Role of the mod(mdg4) common region in homolog segregation ... · over TM6, Tb e and maintained by...
69
1 Role of the mod(mdg4) common region in homolog segregation in Drosophila male meiosis Morvarid Soltani-Bejnood * , Sharon E. Thomas † , Louisa Villeneuve † , Kierstyn Schwartz † , Chia-sin Hong † and Bruce D. McKee *† *Genome Science and Technology Program, University of Tennessee and Oak Ridge National Laboratory. † Department of Biochemistry, Cellular and Molecular Biology, University of Tennessee, Knoxville. Genetics: Published Articles Ahead of Print, published on February 4, 2007 as 10.1534/genetics.106.063289
Transcript of Role of the mod(mdg4) common region in homolog segregation ... · over TM6, Tb e and maintained by...

1
Role of the mod(mdg4) common region in homolog segregation
in Drosophila male meiosis
Morvarid Soltani-Bejnood*, Sharon E. Thomas†, Louisa Villeneuve†, Kierstyn Schwartz†,
Chia-sin Hong† and Bruce D. McKee*†
*Genome Science and Technology Program, University of Tennessee and Oak Ridge
National Laboratory. †Department of Biochemistry, Cellular and Molecular Biology,
University of Tennessee, Knoxville.
Genetics: Published Articles Ahead of Print, published on February 4, 2007 as 10.1534/genetics.106.063289

2
Running Title: Meiotic functions of mod(mdg4) Keywords: meiosis, homolog pairing, Drosophila, modifier of mdg4 Address correspondence to: Bruce D. McKee Department of Biochemistry, Cellular and Molecular Biology M407 Walters Life Sciences Building University of Tennessee Knoxville, TN 37996-0840 Phone: 865-974-5148 FAX: 865-974-6306 email: [email protected]

3
ABSTRACT
Homologous chromosomes must pair and establish stable connections during
prophase I of meiosis in order to segregate reliably from each other at anaphase I. In
most organisms the stable connections, called chiasmata, arise from crossovers. In
Drosophila males, homologs pair and segregate without crossing over. Chiasmata are
replaced by a homolog conjunction complex that includes the Stromalin in Meiosis
(SNM) and Modifier of Mdg4 in Meiosis (MNM) proteins. MNM is one of 31 alternative
splice products of mod(mdg4), all of which share a common 402 amino acid N-terminus
and differ at their C-termini. Previous data demonstrated that an MNM-specific exon is
required for homolog conjunction, but did not address whether the N-terminal common
region, which includes a BTB domain that can mediate coalescence of protein-DNA
complexes, is also required. Here we describe a mutation in the common region of
mod(mdg4), Z3-3401, that causes qualitatively similar phenotypes as the MNM-specific
alleles but disrupts X-Y segregation much more drastically than autosomal segregation.
The mutant MNM protein in Z3-3401 is expressed throughout prophase I in
spermatocytes but the protein is confined to the cytoplasm, suggesting that the Z3-3401
mutation disrupts a signal required for nuclear localization or retention. Z3-3401 fails to
complement a large battery of lethal and semi-lethal alleles in the common region for
meiotic nondisjunction, including an allele containing an amino acid substitution at a
conserved residue in the BTB/POZ domain, consistent with a general requirement for
the mod(mdg4) common region in homolog segregation.

4
INTRODUCTION
The segregation of homologous chromosomes at meiosis I is an essential step in
sexual reproduction, and must be accomplished accurately to prevent the generation of
aneuploid gametes. Mis-segregation of homologs is a major cause of spontaneous
abortion and genetic illness in humans (HASSOLD and HUNT 2001).
Segregation of homologs at anaphase I depends upon their prior alignment and
pairing during early prophase I (PAGE and HAWLEY 2003; McKEE 2004). In most
eukaryotes, the initial homologous pairing interactions are quickly followed by the
formation of elaborate homolog linking structures known as synaptonemal complexes
(SCs) and by the onset of meiotic recombination (ROEDER 1997; PAGE and HAWLEY
2003, 2004). The crossovers that occur between homologous chromatids during this
stage are in turn essential for generation of chiasmata, the stable linkers that connect
homologs throughout late prophase I and metaphase I and that enable the homologs to
segregate reliably from one another at anaphase I (HAWLEY 1988; CARPENTER
1994).
Homolog pairing is essential for segregation even in variant forms of meiosis that
do not involve recombination and chiasmata (WOLF, 1994). In Drosophila males,
homologs are intimately paired throughout the first half of meiotic prophase but do not
recombine or form SCs. Pairing is lost in mid-prophase I but homologs remain together
in discrete nuclear territories until the onset of prometaphase I when they condense into
tight “achiasmate” bivalents, which then segregate with exceptional fidelity at anaphase
I (VAZQUEZ et al. 2002). The central role of pairing in this process has been well-

