
Sequence analysis and evolution of group B rotaviruses
7
Virus Research 125 (2007) 219–225 Sequence analysis and evolution of group B rotaviruses Mustafizur Rahman a,b,∗ , Zahid M. Hassan a , Hasan Zafrul a , Farjana Saiada a , Sukalyani Banik a , Abu S.G. Faruque a , Thomas Delbeke b , Jelle Matthijnssens b , Marc Van Ranst b , Tasnim Azim a a Laboratory of Virology, ICDDR,B, GPO Box-128, Dhaka 1000, Bangladesh b Laboratory of Clinical and Epidemiological Virology, Rega Institute for Medical Research, University of Leuven, Minderbroedersstraat 10, B-3000 Leuven, Belgium. Received 29 July 2006; received in revised form 10 January 2007; accepted 12 January 2007 Available online 12 February 2007 Abstract Human group B rotaviruses were isolated from hospitalized patients in Bangladesh between July 2003 and December 2004. Phylogenetic analyses of the gene segments encoding the hemagglutinin (VP4), glycoprotein (VP7) and RNA-binding protein (NSP2) of group B rotaviruses showed that Bangladeshi strains were more similar to the Indian strains than to the prototype Chinese strains. Moreover, all human strains were clustered together and were distantly related to the animal strains. With limited sequence data, the evolutionary rate of the glycoproteins (VP7) of human group B rotaviruses was estimated to be 1.57 × 10 −3 nucleotide substitutions/(site year), which was comparable to other rapidly evolving RNA viruses. The most recent common ancestor (MRCA) of the extant human group B rotaviruses was calculated to date to around 1976. © 2007 Elsevier B.V. All rights reserved. Keywords: Group B rotavirus; VP4; VP7; NSP2; Evolution; Phylogeny 1. Introduction Rotaviruses are a major causative agent of severe diarrhea in humans and animals. They are classified into seven antigeni- cally distinct groups (A–G) on the basis of a common group antigen VP6 (Estes, 2001). Groups A–C infect humans and animals but groups D–G infect animals only (Kapikian et al., 2001). Group B rotaviruses are called adult diarrhea rotavirus (ADRV) because they mostly infect adults and older children (Hung et al., 1983; Mackow et al., 1993). The human group ∗ Corresponding author at: Virology Laboratory, Laboratory Sciences Divi- sion, ICDDR,B, GPO Box-128, Dhaka 1000, Bangladesh. Tel.: +880 2 8860523–32x2409; fax: +880 2 881 2529. E-mail addresses: mustafi[email protected] (M. Rahman), [email protected] (Z.M. Hassan), [email protected] (H. Zafrul), [email protected] (F. Saiada), sukalyani [email protected] (S. Banik), [email protected] (A.S.G. Faruque), [email protected] (T. Delbeke), [email protected] (J. Matthijnssens), [email protected] (M. Van Ranst), [email protected] (T. Azim). B rotaviruses were first detected in China in 1982 and since then they have been causing large waterborne epidemics infect- ing thousands of people there (Chen et al., 1985; Hung et al., 1984; Fang et al., 1989; Yang et al., 1998). For many years they were restricted to China, but recently they have been isolated sporadically in India (since 1998) and Bangladesh (since 2001) (Ahmed et al., 2001; Sanekata et al., 2003; Kelkar et al., 2004; Kobayashi et al., 2001). The animal group B rotaviruses were isolated from lambs (United Kingdom and the United States), pigs (United States and Japan), cows (the United States, Japan and India), goat (China) and rats (the United States) (Chasey and Banks, 1984; Theil et al., 1985, 1995; Parwani et al., 1996; Sanekata et al., 1996; Barman et al., 2004; Tsunemitsu et al., 1999; Eiden et al., 1991). Interestingly, like human group B strains, the animal group B rotaviruses were mostly isolated from adult animals. The scientific mystery surround- ing the reason why they infect mainly adults remains to be addressed. The importance of the viruses in causing diarrhea in humans is poorly understood because no diagnostic method for detecting group B rotaviruses is commercially available at this moment. 0168-1702/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.virusres.2007.01.008
-
Upload
mustafizur-rahman -
Category
Documents
-
view
222 -
download
3
Transcript of Sequence analysis and evolution of group B rotaviruses

A
asw(e1©
K
1
icaa2((
sT
zfg(m(
0d
Virus Research 125 (2007) 219–225
Sequence analysis and evolution of group B rotaviruses
Mustafizur Rahman a,b,∗, Zahid M. Hassan a, Hasan Zafrul a, Farjana Saiada a,Sukalyani Banik a, Abu S.G. Faruque a, Thomas Delbeke b,
Jelle Matthijnssens b, Marc Van Ranst b, Tasnim Azim a
a Laboratory of Virology, ICDDR,B, GPO Box-128, Dhaka 1000, Bangladeshb Laboratory of Clinical and Epidemiological Virology, Rega Institute for Medical Research, University of Leuven,
Minderbroedersstraat 10, B-3000 Leuven, Belgium.
