Roles of Extracellular Matrix Components in …Roles of Extracellular Matrix Components in...
7
0 1985 hy The American Society of Biological Chemists, Inc. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 260, No. 22, Issue of October 5, pp. 12252-12258,1985 Printed in U. S. A. Roles of Extracellular MatrixComponents in Differentiating Teratocarcinoma Cells* (Received for publication, February 4,1985) Amita Grover and Eileen D. Adamson From the La Jolla Cancer Research Foundation, La Jolla, California 92037 F9 embryonal carcinoma cells treated with 5 X M retinoic acid and cultured in suspension for 8 days form aggregates consisting of an outer epithelial layer of a-fetoprotein-producing visceral endodermcells. We have previously shown (Grover, A., Oshima, R. G., and Adamson, E. D. (1983) J. Cell Biol. 96, 1690-1696) that the differentiation of F9 cells to visceral endoderm is accompanied by the activation of several genes, and increased laminin synthesis is one of the earliest events. Here we analyze in detail the syntheses and secretion of fibronectin, type IV collagen, and laminin during the 8-day process. Employing immunoprecipi- tationandenzyme-linkedimmunosorbent assay, we show that the levels of all three components change with different patterns. Unstimulated F9 cells synthe- size and secrete relatively high levels of fibronectin and low levels of type IV collagen. Fibronectin synthe- sis and secretion decreases to 10% of its original level whereas type IV collagen synthesis rises approxi- mately 3-fold during the differentiationprocess. Lam- inin synthesis also rises at least 2-fold, and the propor- tions of its subunits change as the syntheses of B1 and A accelerate starting on day 2. However, unlike fibro- nectin and type IV collagen, laminin is largely accu- mulated in the aggregates.The data suggest that fibro- nectin has a role in aggregation whereas laminin is important in the differentiationprocess. The tumorigenic stem cells of murine teratocarcinomas or embryonal carcinoma cells have the capacity to differentiate into multiple types of (non-tumorigenic) cell products (1). Embryonal carcinoma cells have proved to be good models for studies of events during early embryo development (2). One of these cell lines, F9, when stimulated by retinoic acid (3) differentiates in vitro into endodermal cell types only (4). F9 cells treated with 5 x lo-’ M retinoic acid and cultured in suspension form embryoid bodies consisting of an outer layer of epithelial cells which morphologically and biochemi- cally resemble visceral endoderm of the early postimplanta- tion mouse embryo (5, 6). Typically, these endoderm cells synthesize and secrete a-fetoprotein. The differentiation of F9 cells to visceral endoderm is an 8-day process (5, 7) in which the production of high levels of a-fetoprotein is a late event (day 7). We have previously shown that maximum synthesis and secretion of a-fetoprotein correlates with the formation of a mature epithelial layer properly aligned on a thin basement membrane (7). The organization of the outer * This work was supported by Grants HD 18772 and P30 CA 30199 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solelyto indicate this fact. epithelial layer on aggregates of differentiated F9 cells in- volves many overlapping steps consisting of early events such as increased synthesis of laminin on day 2, appearance of endodermal markers (Endo A and B) and small amounts of a-fetoprotein polypeptides on day 3, an asymmetric accumu- lation of extracellular matrix materials on day 4, and a mor- phological appearance of an outer layer of cells in aggregates on days 4-5. a-Fetoprotein secreted into the medium rises rapidly to reach high levels on day 8. The crucial phase in the differentiation process appears to be between days 2 and 4, and since increased laminin synthesis is one of the earliest events, we hypothesize that extracellular matrix production may trigger the processes leading to outer layer asymmetry, epithelial cell polarity, and visceral endoderm expression. We describe here analyses of the synthesis andsecretion of the components of extracellular matrix, laminin (8-lo), fibro- nectin (10-13), and type IV collagen (10,14) during the 8-day process. Using radiolabeling and immunoprecipitation studies together with ELISA’ (for laminin), we show that the synthe- sis and secretion of fibronectin decreases, whereas that of type IV collagen and laminin increases. A significant increase in the level of laminin is observed between days 2 and 4. Analyses of the subunits of laminin (A, B1, and B2) show a change in theproportion of B1 and B2 polypeptides between days 2 and 4 and increased synthesis of polypeptide A on day 3. The evidence suggests that fibronectin may have a role in very early events in aggregate formation, whereas laminin may play a key role in the establishment and maintenance of a basement membrane underlying the epithelium of visceral endoderm cells. MATERIALS AND METHODS Cells and Culture Conditions-B1 subclones of F9 cells were main- tained as described (7). For differentiation, the cultures were grown in DMEM:Ham’s F-12 (1:l) supplemented with 10% heat-inactivated fetal bovine serum (7). In most cases, cultures were induced with 5 X lo-’ M retinoic acid; however, later passages of F9 cells gave a similar response at 5 x lo-’ M. For testing laminin production, F9 cells were grown in 96-well, untreated plastic plates, seeding with lo3 cells/well in 0.2 ml medium. For radioactive labeling, aggregates were grown in lOO-mm, untreated plastic dishes starting with 5 X lo6 F9 cells/lO ml for 1-day cultures, 3 X lo6 cells/lO ml for 2- and 3-day cultures, and 1 X lo6 cells/lO ml for 4-, 6-, and 8-day cultures. fibronectin, absorbed with human fibronectin (151, rabbit antiserum Antisera-For immunoprecipitation, rabbitantiserum to mouse to mouse type IV collagen (16), and rabbit antiserum to rat laminin (17) were generously supplied by Dr. Eva Engvall of this institute. Rabbit antiserum to C chain of mouse laminin (18) was kindly provided by Dr. Brigid Hogan (National Institute for Medical Re- search, Mill Hill, London, United Kingdom). For ELISA, we prepared rabbit antiserum to laminin by immunizing rabbits with highly puri- fied rat laminin from L2 rat yolk sac tumors (17,191. After absorption The abbreviations used are: ELISA, enzyme-linked immunosor- bent assay; DMEM, Dulbecco’s modified Eagle’s medium; SDS, so- dium dodecyl sulfate; PMSF, phenylmethylsulfonyl fluoride. 12252
Transcript of Roles of Extracellular Matrix Components in …Roles of Extracellular Matrix Components in...

0 1985 hy The American Society of Biological Chemists, Inc. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 260, No. 22, Issue of October 5, pp. 12252-12258,1985
Printed in U. S. A.
