
Semisynthetic protein nanoreactor for single-molecule ... · Semisynthetic protein nanoreactor for...
6
Semisynthetic protein nanoreactor for single-molecule chemistry Joongoo Lee and Hagan Bayley 1 Department of Chemistry, University of Oxford, Oxford OX1 3TA, United Kingdom Edited by Cynthia J. Burrows, University of Utah, Salt Lake City, UT, and approved September 16, 2015 (received for review June 2, 2015) The covalent chemistry of individual reactants bound within a protein pore can be monitored by observing the ionic current flow through the pore, which acts as a nanoreactor responding to bond-making and bond-breaking events. In the present work, we incorporated an unnatural amino acid into the α-hemolysin (αHL) pore by using solid-phase peptide synthesis to make the central segment of the polypeptide chain, which forms the transmembrane β-barrel of the assembled heptamer. The full-length αHL monomer was obtained by native chemical ligation of the central synthetic peptide to flanking recombinant polypeptides. αHL pores with one semisynthetic subunit were then used as nanoreactors for single-molecule chemistry. By in- troducing an amino acid with a terminal alkyne group, we were able to visualize click chemistry at the single-molecule level, which revealed a long-lived (4.5-s) reaction intermediate. Additional side chains might be introduced in a similar fashion, thereby greatly expanding the range of single-molecule covalent chemistry that can be investigated by the nanoreactor approach. semisynthesis | alpha-hemolysin | single-molecule chemistry | click chemistry T he staphylococcal α-hemolysin (αHL) pore has been engi- neered for a variety of applications in biotechnology. For example, the pore can function as a detector for stochastic sensing (1–3) and DNA sequencing (4, 5), and act as a “nanoreactor” for single-molecule covalent chemistry (6). In the nanoreactor ap- proach, individual bond-making and bond-breaking steps can be observed with submillisecond time resolution by monitoring the ionic current carried by the pore under an applied potential. Sin- gle-molecule chemistry has attracted interest because it provides insights into fundamental chemical processes that cannot be ob- served at the ensemble scale (6–8). However, chemistry carried out within the αHL nanoreactor has so far been confined to the re- actions of thiolates (i.e., deprotonated Cys side chains) (7–12) and derivatives of the side chains of Cys residues (13, 14). The in- corporation of unnatural amino acids into the αHL pore would substantially advance single-molecule chemistry, through the in- corporation of a large variety of individual side chains as well as multiple substitutions. We have produced αHL polypeptides with unnatural alkyl and aryl amino acids (15) in vitro by using chem- ically aminoacylated tRNAs. However, the method is demanding and often gives poor results, e.g., when more than one amino acid is introduced. In vivo methods that use engineered synthetase/ tRNA pairs also enable the incorporation of unnatural amino acids (16, 17). Two different amino acids (17, 18), but so far not more, can be incorporated. However, not all amino acids are successfully handled, and a new one often requires additional synthetase development. Further, various amino acid side chains are destroyed in vivo, often by reduction (18). Another means to incorporate unnatural amino acids is through native chemical ligation (NCL) (19–21), wherein the amino acid of interest is introduced in a peptide obtained by chemical synthesis (22, 23). The advantage of NCL is that virtually any amino acid or collection of amino acids can be incorporated into the peptide chain, provided the side chain is stable toward the conditions of solid-phase peptide synthesis (SPPS) and NCL, or can be pro- tected and deprotected. An extension of NCL that has seen widespread success is expressed protein ligation (24–26), a method for semisynthesis in which large segments of the polypeptide chain are bacterially expressed. However, relatively little work has been reported on the semisynthesis of membrane proteins (27–31). This deficiency can be attributed to the hydrophobic nature of membrane proteins, which are poorly soluble in either folded or unfolded form (32). For example, a polypeptide segment encompassing the trans- membrane region of the αHL pore precipitates in neutral buffer. The use of a denaturing environment during NCL can solve the solubility problem (20). However, it raises the question of whether the semisynthetic protein can be refolded into the native form. Here, we incorporate an unnatural amino acid with a terminal al- kyne side chain into the αHL polypeptide and demonstrate the functional activity of heptameric pores made from it. Further, we investigate click chemistry of the alkyne at the single-molecule level by using the semisynthetic pore as a nanoreactor. Results Preparation of Polypeptide Fragments. We designed polypeptide fragments that would provide for the efficient semisynthesis of a full-length αHL monomer by a three-way ligation strategy. In our design (Fig. 1A), a central segment [SP (synthetic peptide): SP[Thz 114 - Gly 126 ]-N-acyl-benzimidazolinone(Nbz)] was obtained by chemical synthesis. An N-terminal fragment (NTF: NTF[Ala 1 -Met 113 ]- α thioester) and a C-terminal fragment (CTF: CTF[Cys 127 -Asn 293 ]- D 8 H 6 ) were obtained after expression in Escherichia coli. Recombinant NTF[Ala 1 -Met 113 ]- α thioester was produced by thiolysis of a fusion protein, NTF-GyrA intein-CBD (NTF[Ala 1 -Met 113 ]- Mycobacterium xenopi DNA gyrase A (Mxe GyrA) intein-chitin binding domain (CBD)) (SI Appendix, Fig. S1) derived from the plasmid pTXB3. The DNA encoding residues 1–113 of αHL were PCR-amplified from a plasmid bearing a full-length αHL gene and cloned into pTXB3. After expression at 37 °C, high levels of the fusion protein were found within inclusion bodies in the E. coli host. Following cell lysis and centrifugation, most of the pellet Significance The modulation of ionic current flowing through an individual protein pore provides information at the single-molecule level about chemical reactions occurring within the pore. However, chemistry investigated in this way has been largely confined to the reactions of thiolates, presented by the side chains of cysteine residues. The introduction of unnatural amino acids would pro- vide a large variety of reactive side chains with which additional single-molecule chemistry could be investigated. Here we have produced semisynthetic protein pores containing a terminal al- kyne and used the pores as a nanoreactor to study Cu(I)-catalyzed azide-alkyne cycloaddition. A long-lived intermediate (4.5 s) in the reaction was directly observed. Author contributions: J.L. and H.B. designed research; J.L. performed research; J.L. ana- lyzed data; and J.L. and H.B. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. Email: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1510565112/-/DCSupplemental. 13768–13773 | PNAS | November 10, 2015 | vol. 112 | no. 45 www.pnas.org/cgi/doi/10.1073/pnas.1510565112 Downloaded by guest on January 9, 2020
Transcript of Semisynthetic protein nanoreactor for single-molecule ... · Semisynthetic protein nanoreactor for...