Received 29 July 2006; received in revised form 10 January 2007; accepted 12 January 2007Available online 12 February 2007
bstract
Human group B rotaviruses were isolated from hospitalized patients in Bangladesh between July 2003 and December 2004. Phylogeneticnalyses of the gene segments encoding the hemagglutinin (VP4), glycoprotein (VP7) and RNA-binding protein (NSP2) of group B rotaviruseshowed that Bangladeshi strains were more similar to the Indian strains than to the prototype Chinese strains. Moreover, all human strainsere clustered together and were distantly related to the animal strains. With limited sequence data, the evolutionary rate of the glycoproteins
VP7) of human group B rotaviruses was estimated to be 1.57 × 10−3 nucleotide substitutions/(site year), which was comparable to other rapidlyvolving RNA viruses. The most recent common ancestor (MRCA) of the extant human group B rotaviruses was calculated to date to around976.
2007 Elsevier B.V. All rights reserved.
Bti1ws(
eywords: Group B rotavirus; VP4; VP7; NSP2; Evolution; Phylogeny
. Introduction
Rotaviruses are a major causative agent of severe diarrhean humans and animals. They are classified into seven antigeni-ally distinct groups (A–G) on the basis of a common groupntigen VP6 (Estes, 2001). Groups A–C infect humans andnimals but groups D–G infect animals only (Kapikian et al.,
001). Group B rotaviruses are called adult diarrhea rotavirusADRV) because they mostly infect adults and older childrenHung et al., 1983; Mackow et al., 1993). The human group∗ Corresponding author at: Virology Laboratory, Laboratory Sciences Divi-ion, ICDDR,B, GPO Box-128, Dhaka 1000, Bangladesh.el.: +880 2 8860523–32x2409; fax: +880 2 881 2529.
E-mail addresses: [email protected] (M. Rahman),[email protected] (Z.M. Hassan), [email protected] (H. Zafrul),[email protected] (F. Saiada), sukalyani [email protected] (S. Banik),[email protected] (A.S.G. Faruque), [email protected]. Delbeke), [email protected] (J. Matthijnssens),[email protected] (M. Van Ranst), [email protected]
T. Azim).
Kipaa1egiia
ig
168-1702/$ – see front matter © 2007 Elsevier B.V. All rights reserved.oi:10.1016/j.virusres.2007.01.008
rotaviruses were first detected in China in 1982 and sincehen they have been causing large waterborne epidemics infect-ng thousands of people there (Chen et al., 1985; Hung et al.,984; Fang et al., 1989; Yang et al., 1998). For many years theyere restricted to China, but recently they have been isolated
poradically in India (since 1998) and Bangladesh (since 2001)Ahmed et al., 2001; Sanekata et al., 2003; Kelkar et al., 2004;obayashi et al., 2001). The animal group B rotaviruses were
solated from lambs (United Kingdom and the United States),igs (United States and Japan), cows (the United States, Japannd India), goat (China) and rats (the United States) (Chaseynd Banks, 1984; Theil et al., 1985, 1995; Parwani et al.,996; Sanekata et al., 1996; Barman et al., 2004; Tsunemitsut al., 1999; Eiden et al., 1991). Interestingly, like humanroup B strains, the animal group B rotaviruses were mostlysolated from adult animals. The scientific mystery surround-ng the reason why they infect mainly adults remains to be
ddressed.The importance of the viruses in causing diarrhea in humanss poorly understood because no diagnostic method for detectingroup B rotaviruses is commercially available at this moment.

2 Resea
NAKt11
dsrR(tda
rtBmgtwef
2
2
rio2ham2t2nSVpBis
2
paitt
2
QTtRgT3wf37elp
2
ptwRCTTn3TACwge
2
CsMlfHTaCme
2
20 M. Rahman et al. / Virus
onetheless, few sero-prevalence studies in China, Hong Kong,ustralia, the United States, Canada, Kenya, and the Unitedingdom suggest that they are present in low proportion of
he global population (Penaranda et al., 1989; Nakata et al.,987; Krishnan et al., 1999; Mackow, 1995; Ushijima et al.,992).
Group B rotaviruses belong to the Reoviridae and have aouble stranded RNA genome made up of 11 independent geneegments. They can be differentiated from groups A and Cotaviruses by tracking the migration pattern of their segmentedNA (4-2-1-1-1-1-1) on polyacrylamide gel electrophoresis
PAGE) (Saif and Jiang, 1994). No serotyping/genotyping sys-em has been described for group B rotaviruses due to theifficulties in adapting them in cell culture and availability ofdequate amount of gene sequence data.
During July 2003–December 2004 we detected group Botaviruses in diarrhea patients attended the Dhaka hospi-al of International Centre for Diarrhoeal Disease Research,angladesh (ICDDR,B). We partially sequenced the gene seg-ents encoding VP4, VP7 and NSP2 proteins of Bangladeshi
roup B rotaviruses to investigate their genetic relationships withhe cognate genes of other group B rotavirus strains circulatingorldwide. The molecular dating and evolutionary rates were
stimated only for the VP7 genes since adequate sequence dataor other gene segments were unavailable.
