
Seagrasses and eutrophication - DiVA Portal
44
Seagrasses and eutrophication Interactions between seagrass photosynthesis, epiphytes, macroalgae and mussels Esther Francis Mvungi
Transcript of Seagrasses and eutrophication - DiVA Portal

Seagrasses and eutrophication
Interactions between seagrass photosynthesis,
epiphytes, macroalgae and mussels
Esther Francis Mvungi

©Esther F. Mvungi, Stockholm 2011
Front cover: Photo showing eutrophic seagrass bed in the Western
Indian Ocean. Photo: Mats Björk
ISBN 978-91-7447-250-9
Printed in Sweden by Universitetservice, US-AB, Stockholm 2011
Distributor: Department of Botany, Stockholm University

To my husband, Denis and
my daughter, Doreen

Abstract
Seagrass meadows are highly productive, ecologically and economically
valuable ecosystems. However, increased human activities along the
coastal areas leading to processes such as eutrophication have resulted in
the rapid loss and deterioration of seagrass ecosystems worldwide. This
thesis focuses on the responses of seagrasses to increases in nutrients,
subsequent increases in ephemeral algae, and changes in the physical-
chemical properties of seawater induced by interaction with other marine
biota. Both in situ and laboratory experiments conducted on the tropical
seagrasses Cymodocea serrulata and Thalassia hemprichii revealed that
increased concentrations of water column nutrients negatively affected
seagrass photosynthesis by stimulating the growth of the epiphytic
biomass on the seagrass leaves. Interaction between seagrasses and other
marine organisms induced different responses in seagrass photosynthesis.
Ulva intestinalis negatively affected the photosynthetic performance of
the temperate seagrass Zostera marina both by reducing the light and by
increasing the pH of the surrounding water. On the other hand, the
coexistence of mussels Pinna muricata and seagrass Thalassia
hemprichii enhanced the photosynthetic activity of the seagrass, but no
effect on the mussels’ calcification was recorded. This study
demonstrates that seagrass productivity is affected by a multitude of
indirect effects induced by nutrient over-enrichment, which act singly or
in concert with each other. Understanding the responsive mechanisms
involved is imperative to safeguard the ecosystem by providing
knowledge and proposing measures to halt nutrient loading and to predict
the future performance of seagrasses in response to increasing natural and
human perturbations.
Keywords: CO2, epiphytes, eutrophication, mussels, pH, photosynthetic
activities, seagrasses, Ulva

List of Papers
This thesis is based on the following four papers, which will be referred
to in the text by their Roman numerals.
I. Mvungi EF, Lyimo TJ and Björk M (2011). When Zostera
marina is intermixed with Ulva, its photosynthesis is reduced by
increased pH and lower light, but not by changes in light quality
(Submitted).
II. Hamisi MI, Mvungi EF, Lyimo TJ, Mamboya FA, Österlund K,
Björk M, Bergman B and Diez B (2011). Nutrient enrichment
affects the seagrass Cymodocea serrulata and induces changes to
its epiphytic cyanobacterial community (Manuscript).
III. Mvungi EF and Mamboya FA (2011). Photosynthetic
performance, epiphyte biomass, and nutrient content of two
seagrass species in areas with different levels of nutrient along
the Dar es Salaam coast (Submitted).
IV. Semesi IS, Mvungi EF, Beer S, Jidawi N and Björk M (2011).
Interactions between seagrasses and mussels: CO2, pH and
calcification (Submitted).
My contributions were: for papers I and III planning, performing all
experiments, executing fieldwork, analysing data, and writing the
manuscripts; for paper II planning, performing experiments (except the
morphological and molecular characterization of epiphytic
cyanobacteria), conducting part of the data analysis, and writing part of
the manuscript in cooperation with the first co-author; and for paper IV
planning, executing fieldwork, analysing data, and writing the manuscript
in cooperation with the first co-author.

Table of Contents
Abstract .................................................................................................... iv
List of Papers .......................................................................................... v
Table of Contents ................................................................................... vi
Abbreviations ......................................................................................... vii
1. Introduction ........................................................................................ 8 1.1 General overview of seagrasses ............................................................. 8 1.2 Nutrient fluxes in marine environments ............................................... 9 1.3 Photosynthesis in seagrass meadows ................................................. 10 1.4 Chlorophyll a fluorescence .................................................................... 11 1.5 Inorganic carbon flux in marine environments ................................. 12 1.6 Seagrasses interact with other marine organisms ........................... 14
2. Aim ..................................................................................................... 16
3. Comments on the methods used ................................................. 17 3.1 Species used ............................................................................................ 17 3.2 Experimental setups ............................................................................... 19 3.3 Chlorophyll fluorescence measurements ............................................ 20 3.4 Nutrient content (CNP) analysis ........................................................... 21 3.5 Physical-chemical characteristics of seawater ................................... 21
4. Results and Discussion .................................................................. 22 4.1 Effects of water column nutrient gradients ........................................ 22
4.1.1 Effects on seagrass photosynthesis .......................................... 23 4.1.2 Effects on epiphyte biomass and composition ........................ 26 4.1.3 Effects on nutrient contents of plant tissues ........................... 26
4.2 Ulva overgrowth affects seagrass photosynthesis ............................ 28 4.3 Mussels enhance seagrass photosynthesis ........................................ 30
5. Conclusion ........................................................................................ 32
Acknowledgements .............................................................................. 34
References ............................................................................................. 36

Abbreviations
AF Absorption factor
C:N:P ratio Carbon:nitrogen:phosphorus ratio
Ci Inorganic carbon
ETR Electron transport rate
Fv/Fm Maximum quantum yield
Ik Minimum saturation irradiance
PAM Pulse amplitude modulation/modulated
PAR Photosynthetically active radiation
PSI Photosystem one
PSII Photosystem two
rETR Relative electron transport rate
RLC Rapid light curve
TA Total alkalinity

8
1. Introduction
1.1 General overview of seagrasses
Seagrasses are defined as flowering plants (angiosperms) living their full
lifecycle submersed in marine environments. They are found in all coastal
areas of the world except along the Antarctic shores (Green and Short 2003,
Duarte and Gattuso 2010, Fig. 1). They are descendants of terrestrial plants
that re-entered the ocean about 100 million years ago (Duarte and Gattuso
2010). Thus, unlike other marine vegetation, they maintained both their
vascular system (Kuo and den Hartog 2006) and their ability to produce
flowers, fruits, and seeds (Ackerman 2006). They comprise less than 0.02 %
of the angiosperm species, and have relatively fewer species than other
marine organisms (Short et al. 2007).
Despite their low diversity, seagrass beds are ecologically and economically
highly important. They are among the world’s most productive coastal
ecosystems (Duarte and Chiscano 1999) and support a high diversity of
autotrophic and heterotrophic organisms. They serve as habitats for a broad
diversity of invertebrate and fish species, many of which are of commercial
and recreational importance, as food source for marine herbivores (Heck and
Valentine 2006), they stimulate nutrient recycling and buffer wave action,
hence minimizing coastal erosion (Heiss et al. 2000). They also serve as
substrata for a diverse array of epiphytic organisms that contribute to the
productivity of the seagrass beds in different ways (Borowitzka et al. 2006).
Seagrasses are not only supportive to marine biota, but also to human
populations, especially in the tropical regions (in particular the Western
Indian Ocean region) where people along the coasts depend largely on the
seagrass ecosystem for their daily subsistence in terms of food and per capita
income (de la Torre-Castro and Rönnbäck 2004).
Regardless of their biological and economical importance, reports on
seagrass decline have increased in numbers in recent years and the major
cause attributed is the increase in human-related activities along the coastal
zone (Short and Wyllie-Echeverria 1996, Orth et al. 2006, Waycott et al.
2009). Nutrient over-enrichment (eutrophication) from various sources, such
as domestic waste, industrial discharge, agriculture, and urbanization of
coastal zones (Moore and Wetzel 2000, Burkholder et al. 2007, Duarte et al.
2008), concomitant with global climate change (Short and Neckles 1999),
pose serious threats to seagrass ecosystems worldwide.

9
Fig. 1. Global seagrass distribution (shown in blue) and their geographic bioregions:
1. Temperate North Atlantic, 2. Tropical Atlantic, 3. Mediterranean, 4. Temperate
North Pacific, 5. Tropical Indo-Pacific, 6. Temperate Southern Oceans. The figure is
reproduced from Short et al. (2007), with the kind permission from the publisher,
Elsevier. Copyright © (2007), Elsevier.
1.2 Nutrient fluxes in marine environments
Seagrasses as any other plants require nutrients for their growth,
development, and metabolism, but too much of some nutrients may limit
rather than promote growth. The primary and most common nutrients that
limit seagrass growth in natural systems are nitrogen and phosphorus
(Duarte 1990, Romero et al. 2006). These essential nutrients may be derived
from the decomposition of organic matter in the water column and sediments
(Holmer and Olsen 2002, Kilminster et al. 2006), in which seagrass litters
are the main source (Holmer and Olsen 2002, Romero et al. 2006).
However, in tropical areas, bacterial nitrogen fixation may also provide
substantial amount of nitrogen in the seagrass meadows (Moriarty and
O’Donohue 1993, Hamisi et al. 2009). Unlike terrestrial plants, seagrass can
absorb nutrients through both leaves and roots (Touchette and Burkholder
2000a, Gras et al. 2003, Nielsen et al. 2006).
Seagrasses can be adversely affected by eutrophication of the water in
different ways. Increased nutrient levels in the water column promote
proliferation of phytoplanktonic, epiphytic, and bloom-forming macroalgal
species (Short et al. 1995, Borum and Sand-Jensen 1996, Moore and Watzel

10
2000, McGlathery 2001). Blooms formed lead to reduction in the light
available to seagrasses (Brun et al. 2003, Liu et al. 2005, Lamonte and
Dunton 2006), and alteration of the surrounding water chemistry (Björk et
al. 2004, Beer et al. 2006). Thus, nutrient over-enrichment caused by
anthropogenic activities has been considered to be a major cause of seagrass
decline worldwide (Green and Short 2003, Orth et al. 2006). Many studies
have been carried out to investigate the effects of increased nutrient levels on
seagrass performance (e.g. Short et al. 1990, Moore and Wetzel 2000,
Touchette and Burkholder 2000a; 2000b, Romero et al. 2006, Burkholder et
al. 2007, Lee et al. 2007) but still not conclusive.
When the nutrient is absorbed into the cell, it can either be assimilated
immediately into organic matter or stored in inorganic form in the plant
tissues for future use. Increased nutrient availability has been shown to
correlate with increased nutrient acquisition and assimilation in seagrasses of
both N and P, which is reflected by the increased levels of N and P content
in the plant tissues (Duarte 1990, Bulthuis et al. 1992, Peréz-Lloréns and
Niell 1995, Borum and Sand-Jensen 1996, Pedersen et al. 1997, Alcoverro et
al. 1997, Udy and Dennison 1997, Johnson et al. 2006). Therefore, analysis
of tissue nutrient content is thought to be a more reliable way of determining
fluxes in nutrient availability in the ecosystem than water nutrient analysis,
as the latter is influenced by short-lived temporal fluctuations which are
difficult to trace (Sjöö and Mönk 2009, Lourenço et al. 2006).
1.3 Photosynthesis in seagrass meadows
Oxygenic photosynthesis is responsible for all biochemical production of
organic matter in marine and terrestrial ecosystems, and about 45% of the
photosynthesis on the earth occurs in aquatic environments (Field et al.
1998). The photosynthetic reaction in plants starts with absorption of light
by chlorophylls, which are composed of large protein complexes with light
harvesting antennae where light energy is converted into chemical energy
and charge separations of electrons take place. The electron is then
transported through and along the thylakoid membrane via series of electron
carriers to the photosystem I (PSI), where trans-membrane proton gradient
and reduction of NADP+ to NADPH occurs and ATP is formed via proton
gradient. The electrons are resupplied from the water-splitting complex in
photosystem II (PSII), in which water molecule is cleaved, yielding 4
electrons and 4 protons for each molecule of O2 generated. The NADPH and
ATP formed through these processes are then used in a Calvin cycle where
CO2 is reduced into sugars.