Semisynthetic protein nanoreactor forsingle-molecule chemistryJoongoo Lee and Hagan Bayley1
Department of Chemistry, University of Oxford, Oxford OX1 3TA, United Kingdom
Edited by Cynthia J. Burrows, University of Utah, Salt Lake City, UT, and approved September 16, 2015 (received for review June 2, 2015)
The covalent chemistry of individual reactants bound within a proteinpore can be monitored by observing the ionic current flow throughthe pore, which acts as a nanoreactor responding to bond-making andbond-breaking events. In the present work, we incorporated anunnatural amino acid into the α-hemolysin (αHL) pore by usingsolid-phase peptide synthesis to make the central segment of thepolypeptide chain, which forms the transmembrane β-barrel of theassembled heptamer. The full-length αHL monomer was obtained bynative chemical ligation of the central synthetic peptide to flankingrecombinant polypeptides. αHL pores with one semisynthetic subunitwere then used as nanoreactors for single-molecule chemistry. By in-troducing an amino acid with a terminal alkyne group, we were ableto visualize click chemistry at the single-molecule level, which revealeda long-lived (4.5-s) reaction intermediate. Additional side chains mightbe introduced in a similar fashion, thereby greatly expanding therange of single-molecule covalent chemistry that can be investigatedby the nanoreactor approach.
semisynthesis | alpha-hemolysin | single-molecule chemistry | click chemistry
The staphylococcal α-hemolysin (αHL) pore has been engi-neered for a variety of applications in biotechnology. For
example, the pore can function as a detector for stochastic sensing(1–3) and DNA sequencing (4, 5), and act as a “nanoreactor” forsingle-molecule covalent chemistry (6). In the nanoreactor ap-proach, individual bond-making and bond-breaking steps can beobserved with submillisecond time resolution by monitoring theionic current carried by the pore under an applied potential. Sin-gle-molecule chemistry has attracted interest because it providesinsights into fundamental chemical processes that cannot be ob-served at the ensemble scale (6–8). However, chemistry carried outwithin the αHL nanoreactor has so far been confined to the re-actions of thiolates (i.e., deprotonated Cys side chains) (7–12) andderivatives of the side chains of Cys residues (13, 14). The in-corporation of unnatural amino acids into the αHL pore wouldsubstantially advance single-molecule chemistry, through the in-corporation of a large variety of individual side chains as well asmultiple substitutions. We have produced αHL polypeptides withunnatural alkyl and aryl amino acids (15) in vitro by using chem-ically aminoacylated tRNAs. However, the method is demandingand often gives poor results, e.g., when more than one amino acidis introduced. In vivo methods that use engineered synthetase/tRNA pairs also enable the incorporation of unnatural aminoacids (16, 17). Two different amino acids (17, 18), but so far notmore, can be incorporated. However, not all amino acids aresuccessfully handled, and a new one often requires additionalsynthetase development. Further, various amino acid side chainsare destroyed in vivo, often by reduction (18).Another means to incorporate unnatural amino acids is through
native chemical ligation (NCL) (19–21), wherein the amino acid ofinterest is introduced in a peptide obtained by chemical synthesis(22, 23). The advantage of NCL is that virtually any amino acid orcollection of amino acids can be incorporated into the peptidechain, provided the side chain is stable toward the conditions ofsolid-phase peptide synthesis (SPPS) and NCL, or can be pro-tected and deprotected. An extension of NCL that has seenwidespread success is expressed protein ligation (24–26), a method
for semisynthesis in which large segments of the polypeptide chainare bacterially expressed. However, relatively little work has beenreported on the semisynthesis of membrane proteins (27–31). Thisdeficiency can be attributed to the hydrophobic nature of membraneproteins, which are poorly soluble in either folded or unfolded form(32). For example, a polypeptide segment encompassing the trans-membrane region of the αHL pore precipitates in neutral buffer.The use of a denaturing environment during NCL can solve thesolubility problem (20). However, it raises the question of whetherthe semisynthetic protein can be refolded into the native form.Here, we incorporate an unnatural amino acid with a terminal al-kyne side chain into the αHL polypeptide and demonstrate thefunctional activity of heptameric pores made from it. Further, weinvestigate click chemistry of the alkyne at the single-molecule levelby using the semisynthetic pore as a nanoreactor.
ResultsPreparation of Polypeptide Fragments. We designed polypeptidefragments that would provide for the efficient semisynthesis of afull-length αHL monomer by a three-way ligation strategy. In ourdesign (Fig. 1A), a central segment [SP (synthetic peptide): SP[Thz114-Gly126]-N-acyl-benzimidazolinone(Nbz)] was obtained by chemicalsynthesis. An N-terminal fragment (NTF: NTF[Ala1-Met113]-αthioester) and a C-terminal fragment (CTF: CTF[Cys127-Asn293]-D8H6) were obtained after expression in Escherichia coli.Recombinant NTF[Ala1-Met113]-αthioester was produced by thiolysis
of a fusion protein, NTF-GyrA intein-CBD (NTF[Ala1-Met113]-Mycobacterium xenopi DNA gyrase A (Mxe GyrA) intein-chitinbinding domain (CBD)) (SI Appendix, Fig. S1) derived from theplasmid pTXB3. The DNA encoding residues 1–113 of αHL werePCR-amplified from a plasmid bearing a full-length αHL gene andcloned into pTXB3. After expression at 37 °C, high levels of thefusion protein were found within inclusion bodies in the E. colihost. Following cell lysis and centrifugation, most of the pellet
Significance
The modulation of ionic current flowing through an individualprotein pore provides information at the single-molecule levelabout chemical reactions occurring within the pore. However,chemistry investigated in this way has been largely confined tothe reactions of thiolates, presented by the side chains of cysteineresidues. The introduction of unnatural amino acids would pro-vide a large variety of reactive side chains with which additionalsingle-molecule chemistry could be investigated. Here we haveproduced semisynthetic protein pores containing a terminal al-kyne and used the pores as a nanoreactor to study Cu(I)-catalyzedazide-alkyne cycloaddition. A long-lived intermediate (4.5 s) inthe reaction was directly observed.
Author contributions: J.L. and H.B. designed research; J.L. performed research; J.L. ana-lyzed data; and J.L. and H.B. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1510565112/-/DCSupplemental.