. Materials and methods
.1. Study population and data collection
The Dhaka hospital of ICDDR,B treats over 100,000 diar-hea patients each year and includes every fiftieth (2%) patientn the hospital surveillance system to determine the presencef various common enteric pathogens. During July–December003, a total of 856 stool specimens which were included in theospital surveillance system were tested for group A rotavirusntigen and 245 (28.6%) were positive. The negative speci-ens (n = 611) were tested for group B rotaviruses. In contrast,
188 stool specimens from hospital surveillance system wereested for various enteric pathogens during January–December004, which included group A rotaviruses, Salmonella typhi,on-typhi Salmonella, Shigella dysenteriae, Shigella flexneri,higella boydii, Shigella sonnei, Vibrio cholerae O139, andibrio cholerae O1. No pathogen was detected in 1050 sam-les from which 10% samples (n = 100) were tested for group
rotaviruses. The clinical data of the patients includedn the study were collected from the hospital surveillanceystem.
.2. Electropherotyping
One hundred microliters of 10% stool suspensions inhosphate buffered saline were treated with sodium acetate
nd extracted with an equal volume of phenol:chloroform:soamylalcohol (25:24:1) mixture. The extracted RNA wasested for electropherotype (E-type) by polyacrylamide gel elec-rophoresis (PAGE) as described by Herring et al. (1982).Vd
rch 125 (2007) 219–225
.3. Reverse transcription (RT)-PCR
RNA was extracted from the stool suspension using theIAamp Viral RNA mini kit (Qiagen/Westburg, Leusden,he Netherlands) according to the manufacturer’s instruc-
ions. RT-PCR was carried out using the Qiagen OneStepT-PCR Kit (Qiagen/Westburg). The primers specific for theroup B rotavirus NSP2 gene (Bl, 5′-CTATTCAGTGTGTCG-GAGAGG-3′; and B4, 5′-CGTGGCTTTGGAAAATTCTTG-′) were used as described by Gouvea et al. (1991). The reactionas carried out with an initial reverse transcription step at 45 ◦C
or 30 min, followed by 40 cycles of amplification (30 s at 94 ◦C,0 s at 50 ◦C, 1 min at 72 ◦C), and a final extension of 7 min at2 ◦C in a thermal cycler. PCR products were run on a 1.5%thidium bromide stained agarose gel and visualized under UV-ight. Specific segment size (489 bp) for group B rotavirus DNAroduct was observed on stained gel.
.4. Nucleotide sequencing
The PCR products were purified with the QIA quick PCRurification kit (Qiagen/Westburg), and sequenced in both direc-ions using the dideoxy-nucleotide chain termination methodith the ABI PRISM® BigDye Terminator Cycle Sequencingeaction kit (Perkin-Elmer Applied Biosystems, Foster City,alifornia) on an automated sequencer (ABI PRISMTM 310).he consensus primers GrB VP7 25F (5′-CTTCTCGTCCT-GCTGCTG-3′; strain Bang373, accession number AY238385,t 25–43), GrB VP7 814R (5′-GGGTTTTTACAGCTTCGGC-′; strain Ban373, nt 796–814), GrB VP4 13F (5′-GCTATG-TGACGTATTTACG-3′; strain Bang373, accession numberY238388, nt 13–32), and GrB VP4 1178R (5′-GTATAA-CAGAAGCGTC CAC-3′; strain Bang373, nt 1159–1178)ere used for amplifying and sequencing of the VP7 and VP4ene segments. The NSP2 primers used in the RT-PCR weremployed for NSP2 gene sequencing.
.5. DNA and protein sequence analysis
The chromatogram sequencing files were inspected usinghromas 2.23 (Technelysium, Queensland, Australia), and con-
ensus sequences were prepared using SeqMan II (DNASTAR,adison, WI). Nucleotide and amino acid sequence simi-
arity searches were performed using the National Centeror Biotechnology Information (NCBI, National Institutes ofealth, Bethesda, MD) BLAST (Basic Local Alignment Searchool) server on GenBank database, release 153.0 (Altschul etl., 1990). Multiple sequence alignments were calculated usingLUSTALX 1.81 (Thompson et al., 1997). Sequences wereanually edited in the GeneDoc Version 2.6.002 alignment
ditor (Nicholas et al., 1997).
.6. Phylogenetic analysis
Phylogenetic analyses were conducted using the MEGAersion 2.1 software package (Kumar et al., 2004). The den-rograms were constructed using the neighbor-joining method.

Research 125 (2007) 219–225 221
Gm
2
PUafoslws
2
mIa
3
3
fJScm(bssifs
fcrR
3p
ftt4rpp
Fig. 1. Polyacrylamide gel electrophoresis (PAGE) showing the characteristicgBi
T
M. Rahman et al. / Virus
enetic distances were calculated using the Kimura-2 parameterodel.
.7. Estimation of evolutionary rate
Evolutionary rate and dating were calculated by usingath-O-Gen software (provided by Andrew Rambaut, Oxfordniversity, UK). This program takes a phylogenetic tree as input
nd performs a linear regression between the genetic distancerom the root and the sampling time for each sequence. The rootf the tree was assigned on the long-branch separating the earlyamples from the others. A more accurate estimate for the evo-utionary rate and the most recent common ancestor (MRCA)as obtained using the single rate dated tips (SRDT) model, a
tatistically well-founded likelihood method (Rambaut, 2000).
.8. DNA sequence submission
The nucleotide sequences reported in this paper were sub-itted in GenBank using the National Center for Biotechnology
nformation (NCBI, Bethesda, MD) Sequin, Version 6.15 underccession numbers DQ869562–DQ869567.