11
In the seagrass beds, however, light required for photosynthesis must
penetrate the water column. With increasing water depth, light becomes
more and more attenuated and scattered due to dissolved substances,
phytoplankton, algal blooms, and physical properties of water itself.
Consequently, light is also one of the limiting factors for seagrass
productivity (Hemminga and Duarte 2000).
1.4 Chlorophyll a fluorescence
When light energy is absorbed by PSII, it can be used through one of three
directly competing pathways: in photochemistry, as heat dissipation or can
be re-emitted back as fluorescence (Krause and Weiss 1991). When
absorbing a photon, the chlorophyll molecule becomes excited and moves to
a higher energy level. If the energy of the excited electron is not utilized in
photochemistry (or heat dissipation), the electron falls back to the ground
state and the energy will be emitted as fluorescence (Krause and Weiss
1991).
Chlorophyll fluorescence techniques are a convenient and non-destructive
way to assess processes of photosynthesis, both in situ and in the laboratory,
and have been used extensively in recent years (Krause and Weiss 1991,
Maxwell and Johnson 2000, Schreiber 2004). When a leaf is exposed to a
saturating irradiance the fluorescence yield initially increases rapidly to a
peak and then slowly declines. This fluctuation in the fluorescence signal is
known as the “Kautsky” curve (Kaustsky et al 1960, Fig. 2), from which
dynamic changes in photosynthesis can be estimated.
When a plant sample is dark-adapted for sufficient time (ca.10–15 minutes),
all its PSII reaction centres open, providing the maximum quantum yield of
PSII (Maxwell and Johnson 2000), estimated as: Fv/Fm = (Fm−Fo)/Fm
where Fv is the variable fluorescence, Fm is the maximum fluorescence
following a saturating light pulse, and Fo is the minimum fluorescence (Fig.
2). The Fv/Fm ratio has been used as a simple and rapid measure of stress
status of the plants (Krause and Weiss 1991, Baker 2008).
When a sample is light-adapted, its reaction centres are closed, and it
provides the proportion of absorbed energy that being used for
photochemistry (Genty et al. 1989, Baker 2008), and is calculated as ФPSII
(Y) = (Fm′ – F′) / Fm′, where F′ and Fm′ are the corresponding light-
acclimated minimum and maximum fluorescence (Fig. 2). The electron
transport rate (ETR) can be obtained from the ФPSII value as follows: ETR =
Y * PAR * 0.5 * AF; where Y is the effective quantum yield, PAR is the
irradiance at the leaf surface, 0.5 is a factor that accounts for partitioning of
12
absorbed photons between PSII and PSI, and AF is the proportion of incident
light that is absorbed by photosynthetic pigments (Beer et al. 2001).
Photosynthesis has long been measured using gas exchange techniques,
which are destructive because they require the removal of the plant from its
natural environment. However, the development of underwater pulse
amplitude modulated (PAM) fluorometer made it possible to conduct in
situ photosynthetic measurements in the ambient environments (Schwarz et
al. 2000). Because PAM has the advantages of being non-intrusive, fast,
reliable, and able to provide real-time data, it has become very popular.
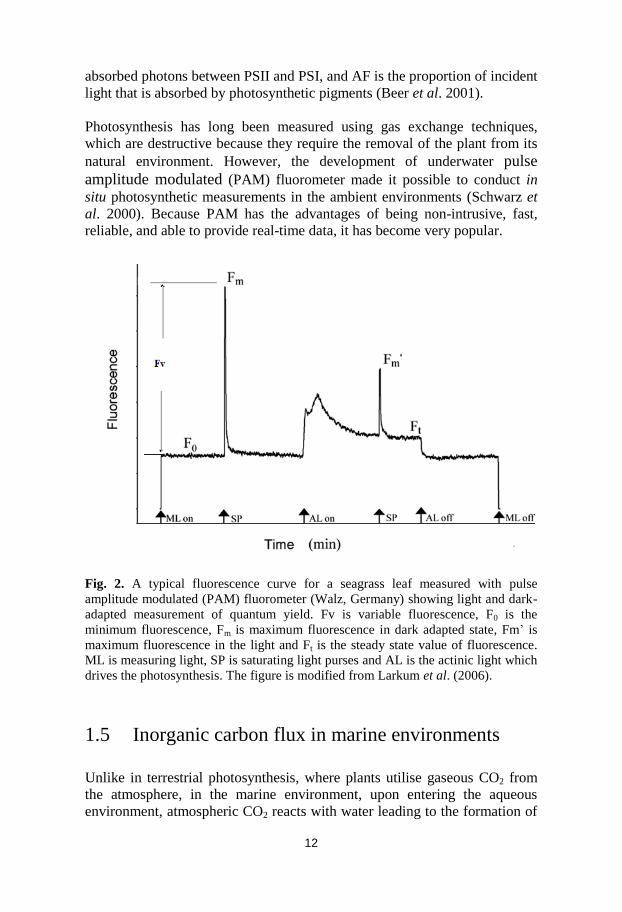
Fig. 2. A typical fluorescence curve for a seagrass leaf measured with pulse
amplitude modulated (PAM) fluorometer (Walz, Germany) showing light and dark-
adapted measurement of quantum yield. Fv is variable fluorescence, F0 is the
minimum fluorescence, Fm is maximum fluorescence in dark adapted state, Fm’ is
maximum fluorescence in the light and Ft is the steady state value of fluorescence.
ML is measuring light, SP is saturating light purses and AL is the actinic light which
drives the photosynthesis. The figure is modified from Larkum et al. (2006).
1.5 Inorganic carbon flux in marine environments
Unlike in terrestrial photosynthesis, where plants utilise gaseous CO2 from
the atmosphere, in the marine environment, upon entering the aqueous
environment, atmospheric CO2 reacts with water leading to the formation of

13
ionic forms of carbon (carbonic acid, bicarbonate, and carbonate) in
equilibrium with CO2, as described by Falkowski and Raven (2007):
H2O + CO2 ↔ H2CO3 ↔ H+ + HCO3
- ↔ 2H
+ + CO3
2-
However, depending on pH changes in the seawater, the equilibrium reaction
can shift, with high concentrations of CO2 at lower pH and relatively more
bicarbonate and carbonate at higher pH. At pH 8.2, salinity 35, and water
temperature 25oC, more than 95% of inorganic carbon present is in the form
of bicarbonate (Falkowski and Raven 2007).
Unlike in the terrestrial ecosystem, in marine photosynthesis CO2 can often
be a limiting factor due to its low solubility and slow diffusion rates (Björk
et al. 1997, Beer et al. 2002). Some species have developed mechanisms that
enable them to use the readily available bicarbonate (Beer and Rehnberg
1997, Invers et al. 2001). One example is the use of extracellular carbonic
anhydrase (CA), which catalyses the conversion of HCO3- to CO2 and OH
-.
This can be further accelerated by the use of proton pump-mediated acid
zones and by active uptake of HCO3- (possibly by an AE protein, Drechsler
et al. 1993). Although most marine macrophytes have been reported to be
able to use both CO2 and HCO3− for photosynthetic carbon fixation (Beer
and Rehnberg 1997, Invers et al. 2001), the use of bicarbonate in some
seagrass species is still under some dispute and is thought to be less efficient
as compared to the macroalgae (Beer and Rehnberg 1997, Björk et al. 1997).
Seagrass photosynthesis has been shown to be carbon-limited for both
temperate and tropical species (Björk et al. 1997, Invers et al. 1997).
Effect of increasing global CO2 on marine biota
The concentration of atmospheric CO2 has increased in the past 200 years
due to anthropogenic activities, and almost one third of it goes into the ocean
(Sabine et al. 2004). There it causes ocean acidification, in which the pH and
concentration of carbonate ions decrease while concentrations of hydrogen
and bicarbonates increase. These chemical changes brought about by the
dissolution of CO2 are expected to affect many biological processes in
marine organisms (Fabry et al. 2008). Despite the fact that these changes are
predicted to continue rapidly as the oceans take in more CO2 from the
atmosphere, little yet is known about how marine organisms will cope with
such changes at individual or ecosystem levels.
Several studies have been carried out focusing on the processes likely to be
affected by ocean acidification. For instance, marine calcifying organisms
have been shown to be negatively affected by the increase in CO2
concentration through the marked decrease in rates of calcification (e.g.
Fabry et al. 2008, Ries et al. 2009). Unlike calcifying organisms, marine

14
photosynthetic organisms have been shown to have enhanced photosynthetic
activities in response to higher levels of CO2 (Palacios and Zimmerman
2007, Riebesell et al. 2000, Schneider and Erez 2006). However, this
observation is based on only a few studies on a few species, hence more
investigations at individual and ecosystem levels are called for.
1.6 Seagrasses interact with other marine organisms
Seagrass–mussel interactions Seagrass ecosystem forms a highly complex habitat that harbours more
faunal assemblages than unvegetated habitats (Duarte and Gattuso 2010).
Mussels, for example, are a conspicuous feature of intertidal habitats on both
soft and hard substrata in many coastal areas. Through their filter-feeding
activity, mussels may be capable of lessening both light and nutrient
limitation in the seagrass meadows (Newell 2004). The presence of filter-
feeding mussels can significantly reduce suspended algal biomass and
inorganic particles, thereby increasing light availability and penetration
through the water column, and subsequently increasing the growth rates of
seagrasses (Wall et al. 2008). They also link water column productivity to
the benthos by removing pelagic organisms, increase deposition of solid and
faecal materials, and therefore promote a flux of organic matter and nutrient
recycling from the water column into the sediments, thus stimulating the
uptake and growth of seagrasses (Kotta and Møhlenberg 2002, Lauringson et
al. 2007, Peterson and Heck 1999, 2001a & 2001b).
Co-habitation of mussels and seagrasses is commonly seen in the coastal
areas of Tanzania; however knowledge about their interactions is generally
scarce. Thus, paper IV was designed to investigate the potential influence
that the dissolved CO2 released during the respiration of mussels (Pinna
muricata) might have on seagrass photosynthesis.
Seagrass–epiphytes and –macroalgae interactions
Seagrasses grow in a variety of sediment types in shallow water and usually
provide the solid substrata for the attachment of a diversity of epiphytic
organisms. Most seagrass epiphytes are primary producers and hence an
important component of seagrass ecosystems. They account for up to 30% of
the ecosystem’s productivity and the epiphytic algae contribute more than
30% of the total above-ground biomass in many seagrass ecosystems
(Borowitzka et al. 2006). Epiphytes constitute a valuable food source for
herbivores as they can be consumed selectively, accidentally, or through
detritus food webs (Borowitzka et al. 2006). Higher levels of N2-fixation in