13768–13773 | PNAS | November 10, 2015 | vol. 112 | no. 45 www.pnas.org/cgi/doi/10.1073/pnas.1510565112
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020
could be solubilized in denaturing buffer containing 8 M urea.To allow binding to chitin beads, the concentration of theurea was reduced to 1 M. Unbound proteins were washed offthe column and the bound fusion protein was treated with2-mercaptoethane sulfonate under gravity flow, yielding theNTF[Ala1-Met113]-αthioester in high purity as determined by SDS/PAGE (SI Appendix, Fig. S1B) and LC-MS (liquid chromatography–mass spectrometry) (SI Appendix, Fig. S2).We prepared another plasmid (pT7-SC1-CDH, SI Appendix,
Fig. S3) for the production of CTF[Cys127-Asn293]-D8H6 undercontrol of the T7 promoter. In this plasmid, the codons for residues1–126 in the pT7-SC1 plasmid, which contains a full-length αHLgene, are replaced with codons for Met-Cys. CTF was againobtained from inclusion bodies, and then purified in 6 M guani-dine hydrochloride by use of the His6 tag at the C terminus. TheN-terminal Met was found to be cleaved and the unmasked Cysresidue had undergone condensation with pyruvic acid, an abun-dant metabolite (33). We regenerated free Cys at the N terminus bycleaving the thiazolidine (Thz) with methoxylamine (MeONH2) atpH 4 (SI Appendix, Fig. S4B). The purified CTF (SI Appendix, Fig.S4A) was characterized by LC-MS (SI Appendix, Fig. S5).The central synthetic peptide (SP: Thz114ThrLeuPrpTyrGly-
PheAsnGlyAsnValThrGly126-Nbz), in which Thr-117 is replacedwith L-propargylglycine (Prp), was prepared by SPPS (34) withFmoc-protected amino acids (SI Appendix, Fig. S6). The alkyneside chain of residue 117 was predicted to point into the trans-membrane β-barrel of the αHL pore (35); residue 117 is a sen-sitive position for the detection of chemical reactions (6). Thepeptide as released from the resin contained a C-terminal Nbzgroup, which undergoes thiolysis, with (4-mercaptopropionicacetic acid, MPAA), to yield a peptide-αarylthioester (34). TheHPLC-purified peptide (SI Appendix, Fig. S7A) was character-ized by LC-MS (SI Appendix, Fig. S7B).
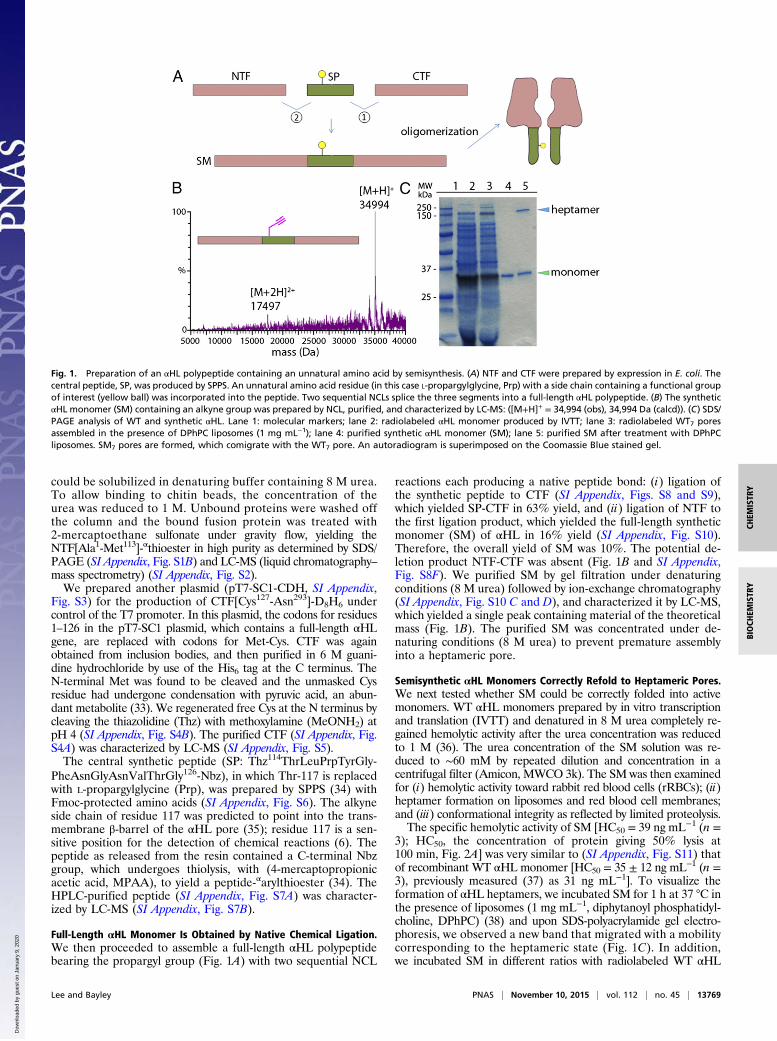
Full-Length αHL Monomer Is Obtained by Native Chemical Ligation.We then proceeded to assemble a full-length αHL polypeptidebearing the propargyl group (Fig. 1A) with two sequential NCL
reactions each producing a native peptide bond: (i) ligation ofthe synthetic peptide to CTF (SI Appendix, Figs. S8 and S9),which yielded SP-CTF in 63% yield, and (ii) ligation of NTF tothe first ligation product, which yielded the full-length syntheticmonomer (SM) of αHL in 16% yield (SI Appendix, Fig. S10).Therefore, the overall yield of SM was 10%. The potential de-letion product NTF-CTF was absent (Fig. 1B and SI Appendix,Fig. S8F). We purified SM by gel filtration under denaturingconditions (8 M urea) followed by ion-exchange chromatography(SI Appendix, Fig. S10 C and D), and characterized it by LC-MS,which yielded a single peak containing material of the theoreticalmass (Fig. 1B). The purified SM was concentrated under de-naturing conditions (8 M urea) to prevent premature assemblyinto a heptameric pore.
Semisynthetic αHL Monomers Correctly Refold to Heptameric Pores.We next tested whether SM could be correctly folded into activemonomers. WT αHL monomers prepared by in vitro transcriptionand translation (IVTT) and denatured in 8 M urea completely re-gained hemolytic activity after the urea concentration was reducedto 1 M (36). The urea concentration of the SM solution was re-duced to ∼60 mM by repeated dilution and concentration in acentrifugal filter (Amicon, MWCO 3k). The SM was then examinedfor (i) hemolytic activity toward rabbit red blood cells (rRBCs); (ii)heptamer formation on liposomes and red blood cell membranes;and (iii) conformational integrity as reflected by limited proteolysis.The specific hemolytic activity of SM [HC50 = 39 ng mL−1 (n =
3); HC50, the concentration of protein giving 50% lysis at100 min, Fig. 2A] was very similar to (SI Appendix, Fig. S11) thatof recombinant WT αHL monomer [HC50 = 35 ± 12 ng mL−1 (n =3), previously measured (37) as 31 ng mL−1]. To visualize theformation of αHL heptamers, we incubated SM for 1 h at 37 °C inthe presence of liposomes (1 mg mL−1, diphytanoyl phosphatidyl-choline, DPhPC) (38) and upon SDS-polyacrylamide gel electro-phoresis, we observed a new band that migrated with a mobilitycorresponding to the heptameric state (Fig. 1C). In addition,we incubated SM in different ratios with radiolabeled WT αHL
Fig. 1. Preparation of an αHL polypeptide containing an unnatural amino acid by semisynthesis. (A) NTF and CTF were prepared by expression in E. coli. Thecentral peptide, SP, was produced by SPPS. An unnatural amino acid residue (in this case L-propargylglycine, Prp) with a side chain containing a functional groupof interest (yellow ball) was incorporated into the peptide. Two sequential NCLs splice the three segments into a full-length αHL polypeptide. (B) The syntheticαHL monomer (SM) containing an alkyne group was prepared by NCL, purified, and characterized by LC-MS: ([M+H]+ = 34,994 (obs), 34,994 Da (calcd)). (C) SDS/PAGE analysis of WT and synthetic αHL. Lane 1: molecular markers; lane 2: radiolabeled αHL monomer produced by IVTT; lane 3: radiolabeled WT7 poresassembled in the presence of DPhPC liposomes (1 mg mL−1); lane 4: purified synthetic αHL monomer (SM); lane 5: purified SM after treatment with DPhPCliposomes. SM7 pores are formed, which comigrate with the WT7 pore. An autoradiogram is superimposed on the Coomassie Blue stained gel.
