin 6; SOCS, suppressor of cytokine signaling;STB, syncytiotrophoblast; VEGFR, vascular endothelial growth factor receptor.
R E P R O D U C T I O N - D E V E L O P M E N T
452 endo.endojournals.org Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

specialized progenitor cells, termed cytotrophoblasts (CTBs), aprocess that establishes the architecture of the placenta and thematernal-fetal interface or basal plate (Fig. 1). During CTB dif-ferentiation, progenitors assume one of two fates. In floatingvilli, they fuse to form multinucleate syncytiotrophoblasts(STBs), whose primary functions are transport and hormoneproduction. In anchoring villi, mononuclear CTBs acquire tu-mor-like properties that enable them to invade the decidua, theendometrium of pregnancy, and the adjacent third of the myo-metrium (interstitial invasion). They also breach the small uter-ine vessels they encounter, completely replacing the resident ma-ternal endothelial lining and intercalating within the muscularwalls, a process that involves nearly the entire intrauterine courseof arterioles, but only the termini of veins (endovascular inva-sion). As a result, high-resistance spiral arterioles are trans-formed into low-resistance, high-capacitance vessels that divertuterine blood flow to the floating villi. This process is most activeduring 10–20 wk of gestation, before the onset of the clinicalsymptoms of PE.
During the last several years, a clearer picture of the patho-genesis of PE has begun to emerge. A two-stage model has beenproposed in which the initiating event, poor placentation, isthought to occur early in gestation (1). This concept is supportedby several studies that document the association between re-duced blood flow to the placenta before 20 wk gestation, asdetermined by color Doppler ultrasound evaluation of terinearterial blood flow, and a greatly increased risk of developing PE(2, 3). Anatomic examination shows that the specific area of theplacenta most affected by this syndrome is the basal plate, the siteof CTB invasion. Interstitial CTB invasion is often shallow, andendovascular invasion does not proceed beyond the terminalportions of the spiral arterioles. Thus, the maternal vessels do notundergo the complete spectrum of physiological changes thatnormally occur (e.g. loss of endothelial lining and musculoelastic
tissue); the mean external diameter of the myometrial vessels isless than half that of equivalent vessels from uncomplicatedpregnancies (4 – 6). In addition, fewer vessels show evidence ofCTB invasion (7). Thus, their architecture precludes an ade-quate response to gestation-related fetal demands for in-creased blood flow.
The second stage of PE is thought to be the maternal responseto abnormal placentation. Systemic endothelial dysfunction ap-pears to be an important common denominator (1, 8, 9). Recentdata point to an imbalance in circulating factors with angiogenic/vasculogenic functions, such as soluble vascular endothelialgrowth factor receptor-1 (VEGFR-1, sFlt-1), placental growthfactor, and the TGF-� receptor endoglin (10–16).
The studies reported here employed an unbiased approachthat required no previous knowledge of the molecules involvedto achieve a better understanding of PE pathogenesis. Specifi-cally, we used a microarray platform to evaluate gene expressionpatterns at a global level in the basal plate region, the area of CTBinvasion, of affected individuals compared with control subjectswho were matched for gestational age at the time of delivery. Theimportance of controlling for gestational age is illustrated by ourrecent work on gene expression changes at the maternal-fetalinterface that revealed dramatic differences between migestationand term (17). In this regard, we exploited our recent observationthat preterm labor (PTL) without signs of inflammation is asso-ciated with normal CTB differentiation/invasion. We did this byusing basal plate samples collected from patients who delivereddue to this pregnancy complication as gestation-matched con-trols (18). The results of the current study revealed 55 differen-tially expressed genes, of which the majority were not known tobe dysregulated in severe PE. The results of this work highlightthe complex pathophysiology of this condition and the manypathways it impacts.
FIG. 1. Diagram of the human maternal-fetal interface. A, Representation of the human placenta after delivery. The placental surface that was adjacent to the uterinewall is termed the basal plate. The boxed area denotes the region biopsied for these studies. B, View of the basal plate at the cellular level. This chimeric region of theplacenta is composed of both maternal and fetal components: extravillous (invasive) cytotrophoblasts (dark gray), decidual cells (light gray), remodeled vasculature (bothinvasive CTBs and maternal endothelium), and maternal immune cells (white). [Reproduced with permission from V. D. Winn et al.: Endocrinology 148:1059, 2007 (17).©The Endocrine Society.]
Endocrinology, January 2009, 150(1):452–462 endo.endojournals.org 453
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

Materials and Methods
Tissue collectionThe University of California, San Francisco (UCSF), Committee on
Human Research approved this study. Informed consent was obtainedfrom each parturient tissue donor before delivery. Basal plate biopsyspecimens of the maternal-fetal interface were collected from individualswho developed severe PE or PTL without signs of infection. Pregnanciescomplicated by multiple gestations, fetal anomalies, premature ruptureof the membranes, infection, diabetes, or other autoimmune diseaseswere excluded. Gestational age was determined by using standard datingcriteria (19). The basal plate was dissected and processed from the pla-centa proper as previously described (17). All samples were processedwithin 2 h of delivery.
Total RNA extractionRNA was isolated from snap-frozen basal plate specimens using a
modified Trizol method as previously described (20). Aliquots were eval-uated by using the Agilent RNA 6000 Nano LabChip kit (Agilent Tech-nologies, Santa Clara, CA) on an Agilent Bioanalyzer 2100 system. Cap-illary electrophoresis data in commaseparated value files were analyzedby using the Degradometer version 1.41 software (available at http://www.dnaarrays.org) (21). Only RNA with a degradation factor of lessthan 11 was used in subsequent microarray experiments.
Microarray hybridizationThe microarray platform was the high-density HG-U133A and HG-
U133B GeneChips (Affymetrix, Santa Clara, CA). Sample processingand hybridization was accomplished by using the protocols devised bythe UCSF Gladstone (National Heart, Lung, and Blood Institute)Genomics Core Facility (http://www.gladstone.ucsf.edu/gladstone/php/section) as previously described (17). Samples from individual basalplates were analyzed separately. Specifically, the HG-U133A and HG-U133B Affymetrix GeneChips were each hybridized with 15 �g cRNAand then washed, stained, and imaged at the Gladstone Genomics CoreFacility by using standard Affymetrix protocols. Data files were depos-ited in the GEO (Gene Expression Omnibus) data repository (accessionno. pending).
Data analysisThe raw image data were analyzed by using GeneChip Expression
Analysis software (Affymetrix) to produce perfect match and mismatchvalues. Subsequently, quality control, preprocessing, and linear model-ing were performed using Bioconductor (22), an open source and opendevelopment software project based on the R statistical package (http://www.r-project.org). Initial hybridization quality was assessed using theBioconductor package affyPLM, and slight variations in quality werecompensated for during the preprocessing stage, which was performedin two steps. First, we used a Probe Level robust linear model (23) toobtain separate normalized log intensities for each chip (i.e. backgroundsubtraction, quantile normalization, and probe set summarization). Sec-ond, we applied a global median normalization at the probe set level toall A and B GeneChips (n � 46) and then combined these data into amatrix of log2-based gene expression measures, in which columns cor-responded to different cRNA samples, and rows corresponded to thedifferent probe sets.
Estimated log ratios (M value) between cases and controls were de-termined by using the limma software package in R (24). Then differ-entially expressed genes were selected by statistical analysis of LODratios, and B �0 was set as significant. The results showed that theexpression of 55 genes (71 probe sets) was significantly modulated. Thenthe normalized intensity values for this data set were centered to themedian intensity value for each probe set, after which the probe sets wereranked according to their M values (representing fold change) and de-picted as a gene expression color map. Evaluation of these filtered probes
in the context of both clinical and experimental variables was performedto assess confounding factors.
