
Selective activation of heteromeric SK channels contribute to action potential repolarization in...
8
Selective activation of heteromeric SK channels contribute to action potential repolarization in mouse atrial myocytes Jane M Hancock BSc PhD, Kate L Weatherall BSc PhD, Andrew F James BSc PhD, Jules C Hancox BSc PhD, Neil V Marrion BSc PhD School of Physiology & Pharmacology, University of Bristol, Bristol, BS8 1TD, UK. Supporting Information Experimental Procedures Constructs and cell culture. Channels were transiently expressed in HEK293 cells, with cells maintained as described previously 1 . Cells were transiently transfected separately with the following constructs: mouse Na V 1.5 (accession number AJ271477), rat Ca V 1.2 (accession number AF394940), rat Ca V 1.3 (accession number AF370009), human K V 1.5 (accession number NG_012198.1), K V 2.1 (accession number NP_037318.1) and mouse K V 4.2 (accession number NM019697) and hSK3_ex4 2 . Rat Ca V 1.2 and 1.3 were co-transfected with rat Ca V 2A (accession number M80545), rat Ca V 21 (accession number AF286488) and enhanced green fluorescent protein (eGFP) (channel constructs were used in an equal ratio to a total plasmid concentration of 8 g per 25 cm 3 flask, eGFP plasmid was used at a ratio of 1:20) using lipofectamine 2000, with cells transfected in 25 cm flasks for 24 hours before being plated in 35 mm dishes. Transient transfection of all other constructs was achieved using polyethylenimine (Alfa Aesar, Inc.) by combining channel plasmid DNA with enhanced eGFP DNA in a ratio of 1:5 to 1:10 (maximal plasmid content: 1 μg/35 mm dish). In all cases, cells were used 24-48 hr after transfection. Electrophysiology. Recording of Na V -mediated, K V -mediated or mouse atrial myocyte currents was achieved by bathing cells in a solution of composition: NaCl (138 mM), KCl (4 mM), HEPES acid (10 mM), D-glucose (10 mM), MgCl 2 (1.2 mM), CaCl 2 (2.5 mM) (pH 7.4). Fire-polished electrodes (3-5 M) pulled from borosilicate glass contained: K Aspartate (130 mM), KCl (20 mM), MgCl 2 (1.5 mM), EGTA (0.01 mM), MgATP (5 mM), HEPES Na (5 mM), NaOH (3 mM) (pH 7.4). A junction potential of +10 mV was calculated but not compensated for. Ca V currents were recorded from cells bathed in a solution of composition: TEACl (140 mM), HEPES (10 mM), D-glucose (10 mM), MgCl 2 (1 mM) and CaCl 2 (5 mM) (pH 7.4) and using electrodes were filled with solution of composition: CsMeSO 4 (120 mM), TEACl (20 mM), HEPES Na (10 mM), EGTA (5 mM), Mg 2 ATP (4 mM), MgCl 2 (1 mM) (pH 7.3). A junction potential of +14 mV was calculated but not compensated for. Cells were voltage-clamped at -80 mV using an Axopatch 200A amplifier (Molecular Devices, CA, USA) and the voltage stepped to positive levels in 10 mV increments. Capacitance and series resistance compensation (>90%) was used throughout, with currents filtered at 1 kHz (8- pole low pass Bessel filter, Frequency Devices, CT, USA) and sampled at 5-10 kHz using Pulse (HEKA). All recombinant channel currents were leak- subtracted using a P/4 protocol from a holding potential of -80 mV. Recording of hSK3_ex4-mediated currents were obtained from outside-patches bathed in a solution of composition: KCl (127 mM), HEPES Na (10 mM), EGTA (10 mM), CaCl 2 (6.19 mM, calculated free [Ca 2+ ] 60 nM), MgCl 2 (1.44 mM,
-
Upload
bartholomew-young -
Category
Documents
-
view
218 -
download
0
Transcript of Selective activation of heteromeric SK channels contribute to action potential repolarization in...

