
Semiario 4 bq
58
Lisozima, Ribonucleasa A, Carboxipeptidas a A, Serinproteasas y Bioquímica del ejercicio ESTRATÉGIAS CATALÍTICAS DE LAS ENZIMAS Y MECANISMO DE ACCIÓN Universidad Nacional de Trujillo Facultad de Farmacia y Bioquímica Bioquímica I Seminario IV Grupo N° 06
-
Upload
adriana-samillan-moya -
Category
Documents
-
view
153 -
download
2
Transcript of Semiario 4 bq

Lisozima, Ribonucleasa A,
Carboxipeptidasa A,
Serinproteasas y Bioquímica del
ejercicio
ESTRATÉGIAS CATALÍTICAS DE LAS
ENZIMAS Y MECANISMO DE ACCIÓN
Universidad Nacional de TrujilloFacultad de Farmacia y Bioquímica
Bioquímica I
Seminario IVGrupo N° 06

Es la facilitación de la hidrolisis de una
molécula biológica, por la acción de una
enzima. Un ejemplo muy interesante es la
hidrolisis del glucógeno por la alfa amilasa,
esta actúa sobre los enlaces glucosídicos
alfa (1-4), y no afecta los enlaces alfa (1-
6), dando lugar a la ruptura de la molécula
de glucógeno en productos como: glucosa,
maltosa e isomaltosa.
CATÁLISIS ENZIMATICA
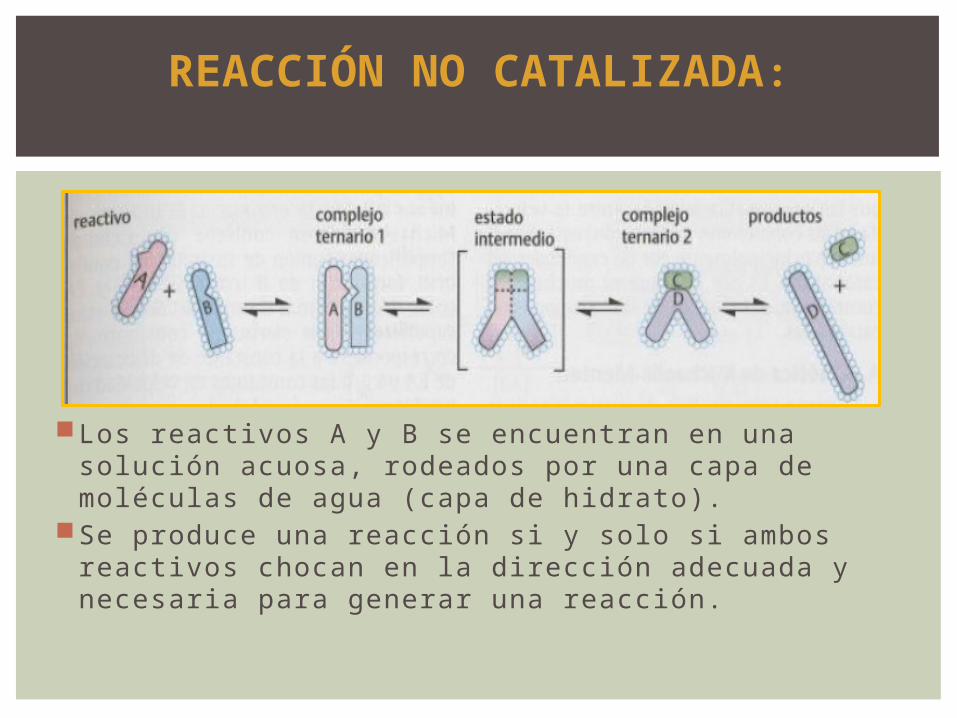
REACCIÓN NO CATALIZADA:
Los reactivos A y B se encuentran en una solución acuosa, rodeados por una capa de moléculas de agua (capa de hidrato).
Se produce una reacción si y solo si ambos reactivos chocan en la dirección adecuada y necesaria para generar una reacción.
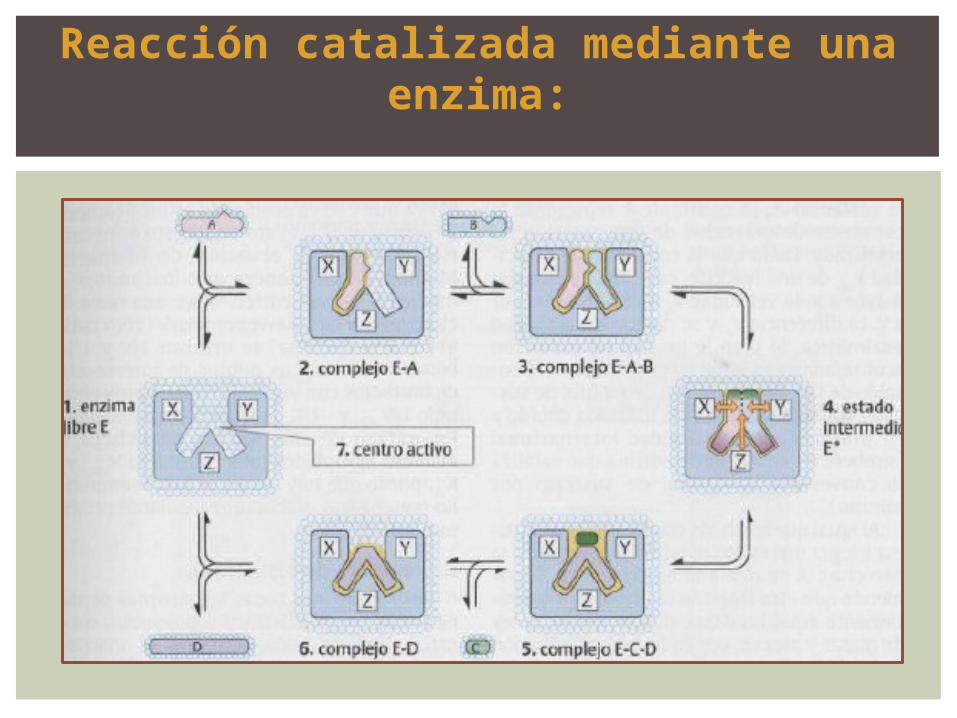
Reacción catalizada mediante una enzima:

el posicionamiento y la orientación de los sustratos
incrementan drásticamente la probabilidad de que se
generen complejos A-B productivos.
Durante la unión de sustrato se eliminan asimismo sus capas
de hidrato. Al expulsar las moléculas de agua se modifican
completamente las condiciones dentro del centro activo de
la enzima durante la catálisis.
Otro factor importante es la estabilización del estado
intermedio a través de la interacción entre los aminoácidos
y los restos de proteína y sustratos
Las enzimas pueden unir los reactivos (específ icamente sus sustratos)
en el centro activo, proceso en el cual los sustratos se or ientan en una
posición que les permite alcanzar el estado intermedio.

ESTRATEGIAS CATALITICAS DE LAS
ENZIMAS

La catálisis enzimática comienza con la unión del sustrato.
Las interacciones solamente se forma cuando el sustrato se encuentra en el estado de transición.
Sólo el sustrato correcto puede participar en la mayor parte de las interacciones con la enzima (especificidad)
GENERALIDADES

• LIZOSIMAEs una enzima (glucosidasa) que destruye
paredes celulares bacterianas por
hidrólisis de los enlaces glucosídico ß (1-
4) de ácido n-acetilmurámico (NAM o
MurNAc) a n-acetilglucosamina (NAC o
GlcNAc) en los peptidoglucanos de la
pared celular .De modo similar, hidroliza
los enlaces ß (1-4) de poli (NAG) quitina,
un componente de la pared celular de la
mayoría de los hongos, además de un
componente importante de los
exoesqueletos de los insectos y los
crustáceos.


La forma de la molécula de proteína
es casi elipsoidal, y sus dimensiones
son 30x30x45A .Su característica
más notable es una hendidura
prominente, el sitio de fijación del
sustrato, que atraviesa una cara de
la molécula. La cadena poli peptídica
forma cinco segmentos helicoidales,
además una hoja B antiparalelas
tricatenaria que comprende una
pared de la hendidura de fijación.
Estructura de enzima:

• RIBONUCLEASA A
Las ribonucleasas están divididas en tres grandes
familias: la superfamilia RNasa A, la familia T1 y la
familia T2. La superfamilia Ribonucleasa A está
constituida por las RNasas de mamíferos y otros
vertebrados como aves, reptiles y anfibios
(Soochin et al., 2005). Este grupo de proteínas
presenta altas tasas de duplicación génica y pérdida
de genes de lo que ha resultado un número variable
de genes en diferentes especies (Soochin et
al., 2005). En el ser humano, los genes que las
codifican están ubicados en el brazo largo del
cromosoma 14




• CARBOXIPEPTIDASA A

Es una enzima proteolítica que cataliza la ruptura del enlace pepitico, denominada, proteinasas.
Es una enzima digestiva secretada en el jugo pancreático que hidroliza el enlace peptídico carboxiterminal en las cadenas polipeptídicas
La hidrólisis se produce más fácilmente en el residuo carboxiterminal que tiene una cadena lateral aromática o una cadena alifática. Esta enzima contiene regiones α y la hoja plegada β
GENERALIDADES

• constituye una exopeptidasa ya que cataliza la remoción del aminoácido situado en el extremo carbonilo del sustrato, siendo específica para aminoácidos voluminosos
SEGÚN SU FUNCION
• cataliza la hidrolisis del enlace peptídico carboxiloterminal de la cadena polipeptídica. con la ayuda de metales, generalmente Zn+2, unidos a su sitio activo, este ion actúa como una catalizador metálico para las reacciones hidrolíticas.
HIDRÓLISIS
• como intermediario de la hidrólisis de una forma muy similar a la acción de la serina en la quimiotripsina.
ESTABILIZACION

ESTRUCTURA DE SITIO ACTIVO DE LA CARBOXIPEPTIDASA A
Sitio Activo
• consta de un surco poco profundo en la superficie de la enzima que conduce a una bolsa profunda, forrada con cadenas laterales alifáticas y polares
Cadena Lateral
• La cadena lateral de este residuo es conformacionalmente muy móvil
Cadena Lateral Aromática
• se fija en la bolsa de fijación• el ion carboxilato del C-terminal forma un
enlace salino con la Arg-145• el oxígeno carbonílico del enlace
escindible se constituye en el cuarto ligando del ion zinc.
La Cadena Lateral Fenólica
• puede rotar alrededor del enlace Cα-Cβ y un 80%
• su densidad electrónica en el mapa de la estructura cristalina se encuentra en una orientación en la superficie de la molécula apuntando hacia la solución

• SERINPROTEASASEstas enzimas emplean las siguientes estrategias:
Catálisis covalente. El centro activo contiene un
grupo reactivo, habitualmente un nucleófilo
potente, un residuo de serina que en el
transcurso de la catálisis llega a ser modificado
covalentemente de forma temporal.
Catálisis general acido-base. Una molécula
diferente al agua desempeña el papel de dador
o aceptor de un protón. Ej. La Quimotripsina
utiliza un residuo de histidina como un
catalizador básico para incrementar el poder
nucleófilo de la serina.

