Serum effects on the response of mammalian cells to the exotoxins of Pseudomonas...
8
Serum effects on the response of mammalian cells to the exotoxins of Pseudomonas aeruginosa and Corynebacterium diphtheriae JOHN L. MIDDLEBROOK AND REBECCA B. DORLAND Potlrology Divisior~, Utritecl Stcrtrs A I Y ~ I ) . Merlic.crl Resenrc.11 It~stitrrtr (~f'Ir;fc~.tiorrs Disecrses. Fredrrick, M D , U.S.A. 21701 Accepted November 2, 1976 MIDDLEBROOK, J. L., and R. B. DORLAND. 1977. Serum effects on the response of mammalian cells to the exotoxins of Pserrclor17or1crsrrrrrrgitroscl and Coryrreherctc,rirr,,~ eliplrtlro.ioe. Can. J. Microbiol. 23: 175-1232, The response of mammalian cells to P~erlclor?lorrns and diphtheria exotoxins was studied. A method was developed whereby the sensitivity of cells to these two toxins could be quantitated. The method is versatile and can be used to study the effects of toxins on many cellular metabolic or transport processes. The type of serum used in the culture medium significantly influenced the response of cells to the toxins. Calf, horse, and human sera protected cells while fetal calf serum did not. Precipitation with (NH4)IS0, demonstl.ated the probable presence of toxin-specific antibody in the protective calf serum while none was detected in the nonprotective fetal calf selum. The level of antibody in calf semm, as titrated by hemagglutination, was sufficient to account for all the obselved protection. It is suggested that fetal calf serum be used for all fi~tlr~e cell culture studies of bacterial toxins. MIDDLEBROOK. J. L.. et R. B. DORLAND. 1977. S e ~ u m effects on the response of mammalian cells to the exotoxins of P.sc,11dor11orros ctc,rrrgirroso and Coryrrc~hac~~rittr~~ clil)lrrlrc,rioe. Can. J. Microbiol. 23: 175-1232, I1 s'agit d'une etude de la riponse de cellules de mammiferes aux exotoxines de P.s~rrtlor1rorrtr.s et du bacille diphterique. On amis au point une methode qui permet de quantifier la sensibilite des cellules B ces deux toxines. Cette methode est versatile et peut s'appliqueri l'etude de I'effet des toxines surplusieurs voies metaboliques ou de transport. La nature du selum incorport au milieu de culture semble avoir une influence significative sur la reponse des cellules aux toxines. Les s61ums de veau et de cheval et le s61um humain protegent les cellules ce qui n'est pas le cas du se~um de veau fetal. La pllcipitation par le (NH4)?S0, revele la presence probable d'antico~ps specifiques i la toxine dans le s e ~ u m de veau protecteuralors qu'on ne detecte rien dans le serum de veau fetal non-protecteur. Le taux d'anticorpsdans le s e ~ u m de veau, mesure par hemaggluti- nation passive, est assezelevi pourjustifierlaprotection observee. On recommandequele s61um de veau fetal soit desormaisemploye dans toutes les cultures cellulairesutilisCes dans I'etudedes toxines bacteriennes. [Traduit par le journal] Studies of the effects of bacterial toxins on cells in culture have made substantial contribu- tions towards our understanding of the mode of toxin action (23). Generally, two approaches have been used. In the first, workers have at- tempted to correlate the toxin sensitivity of cell lines derived from various tissues and species with the histopathology of animal intoxication. In the second, a single cell line has been used death in human diphtheria infection, Bonventre and Imhoff (1) examined heart cells in culture and found them to be unusually sensitive to the toxin. Finally, many of the insights leading to the currently accepted mechanism of action of diphtheria toxin (DE) were obtained using cultured HeLa cells (3, 6, 18, 24). Our laboratory is engaged in an in-depth com- parative study of diphtheria exotoxin and as a model to work out details of the toxin's exotoxin A (PE) from Pseudo~nonns oer~/ginosa mechanism of action. Probably the toxin most (12, 13). We began by coinparing the sensitivities successfully studied using these two approaches of many mammalian cell lines to both toxins is the exotoxin of Corynebacteriunz diphtheriae. (15). During the course of our work we found Solotorovsky and Gabliks (22) reported a close that some types of sera commonly used in cell correlation between the relative sensitivity of a culture could partially protect cells from the cell line to the toxin and the sensitivity of its cytotoxic actions of both toxins. We investigated animal donor. Moreover, following reports the nature of this artifact and this report that cardiac failure was a frequent cause of describes our findings. We believe that low Can. J. Microbiol. Downloaded from www.nrcresearchpress.com by Ryerson University Library on 04/16/13 For personal use only.
Transcript of Serum effects on the response of mammalian cells to the exotoxins of Pseudomonas...

Serum effects on the response of mammalian cells to the exotoxins of Pseudomonas aeruginosa and Corynebacterium diphtheriae
JOHN L. MIDDLEBROOK A N D REBECCA B. DORLAND Potlrology Divisior~, Utritecl Stcrtrs A I Y ~ I ) . Merlic.crl Resenrc.11 It~stitrrtr (~f'Ir;fc~.tiorrs Disecrses.