Selective activation of heteromeric SK channels contribute to action potential repolarization in mouse atrial myocytes Jane M Hancock BSc PhD, Kate L Weatherall BSc PhD, Andrew F James BSc PhD, Jules C Hancox BSc PhD, Neil V Marrion BSc PhDSchool of Physiology & Pharmacology, University of Bristol, Bristol, BS8 1TD, UK.
Supporting Information
Experimental ProceduresConstructs and cell culture. Channels were transiently expressed in HEK293 cells, with cells maintained as described previously1. Cells were transiently transfected separately with the following constructs: mouse NaV1.5 (accession number AJ271477), rat CaV1.2 (accession number AF394940), rat CaV1.3 (accession number AF370009), human KV1.5 (accession number NG_012198.1), KV2.1 (accession number NP_037318.1) and mouse KV4.2 (accession number NM019697) and hSK3_ex42. Rat CaV1.2 and 1.3 were co-transfected with rat CaV2A (accession number M80545), rat CaV21 (accession number AF286488) and enhanced green fluorescent protein (eGFP) (channel constructs were used in an equal ratio to a total plasmid concentration of 8 g per 25 cm3 flask, eGFP plasmid was used at a ratio of 1:20) using lipofectamine 2000, with cells transfected in 25 cm flasks for 24 hours before being plated in 35 mm dishes. Transient transfection of all other constructs was achieved using polyethylenimine (Alfa Aesar, Inc.) by combining channel plasmid DNA with enhanced eGFP DNA in a ratio of 1:5 to 1:10 (maximal plasmid content: 1 μg/35 mm dish). In all cases, cells were used 24-48 hr after transfection.
Electrophysiology. Recording of NaV-mediated, KV-mediated or mouse atrial myocyte currents was achieved by bathing cells in a solution of composition: NaCl (138 mM), KCl (4 mM), HEPES acid (10 mM), D-glucose (10 mM), MgCl2 (1.2 mM), CaCl2 (2.5 mM) (pH 7.4). Fire-polished electrodes (3-5 M) pulled from borosilicate glass contained: K Aspartate (130 mM), KCl (20 mM), MgCl2 (1.5 mM), EGTA (0.01 mM), MgATP (5 mM), HEPES Na (5 mM), NaOH (3 mM) (pH 7.4). A junction potential of +10 mV was calculated but not compensated for. CaV currents were recorded from cells bathed in a solution of composition: TEACl (140 mM), HEPES (10 mM), D-glucose (10 mM), MgCl2 (1 mM) and CaCl2 (5 mM) (pH 7.4) and using electrodes were filled with solution of composition: CsMeSO4 (120 mM), TEACl (20 mM), HEPES Na (10 mM), EGTA (5 mM), Mg2ATP (4 mM), MgCl2 (1 mM) (pH 7.3). A junction potential of +14 mV was calculated but not compensated for. Cells were voltage-clamped at -80 mV using an Axopatch 200A amplifier (Molecular Devices, CA, USA) and the voltage stepped to positive levels in 10 mV increments. Capacitance and series resistance compensation (>90%) was used throughout, with currents filtered at 1 kHz (8-pole low pass Bessel filter, Frequency Devices, CT, USA) and sampled at 5-10 kHz using Pulse (HEKA). All recombinant channel currents were leak-subtracted using a P/4 protocol from a holding potential of -80 mV. Recording of hSK3_ex4-mediated currents were obtained from outside-patches bathed in a solution of composition: KCl (127 mM), HEPES Na (10 mM), EGTA (10 mM), CaCl2 (6.19 mM, calculated free [Ca2+] 60 nM), MgCl2 (1.44 mM, calculated free [Mg2+] 1 mM) and electrodes containing: KCL (117 mM), HEPES Na (10 mM), EGTA ( 10 mM), Na2ATP (10), CaCl2 (9.65, mM, calculated free [Ca2+] 1 µM), MgCl2 (2.5 mM, calculated free [Mg2+] 1 mM). Currents were revealed by voltage ramps (-100 to 100 mV, 1 s duration) from a holding potential of 0 mV. All numerical values are expressed as mean ± S.E.M. Statistical analysis was performed using Prism 5.02 (GraphPad Software). All salts were purchased from Sigma-Aldrich except HEPES (Calbiochem, USA). UCL1684 (100 M) and NS8593 (10 mM) were dissolved in DMSO and diluted (1:1000) into the extracellular solution when required. The chosen final concentration of each SK channel inhibitor provided supramaximal block of expressed recombinant SK channels3,4.