Roles of Extracellular Matrix Components in Differentiating Teratocarcinoma Cells*
(Received for publication, February 4,1985)
Amita Grover and Eileen D. Adamson From the La Jolla Cancer Research Foundation, La Jolla, California 92037
F9 embryonal carcinoma cells treated with 5 X M retinoic acid and cultured in suspension for 8 days form aggregates consisting of an outer epithelial layer of a-fetoprotein-producing visceral endoderm cells. We have previously shown (Grover, A., Oshima, R. G., and Adamson, E. D. (1983) J. Cell Biol. 96, 1690-1696) that the differentiation of F9 cells to visceral endoderm is accompanied by the activation of several genes, and increased laminin synthesis is one of the earliest events. Here we analyze in detail the syntheses and secretion of fibronectin, type IV collagen, and laminin during the 8-day process. Employing immunoprecipi- tation and enzyme-linked immunosorbent assay, we show that the levels of all three components change with different patterns. Unstimulated F9 cells synthe- size and secrete relatively high levels of fibronectin and low levels of type IV collagen. Fibronectin synthe- sis and secretion decreases to 10% of its original level whereas type IV collagen synthesis rises approxi- mately 3-fold during the differentiation process. Lam- inin synthesis also rises at least 2-fold, and the propor- tions of its subunits change as the syntheses of B1 and A accelerate starting on day 2. However, unlike fibro- nectin and type IV collagen, laminin is largely accu- mulated in the aggregates. The data suggest that fibro- nectin has a role in aggregation whereas laminin is important in the differentiation process.
The tumorigenic stem cells of murine teratocarcinomas or embryonal carcinoma cells have the capacity to differentiate into multiple types of (non-tumorigenic) cell products (1). Embryonal carcinoma cells have proved to be good models for studies of events during early embryo development (2). One of these cell lines, F9, when stimulated by retinoic acid (3) differentiates in vitro into endodermal cell types only (4).
F9 cells treated with 5 x lo-’ M retinoic acid and cultured in suspension form embryoid bodies consisting of an outer layer of epithelial cells which morphologically and biochemi- cally resemble visceral endoderm of the early postimplanta- tion mouse embryo (5, 6). Typically, these endoderm cells synthesize and secrete a-fetoprotein. The differentiation of F9 cells to visceral endoderm is an 8-day process (5, 7 ) in which the production of high levels of a-fetoprotein is a late event (day 7). We have previously shown that maximum synthesis and secretion of a-fetoprotein correlates with the formation of a mature epithelial layer properly aligned on a thin basement membrane (7). The organization of the outer
* This work was supported by Grants HD 18772 and P30 CA 30199 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
epithelial layer on aggregates of differentiated F9 cells in- volves many overlapping steps consisting of early events such as increased synthesis of laminin on day 2, appearance of endodermal markers (Endo A and B) and small amounts of a-fetoprotein polypeptides on day 3, an asymmetric accumu- lation of extracellular matrix materials on day 4, and a mor- phological appearance of an outer layer of cells in aggregates on days 4-5. a-Fetoprotein secreted into the medium rises rapidly to reach high levels on day 8. The crucial phase in the differentiation process appears to be between days 2 and 4, and since increased laminin synthesis is one of the earliest events, we hypothesize that extracellular matrix production may trigger the processes leading to outer layer asymmetry, epithelial cell polarity, and visceral endoderm expression.
We describe here analyses of the synthesis and secretion of the components of extracellular matrix, laminin (8-lo), fibro- nectin (10-13), and type IV collagen (10,14) during the 8-day process. Using radiolabeling and immunoprecipitation studies together with ELISA’ (for laminin), we show that the synthe- sis and secretion of fibronectin decreases, whereas that of type IV collagen and laminin increases. A significant increase in the level of laminin is observed between days 2 and 4. Analyses of the subunits of laminin (A, B1, and B2) show a change in the proportion of B1 and B2 polypeptides between days 2 and 4 and increased synthesis of polypeptide A on day 3. The evidence suggests that fibronectin may have a role in very early events in aggregate formation, whereas laminin may play a key role in the establishment and maintenance of a basement membrane underlying the epithelium of visceral endoderm cells.
MATERIALS AND METHODS
Cells and Culture Conditions-B1 subclones of F9 cells were main- tained as described (7). For differentiation, the cultures were grown in DMEM:Ham’s F-12 (1:l) supplemented with 10% heat-inactivated fetal bovine serum (7). In most cases, cultures were induced with 5 X lo-’ M retinoic acid; however, later passages of F9 cells gave a similar response at 5 x lo-’ M. For testing laminin production, F9 cells were grown in 96-well, untreated plastic plates, seeding with lo3 cells/well in 0.2 ml medium. For radioactive labeling, aggregates were grown in lOO-mm, untreated plastic dishes starting with 5 X lo6 F9 cells/lO ml for 1-day cultures, 3 X lo6 cells/lO ml for 2- and 3-day cultures, and 1 X lo6 cells/lO ml for 4-, 6-, and 8-day cultures.
fibronectin, absorbed with human fibronectin (151, rabbit antiserum Antisera-For immunoprecipitation, rabbit antiserum to mouse
to mouse type IV collagen (16), and rabbit antiserum to rat laminin (17) were generously supplied by Dr. Eva Engvall of this institute. Rabbit antiserum to C chain of mouse laminin (18) was kindly provided by Dr. Brigid Hogan (National Institute for Medical Re- search, Mill Hill, London, United Kingdom). For ELISA, we prepared rabbit antiserum to laminin by immunizing rabbits with highly puri- fied rat laminin from L2 rat yolk sac tumors (17,191. After absorption
The abbreviations used are: ELISA, enzyme-linked immunosor- bent assay; DMEM, Dulbecco’s modified Eagle’s medium; SDS, so- dium dodecyl sulfate; PMSF, phenylmethylsulfonyl fluoride.
12252

Extracellular Matrix in Development 12253
with rat serum proteins, the antiserum reacted specifically with laminin.
Radioactive Labeling-F9 aggregates were washed twice in DMEM medium lacking methionine and labeled with [35S]methionine either continuously for 4 h or pulsed for 30 min at 37 "C. For continuous labeling experiments, F9 aggregates were suspended in 0.5-1.0 ml of DMEM (minus methionine) and supplemented with 10% fetal bovine serum and 50-300 pCi/ml [35S]methionine (specific activity >lo00 Ci/mmol; New England Nuclear). After incubation, the medium was collected, and 2 mM PMSF was immediately added. Aggregates were washed with phosphate-buffered saline and solubilized in 1 ml of RIPA buffer (RIPA buffer, 0.01 M Tris-HC1, pH 7.4, 0.15 M NaCl, 1% Nonidet P-40, 1% (w/v) sodium deoxycholate, 0.1% (w/v) SDS, 0.02% (w/v) sodium azide). PMSF (2 mM) was added and cell lysates were incubated for 30 min on ice. After centrifugation at 105,000 X g for 30 min at 4 "C, the cell extract was separated from the pellet. These cell extracts included both intracellular and cell-associated extracellular protein. Aliquots (2 pl) of media and cell extract were removed for determination of trichloroacetic acid-precipitable counts (described below). The remaining medium and cell extract were stored at -20 "C.