Quantitative PCRReverse transcription of RNA (total) samples isolated from basal
plate biopsies was carried out as previously described (17). All templateswere amplified with Assay-on-Demand kits (Applied Biosystems, FosterCity, CA) or primer/probe sets designed by the UCSF Biomolecular Re-search Center (see supplemental Table SI, published as supplementaldata on The Endocrine Society’s Journals Online web site at http://endo.endojournals.org). Quantitative PCR (Q-PCR) was carried out as pre-viously described (17). Negative control RNA samples were not reversetranscribed or lacked template inputs. Relative quantification was de-termined by using the standard curve method (see Applied BiosystemsUser Bulletin no. 2; http://www3.appliedbiosystems.com). In prelimi-nary experiments, we investigated the utility of 11 potential targets asendogenous controls (endogenous control plate; Applied Biosystems).The results showed that the 18S rRNA did not vary with gestational ageor disease state. Accordingly, the levels of this transcript were used toobtain normalized values for the target amplicons. A 24-wk PTL sample,the earliest gestational age included in our analysis, was used as thecalibrator for the relative quantification using the standard curvemethod. Results are reported as the relative fold mRNA levels � SD foreach basal plate specimen. The mean values of the PE and PTL sampleswere compared using a two-tailed Student’s t test (P � 0.05).
Sialic acid binding Ig-like lectin 6 (Siglec-6)immunohistochemistry
Serial sections (5 �m) from formalin-fixed tissues were deparaf-finized and rehydrated. Staining was performed as previously published(25). The tissue sections were incubated overnight at 4 C with one of theprimary antibodies [anti-Siglec-6 (26), 1:25, or anti-cytokeratin-7 (anti-CK-7) clone OV-TL 12/30 (Dako, Carpinteria, CA), diluted 1:50 inDakoCytomatin antibody diluent (Dako)]. Negative control tissue sec-tions were incubated without primary antibody. Visualization wasachieved by incubation with diaminobenzidine for 2 min following themanufacturer’s instructions (Vector Laboratories, Burlingame, CA), andnuclei were counterstained with hematoxylin QS (Vector Laboratories).The tissue sections were imaged by using a Nikon eclipse 80i microscopeand photographed with a Q-imaging Retiga 2000R digital camera.
For immunofluorescence staining, frozen sections (5 �m) from OCT-embedded tissues were washed in PBS, and nonspecific reactivity wasblocked by incubating with 3% BSA (wt/vol), 0.1% Triton X-100 (vol/vol), and 0.5% Tween 20 (vol/vol) in PBS for 30 min. Then sections wereincubated with anti-Siglec-6 (1:100 in blocking buffer) for 1 h andwashed in PBS. Negative controls were incubated without the primaryantibody. Sections were then exposed to anti-CK-7 [clone 7D3 (27),1:100] for 1 h and washed in PBS as described above. The sections werethen incubated with Alexa Fluor 594-conjugated goat antimouse IgG(1:1000; Molecular Probes Inc., Eugene, OR) and fluorescein isothio-cyanate-labeled donkey antirat IgG (1:200; Jackson ImmunoResearchLaboratories, West Grove, PA) antibodies for 30 min and washed in PBS.Tissue sections were mounted in Vectashield containing 4�,6-diamidino-2-phenylindole (Vector Laboratories), which allowed visualization ofthe nuclei. Imaging was done using a Leica DM 5000B fluorescencemicroscope equipped with a Leica DFC 350FX digital camera (LeicaMicrosystems, Bannockburn, IL; and Leica-Camera, Solms, Germany).
Generation of antihuman pappalysin-2 (PAPP-A2)polyclonal serum
A fragment encoding Ser-234 to Gln-495 of PAPP-A2 was ampli-fied by PCR using the plasmid pPA2 as the template (28). The forwardprimer was 5�-CGATAGATCTATCGAGGGTAGGAGTCCACCG-GAGGAAAGCAAC-3� (a BglII site is underlined, and a factor Xa rec-ognition site is shown in bold). The reverse primer was 5�-CGATA-AGCTTTCACTGCAAAGGCGACAGAATCTC-3� (a HindIII site is
454 Winn et al. PE Effects on Basal Plate Gene Expression Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

underlined, and a stop codon is shown in bold). The 821-bp product wasdigested with BglII and HindIII, and cloned into the BamHI/HindIII sitesof the Escherichia coli expression vector pT7H6UB (29). The resultingconstruct, pT7H6UBFX_P2_234–495, encoded a fusion protein withan N-terminal hexa-His tag followed by residues 2–76 of human ubiq-uitin, a factor Xa recognition site, and residues 234–495 of humanPAPP-A2. E. coli strain BL21 (DE3), transformed with
pT7H6UBFX_P2_234–495, was grown at 37 Cto an OD600 of approximately 0.8. Expressionwas induced by the addition of isopropyl-�-D-thiogalactopyranoside to a final concentration of1 mM. After 4 h at 37 C, the bacteria were har-vested by centrifugation, resuspended in 20 mM
Tris-HCl, 0.5 M NaCl (pH 8.0) (buffer A), anddisrupted by sonication, and the lysate was cen-trifuged. The pellet containing inclusion bodieswas washed three times with buffer A containing(2 M urea and 2% Triton X-100), and then redis-solved in buffer A containing 8 M urea, and loadedonto a Ni-NTA column (GE Healthcare, Piscat-away, NJ). Bound protein was eluted with bufferA containing 8 M urea and 20 mM EDTA andrefolded by rapid dilution into 0.4 M L-arginine,10 mM Tris-HCl (pH 8.0) (30). After concentra-tion, centrifugation, and dialysis into 20 mM Tris-HCl, 100 mM NaCl (pH 8.0), the fusion proteinwas cleaved with factor Xa (Sigma Chemical Co.,St. Louis, MO) to remove the ubiquitin domain.The recombinant PAPP-A2 domain was purifiedby ion-exchange chromatography on Q-Sepha-rose (GE Healthcare). Rabbits were immunizedwith antigen dissolved in Freund’s complete ad-juvant (Sigma) essentially as described (31).Freund’s incomplete adjuvant (Sigma) was usedas the vehicle in subsequent injections. Serum wascollected at 6-wk intervals.