Fredrrick, M D , U . S . A . 21701
Accepted November 2, 1976
MIDDLEBROOK, J . L., and R . B. DORLAND. 1977. Serum effects on the response of mammalian cells to the exotoxins of Pserrclor17or1crs rrrrrrgitroscl and Coryrreherctc,rirr,,~ eliplrtlro.ioe. Can. J . Microbiol. 23: 175-1232,
The response of mammalian cells to P~erlclor?lorrns and diphtheria exotoxins was studied. A method was developed whereby the sensitivity of cells to these two toxins could be quantitated. The method is versatile and can be used to study the effects of toxins on many cellular metabolic or transport processes. The type of serum used in the culture medium significantly influenced the response of cells to the toxins. Calf, horse, and human sera protected cells while fetal calf serum did not. Precipitation with (NH4)IS0, demonstl.ated the probable presence of toxin-specific antibody in the protective calf serum while none was detected in the nonprotective fetal calf selum. The level of antibody in calf semm, as titrated by hemagglutination, was sufficient to account for all the obselved protection. It is suggested that fetal calf serum be used for all f i ~ t l r ~ e cell culture studies of bacterial toxins.
MIDDLEBROOK. J . L.. et R. B. DORLAND. 1977. S e ~ u m effects on the response of mammalian cells to the exotoxins of P.sc,11dor11orros ctc,rrrgirroso and Coryr rc~hac~~r i t t r~~ clil)lrrlrc,rioe. Can. J . Microbiol. 23: 175-1232,
I1 s'agit d'une etude de la riponse d e cellules de mammiferes aux exotoxines de P.s~rrtlor1rorrtr.s et du bacille diphterique. On amis au point une methode qui permet de quantifier la sensibilite des cellules B ces deux toxines. Cette methode est versatile et peut s 'appliqueri l'etude de I'effet des toxines surplusieurs voies metaboliques ou de transport. La nature du selum incorport au milieu de culture semble avoir une influence significative sur la reponse des cellules aux toxines. Les s61ums de veau et de cheval et le s61um humain protegent les cellules ce qui n'est pas le cas du s e ~ u m de veau fe ta l . La pllcipitation par le (NH4)?S0, revele la presence probable d 'antico~ps specifiques i la toxine dans le s e ~ u m de veau protecteuralors qu'on ne detecte rien dans le serum de veau fetal non-protecteur. Le taux d'anticorpsdans le s e ~ u m de veau, mesure par hemaggluti- nation passive, est assezelevi pourjustifierlaprotection observee. On recommandequele s61um de veau fe ta l soit desormaisemploye dans toutes les cultures cellulairesutilisCes dans I'etudedes toxines bacteriennes.
[Traduit par le journal]
Studies of the effects of bacterial toxins on cells in culture have made substantial contribu- tions towards our understanding of the mode of toxin action (23). Generally, two approaches have been used. In the first, workers have at- tempted to correlate the toxin sensitivity of cell lines derived from various tissues and species with the histopathology of animal intoxication. In the second, a single cell line has been used
death in human diphtheria infection, Bonventre and Imhoff (1) examined heart cells in culture and found them to be unusually sensitive to the toxin. Finally, many of the insights leading to the currently accepted mechanism of action of diphtheria toxin (DE) were obtained using cultured HeLa cells (3, 6, 18, 24).
Our laboratory is engaged in an in-depth com- parative study of diphtheria exotoxin and
as a model to work out details of the toxin's exotoxin A (PE) from Pseudo~nonns oer~/ginosa mechanism of action. Probably the toxin most (12, 13). We began by coinparing the sensitivities successfully studied using these two approaches of many mammalian cell lines to both toxins is the exotoxin of Corynebacteriunz diphtheriae. (15). During the course of our work we found Solotorovsky and Gabliks (22) reported a close that some types of sera commonly used in cell correlation between the relative sensitivity of a culture could partially protect cells from the cell line to the toxin and the sensitivity of its cytotoxic actions of both toxins. We investigated animal donor. Moreover, following reports the nature of this artifact and this report that cardiac failure was a frequent cause of describes our findings. We believe that low
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

176 CAN. J . MICROBI OL. VOL. 23. 1977
levels of toxin-specific antibody in certain types of sera call account for the observed protection. Our work indicates that previous studies of either toxin using cell culture should be examined carefully. Moreover, we would like to raise the possibility that any studies of bacterial toxins in cell culture might encounter this problem.