15
the seagrass ecosystems have been linked to bacteria and cyanobacteria, both
in the rhizosphere and the phyllosphere (Welsh et al. 2000, Hamisi et al.
2009), of which epiphytic cyanobacteria have been estimated to supply up to
38% of the nitrogen needed for primary production in some seagrass beds
(Welsh et al. 1996).
Calcareous algae on the other hand, are also common epiphytes of
seagrasses and contribute to the production of calcareous sediments
(Chisholm 2000). It has been shown that seagrass photosynthesis influences
calcification rates in coralline algae (Semesi et al. 2009), thus it was
interesting to explore whether the same effect occurred on mussels
calcification when grow intermixed with seagrasses (paper IV).
Despite their important beneficial roles in seagrass ecosystems, epiphytes
can also have detrimental effects on seagrasses. Epiphyte loads reduce the
productivity of the seagrass by shading, competing for nutrient availability,
and creating physical barriers around the leaf surface, which may impede
light absorption (Romero et al. 2006). Similarly, in eutrophic environments,
green macroalgae frequently occur in masses, forming dense canopies or
intermixed with seagrasses, reducing the amount of light reaching the
seagrasses beneath (Brun et al. 2003, McGlathery 2001). They also tend to
alter the light spectrum as they absorb mainly in the blue and red regions,
meaning that the light transmitted through the algal mats is mainly in the
green spectrum (Vergara et al. 1997), which is regarded to be less effective
for photosynthesis (e.g. Mazzella and Alberte 1986). Based on this, paper I
investigated how photosynthetic activity in the temperate seagrass Zostera
marina is affected by changes both in water chemistry and in light quality
due to shading attributable to the green macroalgae Ulva intestinalis. Paper
III addresses the photosynthetic activities of two seagrasses in relation to
epiphytic biomass in two areas with different levels of nutrients.

16
2. Aim
The major aim of this thesis was to achieve a better understanding of the
response of seagrass species to elevated levels of nutrients in the water
column and the possible influences of interactions between seagrasses and
other organisms in the seagrass beds. To achieve this, in situ and laboratory
experiments were designed and conducted with the following specific
objectives:
i. To evaluate the effect of increased nutrient concentrations in the
water column and the associated effects of increased epiphytic
loads on seagrass performance (papers I, II, and III).
ii. To investigate effects on seagrass photosynthesis by interactions
between seagrasses and other flora and fauna (papers I and IV).

17
3. Comments on the methods used
This thesis is based on experiments conducted both in situ and in the
laboratory. This section summarises the methodology used in the different
experiments (refer to papers I–IV for detailed descriptions).
3.1 Species used
Zostera marina (Eelgrass) is the dominant seagrass species in the temperate
coastal waters of both the Pacific and the Atlantic oceans (Fig. 1). Eelgrass is
very abundant in the Baltic sea
and can colonize depths up to 30
m in clear waters, but is typically
restricted to shallows or the
intertidal depths of estuaries
(Borum and Greve, 2004). It
grows on a wide range of soft and
mixed sediments. (Photo: Fort
Wetherill)
Cymodocea serrulata belongs to the family Potamogetonaceae, commonly
found along the coasts of the tropical Indo-West Pacific region (Fig. 1). It is
one of the twelve species in the
Western Indian Ocean. C.
serrulata can be distinguished
from other seagrass species by
their shoots with distinctive
open leaf scars, triangular, flat
leaf sheath, fibrous roots on the
shoot, and serrated leaf tips
(Oliviera et al. 2005, Photo:
Katrin Osterlund).

18
Thalassia hemprichii is a member of the family Hydrocharitaceae, widely
distributed in Indo-Pacific regions (Fig. 1). It is a common species in many
coastal areas of the Western Indian Ocean, growing within soft substrates in
the intertidal and subtidal areas. The distinctive features of T. hemprichii is
the presence of short black bars of tannin cells on the leaf blade running
parallel to the long axis of the leaf and the thick rhizome with conspicuous
scars between successive erect shoots (Oliviera et al. 2005, photo: Katrin
Osterlund).
Ulva intestinalis is a cosmopolitan green macroalga species frequently found
in many habitats along the coastal zones of most oceans. In favourable
conditions U. intestinalis can quickly occupy any empty space in the
uppermost littoral zone (Martins and Marques 2002, photo: Satogawa-cho).
Pinna muricata: is a bivalve species in the family Pinnidae, which occurs
intermixed in the seagrass beds and in unvegetated areas in the Indo-Pacific
region. P. muricata in the Western Indian Ocean region live buried in sandy

19
muddy substrates in subtidal and shallow areas covered by different seagrass
species including Thalassia hemprichii (Richmond 2002, photo: E. Mvungi).
3.2 Experimental setups
Experiments to investigate the combined effects of Ulva-induced changes in
light quality and changes in water chemistry on the photosynthetic response
of Zostera marina were conducted in the laboratory, at the Botany
Department of Stockholm University. Shoots of Zostera marina were
collected from west coast of Sweden. At the laboratory, the photosynthetic
characteristics of the seagrass and the changes in water chemistry were
assessed under normal light and under Ulva-filtered light (paper I).
To investigate the effects of increased nutrient levels on performance of the
seagrass Cymodocea serrulata and the associated diazotrophic community,
an outdoor laboratory experiment was set at the Institute of Marine Sciences
(IMS), Zanzibar, Tanzania (paper II). In this setting, seagrass samples were
collected from the Chapwani seagrass bed in the Indian Ocean, about 2.5 km
from IMS and transported to the laboratory, where they were transplanted to
buckets containing sediments in a flow-through water system. Seagrass
growth, photosynthetic characteristics, and epiphytic characterization were
evaluated during the experiments.
Paper III was designed to assess the response of seagrasses in their natural
environments to varying nutrient levels in relation to epiphytic loads. A field
experiment was conducted during low tides in the intertidal seagrass beds at
Mjimwema and Ocean Road along the Dar es Salaam coast. Ocean Road is
located at 6o49' S, 39
o19' E. The area is considered to be a nutrient-rich site
as it receives untreated domestic sewage through the pipes that drain the city
centre. Mjimwema is located on the same coast about 4 km south of Ocean
Road at 6o50' S, 39
o21' E. This area is considered to be nutrient poor and less
impacted by anthropogenic disturbances from the coastal population. These
sites were selected based on their contrasting nutrient status (paper III).

20
Seagrass interaction with other marine animals is common in many parts of
the world; however the nature of these interactions has not been fully
explored, in particular in the Western Indian Ocean region. Thus, to
investigate the potential influence of the mussels Pinna muricata on the
photosynthesis of the seagrass Thalassia hemprichii, and consequently on
the water chemistry, a field experiment was set at the Fumba Bay seagrass
beds (paper IV). Fumba Bay is located on the south-west of Unguja Island,
Zanzibar, Tanzania (06o 19´S, 39
o 17´ E). The bay is characterized by
shallow waters, and during low tides it has large exposed intertidal flats,
dominated by large monospecific stands of Thalassia hemprichii and mixed
patches of other seagrass species.
3.3 Chlorophyll fluorescence measurements
In all experiments (papers I–IV) chlorophyll a fluorescence was measured
with a diving pulse amplitude modulated (PAM) fluorometer (Walz,
Germany). Photosynthetic activities of seagrass were measured in terms of
rapid light curve (RLC), maximum quantum yield of PSII (Fv/Fm), and
effective quantum yield at different parts of the leaves depending on the
study question (see papers I–IV). Prior to any chlorophyll measurements, the
PAM flourometer’s light sensor was calibrated against a light meter
(IL1400A, International Light Technologies).
Rapid light curves were generated automatically by the diving PAM in a pre-
programmed incremental sequence of actinic illumination periods, with light
intensities increasing in 8 steps of photon m-2
s-1
according to the methods
described by Ralph and Gademann (2005). The maximum rate of electron
transport (ETRmax) and initial slope (α) were calculated by fitting the RLC
data to an exponential function described by Jassby and Platt (1976). The
onset of light saturation (Ik) was calculated by dividing the ETRmax by initial
slope (papers I, II, and III).
The Fv/Fm was determined by dark-adapting the seagrass leaves for 10 min
using the dark leaf clip (Walz, Germany) and saturating pulses of light from
the diving PAM (papers II and III). The effective quantum yield (Y) was
determined under ambient light conditions by the saturating-light method
(Beer and Björk 2000). The value of Y was then used to estimate the
electron transport rate (ETR) through photosystems II and I (paper IV). The
use of PAM reflects photosynthetic rates in the light reaction only, but it
correlates well with oxygen evolution technique (Beer and Björk 2000, Silva
and Santos 2004). Thus, it permits estimation of primary productivity in the
field, on intact plants, which is very important.

21
3.4 Nutrient content (CNP) analysis
Samples used for elemental content analysis were dried at 60oC and ground
to fine powder (papers II and III). Carbon and nitrogen (CN) contents of
seagrass tissues (and epiphytes in paper I) were analysed using a CHN
analyser (CHNS 932) and phosphorus (P) content was determined by dry-
oxidation acid hydrolysis extraction followed by colourimetric analysis
(Fourqurean et al. 1992). The carbon, nitrogen, and phosphorus contents are
given in percentages of dry weight, and ratios are mol:mol.
3.5 Physical-chemical characteristics of seawater
In both in situ and laboratory experiments (papers I and IV), pH
measurements were performed with a Multi 340i pH meter (WTW,
Germany). Concentrations of total inorganic carbon and total alkalinity (TA)
in the seawater were determined following the procedures of Anderson and
Robinson (1946): whereby 4 ml of water sample was taken, the pH was
measured before addition HCL, then 1 ml of 0.01 M HCL was added and the
pH was recorded again. The pH values obtained before and after addition of
HCL were used to calculate TA and total carbon using the formula and
constants of Riley and Skirrow (1965) and Smith and Kinsey (1978).
The calcification rates of mussels were estimated using alkalinity
anomaly technique (Smith and Key 1975) by dividing changes in total
alkalinity (TA) by 2. Rapid titration method is not as precise as an end
point titration, but: it is easy, possible to perform in the field and good
enough for our purposes.
For analyses of nutrient concentrations in the water, samples were collected
from the experimental buckets (paper II) and from the field (paper III). The
concentrations of nitrate, nitrite, and phosphates were determined following
the procedures described by Parsons et al. (1989). Samples not immediately
analysed were stored at −20oC.