. Results
.1. Detection of group B rotaviruses
Of 611 group A rotavirus antigen negative specimensrom episodes of diarrhea among hospitalized patients duringuly–December 2003, enteric pathogens (S. typhi, non-typhialmonella, S. dysenteriae, S. flexneri, S. boydii, S. sonnei, V.holerae O139, and V. cholerae O1) were detected in 218 speci-ens (35.7%). Group B rotaviruses were detected in 13 samples
2.1% of the subset tested) by using RT-PCR and were confirmedy using nucleotide sequencing. Four group B rotavirus positiveamples were found to be co-infected with V. cholerae O1 (threeamples) and S. flexneri (one sample). On the other hand, dur-ng January–December 2004, 100 samples which were negativeor any pathogen, were tested for group B rotaviruses and fouramples (4% of the subset tested) gave positive results.
A polyacrylamide gel electrophoresis (PAGE) was also per-ormed on all 711 samples. Only six samples showed theharacteristic RNA migration pattern specific for group Botavirus (4-2-1-1-1-1-1). Fig. 1 shows the comparison of theNA migration patterns of groups A–C rotaviruses.
.2. Clinical features of the group B rotavirus infectedatients
Clinical features of the study patients (n = 711) were collectedrom hospital surveillance system. The age ranged from 3 dayso 80 years, mean 16.0 years and median 7.0 years. 53.6% ofhe patients were male. Fever (>37.8 ◦C) was detected in only
.5% and vomiting in 82.2%. Since, the number of group Botavirus infected patients were low, it was not practical to com-are them with other diarrhea patients. Clinical data of the 17atients infected with group B rotaviruses are listed in Table 1.yape
enomic double-stranded RNA migration patterns of group A (lane A), group(lane B), and group C (lane C) rotaviruses. Segment numbers for group B are
ndicated.
he age ranged from 9 months to 70 years, mean age, 27.8
ears and median, 26 years. Fourteen (82.4%) patients weredults (more than 16 years). Vomiting (82.4%) and abdominalain (65%) were common features while fever was not detectedxcept in one patient.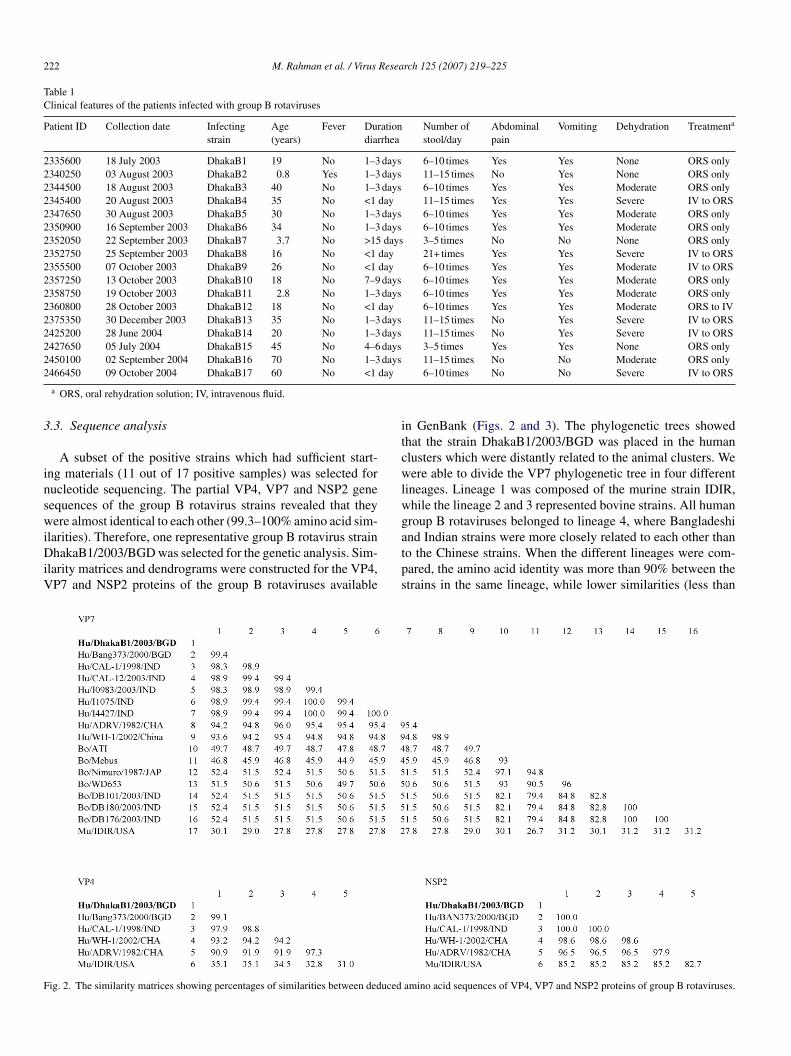
222 M. Rahman et al. / Virus Research 125 (2007) 219–225
Table 1Clinical features of the patients infected with group B rotaviruses
Patient ID Collection date Infectingstrain
Age(years)
Fever Durationdiarrhea
Number ofstool/day
Abdominalpain
Vomiting Dehydration Treatmenta