La Quimotripsina rompe selectivamente los enlaces peptídicos
contiguos al extremo carboxílico de aminoácidos hidrofóbicos
voluminosos, como triptófano, tirosina, fenilalanina y metionina.
La tripsina corta en el lado carboxilato de los residuos básicos
como la lisina o la arginina.
Reacción de la Quimotripsina
Quimotripsina
Quimotripsina

MECANISMO DE ACCION ENZIMATICA DE LAS LISOZIMAS

Los únicos grupos funcionales en la vecindad inmediata del centro react ivo de la
l isoz ima que t ienen las propiedades catal í t icas requer idas son las cadenas
laterales de Glu 35 y Asp 52.estas cadenas laterales ,d ispuestas a ambos lados
del enlace glucosídico que se desea escindir , t ienen entornos con di ferencias
marcadas.asp 52 esta rodeado por var ios residuos polares conservados, con los
que forma una red compleja unidas por puentes de hidrogeno.
En consecuencia se prevé que Asp 52 t iene un pk normal , esto es , no deber ía estar
protonado, y por lo tanto tener carga negat iva en el rango de pH de 3 a 8 para el
cual la l i soz ima t iene act iv idad enzimát ica. Por ende, Asp 52 puede actuar en la
estabi l i zación electrostát ica de un ion oxonio. Por e l contrar io , e l grupo carboxi lo
de Glu 35 se anida en un bols i l lo con predominio no polar , donde es probable que
se mantenga protonado con valores de pH inusualmente elevados de los grupos
carboxi lo. Por lo tanto, este res iduo puede actuar como catal izador acido.
Glu 35 y Asp 52 son los residuos catalíticos de la lisozima:

LA REACCIÓN DE LA LISOZIMA PROCEDE POR MEDIO DE UN INTERMEDIARIO COVALENTE:
Según Phi l l ips, en el mecanismo de l isozima se incluye la catál is is acida por
Glu 35 y la estabi l ización de un intermediario de ion oxonio por el grupo
carboxi lato de Asp 52. La formación de 1 enlace covalente entre el O2 de
carboxi lato de Asp 52 y C1 del residuo D del sustrato no parecía posible,
debido a que estos átomos están separados por -3 Å(la longitud del enlace
covalente C-O es de -1,4 Å ) .
En investigaciones más reciente se reveló que, de hecho, la reacción de
l isozima t iene lugar a través de un intermediario covalente. La reacción
comienza cuando la l isozima se une a una pared celular bacteriana, por unión
a una unidad de hexasacárido. Esto distorsiona al residuo D para adoptar la
conformación de semisi l la. El resto del mecanismo catal í t ico se produce como
se sigue.

1.-Glu 35 transfiere su protón a O1 del anil lo D (un ejemplo de catál isis ácida).este paso comprende un estado de transición de ion oxonio estabil izado por resonancia, cuya formación se faci l ita por la tensión en el anil lo D que lo distorsiona para dar la forma de semisi l la en la que C1, C2, C5 y O5 son coplanares (catálisis por el enlace preferencial del estado de transición)
• 2.-El grupo carboxilato de Asp 52 que actúa como un nucleófilo ataca el C1 ahora reducido del anillo D para formar un intermediario glucosilo-emzima (catálisis covalente)
• 3.-Después de la extracción del grupo que se elimina, que contiene el anillo E, entra agua en el sitio activo para hidrolizar el enlace covalente y generar los grupos del sitio activo con la ayuda de Glu 35(catálisis general básica).
• 4.- La enzima luego libera el producto del anillo D, para completar la reacción catalítica. En principio la enzima se une a la molécula de glúcido en conformación "silla" (a), que luego pasa a otra forma (b) al formar el complejo ES. En el estado de transición, actúa un resto glutámico del Sitio Activo de la enzima, que genera un carbocatión en el anillo D, provocando el colapso del enlace.
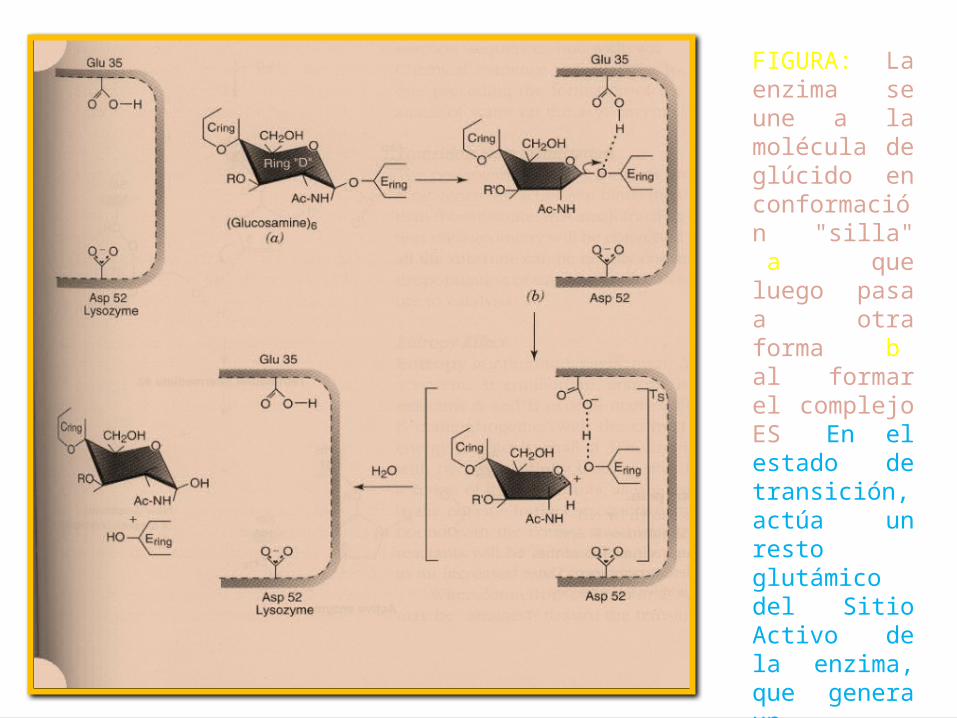
FIGURA: La enzima se une a la molécula de glúcido en conformación "silla" (a), que luego pasa a otra forma (b) al formar el complejo ES. En el estado de transición, actúa un resto glutámico del Sitio Activo de la enzima, que genera un carbocatión en el anillo D, provocando el colapso del enlace.