5
documented for the X and Y chromosomes, which pair only within a discrete
heterochromatic region encompassing the rDNA. X heterochromatic deletions that
remove all of the rDNA prevent pairing of the X and Y and lead to their random
assortment at anaphase I (McKEE and LINDSLEY 1987; PARK and YAMAMOTO
1995; McKEE 1996). Moreover, transgenic rDNA insertions on such
heterochromatically deficient X chromosomes substantially restore both pairing and
disjunction of the X-Y pair (McKEE and KARPEN 1990; McKEE 1996).
The means by which achiasmate homologs in Drosophila remain stably
connected until anaphase I despite the absence of synaptonemal complexes and
chiasmata has been an enigma. Recently, however, two proteins, Modifier of Mdg4 in
Meiosis (MNM) and Stromalin in Meiosis (SNM), were shown to be essential for stable
connections between achiasmate homologs. mnm and snm mutations cause high
frequencies of univalents and random segregation of homologs during meiosis I
(THOMAS et al. 2005). Ectopically expressed, GFP-tagged MNM was shown to
suppress the meiotic phenotypes of the two mnm mutations and to localize to meiotic
chromosomes throughout prophase I and metaphase I. MNM-GFP colocalizes with
native SNM protein to nucleoli of prophase I spermatocytes, where the rDNA genes are
sequestered, and to the pairing region of condensed X-Y bivalents during
prometaphase I and metaphase I. Both proteins disappear at the onset of anaphase I,
strongly implying that they play a structural role in maintaining homolog connections.
Mutations in a third gene (teflon (tef)) cause similar phenotypes but affect only the
autosomes (TOMKIEL et al. 2001).

6
Despite these recent advances, several key issues related to the mechanism of
achiasmate homolog segregation remain unresolved. Perhaps the most important is
the molecular basis for homolog conjunction. SNM is a distant homolog of the SCC3
family of cohesin proteins, raising the possibility that achiasmate homologs are
connected by a cohesin complex of some kind. However, MNM and SNM do not visibly
co-localize with the cohesin protein SMC1 on male meiotic chromosomes (THOMAS et
al. 2005).
An alternative mechanism is suggested by the domain structure of MNM. MNM
is encoded by the complex mod(mdg4) locus, which is thought to produce 31 distinct
chromosomal proteins with a common 402 amino-acid N-terminus but different C-
termini encoded by alternatively spliced exons in the variable region (VR) (see Fig. 1A).
The common region (CR) includes an N-terminal BTB/POZ domain and most of the VR
C-termini, including that of MNM, contain a C2H2 motif (DORN and KRAUSS 2003;
LABRADOR and CORCES 2003). BTB/POZ domains are strong protein interaction
domains found in many transcriptional regulatory proteins, where they function in
mediating homo-dimerization and multimerization (BARDWELL and TREISMANN
1994; ZOLLMAN et al. 1994; IGARISHI et al. 1998; MULLER et al. 1999; MELNICK et
al. 2000; GAUSE et al. 2001; STOGIOS et al. 2005). The BTB domain of mod(mdg4)
is most similar to that of Drosophila GAGA factor, an abundant transcription regulator
required for chromatin remodeling of many developmentally regulated promoters and for
pairing-dependent silencing (GRANOK et al. 1995). Indeed the BTB domain of
Mod(mdg4) can substitute for that of GAGA with little loss of function (READ et al.

7
2000). GAGA utilizes a C-terminal C2H2 zinc-finger motif along with its N-terminal BTB
domain to bind cooperatively to DNAs containing multiple GAGAA sequences, forming
large multimeric complexes held together by BTB-BTB interactions (KATSANI et al.
1999; ESPINAS et al. 1999). Mod(mdg4) proteins also form multimers and both MNM
and Mod(mdg4)67.2, which is required in Drosophila somatic cells for the function of
gypsy insulators (GERASIMOVA et al. 1995), form prominent nuclear foci that
presumably arise via coalescence of multiple chromosome sites bound by Mod(mdg4)-
containing complexes (GERASIMOVA and CORCES 1998; GERASIMOVA et al. 2000;
GHOSH et al. 2001; GAUSE et al. 2001; THOMAS et al. 2005). Moreover,
comparisons of polytene chromosome localization patterns of different Mod(mdg4)
proteins indicate that the variable C-termini specify distinct localization patterns
(BUCHNER et al. 2000). Thus, a plausible mechanism for MNM-mediated conjunction
would involve binding to chromosome pairing sites via its C-terminal C2H2 motif and
coalescence of bound sites on homologous chromosomes via BTB-mediated
multimerization.
The first step in this scenario has experimental support. Both of the mnm
mutations disrupt the C2H2 motif of MNM, Z3-5578 truncating MNM upstream of this
motif and Z3-3298 substituting a Y for the upstream H (Fig. 1A,B), and both mutations
abrogate localization of MNM (and its partner, SNM) to meiotic chromosomes
(THOMAS et al. 2005). However, there is no direct evidence as yet for a role of the
mod(mdg4) BTB domain in homolog conjunction. Indeed, since the two mnm mutations
affect only the unique C-terminus of MNM, it is not known whether any part of the 402