2335600 18 July 2003 DhakaB1 19 No 1–3 days 6–10 times Yes Yes None ORS only2340250 03 August 2003 DhakaB2 0.8 Yes 1–3 days 11–15 times No Yes None ORS only2344500 18 August 2003 DhakaB3 40 No 1–3 days 6–10 times Yes Yes Moderate ORS only2345400 20 August 2003 DhakaB4 35 No <1 day 11–15 times Yes Yes Severe IV to ORS2347650 30 August 2003 DhakaB5 30 No 1–3 days 6–10 times Yes Yes Moderate ORS only2350900 16 September 2003 DhakaB6 34 No 1–3 days 6–10 times Yes Yes Moderate ORS only2352050 22 September 2003 DhakaB7 3.7 No >15 days 3–5 times No No None ORS only2352750 25 September 2003 DhakaB8 16 No <1 day 21+ times Yes Yes Severe IV to ORS2355500 07 October 2003 DhakaB9 26 No <1 day 6–10 times Yes Yes Moderate IV to ORS2357250 13 October 2003 DhakaB10 18 No 7–9 days 6–10 times Yes Yes Moderate ORS only2358750 19 October 2003 DhakaB11 2.8 No 1–3 days 6–10 times Yes Yes Moderate ORS only2360800 28 October 2003 DhakaB12 18 No <1 day 6–10 times Yes Yes Moderate ORS to IV2375350 30 December 2003 DhakaB13 35 No 1–3 days 11–15 times No Yes Severe IV to ORS2425200 28 June 2004 DhakaB14 20 No 1–3 days 11–15 times No Yes Severe IV to ORS2427650 05 July 2004 DhakaB15 45 No 4–6 days 3–5 times Yes Yes None ORS only2 days2 day
3
inswiDiV
itcwlwg
F
450100 02 September 2004 DhakaB16 70 No 1–3466450 09 October 2004 DhakaB17 60 No <1
a ORS, oral rehydration solution; IV, intravenous fluid.
.3. Sequence analysis
A subset of the positive strains which had sufficient start-ng materials (11 out of 17 positive samples) was selected forucleotide sequencing. The partial VP4, VP7 and NSP2 geneequences of the group B rotavirus strains revealed that theyere almost identical to each other (99.3–100% amino acid sim-
larities). Therefore, one representative group B rotavirus strainhakaB1/2003/BGD was selected for the genetic analysis. Sim-
larity matrices and dendrograms were constructed for the VP4,P7 and NSP2 proteins of the group B rotaviruses available
atps
ig. 2. The similarity matrices showing percentages of similarities between deduced
11–15 times No No Moderate ORS only6–10 times No No Severe IV to ORS
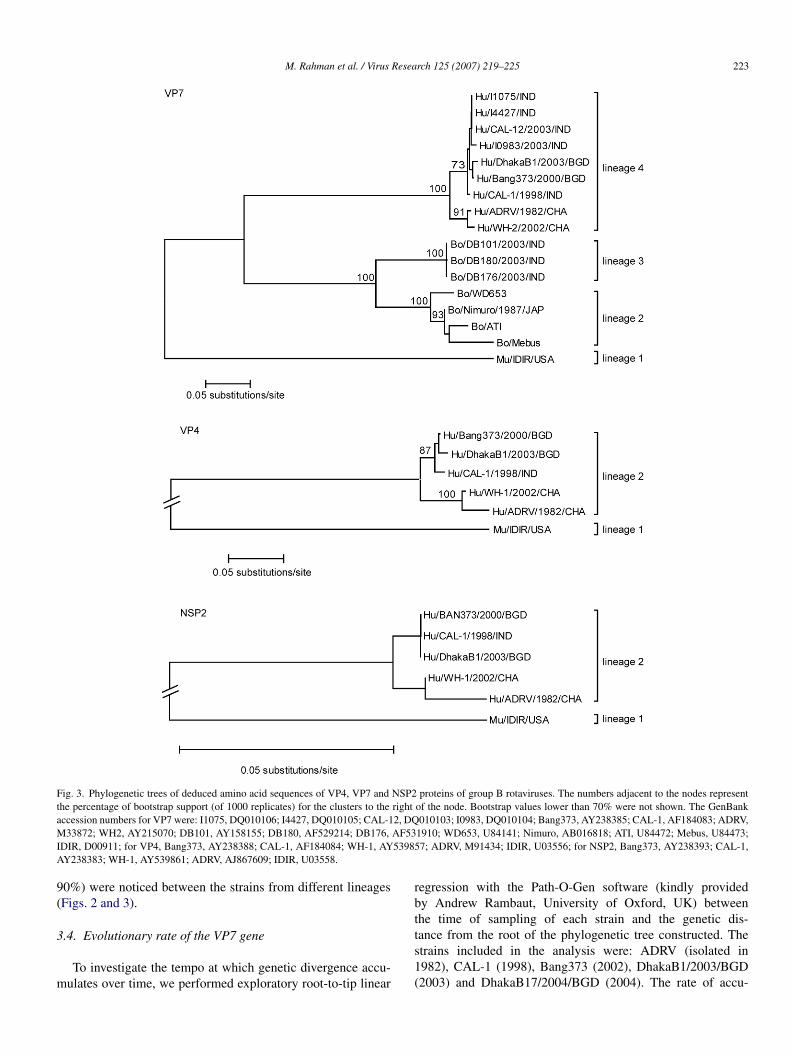
n GenBank (Figs. 2 and 3). The phylogenetic trees showedhat the strain DhakaB1/2003/BGD was placed in the humanlusters which were distantly related to the animal clusters. Weere able to divide the VP7 phylogenetic tree in four different
ineages. Lineage 1 was composed of the murine strain IDIR,hile the lineage 2 and 3 represented bovine strains. All humanroup B rotaviruses belonged to lineage 4, where Bangladeshi
nd Indian strains were more closely related to each other thano the Chinese strains. When the different lineages were com-ared, the amino acid identity was more than 90% between thetrains in the same lineage, while lower similarities (less thanamino acid sequences of VP4, VP7 and NSP2 proteins of group B rotaviruses.