Lee and Bayley PNAS | November 10, 2015 | vol. 112 | no. 45 | 13769
CHEM
ISTR
YBIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020

produced by IVTT in the presence of rabbit red blood cell mem-branes (rRBCm). SM oligomerized to form heteroheptamers withdifferent stoichiometries (WT7−nSMn, n = 0–7) (Fig. 3A). To-gether, these data show that SM forms functional heptamericpores. A limited proteolysis experiment was carried out withproteinase K, to determine the conformational state of the αHLpolypeptide in the monomer and the heptamer (39–41). We foundthat the monomer was cleaved into two fragments of 20 and15 kDa, indicating that it is correctly folded (Fig. 2B, lane 4).When the heptamer formed on liposomes was similarly treated, alarge fraction of the polypeptide chains was protected from theprotease (Fig. 2B, lane 5), indicating that the heptamer was cor-rectly assembled (SI Appendix, Fig. S12).
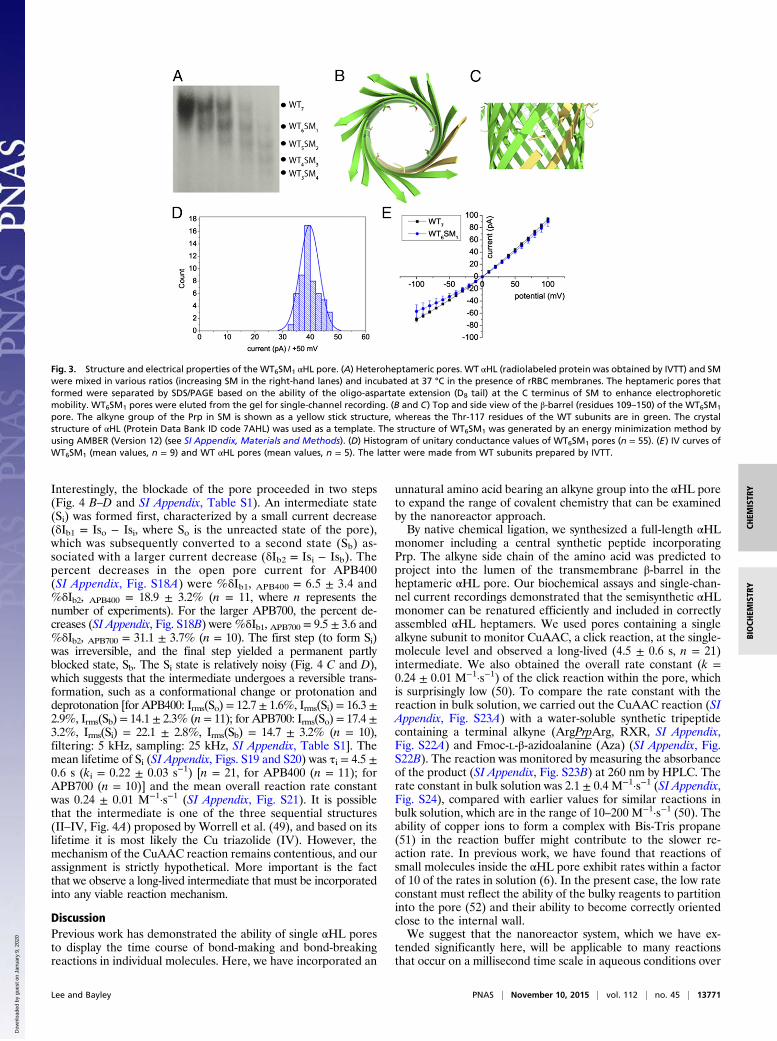
The Electrical Properties of WT and Semisynthetic Pores Are Similar.We examined the electrical properties of WT6SM1 pores bydetermining the mean conductance under defined conditions
(Fig. 3D) and measuring IV curves (Fig. 3E). The conductance ofWT6SM1 (0.79 ± 0.07 nS at +50 mV in 1 M KCl, 20 mM Bis-Trispropane, pH 8.5) was similar to that of the WT7 pore (∼0.88 nS,same conditions). We also investigated the binding kinetics ofβ-cyclodextrin (βCD) with WT6SM1 to confirm that the structureof the transmembrane β-barrel was intact. βCD is a noncovalentmolecular adaptor that acts as a transient channel blocker when itis lodged in the lumen of the αHL pore (42, 43). In single-channelrecordings, the addition of βCD to the trans compartment pro-duced reversible, transient, partial blockades of the ionic current.The mean dwell time of βCD in the pore (τoff) did not vary withβCD concentration, whereas the mean lifetime of the unoccupiedstate (τon) was inversely proportional to the concentration, sug-gesting a bimolecular interaction (SI Appendix, Fig. S13). Wedetermined the association rate constant (kon) and dissociationrate constant (koff) of βCD binding from the τ-values: kon = 1.1 ±0.3 × 105 M−1·s−1; koff = 1.4 ± 0.1 × 103 s−1; KD = koff/kon = 1.3 ±0.4 × 10−2 M. Similar values were obtained previously for the WTαHL pore (43). Therefore, the alkyne residue presented by the SMsubunit neither affects the electrical properties of the αHL porenor its ability to bind the βCD adaptor.