For pulse-chase experiments, F9 aggregates were preincubated for 30 min in DMEM lacking methionine and suspended in 0.5 ml of fresh DMEM supplemented with 10% dialyzed fetal bovine serum, 50 pg/ml ascorbic acid, and 500 pCi/ml [35S]methionine. After 30 min of incubation at 37 "C, culture medium was collected and aggregates were washed twice in DMEM containing 10% fetal bovine serum and 2 mM methionine. The aggregates were divided into 5 fractions of equal sizes and incubated for 0, 2, 4,8, and 10 h in 0.5 ml of DMEM supplemented with 10% fetal bovine serum and excess (2 mM) me- thionine. At the end of each incubation period, the medium was collected and the cell extracts were prepared as described above. Cell- associated fibronectin was poorly solubilized in RIPA buffer, and in some experiments F9 aggregates were solubilized with SDS (0.5% final, w/v), treated with DNase I, and boiled for 3 min (20).
The incorporation of [35S]methionine in total solubilized cellular protein and protein secreted into the medium was determined by trichloroacetic acid precipitation. Aliquots (2 pl) were dried on filters (Whatman No. 1) and incubated in 5% ice-cold trichloroacetic acid for 30 min, followed by 10% hot trichloroacetic acid (90 "C) for 5 min to hydrolyze methionine tRNA (21). The filters were subsequently washed with absolute ethanol, ethanol and ether (3:1), and finally with ether. The filters were dried and the radioactivity measured in Betamax scintillation fluid (Westchem Products).
Immunoprecipitation-For continuous labeling experiments, ali- quots of cell extracts or media consisted of equal numbers of trichlo- roacetic acid precipitable counts, and for pulse-chase experiments, aliquots consisted of equal fractions of cell extracts or media. Samples were diluted with ice-cold immunoprecipitation buffer (0.05 M Tris- HCl, pH 8.0, 0.4 M NaCl, 0.005 M EDTA, 1% Nonidet P-40, 0.02% (w/v) sodium azide, and 0.1 mM PMSF) to a final volume of 1 ml, and [35S]methionine-labeled fibronectin, laminin, and type IV colla- gen were removed sequentially. First, fibronectin was isolated on gelatin-Sepharose beads (22). Between 30 and 50 pl of washed gelatin- Sepharose was added to the fraction and mixed overnight a t 4 "C. The supernatant was separated from the beads and laminin and type IV collagen were removed in sequence by immunoprecipitation using 3-5 pl of the appropriate antiserum and 30-50 pl of Protein A-agarose (Sigma) as described (23). Gelatin-Sepharose beads failed to isolate fibronectin from cell extracts (see "Results"), and in some experi- ments antibodies were employed to extract fibronectin from cell extracts. Beads and adsorbed proteins were washed with immunopre- cipitation buffer and solubilized in 50 pl of electrophoresis sample buffer (24) containing 5% 2-mercaptoethanol with heating to 100 "C for 3 min. Approximately 80-95% of specific proteins were recovered by immunoprecipitation or by gelatin-Sepharose.
Polyacrylamide Gel Electrophoresis-Samples were analyzed ac- cording to the method of Laemmli (24) using 5% polyacrylamide slab gels containing SDS. Molecular markers included in gels were myosin (Mr = 200,000), phosphorylase b (Mr = 94,000), and albumin (M, = 67,000). In addition, human or bovine fibronectin (Mr = 220,000) and rat laminin (Mr = 400,000 and M, = 210,000 and 220,000) were also used as standards. The gels were stained, fixed, and then impregnated with Autofluor (National Diagnostics), dried, and exposed to x-ray film (Kodak XAR-5) at -70 "C. For quantification of radioactivity, gel bands on the film were scanned on an Automatic Computing
Densitometer (Gelman Sciences), and relative percentages were plot- ted.
Collagenase Treatment to Solubilize Laminin from Extracellular Deposits for ELISA-On various days of culture, the medium from each well was collected and aggregates were harvested, washed with phosphate-buffered saline, and suspended in 0.1 ml of 0.05 M Tris- HCl, pH 7.5, 0.5 M NaCl, 0.005 M CaC12, 1% Nonidet P-40, and 0.2 mM PMSF. Bacterial collagenase form I11 (Advance Biofactures Corp.) was added to the suspension (5 pl of collagenase containing approximately 12 units/O.l ml of cell suspension). After incubation for 2 h at 37 "C, the cell suspension was centrifuged at 15,000 rpm in an Eppendorf microcentrifuge for 3 min to separate the cell extract from the undigested pellet, and 20 mM EDTA was added to the cell extract. The pellet was suspended in 0.05 M Tris-HC1, pH 7.5, containing 0.02 M EDTA, 0.5 M NaC1, 0.2 mM PMSF, and 0.1% Triton X-100. The medium, cell extract, and pellet extract were tested in ELISA for determination of laminin.
Quantitation of Laminin and Growth Rates-Laminin secreted into the medium and deposited into the extracellular matrix (see above) was determined by sandwich ELISA assay (25) using affinity-purified rabbit anti-rat laminin. The lower limit of detection was 25 ng/ml laminin. The growth of embryoid bodies was measured by disrupting them in 1 M glycine, 2 mM EDTA for 3 h at 37 "C and determining the cell numbers as described (7).
RESULTS
Measurement of Total Metabolically Labeled Proteins-F9 monolayers and aggregates cultured with retinoic acid over an 8-day time course were labeled with [35S]methionine for 4 h and the radioactivity incorporated into trichloroacetic acid- insoluble precipitates was determined as described under "Materials and Methods." The incorporation of [35S]methio- nine into proteins synthesized by F9 aggregates cultured with retinoic acid was highest on the first day and then declined approximately 6-10-fold over an 8-day period in several ex- periments examined, Fig. 1 shows data from one such exper- iment. The level of trichloroacetic acid-insoluble radioactivity found in the cells was consistently more than 10-fold that in the medium at all stages. Although the number of cells is increasing (7) , total incorporation into cell cultures declined during the differentiation process. The nature of the meta- bolically labeled proteins was further examined by immuno- precipitation.