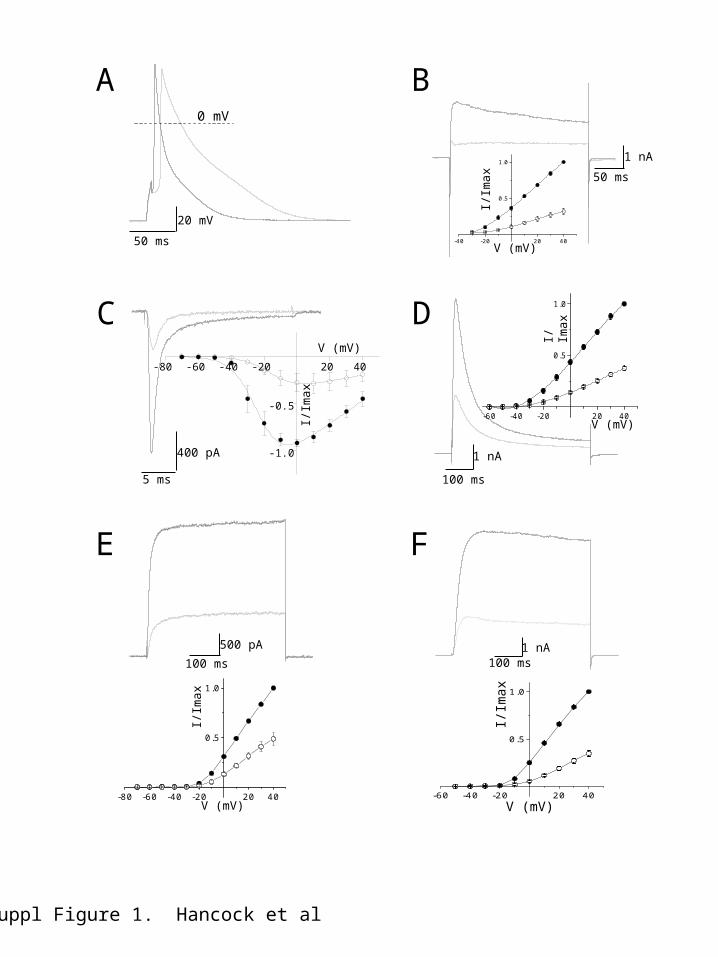
ResultsAction potentials in the presence of NS8593 (Figure 1A) exhibited a higher threshold, with the control threshold being reached by a depolarising current injection of 35 ± 1 pA, which increased to 48 ± 2 pA in the presence of NS8593 (p < 0.001, n = 5). In addition, NS8593 significantly reduced the rate of rise during the initial upstroke of the action potential (control: 152.2 ± 18.2 mV/ms, NS8593: 75.8 ± 6.3 mV/ms, p < 0.01, n = 5). The most apparent effect was on APD, with NS8593 prolonging action potential early and late repolarisation, extending the action potential at 30 % repolarisation (APD30) from 5.1 ± 1.0 ms to 13.6 ± 2.4 ms, at 60 % repolarisation (APD60) from 16.7 ± 2.7 ms to 43.5 ± 2.9 ms, and at 90 % repolarisation (APD90) from 67.1 ± 7.9 ms to 123.6 ± 9.2 ms (Figure 1A). The APD90 prolongation was significantly greater than observed with UCL1684 (p < 0.01, n = 5). The difference between the effects of the two SK channel current inhibitors suggests that NS8593 is likely to be affecting channel subtypes in addition to SK. This suggestion was reinforced by comparison of the effects of the two SK current inhibitors on outward current native to mouse atrial myocytes. Dissociated atrial myocytes were voltage-clamped at -80 mV and outward current was evoked by step depolarizations up to +40 mV. Application of NS8593 blocked outward current evoked by membrane depolarization positive to -20 mV, with sensitive current increasing in amplitude with depolarization (Figure 1B). In addition, NS8593 blocked more current at +40 mV than did UCL1684 (p < 0.001, NS8593: n = 6, UCL1684: n = 8). These data indicate that the two SK channel current inhibitors block different components of outward current native to atrial myocytes and suggest that the action of NS8593 is not selective for SK channel current. It is impossible to discriminate between some potassium and calcium channel current subtypes in atrial myocytes, because the pharmacology is poor and/or the biophysical properties do not permit separation by voltage protocols. We elected therefore, to express individually those channel subtypes that are known to underlie the atrial myocyte action potential5.