Materials and Methods Toxit~s
Pserrrlor~~or~rrs oe~.rtgir~oso, strain PA103, was obtained from P. V. Liu. Pse~rrlo~not~ns aer~rgit~oscr exotoxin A was produced and purified by Dr. Stephen Leppla of this Institute using methods recently described (I I). The final product behaves as a single polypeptide of 66 000 daltons, contains no nleasurable carbohydrate, and is 295% pure as judged by sodium dodecyl sulfate poly- acrylanlide gel electrophoresis. Heating the purified toxin at 70 "C for I h conlpletely destroyed its cytotoxic activity. The mean lethal dose (LD,,) of purified PE administered intraperitoneally was 0.1 pg/ZOg CD-1 mouse (11). Purified C. rliplrtlreriae exotoxin (25 000 median lethal dose (MLD)/nll) (19) was the generous gift of Dr. A. M. Pappenheimer, Jr., Harvard University. Toxin concentra- tions were determined using extinction coefficients (EL%, cm) at 280 nm of 10.5 and 11.9 for PE and DE, respectively. PE-specific antisera, obtained by hyper- immunization of a pony using purified toxin, was pre- pared by Dr. Leppla.
Mcciio atirl Sera Trypsin, media, vitamins, antibiotics, and amino acids
were obtained from Grand Island Biological Company, Grand Island, NY. Calf and fetal calf sera were obtained from Grand Island Biological Company; Flow Labora- tories, Rockville, MD; Microbiological Associates, Bethesda, M D ; and Reheis Chemical Company, Phoenix, AZ. Human and horse sera were also obtained from Grand Island. All sera were heat-inactivated for 30 min at 56 "C before use in tissue culture.
Cells otrrl Cell Clrltlrre Mouse L-929 fibroblasts (CCL-I), human cervical
carcinoma (HeLa) cells (CCL-2), and monkey kidney (Vero) cells (CCL-81) were obtained from the American Type Culture Collection, Rockville, MD, and cultured as recommended by their registry. Primary chick fibro- blasts were prepared in the following manner. De- capitated 10-day-old embryos were washed in Puck's saline A (0.4 g/litre KCI, 8 g/litre NaCI, 0.35 g/litre NaHC03, 1 gllitre glucose), minced well, and agitated for 2min in 0.25z trypsin. The supernatant fraction was decanted, more trypsin was added (5 nll/enlbryo), and the solution was stirred for 30min at 37°C. The resulting supernatant fraction was filtered through sterile gauze and cells in the filtrate were washed twice with Eagle's mininlum essential medium containing 10% serum. The cells were counted and plated as indicated.
Ciret~~icals Tritiated thymidine (meth~l- [~H], specific activity 3 Ci/
mmol) and carrier-free, low pH iodine-125 were pur- chased from Schwarz/Mann, Orangeburg, NY, and New
England Nuclear, Boston, MA, respectively. Liquifluor liquid scintillation counting mixture was obtained from New England Nuclear and NCS tissue solubilizer from Amersham/Searle, Arlington Heights, IL. All other chemicals were reagent grade and were used without further purification.
Cytotoxic Assay Cells obtained as monolayers in Falcon T-75 tissue-
culture flasks were detached by trypsinization, counted with a Coulter counter model B, and resuspended at 5 x 10' cells/ml in medium 199 with Earle's salts con- taining 2.2 g/litre bicarbonate, 0.1 g/litre glutamine, 100 unitslml penicillin, 100 pg/ml streptomycin, and supplemented with 10% vol./vol. serum. Twenty-four well tissue culture plates (Linbro Scientific Corp., Ham- den, CT or Costar, Cambridge, MA) were seeded with 1 ml of cell suspension per well and the plates incubated at 37 "C for 24 h in a humidified 5% COz, 95% air at- mosphere. Toxins serially diluted in a sterile buffer (0.85% NaCI, 10 m M sodium phosphate, pH 7.1) were added in 0.1-ml volumes to the wells; the cells were incubated an additional 48 h. The monolayers were then washed twice with warm (37 "C) Hanks' balanced salt solution (HBSS) to remove detached cells and the remaining cells solubilized in 0.5 ml of 0.1 M NaOH. An aliquot from each well was removed for protein assay by the method of Lowry et a/. (14) using a bovine serum albumin standard (Miles Laboratories, Kankakee, IL). Each symbol in the figures is the average of triplicate samples; standard errors were usually 5-10%.
Cell~tlar T\rymin'inc Incorporntiot~ Cells were treated exactly as in the cytotoxic assay
above until harvest. At this point, each sample was first incubated with [3H]thymidine (2 pCi/well), then washed three times with HBSS, and further incubated with a solution of 0.05% trypsin and 0.02z ethylenediamine- tetraacetic acid (EDTA) in modified Puck's saline A (0.5 ml/well). The contents of each well were then trans- ferred to centrifuge tubes; the trichloroacetic acid (TCA) precipitable fraction was obtained by addition of an equal volume of 10% TCA. Precipitates were then washed twice with 5% TCA and once with methanol. The final pellets were solubilized with NCS and the entire sample assayed for radioactivity using a toluene-Liquifluor scintillator and a Mark I1 liquid scintillation spectrometer (Nuclear Chicago).
Ioditrotiot~ Procef/rrre Iodination of the toxins was performed by the method
of David (4) as more recently described by Bonventre ct ol. (2). Our pro-edure differed from that of Bonventre et ol. in that centrifugation was used to remove the Sepharose beads and dialysis to remove the free Na['ZSIl.