Immunoblot analysisTo assess the specificity of the anti-PAPP-A2
sera, reduced recombinant PAPP-A2 (0.1 ng), hu-man term pregnancy serum (1 �l), purified PAPP-A/pro-major basic protein complex (0.1 �g) (32),and reduced recombinant PAPP-A (0.1 �g) (33)were separated on a 7% TA Novex gel under re-ducing conditions and transferred to a polyvinyli-dene difluoride (PVDF) membrane. Blocking wasperformed in 2% Tween 20 for 20 min. The mem-brane was incubated overnight at 4 C with eitheranti-PAPP-A2 serum or the preimmune serum (1:10,000 dissolved in blocking buffer). After wash-ing, the membrane was incubated with a second-ary antibody (antirabbit IgG Dako P217; 1:4000dissolved in blocking buffer) for 1 h at room tem-perature. To evaluate placental expression, 10 �gof a protein lysate prepared in RIPA lysis buffer[50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 0.5%Na deoxycholate (wt/vol), 0.1% sodium dodecylsulfate (wt/vol), and 1% Nonidet P-40 (vol/vol)]supplemented with 1% protease inhibitor cock-tail (Sigma) from basal plate specimens obtainedfrom PE (n � 10) or PTL (n � 9) patients wasseparated on a 4–20% gradient SDS-PAGE geland transferred to a PVDF membrane. Afterblocking nonspecific reactivity by incubating in5% BSA in Tris-buffered saline with Tween 20(10 mM Tris base; 150 mM NaCl; and 0.05%Tween 20, pH 8.4) for 1 h at room temperature,the membrane was incubated with anti-PAPP-A2
serum (1:10,000 diluted in blocking buffer) overnight at 4 C, washedwith Tris-buffered saline with Tween 20, followed by incubation withantirabbit horseradish peroxidase-conjugated antibody (1:10,000 dis-solved in blocking buffer) for 1 h at room temperature. Immunoreactivebands were visualized by using chemiluminescence (Pierce, Rockford, IL)and exposure to XR film (Kodak, Rochester, NY). To evaluate for pro-
FIG. 2. Heat map of differentially expressed genes in basal plate of PE placentas compared with controls.The normalized log intensity values for 71 differentially expressed probe sets were centered to the medianvalue of each probe set and colored on a range of �2.5 to �2.5. Red denotes up-regulated expressionlevels, and blue denotes down-regulated expression levels as compared with the median value. Columnscontain data from a single basal plate specimen, and rows correspond to a single probe set. Samples arearranged in order of increasing gestational age from left to right for each category. Rows are ranked byfold change [mean PE value (n � 12) divided by mean PTL value (n � 11)].
Endocrinology, January 2009, 150(1):452–462 endo.endojournals.org 455
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

tein loading, the PVDF membrane was stripped and incubated with anti-actin antibody (C4, 1:10,000; Sigma) for 1 h at room temperature andthen processed as described above. Densitometry was performed usingQuantityOne software (Bio-Rad, Hercules, CA).
PAPP-A2 immunohistochemistrySerial sections (5 �m) were deparaffinized and rehydrated before
antigen retrieval in citrate buffer (pH 6.8) for 20 min at 95–100 C.Endogenous peroxidase activity was quenched as described above. Non-specific reactivity was blocked by incubating the tissue sections withpreimmune serum (ABC Elite kits; Vector Laboratories) according to themanufacturer’s instructions before overnight exposure at 4 C to a pri-mary antibody dissolved in blocking buffer at the concentrations indi-cated [anti-PAPP-A2, 1:30,000; anti-PAPP-A (34), 1:50; anti-CK-7,1:100; and anti-HLA-G, 1:50 (35)]. Binding of the primary antibody wasdetected by incubating the tissue sections in a solution of the species-appropriate secondary antibodies provided in the ABC Elite kits (VectorLaboratories). Control sections were incubated without primary anti-body. Visualization, counterstaining, and imaging were performed asdescribed above.
Results
Using a microarray approach to analyze the basal plate regionsof placentas obtained from women who experienced PTL (n �
11) and women whose pregnancies were complicated by PE (n �
12), we identified 55 genes (71 probe sets) that were differentiallyexpressed (Fig. 2). Inclusion criteria for the PTL patients in-cluded a singleton pregnancy, no evidence of infection (clinical orhistological), absence of premature rupture of membranes, andno significant maternal disease. Approximately one third of thewomen delivered due to a diagnosis of cervical insufficiency. ThePE patients fulfilled the criteria for PE as defined in a 2002 Amer-ican College of Obstetrics and Gynecology bulletin (36). Clinicalcharacteristics of the two groups are compared in Table 1. Themajority of PE patients (10 of 12) experienced labor (with orwithout a cesarean section). Although more patients with PE(n � 6) than with PTL (n � 2) delivered by cesarean section, posthoc statistical analyses of the data showed that the method ofdelivery was not a confounding factor. The characteristics thatdefined the PE group are shown in Table 2. Among the mosthighly up-regulated genes were those that encode moleculeswhose expression was previously reported to be elevated in PE:leptin, VEGFR-1 (the gene that is alternatively spliced to producesFlt-1), and CRH. In addition, this analysis revealed more than40 novel transcripts, which to our knowledge have not previ-ously been reported to be differentially expressed in PE or asso-ciated with the pathogenesis of this syndrome.
To rule out the possibility that clinical and/or experimentalvariables accounted for the observed expression patterns, wesearched for possible correlations. We found no association withgestational age, parity, labor, maternal age, or fetal sex. Addi-tional variables that were considered included time between be-tamethasone injection and delivery, other common medicationsthat are used in these patient populations, mode of delivery,criteria for diagnosing PE, RNA degradation value, and tissue-processing time. Furthermore, we evaluated the level of the CTB-specific molecule CK-7, which is expressed by only trophoblasts,to determine that differences did not simply reflect different ra-tios of fetal to maternal tissue contribution to the samples. CK-7levels were not different between the PE and PTL samples or withgestational age.
Next, we used a Q-PCR approach to validate the expressionpatterns of nine genes. They included those genes whose normalexpression patterns are known to be dysregulated in PE, whichserved as positive controls, and a number of novel molecules overa range of expression. The results of these experiments confirmedthe expression patterns observed in the microarray analysis; allwere statistically significant except hydroxysteroid (17-�) dehy-drogenase 1, which showed the same trend (Fig. 3). For com-parison, the microarray data are plotted as insets to the individ-ual bar graphs. In accordance with many published studies thathave profiled the expression patterns of individual molecules inPE (37, 38), considerable variation among individual sampleswas apparent. This may be a reflection of the heterogeneity of thefetal and maternal responses to PE.
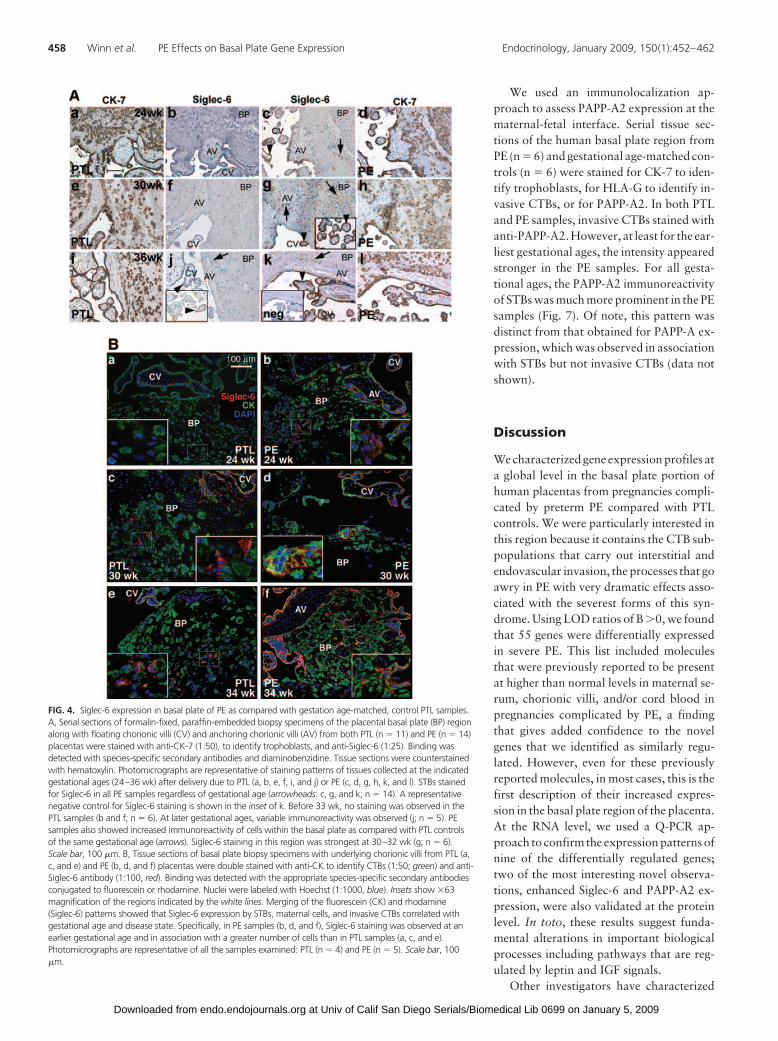
To further validate our findings, we performed a more ex-tensive, protein-level analysis of novel molecules with PE-asso-ciated alterations in their expression patterns that potentiallycould play important functions in the pathogenesis of thissyndrome. In particular, we focused on Siglec-6, which was orig-inally cloned on the basis of its ability to bind leptin (26). Inter-estingly, leptin expression, known to be dysregulated in PE (39–44), was also markedly increased in our microarray data set (Fig.2), but the classical leptin receptors (e.g. ObR) were not. Byapplying an immunohistochemical approach, we showed thatSiglec-6 expression was in general more pronounced in the basalplate of PE samples (n � 14) than in the PTL controls (n � 11,Fig. 4A). Additionally, we found that PE was associated with adramatic increase in STB expression of Siglec-6, particularly ear-lier in gestation (�33 wk), when there was no immunoreactivityin the PTL samples and clear immunoreactivity in PE.