MECANISMO DE ACCIÓN ENZIMATICA
DE LA RIBONUCLEASA A

La acción catalítica de la Ribonucleasa pancreática se debe a tres aminoácidos absolutamente conservados: His 12, His 119 y Lys 41. No está aún bien determinado el papel de esta última, pero la acción de los dos residuos de histidina ilustra cómo este aminoácido se comporta a la vez como ácido y como base. Así, en el estado basal de la enzima, el Centro Activo consta de esos tres aminoácidos, de tal manera que la His 12 aparece como base y la His 119 como ácido (protonada, imidazolio)
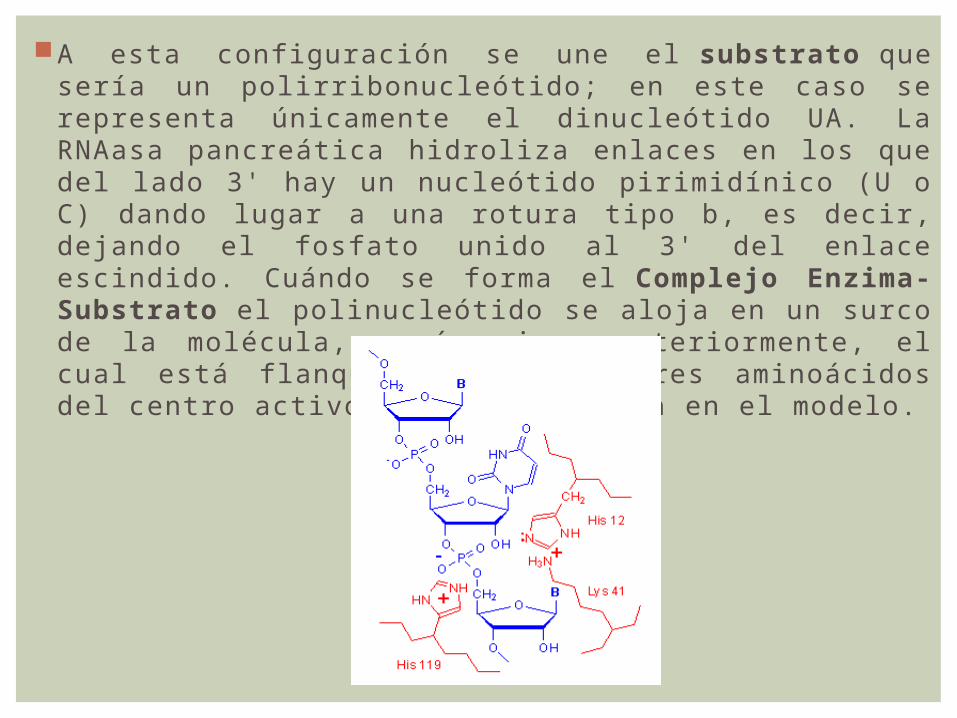
A esta configuración se une el substrato que sería un polirribonucleótido; en este caso se representa únicamente el dinucleótido UA. La RNAasa pancreática hidroliza enlaces en los que del lado 3' hay un nucleótido pirimidínico (U o C) dando lugar a una rotura tipo b, es decir, dejando el fosfato unido al 3' del enlace escindido. Cuándo se forma el Complejo Enzima-Substrato el polinucleótido se aloja en un surco de la molécula, según vimos anteriormente, el cual está flanqueado por los tres aminoácidos del centro activo, según se indica en el modelo.

A continuación la His 12 captura un protón del grupo 2'OH del nucleótido pirimidínico, el cual lleva a cabo un ataque nucleofílico sobre el substrato de manera que se forma el Estado de transición inestable (con P pentavalente), al tiempo que la His 119 cede su protón al oxígeno en 5' del enlace atacado; el estado de ionización de las histidinas es justo el contrario que el visto en el estado basal de la enzima.

Este estado de transición se resuelve con la Liberación del primer producto (3'-AMP) quedando unido a la enzima el nucleótido cícl ico 3',5' UMP.
Entra entonces al centro activo una molécula de agua , que es el segundo substrato de la reacción.

La histidina 119 sustrae uno de los dos protones del agua, que queda convertida en un ion hidroxilo,
que a su vez ataca al fosfato cíclico, dando lugar al Estado de transición 2.

Éste se resuelve rompiéndose el enlace entre el fosfato y el oxígeno que sustituye al carbono 2' de la ribosa, el cual recibe a su vez el protón de la histidina 12, para dar lugar a la liberación del segundo producto, que es el nucleótido 3',5' uridin bisfosfato.

MECANISMO DE ACCIÓN ENZIMATICA
DE LA CARBOXIPEPTIDASA
A

La carboxipeptidasa A cataliza la hidrolisis del enlace peptídico carboxilo terminal de la cadena polipeptídica.
La hidrolisis es más fácil si el residuo carboxílico terminal tiene una cadena lateral aromática o una
cadena lateral alifática voluminosa
Hay una adaptación inducida del centro activo cuando se une el substrato. La enzima es una cadena polipeptídica de 307 aminoácidos.
Tiene un ion Zn2+ fuertemente unido, que es esencial para la actividad enzimática y que está localizado en un surco de la superficie de la molécula donde coordina a dos cadenas laterales de histidina, a una cadena lateral de glutamato y a una molécula de agua