The Semisynthetic Pore Reveals an Intermediate in the Copper(I)-Catalyzed Azide-Alkyne Cycloaddition Reaction. We then used theprotein nanoreactor approach to investigate click chemistry ofthe alkyne side chain in WT6SM1 at the single-molecule level.The copper(I)-catalyzed azide-alkyne cycloaddition (CuAAC), aclick reaction (44), has been widely studied in materials science(45), bioconjugation (46), and drug discovery (47). We first ex-amined the unfunctionalized WT7 pore (SI Appendix, Fig. S15) inthe presence of the reagents used for the click reaction, namelythe Cu(I) catalyst and azide substrates (SI Appendix, Fig. S14,APB400: 0.4 kDa azide-PEG-biotin; APB700: 0.7 kDa azide-PEG-biotin). Cu(I) (20 mM) and APB400 (0.5–2.0 mM), neitherseparately nor together, affected the WT7 pore (SI Appendix, Fig.S15 A–G), whereas APB700, either by itself (SI Appendix, Fig. S15H–K) or with Cu(I) (SI Appendix, Fig. S15L), produced reversibleevents, which increased in frequency with APB700 concentration.These events most likely arise from entry of APB700 into thelumen of the pore, without covalent attachment, for periodssufficient to be detected (48). By examining the concentrationdependence of the blockades (APB700, 0.5 to 2 mM) in theabsence of Cu(I), we obtained kon = 1.3 ± 0.2 × 105 M−1·s−1;koff = 8.7 ± 0.3 × 103 s−1; KD = koff/kon = 6.7 ± 1.0 × 10−2 M (SIAppendix, Fig. S17), similar to the values obtained by Movileanuet al. for a 940-Da PEG (48). The mean lifetime of the blockadeswas short (∼100 μs), resulting in a broad distribution of amplitudesunder our data acquisition conditions. No permanent current de-crease was observed with the WT7 pore during 3 h of monitoringafter the addition of Cu(I), with either of the azides present inboth compartments.We then carried out chemistry with single WT6SM1 pores. As
in the case of the WT7 pore, Cu(I) added to both the cis andtrans compartments at the same time did not generate significantchanges in the current (SI Appendix, Fig. S16 A and B), sug-gesting that if a Cu acetylide (Fig. 4 A, I) is formed it is short-lived. When APB700 was added to the trans compartment in theabsence of Cu(I), short reversible current blockades character-istic of entry and exit from the pore were seen, which weresimilar to those observed with the WT7 pore: kon = 1.6 ± 0.2 ×105 M−1·s−1; koff = 5.6 ± 1.1 × 103 s−1; KD = koff/kon = 3.5 ± 0.8 ×10−2 M (SI Appendix, Fig. S16 C–L). In the case of APB400,the events were presumably too short to be registered at thedata acquisition rate used (48). By contrast with WT7, the additionof Cu(I) to both compartments in the presence of 2 mM APB400or APB700 (trans) eventually led to an irreversible partialblockade of the WT6SM1 pore, suggesting that Cu(I) triggered areaction between the substrate and the alkyne within the pore.
Fig. 2. Functional properties of the semisynthetic αHL polypeptide. (A) He-molysis assays. Row 1: hemolysis by recombinant WT αHL; row 2: hemolysis bySM; row 3: rRBCs with no αHL addition. The decrease in light scattering overtime was recorded in a microplate reader. WT (1 μL, 1.08mgmL−1) and SM αHLmonomer (1 μL, 4.02 mg mL−1) were added to the first column of row 1 androw 2 of the plate, respectively, and each sample was serially twofold dilutedin a final volume of 100 μL per well. (B) Limited proteolysis with proteinase Kof SM in solution and in the presence of DPhPC liposomes (1 mg mL−1). Lane 1:molecular markers; lane 2: SM; lane 3: SM treated with DPhPC liposomes; lane4: SM in solution digested by proteinase K. Two fragments (∼20 and 15 kDa)were generated (b and c); lane 5: SM treated with DPhPC liposomes wasfurther treated with proteinase K and then heat-denatured at 95 °C to dis-assemble the heptamers. The central proteolytic site is inaccessible in SM7, andmost of the SM polypeptide is undigested. Fragment a is generated aftercleavage near the N terminus in the prepore form of SM7 (∼32 kDa) (41). Notethat a single irrelevant lane was excised between lanes 4 and 5.
13770 | www.pnas.org/cgi/doi/10.1073/pnas.1510565112 Lee and Bayley
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020
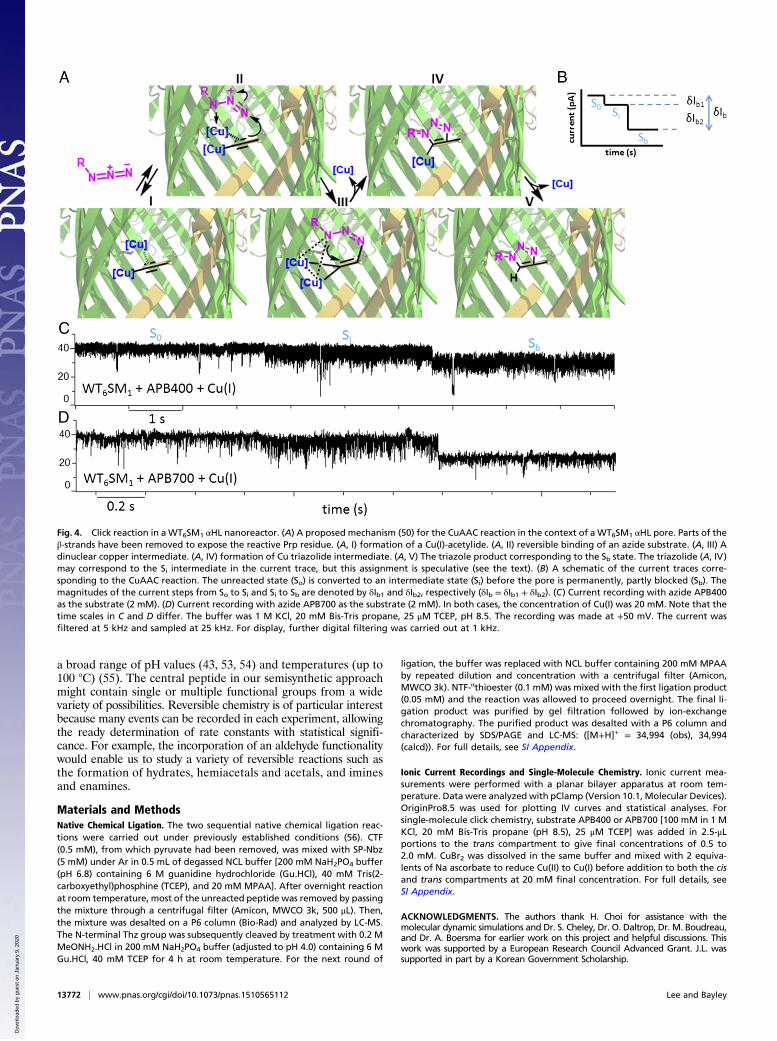
Interestingly, the blockade of the pore proceeded in two steps(Fig. 4 B–D and SI Appendix, Table S1). An intermediate state(Si) was formed first, characterized by a small current decrease(δIb1 = Iso − Isi, where So is the unreacted state of the pore),which was subsequently converted to a second state (Sb) as-sociated with a larger current decrease (δIb2 = Isi − Isb). Thepercent decreases in the open pore current for APB400(SI Appendix, Fig. S18A) were %δIb1, APB400 = 6.5 ± 3.4 and%δIb2, APB400 = 18.9 ± 3.2% (n = 11, where n represents thenumber of experiments). For the larger APB700, the percent de-creases (SI Appendix, Fig. S18B) were %δIb1, APB700 = 9.5 ± 3.6 and%δIb2, APB700 = 31.1 ± 3.7% (n = 10). The first step (to form Si)was irreversible, and the final step yielded a permanent partlyblocked state, Sb. The Si state is relatively noisy (Fig. 4 C and D),which suggests that the intermediate undergoes a reversible trans-formation, such as a conformational change or protonation anddeprotonation [for APB400: Irms(So) = 12.7 ± 1.6%, Irms(Si) = 16.3 ±2.9%, Irms(Sb) = 14.1 ± 2.3% (n = 11); for APB700: Irms(So) = 17.4 ±3.2%, Irms(Si) = 22.1 ± 2.8%, Irms(Sb) = 14.7 ± 3.2% (n = 10),filtering: 5 kHz, sampling: 25 kHz, SI Appendix, Table S1]. Themean lifetime of Si (SI Appendix, Figs. S19 and S20) was τi = 4.5 ±0.6 s (ki = 0.22 ± 0.03 s−1) [n = 21, for APB400 (n = 11); forAPB700 (n = 10)] and the mean overall reaction rate constantwas 0.24 ± 0.01 M−1·s−1 (SI Appendix, Fig. S21). It is possiblethat the intermediate is one of the three sequential structures(II–IV, Fig. 4A) proposed by Worrell et al. (49), and based on itslifetime it is most likely the Cu triazolide (IV). However, themechanism of the CuAAC reaction remains contentious, and ourassignment is strictly hypothetical. More important is the factthat we observe a long-lived intermediate that must be incorporatedinto any viable reaction mechanism.