22
4. Results and Discussion
Tropical marine environments are characterized by low nutrient levels
(Hemminga and Duarte 2000), however, with the increasing rate of human-
related activities in coastal areas, the rate of nutrient loading in marine
environments is increasing, in many parts of the world. Nutrient over-
enrichment stimulates the overgrowth of macroalgae and epiphytic loads on
seagrass leaves, both of which have been found here and in previous studies
(e.g. Short et al. 1995, McGlathery 2001) to affect the performance of
seagrasses.
4.1 Effects of water column nutrient gradients
The results obtained in the studies reported here showed that responses of
seagrasses to nutrient gradients vary, both in the laboratory and in the field.
The nutrient enrichment experiments revealed no significant difference in
growth characteristics between of seagrasses growing in different nutrient
levels. However, the biomass of the seagrass decreased when exposed to the
higher nutrient level for an extended time (paper II). Furthermore, in the
natural environment the studied seagrasses showed different growth
characteristics in the two sites with different levels of nutrient (Table 1).
While Cymodocea serrulata showed significantly higher shoot densities and
canopy height at the nutrient-poor site than at the nutrient-rich site, shoot
density for Thalassia hemprichii did not vary significantly between the two
sites, but its canopy height was significantly higher at the nutrient-rich site
than at the nutrient-poor site (Table 1).
The two species varied in their biomass partitioning in the two sites. Species
at the nutrient-poor site showed significantly higher allocation of biomass in
the below ground than above ground tissues, while at the nutrient-rich site,
the partitioning was more or less the same for above and below ground
tissues. Thus, the above- to below-ground biomass ratio for both species was
lower at the nutrient-poor site than at the nutrient-rich site, reflecting a
strategy to cope in low nutrient availability by expansion of the surface area
for nutrients uptake (paper III).

23
Table 1. Characteristics of the seagrasses Thalassia hemprichii and
Cymodocea serrulata in areas with different levels of anthropogenic nutrient
inputs. Data are means ± Standard deviation).
4.1.1 Effects on seagrass photosynthesis
Photosynthetic performance of the seagrass Cymodocea serrulata was
negatively affected by prolonged exposure to increased level of nutrients in
the water column, as revealed by the reduction of maximum quantum yield
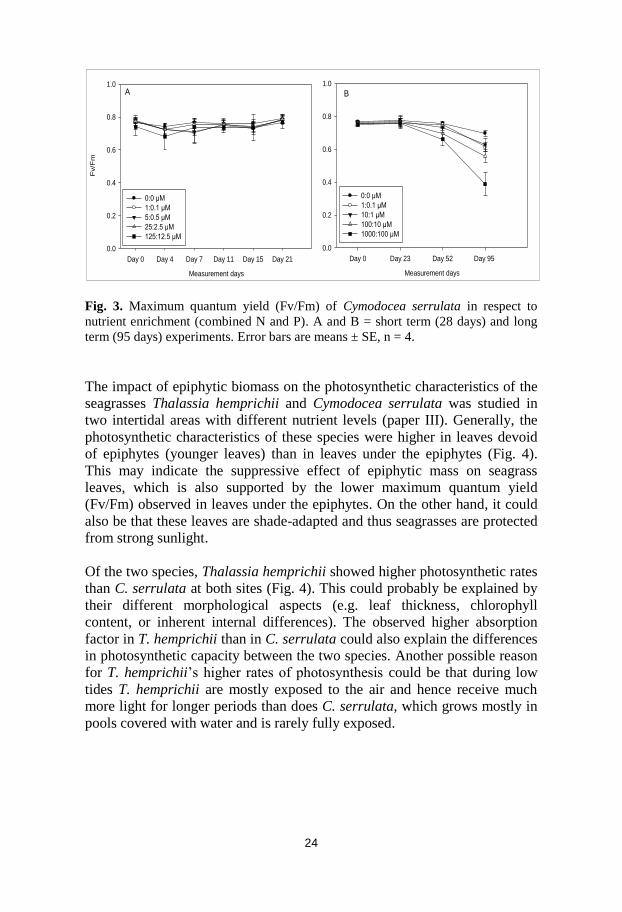
of PSII during long-term enrichment (Fig. 3, paper II). This could probably
be associated with the development of nitrate toxicity (Burkholder et al.
1992), which led to a decrease in photosynthetic capacity. The levels of
nutrient additions used in this study resulted in water concentrations that
were within the range of those previously reported for islands about 3 to 5
km offshore in the area where seagrass were collected (Björk et al. 1995,
Mohammed and Mgaya 2001) and likely to be found in places influenced by
untreated sewage.
Site Species Shoot density
(no. shoots m-2
)
Canopy
height (cm)
Above ground
biomass (gDWm-2
)
Below ground
biomass (gDWm-2
)
Above to below-
ground (AG/BG)
biomass ratios
Dry weight of
epiphytes (gg-1
)
Mjimwema T. hemprichii 1038.1 ± 288.4 14.8 ± 2.1 267.0 ± 43.8 1177.4 ± 265.2
0.23 ± 0.03 0.08 ± 0.01
C. serrulata 1622.7 ± 304.5 16.7 ± 1.8 352.2 ± 144.7 737.2 ± 260.8 0.51 ± 0.21 0.07 ± 0.04
Ocean Road T. hemprichii 792.0 ± 84.1 17.6 ± 2.7 307.0 ± 74.9 412.1 ± 93.3 0.77 ± 0.15 0.25 ± 0.04
C. serrulata 819.8 ± 203.8 14.1 ± 2.0 202.7 ± 69.6 267.7 ± 147.9 0.81 ± 0.13 0.34 ± 0.29
24
Fig. 3. Maximum quantum yield (Fv/Fm) of Cymodocea serrulata in respect to
nutrient enrichment (combined N and P). A and B = short term (28 days) and long
term (95 days) experiments. Error bars are means ± SE, n = 4.
The impact of epiphytic biomass on the photosynthetic characteristics of the
seagrasses Thalassia hemprichii and Cymodocea serrulata was studied in
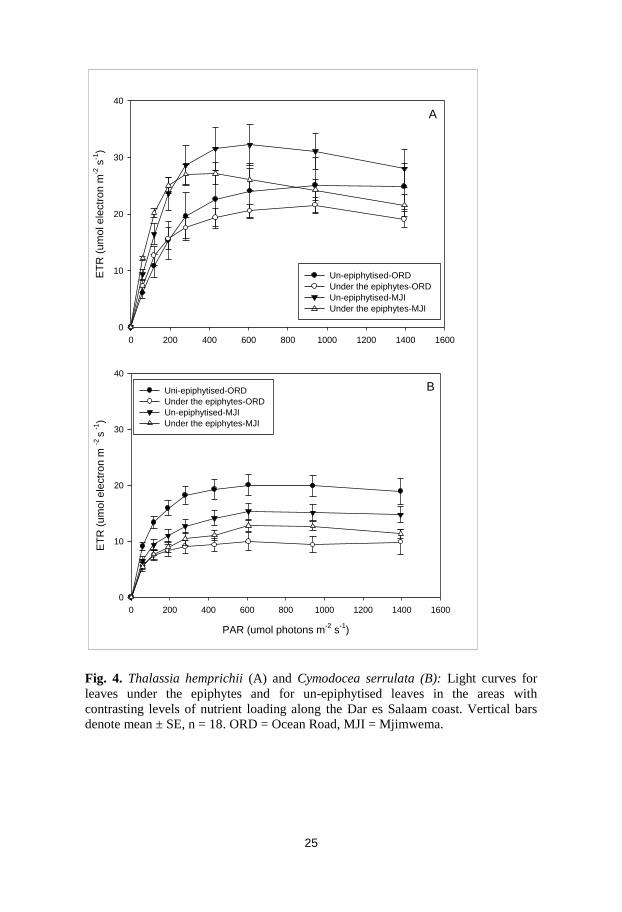
two intertidal areas with different nutrient levels (paper III). Generally, the
photosynthetic characteristics of these species were higher in leaves devoid
of epiphytes (younger leaves) than in leaves under the epiphytes (Fig. 4).
This may indicate the suppressive effect of epiphytic mass on seagrass
leaves, which is also supported by the lower maximum quantum yield
(Fv/Fm) observed in leaves under the epiphytes. On the other hand, it could
also be that these leaves are shade-adapted and thus seagrasses are protected
from strong sunlight.
Of the two species, Thalassia hemprichii showed higher photosynthetic rates
than C. serrulata at both sites (Fig. 4). This could probably be explained by
their different morphological aspects (e.g. leaf thickness, chlorophyll
content, or inherent internal differences). The observed higher absorption
factor in T. hemprichii than in C. serrulata could also explain the differences
in photosynthetic capacity between the two species. Another possible reason
for T. hemprichii’s higher rates of photosynthesis could be that during low
tides T. hemprichii are mostly exposed to the air and hence receive much
more light for longer periods than does C. serrulata, which grows mostly in
pools covered with water and is rarely fully exposed.
A
Measurement days
Day 0 Day 4 Day 7 Day 11 Day 15 Day 21
Fv/F
m
0.0
0.2
0.4
0.6
0.8
1.0
0:0 µM
1:0.1 µM
5:0.5 µM
25:2.5 µM
125:12.5 µM
B
Measurement days
Day 0 Day 23 Day 52 Day 95
0.0
0.2
0.4
0.6
0.8
1.0
0:0 µM
1:0.1 µM
10:1 µM
100:10 µM
1000:100 µM
25
Fig. 4. Thalassia hemprichii (A) and Cymodocea serrulata (B): Light curves for
leaves under the epiphytes and for un-epiphytised leaves in the areas with
contrasting levels of nutrient loading along the Dar es Salaam coast. Vertical bars
denote mean ± SE, n = 18. ORD = Ocean Road, MJI = Mjimwema.
A
0 200 400 600 800 1000 1200 1400 1600
ET
R (
um
ol ele
ctr
on m
-2 s
-1)
0
10
20
30
40
Un-epiphytised-ORD
Under the epiphytes-ORD
Un-epiphytised-MJI
Under the epiphytes-MJI
B
PAR (umol photons m-2
s-1
)
0 200 400 600 800 1000 1200 1400 1600
ET
R (
um
ol e
lectr
on m
-2 s
-1)
0
10
20
30
40
Uni-epiphytised-ORD
Under the epiphytes-ORD
Un-epiphytised-MJI
Under the epiphytes-MJI