TABLE 1. Clinical characteristics of study groups
PTL (n � 11) PE (n � 12)
Maternal age (yr) 30.2 � 7.1 30.7 � 9.1Gestational age (wk) 31.0 � 4.6 32.1 � 3.3Nulliparous 7 (64%) 9 (75%)Cesarean section 2 (18%) 6 (50%)Labored 11 (100%) 10 (83%)
Statistical analysis was by � 2 test. Differences between the two groups were notsignificant.
TABLE 2. Characteristics of patients with severe PE (n � 12)
No. ofpatients
Severe PE blood pressures 9 (83%)Severe PE proteinuria 6 (50%)Severe PE abnormal laboratory results 2 (17%)Eclampsia 2 (17%)Superimposed PE 2 (17%)Fetal abnormalitya 5 (42%)
The median creatinine was 0.9 (range, 0.5–1.3).a Growth restriction, oligohydramnios, or abnormal Doppler ultrasound.
456 Winn et al. PE Effects on Basal Plate Gene Expression Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

To identify the cells in the basal plate region that displayedSiglec-6 immunoreactivity, we costained tissue sections from PE(n � 5) and from PTL controls (n � 4) with anti-CK-7, whichidentifies all subsets of fetal trophoblasts, and with anti-Siglec-6.The results are shown in Fig. 4B. At 24 wk gestation, the PTLtissue exhibited no immunoreactivity. At the same gestationalage, STBs and a subset of the maternal cells stained with anti-Siglec-6 in the PE specimens. By 30 wk, the PTL samples showedSTB and rare maternal cell staining, reminiscent of the pattern inPE at 24 wk. The fact that Siglec-6 was detected by immunoflu-orescence at this gestational age and not by immunohistochem-istry likely reflects differences in the sensitivity of the methods. InPE tissue at 30 wk, a subset of invasive CTBs also expressed theSiglec-6 antigen. At 36 wk, PTL samples displayed invasive CTBSiglec-6 staining for the first time and continued to have moreSTB expression. However, the PE sample at 36 wk still showedmore Siglec-6 staining overall. This staining pattern may reflect
some aspects of premature placental aging or degenerativechanges in PE.
We also validated PAPP-A2 expression at the protein levelbecause of its known role in IGF biology, which plays importantroles in implantation and placentation. Because antibodies thatspecifically reacted with PAPP-A2 were not commercially avail-able, we produced a fragment of the PAPP-A2 protein in E. coliand used it to immunize rabbits for the purpose of producingpolyclonal antibodies. The PAPP-A2 serum, which specificallyrecognized PAPP-A2, did not cross-react with PAPP-A, the onlyhomolog of PAPP-A2 encoded by the human genome (Fig. 5). Insubsequent experiments, we used this antibody for immunoblotanalyses, which confirmed increased levels of PAPP-A2 in the PEbasal plate compared with PTL (Fig. 6). Densitometry ofPAPP-A2 normalized to actin showed roughly a 4.5-fold-highermean expression in PE compared with PTL (31.35 � 11.5 vs.7.05 � 3.53; P � 0.00001).
FIG. 3. Q-PCR confirmation of a subset of differentially expressed genes. Total RNA isolated from basal plate biopsies obtained from PTL (24–36 wk; n � 10) or PE(24–36 wk; n � 10) was analyzed using TaqMan primer/probe sets. Relative RNA levels were normalized to 18S values and then divided by a calibrator, in this case a24-wk PTL sample. Each bar represents the mean � SD of triplicate determinations. PTL, gray; PE, black. Dashed lines are the mean values for the PTL or PE RNAs.Samples are arranged in order of increasing gestational age from left to right for each category. Significance was determined by using Student’s t test (P � 0.05). Forcomparison, the insets show the corresponding microarray log intensity data for the same samples (log2). A, Q-PCR data for transcripts that were expected to be up-regulated in PE: CRH (P � 0.0005), leptin (P � 0.005), and Fms-related tyrosine kinase 1 (Flt-1; P � 0.005). B, Q-PCR data for novel transcripts up-regulated in PE:Siglec-6 (P � 0.004), PAPP-A2 (P � 0.005), and epidermal growth factor receptor pathway substrate 8-like protein 1 (EPS8L1; P � 0.005). C, Q-PCR data for noveltranscripts down-regulated in PE: KIAA1211 protein (P � 0.005), ankyrin repeat, SOCS box-containing 2 (ASB2; P � 0.05), and hydroxysteroid (17-�) dehydrogenase 1(HSD17B1; P � 0.08).
Endocrinology, January 2009, 150(1):452–462 endo.endojournals.org 457
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from
We used an immunolocalization ap-proach to assess PAPP-A2 expression at thematernal-fetal interface. Serial tissue sec-tions of the human basal plate region fromPE (n � 6) and gestational age-matched con-trols (n � 6) were stained for CK-7 to iden-tify trophoblasts, for HLA-G to identify in-vasive CTBs, or for PAPP-A2. In both PTLand PE samples, invasive CTBs stained withanti-PAPP-A2. However, at least for the ear-liest gestational ages, the intensity appearedstronger in the PE samples. For all gesta-tional ages, the PAPP-A2 immunoreactivityof STBs was much more prominent in the PEsamples (Fig. 7). Of note, this pattern wasdistinct from that obtained for PAPP-A ex-pression, which was observed in associationwith STBs but not invasive CTBs (data notshown).