8
amino acid N-terminal CR of mod(mdg4) is required for conjunction. Although the
transgene rescue data and localization patterns of MNM-GFP described above are
consistent with the scenario outlined, those data do not prove that the N-terminal
sequences of MNM must be present for the MNM-specific domain to mediate
conjunction, let alone establish whether or not those sequences play a direct role in the
conjunction process.
Here we provide genetic and cytological evidence that the common region of
mod(mdg4) is required for homolog conjunction. We describe a new mod(mdg4) allele
that causes meiotic phenotypes very similar to those of the mnm alleles but maps to the
common region. We also demonstrate that a large number of mutations in the CR
disrupt meiotic homolog segregation, including one that involves substitution of a
conserved residue in the BTB domain. These findings set the stage for mechanistic
studies of the role of the BTB domain and other domains of Mod(mdg4) in meiotic
conjunction.
MATERIALS AND METHODS
Fly stocks, special chromosomes and Drosophila culture methods: The
Zuker-3 collection consists of >6000 EMS-mutagenized third chromosomes balanced
over TM6, Tb e and maintained by C. Zuker (KOUNDAKJIAN et al. 2004). Z3-3401 and
the mnm and snm lines used in this study were identified in a screen of the Zuker-3
collection for mutations that cause paternal loss of chromosome four (WAKIMOTO et
al. 2004) and were kindly provided by B. Wakimoto. mod(mdg4) alleles were obtained
from V. Corces (John Hopkins University, MD), Rainer Dorn (Institute of Genetics,

9
Martin-Luther-University, Halle, Germany), M. Frasch (Mount Sinai School of Medicine,
New York) and the Bloomington Stock Center at the University of Indiana.
The marked Y chromosome (Dp(1;Y)BSYy+ = BSYy+) carries two transposed
segments from the X chromosome with the markers Bs and y+ appended to the ends of
the left and right arms, respectively (FLYBASE). C(1)RM, y2 su(wa) wa , C(4)RM, ci eyR
and C(2)EN, b pr are attached chromosomes consisting of two genetically complete
copies of the chromosome (X, 4 or 2) attached to a single centromere (FLYBASE).
Dp(1;1)scV1 contains a small duplication from the tip of XL carrying the y+ allele
appended to the small heterochromatic right (XR) arm (RASOOLY and ROBBINS
1991). The attached-XY chromosome used in the recombination crosses was YSX.YL
In(1)EN, y B (X^Y, y B) (FLYBASE).
Unless otherwise specified, the males being tested were crossed singly to two or
three females in shell vials. Crosses were incubated at 23◦C on cornmeal-molasses-
yeast-agar medium. Parents were removed from the vial on day 8 and progeny were
counted between day 13 and day 22.
Mapping of Z3-3401: Z3-3401 was mapped to the mod(mdg4) region by its
failure to complement Df(3R)GC14 (93D7; 93E1) (MOHLER and PARDUE 1994) for X-
Y nondisjunction (NDJ). More detailed mapping was carried out by complementation
against a battery of deletions, transposon insertions and EMS mutations described in
Table 1 using the same assay. To aid in this analysis, the breakpoints of several small
deletions in the mod(mdg4) region were molecularly mapped, as described below. Z3-
3401 failed to complement all deletions that encompass part or all of the CR and all
transposon insertions and mutations in the CR of mod(mdg4).

10
Determination of deletion breakpoints: Deletion breakpoints were mapped
relative to polymorphisms between ORiso3 and CSiso3, which are wild-type lines with
isogenic 3rd chromosomes derived from the Oregon R and Canton S stocks. The DNA
sequences of ORiso3 and CSiso3 differ from each other at multiple sites, some of which
have been identified (HOSKINS et al. 2001). ORiso3 and CSiso3 were crossed with
iso-3rd chromosome stocks carrying the mod(mdg4) deficiencies B2, T16, eGP4,
142∆10, 142∆29, 142∆33,142∆49 and Df(3R)GC14. Genomic DNA was prepared from
F1 adult heterozygotes as described above. Fragments of ~500-800 nt. within and
beyond the mod(mdg4) locus were amplified by PCR from these DNAs, purified and
sequenced as described above. Sequences were analyzed for SNPs and double-
peaks. The logic of the assay is that sequence differences between ORiso3 and
CSiso3 that lie within the deleted region will result in different single peaks on the DNA
sequence electropherograms for the ORiso3/Df and CSiso3/Df samples (e.g., G versus
A at a specific nucleotide position), whereas sequence differences outside of the
deleted region will result in a double peak on the electropherogram for at least one of
the two samples (e.g., a G/A double peak in one sample and a G peak in the other).
This method enabled us to map the relevant breakpoint of each deletion (Table 2, Fig.
1A) with respect to 12 SNPs within the mod(mdg4) locus. The molecular coordinates
and associated primers of the flanking SNPs are available upon request.
Molecular identification of mutations in Z3-3401, mod(mdg4)324 and
mod(mdg4)340: To identify the Z3-3401 mutation, genomic DNA was extracted from
adult flies homozygous for Z3-3401 and for the Zuker-3 progenitor chromosome using
the Wizard genomic DNA purification kit (Promega). Known and conceptual exons of