La cadena lateral del residuo terminal del substrato se acomoda en un gran bolsillo cerca de un ion Zn2+.
La molécula de agua altamente polarizada transfiere un protón al grupo NH escindiendo el enlace. Alternativamente, la molécula de agua puede atacar directamente el carbono carbonílico del enlace.
La polarización del grupo carbonílico por el Zn2+ facilita el ataque, sea por el glutamato 270 o la molécula activada de agua. La inducción de un dipolo es impulsada por el ambiente no polar del ion Zn, el que incrementa su carga.
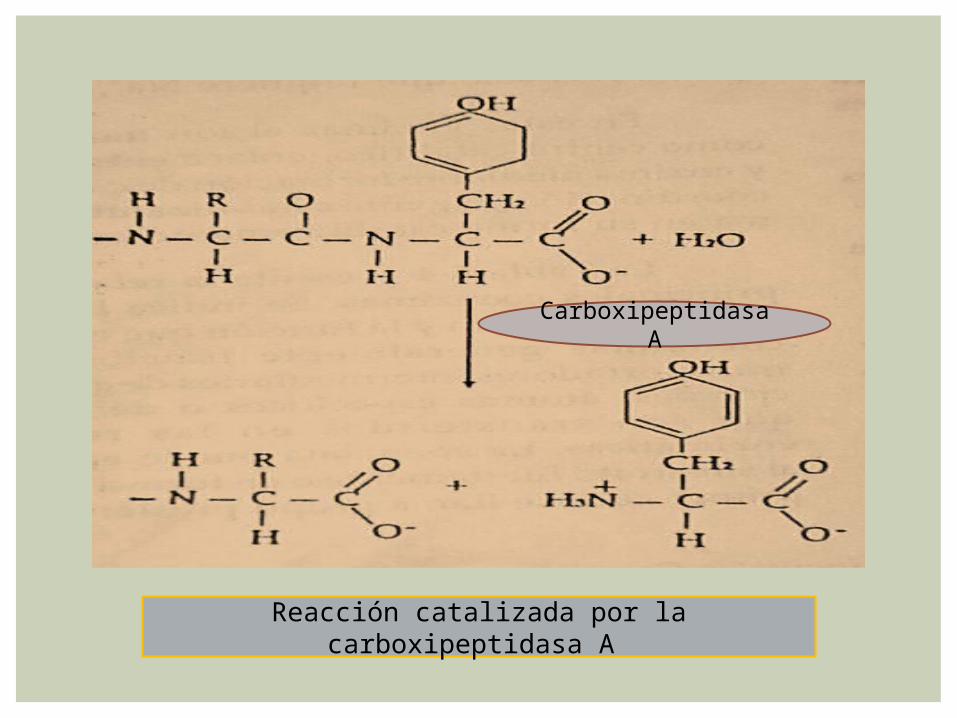
Reacción catalizada por la carboxipeptidasa A
Carboxipeptidasa A

COMPLEJO: EZIMA -
SUSTRATO
INTERMEDIARIO ANHIDRO
CARBOXILICOPRODUCTOS

MECANISMO DE ACCIÓN ENZIMÁTICA
DE LA SERINPROTEASAS A

Las Serinproteasas son hidrolasas que degradan enlaces peptídicos de péptidos y proteínas y que poseen en su centro activo un aminoácido de serina esencial para la catálisis enzimática
la tripsina es específica para un residuo con carga positiva y la elastasa lo es para un residuo neutro pequeño. En conjunto formaron un equipo digestivo muy potente.
La quimiotripsina, tripsina y elastasa son enzimas digestivas sintetizadas por las células de los acinos pancreáticos y segregados por el conducto pancreático en el interior del duodeno (asa superior del intestino delgado).

Las mediciones cinéticas realizadas
por Brian Hartley de la hidrólisis
catalizada por la quimiotripsina del p-
nitrofenilacetato indicaron que la
reacción se produce en dos pasos:
La enzima reacciona con rapidez
con el p-nitrofenilacetato para
liberar el ión p-nitrofenolato que
forma un intermediario acilo-
enzima covalente.
El intermediario acilo-enzima
covalente se hidroliza despacio
para liberar al acetato.

IDENTIFICACIÓN DE RESIDUOS CATALÍTICOS
• Ser 195. Una prueba diagnóstica para la presencia de la ser activa de las serina proteasa es su reacción con diisopropilfosfofluoridato (DIPF): Que inactiva en forma irreversible a la enzima.
• His 57. Un segundo residuo importante por su actividad catalítica se descubrió por medio de la marcación por afinidad, para formar un enlace covalente estable con un grupo susceptible cercano.