Synthesis and Secretion of Fibronectin-Analyses of meta- bolically labeled fibronectin in cell extracts solubilized in RIPA buffer failed to show a band of radioactivity correspond- ing to fibronectin even when autoradiographs were exposed for a long period of time for both methods of isolation, namely, by affinity to gelatin-Sepharose or by immunoprecipitation using a specific antiserum. Aggregates could be more com- pletely solubilized in 0.5% (w/v) SDS-containing lysis buffer (20), since no visible pellet was obtained after centrifugation at 105,000 X g. However, inclusion of SDS in lysis buffer destroyed the ability of fibronectin to bind to gelatin-sepha- rose, but radioactive fibronectin could be detected by immu- noprecipitation with its specific antiserum (Fig. 2a).
Fibronectin synthesized and either retained or secreted into the medium by aggregates of different developmental stages is shown in Fig. 2. The level of 35S-labeled fibronectin showed an initial rise with a peak on day 2 in cells (Fig. 2a, lane 3) and on day 1 in medium (Fig. 2b, lane 2) and then decreased upon further differentiation both in cells and in medium. On day 3, the fibronectin band was similar in intensity to day 1 in cells (Fig. 2a, compare lane 2 with 4 ) but weaker in the medium (Fig. 2b, compare lane 2 with 4 ) . Radioactive fibro- nectin was greatly reduced in cells and had disappeared from medium by day 7 (Fig. 2, a, and b, lane 6). The relative amounts of 35S-labeled fibronectin secreted into the medium from aggregates after treatment with retinoic acid for various
12254 Extracellular Matrix in Development
n \ medium
I I I I
2 4 6 8 Days of Culture in RA
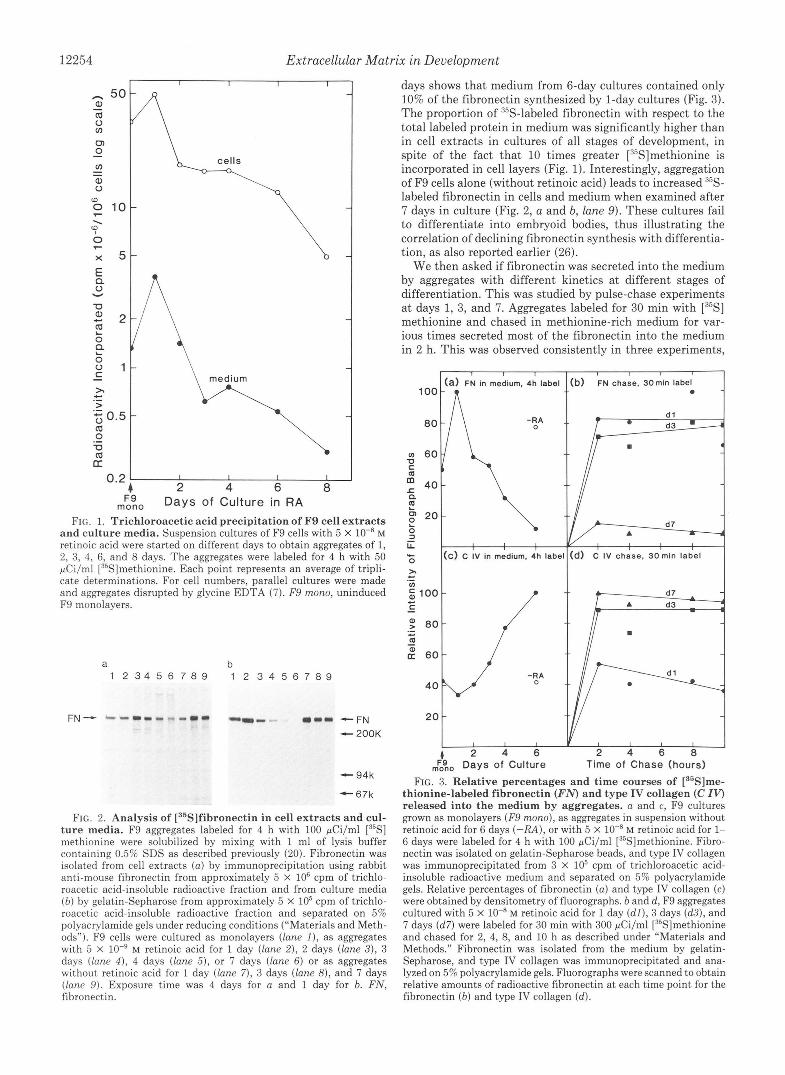
FIG. 1. Trichloroacetic acid precipitation of F9 cell extracts and culture media. Suspension cultures of F9 cells with 5 X lo-* M retinoic acid were started on different days to obtain aggregates of 1, 2, 3, 4, 6, and 8 days. The aggregates were labeled for 4 h with 50 pCi/ml [35S]methionine. Each point represents an average of tripli- cate determinations. For cell numbers, parallel cultures were made and aggregates disrupted by glycine EDTA (7). F9 mono, uninduced F9 monolayers.
a b 1 2 3 4 5 6 7 8 9 1 2 3 4 5 6 7 8 9
- 94k
-67k
FIG. 2. Analysis of [35S]fibronectin in cell extracts and cul- ture media. F9 aggregates labeled for 4 h with 100 pCi/ml [35S] methionine were solubilized by mixing with 1 ml of lysis buffer containing 0.5% SDS as described previously (20). Fibronectin was isolated from cell extracts ( a ) by immunoprecipitation using rabbit anti-mouse fibronectin from approximately 5 X lo6 cpm of trichlo- roacetic acid-insoluble radioactive fraction and from culture media ( b ) by gelatin-Sepharose from approximately 5 X lo5 cpm of trichlo- roacetic acid-insoluble radioactive fraction and separated on 5% polyacrylamide gels under reducing conditions (“Materials and Meth- ods”). F9 cells were cultured as monolayers (lane I ) , as aggregates with 5 X lo-’ M retinoic acid for 1 day (lane 2), 2 days (lane 3), 3 days (lane 4 ) . 4 days (lane 5 ) , or 7 days (lane 6) or as aggregates without retinoic acid for 1 day (lane 7) , 3 days (lane 8), and 7 days (lane 9). Exposure time was 4 days for a and 1 day for b. FN, fibronectin.
days shows that medium from 6-day cultures contained only 10% of the fibronectin synthesized by 1-day cultures (Fig. 3). The proportion of “S-labeled fibronectin with respect to the total labeled protein in medium was significantly higher than in cell extracts in cultures of all stages of development, in spite of the fact that 10 times greater [3sS]methionine is incorporated in cell layers (Fig. 1). Interestingly, aggregation of F9 cells alone (without retinoic acid) leads to increased %- labeled fibronectin in cells and medium when examined after 7 days in culture (Fig. 2, a and b, lane 9). These cultures fail to differentiate into embryoid bodies, thus illustrating the correlation of declining fibronectin synthesis with differentia- tion, as also reported earlier (26).