11
mod(mdg4) were amplified from the genomic DNAs using primer pairs complementary
to intronic sequences immediately flanking the exons, and where necessary, exon-
internal primers. The polymerase chain reaction (PCR) parameters were 1 minute at
94oC, 35 cycles of 94oC for 1 minute, 55oC for 1.5 minutes, and 72oC for 2 minutes in a
Perkin-Elmer thermocycler. Reaction mixtures contained 0.2µM of each primer, 50 ng
Drosophila genomic DNA (Zuker-3 or y w), 1.5 mM MgCl2, 0.2 mM dNTP mix and 2.5 U
Taq DNA polymerase (Promega) in a total volume of 50 microliters. The amplicons
were sequenced directly using an ABI 373 sequencer. We identified a single
nucleotide substitution (C to T) in exon 4 of Z3-3401 predicted to result in substitution of
cysteine (C) for arginine (R) at residue 224 (R224C).
To identify the mutations in mod(mdg4)324 and mod(mdg4)340, genomic DNA
from mod(mdg4)324/Zuker-3 and mod(mdg4)340/Zuker-3 flies was extracted and
analyzed as above. Both mutations were identified as double peaks on the resulting
DNA sequence electropherograms. The mod(mdg4)324 mutation is a G to A
substitution predicted to result in replacement of the glycine at residue 92 with aspartic
acid (G92D). The mod(mdg4)340 mutation is a G to T substitution predicted to result in
replacement of the codon for glutamine 177 (Q177) with a nonsense codon. Additional
double peaks were present on the electropherograms of the CR sequences derived
from both of the mod(mdg4)/Zuker-3 DNA samples, but all except the mutations cited
above proved to represent synonymous substitutions.
Measuring X-Y nondisjunction (NDJ): +/BsYy+ males were crossed singly to
two X/X y w (yellow1 white1118) females in shell vials. The X, Y, XY and O sperm
classes yield: + (y w/+) females, w Bs (y w/BSYy+) males, BS (y w/+/BSYy+) females, and

12
y w (y w/O) males, respectively. The nondisjunction frequency (%X-Y NDJ) = 100 x (BS
females + y w males)/N. N = # of progeny scored.
Measuring 4-loss frequencies: Males were crossed singly to two C(4)RM, ci
eyR (4^4/O) females and the progeny scored for the recessive ci (cubitus-interruptus)
and eyR (eyeless) markers. 4^4/O females generate only 4^4 and nullo-4 (O) eggs,
which when fertilized by regular sperm carrying a wild-type 4th chromosome yield only
ci+ ey+ progeny (viable triplo-4 and poorly viable, Minute haplo-4 progeny). Nullo-4 (O)
sperm from paternal NDJ or chromosome 4 loss yield viable disomic 4^4/O, ci ey
progeny. Paternal NDJ generates 44 sperm as well but these yield only ci+ ey+ progeny
that cannot be distinguished from the regular progeny. % 4-loss = 100 x (ci ey)/N.
Measuring 2nd chromosome NDJ: Males were +/BSYy+; bw/+; Z3/(Df or +).
Sibling mutant (Z3/Df) and control (Z3/+) males were crossed either to C(2)EN, b pr
(2^2/O) females or y w (2/2) females with unattached 2nd chromosomes at a ratio of 2
males to 4 females, or 1 male to 2 females, respectively. Males and females were left
together for 6-8 days, then the females were transferred to fresh vials every 3-6 days
and allowed to continue laying eggs until fertilized eggs were exhausted. Males were
transferred to vials with fresh virgin females and the procedure repeated as long as the
males remained fertile. Progeny were counted to completion, and scored for relevant
markers. 2^2/O females generate eggs that are nullosomic (O) or disomic (2^2) for
chromosome 2, so the only viable progeny are the products of paternal chromosome 2
NDJ (22 and O sperm). Overall 2nd chromosome NDJ was estimated from progeny
per male in the 2^2/O and 2/2 crosses (F(2^2/O) and F(2/2)). The formula is (%NDJ) =
100 x (2 x f x F(2^2)/(2 x f x F(2^2) + F(2/2)). F(2^2/O) is doubled in the formula to

13
compensate for the deaths of ½ the nondisjunctional progeny in the 2^2/O cross, due to
fertilization of the wrong eggs (e.g., O sperm fertilizing O eggs). f is a fertility correction
(=1.8 in these data), based on an independent estimate of the relative fertility of 2^2/O
and 2/2 females in crosses to males of like genotype (2^2/O x 2^2/O and 2/2 x 2/2),
again doubling the progeny from the 2^2/O cross to account for loss of 50% of the
aneuploid fertilization products.
The frequency of sister chromatid NDJ relative to homolog NDJ (% sis-2 NDJ) for
the 2nd chromosomes was estimated from the ratio of bw (brown-eyed) progeny to bw+
(red-eyed) progeny. Both bw and bw+ progeny carry two paternal 2nd chromosomes;
(progeny from fertilization of 2^2 eggs by O sperm are b pr (black body, purple eyes)).
Since the paternal genotype is heterozygous bw/bw+, the 22/O progeny can be bw/bw,
bw+/bw+ or bw/bw+, the former two genotypes resulting from sister chromatid NDJ and
the latter from homolog NDJ. The formula for % sis-2 NDJ is 100 x 2 x bw/(bw + bw+).
(The bw progeny are doubled to account for a presumed equal number of bw+ progeny
that are homozygous for the bw+ chromatid.)
Assaying homologous pairing in spermatogonia and spermatocytes:
Pairing was assayed by counting GFP spots in spermatogonia and spermatocytes from
males homozygous for a chromosome 2 transgene carrying a 256mer tandem array of
lacO repeats and heterozygous for a transgene (also on chromosome 2) expressing a
GFP-LacI chimeric protein under control of the hsp83 promoter (ROBINETT et al. 1996;
STRAIGHT et al. 1996; VAZQUEZ et al. 2002; THOMAS et al. 2005). Testes were
dissected from third instar larvae, pupae or young adults in testes buffer (183 mM KCl,
47 mM NaCl, 10 mM Tris-HCl, 1 mM EDTA, 1 mM PMSF) and gently squashed in