Effect of NS8593 and UCL1684 on inward sodium and calcium currentsNaV1.5-mediated inward currentThe rising phase of the atrial action potential results from activation of NaV1.5 sodium (Na+) channels5,6. Step depolarization more positive than -50 mV from a holding potential of -80 mV evoked the inward Na+ current (Figure 1C) in HEK293 cells expressing NaV1.5 channels. Peak macroscopic current was observed at -10 mV (Figure 1C, inset). The rapidly inactivating current was reduced in amplitude by addition of NS8593 (10 M) (Figure 1C), with current evoked at -10 mV being reduced by 69 ± 10 % (n = 6). Current was reduced throughout the voltage range, with current reduction being accompanied by a trend to shift the voltage dependence of activation to more depolarized levels (control V0.5 -20.3 ± 5.4 mV, NS8593 V0.5 -8.7 ± 6.2 mV, p > 0.1, n = 6) and a reduction in slope of activation (control: k ~ 7.1 ± 1.8, NS8593: k ~ 12.2 ± 1.5, n = 6, p < 0.05), suggesting that current inhibition is associated with the channel becoming less sensitive to voltage and requiring stronger depolarization to open.
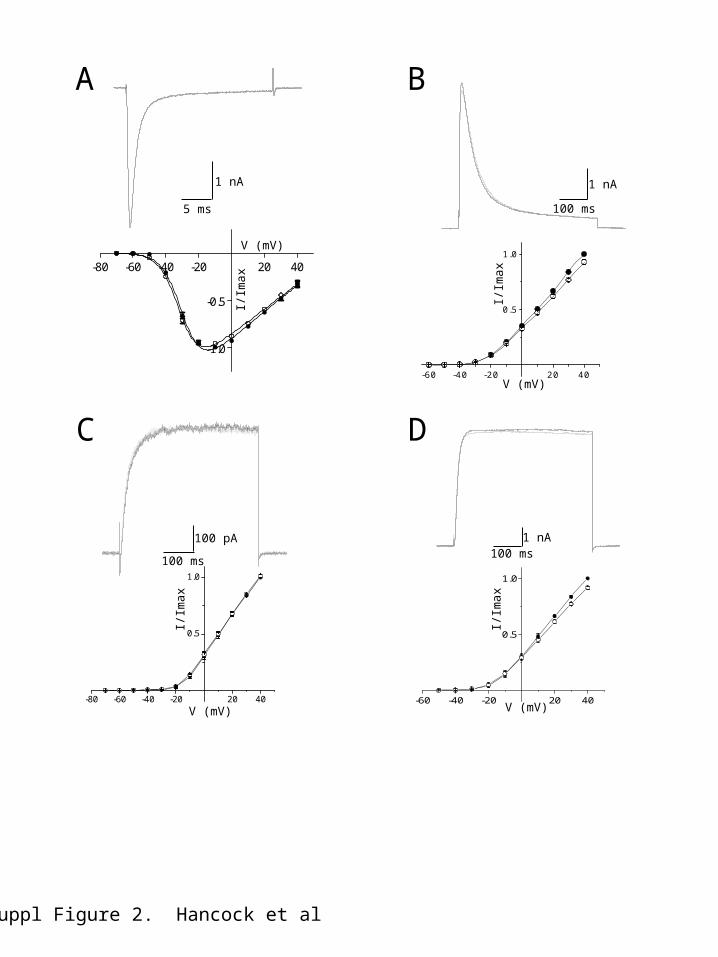
In contrast, NS8593 had no effect on the voltage-dependent rate of inactivation of NaV1.5 current at 0 mV (control : 1.1 ± 0.2 ms, NS : 1.3 ± 0.1 ms, P > 0.05, n = 6). The effect of the SK channel NAM on recombinant NaV1.5-mediated channels is consistent with the observed effect of the drug to increase the threshold and reduce the rate of rise of the atrial myocyte action potential. In comparison, macroscopic NaV1.5-mediated inward current was not affected by addition of the SK channel inhibitor, UCL1684 (100nM; Figure 2A).