DiscussionPrevious work has demonstrated the ability of single αHL poresto display the time course of bond-making and bond-breakingreactions in individual molecules. Here, we have incorporated an
unnatural amino acid bearing an alkyne group into the αHL poreto expand the range of covalent chemistry that can be examinedby the nanoreactor approach.By native chemical ligation, we synthesized a full-length αHL
monomer including a central synthetic peptide incorporatingPrp. The alkyne side chain of the amino acid was predicted toproject into the lumen of the transmembrane β-barrel in theheptameric αHL pore. Our biochemical assays and single-chan-nel current recordings demonstrated that the semisynthetic αHLmonomer can be renatured efficiently and included in correctlyassembled αHL heptamers. We used pores containing a singlealkyne subunit to monitor CuAAC, a click reaction, at the single-molecule level and observed a long-lived (4.5 ± 0.6 s, n = 21)intermediate. We also obtained the overall rate constant (k =0.24 ± 0.01 M−1·s−1) of the click reaction within the pore, whichis surprisingly low (50). To compare the rate constant with thereaction in bulk solution, we carried out the CuAAC reaction (SIAppendix, Fig. S23A) with a water-soluble synthetic tripeptidecontaining a terminal alkyne (ArgPrpArg, RXR, SI Appendix,Fig. S22A) and Fmoc-L-β-azidoalanine (Aza) (SI Appendix, Fig.S22B). The reaction was monitored by measuring the absorbanceof the product (SI Appendix, Fig. S23B) at 260 nm by HPLC. Therate constant in bulk solution was 2.1 ± 0.4 M−1·s−1 (SI Appendix,Fig. S24), compared with earlier values for similar reactions inbulk solution, which are in the range of 10–200 M−1·s−1 (50). Theability of copper ions to form a complex with Bis-Tris propane(51) in the reaction buffer might contribute to the slower re-action rate. In previous work, we have found that reactions ofsmall molecules inside the αHL pore exhibit rates within a factorof 10 of the rates in solution (6). In the present case, the low rateconstant must reflect the ability of the bulky reagents to partitioninto the pore (52) and their ability to become correctly orientedclose to the internal wall.We suggest that the nanoreactor system, which we have ex-
tended significantly here, will be applicable to many reactionsthat occur on a millisecond time scale in aqueous conditions over
Fig. 3. Structure and electrical properties of the WT6SM1 αHL pore. (A) Heteroheptameric pores. WT αHL (radiolabeled protein was obtained by IVTT) and SMwere mixed in various ratios (increasing SM in the right-hand lanes) and incubated at 37 °C in the presence of rRBC membranes. The heptameric pores thatformed were separated by SDS/PAGE based on the ability of the oligo-aspartate extension (D8 tail) at the C terminus of SM to enhance electrophoreticmobility. WT6SM1 pores were eluted from the gel for single-channel recording. (B and C) Top and side view of the β-barrel (residues 109–150) of the WT6SM1
pore. The alkyne group of the Prp in SM is shown as a yellow stick structure, whereas the Thr-117 residues of the WT subunits are in green. The crystalstructure of αHL (Protein Data Bank ID code 7AHL) was used as a template. The structure of WT6SM1 was generated by an energy minimization method byusing AMBER (Version 12) (see SI Appendix, Materials and Methods). (D) Histogram of unitary conductance values of WT6SM1 pores (n = 55). (E) IV curves ofWT6SM1 (mean values, n = 9) and WT αHL pores (mean values, n = 5). The latter were made from WT subunits prepared by IVTT.
Lee and Bayley PNAS | November 10, 2015 | vol. 112 | no. 45 | 13771
CHEM
ISTR
YBIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020
a broad range of pH values (43, 53, 54) and temperatures (up to100 °C) (55). The central peptide in our semisynthetic approachmight contain single or multiple functional groups from a widevariety of possibilities. Reversible chemistry is of particular interestbecause many events can be recorded in each experiment, allowingthe ready determination of rate constants with statistical signifi-cance. For example, the incorporation of an aldehyde functionalitywould enable us to study a variety of reversible reactions such asthe formation of hydrates, hemiacetals and acetals, and iminesand enamines.
Materials and MethodsNative Chemical Ligation. The two sequential native chemical ligation reac-tions were carried out under previously established conditions (56). CTF(0.5 mM), from which pyruvate had been removed, was mixed with SP-Nbz(5 mM) under Ar in 0.5 mL of degassed NCL buffer [200 mM NaH2PO4 buffer(pH 6.8) containing 6 M guanidine hydrochloride (Gu.HCl), 40 mM Tris(2-carboxyethyl)phosphine (TCEP), and 20 mM MPAA]. After overnight reactionat room temperature, most of the unreacted peptide was removed by passingthe mixture through a centrifugal filter (Amicon, MWCO 3k, 500 μL). Then,the mixture was desalted on a P6 column (Bio-Rad) and analyzed by LC-MS.The N-terminal Thz group was subsequently cleaved by treatment with 0.2 MMeONH2.HCl in 200 mM NaH2PO4 buffer (adjusted to pH 4.0) containing 6 MGu.HCl, 40 mM TCEP for 4 h at room temperature. For the next round of
ligation, the buffer was replaced with NCL buffer containing 200 mM MPAAby repeated dilution and concentration with a centrifugal filter (Amicon,MWCO 3k). NTF-αthioester (0.1 mM) was mixed with the first ligation product(0.05 mM) and the reaction was allowed to proceed overnight. The final li-gation product was purified by gel filtration followed by ion-exchangechromatography. The purified product was desalted with a P6 column andcharacterized by SDS/PAGE and LC-MS: ([M+H]+ = 34,994 (obs), 34,994(calcd)). For full details, see SI Appendix.