26
4.1.2 Effects on epiphyte biomass and composition
The occurrence and abundance of epiphytes on seagrass leaves has been
shown to vary depending on environmental conditions (Hays 2005, Ruíz et
al. 2009). Epiphytic composition and biomass varied in relation to different
levels of nutrient in both laboratory and field experiments (papers II and III).
In the nutrient enrichment experiments, epiphyte biomass did not vary
significantly during short term exposure to increased levels of nutrients.
However, in the long term experiments, epiphyte biomass varied
significantly in relation to nutrient added (paper II). Moreover, in the in situ
experiment, higher epiphyte biomass was observed for both species at the
nutrient rich-site than at the nutrient-poor site, indicating that epiphytes
respond more quickly to nutrient availability than their host seagrasses. A
similar trend has been reported in many studies (e.g. Hauxwell et al. 2003,
Hays 2005, McGlathery 2001).
The composition of epiphytes varied between species and between sites.
Cymodocea serrulata had more epiphytic species at the nutrient-poor site,
while T. hemprichii had more species at the nutrient-rich site (paper III),
probably indicating the specific preferences of the various epiphytes for
different structural and physical-chemical factors (Frankovich and
Fourqurean 1997). In general, there were more cyanobacteria species at the
nutrient-poor site than at the nutrient-rich site, indicating their contribution
to sustained nitrogen demand in this area, supporting the previous
observation by Hamisi et al. (2004).
4.1.3 Effects on nutrient contents of plant tissues
The common response of the plants to nutrient fluctuation is storage of
respective nutrient in plant tissues (Romero et al. 2006), however the
response seems to vary between species. There was no clear pattern in
nutrient content in relation to the levels of nutrient added in the flow-through
experiments (paper II), even though an increase in %N and %P content of
the leaves was observed following nutrient addition in all treatments.
Changes in nutrient contents were also observed in the control plants,
reflecting the fact that the nutrient concentration in the incoming water was
higher than concentrations in the habitat from which they were collected.
The in situ study showed variation in %C, %N, and %P content both within
and between species and between sites (paper III). Percentages of N and P,
as well as C:N:P ratios, correlated well with the nutrient concentration in the
water column, indicating more availability of dissolved nutrient levels at
Ocean Road than at Mjimwema, and hence increased acquisition and
retention in the plant parts (Duarte 1990, Peréz-Lloréns and Niell 1995,
27
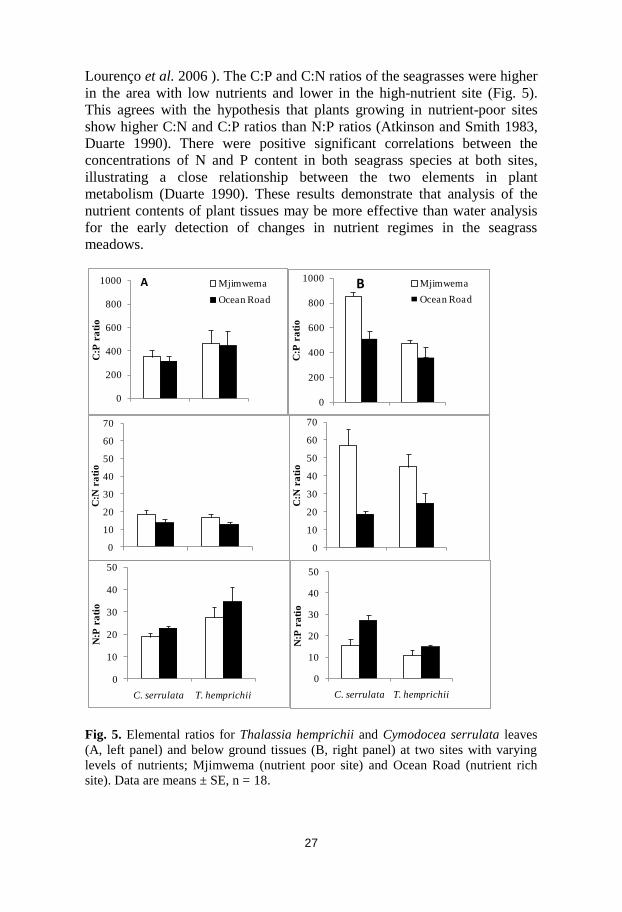
Lourenço et al. 2006 ). The C:P and C:N ratios of the seagrasses were higher
in the area with low nutrients and lower in the high-nutrient site (Fig. 5).
This agrees with the hypothesis that plants growing in nutrient-poor sites
show higher C:N and C:P ratios than N:P ratios (Atkinson and Smith 1983,
Duarte 1990). There were positive significant correlations between the
concentrations of N and P content in both seagrass species at both sites,
illustrating a close relationship between the two elements in plant
metabolism (Duarte 1990). These results demonstrate that analysis of the
nutrient contents of plant tissues may be more effective than water analysis
for the early detection of changes in nutrient regimes in the seagrass
meadows.
Fig. 5. Elemental ratios for Thalassia hemprichii and Cymodocea serrulata leaves
(A, left panel) and below ground tissues (B, right panel) at two sites with varying
levels of nutrients; Mjimwema (nutrient poor site) and Ocean Road (nutrient rich
site). Data are means ± SE, n = 18.
0
200
400
600
800
1000
C:P
ra
tio
Mjimwema
Ocean Road
A
0
10
20
30
40
50
60
70
C:N
ra
tio
0
10
20
30
40
50
C. serrulata T. hemprichii
N:P
ra
tio
0
200
400
600
800
1000
C:P
ra
tio
Mjimwema
Ocean Road
B
0
10
20
30
40
50
60
70
C:N
ra
tio
0
10
20
30
40
50
C. serrulata T. hemprichii
N:P
ra
tio
28
4.2 Ulva overgrowth affects seagrass photosynthesis
Seagrass photosynthesis depends upon light penetrating through the water
column to reach the seagrass at the bottom. However, much light is lost
before it reaches the seagrass. Shading by macroalgae blooms attributable to
eutrophication has been implicated as a major cause of seagrass decline
(Short et al. 1995). Results in this thesis (paper I) conform to the previous
studies, by showing that shading by an algal thallus resulted in a significant
reduction of available irradiance (Table 2). This indicates that, in the natural
eutrophic environment where other factors contribute to light attenuation, the
situation is likely to be intensified.
Table 2. Percentage reduction of light when passing through Ulva
intestinalis thalli and absorption factor of Zostera marina in that light (Mean
± S.E). ND = not determined.
In addition to reducing light reaching the seagrasses, Ulva thalli also affects
the quality of that light by absorbing much of the useful spectrum, resulting
in filtered light mainly in the green area of the spectrum, which is considered
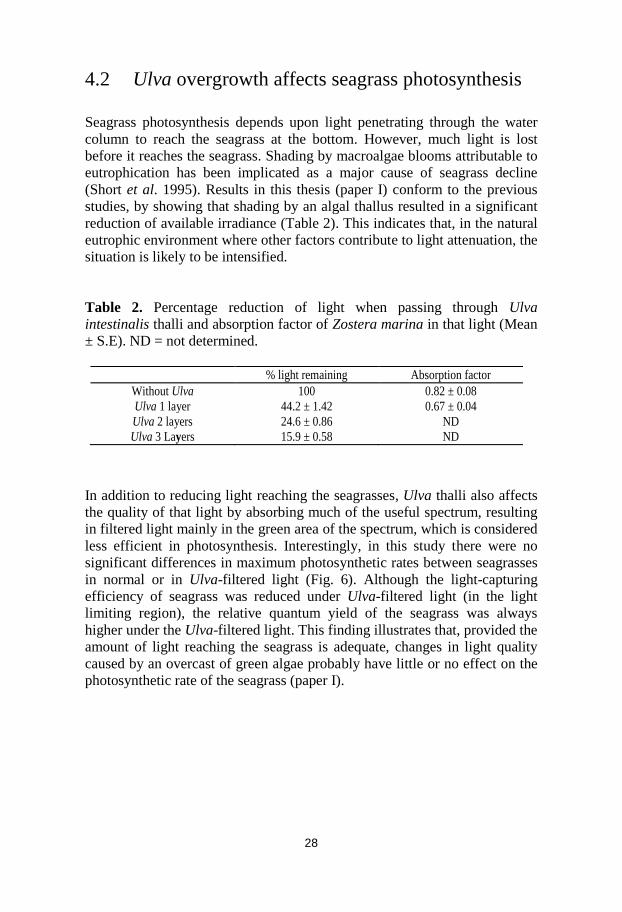
less efficient in photosynthesis. Interestingly, in this study there were no
significant differences in maximum photosynthetic rates between seagrasses
in normal or in Ulva-filtered light (Fig. 6). Although the light-capturing
efficiency of seagrass was reduced under Ulva-filtered light (in the light
limiting region), the relative quantum yield of the seagrass was always
higher under the Ulva-filtered light. This finding illustrates that, provided the
amount of light reaching the seagrass is adequate, changes in light quality
caused by an overcast of green algae probably have little or no effect on the
photosynthetic rate of the seagrass (paper I).
% light remaining Absorption factor
Without Ulva 100 0.82 ± 0.08
Ulva 1 layer 44.2 ± 1.42 0.67 ± 0.04
Ulva 2 layers 24.6 ± 0.86 ND
Ulva 3 Layers 15.9 ± 0.58 ND
29
Fig. 6. Zostera marina. Electron transport rate in relation to irradiance levels under
normal (filled circle) and Ulva-filtered light (open circle). Vertical bars represent the
standard error, n = 40.
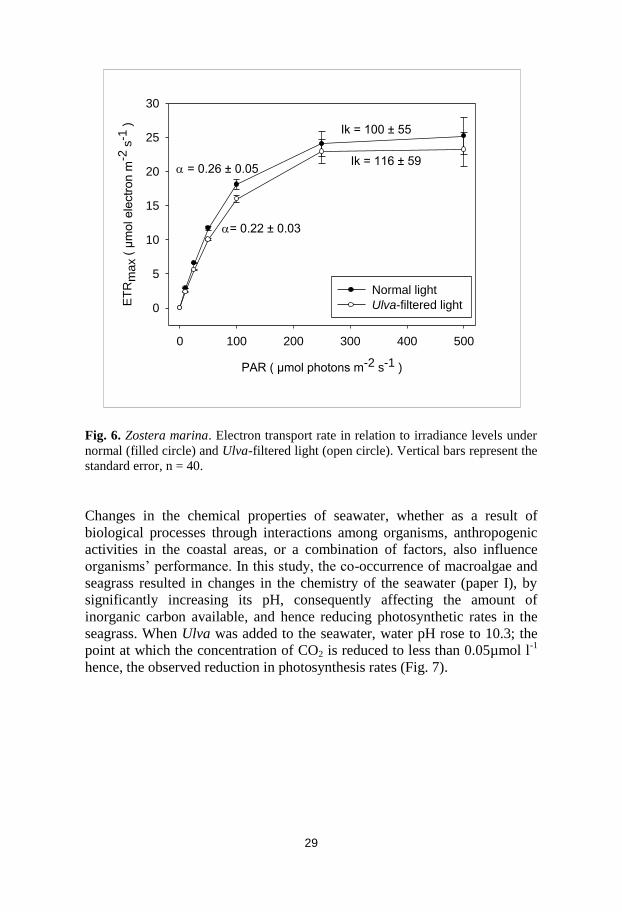
Changes in the chemical properties of seawater, whether as a result of
biological processes through interactions among organisms, anthropogenic
activities in the coastal areas, or a combination of factors, also influence
organisms’ performance. In this study, the co-occurrence of macroalgae and
seagrass resulted in changes in the chemistry of the seawater (paper I), by
significantly increasing its pH, consequently affecting the amount of
inorganic carbon available, and hence reducing photosynthetic rates in the
seagrass. When Ulva was added to the seawater, water pH rose to 10.3; the
point at which the concentration of CO2 is reduced to less than 0.05µmol l-1
hence, the observed reduction in photosynthesis rates (Fig. 7).
PAR ( µmol photons m-2 s-1 )
0 100 200 300 400 500
ET
Rm
ax (
µm
ol e
lectr
on
m-2
s-1
)
0
5
10
15
20
25
30
Normal light
Ulva-filtered light
Ik = 100 ± 55
Ik = 116 ± 59
= 0.26 ± 0.05
= 0.22 ± 0.03
30
Fig. 7. Photosynthetic rates of Zostera marina in relation to changes in seawater pH
induced by addition of Ulva intestinalis thalli under normal (closed circles, solid
trendline) and Ulva-filtered (open circles, dashed trendline) lights. Data points are
averages of different measurements pooled together, n = 41.
4.3 Mussels enhance seagrass photosynthesis
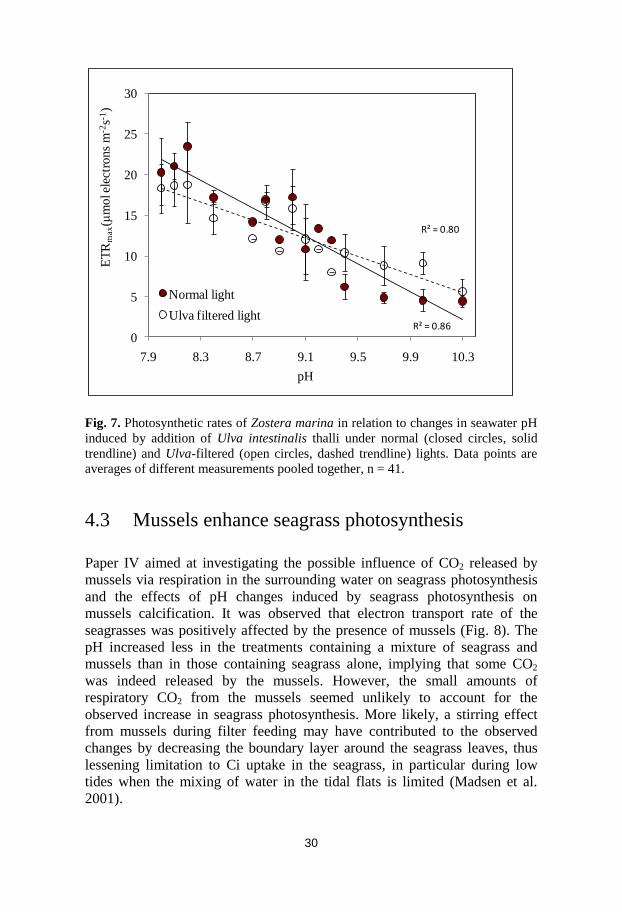
Paper IV aimed at investigating the possible influence of CO2 released by
mussels via respiration in the surrounding water on seagrass photosynthesis
and the effects of pH changes induced by seagrass photosynthesis on
mussels calcification. It was observed that electron transport rate of the
seagrasses was positively affected by the presence of mussels (Fig. 8). The
pH increased less in the treatments containing a mixture of seagrass and
mussels than in those containing seagrass alone, implying that some CO2
was indeed released by the mussels. However, the small amounts of
respiratory CO2 from the mussels seemed unlikely to account for the
observed increase in seagrass photosynthesis. More likely, a stirring effect
from mussels during filter feeding may have contributed to the observed
changes by decreasing the boundary layer around the seagrass leaves, thus
lessening limitation to Ci uptake in the seagrass, in particular during low
tides when the mixing of water in the tidal flats is limited (Madsen et al.
2001).
R² = 0.86
R² = 0.80
0
5
10
15
20
25
30
7.9 8.3 8.7 9.1 9.5 9.9 10.3
ET
Rm
ax(μ
mol e
lect
rons
m-2
s-1)
pH
Normal light
Ulva filtered light
31
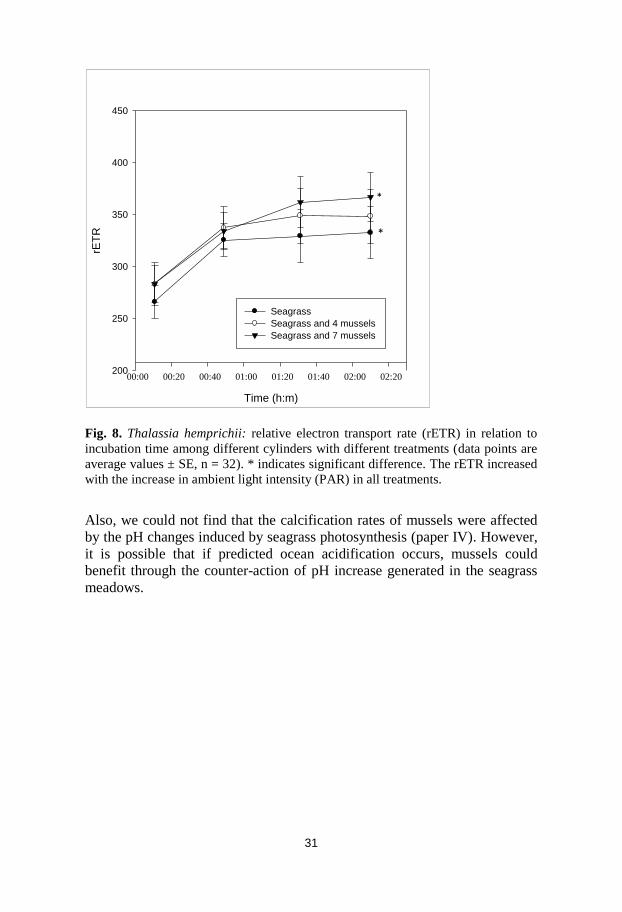
Fig. 8. Thalassia hemprichii: relative electron transport rate (rETR) in relation to
incubation time among different cylinders with different treatments (data points are
average values ± SE, n = 32). * indicates significant difference. The rETR increased
with the increase in ambient light intensity (PAR) in all treatments.
Also, we could not find that the calcification rates of mussels were affected
by the pH changes induced by seagrass photosynthesis (paper IV). However,
it is possible that if predicted ocean acidification occurs, mussels could
benefit through the counter-action of pH increase generated in the seagrass
meadows.
Time (h:m)
00:00 00:20 00:40 01:00 01:20 01:40 02:00 02:20
rET
R
200
250
300
350
400
450
Seagrass
Seagrass and 4 mussels
Seagrass and 7 mussels
*
*