14
testes buffer. Spot frequencies were compared between Z3-3401/mod(mdg4)T16 and
sibling control (Z3-3401/+ and mod(mdg4)T16/+) males. The mean distance among the
four GFP spots (spot dispersion) during late prophase I was determined by measuring
and averaging the four shortest pairwise distances using Metamorph. The resulting
values were averaged over the N nuclei scored to obtain the mean spot dispersion.
Transgene rescue crosses: Two insertions of [hs::MNM-GFP] (THOMAS et al.
2005), one each on chromosome 2 and 3, were used in the rescue experiments. For the
chromosome 2 transgene, +/BsYy+; [hs::MNM-GFP]2, bw+/bw; Z3-3401, st/ Z3-3401, st
males and their +/BsYy+; bw/bw; Z3-3401, st/ Z3-3401, st brothers (scarlet versus white
eyes, respectively) were subjected to 0-2 heat shocks (39◦C for 1 hr.) during larval
stages, then crossed as adults to y w females to assay X-Y NDJ, or their testes
dissected and analyzed cytologically. Similar methods were used to test a 3rd
chromosome insertion of [hs::MNM-GFP] and a 2nd chromosome insertion of [CR-7.5] (=
P[w+ 7.5Kb BamHI] (BUCHNER et al. 2000)).
Testis Immunostaining: Testes were dissected from third instar larvae, pupae
or young adults. For anti-α-tubulin/DAPI experiments, testes were fixed according to
CENCI et al. (1994). Before incubation with antibodies, slides were rinsed twice in PBS
and blocked in PBS, 1% BSA for 45 min. Testes preparations were incubated overnight
at 4ºC with FITC conjugated monoclonal anti-α-tubulin (Sigma) diluted 1:150 in PBT
(PBS with 0.1% Triton X-100) containing 1% BSA. Slides were rinsed twice with PBT,
once with PBS, stained with DAPI (1 µg/mL) for 5 min, rinsed twice in PBS, and
mounted with Vectashield mounting medium.

15
For the SNM staining, the procedure of THOMAS et al. (2005) was followed.
Briefly, testes were fixed according to GUNSALUS et al (1995). Before incubation with
antibodies, slides were washed twice in PBT + DOC (PBS with 0.3% Triton X-100 with
0.3% sodium deoxycholate) for 15 min and once in PBT (PBS with 0.1% Triton X-100)
for 10 min, and blocked in TNB (0.1M Tris-HCl, ph 7.5, 0.15M NaCl, 0.5% Blocking
Reagent (PerkinElmer) for 30 min. Testes preparations were incubated overnight at
4ºC with FITC conjugated monoclonal anti-α-tubulin (Sigma) diluted 1:150 with either
undiluted anti-SNM N-terminus antibody or anti-SNM C-terminus diluted 1:250. Slides
were rinsed three times with TNT (0.1M Tris-HCl, ph 7.5, 0.15M NaCl, 0.05% Tween-
20) and then incubated with Alexa Fluor® 647 goat anti-rabbit IgG (H+L) diluted 1:500
in TNB for 30 min. Slides were rinsed three times with TNT, stained with DAPI (1
µg/mL) for 5 min and rinsed two more times with TNT, and mounted with Vectashield
mounting medium. Anti-ModC (BUCHNER et al. 2000; THOMAS et al. 2005) was
diluted 1:4000 in PBS and visualized using Alexa Fluor 546 goat anti-rabbit IgG (H+L)
diluted 1:5000.
Microscopy: All testis preparations were examined with an Axioplan (ZEISS)
microscope equipped with an HBO 100-W mercury lamp for epifluorescence and with a
scientific grade cooled charge-coupled device (CCD; Roper). Grayscale digital images
were collected, pseudocolored and merged using Metamorph Software (Universal
Imaging Corporation).
Measuring recombination and nondisjunction in female meiosis: To
measure sex chromosome NDJ and recombination, Dp(1;1)scV1, y pn cv m f . y+/y
females were crossed with YSX.YL In(1)EN, y B/Y (X^Y, y B/Y) males. The regular