Assoy for Atllibociy Sera were tested for antibody by the method of Farr
(5). [L2'I]toxin was incubated with medium or medium plus 10% serum for 1 h at 37 "C followed by the addition of saturated (NH,),SO, sufficient to bring the mixture to 40% saturation. After 21 h at 4 "C the solution was centrifuged and an aliquot of the supernatant fraction assayed for radioactivity in a Nuclear Chicago 1185 series gamma-counting system. The pellet was washed once with 40% (NH,),SO, and assayed for radioactivity.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.
MIDDLEBROOK AND DORLAND
TOXIN CONCENTRATION (ng /ml )
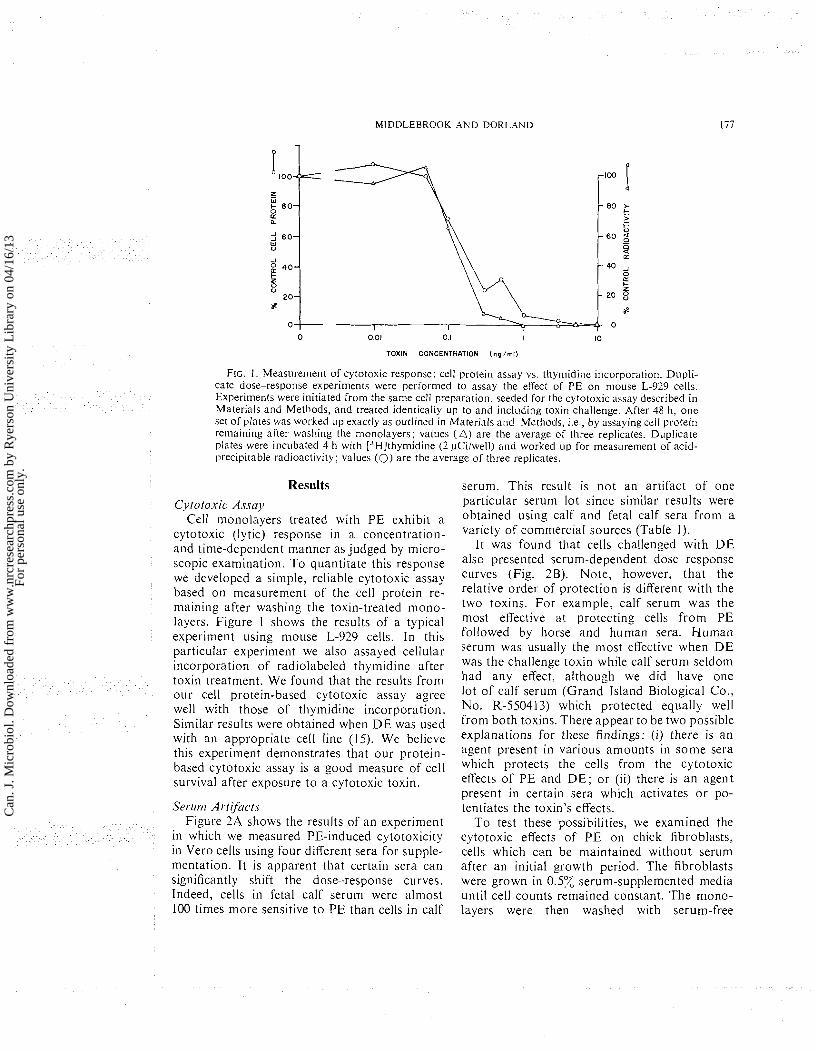
FIG. I. Measurement of cytotoxic response: cell protein assay vs, thymidine incorporation. Dupli- cate dose-response experiments were performed to assay the effect of PE on mouse L-929 cells. Experiments were initiated from the same cell preparation, seeded for the cytotoxic assay described in Materials and Methods, and treated identically up to and inclilding toxin challenge. After 48 h, one set of plates was worked up exactly as outlined in Material5 and Methods, 1.e , by assaying cell protein renla~ning after washing the monolayers; values ( A ) are the average of three replicates. Duplicate plates were incubated 4 h with [3H]thymidine (2 pCi/well) and worked up for measurement of acid- precipitable radioactivity; values (0) are the average of three replicates.
Results
Cyfotoxic Assay Cell monolayers treated with PE exhibit a
cytotox~c (ly tic) response in a concentratlon- and time-dependent manner as judged by micro- scopic examination. To quantitate this response we developed a simple, rel~able cytotoxic assay based on measurement of the cell protein re- maining after washing the toxin-treated mono- layers. Figure 1 shows the results of a typical experiment using mouse L-929 cells. In this particular experiment we also assayed cellular incorporation of radiolabeled thymidine after toxin treatment. We found that the results from our cell protein-based cytotoxic assay agree well with those of thymidine incorporation. Similar results were obtained when D E was used with an appropriate cell line (15). We believe this experiment demonstrates that our protein- based cytotoxic assay is a good measure of cell survival after exposure to a cytotoxic toxin.