32
5. Conclusion
The results presented in this thesis demonstrate that increased levels of
nutrients in the water column affect the performance of seagrasses, but
primarily indirectly.
Co-occurrence of green macroalgae in seagrass meadows negatively
affects the photosynthetic activity of seagrasses through competition for
available CO2. On the other hand, photosynthetic capacity of seagrasses
was boosted in the presence of mussels, while seagrass photosynthesis
had no observable effects on mussel calcification.
Nutrient content of the seagrass tissues reflect the nutrient
concentrations in the water column of the studied sites. This suggests
analysis of plant tissues as a reliable method for monitoring changes in
the nutrient status of seagrass ecosystems in regions where such data are
scarce.
Photosynthetic capacity of seagrasses is affected by long term exposure
to enhanced nutrient levels and the subsequent overgrowth of micro- and
macro-epiphytes, that cause substantial reductions of light available for
seagrass photosynthesis.
This thesis contributes to the understanding of multiple effects of nutrient
over-enrichment on seagrass performance, with focus on the Western Indian
Ocean region, where seagrass ecophysiological studies are still scarce.
Nutrient over-enrichments induce multiple indirect effects on seagrass
functioning, which act singly or in combination. Because the borderline
between direct and indirect effects of nutrient over-enrichment in the field is
challenging to define, major efforts should be directed towards minimizing
the rates of nutrient loading in coastal environments and monitoring nutrient
flux in coastal environments through analyses of water and plant tissues.
These efforts must be made before remarkable harmful effects are evident, to
reduce or prevent their impact and to predict their future responses to
unanticipated natural impacts.

33
In the future....
As nutrient loading and CO2 emission is intensified through anthropogenic
activities, it will be important to investigate the combined effects of
increased nutrients and CO2 in seawater on the physiological responses of
seagrasses. Changes in the chemical properties of seawater may influence
the availability of nutrients.
To evaluate the functioning of the key enzymes involved in nutrient
assimilation in seagrasses in response to the changing CO2 status of the
marine environments. This could be addressed by measuring the activity of
enzymes involved in nutrient assimilation in relation to varying levels of
CO2 concentrations.
There exists limited research on seagrass physiological response in the
Western Indian Ocean region; thus, there is a need to study other seagrass
species within the region in regard to the interactive effects of nutrients and
associated epiphytic loads on seagrass photosynthesis and consequent effects
on the functioning of the below-ground parts.
The present study showed the importance of mussels and seagrass
interaction in a short-term experiment. It will be important in future to
continue to assess the long-term influences of mussels assemblages in the
seagrass beds.

34
Acknowledgements
First and foremost, praise goes to God, for blessing me with the wellbeing,
courage, strength, and endurance to finally finish this work.
To my supervisor, Prof. Mats Björk: Thank you for bringing me into the
world of sciences. Your constructive ideas, critical approaches to data
interpretation, and tireless guidance were invaluable to the accomplishment
of this work.
To my co-supervisor, Dr. Thomas J. Lyimo: Thank you for introducing
me to the world of marine sciences. Through you I found myself in the
middle of intertidal seagrass meadows! I appreciate your support.
I also extend my sincere gratitude to Prof. Birgitta Bergman for accepting
me as a PhD student in the Physiology section, Botany Department,
Stockholm University and for your valuable comments on my thesis.
My scholarship was provided by the Swedish government through the
Swedish Agency for Research Cooperation (Sida/SAREC) marine bilateral
programme, to which I am sincerely grateful.
Also my sincere appreciation goes to the Director, Coordinator, and
Heads at the University of Dar es Salaam, Institute of Marine Sciences, who
made funds available whenever needed.
Many thanks to the managements of the Western Indian Ocean Marine
Science Association (WIOMSA) and the International Foundation for
Sciences (IFS). Through your support I was able to attend different courses,
workshops, and conferences, through which I learnt new things and met new
people in my life. Without your support, I could not have managed.
To my colleagues, students, and friends at the University of Dar es
Salaam Departments of Botany, Zoology and Wildlife Conservation,
Aquatic Sciences, Molecular Biology and Biotechnology, and the Institute of
Marine Sciences: Thank you. You have made my journey very enjoyable
and colourful.
To my fellow students, group mates, friends, group leaders, and
everybody in the physiology section of the Botany Department, Stockholm
University: Thank you very much for sharing all the moments that finally I
accomplish this study. The working environment was very pleasant because
of you.