16
progeny are B females and B+ males; X-X NDJ yields B+ females and B males. %NDJ
= 100 x 2 x (B+ females + y B males)/(N + B+ females + y B males). Recombination was
scored in the regular (B+) sons. Since both X chromosomes carry mutant y alleles at
the native y locus, the duplicated y+ allele on the X chromosome right arm in the y pn cv
m f .y+ homolog serves as a centromere marker. pn, which is less than 1 cM from the
XL tip, is the distal-most marker. Recombination on chromosomes 2 and 3 was
measured as described in the legend to Table 6. Map distances were calculated by
standard formulae and expressed in centiMorgans (cM).
Analyzing pigmentation of y2 flies: The body, wing and bristle colors of adult
flies were scored visually at 20X magnification; a minimum of 20 flies of each genotype
were scored.
Analysis of MNM RNAs: Total RNA was isolated using RNAwiz (Ambion) from
the following genotypes: Z3-5578/Z3-3401, Z3-5578/Df(3R)GC14 and Z3-
3401/Df(3R)GC14. For each RNA, Oligo(dT) and gene specific primer reverse
transcription (RT) reactions were performed using SuperScript First-Strand Synthesis
System for RT-PCR (Invitrogen). Primers for gene specific primer RT are as follows:
5’gattgttagatgtcttatgg 3’ and 5’ tgtaagcctatgacgcatcc 3’. The three RT reactions were
combined into a cocktail and were used in PCR. To determine if trans-splicing occurs,
PCR was performed using the RT cocktail from trans-heterozygotes (Z3-5578/Z3-3401).
As a control for template switching, PCR was performed on the combined cocktails of
hemizygotes (Z3-5578/Df(3R)GC14 and Z3-3401/Df(3R)GC14). The primers used in
PCR are as follows: 5’tgaaatggctacatatgtgg 3’ and 5’cggcatctgagtgaacatct 3’. PCR
products were run on an agarose gel, gel purified using QIAquick Gel Extraction Kit

17
(Qiagen) and TA cloned (Invitrogen). Minipreps were performed on individual colonies
and the DNA was sequenced using standard techniques.
The parental (P) and recombinant (R) RNA frequencies in the trans-
heterozygous sample were estimated from the frequencies of the corresponding
parental (P1 + P2) and recombinant (R1 + R2) cDNAs by correcting for the observed
frequency of template switching (0.16) in the control reaction, as follows. The frequency
of template switching for both P and R templates in the trans-heterozygous reaction was
assumed to be the same as for P templates in the control reaction. Therefore, R1 + R2
cDNAs originate from R templates at a frequency of (1 - .16)R and from P templates at
a frequency of .16(P). Since P = 1 - R, R1 + R2 = (1 - .16)R + .16(1 - R) = .16 + .68R.
Plugging in R1 + R2 = .40, we obtain R = (.40 - .16)/.68 = .35. 95% confidence interval
= +/- 1.96(R(1-R)/N)½.
RESULTS
An amino acid substitition in the mod(mdg4) CR that causes elevated X-Y
NDJ: The meiosis-specific mod(mdg4) mutations described previously, Z3-5578 and
Z3-3298 (THOMAS et al. 2005), were recovered in a screen for paternal chromosome
loss mutants among the Zuker-3 (Z3) collection of EMS-mutagenized autosomes
(KOUNDAKJIAN et al. 2004; WAKIMOTO et al. 2004). That screen yielded an
additional viable mutation, Z3-3401, that caused high frequencies of X-Y nondisjunction
(NDJ) in male meiosis (Table 3) but no NDJ of sister X chromatids (data not shown),
similar to the phenotypes of the mnm alleles, Z3-5578 and Z3-3298. Deficiency
complementation (Table 1, analyzed more fully below) and DNA sequence analysis of

18
Z3-3401 led to the identification of a single base-pair substitution in exon 4 of the
mod(mdg4) CR predicted to result in a R224C substitution in all Mod(mdg4) proteins
(Fig. 1B). This mutation lies downstream of the BTB-domain, in a region of the protein
rich in T residues and of unknown function (BUCHNER et al. 2000; DORN and
KRAUSS 2003; BARDWELL and TREISMAN 1994; ZOLLMANN et al. 1994).
Z3-3401 disrupts segregation of sex chromosomes more severely than
autosomes: As shown in Table 3, the X-Y NDJ frequency in Z3-3401/Df males was
similar to the frequencies in Z3-5578/Df and Z3-3298/Df males. However, unlike the
mnm alleles which behave as genetic nulls, the X-Y NDJ frequency in Z3-3401/Z3-3401
males, although substantial (circa 30%), was significantly lower than in Z3-3401/Df
males, suggesting that Z3-3401 may be a hypomorphic allele.
Both mnm and snm mutations disrupt autosomal segregation as strongly as X-Y
segregation. To ascertain if Z3-3401 also disrupts segregation of the autosomes, 4th
chromosome loss (production of nullo-4 sperm) was assayed by crossing Z3-3401
males carrying wild-type 4th chromosomes to females carrying C(4)RM, an attached 4th
chromosome marked with the recessive mutations ci and eyR (Table 4). Z3-3401/Df
males generated ci ey progeny (C(4)RM/0), indicative of nullo-4 sperm, at frequencies
of 5.2% and 7.9% over two different mod(mdg4) deficiencies, frequencies much lower
than those of hemizygous mnm males (22-28%). An even lower 4-loss frequency
(3.3%) was exhibited by Z3-3401 homozygotes, consistent with the above suggestion
that it is a hypomorphic allele. The difference between the 4-loss and X-Y NDJ
frequencies suggests that Z3-3401 affects sex chromosomes more strongly than
autosomes, or, alternatively, large chromosomes more strongly than small ones (the