Ionic Current Recordings and Single-Molecule Chemistry. Ionic current mea-surements were performed with a planar bilayer apparatus at room tem-perature. Data were analyzed with pClamp (Version 10.1, Molecular Devices).OriginPro8.5 was used for plotting IV curves and statistical analyses. Forsingle-molecule click chemistry, substrate APB400 or APB700 [100 mM in 1 MKCl, 20 mM Bis-Tris propane (pH 8.5), 25 μM TCEP] was added in 2.5-μLportions to the trans compartment to give final concentrations of 0.5 to2.0 mM. CuBr2 was dissolved in the same buffer and mixed with 2 equiva-lents of Na ascorbate to reduce Cu(II) to Cu(I) before addition to both the cisand trans compartments at 20 mM final concentration. For full details, seeSI Appendix.
ACKNOWLEDGMENTS. The authors thank H. Choi for assistance with themolecular dynamic simulations and Dr. S. Cheley, Dr. O. Daltrop, Dr. M. Boudreau,and Dr. A. Boersma for earlier work on this project and helpful discussions. Thiswork was supported by a European Research Council Advanced Grant. J.L. wassupported in part by a Korean Government Scholarship.
Fig. 4. Click reaction in aWT6SM1 αHL nanoreactor. (A) A proposed mechanism (50) for the CuAAC reaction in the context of a WT6SM1 αHL pore. Parts of theβ-strands have been removed to expose the reactive Prp residue. (A, I) formation of a Cu(I)-acetylide. (A, II) reversible binding of an azide substrate. (A, III) Adinuclear copper intermediate. (A, IV) formation of Cu triazolide intermediate. (A, V) The triazole product corresponding to the Sb state. The triazolide (A, IV)may correspond to the Si intermediate in the current trace, but this assignment is speculative (see the text). (B) A schematic of the current traces corre-sponding to the CuAAC reaction. The unreacted state (So) is converted to an intermediate state (Si) before the pore is permanently, partly blocked (Sb). Themagnitudes of the current steps from So to Si and Si to Sb are denoted by δIb1 and δIb2, respectively (δIb = δIb1 + δIb2). (C) Current recording with azide APB400as the substrate (2 mM). (D) Current recording with azide APB700 as the substrate (2 mM). In both cases, the concentration of Cu(I) was 20 mM. Note that thetime scales in C and D differ. The buffer was 1 M KCl, 20 mM Bis-Tris propane, 25 μM TCEP, pH 8.5. The recording was made at +50 mV. The current wasfiltered at 5 kHz and sampled at 25 kHz. For display, further digital filtering was carried out at 1 kHz.
13772 | www.pnas.org/cgi/doi/10.1073/pnas.1510565112 Lee and Bayley
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020

1. Bayley H, Cremer PS (2001) Stochastic sensors inspired by biology. Nature 413(6852):226–230.
2. Boersma AJ, Bayley H (2012) Continuous stochastic detection of amino acid enan-tiomers with a protein nanopore. Angew Chem Int Ed Engl 51(38):9606.
3. Harrington L, Cheley S, Alexander LT, Knapp S, Bayley H (2013) Stochastic detection ofPim protein kinases reveals electrostatically enhanced association of a peptide sub-strate. Proc Natl Acad Sci USA 110(47):E4417–E4426.
4. Branton D, et al. (2008) The potential and challenges of nanopore sequencing. NatBiotechnol 26(10):1146–1153.
5. Bayley H (2015) Nanopore sequencing: From imagination to reality. Clin Chem 61(1):25–31.
6. Bayley H, Luchian T, Shin S-H, Steffensen M (2008) Single-molecule covalent chemistryin a protein nanoreactor. Single Molecules and Nanotechnology, Springer Series inBiophysics, eds Rigler R, Vogel H (Springer, Berlin), Vol 12, pp 251–277.
7. Steffensen MB, Rotem D, Bayley H (2014) Single-molecule analysis of chirality in amulticomponent reaction network. Nat Chem 6(7):603–607.
8. Pulcu GS, Mikhailova E, Choi L-S, Bayley H (2015) Continuous observation of the sto-chastic motion of an individual small-molecule walker. Nat Nanotechnol 10(1):76–83.
9. Shin SH, Luchian T, Cheley S, Braha O, Bayley H (2002) Kinetics of a reversible co-valent-bond-forming reaction observed at the single-molecule level. Angew Chem IntEd Engl 41(19):3707–3709, 3523.
10. Luchian T, Shin SH, Bayley H (2003) Single-molecule covalent chemistry with spatiallyseparated reactants. Angew Chem Int Ed Engl 42(32):3766–3771.
11. Lu S, Li WW, Rotem D, Mikhailova E, Bayley H (2010) A primary hydrogen-deuteriumisotope effect observed at the single-molecule level. Nat Chem 2(11):921–928.
12. Choi LS, Bayley H (2012) S-nitrosothiol chemistry at the single-molecule level. AngewChem Int Ed Engl 51(32):7972–7976.
13. Boersma AJ, Brain KL, Bayley H (2012) Real-time stochastic detection of multipleneurotransmitters with a protein nanopore. ACS Nano 6(6):5304–5308.
14. Hammerstein AF, Shin SH, Bayley H (2010) Single-molecule kinetics of two-step di-valent cation chelation. Angew Chem Int Ed Engl 49(30):5085–5090.
15. Banerjee A, et al. (2010) Molecular bases of cyclodextrin adapter interactions withengineered protein nanopores. Proc Natl Acad Sci USA 107(18):8165–8170.
16. Chin JW (2014) Expanding and reprogramming the genetic code of cells and animals.Annu Rev Biochem 83:379–408.
17. Wang K, et al. (2014) Optimized orthogonal translation of unnatural amino acidsenables spontaneous protein double-labelling and FRET. Nat Chem 6(5):393–403.
18. Sachdeva A, Wang K, Elliott T, Chin JW (2014) Concerted, rapid, quantitative, and site-specific dual labeling of proteins. J Am Chem Soc 136(22):7785–7788.
19. Dawson PE, Muir TW, Clark-Lewis I, Kent SBH (1994) Synthesis of proteins by nativechemical ligation. Science 266(5186):776–779.
20. Kent SBH (2009) Total chemical synthesis of proteins. Chem Soc Rev 38(2):338–351.21. Kent SBH (2013) Bringing the science of proteins into the realm of organic chemistry:
Total chemical synthesis of SEP (synthetic erythropoiesis protein). Angew Chem Int EdEngl 52(46):11988–11996.
22. Hackeng TM, Griffin JH, Dawson PE (1999) Protein synthesis by native chemical li-gation: Expanded scope by using straightforward methodology. Proc Natl Acad SciUSA 96(18):10068–10073.