CaV1.2- & CaV1.3-mediated inward currentThe plateau phase of the atrial action potential (Figure 1A) results from the activation of CaV1.2 and 1.3 calcium (Ca2+) current5,6. Application of either NS8593 (10 µM) or UCL1684 (100 nM) had no effect on inward Ca2+-carried current carried by either CaV1.2 or CaV1.3 (co-expressed with CaV2A and CaV21 subunits, the auxiliary subunits known to be expressed in cardiac tissue7,8.
Effect of NS8593 and UCL1684 on outward potassium currentsThe atrial action potential configuration is controlled by the activity of a number of potassium (K+) channel subtypes. Isolation of current resulting from activation of each K+ channel subtype is problematic in native cells, so we expressed individual channel subtypes in HEK293 cells to determine sensitivity to SK channel inhibitors. Kv4.2-mediated outward currentActivation of KV4.2 and/or KV4.2-KV4.3 channels generates the rapidly activating transient outward current (Ito) that contributes to early rapid repolarization of the atrial action potential5,6. Expression of homomeric KV4.2 channels produced a transient outward current when activated depolarized from -30 mV (holding potential -80 mV; Figure 1D, inset), displayed half activation at -4.7 ± 1.7 mV (slope factor 15.3 ± 0.5, n = 5), and inactivated with an exponential time-course that was voltage-sensitive (: 52.1 ms ± 2.5 ms at 0 mV). Application of NS8593 (10 M) reduced the amplitude of evoked KV4.2-mediated currents by 63.2 ± 3.0 % (at +40 mV, n = 5) (Figure 1D). Inhibition by NS8593 was not accompanied by any changes in the voltage-dependence of activation (V0.5 -3.3 ± 3.7 mV, p > 0.26; slope factor 16.8 ± 0.5, p > 0.08), or the rate of inactivation (control : 52.1 ± 2.5 ms, NS8593 : 54.3 ± 3.5, p > 0.5, n = 5 at 0 mV), suggesting that current inhibition might result from pore block. In contrast, application of UCL1684 (100 nM) produced a small decrease in the amplitude of KV4.2-mediated outward current that was only seen at potentials positive to +10 mV, inhibiting current amplitude by 7 ± 2 % at +40 mV (p < 0.001, n = 6) (Figure 2B).