We then asked if fibronectin was secreted into the medium by aggregates with different kinetics at different stages of differentiation. This was studied by pulse-chase experiments a t days 1, 3, and 7. Aggregates labeled for 30 min with [35S] methionine and chased in methionine-rich medium for var- ious times secreted most of the fibronectin into the medium in 2 h. This was observed consistently in three experiments,
I I
(a) FN in medium, 4h label
P
L 0
U -
( C ) C ;V in medium, ;h lab#
2o t - ( 2 4 6
,$$, Days of Culture
I
:b) FN chase, 30min label
I I I I I 2 4 6 8
Time of Chase (hours) FIG. 3. Relative percentages and time courses of [3SS]me-
thionine-labeled fibronectin (FN) and type IV collagen (C IV) released into the medium by aggregates. a and c, F9 cultures grown as monolayers (F9 mono), as aggregates in suspension without retinoic acid for 6 days (-RA), or with 5 X lo-* M retinoic acid for 1- 6 days were labeled for 4 h with 100 pCi/ml [35S]methionine. Fibro- nectin was isolated on gelatin-Sepharose beads, and type IV collagen was immunoprecipitated from 3 x lo5 cpm of trichloroacetic acid- insoluble radioactive medium and separated on 5% polyacrylamide gels. Relative percentages of fibronectin ( a ) and type IV collagen (c) were obtained by densitometry of fluorographs. b and d, F9 aggregates cultured with 5 X lo-* M retinoic acid for 1 day ( d l ) , 3 days (d3) , and 7 days (d7) were labeled for 30 min with 300 pCi/ml [35S]methionine and chased for 2, 4, 8, and 10 h as described under “Materials and Methods.” Fibronectin was isolated from the medium by gelatin- Sepharose, and type IV collagen was immunoprecipitated and ana- lyzed on 5% polyacrylamide gels. Fluorographs were scanned to obtain relative amounts of radioactive fibronectin a t each time point for the fibronectin ( b ) and type IV collagen (d).

Extracellular Matrix in Development 12255
and there was no difference between the stages examined as shown in Fig. 3b which represents data from one of the experiments. In addition, fibronectin levels in the medium remained approximately constant between 2 and 10 h of chase, thus indicating its stability.
Synthesis and Secretion of Type IV Collagen-Fig. 4 shows the synthesis and secretion of type IV collagen polypeptides. Since it is known that type IV collagen is not processed before deposition into the matrix (27), it is detected as a "procolla- gen" doublet with a molecular weight of about 200,000 on these gels. The bands obtained were sensitive to digestion by bacterial collagenase (data not shown). Relatively low levels of type IV collagen were observed in aggregates (Fig. 4a) compared to medium (Fig. 4b). Low levels of type IV collagen observed in cell aggregates may be due in part to incomplete solubilization of aggregates in RIPA buffer as described above.
The level of newly synthesized type IV collagen increased during differentiation. Significantly lower amounts of 35S- labeled type IV collagen were synthesized and secreted by 1- day aggregates (lane 2) than by 6-day aggregates (lane 6) , and comparison of relative amounts of 35S-labeled type IV collagen secreted into the medium showed an approximately 3-fold increase in collagen synthesis as a result of differentiation (Fig. 3c). In contrast to fibronectin synthesis, type IV collagen synthesis is not stimulated by aggregation in the absence of retinoic acid (compare Figs. 2b with 4b and 3a with 3c). Pulse- chase experiments indicated that most of the newly synthe- sized collagen was secreted into the medium in 2 h at all stages of differentiation (Fig. 3d). These studies show that during 4 h of labeling as in Fig. 3, b and d, a steady state of synthesis and secretion has been reached for all stages.
Synthesis and Secretion of Larninin-Fig. 5 shows autora- diographs of immunoprecipitates from cell extracts and media separated on SDS gels. Synthesis of all three chains together (A, B1, and B2) was first detected in 2-day aggregates (Fig. 5a, lane 3) . Laminin synthesis usually fell on the first day (Fig. 6) and then increased steadily above the level seen in uninduced monolayer cultures and in control aggregates (without retinoic acid). The synthesis of laminin radically differed from that of fibronectin and type IV collagen since more laminin was found in the cell aggregates compared to the medium (Fig. 5). Note that smaller fractions of total lysates were needed to examine cell extracts compared to
a b 1 2 3 4 5 6 7 1 2 3 4 5 6 7
LN - C = -200K
-94K
FIG. 4. Immunoprecipitation of [35S]methionine-labeled type IV collagen. Aggregates labeled for 4 h with 100 pCi/ml [35S] methionine were solubilized and type IV collagen immunoprecipi- tated. The immunoprecipitates from cell extracts and culture media were analyzed on 5% polyacrylamide gels under reducing conditions. a, immunoprecipitation of cell extracts from approximately 2.5 X lo6 cpm of trichloroacetic acid-insoluble radioactive fraction; b, immu- noprecipitation of culture media from approximately 3 x IO5 cpm of trichloroacetic acid-insoluble radioactive fraction. F9 cells were either cultured as monolayers (lane I ) or as aggregates without retinoic acid for 6 days (lane 7). Cultures with 5 X lo-@ M retinoic acid were for 1 day (lane 2), 2 days (lane 3) , 3 days (lane 4 ) , 4 days (lane 5) , or 6 days (lane 6) . Exposure time was 6 days. C, type IV collagen; LN, laminin.
a 1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8
b
A-
..I,:; '4.. :B .2 OK
Ent
94K
67K
FIG, 5. Immunoprecipitation of [36S]methionine-labeled laminin. F9 aggregates labeled for 4 h with 50 pCi/ml [35S]methio- nine were solubilized, and laminin was immunoprecipitated from cell extracts and culture media. The immunoprecipitates were analyzed on 5% polyacrylamide gels under reducing conditions. a, immunopre- cipitation of cell extracts from approximately 2.5 X lo6 cpm of acid- insoluble radioactive fraction; b, immunoprecipitation of culture me- dia from approximately 1 x IO6 cpm of acid-insoluble radioactive fraction. F9 cells were cultured as monolayers (lune I ) , aggregates without retinoic acid for 8 days (lane 8) , or as aggregates with 5 X lo-' M retinoic acid for 1 day (lane 2), 2 days (lane 3), 3 days (lane 4 ) , 4 days (lane 5 ) , 6 days (lane 6 ) , and 8 days (lane 7). Exposure time of fluorographs was 8 days. LN, laminin; Ent , entactin.