M. Rahman et al. / Virus Research 125 (2007) 219–225 223
Fig. 3. Phylogenetic trees of deduced amino acid sequences of VP4, VP7 and NSP2 proteins of group B rotaviruses. The numbers adjacent to the nodes representthe percentage of bootstrap support (of 1000 replicates) for the clusters to the right of the node. Bootstrap values lower than 70% were not shown. The GenBanka 2, DQM AF53I 53985A
9(
3
m
rbt
ccession numbers for VP7 were: I1075, DQ010106; I4427, DQ010105; CAL-133872; WH2, AY215070; DB101, AY158155; DB180, AF529214; DB176,
DIR, D00911; for VP4, Bang373, AY238388; CAL-1, AF184084; WH-1, AYY238383; WH-1, AY539861; ADRV, AJ867609; IDIR, U03558.
0%) were noticed between the strains from different lineagesFigs. 2 and 3).
.4. Evolutionary rate of the VP7 gene
To investigate the tempo at which genetic divergence accu-ulates over time, we performed exploratory root-to-tip linear
ts1(
010103; I0983, DQ010104; Bang373, AY238385; CAL-1, AF184083; ADRV,1910; WD653, U84141; Nimuro, AB016818; ATI, U84472; Mebus, U84473;7; ADRV, M91434; IDIR, U03556; for NSP2, Bang373, AY238393; CAL-1,
egression with the Path-O-Gen software (kindly providedy Andrew Rambaut, University of Oxford, UK) betweenhe time of sampling of each strain and the genetic dis-
ance from the root of the phylogenetic tree constructed. Thetrains included in the analysis were: ADRV (isolated in982), CAL-1 (1998), Bang373 (2002), DhakaB1/2003/BGD2003) and DhakaB17/2004/BGD (2004). The rate of accu-
224 M. Rahman et al. / Virus Resea
Frs
mbdldo
4
ctopsta(Bb
B2Bimwaoptsv
etsfllc
bb
crfeaasm2atww
A
DR
R
A
A
B
B
C
C
E
E
F
G
G
ig. 4. Estimation of evolutionary rate and date of VP7 gene: linear root-to-tipegression plot presenting the correlation between the branch lengths and theampling dates of human group B rotavirus strains.
ulation of nucleotide changes in VP7 gene was estimated toe 1.57 × 10−3 nucleotide substitutions/(site year) (Fig. 4). Theate of the most recent common ancestor (MRCA) was calcu-ated to be 1976 (R2, 0.99). Due to lack of adequate sequenceata, it was not possible to calculate the evolutionary rate ofther genes.
. Discussion
Human group B rotaviruses have only been isolated in Asianountries (China, India and Bangladesh) although antibodieso the viruses were detected in humans from several countriesutside Asia. Besides humans, the viruses were detected in rats,igs, cows, and goats. Sequence analysis revealed that the humantrains were most similar to each other and were distantly relatedo the animal strains. The bovine and murine strains also showed
considerable amino acid sequence diversity between themFig. 2). It can be revealed that the gene segments of group
rotaviruses are very much host-specific and no reassortmentetween them was evident (Sen et al., 2003).
Recently, a considerably high percentage (18.5%) of grouprotaviruses was isolated from Indian children (Barman et al.,
006). In our study, the low rate of detection suggests that grouprotavirus is an uncommon cause of gastroenteritis in hospital-
zed patients in Bangladesh. It is possible that the true burdenight have been underestimated since hospitalized patients onlyere included in our study. It can also be noted that adults
re more likely to be infected by group B rotaviruses thanther diarrhea patients (82.4% versus 41.8% in our study sam-les, P < 0.01) and they usually do not come to hospital unlesshe disease becomes extremely severe. Therefore, a large-scaleurveillance study to understand the true epidemiology of theirus is required.
No classification system for group B rotaviruses has yet beenstablished. For group A rotaviruses, the strains having morehan 89% amino acid identical VP7 sequences belonged to theame G genotype (Gentsch et al., 1996). Taking this cut-off value
or the VP7 genes, group B rotaviruses can be divided into ateast four different G genotypes which also clustered in differentineages in the phylogenetic tree (Fig. 3). All human rotavirusesan be assigned as one distinct G genotype while bovine strainsH
rch 125 (2007) 219–225
elong to two different G genotypes. The murine strain mighte placed in a separate G genotype.