35
Outside of academia, I wish to extend my sincere gratitude to my family,
to my sisters, brothers, aunts, uncles, and in-laws for your love, prayers, and
encouragements. You were always there for me.
My sincere appreciation and special thanks goes to my sister Stella, who
was a real mother to my daughter in my absence. She was so little when I
left her, but she never felt my absence because you were always there for
her. Mungu akubariki!
To my daughter Doreen and your pal Elisha: You are so adorable I could
feel it even when we were far apart. Thank you for your understanding and
for your prayers! It is good that the journey is over and I can spend some
more time with you.
To my husband Denis: you’re the most important and unique gift in my
life. Your love, passion, care, patience, enthusiasm, and encouragements
defined my path. Thanks for being such a great father and companion to our
daughter when I was away.
It is impossible to mention every person who contributed to the successful
accomplishment of this study – that could be another thesis of its own! but I
extend my sincere appreciation to every person (in or out of academia) who
in one way or another facilitated the successful completion of this thesis.
God bless you all!

36
References
Ackerman JD (2006) Sexual reproduction of seagrasses: pollination in the
marine context. In: Larkum AWD, Orth RJ, Duarte CM (Eds.),
Seagrasses: biology, ecology and conservation. Springer, the Netherlands,
pp 89-109.
Alcoverro T, Romero J, Duarte CM, Lopez NI (1997) Spatial and temporal
variations in nutrient limitation of seagrass Posidonia oceanica growth in
the NW Mediterranean. Mar Ecol Prog Ser 146, 155-161.
Anderson DH, Robinson RJ (1946) Rapid electrometric determination of the
alkalinity of seawater. Ind Engin Chem Anal Ed 18:767-769
Atkinson JB, Smith A (1983) C:N:P ratios of benthic marine plants. Limnol
Oceanogr 28, 568-574.
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesis in vivo.
Annu Rev Plant Biol 59, 89–113
Beer S, Björk M (2000) Measuring rates of photosynthesis of two tropical
seagrasses by pulse amplitude modulated (PAM) fluorometry. Aquat Bot
66, 69-76.
Beer S, Björk M, Gademann R, Ralph P (2001) Measurements of
photosynthetic rates in seagrasses. In: Short FT, Coles RG (Eds.), Global
Seagrass Research Methods. Elsevier, New York, pp 183–198.
Beer S, Björk M, Hellblom F, Axelsson L (2002) Inorganic carbon utilization
in marine angiosperms (seagrasses). Funct Plant Biol 29, 349-354.
Beer S, Mtolera M, Lyimo T, Björk M (2006) The photosynthetic
performance of the tropical seagrass Halophila ovalis in the upper
intertidal. Aquat Bot 84, 367-371.
Beer S, Rehnberg J (1997) The acquisition of inorganic carbon by the
seagrass Zostera marina. Aquat Bot 56, 277-283.
Björk M, Axelsson L, Beer S (2004) Why is Ulva intestinalis the only
macroalga inhabiting isolated rockpools along the Swedish Atlantic coast?
Mar Ecol Prog Ser 284, 109-116.

37
Björk M, Mohammad SM, Björklund M, Semesi A (1995) Coralline algae,
important coral reef builders threatened by pollution. Ambio 24, 502-505.
Björk M, Weil A, Semesi S, Beer S (1997) Photosynthetic utilisation of
inorganic carbon by seagrasses from Zanzibar, East Africa. Mar Biol 129,
363-366.
Borowitzka MA, Lavery PS, van Keulen M (2006) Epiphytes of seagrasses.
In: Larkum AWD, Orth RJ, Duarte CM (Eds.), Seagrasses: biology,
ecology and conservation. Springer, the Netherlands: pp 441-461.
Borum J, Greve TM (2004) The four European seagrass species. In: Borum J,
Duarte CM, Krause-Jensen D, Greve TM (Eds.), European Seagrasses: an
introduction to monitoring and management. The M & MS project. pp 1–
7.
Borum J, Sand-Jensen K (1996) Is total primary production in shallow coastal
marine waters stimulated by nitrogen loading? Oikos 76, 406-410.
Brun FG, Hernandez I, Vergara JJ, Perez-Llorens JL (2003) Growth, carbon
allocation and proteolytic activity in the seagrass Zostera noltii shaded by
Ulva canopies. Funct Plant Biol 30, 551-560.
Bulthuis DA, Axelrad DM, Mickelson MJ (1992) Growth of the seagrass
Heterozostera tasmanica limited by nitrogen in Port Phillip Bay, Australia.
Mar Ecol Prog Ser 89, 269-275.
Burkholder JM, Tomasko DA, Touchette BW (2007) Seagrasses and
eutrophication. J Exp Mar Biol Ecol 350, 46-72.
Burkholder JM, Mason KM, Glasgow HBJ (1992) Water-column nitrate
enrichment promotes decline of eelgrass Zostera marina: Evidence from
seasonal mesocosm experiments. Mar Ecol Prog Ser 81, 163–178
Chisholm JRM (2000) Calcification by crustose coralline algae on the
northern Great Barrier Reef, Australia. Limnol Oceanogr 45, 1476-1484.
de la Torre-Castro, M., Rönnbäck, P., 2004. Links between humans and
seagrasses – an example from tropical East Africa. Ocean Coast Manage
47, 361-387.
Drechsler Z, Sharkia R, Cabantchik ZI, Beer S (1993) Bicarbonate uptake in
the marine macroalga Ulva sp. is inhibited by classical probes of anion
exchange by red blood cells. Planta 191, 34-40.
Duarte CM (1990) Seagrass nutrient content. Mar Ecol Prog Ser 67, 201-207.

38
Duarte CM, Chiscano CL (1999) Seagrass biomass and production: a
reassessment. Aquat Bot 65, 159-174.
Duarte CM, Dennison WC, Orth RJW, Carruthers TJB (2008) The charisma
of coastal ecosystems: addressing the imbalance. Estuar Coast 31, 233-
238.
Duarte CM, Gattuso JP (2010) Seagrass meadows. In: Encyclopedia of earth.
Eds. Cutler J. Cleveland (Washington, D.C.: Environmental Information
Coalition, National Council for Science and the Environment).
http://www.eoearth.org/article/Seagrass_meadows.
Fabry VJ, Seibel BA, Feely RA, Orr JC (2008) Impacts of ocean acidification
on marine fauna and ecosystem processes. Ices J Mar Sci 65, 414-432.
Falkowski PG, Raven JA (2007) Aquatic photosynthesis. Princeton University
Press, New Jersey.
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Primary
production of the biosphere: integrating terrestrial and oceanic
components. Science 281, 237-240.
Fourqurean JW, Zieman JC, Powell GVN (1992) Phosphorus limitation of
primary production in Florida Bay evidence from CNP ratios of the
dominant seagrass Thalassia testudinum. Limnol Oceanogr 37, 162-171.
Frankovich TA, Fourqurean JW (1997) Seagrass epiphyte loads along a
nutrient availability gradient, Florida Bay, USA. Mar Ecol Prog Ser 159,
37-50.
Genty B, Briantais JM, Baker NR (1989) The relationship between the
quantum yield of photosynthetic electron transport and quenching of
chlorophyll fluorescence. Biochimi Biophys Acta 990, 87-92.
Gras AF, Koch MS, Madden CJ (2003) Phosphorus uptake kinetics of a
dominant tropical seagrass Thalassia testudinum. Aquat Bot 76, 299-315.
Green EP, Short FT (2003) World atlas of seagrasses. California University
Press, California.
Hamisi MI, Lyimo TJ, Muruke MHS, Bergman B (2009) Nitrogen fixation by
epiphytic and epibenthic diazotrophs associated with seagrass meadows
along the Tanzanian coast, Western Indian Ocean. Aquat Microb Ecol 57:
33-42

39
Hauxwell J, Cebrian J, Valiela I (2003) Eelgrass Zostera marina loss in
temperate estuaries: relationship to land-derived nitrogen loads and effect
of light limitation imposed by algae. Mar Ecol Prog Ser 247, 59-73.
Hays CG (2005) Effect of nutrient availability, grazer assemblage and
seagrass source population on the interaction between Thalassia
testudinum (turtle grass) and its algal epiphytes. J Exp Mar Biol Ecol 314,
53-68.
Heck KL, Valentine JF (2006) Plant-herbivore interactions in seagrass
meadows. J Exp Mar Biol Ecol 330, 420-436.
Heiss WM, Smith AM, Probert PK (2000) Influence of the small intertidal
seagrass Zostera novazelandica on linear water flow and sediment texture.
New Zeal J Mar Fresh 34, 689-694.
Hemminga MA, Duarte CM (2000) Seagrass ecology. Cambridge University
Press, Cambridge.
Holmer M, Olsen AB (2002) Role of decomposition of mangrove and
seagrass detritus in sediment carbon and nitrogen cycling in a tropical
mangrove forest. Mar Ecol Prog Ser 230, 87-101.
Invers O, Romero J, Perez M (1997) Effects of pH on seagrass
photosynthesis: a laboratory and field assessment. Aquat Bot 59, 185-194.
Invers O, Zimmerman RC, Alberte RS, Perez M, Romero J (2001) Inorganic
carbon sources for seagrass photosynthesis: an experimental evaluation of
bicarbonate use in species inhabiting temperate waters. J Exp Mar Biol
Ecol 265, 203-217.
Jassby AD, Platt T (1976) Mathematical formulation of relationship between
photosynthesis and light for phytoplankton. Limnol Oceanogr 21, 540-547.
Johnson MW, Heck JKL, Fourqurean JW (2006) Nutrient content of
seagrasses and epiphytes in the northern Gulf of Mexico: evidence of
phosphorus and nitrogen limitation. Aquat Bot 85, 103-111.
Kautsky H, Appel W, Amann H (1960) Chlorophyllfluoreszenz und
kohlenäureassimilation. Biochemisches zeiterschrift 322, 277-292.
Kilminster KL, Walker DI, Thompson PA, Raven JA (2006) Limited
nutritional benefit to the seagrass Halophila ovalis, in culture, following
sediment organic matter enrichment. Est Coast Shelf Sci 68, 675-685.