Discussion
We characterized gene expression profiles ata global level in the basal plate portion ofhuman placentas from pregnancies compli-cated by preterm PE compared with PTLcontrols. We were particularly interested inthis region because it contains the CTB sub-populations that carry out interstitial andendovascular invasion, the processes that goawry in PE with very dramatic effects asso-ciated with the severest forms of this syn-drome. Using LOD ratios of B �0, we foundthat 55 genes were differentially expressedin severe PE. This list included moleculesthat were previously reported to be presentat higher than normal levels in maternal se-rum, chorionic villi, and/or cord blood inpregnancies complicated by PE, a findingthat gives added confidence to the novelgenes that we identified as similarly regu-lated. However, even for these previouslyreported molecules, in most cases, this is thefirst description of their increased expres-sion in the basal plate region of the placenta.At the RNA level, we used a Q-PCR ap-proach to confirm the expression patterns ofnine of the differentially regulated genes;two of the most interesting novel observa-tions, enhanced Siglec-6 and PAPP-A2 ex-pression, were also validated at the proteinlevel. In toto, these results suggest funda-mental alterations in important biologicalprocesses including pathways that are reg-ulated by leptin and IGF signals.
Other investigators have characterized
FIG. 4. Siglec-6 expression in basal plate of PE as compared with gestation age-matched, control PTL samples.A, Serial sections of formalin-fixed, paraffin-embedded biopsy specimens of the placental basal plate (BP) regionalong with floating chorionic villi (CV) and anchoring chorionic villi (AV) from both PTL (n � 11) and PE (n � 14)placentas were stained with anti-CK-7 (1:50), to identify trophoblasts, and anti-Siglec-6 (1:25). Binding wasdetected with species-specific secondary antibodies and diaminobenzidine. Tissue sections were counterstainedwith hematoxylin. Photomicrographs are representative of staining patterns of tissues collected at the indicatedgestational ages (24–36 wk) after delivery due to PTL (a, b, e, f, i, and j) or PE (c, d, g, h, k, and l). STBs stainedfor Siglec-6 in all PE samples regardless of gestational age (arrowheads: c, g, and k; n � 14). A representativenegative control for Siglec-6 staining is shown in the inset of k. Before 33 wk, no staining was observed in thePTL samples (b and f; n � 6). At later gestational ages, variable immunoreactivity was observed (j; n � 5). PEsamples also showed increased immunoreactivity of cells within the basal plate as compared with PTL controlsof the same gestational age (arrows). Siglec-6 staining in this region was strongest at 30–32 wk (g; n � 6).Scale bar, 100 �m. B, Tissue sections of basal plate biopsy specimens with underlying chorionic villi from PTL (a,c, and e) and PE (b, d, and f) placentas were double stained with anti-CK to identify CTBs (1:50; green) and anti-Siglec-6 antibody (1:100, red). Binding was detected with the appropriate species-specific secondary antibodiesconjugated to fluorescein or rhodamine. Nuclei were labeled with Hoechst (1:1000, blue). Insets show �63magnification of the regions indicated by the white lines. Merging of the fluorescein (CK) and rhodamine(Siglec-6) patterns showed that Siglec-6 expression by STBs, maternal cells, and invasive CTBs correlated withgestational age and disease state. Specifically, in PE samples (b, d, and f), Siglec-6 staining was observed at anearlier gestational age and in association with a greater number of cells than in PTL samples (a, c, and e).Photomicrographs are representative of all the samples examined: PTL (n � 4) and PE (n � 5). Scale bar, 100�m.
458 Winn et al. PE Effects on Basal Plate Gene Expression Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

(human) placental gene expression patterns in PE compared withnormal pregnancy. For example, Reimer et al. (45) demonstrateda large increase in leptin production by chorionic villi. A small-scale study with the same design found a PE-associated up-reg-ulation of the gene encoding the muscle subunit of glycogenphosphorylase (46); an increase in the expression of the solubleform of the disintegrin, ADAM 12, was discovered in the sameway (47). The fact that we did not identify the latter two mol-ecules is likely attributable to differences in the cellular compo-sition of our samples, which contained a much smaller propor-tion of chorionic villi and, consequently, STBs that release amyriad of substances into maternal blood. Other relevant con-siderations include the fact that the causes of PE are complex. Forexample, in some instances, poor placentation leads to intra-uterine growth restriction without PE (7). Conversely, abnormalmaternal responses may occur even if placentation is normal (48,49). In this case, the metabolic, vascular, and immunologicaldemands of pregnancy may synergize with the effects of preex-isting medical conditions, such as arterial disease, hypertension,obesity, and diabetes, which in turn strongly predispose womento develop PE. Interestingly, many of these disease processes areassociated with altered leptin levels. Numerous investigatorshave reported a PE-associated increase in circulating levels ofleptin (39–44, 50–52), and a leptin gene polymorphism has beenlinked to an increased risk of developing this pregnancy com-plication (53). However, a clear picture of how an increase inleptin expression is linked to the pathophysiology of PE has yetto emerge.
Our data demonstrated increased leptin expression in thebasal plate of PE placentas compared with control tissue. Inter-estingly, although the classic leptin receptors were not differen-tially expressed, we observed elevated levels of the mRNA thatencodes Siglec-6, a transmembrane protein that also binds leptin.These findings suggest that this molecule may play an importantrole as a placental leptin receptor and that increased Siglec-6
levels could contribute to the pathogenesis of PE. Although thecloning strategy for Siglec-6 was based on its ability to interactwith leptin, the other Siglec family members bind sialic acid-containing glycans. Some family members have exquisite ligandspecificity, whereas others have less rigid structural requirements(54). For example, Siglec-6 has binding specificity for the sia-lyl-Tn epitope (Sia�2–6Gal-NAc�1-O-R, where R is a serine orthreonine). Published data suggest that in the placenta, leptin isa Siglec-6 ligand, but the endogenous binding partners have yetto be identified (26). Additionally, Siglec-6 expression has otherinteresting features. For example, in humans, it is restricted to theplacenta and B lymphocytes. In other species, including nonhu-man primates, placental cells lack Siglec-6 expression, whereasthe B cells exhibit it (25). The fact that Siglec-6 is expressed onlyin human placentas and not in nonhuman primate placentas (25)is extremely intriguing, because PE is thought to be a uniquelyhuman disease; spontaneous PE has not been reported in otheranimals, even nonhuman primates (55).
The function(s) of Siglec-6 in any biological system, includingB cells, have yet to be determined. A recent study suggested a rolefor Siglec-6 in term labor (25). Interestingly, Siglec-6 expressionwas not observed in our earliest gestation PTL samples but didincrease as pregnancy advanced. Siglec-6 has conserved immu-noreceptor tyrosine-based inhibitory motif (ITIM) and ITIM-like domains, suggesting that ligand binding results in tyrosinephosphorylation and recruitment of Src homology domain-con-taining adapters with inhibitory effects. The effect of leptin hasbeen investigated in mice and bats (56, 57). In both species, leptinappears to have a stimulatory effect on trophoblast invasion invitro, but its mechanism of action is not known. Determiningwhether leptin and Siglec-6 function during human CTB inva-sion is a critical first step in understanding whether these mol-ecules play a functional role in PE pathogenesis.