Based on limited sequence data the evolutionary rate we cal-ulated for the group B rotavirus VP7 genes was in the sameange of other rapidly evolving RNA viruses, which is a millionold higher than the rate calculated for mammalian genes. Inter-stingly, a Chinese group B rotavirus WH-1 isolated in 2002 waslmost similar to the prototype strain ADRV isolated 20 yearsgo in the same region (nucleotide similarities for different geneegments ranged from 97.8 to 98.8%) and showed a much lowerutation rate (7.9 × 10−4 substitutions/(site year)) (Yang et al.,
004). Why the evolutionary rate is different between Chinesend Indian–Bangladeshi lineages is yet to be addressed. Fur-her study is required to understand their evolutionary changes,hich is important for anticipating their pathogenicity andidespread epidemics.
cknowledgments
This work was supported by ICDDR,B, GPO Box-128,haka 1000, Bangladesh, and Rega Institute for Medicalesearch, University of Leuven, Leuven, Belgium.
eferences
hmed, M.U., Kobayashi, N., Wakuda, M., Sanekata, T., Taniguchi, K., Kader,A., Naik, T.N., Ishino, M., Alam, M.M., Kojima, K., Mise, K., Sumi, A.,2001. Genetic analysis of group B human rotaviruses detected in Bangladeshin 2000 and 2001. J. Med. Virol. 72 (1), 149–155.
ltschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basiclocal alignment search tool. J. Mol. Biol. 215 (3), 403–410.
arman, P., Ghosh, S., Das, S., Varghese, V., Chaudhuri, S., Sarkar, S., Krishnan,T., Bhattacharya, S.K., Chakrabarti, A., Kobayashi, N., Naik, T.N., 2004.Sequencing and sequence analysis of VP7 and NSP5 genes reveal emergenceof a new genotype of bovine group B rotaviruses in India. J. Clin. Microbiol.42 (6), 2816–2818.
arman, P., Ghosh, S., Samajdar, S., Mitra, U., Dutta, P., Bhattacharya, S.K.,Krishnan, T., Kobayashi, N., Naik, T.N., 2006. RT-PCR based diagnosisrevealed importance of human group B rotavirus infection in childhooddiarrhoea. J. Clin. Virol. 36 (3), 222–227.
hasey, D., Banks, J., 1984. The commonest rotaviruses from neonatal lambdiarrhoea in England and Wales have atypical electropherotypes. Vet. Rec.115 (13), 326–327.
hen, C.M., Hung, T., Bridger, J.C., McCrae, M.A., 1985. Chinese adultrotavirus is a group B rotavirus. Lancet 2 (8464), 1123–1124.
iden, J.J., Wilde, J., Firoozmand, F., Yolken, R., 1991. Detection of animal andhuman group B rotaviruses in fecal specimens by polymerase chain reaction.J. Clin. Microbiol. 29 (3), 539–543.
stes, M.K., 2001. Rotaviruses and their replication. In: Knipe, D.M.E.A. (Ed.),Field’s Virology, 2.2 vols. Lippincott Williams and Wilkins, pp. 1747–1786.
ang, Z.Y., Ye, Q., Ho, M.S., Dong, H., Qing, S., Penaranda, M.E., Hung, T.,Wen, L., Glass, R.I., 1989. Investigation of an outbreak of adult diarrhearotavirus in China. J. Infect. Dis. 160, 948–953.
entsch, J.R., Woods, P.A., Ramachandran, M., Das, B.K., Leite, J.P., Alfieri,A., Kumar, R., Bhan, M.K., Glass, R.I., 1996. Review of G and P typingresults from a global collection of rotavirus strains: implications for vaccinedevelopment. J. Infect. Dis. 174, S30–S36.
ouvea, V., Allen, J.R., Glass, R.I., Fang, Z.Y., Bremont, M., Cohen, J., McCrae,
M.A., Saif, L.J., Sinarachatanant, P., Caul, E.O., 1991. Detection of groupB and C rotaviruses by polymerase chain reaction. J. Clin. Microbiol. 29,519–523.erring, A.J., Inglis, N.F., Ojeh, C.K., Snodgrass, D.R., Menzies, J.D., 1982.Rapid diagnosis of rotavirus infection by direct detection of viral nucleic

Resea
H
H
K
K
K
K
K
M
M
N
N
P
P
R
S
S
S
S
T
T
T
T
U
Y
M. Rahman et al. / Virus
acid in silver-stained polyacrylamide gels. J. Clin. Microbiol. 16, 473–477.
ung, T., Chen, G.M., Wang, C.G., Chou, Z.Y., Chao, T.X., Ye, W.W., Yao, H.L.,Meng, K.H., 1983. Rotavirus-like agent in adult non-bacterial diarrhoea inChina. Lancet 2 (8358), 1078–1079.
ung, T., Chen, G.M., Wang, C.G., Yao, H.L., Fang, Z.Y., Chao, T.X., Chou,Z.Y., Ye, W., Chang, X.J., Den, S.S., et al., 1984. Waterborne outbreak ofrotavirus diarrhoea in adults in China caused by a novel rotavirus. Lancet 1(8387), 1139–1142.
apikian, A.Z., Hoshino, Y., Chanock, R.M., 2001. Rotaviruses. In: Knipe,D.M.E.A. (Ed.), Field’s Virology, 2.2 vols. Lippincott Williams and Wilkins,pp. 1787–1833.
elkar, S.D., Zade, J.K., 2004. Group B rotaviruses similar to strain CAL-1,have been circulating in Western India since 1993. Epidemiol. Infect. 132(4), 745–749.