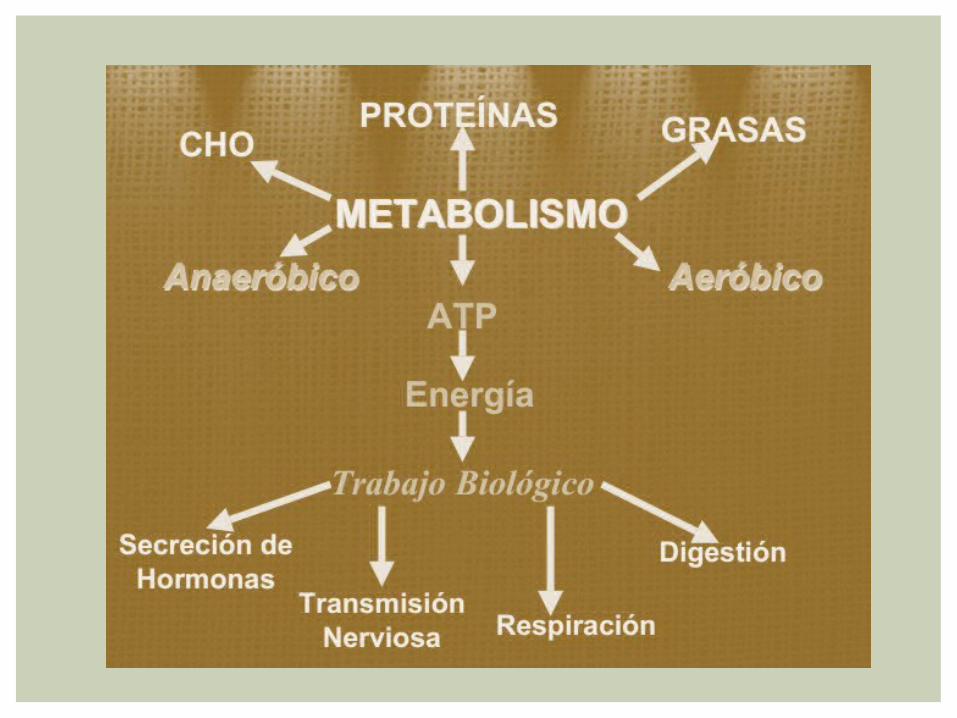
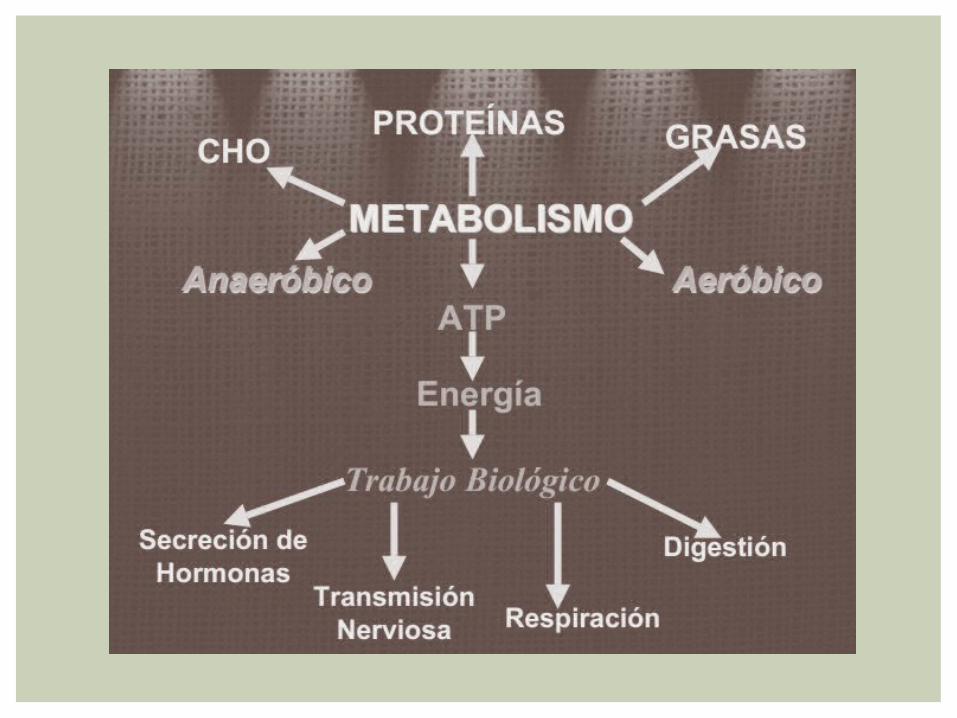
BIOQUÍMICA DEL EJERCICIO
El funcionamiento del organismo humano depende de una variedad de procesos bioquímicos que en conjunto representan el metabolismo de las células corporales
Para el atleta, los procesos liberadores y de síntesis de energía que constituyen el metabolismo facilita la ejecutoria deportiva con dietas efectivas
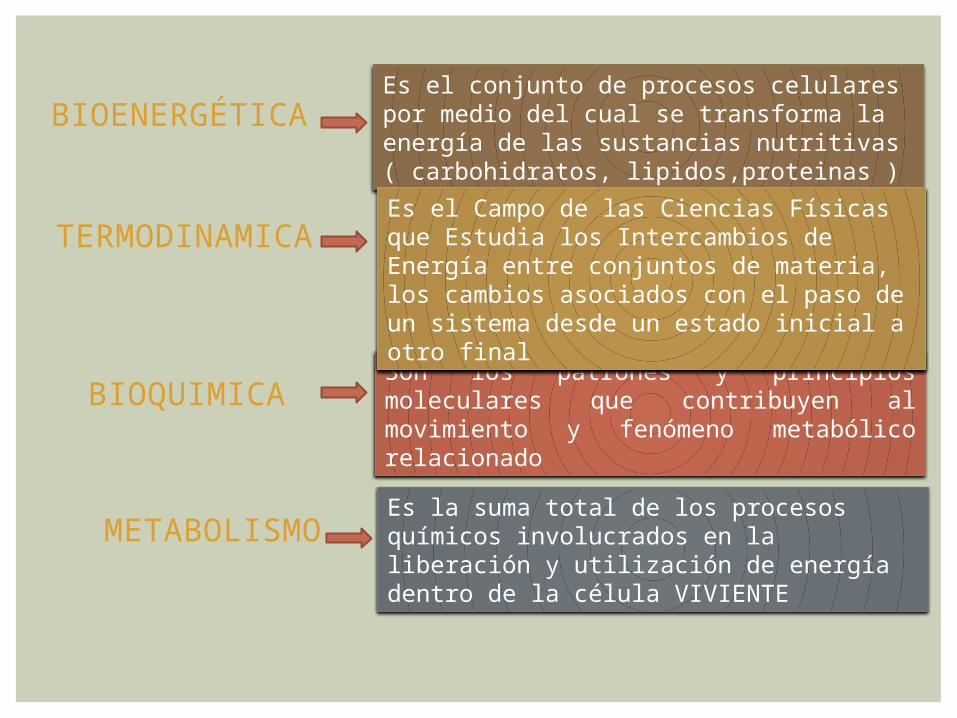
BIOENERGÉTICAEs el conjunto de procesos celulares por medio del cual se transforma la energía de las sustancias nutritivas ( carbohidratos, lipidos,proteinas )
BIOQUIMICA Son los patrones y principios moleculares que contribuyen al movimiento y fenómeno metabólico relacionado
TERMODINAMICA Es el Campo de las Ciencias Físicas que Estudia los Intercambios de Energía entre conjuntos de materia, los cambios asociados con el paso de un sistema desde un estado inicial a otro final
METABOLISMOEs la suma total de los procesos químicos involucrados en la liberación y utilización de energía dentro de la célula VIVIENTE
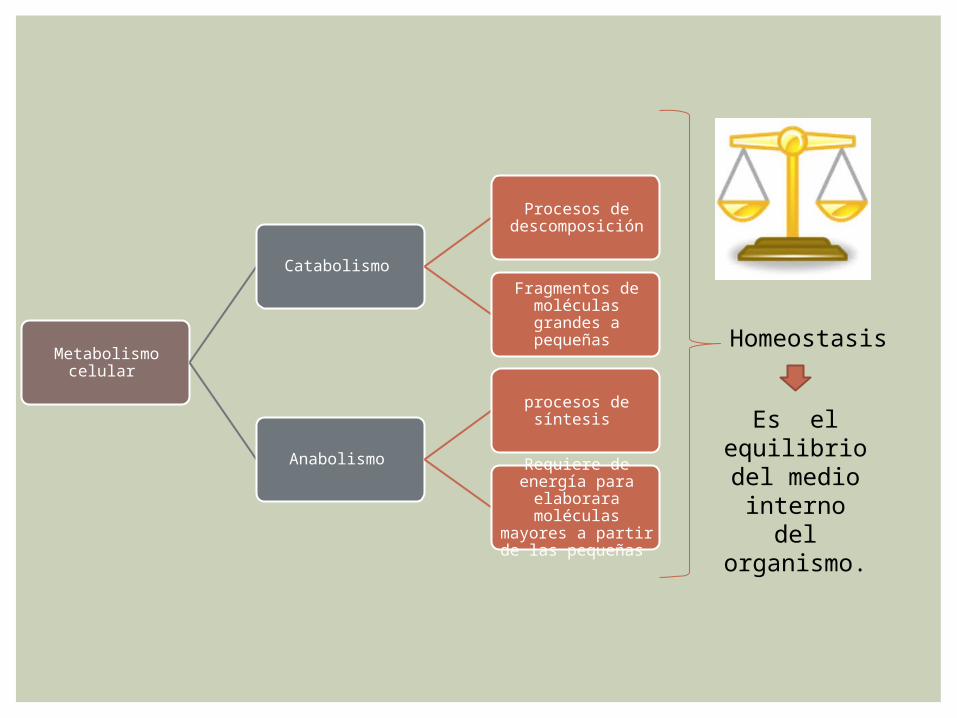
Metabolismo celular
Catabolismo
Procesos de descomposición
Fragmentos de moléculas grandes
a pequeñas
Anabolismo
procesos de síntesis
Requiere de energía para elaborara
moléculas mayores a partir de las
pequeñas
Homeostasis
Es el equilibrio del medio interno del
organismo.