I 1 1 1 I
LN subunits in cell lysates, 4h label
0 0 1 - R A
V A-RA
1 1 1 1 I I
1 2 3 4 6 8 IO Days of Culture
FIG. 6. Comparison of laminin (LN) subunits synthesized and accumulated by F9 aggregates during differentiation. Fluorographs (Fig. 5) were scanned (44) and the peaks integrated to determine relative amounts of immunoprecipitated subunits, A, B1, and B2. The values shown were normalized to B2 on day 4 when the highest levels were found. F9 mono, uninduced F9 monolayers; -RA, control aggregates (without retinoic acid).
media samples in Fig. 5 (see Fig. 7c for the best comparison). The most distinctive change was seen in the proportions of
laminin subunits during the differentiative process. Undiffer- entiated cells, control aggregates (without retinoic acid), and 1-day aggregates incorporate more radioactivity into B2 chains than into B1 chains (Fig. 5a, lanes 1, 2, and 8). This difference declines as differentiation proceeds so that by day 3 the amount of radioactivity in newly synthesized B1 and B2 chains is about the same (Fig. 5a, lane 4). Day 1 aggregates synthesize very low levels of polypeptide A chain (seen only after long exposures), and this increases significantly by day 2 and remains constant between days 3 and 6. The relative proportions of subunits and total laminin synthesis during

12256 Extracellular Matrix in Development
differentiation show an approximately 2-fold rise in the level of total laminin synthesis in day 8 aggregates with an approx- imately 4-fold rise in the level of A and B1 chains (Fig. 6).
Pulse-chase experiments showed that 2 h was sufficient to allow newly synthesized laminin to pass into the medium almost completely by aggregates of different developmental stages (Fig. 7). The level of radiolabeled laminin synthesized by day 3 and day 7 aggregates remained unchanged for up to 10 h of chase, while in day 1 aggregates and in aggregates cultured without retinoic acid it decreased markedly (compare lanes 1 and 2 in Fig. 7, a, b, and c), thus showing the poor stability of laminin produced at the early stages when it is composed mainly of B2 subunits. At days 3 and 7, note that at the later times of chase, laminin in the medium was more completely processed to its high molecular weight forms (Fig. 7, b and c, and see also Fig. 5b). In addition, secreted laminin contained all its associated polypeptide chains, namely A, B1, B2, and entactin chains, suggesting that the subunits are assembled prior to their secretion.
Immunoprecipitates of media with anti-laminin also showed a band of radioactivity at the approximate molecular weight of 150,000. The band was specifically immunoprecipi- tated by anti-mouse C antiserum (18) and therefore appears to be entactin (data not shown). No radioactive polypeptide migrating at the position of the entactin chain was seen in the cell fraction of aggregates even after long exposures of the autoradiographs. The level of entactin in the medium changed during differentiation. The uninduced cells synthesized low levels of entactin which usually increased up to day 2 or 3 and declined thereafter up to day 8 (Fig. 5b).
Quantitation of Laminin by ELISA-Although densito- metric scanning of fluorographs provided information about the relative rates of synthesis a t different stages and also indicated the relative proportions of deposited versus solubi- lized laminin, it did not measure total amounts. We therefore employed ELISA to circumvent this problem and to provide an estimation of total laminin.
Table I shows the amount of laminin produced in culture media and within solubilized F9 aggregates at various stages of development. The biggest increase in laminin production appeared to be between days 1 and 2 of differentiation, and the amounts of laminin continued to rise to a peak at day 4 (1011 ng/106 cells/day), a result which confirms the immu- noprecipitation data of Fig. 6. The undetectable levels of laminin in day 1 aggregates could either be due to incomplete solubilization or to rapid turnover of laminin as indicated in
a b C 1 2 3 4 5 6 7 1 2 3 4 5 6 7 1 2 3 4 5 6 7
. .
- A ] L N
LN[::I Z B
- Ent
FIG. 7. Pulse-chase analyses of [36S]methionine-labeled laminin synthesized by F9 aggregates. Aggregates cultured with 5 X lo-” M retinoic acid for 1 day (a ) , 3 days (b ) , or 7 days ( c ) were labeled with 300 pCi/ml [3sS]methionine for 30 min and chased up to 10 h. Laminin was immunoprecipitated using rabbit anti-rat laminin antiserum by sequential immunoprecipitation as described under “Materials and Methods.” Lanes 1 and 2 are autoradiographs from aggregates after 0 and 10 h of chase. Lanes 3-7 are autoradiographs of‘ media after 0 h (lane 3) , 2 h (lune 4 ) , 4 h (lune 5 ) , 8 h (lane 6), and 10 h (lane 7) of chase. Exposure time of fluorographs was 6 days. LN, laminin; Ent, entactin.
TABLE I The accumulation and secretion of laminin during embryoid body
formation F9 cells were cultured as suspension of aggregates in medium
containing 5 X lo-* M retinoic acid. At each time point, medium and extracts of aggregates were examined by ELISA (see “Materials and Methods”).
Day of Laminin Laminin laminin Proportion culture accumu- lated secreted
Net
Droduction secreted
ng f 10‘ cells ngf106 cellsfday % 1 Bkd“ Bkd“ Bkd” 2 818 154 972 15.8 4 2235 303 1011 30.0 6 3153 366 825 8 2000
44.4 101 Degradation of -b
accumulated laminin?*
a Background, 525 ng/ml. Since laminin accumulation appears to be lower at 8 days and
the amount secreted is also lower, we conclude that either laminin is being degraded to products that do not react in ELISA and/or that laminin is not completely solubilized at the later stages.
the immunoprecipitation data (Fig. 7a). Comparison of the amounts of laminin secreted (Table I, column 3) with the total laminin recovered at each stage (Table I, column 4) shows that at first the laminin is largely (more than 80%) accumulated in the aggregates. As laminin production rises, however, an increasing proportion is found in the medium (Table I, column 5 ) reaching 44% a t day 6. After day 6, accumulated levels of laminin appear to fall and this could be due to falling synthetic rates (Fig. 6) and/or increased degra- dation. Another possibility is that laminin is becoming in- creasingly cross-linked into a basement membrane, and there- fore is difficult to solubilize, and so our measurements may underestimate the accumulated levels. However, immunoflu- orescent staining also supports the notion that laminin dep- osition declines after day 4 (7).