obayashi, N., Naik, T.N., Kusuhara, Y., Krishnan, T., Sen, A., Bhattacharya,S.K., Taniguchi, K., Alam, M.M., Urasawa, T., Urasawa, S., 2001. Sequenceanalysis of genes encoding structural and nonstructural proteins of ahuman group B rotavirus detected in Calcutta. India J. Med. Virol. 64 (4),583–588.
rishnan, T., Sen, A., Choudhury, J.S., Das, S., Naik, T.N., Bhattacharya, S.K.,1999. Emergence of adult diarrhoea rotavirus in Calcutta, India. Lancet 353,380–381.
umar, S., Tamura, K., Nei, M., 2004. MEGA3: integrated software formolecular evolutionary genetics analysis and sequence alignment. BriefingsBioinform. 5, 150–163.
ackow, E.R., Werner-Eckert, R., Fay, M.E., Tao, H., Chen, G., 1993. Identifi-cation and baculovirus expression of the VP4 protein of the human group Brotavirus ADRV. J. Virol. 67 (5), 2730–2738.
ackow, E.R., 1995. Group B and C rotaviruses In: Blaser, M.J., Smith, P.D., Ravdin, J.I., Greenberg, H.B., Guerrant, R.L., (Eds.). Infections of thegastrointestinal tract. NY: Raven Press Ltd., pp. 983–1008.
akata, S., Estes, M.K., Graham, D.Y., Wang, S.S., Gary, G.W., Melnick, J.L.,1987. Detection of antibody to group B adult diarrhea rotaviruses in humans.J. Clin. Microbiol. 25 (5), 812–818.
icholas, K.B., Nicholas, H.B., Deerfield, D.W., 1997. GeneDoc: analysis andvisualization of genetic variation. Embnet News 4, 14.
arwani, A.V., Lucchelli, A., Saif, L.J., 1996. Identification of group Brotaviruses with short genome electropherotypes from adult cows with diar-rhea. J. Clin. Microbiol. 34 (5), 1303–1305.
Y
rch 125 (2007) 219–225 225
enaranda, M.E., Cubitt, W.D., Sinarachatanant, P., Taylor, D.N., Likanonsakul,S., Saif, L., Glass, R.I., 1989. Group C rotavirus infections in patients withdiarrhea in Thailand, Nepal, and England. J. Infect. Dis. 160 (3), 392–397.
ambaut, A., 2000. Estimating the rate of molecular evolution: incorporat-ing non-contemporaneous sequences into maximum likelihood phylogenies.Bioinformatics 16, 395–399.
aif, L.J., Jiang, B., 1994. Non-group A rotaviruses of humans and animals. In:Ramig, R.F. (Ed.), Rotaviruses. Springer-Verlag, pp. 339–371.
anekata, T., Kuwamoto, Y., Akamatsu, S., Sakon, N., Oseto, M., aniguchi, K.,Nakata, S., Estes, M.K., 1996. Isolation of group B porcine rotavirus in cellculture. J. Clin. Microbiol. 34 (3), 759–761.
anekata, T., Ahmed, M.U., Kader, A., Taniguchi, K., Kobayashi, N., 2003.Human group B rotavirus infections cause severe diarrhea in children andadults in Bangladesh. J. Clin. Microbiol. 41, 2187–2190.
en, A., Kobayashi, N., Bhattacharya, S.K., Naik, T.N., 2003. Molecular charac-terization and epidemiology of group B rotaviruses. In: Raghunath, D.E.A.(Ed.), Diarrhoeal Diseases, Current Status, Research Trends and Field Stud-ies. Tata McGraw-Hill Publishing Co. Ltd., pp. 137–178.
heil, K.W., Saif, L.J., Moorhead, P.D., Whitmoyer, R.E., 1985. Porcinerotavirus-like virus (group B rotavirus): characterization and pathogenicityfor gnotobiotic pigs. J. Clin. Microbiol. 21 (3), 340–345.
heil, K.W., Grooms, D.L., McCloskey, C.M., Redman, D.R., 1995. GroupB rotavirus associated with an outbreak of neonatal lamb diarrhea. J. Vet.Diagn. Invest. 7 (1), 148–150.
hompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G.,1997. The CLUSTAL X windows interface: flexible strategies for multi-ple sequence alignment aided by quality analysis tools. Nucl. Acids Res. 25,4876–4882.
sunemitsu, H., Morita, D., Takaku, H., Nishimori, T., Imai, K., Saif, L.J., 1999.First detection of bovine group B rotavirus in Japan and sequence of its VP7gene. Arch. Virol. 144 (4), 805–815.
shijima, H., Shinozaki, T., Fang, Z.Y., Glass, R.I., 1992. Group B rotavirusantibody in Japanese children. J. Diarrhoeal. Dis. Res. 10 (1), 41.
ang, H., Chen, S., Ji, S., 1998. A novel rotavirus causing large scale of adultdiarrhea in Shi Jiazhuang. Zhonghua Liu Xing Bing Xue Za Zhi. 19 (6),
336–338.ang, J.H., Kobayashi, N., Wang, Y.H., Zhou, X., Li, Y., Zhou, D.J., Hu, Z.H.,Ishino, M., Alam, M.M., Naik, T.N., Ahmed, M.U., 2004. Phylogeneticanalysis of a human group B rotavirus WH-1 detected in China in 2002. J.Med. Virol. 74 (4), 662–667.


