Energía
potencial eléctrica Nuclear Radiante solar cineteca
Capacidad para efectuar
trabajo
Clases de energía
Química Osmótica Mecánica
Formas

Ciclo biológico de la energía

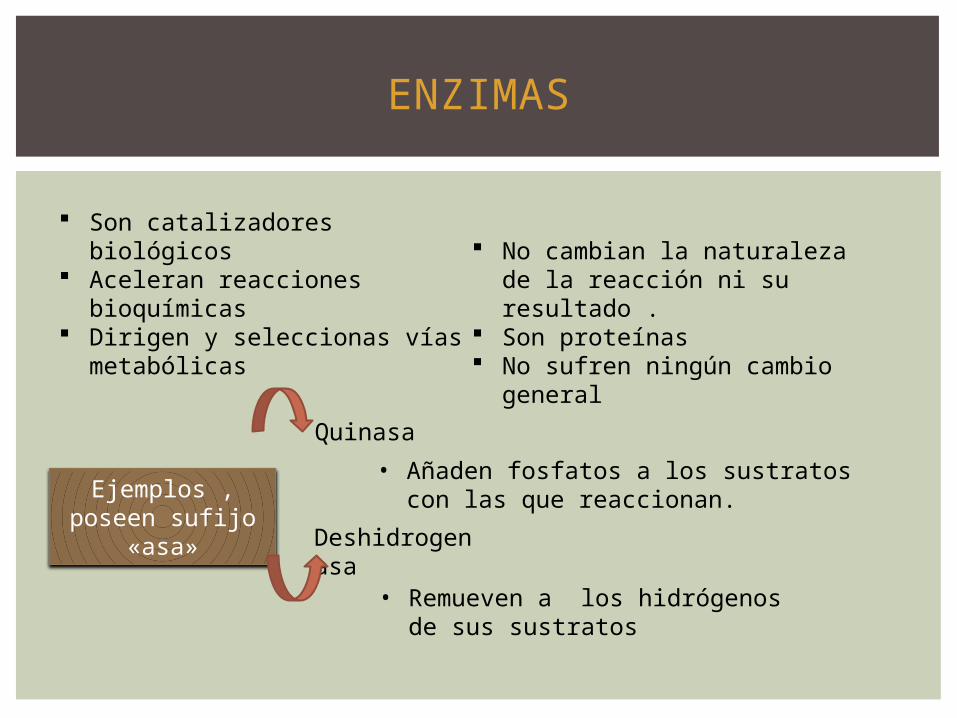
ENZIMAS
Son catalizadores biológicos Aceleran reacciones
bioquímicas Dirigen y seleccionas vías
metabólicas
No cambian la naturaleza de la reacción ni su resultado .
Son proteínas No sufren ningún cambio
general
Ejemplos , poseen sufijo
«asa»
Quinasa
Deshidrogenasa
• Añaden fosfatos a los sustratos con las que reaccionan.
• Remueven a los hidrógenos de sus sustratos


Combustibles Metabólicos para el Ejercicio
Carbohidratos Estructu
ra química
• Átomos de C, H,y O
Función • Provee energía • 4 Kcal / g de R-CHO
Clasificación
• Monosacárido: glucosa ( sangre), galactosa( Glandulas
mamarias),fructosa (frutas y miel de abeja)
• Disacárido: sacarosa( caña de
azúcar),lactosa(leche),maltosa( digestión)• Polisacárido: almidón( tubérculos), celulosa (fibra),glucógeno( reservorio)