19
fourth chromosome being approximately 4% the size of the X chromosome and 2% that
of the 2nd or 3rd chromosome).
To assess the effects of Z3-3401 on segregation of a large autosome pair, Z3-
3401/Df males were crossed to females carrying the attached-2nd chromosome,
C(2)EN, b pr (Table 5). Since all eggs in this cross are either diplo-2 or nullo-2, the only
viable progeny result from fertilization by NDJ sperm (diplo-2 or nullo-2), so that
progeny per male is a rough measure of NDJ. Crosses with wild-type males typically
produce fewer than 0.5 progeny per male, whereas snm and mnm males generate 20-
25 progeny each (Table 5; THOMAS et al. 2005). Z3-3401/Df males generated 15
progeny per male, indicative of a significant amount of chromosome 2 NDJ. The
distribution of paternal chromosome 2 markers among the NDJ progeny indicated that
virtually all of the NDJ occurred at meiosis I.
To obtain quantitative estimates of the frequency of chromosome 2 NDJ, parallel
crosses of Z3-3401/Df sibling males to C(2)EN females (2^2/O) and to females with
normal 2nd chromosomes (2/2) were conducted by a procedure designed to
exhaustively sample sperm (see Materials and Methods). Under these conditions, the
numbers of progeny produced per male (F) in the 2^2 and 2/2 crosses are proportional
to the numbers of NDJ and regular sperm, respectively, produced by the tested males.
Using this method, we estimated the 2nd chromosome NDJ frequency for Z3-
3401/Df males at 17%, a moderate frequency more comparable to that of the 4th
chromosome than to the X-Y pair. By comparison, similar crosses for the two mnm
alleles (Table 5) yielded chromosome 2 NDJ frequencies ranging from 43-45%, very
near to random assortment. These values are in excellent agreement with NDJ

20
estimates from cytological assays (data not shown), thus providing a validation for the
parallel cross method. We conclude that Z3-3401 is significantly more disruptive of sex
chromosome than autosomal segregation, unlike mnm and snm mutations which disrupt
segregation of all chromosome pairs to roughly the same degree.
Z3-3401 causes bivalent instability and missegregation of univalents at MI:
To gain further insight into the nature of the meiotic anomaly in Z3-3401 males, we
examined testis squashes stained with DAPI or acetic orcein to visualize DNA (Fig.
2A,C). Inspection of spermatids from Z3-3401 squashes revealed considerable
variability in size of spermatid nuclei in Z3-3401 (Fig. 2A), an indicator of NDJ. The
cause of the high NDJ in Z3-3401 males was evident from inspection of meiosis I
spermatocytes (Fig. 2A,C). In wild-type spermatocytes, meiotic chromosomes are first
clearly resolved shortly before prometaphase I as three large and one small condensing
masses of chromatin, corresponding to the three large bivalents (X-Y, 2nd and 3rd
chromosomes) and the small 4th chromosome pair, arrayed around the nuclear
periphery. The chromosomes subsequently congress to form a single compact clump
of chromatin on the MI spindle (CENCI et al. 1994). In Z3-3401 spermatocytes,
although the chromosomes condensed normally, the homologs were frequently
unpaired at prometaphase I and metaphase I. There was no sign of chromosome
fragmentation or of breakdown of univalents into their constituent chromatids.
Anaphase I was disorganized, with chromosomes migrating to poles asynchronously
(Fig. 2C). Meiosis I poles frequently (54%) exhibited nuclei of unequal sizes, indicative
of meiosis I NDJ (Fig. 2A,B). Although the meiosis I phenotypes of Z3-3401/Df males
were qualitatively similar to those of mnm males, normal metaphase I configurations

21
and equal telophase I poles were observed at considerably higher frequencies in Z3-
3401 males than in Z3-3298/Df males (Fig. 2B), consistent with the genetic evidence
that autosomes segregate more regularly in Z3-3401/Df than in Z3-3298/Df.
No meiotic abnormalities unique to Z3-3401 were observed. As in Z3-5578 and
Z3-3298 spermatocytes, both spindle structure and kinetochore function appeared
normal at both divisions. The second meiotic division appeared to proceed normally,
consistent with the genetic data (Fig. 2A). Pairing of homologous sequences during
early prophase I as well as in premeiotic gonial cells, as revealed by fusion of
fluorescent spots resulting from recruitment of LacI-GFP proteins to transgenic lacO
arrays at homologous sites in autosomal euchromatin (ROBINETT et al. 1996;
STRAIGHT et al. 1996; VAZQUEZ et al. 2002), also appeared completely normal (Fig.
3), consistent with results of similar analyses of pairing in Z3-5578 and Z3-3298 mutants
(THOMAS et al. 2005). Moreover, in Z3-3401/Df males, as previously reported for wild-
type and mnm males, pairing of homologous and sister chromatids is lost
simultaneously during stage S2b, corresponding to mid-prophase I, and four distinct
GFP spots, corresponding to the four chromatids of the chromosome 2 bivalent, are
seen from then until the conclusion of the meiotic divisions. Finally, as described
previously for both mnm and snm alleles, the separate GFP spots are further apart, on
average, in Z3-3401/Df than in wild-type males during late prophase I (Fig. 3B), a
phenotype referred to as “territorial expansion” (THOMAS et al. 2005).
Thus, we find no genetic or cytological evidence that Z3-3401 has any meiotic
phenotypes qualitatitvely different from those previously described for mnm (and snm)
mutants, namely disruption of homologous chromosome territories during late prophase