A PE-associated increase in the expression of PAPP-A2 wasanother novel observation that emerged from our work. This isthe first report of PAPP-A2 expression at the protein level in theplacenta, experiments that were made possible by the generationof rabbit polyclonal antibodies that specifically reacted with thismolecule. PAPP-A2, which has 46% sequence identity withPAPP-A, is a metalloproteinase that cleaves IGF-binding pro-tein-5 (IGFBP-5) (28). Depending on the cellular context, IGFBPseither stimulate or inhibit IGF-I and -II actions. For example, insmooth muscle cells, IGFBP-5 binds to extracellular matrix com-
FIG. 5. Immunoblotting showed that the anti-PAPP-A2 polyclonal rabbitantiserum reacted with a single band of the appropriate molecular weight. A,The samples were reduced and separated on a 7% TA Novex gel: lane 1,supernatant from mock-transfected HEK293 cells; lane 2, supernatant fromHEK293 cells transfected with PAPP-A2 cDNA; lane 3, term pregnancy serum;lane 4, purified PAPP-A/pro-major basic protein complex; lane 5, supernatantfrom HEK293 cells transfected with PAPP-A cDNA. The samples were transferredto a PVDF membrane that was incubated overnight with anti-PAPP-A2(1:10,000). Immunoreactive bands were visualized by incubating the blot with ahorseradish peroxidase-conjugated secondary antibody (1:4000) for 1 h at roomtemperature. A band of about 250 relative molecular mass was detected only inthe lanes that contained the recombinant PAPP-A2 protein (lane 2) andpregnancy serum (lane 3). There was no cross-reactivity with the related proteinPAPP-A (lanes 4 and 5). B, Identical blot processed the same as in A exceptincubated with rabbit preimmune serum. Nonspecific bands (110 and 135relative molecular mass) were detected in the serum sample (lane 3) only uponoverexposure.
FIG. 6. PAPP-A2 protein levels were increased in the basal plate portion ofplacentas from PE pregnancies. Protein lysates from basal plate biopsy samples(10 �g) of PTL (n � 9) and PE (n � 9) placentas were separated by SDS-PAGE(4–20% gel). Maternal serum collected at term served as a positive control.Immunoblotting with anti-PAPP-A2 (1:30,000) showed increased immunoreactivityfor PAPP-A2 in PE as compared with PTL samples. Immunoblotting with anti-actin(clone C4, 1:10,000) was used as a measure of protein loading. Samples arearranged in order of increasing gestational age from left to right for each category,with numbers corresponding to gestational age (weeks.days).
Endocrinology, January 2009, 150(1):452–462 endo.endojournals.org 459
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

ponents, including thrombospondin-1 and osteopontin, and po-tentiates the cells’ response to IGF-I. In contrast, IGFBP-4, aPAPP-A substrate, largely inhibits IGF actions (58). In a fibro-blast model, an increase in IGFBP-5 proteolysis attenuates itsstimulatory effects on cell migration (59). If CTBs respond in ananalogous manner, then the observed PE-associated increase inPAPP-A2 levels could inhibit CTB invasion by mechanisms thatinclude an increase in IGFBP-5 proteolysis.
Data suggest that there is cross talk between the (classical)leptin and insulin signaling pathways (60). Specifically, suppres-sor of cytokine signaling (SOCS-3) attenuates both pathways(61). Furthermore, related Siglec family members’ (CD33/Siglec-3 and Siglec-7) responses are blocked by SOCS-3 via ac-celerated proteasomal degradation (62, 63). Given that IGF andinsulin signal through the same pathways, it is possible that al-terations in leptin, Siglec-6, and PAPP-A2 levels may be workingin concert to exert inhibitory effects on CTB invasion. Under-standing the pregnancy-related functions of these molecules andthe other genes that were differentially expressed in PE will likelylead to a better understanding of the pathogenesis of this human-specific condition, the crucial first step in the rational design oftreatments (both preventative and therapeutic) that address the
causes, rather than the consequences, of thispregnancy complication. Additionally, weenvision that PE-associated alterations inthe expression of gene products that are se-creted in maternal blood can be used to pre-dict a woman’s risk of developing this syn-drome and/or enable diagnosis during theearly stages of PE when the signs are some-times ambiguous.
Acknowledgments
We thank Ms. Jean Perry, the study research nursecoordinator, who assisted in tissue collection, asdid the nurses, residents, and faculty at the UCSFBirth Center. Dr. Chris Barker, Dr. Chandi Grif-fin, and Ms. Jennifer Gregg, members of the UCSFGladstone (National Heart, Lung, and Blood In-stitute) Shared Microarray Facility, contributedinvaluable technical and intellectual expertise.We also thank Dr. David Erle and MichaelSalazar, members of the Sandler Genomics CoreFacility, and Nissi Varki (University of Califor-nia, San Diego) for helpful discussions. We aregrateful to Mr. Evan Messenger and Dr. KathyIvanetich, UCSF Biomolecular Research Center,for technical assistance in performing the Q-PCRexperiments. We thank Ms. Mary McKenney forcritical review of the manuscript.
Address all correspondence and requests for re-prints to: Virginia D. Winn, M.D., Ph.D., Univer-sity of Colorado Denver, Department of Obstet-rics and Gynecology, Reproductive Sciences,Mail Stop 8309, 12800 East 19th Avenue, P.O.Box 6511, Aurora, Colorado 80045. E-mail:[email protected].
This work was supported by National Insti-tutes of Health Grants R01 HL 64597 (to S.J.F.),
R01 GM 32373 (to A.V.), 5-MO1-RR 00083 (General Clinical ResearchCenter, San Francisco General Hospital), HL 072301 (UCSF, NationalHeart, Lung, and Blood Institute Shared Microarray Facility), and theDanish Medical Research Council (to C.O.). V.D.W. is a March of DimesReproductive Scientists Development Program scholar (National Insti-tute of Child Health and Human Development 5K12HD00849) and anAmerican Board of Obstetrics and Gynecology/American Associationof Obstetricians and Gynecologists Foundation scholar.
Disclosure Statement: M.G., A.G., K.K.-S., A.K., K.R., R.-F.Y.,M.T.O., and A.V. have nothing to declare. V.D.W., R.H.-K., and S.J.F.have a patent filed for the array data. V.D.W., C.O., and S.J.F. haveresearch grants as listed above.
References
1. Roberts JM, Cooper DW 2001 Pathogenesis and genetics of pre-eclampsia.Lancet 357:53–56
2. Gomez O, Martinez JM, Figueras F, Del Rio M, Borobio V, Puerto B, Coll O,Cararach V, Vanrell JA 2005 Uterine artery Doppler at 11–14 weeks of ges-tation to screen for hypertensive disorders and associated complications in anunselected population. Ultrasound Obstet Gynecol 26:490–494
3. Hershkovitz R, de Swiet M, Kingdom J 2005 Mid-trimester placentation as-sessment in high-risk pregnancies using maternal serum screening and uterineartery Doppler. Hypertens Pregnancy 24:273–280
FIG. 7. PE is associated with enhanced staining of invasive CTBs and STBs for PAPP-A2. Serial sections offormalin-fixed, paraffin-embedded basal plate biopsy specimens of placentas from PTL (A–C and G–I) andPE (D–F and J–L) at various gestational ages were stained with anti-CK-7 (1:100) to identify trophoblasts,anti-HLA-G (1:50) to identify invasive CTBs, or anti-PAPP-A2 (1:30,000). Binding of the primary antibodywas detected with the appropriate species-specific secondary antibody and diaminobenzidine. Tissuesections were counterstained with hematoxylin. Invasive CTBs (*) reacted with anti-PAPP-A2 in all thesamples, but the staining intensity (when using HLA-G and CK-7 levels for comparison) was greater in PE atthe earlier gestational ages than in PTL. In contrast, PAPP-A2 immunoreactivity of STB, which was aprominent feature of PE samples (F and L, arrows), was largely absent in PTL specimens (C and I). Negativecontrols for PAPP-A2 staining are shown as insets (C, F, I, and L). Photomicrographs are representative ofmultiple analyses (PTL, n � 6; PE, n � 6). V, Anchoring chorionic villi; CV, floating chorionic villi. Scale bar, 200�m.