Serlit?? Art i f i c f ~ Figure 2A shows the results of an experiment
in which we measured PE-induced cytotoxicity in Vero cells using four different sera for supple- mentation. It is apparent that certain sera can significantly shift the dose-response curves. Indeed, cells in fetal calf serum were almost 100 times more sensitive to PE than cells in calf
serum. This result is not an artifact of one articular serum lot since similar results were obtained using calf and fetal calf sera from a variety of commercial sources (Table 1).
It was found that cells challenged with D E also presented serum-dependent dose-response curves (Fig. 2B). Note, however, that the relative order of orotection is different with the two toxins. For example, calf serum was the most effective at protecting cells from PE followed bv horse and human sera. Human serum was usually the most effective when D E was the challenge toxin while calf serum seldom had any effect, although we did have one lot of calf serum (Grand Island Biological Co., No. R-550413) which protected equally well from both toxins. There appear to be two possible explanations for these findings: (i) there is an agent present in various amounts in some sera which protects the cells from the cytotoxic effects of PE and D E ; or (ii) there is a n agent present in certain sera which activates or po- tentiates the toxin's effects.
To test these possibilities, we examined the cytotoxic effects of PE on chick fibroblasts, cells which can be maintained without serum after an initial growth period. The fibroblasts were grown in 0 .5x serum-supplemented media until cell counts remained constant. The mono- layers were then washed with serum-free
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

178 C A N . J . MICRO
0 0.1 I 10 100 1000 TOXIN tng/ml)
FIG. 2. EEect of different sera on cell response to PE or DE. A. Aliquots of a concentrated Vero cell suspension were diluted to 5 x 10" cells/ml in medium 199 supple- mented with 10% vol./vol. of the serum indicated. A dose- response experiment was performed on each preparation using PE and our standard cytotoxic assay. Each point is the average of quadruplicates. B. HeLa cells and DE were used in the same protocol as part A. Serunl lots were not the same as in A. Symbols: (@), fetal calf serum; (A), human serum; (H), horse serum; (O), calf serum.
TABLE I . Tissue culture LDso of PE in L-929 cells cultured with calf or fetal calf sera from
different sourcesa
TCLDso, Source Lot No. ~ g / n l l
Fetal calf sera Reheis M 1 6703 0 .3 GIBCO R.550324 0 .2 GIBCO A851218 0 . 3 Flow 4055759 0 . 2 M.A. 8671 3 0 . 3
Calf sera GIBCO E054013 30 GIBCO A754019 100 Flow 402038 1 20 M.A. 82539 100
oReplicate cell suspensions were plated using the serum indicated as a supplement. After 24 h cells were challenged with toxin and a cytotoxicity assay was per- formed (see Materials and Methods). Tissue culture LDs0 was defined as that concentration of toxin which produced 50Z ofthe total change in cell protein. Sources are as follows: Reheis, Reheis Chemical Co.; GIBCO, Grand Island Biological Co.; Flow. Flow Laboratories; M.A., Microbiological Associates.
BIOL. VOL. 23. 1977
medium and further incubated in either serum- free medium or medium plus 10% serum. Cells from both these treatments were challenged with PE in the usual manner. The results (Fig. 3) demonstrate that cells exhibited essentially the same sensitivity to PE whether they were chal- lenged in fetal calf serum-supplemented or serum-free media. Cells were about 100 times less sensitive to PE when challenged in calf serum-supplemented medium. We believe the results of this experiment are consistent with the notion that there is some agent in calf serum (and presumably horse and human sera, as well) which can protect cells from the cyto- toxic effects of PE. Apparently fetal calf serum does not contain this agent and should be used for all cell culture studies of PE.
The experiment depicted in Fig. 3 demon- strates another interesting point about the effects of PE on cells. Under the conditions of o u r experiment, cells in 10x fetal calf serum were growing at a rapid rate during the toxin challenge while cells in serum-free medium were in a resting state. Since their dose-response curves are similar, it appears that resting and rapidly growing cells .are equally sensitive to PE's cytotoxic effect.
Tiine Poir~t of Ser~~nl Protection Figure 4 shows the results of an experiment
designed to determine whether the PE protection afforded by calf serum occurs before, during, or after challenge of the cells with toxin. The results indicate that it is only during the t ime of toxin challenge that protection occurred. Cells originally grown in either calf o r fetal calf serum were afforded n o detectable protec- tion if subsequently challenged in fetal calf serum. However, both were protected if chal- lenge took place in the presence of calf serum. By the same token, postchallenge incubation of cells with calf serum did not seem to provide any protection compared t o postchallenge in- cubation with fetal calf serum. These results appear to rule out the possibilities (i) that the cells may incorporate a protective factor from calf serum before challenge, which reduces the toxin's potency or (ii) that there is a nutritional or other factor in calf serum capable of re- ducing or reversing the toxin's effects after challenge.