Kv1.5-mediated outward currentA dominant contributor to the configuration of the action potential is activation of IKur (IK,slow1), which is encoded by KV1.5 subunits5,6. Expression of Kv1.5 subunits in HEK293 cells produced a delayed rectifier-like current, which activated from -20 mV (Figure 1E), with half activation seen at 7.3 ± 3.2 mV (slope factor 12.1 ± 1.0, n=4). Consistent with the dramatic prolongation of the early phase of the atrial myocyte action potential repolarization (Figure 1A), application of NS8593 (10 M) inhibited KV1.5-mediated currents by 51 ± 7 % (at +40 mV, p < 0.001, n = 4, Figure 1E), with no change in the voltage-dependence of activation (V0.5 9.7 ± 1.6 mV, p > 0.19; slope factor 11.9 ± 1.2, p > 0.37; n = 4). Application of UCL1684 (100 nM) had no effect on the amplitude or voltage-dependent properties of the expressed current (n = 6, Figure 2C). Kv2.1-mediated outward currentActivation of Kv2.1 channels is proposed to underlie IK,slow2 in the mouse atrium, which contributes to the late repolarising phase of the mouse atrial action potential9. Expression of Kv2.1 channels in HEK293 cells produced a delayed rectifier-like current activating from -20 mV (Figure 1F). NS8593 blocked the peak outward Kv2.1-mediated current by 63 ± 3 % (p < 0.001, n = 6, Figure 1F). This block of current was accompanied by a depolarizing shift in the voltage-dependence of Kv2.1-mediated current activation (control V0.5 6.9 ± 2.5 mV, NS8593 V0.5 16.3 ± 3.9 mV, p < 0.001, n = 6), with a small but significant reduction in slope of the activation curve (control: k ~ 10.1 ± 0.3, NS8593: k ~ 11.2 ± 0.2, p < 0.03, n = 6). KV2.1 channel current was affected by UCL1684 (Figure 2D), but similar to that observed with expressed KV4.2-mediated current only a small block was observed at depolarized potentials (+10 mV to +40 mV, 8 ± 1 % at +40 mV, p < 0.001, n = 6, Figure 2D, lower).

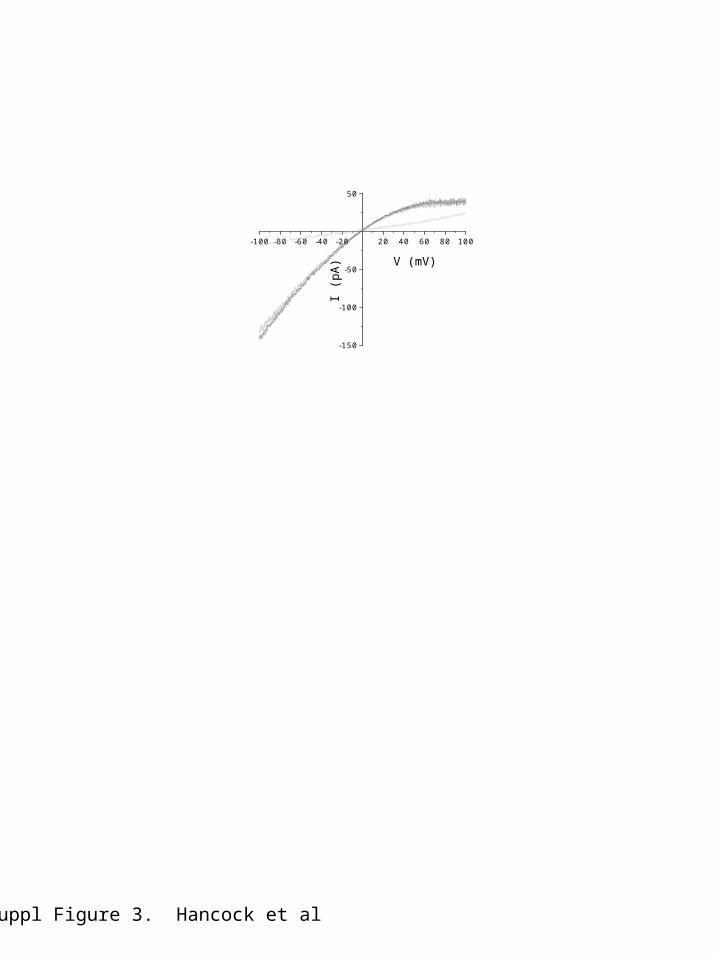
Expression of hSK3_ex4 produces a current insensitive to apaminApplication of a supramaximal concentration of apamin (100 nM) to hSK3-ex4-mediated current did not produce an effect (figure 3). This isoform of hSK3 channels2 is blocked by subsequent addition of TEA (100 mM) (Figure 3).