460 Winn et al. PE Effects on Basal Plate Gene Expression Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

4. Brosens IA, Robertson WB, Dixon HG 1972 The role of the spiral arteries inthe pathogenesis of preeclampsia. Obstet Gynecol Annu 1:177–191
5. Gerretsen G, Huisjes HJ, Elema JD 1981 Morphological changes of the spiralarteries in the placental bed in relation to pre-eclampsia and fetal growthretardation. Br J Obstet Gynaecol 88:876–881
6. Moodley J, Ramsaroop R 1989 Placental bed morphology in black womenwith eclampsia. S Afr Med J 75:376–378
7. Khong TY, De Wolf F, Robertson WB, Brosens I 1986 Inadequate maternalvascular response to placentation in pregnancies complicated by pre-eclampsiaand by small-for-gestational age infants. Br J Obstet Gynaecol 93:1049–1059
8. Roberts JM, Taylor RN, Musci TJ, Rodgers GM, Hubel CA, McLaughlin MK1989 Preeclampsia: an endothelial cell disorder. Am J Obstet Gynecol 161:1200–1204
9. Roberts JM 1998 Endothelial dysfunction in preeclampsia. Semin ReprodEndocrinol 16:5–15
10. Friedman SA, Schiff E, Emeis JJ, Dekker GA, Sibai BM 1995 Biochemicalcorroboration of endothelial involvement in severe preeclampsia. Am J ObstetGynecol 172:202–203
11. Hsu CD, Iriye B, Johnson TR, Witter FR, Hong SF, Chan DW 1993 Elevatedcirculating thrombomodulin in severe preeclampsia. Am J Obstet Gynecol169:148–149
12. Taylor RN, Crombleholme WR, Friedman SA, Jones LA, Casal DC, RobertsJM 1991 High plasma cellular fibronectin levels correlate with biochemicaland clinical features of preeclampsia but cannot be attributed to hypertensionalone. Am J Obstet Gynecol 165:895–901
13. Tyurin VA, Liu SX, Tyurina YY, Sussman NB, Hubel CA, Roberts JM, TaylorRN, Kagan VE 2001 Elevated levels of S-nitrosoalbumin in preeclampsiaplasma. Circ Res 88:1210–1215
14. Clark BA, Halvorson L, Sachs B, Epstein FH 1992 Plasma endothelin levels inpreeclampsia: elevation and correlation with uric acid levels and renal impair-ment. Am J Obstet Gynecol 166:962–968
15. Mills JL, DerSimonian R, Raymond E, Morrow JD, Roberts 2nd LJ, ClemensJD, Hauth JC, Catalano P, Sibai B, Curet LB, Levine RJ 1999 Prostacyclin andthromboxane changes predating clinical onset of preeclampsia: a multicenterprospective study. JAMA 282:356–362
16. Levine RJ, Lam C, Qian C, Yu KF, Maynard SE, Sachs BP, Sibai BM,Epstein FH, Romero R, Thadhani R, Karumanchi SA 2006 Soluble endog-lin and other circulating antiangiogenic factors in preeclampsia. N EnglJ Med 355:992–1005
17. Winn VD, Haimov-Kochman R, Paquet AC, Yang YJ, Madhusudhan MS,Gormley M, Feng KT, Bernlohr DA, McDonagh S, Pereira L, Sali A, FisherSJ 2007 Gene expression profiling of the human maternal-fetal interfacereveals dramatic changes between midgestation and term. Endocrinology148:1059 –1079
18. Zhou Y, Bianco K, Huang L, Nien JK, McMaster M, Romero R, Fisher SJ 2007Comparative analysis of the maternal-fetal interface in preeclampsia and pre-term labor. Cell Tissue Res 329:559–569
19. American College of Obstetricians and Gynecologists 1999 Practice bulletin10. Induction of labor. Washington, DC: American College of Obstetriciansand Gynecologists
20. Haimov-Kochman R, Fisher SJ, Winn VD 2006 Modification of the standardTrizol-based technique improves the integrity of RNA isolated from RNase-rich placental tissue. Clin Chem 52:159–160
21. Auer H, Lyianarachchi S, Newsom D, Klisovic MI, Marcucci G, Kornacker K2003 Chipping away at the chip bias: RNA degradation in microarray analysis.Nat Genet 35:292–293
22. Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, Dudoit S, Ellis B,Gautier L, Ge Y, Gentry J, Hornik K, Hothorn T, Huber W, Iacus S, IrizarryR, Leisch F, Li C, Maechler M, Rossini AJ, Sawitzki G, Smith C, Smyth G,Tierney L, Yang JY, Zhang J 2004 Bioconductor: open software developmentfor computational biology and bioinformatics. Genome Biol 5:R80
23. Bolstad BM 2004 Low level analysis of high-density oligonucleotide arraydata: background, normalization and summarization. Dissertation, Universityof California, Berkeley
24. Smyth GK 2005 Limma: linear models for microarray data. In: GentlemanR, Carey V, Huber W, Irizarry R, Dudoit S, eds. Bioinformatics and com-putational biology solutions Using R and Bioconductor. New York: Spring-er; 397– 420
25. Brinkman-Van der Linden EC, Hurtado-Ziola N, Hayakawa T, Wiggleton L,Benirschke K, Varki A, Varki N 2007 Human-specific expression of Siglec-6in the placenta. Glycobiology 17:922–931
26. Patel N, Brinkman-Van der Linden EC, Altmann SW, Gish K, BalasubramanianS, Timans JC, Peterson D, Bell MP, Bazan JF, Varki A, Kastelein RA 1999 OB-BP1/Siglec-6: a leptin- and sialic acid-binding protein of the immunoglobulin su-perfamily. J Biol Chem 274:22729–22738
27. Damsky CH, Fitzgerald ML, Fisher SJ 1992 Distribution patterns of extra-cellular matrix components and adhesion receptors are intricately modulatedduring first trimester cytotrophoblast differentiation along the invasive path-way, in vivo. J Clin Invest 89:210–222
28. Overgaard MT, Boldt HB, Laursen LS, Sottrup-Jensen L, Conover CA, OxvigC 2001 Pregnancy-associated plasma protein-A2 (PAPP-A2), a novel insulin-like growth factor-binding protein-5 proteinase. J Biol Chem 276:21849–21853
29. Ellgaard L, Holtet TL, Nielsen PR, Etzerodt M, Gliemann J, Thogersen HC1997 Dissection of the domain architecture of the �2macroglobulin-receptor-associated protein. Eur J Biochem 244:544–551
30. Anderson M, Blowers D, Hewitt N, Hedge P, Breeze A, Hampton I, Taylor I1999 Refolding, purification, and characterization of a loop deletion mu-tant of human Bcl-2 from bacterial inclusion bodies. Protein Expr Purif15:162–170
31. Harlow E. LD 1988 Antibodies: a laboratory manual. Cold Spring Harbor,NY: Cold Spring Harbor Laboratory Press
32. Oxvig C, Sand O, Kristensen T, Kristensen L, Sottrup-Jensen L 1994 Isolationand characterization of circulating complex between human pregnancy-asso-ciated plasma protein-A and proform of eosinophil major basic protein. Bio-chim Biophys Acta 1201:415–423