DISCUSSION
We have demonstrated that the patterns of synthesis of all three extracellular matrix components change during the differentiation of F9 cells to visceral endoderm. After treat- ment with retinoic acid, we observed in day 8 aggregates a decrease in the synthesis and secretion of fibronectin to almost undetectable levels (Fig. 2), a steady increase in the level of type IV collagen, especially in the medium (Fig. 4), and an increase in the level of laminin with a maximum increase between days 2 and 4 (Fig. 6, Table I). Concomitant with the increase in the level of laminin, a change in the pattern of laminin subunits was also observed between days 2 and 4. Based on the experimental data, we hypothesize that fibronectin plays an early role in cell-cell interaction during aggregation and may initiate the deposition of matrix, whereas laminin functions mainly in basement membrane organiza- tion and also may play a role in the process of differentiation. Evidence in support of these hypotheses is presented here.
Evidence That Fibronectin Plays a n Early Role in Epithe- lium Formation-Uninduced F9 cells synthesize and secrete the relatively high levels of fibronectin that we detected in monolayer cultures. When single cells are plated in Petri dishes, the cells begin to aggregate and 24 h later, smooth aggregates are seen. At this time, rates of fibronectin synthesis are maximal and if retinoic acid is also present, the rate is even higher (Figs. 2b and 3a). After the first day, however, the proportion of total protein synthesized that is fibronectin declined significantly. It appears that the expression of the

Extracellular Matrix in Development 12257
fibronectin gene is specifically regulated when F9 aggregate cultures are treated with retinoic acid. This hypothesis is currently being examined by analyzing mRNA levels during the differentiative process.
Deposits of fibronectin are visible throughout sections of day 1 aggregates (with or without retinoic acid) by immuno- fluorescent staining? After 1 day, aggregates (in retinoic acid) already display some tight junctions (data not shown), which increase with time and which may impede passage of macro- molecules from the inner cells of aggregates. By the end of the 4th day, tight junction formation is complete, and an epithelium is clearly visible in the phase microscope. We suggest that fibronectin initially plays an important part in aggregate formation and further hypothesize that fibronectin deposits may also trigger the formation of the basement membrane by interacting with collagen and proteoglycans (28) if retinoic acid is present. Similarly, in the 4th day mouse blastocyst, fibronectin is also first detected between the cells of the inner cell mass and later fibronectin becomes concen- trated in the nascent basement membrane of the primitive endoderm layer adjacent to the blastocoel cavity (29, 30). At the same time, intracellular type IV collagen (31) and laminin (32) are seen in the endoderm, and later these components form the main structural elements of the new basement membrane.
Evidence That Laminin Plays a Major Role in Basement Membrane Organization and Epithelium Formation-A role for laminin B2 chains also in cell-cell adhesion is not excluded since B2 is synthesized before induction of F9 cells and during the first day of aggregate formation. Such a role for laminin has been suggested (33). Terranova et al. (34) also show that laminin promotes attachment of metastatic cells to type IV collagen. However, laminin synthesis is still relatively low at the beginning of embryoid body formation, and since it reaches a peak on the 4th day (Fig. 6 and Table I), its appearance and location (7) correlates well with the morpho- logical appearance of an outer epithelial layer on the 4th day. We suggest that not only is laminin a major structural com- ponent of the basement membrane, but also assists in the polarization of the outer epithelial layer and hence its ability to express differentiated function (35).
Undifferentiated F9 cells and differentiating aggregates manifest an asynchronous synthesis of B1, B2, and A chains of laminin (Figs. 5 and 6). Asynchronous synthesis of B1, B2, and A chains of laminin is also observed in developing em- bryos. For example, oocytes, unfertilized eggs, and 2-cell mu- rine embryos synthesize only B1 chains; B2 synthesis starts at the 4-8-cell stage and A chains in the 16-cell embryo (36). Only by the 8-cell stage can laminin deposits be detected by immunohistochemistry (37). In this case, the laminin is de- posited as a matrix between the blastomeres rather than as a basement membrane. It appears that the correct proportions of subunits are necessary both for accumulation in a matrix as well as for secretion into the medium which is detected on day 2 (Fig. 5b, lane 3) and that the change in subunit ratio seen here is a developmental phenomenon that leads to lam- inin deposition.
Roles for laminin in induction of polarity (38), intercellular flattening (39), and formation of specialized junctions (40) have been suggested, since these events lead to the ensuing differentiation of the outer trophectoderm layer from the inner cell mass. Laminin may play similar roles during the formation of the epithelial layer on F9 aggregates. The ap- proximately 900,000 Da laminin complex of A, B1, and B2 is
A. Grover and E. D. Adamson, unpublished observations.
known to contain several different functional domains such as cell binding, cell spreading, and type IV collagen and heparin-binding (41), and it is possible that each subunit plays specialized and independent roles in embryogenesis and embryoid body formation. It is not known what specific role the asynchronous synthesis of laminin subunits serves either in the embryo or in F9 cells, but it involves distinct gene regulatory signals since each of these subunits is encoded by a different gene (42).
We showed earlier (35) that exogenous laminin added to F9 suspension cultures stimulates epithelium formation at low laminin concentrations with sparse cell seedings, and, furthermore, that cultures in the presence of anti-laminin antibodies do not differentiate normally, but form aggregates that do not secrete the normal levels of a-fetoprotein and have no distinct outer visceral endoderm layer with little or no basement membrane. However, when exogenous soluble laminin is added to aggregate cultures at high concentrations (10 pglml), embryoid bodies also fail to form, and this is interpreted as the failure of the outer layer of cells to organize into an epithelium when faced with excess laminin on both cell surfaces. Two other F9 culture conditions also fail to form embryoid bodies, one is when 2.5 mM sodium butyrate is present in the culture medium. This chemical prevents the differentiation (43) of F9 monolayer cultures in the presence of retinoic acid and prevents embryoid body formation (al- though not aggregation) as well as epithelium formation and a-fetoprotein prod~ction.~ Such aggregates fail to activate the production of B1 and A chains and therefore mimic aggregate cultures without retinoic acid. F9 aggregate cultures in the presence of 0.1 mM dibutyryl cyclic AMP (and retinoic acid) also fail to form embryoid bodies, but in this case, very high levels of laminin are made after day 2 of culture, and even- tually aggregates differentiate into parietal-type endoderm cells. We find that the level and position of laminin might be the determining factors for differentiation of F9 cells into the alternate endoderm p a t h ~ a y . ~ Taken together, these data demonstrate that laminin may affect F9 cultures in 3 areas: differentiative direction, basement membrane production, and basement membrane and epithelial organization.