Assay for Ailtibody We tested calf and fetal calf sera for the
presence of antibody using a method originally
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

MIDDLEBROOK AND DORLAND
TOXIN CONCENTRATION (ng/ml)
FIG. 3 . Effect of sera on the response of chick fibroblasts to PE. Replicate cell suspensions in Eagle's minimal essential medium containing either 0.5% calf or fetal calf serum were seeded into niultiwell tissue culture plates. After 6 days of incubation, cell counts tripled. No further increase was observed during the next 2 days. On day 8 monolayers were washed twice with serum-free media and refed with either serum-free or 10% honiologous serum-supplenlented media. PE was added to the concentration indicated and the usual 48-h cytotoxicity assay was performed. After 24 h cell numbers in 10% serum had doubled and those in ~~nsupplemented medium had declined slightly. Symbols: (D), cells originally in fetal calf, challenged in serum-free media; (O), cells originally in fetal calf, challenged in 10% fetal calf; (O), cells originally in calf, challenged in serum-free media; (A), cells originally in calf, challenged in 10% calf.
I 10 100 1000
TOXIN CONCENTRATION (ng / rn l )
F IG . 4. Time point of serum protection. Replicate L-929 cell suspensions were seeded and grown for 24 h in medium 199 plus 10% calf o r fetal calf serum. Monolayers were washed and refed with m e d i ~ ~ n i plus 10% homologous o r heterologous serum, and PE added. After 2 h, cells were washed three times with ~~nsupplemented medium followed by the addition of medium with 10% serum. After 48 h of fur- ther incubation, surviving cells were measured by our cytotoxicity methodology. Symbols: (A), cells seeded in fetal calf, challenged in calf, and postincubated in fetal calf serum; (O), cells seeded in fetal calf, challenged in fetal calf, and post-incubated in fetal calf serum; ( x ) , cells seeded in fetal calf, challenged in fetal calf, and postincubated in calf serum; (O), cells seeded in calf, challenged in calf, and postinc~~bated in fetal calf serum; (n), cells seeded in calf, challenged in fetal calf, and post- incubated in fetal calf serum.
developed by Farr (5) and more recently used to significant levels of toxin. The results of a detect antibody to bacterial toxins (8). Briefly, typical experiment are shown in Fig. 5. I t was this method involves incubating the suspect found that only low levels of radioactivity serum with radiolabeled toxin followed by the precipitated in both the control and the 10% addition of (NH,),SO, at a concentration fetal calf serum assays. On the other hand, the sufficient to precipitate antibody (and antibody- (NH,),SO, precipitates in 10% calf serum con- toxin complexes) but insufficient to bring down tained substantial levels of radioactivity. More-
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

CAN. J . MICROBIOL. VOL. 23. 1977
0 1 10 100
TOXIN CONCENTRATION (nq / rn l )
F I G . 5. Assay for PE-specific antibody ~n calf and fetal calf sera. Radiolabeled P E was added t o medium 199 containing the sera indicated and inc~ibated at 37 "C for 1 h. The 100 ng/mi samples initially contained a total of 1.6 x lo3 cpm and the lower concentrations proportionally less. Samples were made 4 0 z vol./vol. in (NH,)2S0, and placed a t 4 C for 21 h. Each sample was then centrifuged, the resulting pellet washed once with 4 0 z (NH,)2S0, , and assayed for radioactivity. Symbols: (O), unsupplemented media; (O), media + 10% fetal calf serum; (A), media + 1 0 z calf serum; ( x ), media + a small volume of pony PE-specific antisera.
over, when a small amount of PE-specific extent, and support the view that toxin-specific antiserum was assayed as a positive control, a antibody is the only significant PE-protective result similar to that of 10% calf serum was agent in calf sera. obtained. This experiment demonstrates the like- lihood that toxin-specific antibody in calf sera Discussion is responsible for at least a portion of its protec- tive effect on cells challenged with PE. In this coinmunication we have described a
method which can be used to measure the Protective Effect of Toxitl-specific Antiserrr
The results of the experiment in Fig. 5 demon- strate the probable presence of toxin-specific antibody in calf serum. One remaining question is whether this agent alone can adequately ex- plain the PE-protective effect of calf serum. T o examine this question we performed the ex- periment shown in Fig. 6. Particular lots of fetal calf and calf sera and PE-specific pony antisera were titrated for the presence of anti- body by a standard hemagglutination (HA) assay (7) (see insert, Fig. 6). Toxin dose- response curves were then obtained for cells cultured in 10% fetal calf serum, 10% calf serum, and 10% fetal calf serum plus a (negligible) volume of pony serum calculated by H A assay to contain the same level of antibody as the 1 0 z calf serum. The results indicate that levels of PE-specific antiserum equal to those titrated in calf serum protected cells to about the same
cytotoxic response of cells to bacterial toxins (or other cytotoxic agents). W e believe that this system affords a number of advantages over previously described methods (9, 10, 2 1, 22). Our ultimate measure was a protein assay which most laboratories are equipped to do and which is automated in many. Moehring and Moehring (16) recently used a similar system to measure the effect of diphtheria toxin on cellular in- corporation of radiolabeled amino acids. How- ever, we emphasize the versatility of this system. We have measured the effects of bacterial toxins on many cellular processes including uptake and incorporation of deoxyribonucleic acid (DNA), ribonucleic acid (RNA), or protein precursors, ion transport, uptake of sugars, and uptake of radiolabeled toxin. All of these studies can be done using cells in culture tubes (22), but our system allows the convenience of handling 24 samples simultaneously. The micro-
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

MIDDLEBROOK A N D DORLAND
FETAL C A I S E W 0
TOXIN CONCENTRATION (ng/ml)
FIG. 6 . Comparison of cell protect~on sing calf serum or PE-spec~fic antiserum. Replicate L-929 cell s~~spens ions were seeded in multiwell tissue c u l t ~ ~ r e plates ~lsing medium 199 plus 10% calf serum (A) ; 10% fetal calf serum (0); or 1 0 z fetal calf serum plus a volume (negl~gible) of PE-spec~fic antisera (0). A standard cytotoxic~ty assay was carried out on all three ci~ltures. Hemagglutination t~trat ions were performed as prev~ously described (7).