33. Overgaard MT, Haaning J, Boldt HB, Olsen IM, Laursen LS, Christiansen M,Gleich GJ, Sottrup-Jensen L, Conover CA, Oxvig C 2000 Expression of re-combinant human pregnancy-associated plasma protein-A and identificationof the proform of eosinophil major basic protein as its physiological inhibitor.J Biol Chem 275:31128–31133
34. Qin QP, Christiansen M, Oxvig C, Pettersson K, Sottrup-Jensen L, Koch C,Norgaard-Pedersen B 1997 Double-monoclonal immunofluorometric assaysfor pregnancy-associated plasma protein A/proeosinophil major basic protein(PAPP-A/proMBP) complex in first-trimester maternal serum screening forDown syndrome. Clin Chem 43:2323–2332
35. McMaster MT, Librach CL, Zhou Y, Lim KH, Janatpour MJ, DeMars R,Kovats S, Damsky C, Fisher SJ 1995 Human placental HLA-G expression isrestricted to differentiated cytotrophoblasts. J Immunol 154:3771–3778
36. American College of Obstetrics and Gynecology 2002 ACOG Practice Bulle-tin: diagnosis and management of preeclampsia and eclampsia. Obstet Gy-necol 99:159–167
37. Muttukrishna S, Knight PG, Groome NP, Redman CW, Ledger WL 1997Activin A and inhibin A as possible endocrine markers for pre-eclampsia.Lancet 349:1285–1288
38. Perkins AV, Linton EA, Eben F, Simpson J, Wolfe CD, Redman CW 1995Corticotrophin-releasing hormone and corticotrophin-releasing hormonebinding protein in normal and pre-eclamptic human pregnancies. Br J ObstetGynaecol 102:118–122
39. Laivuori H, Gallaher MJ, Collura L, Crombleholme WR, Markovic N,Rajakumar A, Hubel CA, Roberts JM, Powers RW 2006 Relationships be-tween maternal plasma leptin, placental leptin mRNA and protein in normalpregnancy, pre-eclampsia and intrauterine growth restriction without pre-eclampsia. Mol Hum Reprod 12:551–556
40. Li RH, Poon SC, Yu MY, Wong YF 2004 Expression of placental leptin andleptin receptors in preeclampsia. Int J Gynecol Pathol 23:378–385
41. Lu D, Yang X, Wu Y, Wang H, Huang H, Dong M 2006 Serum adiponectin,leptin and soluble leptin receptor in pre-eclampsia. Int J Gynaecol Obstet95:121–126
42. McCarthy JF, Misra DN, Roberts JM 1999 Maternal plasma leptin is increasedin preeclampsia and positively correlates with fetal cord concentration. Am JObstet Gynecol 180:731–736
43. Tommaselli GA, Pighetti M, Nasti A, D’Elia A, Guida M, Di Carlo C, BifulcoG, Nappi C 2004 Serum leptin levels and uterine Doppler flow velocimetry at20 weeks’ gestation as markers for the development of pre-eclampsia. GynecolEndocrinol 19:160–165
44. Vitoratos N, Chrystodoulacos G, Kouskouni E, Salamalekis E, Creatsas G2001 Alterations of maternal and fetal leptin concentrations in hypertensivedisorders of pregnancy. Eur J Obstet Gynecol Reprod Biol 96:59–62
45. Reimer T, Koczan D, Gerber B, Richter D, Thiesen HJ, Friese K 2002Microarray analysis of differentially expressed genes in placental tissue ofpre-eclampsia: up-regulation of obesity-related genes. Mol Hum Reprod8:674 – 680
46. Tsoi SC, Cale JM, Bird IM, Kay HH 2003 cDNA microarray analysis of geneexpression profiles in human placenta: up-regulation of the transcript encod-ing muscle subunit of glycogen phosphorylase in preeclampsia. J Soc GynecolInvestig 10:496–502
47. Gack S, Marme A, Marme F, Wrobel G, Vonderstrass B, Bastert G, Lichter P,Angel P, Schorpp-Kistner M 2005 Preeclampsia: increased expression of sol-uble ADAM 12. J Mol Med 83:887–896
Endocrinology, January 2009, 150(1):452–462 endo.endojournals.org 461
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from

48. Ness RB, Roberts JM 1996 Heterogeneous causes constituting the single syn-drome of preeclampsia: a hypothesis and its implications. Am J Obstet Gynecol175:1365–1370
49. Redman CW, Sargent IL 2005 Latest advances in understanding preeclampsia.Science 308:1592–1594
50. Laivuori H, Kaaja R, Koistinen H, Karonen SL, Andersson S, Koivisto V,Ylikorkala O 2000 Leptin during and after preeclamptic or normal pregnancy:its relation to serum insulin and insulin sensitivity. Metabolism 49:259–263
51. Mise H, Sagawa N, Matsumoto T, Yura S, Nanno H, Itoh H, Mori T,Masuzaki H, Hosoda K, Ogawa Y, Nakao K 1998 Augmented placental pro-duction of leptin in preeclampsia: possible involvement of placental hypoxia.J Clin Endocrinol Metab 83:3225–3229
52. Ramsay JE, Ferrell WR, Crawford L, Wallace AM, Greer IA, Sattar N 2004Divergent metabolic and vascular phenotypes in pre-eclampsia and intrauter-ine growth restriction: relevance of adiposity. J Hypertens 22:2177–2183
53. Muy-Rivera M, Ning Y, Frederic IO, Vadachkoria S, Luthy DA, Williams MA2005 Leptin, soluble leptin receptor and leptin gene polymorphism in relationto preeclampsia risk. Physiol Res 54:167–174
54. Varki A, Angata T 2006 Siglecs: the major subfamily of I-type lectins. Gly-cobiology 16:1R–27R
55. Chez RA 1976 Nonhuman primate models of toxemia of pregnancy. PerspectNephrol Hypertens 5:421–424
56. Schulz LC, Townsend K, Kunz TH, Widmaier EP 2007 Inhibition of tropho-blast invasiveness in vitro by immunoneutralization of leptin in the bat, Myotislucifugus (Chiroptera). Gen Comp Endocrinol 150:59–65
57. Schulz LC, Widmaier EP 2004 The effect of leptin on mouse trophoblast cellinvasion. Biol Reprod 71:1963–1967
58. Firth SM, Baxter RC 2002 Cellular actions of the insulin-like growth factorbinding proteins. Endocr Rev 23:824–854
59. Xu Q, Yan B, Li S, Duan C 2004 Fibronectin binds insulin-like growth factor-binding protein 5 and abolishes its ligand-dependent action on cell migration.J Biol Chem 279:4269–4277
60. Fruhbeck G 2006 Intracellular signalling pathways activated by leptin. Bio-chem J 393:7–20
61. Howard JK, Flier JS 2006 Attenuation of leptin and insulin signaling by SOCSproteins. Trends Endocrinol Metab 17:365–371
62. Orr SJ, Morgan NM, Buick RJ, Boyd CR, Elliott J, Burrows JF, JefferiesCA, Crocker PR, Johnston JA 2007 SOCS3 targets Siglec 7 for proteasomaldegradation and blocks Siglec 7-mediated responses. J Biol Chem 282:3418 –3422
63. Orr SJ, Morgan NM, Elliott J, Burrows JF, Scott CJ, McVicar DW, JohnstonJA 2007 CD33 responses are blocked by SOCS3 through accelerated protea-somal-mediated turnover. Blood 109:1061–1068
462 Winn et al. PE Effects on Basal Plate Gene Expression Endocrinology, January 2009, 150(1):452–462
at Univ of Calif San Diego Serials/Biomedical Lib 0699 on January 5, 2009 endo.endojournals.orgDownloaded from


