40
Kotta J, Møhlenberg F (2002) Grazing impact of Mytilus edulis L. and
Dreissena polymorpha (Pallas) in the Gulf of Riga, Baltic Sea estimated
from biodeposition rates of algal pigments. Ann Zool Fenn 39, 151-160.
Krause GH, Weiss E (1991) Chlorophyll fluorescence and photosynthesis: the
basics. Annu Rev Plant Phys 42, 313-349.
Kuo J, den Hartog C (2006) Seagrass morphology, anatomy, and
ultrastructure. In: Larkum AWD, Orth RJ, Duarte CM (Eds.), Seagrasses:
biology, ecology and conservation. Springer, the Netherlands, pp51-87.
Lamonte M, Dunton KH (2006) Effects of drift macroalgae and light
attenuation on chlorophyll fluorescence and sediment sulfides in the
seagrass Thalassia testudinum. J Exp Mar Biol Ecol 334:174–186.
Larkum AWD, Drew EA, Ralph PJ (2006) Photosynthesis and metabolism of
seagrasses at the cellular level. In Seagrasses: Biology, ecology and
conservation, eds. W.D. Larkum, R.J. Orth, and C.M. Duarte, 323–345.
Dordrecht, The Netherlands: Springer.
Lauringson V, Malton E, Kotta J, Kangur K, Orav-Kotta H, Kotta I (2007)
Environmental factors influencing the biodeposition of the suspension
feeding bivalve Dreissena polymorpha (Pallas): comparison of brackish
and freshwater populations. Est Coast Shelf Sci 75, 459-467.
Lee KS, Park SR, Kim YK (2007) Effects of irradiance, temperature, and
nutrients on growth dynamics of seagrasses: a review. J Exp Mar Biol Ecol
350, 144-175.
Liu SL, Wang WL, Dy DT, Fu CC (2005) The effect of ulvoid macroalgae on
the inorganic carbon utilization by an intertidal seagrass Thalassia
hemprichii. Bot Bulle Acad Sinica 46, 197-203.
Lourenço SO, Barbarino E, Nascimento A, Freitas JNP, Diniz GS (2006)
Tissue nitrogen and phosphorus in seaweeds in a tropical eutrophic
environment: What a long-term study tells us. J Appl Phycol 18: 389-98.
Martins I, Marques JC (2002) A model for the growth of opportunistic
macroalgae (Enteromorpha sp.) in tidal estuaries. Est Coast Shelf Sci 55,
247-257.
Maxwell K, Johnson GL (2000) Chlorophyll fluorescence: a practical guide. J
Exp Bot 51(345), 659-668.
Mazzella L, Alberte RS (1986) Light adaptation and the role of autotrophic
epiphytes in primary production of the temperate seagrass, Zostera marina
L. J Exp Mar Biol Ecol 100, 165-180.

41
McGlathery KJ (2001) Macroalgal blooms contribute to the decline of
seagrass in nutrient-enriched coastal waters. J Phycol 37,453-456.
Mohammed MS, Mgaya YD (2001) Nutrients and their dynamics in the coral
reefs off Zanzibar town. In: Richmond M, Francis J (eds), Marine science
development in Tanzania and eastern Africa. Proceedings of the 20th
Anniversary Conference on Advances in Marine Sciences in Tanzania.
Institute of Marine Sciences. University of Dar es Salaam and Western
Indian Ocean Marine Science Association, Zanzibar, Tanzania, pp 171-
183.
Moore KA, Wetzel RL (2000) Seasonal variations in eelgrass (Zostera marina
L.) responses to nutrient enrichment and reduced light availability in
experimental ecosystems. J Exp Mar Biol Ecol 244, 1-28.
Moriarty DJW, O’Donohue MJ (1993) Nitrogen-fixation in seagrass
communities during summer in the Gulf of Carpentaria, Australia. Aust J
Mar Freshw Res 44, 117-125.
Newell RIE (2004) Ecosystem influences of natural and cultivated
populations of suspension-feeding bivalve molluscs: a review. J Shellfish
Res 23:51-61.
Nielsen OI, Koch MS, Jensen HS, Madden CJ (2006) Thalassia testudinum
phosphate uptake kinetics at low in situ concentrations using a IT
radioisotope technique. Limnol Oceanogr 51, 208-217.
Oliviera EC, Österlund K, Mtolera MSP (2005). Marine plants of Tanzania. A
field guide to seaweeds and seagrasses. Stockholm University, Sweden,
229-236 pp.
Orth RJ, Carruthers TJB, Dennison WC, Duarte CM, Fourqurean JW, Heck
KL, Hughes AR, Kendrick GA, Kenworthy WJ, Olyarnik S, Short FT,
Waycott M, Williams SL (2006) A global crisis for seagrass ecosystems.
Bioscience 56, 987-996.
Palacios SL, Zimmerman RC (2007) Response of eelgrass Zostera marina to
CO2 enrichment: possible impacts of climate change and potential for
remediation of coastal habitats. Mar Ecol Prog Ser 344, 1-13.
Parsons RT, Maita Y, Lalli CM (1989) A manual of chemical and biological
methods for seawater analysis. Pergamon Press, Oxford, pp 173.
Pedersen MF, Borum J, Fotel FL (1997) Phosphorus dynamics and limitation
of fast- and slow-growing temperate seaweeds in Oslo Fjord, Norway. Mar
Ecol Prog Ser 399, 103-115.

42
Pérez-Lloréns JL, Niell FX (1995) Short-term phosphate uptake kinetics in
Zostera noltii Hornem: a comparison between excised leaves and
sediment-rooted plants. Hydrobiologia 297, 17-27.
Peterson BJ, Heck KL (1999) The potential for suspension feeding bivalves to
increase seagrass productivity. J Exp Mar Biol Ecol 240, 37-52.
Peterson BJ, Heck KL (2001a) An experimental test of the mechanism by
which suspension feeding bivalves elevate seagrass productivity. Mar Ecol
Prog Ser 218, 115-125.
Peterson BJ, Heck KL (2001b) Positive interactions between suspension-
feeding bivalves and seagrass – a facultative mutualism. Mar Ecol Prog
Ser 213, 143-155.
Ralph PJ, Gademann R (2005) Rapid light curves: a powerful tool to assess
photosynthetic activity. Aquat Bot 82, 222-237.
Richmond MD (2002). A field guide to the seashores of Eastern Africa and
the Western Indian Ocean Islands. Sida/SAREC-UDSM, Dar es Salaam,
Tanzania.
Riebesell U, Zondervan I, Rost B, Tortell PD, Zeebe RE, Morel FMM (2000)
Reduced calcification in marine plankton in response to increased
atmospheric CO2. Nature 407, 634-637.
Ries JB, Cohen AL, McCorkle DC (2009) Marine calcifiers exhibit mixed
responses to CO2-induced ocean acidification. Geology 37, 1131-1134.
Riley JP, Skirrow G (1965) Chemical oceanography. Academic Press, New
York.
Romero J, Lee KS, Pérez M, Mateo MA, Alcoverro T (2006) Nutrient
dynamics in seagrass ecosystems. In: Larkum AWD, Orth RJ, Duarte CM
(Eds.), Seagrasses: biology, ecology and conservation. Springer, the
Netherlands, pp 227–254.
Ruíz JM, Pérez M, Romero J, Tomas F (2009) The importance of herbivory in
the decline of a seagrass (Posidonia oceanica) meadow near a fish farm:
an experimental approach. Bot Mar 52, 449-458.
Sabine CL, Feely RA, Gruber N, Key RM, Lee K, Bullister JL, Wanninkhof
R, Wong CS, Wallace DWR, Tilbrook B, Millero FJ, Peng TH, Kozyr A,
Ono T, Rios AF (2004) The oceanic sink for anthropogenic CO2. Science
305, 367-371.

43
Semesi S, Beer S, Björk M (2009) Seagrass photosynthesis controls rates of
calcification and photosynthesis of calcareous macroalgae in a tropical
seagrass beds. Mar Ecol Prog Ser 382, 41-47.
Schneider K, Erez J (2006) The effect of carbonate chemistry on calcification
and photosynthesis in the hermatypic coral Acropora eurystoma. Limnol
Oceanogr 51(3), 1284-1293.
Schreiber U (2004) Pulse-amplitude (PAM) fluorometry and saturation pulse
method. In: Papageorgiou G, Govindjee (Eds.), Chlorophyll fluorescence:
a signature of photosynthesis. Advances in Photosynthesis and Respiration
Series. Kluwer Academic Publishers, Dordrecht, The Netherlands.
Schwarz AM, Bjork M, Buluda T, Mtolera H, Beer S (2000) Photosynthetic
utilisation of carbon and light by two tropical seagrass species as measured
in situ. Mar Biol 137, 755-761.
Short FT, Burdick DM, Kaldy JE (1995) Mesocosm experiments quantify the
effects of eutrophication on eelgrass, Zostera marina. Limnol Oceanogr
40, 740-749.
Short FT, Carruthers T, Dennison W, Waycott M (2007) Global seagrass
distribution and diversity: a bioregional model. J Exp Mar Biol Ecol 350,
3-20.
Short FT, Dennison WC, Capone DG (1990) Phosphorus-limited growth of
the tropical seagrass Syringodium filiforme in carbonate sediments. Mar
Ecol Prog Ser 62, 169-174.
Short FT, Neckles HA (1999) The effects of global climate change on
seagrasses. Aquat Bot 63, 169-196.
Short FT, Wyllie-Echeverria S (1996) Natural and human-induced
disturbance of seagrasses. Environ Conserv 23, 17-27.
Silva J, Santos R (2004) Can chlorophyll fluorescence be used to estimate
photosynthetic production in the seagrass Zostera noltii? J Exp Mar Biol
Ecol 307, 207-216.
Sjöö GL, Mörk E (2009) Tissue nutrient content in ulva spp. (chlorophyceae)
as bioindicator for nutrient loading along the Coast of East Africa. Open
Environ Biol Monit J 2, 11-17.
Smith SV, Kinsey DW (1978) Calcification and organic carbon metabolism as
indicated by carbon dioxide. In: Stoddart DR (ed), Coral reef: research
methods. UNESCO, Paris.

44
Smith SV, Key GS (1975) Carbon dioxide and metabolism in marine
environments. Limnology and Oceanography 20, 493-495.
Touchette BW, Burkholder JM (2000a) Review of nitrogen and phosphorus
metabolism in seagrasses. J Exp Mar Biol Ecol 250, 133-167.
Touchette BW, Burkholder JM (2000b) Overview of the physiological
ecology of carbon metabolism in seagrasses. J Exp Mar Biol Ecol 250,
169-205.
Udy JW, Dennison WC (1997) Physiological responses of seagrasses used to
identify anthropogenic nutrient inputs. Mar Freshw Res 48, 605-614.
Vergara JJ, Peralta G, Pérez-Lloréns JL, Hernández I (1997) Seasonal
variation of photosynthetic performance and light attenuation in Ulva
canopies from Palmones river estuary. J Phycol 33, 773-779.
Wall CC, Peterson BJ, Gobler CJ (2008) Facilitation of seagrass Zostera
marina productivity by suspension-feeding bivalves. Mar Ecol Prog Ser
357, 165-174.
Waycott M, Duarte CM, Carruthers TJB, Orth RJ, Dennison WC, Olyarnik S,
Calladine A, Fourqurean JW, Heck KL, Hughes AR, Kendrick GA,
Kenworthy WJ, Short FT, Williams SL (2009) Accelerating loss of
seagrasses across the globe threatens coastal ecosystems. Proc Natl Acad
Sci U S A 106, 12377-12381.
Welsh DT, Bartoli M, Nizzoli D, Castaldelli G, Riou SA, Viaroli P (2000)
Denitrification, nitrogen fixation, community primary productivity and
inorganic N and oxygen fluxes in an intertidal Zostera noltii meadow. Mar
Ecol Prog Ser 208, 65-77.
Welsh DT, Bourgues S, deWit R, Herbert RA (1996) Seasonal variations in
nitrogen-fixation (acetylene reduction) and sulphate-reduction rates in the
rhizosphere of Zostera noltii: Nitrogen fixation by sulphate reducing
bacteria. Mar Biol 125, 619-628.


