23. Bang D, Makhatadze GI, Tereshko V, Kossiakoff AA, Kent SB (2005) Total chemicalsynthesis and X-ray crystal structure of a protein diastereomer: [D-Gln 35]ubiquitin.Angew Chem Int Ed Engl 44(25):3852–3856.
24. Muir TW (2003) Semisynthesis of proteins by expressed protein ligation. Annu RevBiochem 72:249–289.
25. Muralidharan V, Muir TW (2006) Protein ligation: An enabling technology for thebiophysical analysis of proteins. Nat Methods 3(6):429–438.
26. Shah NH, Muir TW (2014) Inteins: Nature’s gift to protein chemists. Chem Sci (Camb)5(1):446–461.
27. Olschewski D, Becker CF (2008) Chemical synthesis and semisynthesis of membraneproteins. Mol Biosyst 4(7):733–740.
28. Brenzel S, Cebi M, Reiss P, Koert U, Mootz HD (2009) Expanding the scope of proteintrans-splicing to fragment ligation of an integral membrane protein: Towardsmodulationof porin-based ion channels by chemical modification. ChemBioChem 10(6):983–986.
29. Komarov AG, Linn KM, Devereaux JJ, Valiyaveetil FI (2009) Modular strategy for thesemisynthesis of a K+ channel: Investigating interactions of the pore helix. ACS ChemBiol 4(12):1029–1038.
30. Grosse W, Essen LO, Koert U (2011) Strategies and perspectives in ion-channel engi-neering. ChemBioChem 12(6):830–839.
31. Devaraneni PK, et al. (2013) Semisynthetic K+ channels show that the constrictedconformation of the selectivity filter is not the C-type inactivated state. Proc NatlAcad Sci USA 110(39):15698–15703.
32. Bayley H, Cheley S, Harrington L, Syeda R (2009) Wrestling with native chemical li-gation. ACS Chem Biol 4(12):983–985.
33. Gentle IE, De Souza DP, Baca M (2004) Direct production of proteins with N-terminalcysteine for site-specific conjugation. Bioconjug Chem 15(3):658–663.
34. Blanco-Canosa JB, Dawson PE (2008) An efficient Fmoc-SPPS approach for the gen-eration of thioester peptide precursors for use in native chemical ligation. AngewChem Int Ed Engl 47(36):6851–6855.
35. Song L, et al. (1996) Structure of staphylococcal alpha-hemolysin, a heptamerictransmembrane pore. Science 274(5294):1859–1866.
36. Henricus M, Bayley H (2010) Unfolding and refolding of monomeric and heptamericα-hemolysin. Master of Science thesis (University of Oxford, Oxford).
37. Walker B, Krishnasastry M, Zorn L, Kasianowicz J, Bayley H (1992) Functional ex-pression of the alpha-hemolysin of Staphylococcus aureus in intact Escherichia coliand in cell lysates. Deletion of five C-terminal amino acids selectively impairs hemo-lytic activity. J Biol Chem 267(15):10902–10909.
38. Mantri S, Sapra KT, Cheley S, Sharp TH, Bayley H (2013) An engineered dimeric pro-tein pore that spans adjacent lipid bilayers. Nat Commun 4:1725.
39. Walker B, Braha O, Cheley S, Bayley H (1995) An intermediate in the assembly of a pore-forming protein trapped with a genetically-engineered switch. Chem Biol 2(2):99–105.
40. Cheley S, et al. (1997) Spontaneous oligomerization of a staphylococcal alpha-hemolysinconformationally constrained by removal of residues that form the transmembrane beta-barrel. Protein Eng 10(12):1433–1443.
41. Olson R, Nariya H, Yokota K, Kamio Y, Gouaux E (1999) Crystal structure of staphy-lococcal LukF delineates conformational changes accompanying formation of atransmembrane channel. Nat Struct Biol 6(2):134–140.
42. Gu LQ, Braha O, Conlan S, Cheley S, Bayley H (1999) Stochastic sensing of organic an-alytes by a pore-forming protein containing a molecular adapter. Nature 398(6729):686–690.
43. Gu LQ, Bayley H (2000) Interaction of the noncovalent molecular adapter, beta-cyclodextrin,with the staphylococcal alpha-hemolysin pore. Biophys J 79(4):1967–1975.
44. Kolb HC, Finn MG, Sharpless KB (2001) Click chemistry: Diverse chemical function froma few good reactions. Angew Chem Int Ed Engl 40(11):2004–2021.
45. Iha RK, et al. (2009) Applications of orthogonal “click” chemistries in the synthesis offunctional soft materials. Chem Rev 109(11):5620–5686.
46. Debets MF, et al. (2011) Bioconjugation with strained alkenes and alkynes. Acc ChemRes 44(9):805–815.
47. Kolb HC, Sharpless KB (2003) The growing impact of click chemistry on drug discovery.Drug Discov Today 8(24):1128–1137.
48. Movileanu L, Cheley S, Bayley H (2003) Partitioning of individual flexible polymersinto a nanoscopic protein pore. Biophys J 85(2):897–910.
49. Worrell BT, Malik JA, Fokin VV (2013) Direct evidence of a dinuclear copper in-termediate in Cu(I)-catalyzed azide-alkyne cycloadditions. Science 340(6131):457–460.
50. Lang K, Chin JW (2014) Bioorthogonal reactions for labeling proteins. ACS Chem Biol9(1):16–20.
51. Sadeghi SM, Ferreira CMH, Vandenbogaerde S, Soares HMVM (2014) Graphic dataanalysis and complex formation curves as modeling and optimization tools forcharacterization of Cu–(buffer)x–(OH)y systems involving BTP or BES in aqueous so-lution. J Coord Chem 68(5):777–793.
52. Movileanu L, Bayley H (2001) Partitioning of a polymer into a nanoscopic protein poreobeys a simple scaling law. Proc Natl Acad Sci USA 98(18):10137–10141.
53. Maglia G, et al. (2009) DNA strands from denatured duplexes are translocatedthrough engineered protein nanopores at alkaline pH. Nano Lett 9(11):3831–3836.
54. Franceschini L, Mikhailova E, Bayley H, Maglia G (2012) Nucleobase recognition atalkaline pH and apparent pKa of single DNA bases immobilised within a biologicalnanopore. Chem Commun (Camb) 48(10):1520–1522.
55. Kang XF, Gu LQ, Cheley S, Bayley H (2005) Single protein pores containing molecularadapters at high temperatures. Angew Chem Int Ed Engl 44(10):1495–1499.
56. Bang D, Chopra N, Kent SB (2004) Total chemical synthesis of crambin. J Am Chem Soc126(5):1377–1383.
Lee and Bayley PNAS | November 10, 2015 | vol. 112 | no. 45 | 13773
CHEM
ISTR
YBIOCH
EMISTR
Y
Dow
nloa
ded
by g
uest
on
Janu
ary
9, 2
020


