Further detailed study is required to understand the precise role(s) of fibronectin, laminin, and collagen in the differentia- tive process. Since many major changes in gene expression occur in the first 48 h after aggregate culture in retinoic acid, it will be necessary to record changes in mRNA expression at these early times. It may be possible to determine if a set of genes (for example, laminin B1 and A; collagen type IV; tight junction products) is activated simultaneously or if declining levels of, for instance, fibronectin may help to trigger the induction of a set of genes needed to form an epithelium.
Acknowledgments-We are grateful to Drs. E. Engvall, E. Ruos- lahti, and B. Hogan for the gifts of antisera and other reagents. We thank Dr. E. Engvall for helpful discussions and suggestions during the course of this work and Drs. S. Edwards, E. Engvall, R. Oshima, K. Trevor, D. Mercola, and J. Meek for critical review of the manu- script. We are thankful to D. Lowe for typing this manuscript. Our special thanks to G. Sandford for painstaking artwork and photog- raphy.
REFERENCES 1. Silver, L. M., Martin, G. R., and Strickland, S. (1983) Cold Spring
Harbor Conf. Cell Proliferation, 10, 3-743 2. Martin, G. R. (1980) Science 209, 768-776
A. Grover and E. D. Adamson, unpublished data. A. Grover and E. D. Adamson, manuscript in preparation.

Extracellular Matrix in Development 3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13. 14.
15.
16.
17.
18.
19.
20. 21.
22.
De Luca, L. M., and Shapiro, S. S. (1981) Ann. N. Y. Acad. Sci.
Hogan, B. L. M., Barlow, D., and Tilly, R. (1983) Cancer Surueys
Hogan, B. L. M., Taylor, A., and Adamson, E. (1981) Nature
Hogan, B. L. M., and Tilly, R. (1981) J. Embryol. Exp. Morphol.
Grover, A., Oshima, R. G., and Adamson, E. D. (1983) J. Cell Bid. 96,1690-1696
Chung, A. E., Freeman, I. L., and Braginski, J. E. (1977) Biochem. Biophys. Res. Commun. 79,859-868
Timpl, R., Rohde, H., Robey, P. G., Rennard, S. I., Foidart, J.- M., and Martin, G. R. (1979) J. Bid. Chem. 254,9933-9937
Wartiovaara, J., Leivo, I., and Vaheri, A. (1980) in The Cell Surface: Mediator of Developmental Processes (Subtelny, S., and Wessells, N. K., eds) pp. 305-324, Academic Press, New York
Ruoslahti, E., Engvall, E., and Hayman, E. G. (1981) Collagen Relat. Res. 1,95-128
Hynes, R. O., and Yamada, K. M. (1982) J. Cell Biol. 95, 369- 377
Furcht, L. T. (1983) Mod. Cell Biology 1,53-117 Kefalides, N. A., and Denduchis, B. (1969) Biochemistry 8,4613-
Ruoslahti, E., Hayman, E. G., Pierschbacher, M., and Engvall,
Engvall, E., Bell, M. L., Carlsson, R. N. K., Miller, E. J., and
Engvall, E., Krusius, T., Wewer, U., and Ruoslahti, E. (1983)
Hogan, B. L. M., Taylor, A., Kurkinen, M., and Couchman, J. R.
Alhrechtsen, R., Nielsen, M., Wewer, U., Engvall, E., and Ruos-
Oshirna, R. G. (1982) J. Biol. Chem. 257,3414-3421 Mans, R. J., and Novelli, G. D. (1961) Arch. Biochem. Biophys.
Engvall, E., and Ruoslahti, E. (1977) Int. J . Cancer 20, l -5
359, 1-429
2 , 115-140
(Lond.) 291,235-237
62,379-394
4621
E. (1982) Methods Enzymol. 82,803-831
Ruoslahti, E. (1982) Cell 29, 475-482
Arch. Biochem. Biophys. 222,649-656
(1982) J. Cell Biol. 95, 197-204
lahti, E. (1981) Cancer Res. 41,5076-5081
94,48-53
23. Cooper, A. R., Kurkinen, M., Taylor, A., and Hogan, B. L. M.
24. Laemmli, U. K. (1970) Nature (Lond.) 227, 680-685 25. Engvall, E. (1980) Methods Enzymol. 70,419-439 26. Carlin, B. E., Durkin, M. E., Bender, B., Jaffe, R., and Chung, A.
E. (1983) J. Bid. Chem. 258, 7729-7737 27. Minor, R. R., Clark, C. C., Strause, E. L., Koszalka, T. R., Brent,
R. L., and Kefalides, N. A. (1976) J. Bid. Chem. 251, 1789- 1794
28. Ruoslahti, E., Hayman, E. G., and Engvall, E. (1980) in Tumor Cell Surfaces and Malignancy (Hynes, R. O., and Fox, C. F., eds) pp. 821-828, Alan R. Liss, New York
29. Wartiovaara, J., Leivo, I., and Vaheri, A. (1979) Deu. Bid. 69,
30. Zetter, B. R., and Martin, G. R. (1978) Proc. Natl. Acad. Sci. U.
31. Adamson, E. D., and Ayers, S. E. (1979) Cell 16,953-965 32. Leivo, I., Vaheri, A., Timpl, R., and Wartiovaara, J. (1980) Deu.
33. Ozawa, M., Sato, M., and Muramatsu, T. (1983) J. Biochem. 94,
34. Terranova, V. P., Liotta, L. A., Russo, R. G., and Martin, G. R.
35. Grover, A., Andrews, G., and Adamson, E. D. (1983) J. Cell Biol.
36. Cooper, A. R., and MacQueen, H. A. (1983) Deu. Bid. 96, 467-
(1981) Eur. J. Biochem. 119, 189-197
247-257
S. A. 75,2324-2328
Bid. 76,100-114
479-485
(1982) Cancer Res. 42,2265-2269
97,137-144
471 37. Wu, T.-C., Wan, Y.-J., Chung, A. E., and Damjanov, I. (1983)
38. Ziomek, C. A., and Johnson, M. H. (1980) Cell 21,935-942 39. Ducibella, T., and Anderson, E. (1975) Deu. Biol. 47, 45-58 40. Lo, C. W., and Gilula, N. B. (1979) Cell 18, 399-409 41. Timpl, R., Engel, J., and Martin, G. R. (1983) Trends Biochem.
42. Barlow, D. P., Green, N. M., Kurkinen, M., and Hogan, B. L. M.
43. Levine, R. A., Campisi, J., Wang, S.-Y., and Gudas, L. J. (1984)
44. Laskey, R. A., and Mills, A. D. (1975) Eur. J. Biochem. 36,335-
Deu. Bwl. 100,496-505
Sci. 8,207-209
(1984) EMBO J. 3,2355-2362
Deu. Biol. 105, 443-450
341