titer system with a multiple-harvesting apparatus also offers a means of rapidly processing many samples (10). However, we found that the micro- titer system presented technical difficulties in washing out toxin or changing medium for kinetic studies. These difficulties do not exist in our system.
An important finding is that the type of serum used for cell culture can significantly influence the results of cytotox~city studies using Pseurlo- 1not7us or diphtheria exotoxins. Cells cultured and toxin-challenged in various sera yielded dose-response curves varying over two orders of magnitude (Fig. 2A and B). By comparing the effects of PE on cells which could be main- tained with or without serum, we determined that some types of sera probably contained a protective agent. Of the sera examined, calf serum appeared to contain the most and fetal calf serum the least of this agent. We therefore carried out a comparat~ve study of these two sera. We found that the PE protection afforded by calf serum apparently did not result from the cellular incorporation of an agent before toxin challenge, nor from an agent capable of re- versing toxin effects after challenge (Fig. 4). Only when calf serum was present during PE challenge were cells protected. Since this finding suggested the possible presence of antibody, we assayed the two sera using a method recently shown (8) to be very sens~tive for bacterial
antitoxins. The results of this experiment (Fig. 5) indicated the probable presence of PE-specific antibody in calf serum while none could be detected in fetal calf serum. Another standard assay (hemagglutination) yielded the same re- sults (Fig. 6). Finally, using PE-specific hyper- immune serum we demonstrated that the level of HA antibody-like material in calf serum was sufficient to account entirely for its PE-protective effect. We have not performed similar experi- ments with DE but we believe it reasonable to speculate that a similar situation probably exists for tissue-culture studies with this toxin.
Although all the above data are consistent with the notion that sera other than fetal calf may contain various levels of antibody to PE, DE, or both, it is not our intent to unequivocally prove this point. It is our contention, however, that the use of certain sera for cell culture studies of PE or DE may lead to artifactual results. This finding is more than just of academic interest. Although it is unlikely that any con- clusions arising from mechanistic studies would be affected, we believe that studies comparing the sensitivities of different cell lines to these two toxins should be reexamined in light of our results. For example, Pavlovskis and Gordon have reported that Vero cells were more sensitive to PE than were LLC-MK, cells (20). We came to the opposite conclusion (see companion paper (15)) and believe that the explanation for this
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.

182 CAN. J . MlCROBl OL. VOL. 23. 1977
contradiction is that Pavlovskis and Gordon cultured Vero cells with fetal calf serum and LLC-MK, with calf serum, whereas we used fetal calf serum for both.
We suggest that all future studies concerning the effects of bacterial toxins on cells in culture be performed with fetal calf serum. Although we have no experiments to demonstrate the point here, we believe it possible that antibodies to other toxins might also be present in various sera. Since antibodies do not normally cross the bovine placenta (17), fetal calf sera would offer the best potentially antitoxin-free source of serum. However, since an in utero infection would be possible, it is best to test even fetal calf serum before beginning detailed toxin-cell studies.
Acknowledgments The authors express their appreciation to
John Kondig for supplying the cells used in this work, to Mary Jennings and Karen Bostian for performing the protein assays, and to Dennis Leatherman for his excellent technical assistance. We would also like to acknowledge many helpful comments and suggestions from Dr. Stephen Leppla.
1. BONVENTRE, P. F., and J. G. IMHOFF. 1967. Studies on the mode of action of diphtheria toxin. 11. Protein synthesis in primary heart cell cultures. J. Exp. Med. 126: 1079- 1086.
2. BONVENTRE. P. F . . C . B. S A E L I N G E R . B. IVINS, C . WOSCINSKI, and M. A M O K I N I . 1975. Interaction of cultured mammalian cells with [1251]diphthe~ia toxin. Infect. Immun. 11: 675-684.