References1.Goodchild SJ, Lamy C, Seutin V, Marrion NV. Inhibition of K(Ca)2.2 and K(Ca)2.3 channel currents by protonation of outer pore histidine residues. J Gen Physiol 2009; 4:295-308.2.Wittekindt OH, Visan V, Tomita H, Imtiaz F, Gargus JJ, Lehmann-Horn F, Grissmer S, Morris-Rosendahl DJ. An apamin- and scyllatoxin-insensitive isoform of the human SK3 channel. Mol Pharmacol 2004; 65: 788-801.3.Strobaek D, Hougaard C, Johansen TH, Sorensen US, Nielsen EO, Nielsen KS, Taylor RD, Pedarzani P, Christophersen P. Inhibitory gating modulation of small conductance Ca2+-activated K+ channels by the synthetic compound (R)-N-(benzimidazol-2-yl)-1,2,3,4-tetrahydro-1-naphtylamine (NS8593) reduces afterhyperpolarizing current in hippocampal CA1 neurons. Mol Pharmacol 2006; 70:1771-1782.4.Weatherall KL, Goodchild SJ, Jane DE, Marrion NV. Small conductance calcium-activated potassium channel: From structure to function. Prog Neurobiol 2010; 91:242-255.5.Nerbonne JM, Kass RS. Molecular Physiology of Cardiac Repolarization. Physiol Rev 2005; 85:1205-1293.6.Ravens U. Antiarrhythmic therapy in atrial fibrillation. Pharmacol & Therapeutics 2010; 128:129-145.7.Hullin R, Singer-Lahat D, Freichel M, Biel M, Dascal N, Hofmann F, Flockerzi V. Calcium Channel Beta Subunit Heterogeneity: Functional expression of cloned cDNA from heart, aorta and brain. EMBO J 1992; 11:885-90.8.Marionneau C, Couette B, Liu J, Li H, Mangoni ME, Nargeot J, Lei M, Escande D, Demolombe S. Specific Pattern Of Ionic Channel Gene Expression Associated With Pacemaker Activity In The Mouse Heart. J Physiol 2005; 562:223-234.9.Bou-Abboud E, Li HL, Nerbonne JM. Molecular diversity of the repolarizing voltage-gated K+ currents in mouse atrial cells. J Physiol 2000; 529:345-358.
400 pA
5 ms
20 mV
50 ms
0 mV
A
1 nA
50 ms
C
Suppl Figure 1. Hancock et al
I/Im
ax
1 nA
100 ms
-60 -40 -20 20 40
0.5
1.0
V (mV)
I/Im
ax
-80 -60 -40 -20 20 40
-1.0
-0.5
V (mV)
1 nA100 ms
500 pA
100 ms
-80 -60 -40 -20 20 40
0.5
1.0
V (mV)
I/Im
ax
-60 -40 -20 20 40
0.5
1.0
I/Im
ax
V (mV)
E
B
D
F
-40 -20 20 40
0.5
1.0
I/Im
ax
V (mV)
1 nA100 ms
100 pA
100 ms
B
1 nA
5 ms
1 nA
100 ms
Suppl Figure 2. Hancock et al
A
C D
-80 -60 -40 -20 20 40
-1.0
-0.5
V (mV)I/
Ima
x
-60 -40 -20 20 40
0.5
1.0
V (mV)
I/Im
ax
-80 -60 -40 -20 20 40
0.5
1.0
I/Im
ax
V (mV)-60 -40 -20 20 40
0.5
1.0
I/Im
ax
V (mV)
-100 -80 -60 -40 -20 20 40 60 80 100
-150
-100
-50
50
I (p
A)
V (mV)
Suppl Figure 3. Hancock et al



