Lípidos
Características
No son solubles en agua
Función
9 Kcal de energía por gramo de lípido
Tipos
-Simples: triglicéridos-Compuestas: fosfolípidos y lipoproteínas-Derivadas: colesterol
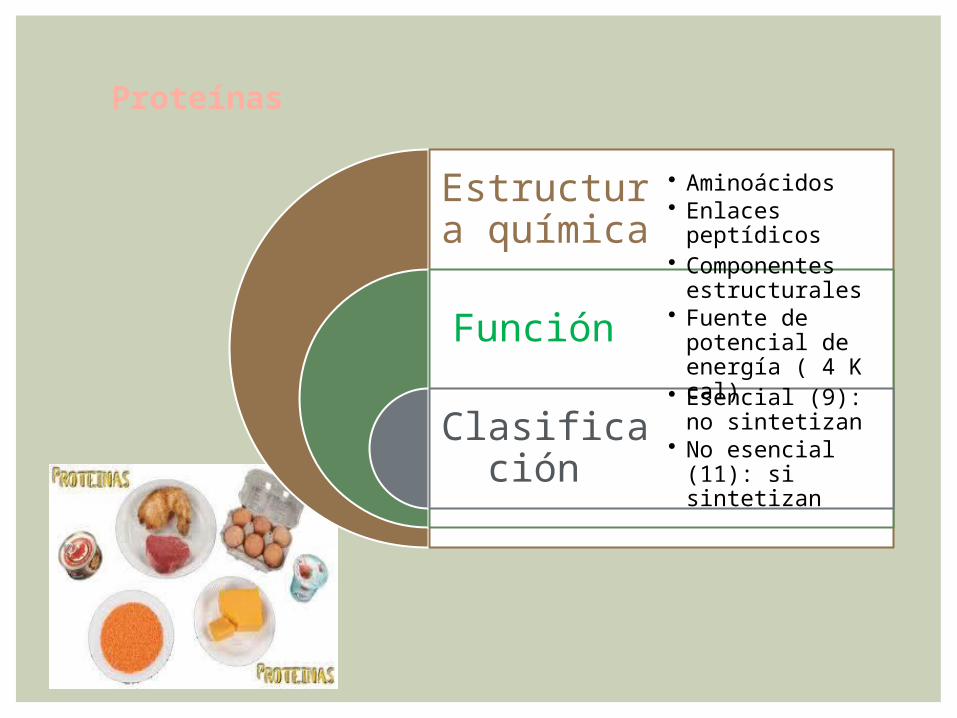
Estructura química
Función
Clasificación
• Aminoácidos • Enlaces
peptídicos
• Componentes estructurales
• Fuente de potencial de energía ( 4 K cal)
• Esencial (9): no sintetizan
• No esencial (11): si sintetizan
Proteínas

El ATP

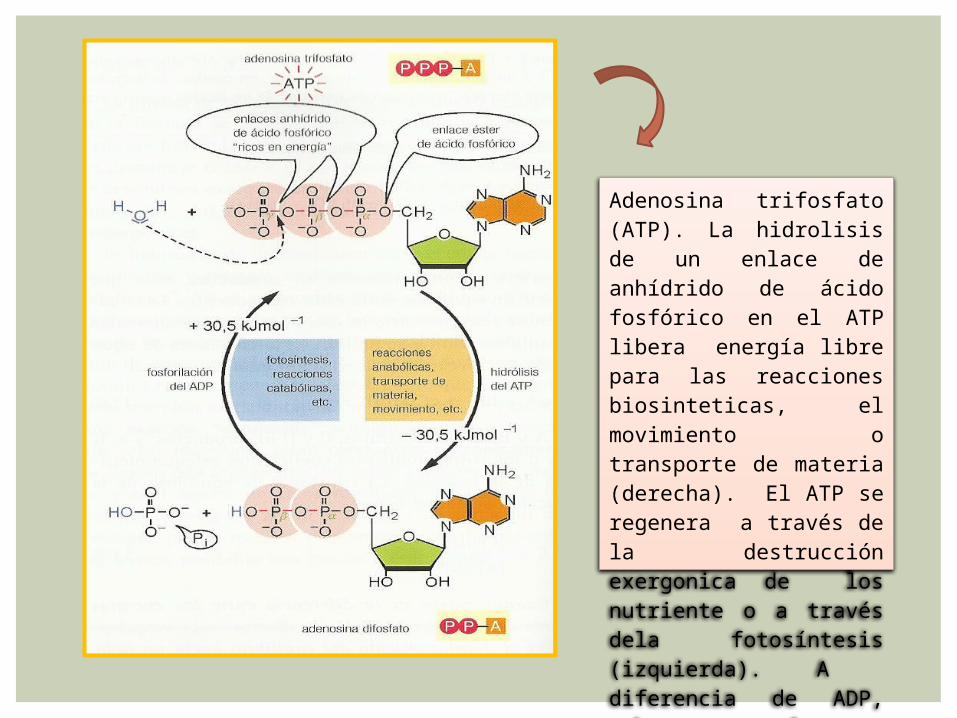
Adenosina trifosfato (ATP). La hidrolisis de un enlace de anhídrido de ácido fosfórico en el ATP libera energía libre para las reacciones biosinteticas, el movimiento o transporte de materia (derecha). El ATP se regenera a través de la destrucción exergonica de los nutriente o a través dela fotosíntesis (izquierda). A diferencia de ADP, pobre en energía.

GRACIAS
!!!












![02 Feinhals - IAEA and EC Regulations for Clearance of Materials … · 2019. 4. 2. · [Bq] Specific activity [Bq/g] [Bq/cm 2] Solid sub-stances, liquids, with the exception of column](https://static.fdocuments.in/doc/165x107/602ee7a9f8f00166e4555cae/02-feinhals-iaea-and-ec-regulations-for-clearance-of-materials-2019-4-2-bq.jpg)