3. COLLIER, R. J. 1975. Diphtheria toxin: mode ofaction and structure. Bacteriol. Rev. 39: 54-85.
4. DAVID, G. S. 1972. Solid state lactoperoxidase: a highly stable enzyme for simple, gentle iodination of proteins. Biochem. Biophys. Res. Commun. 48: 464-47 1.
5. FARR, R. S. 1958. A quantitative immunochemical measure of the primary interaction between I*BSA andantibody. J. Infect. Dis. 103: 239-262.
6. GILL, D. M., A. M. PAPPENHEIMER, JR.. and T . UCHIDA. 1973. Diphtheria toxin, protein synthesis. and the cell. Fed. Proc. 32: 1508-1515.
7. GORDON, J . , B. ROSE, and A. H . SEHON. 1958. Detec- tion of "non-precipitating" antibodies in sera of indi- viduals allergic to ragweed pollen by an in vitro method. J. Exp. Med. 108: 37-51.
8. GRUBER. J . , and G. G . WRIGHT. 1969. Ammonium sulfate coprecipit'ation antibody determination with
purified staphylococcal enterotoxins. J. Bacteriol. 99: 18-24.
9. KEUSCH, G. T. , M. JACEWICZ, and S. 2. HIRSCHMAN. 1972. Quantitative microassay of cell culture for en- terotoxin ofS11igelln (Ixserltericre 1. J. Infect. Dis. 125: 539-54 1 .
10. KNUDSEN. R . C.. L. T . C A L L A H A N , 111. A. AHMED. and K . W. SELL. 1974. Use of microculture plates and the multiple automated sample harvester for in vitro microassay of bacterial toxins. Appl. Microbiol. 28: 326327.
11. LEPPLA. S . H . 1976. Large-scale purification a n d characterization of the exotoxin of P s e r r d o t ~ ~ o ~ ~ t r s oerirgitlostr. Infect. Immunol. 14: 1077-1086.
12. L I U , P. V. 1966. The roles of v a ~ i o u s fractions of P.serrdor?7ot1ns trrrrrgirlostr in its pathogenesis. 11. EE fects of lecithinase and protease. J . Infect. Dis. 116: 112-1 16.
13. L I U , P. V. 1966. The roles of various fractions of Pserrdot?~ontrs oer?igirlo~cr in its pathogenesis. 111. Identity of the lethal toxins produced ill i,it~.o and i r ~ 1.i1.o. J . Infect. Dis. 116: 481-489.
14. LOWRY, 0. H. , N. J. ROSEBROUGH, L. A. FARR. and R. J. RANDALL. 1951. Protein measurementwith Folin phenol reagent. J . Biol. Chem. 193: 265-275.
15. MIDDLEBROOK. J . L.. and R . B. DORLAND. 1977. Re- sponse of cultured mammalian cells to the exotoxins of P.ser/do~?ro~lrrs c1errrgi11osci and Co~ytrebocte~~irrm d i ~ ~ l ~ t h o i n e : differential cytotoxicity. Can. J . Micro- biol. 23. This issue.
16. MOEHRING, J . M., and T . J . MOEHRING. 1976. Com- parison of diphtheria intoxication in human and non- human cell line's and their resistant variants. Infect. Immun. 13: 221-228.
17. OSBURN, B. I. 1973. Immune responsiveness of the fetus and neonate. J . Am. Vet. Med. Assoc. 163: 80 1-803.
18. PAPPENHEIMER. A. M.. JR. , and D. M. G I L L . 1973. Diphtheria. Science. 182: 353-358.
19. PAPPENHEIMER, A. M., JR. . T . UCHIDA, and A. A. HARPER. 1972. An immunological study of the diph- theria toxin molecule. Immunochemistry. 9: 891-906.
20. PAVLOVSK~S, 0 . R., and F . B. GORDON. 1972. Pseridomorlc~,s cro.rrgirloscr exotoxin: effect on cell cul- tures. J. Infect. Dis. 125: 631-636.
21. SCHAEFFER, W. I.. J. GABLIKS, and R. C A L ~ T ~ S . 1966. Interaction of staphylococcal enterotoxin B with cell cultures of human embryonic intestine. J. Bacteriol. 91: 21-26.
22. SOLOTOROVSKY, M., and J . GABLIKS. 1963. T h e biological action of diphthel-ia toxin in cell and tissue culture. 111 Cell culture in the study of bacterial dis- ease. Edited by M . Solotorovsky. Rutgers University Press, New Blunswick, NJ. pp. 5-15.
23. SOLOTOROVSKY. M., and W. JOHNSON. 1970. Tissue culture and bacterial protein toxins. 111 Microbial tox- ins, vol. I . Edited By S. J. Ajl. S. Kadis, and T . C . Montie. Academic Press, New York. pp. 277-327.
24. STKAUSS, N., and E. D. HENDEE. 1959. The effect of diphtheria toxin on the metabolism of HeLa cells. J . Exp. Med. 109: 145-163.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Rye
rson
Uni
vers
ity L
ibra
ry o
n 04
/16/
13Fo
r pe
rson
al u
se o
nly.