22
I and premature dissociation of bivalents, leading to random assortment of the resulting
univalents at anaphase I. However, Z3-3401 differs from mnm and snm alleles in that it
affects sex chromosome segregation more strongly than autosomal segregation.
Does ectopically expressed MNM-GFP fully complement the meiotic
phenotypes of Z3-3401?: Previous data showed that ectopic, heat-shock-driven
expression of MNM-GFP fully complements the meiotic phenotypes of both Z3-5578
and Z3-3298, thus verifying that the mutations in the mnm exons of those two alleles
are responsible for their mutant phenotypes (THOMAS et al. 2005). However, the
R224C mutation in Z3-3401 should affect the sequences of all 31 Mod(mdg4) isoforms,
the functions of most of which are unknown. Ectopic expression of transgenic MNM-
GFP would be expected to fully complement the meiotic phenotypes of Z3-3401 only if
those phenotypes are caused by disruption of the MNM isoform alone.
To test for rescue of Z3-3401, +/BSYy+; Z3-3401/Df males carrying a single copy
of the [hs::MNM-GFP] transgene on chromosome 2 or 3 were generated, exposed to
variable numbers of heat shocks during development, and tested for X-Y NDJ. The
results were clearcut; both the chromosome 2 and chromosome 3 insertion suppressed
X-Y NDJ below 1% when two heat shocks were given (Table 6). In fact, the
chromosome 2 transgene afforded full rescue even in the absence of heat shock. In
addition, DAPI-stained spermatocytes from transgenic, heat-shocked males and from
non-transgenic males were compared. Spermatocytes from the heat-shocked,
transgene-bearing males appeared indistinguishable from wild-type spermatocytes with
respect to organization and uniformity of spermatid nuclei, absence of univalents at PMI

23
and MI, and equality of telophase 1 poles, whereas their non-transgene-bearing
brothers exhibited the expected array of meiotic anomalies (Fig. 2D).
These results confirm that the R224C mutation in exon 4 is responsible for the
male meiotic phenotypes seen in Z3-3401 males. More interestingly, however, they
indicate that the male meiotic phenotypes of Z3-3401 are due entirely to effects of the
R224C mutation on functioning of the MNM isoform, even though the mutation should
be present in all of the 31 Mod(mdg4) isoforms. These findings could indicate that
MNM is the only mod(mdg4) isoform involved in homolog conjunction in male meiosis.
However, they do not rule out the possibility of other meiotic isoforms of Mod(mdg4) that
are not functionally disrupted by the R224C substitution.
The Z3-3401 mutation disrupts a nuclear localization or retention signal: To
gain additional insight into how the Z3-3401 mutation affects conjunction, we used
antibodies against SNM and against the CR of Mod(mdg4) (anti-ModC) to examine the
localization of MNM and SNM proteins in primary spermatocytes (THOMAS et al. 2005;
BUCHNER et al. 2000). We previously showed that these antibodies co-localize to
multiple nucleolar foci throughout prophase I and to a prominent dense focus
associated with the X-Y bivalent during prometaphase I and metaphase I in wild-type
spermatocytes and (THOMAS et al. 2005). Although the anti-ModC antibody is not
specific for MNM, the dense anti-ModC signal on the X-Y bivalent reflects MNM-specific
staining since it, (along with the anti-SNM signal), is completely abolished in
spermatocytes from Z3-5578 and Z3-3298 males. Very similar results were observed
in Z3-3401 spermatocytes at prometaphase I and metaphase I (Fig. 4). No detectable
staining of the X-Y bivalent with either anti-SNM or anti-ModC was observed in Z3-

24
3401/Df spermatocytes during prometaphase I or metaphase I, although robust staining
was evident in the Z3-3401/+ sibling controls. Thus, Z3-3401 abolishes staining of the
X-Y pairing structure as thoroughly as do snm and mnm alleles. This finding suggests
that the Mod(mdg4) CR as well as the MNM-specific VR exon might be required for
chromosomal localization of MNM and SNM. It is important to note that the failure to
observe staining of autosomal bivalents at prometaphase I in Z3-3401 spermatocytes
does not imply that MNM and SNM are absent from autosomal bivalents at this stage
(which would be difficult to reconcile with the relatively mild disruption of autosomal
segregation in Z3-3401 males) because neither antibody yields detectable staining of
autosomes at this stage in wild-type spermatocytes either (THOMAS et al. 2005).
The absence of anti-MNM staining on condensed X-Y bivalents in Z3-3401
spermatocytes was unexpected since the mutation in Z3-3401 lies far upstream of the
C-terminal domain we have postulated to be responsible for chromosome binding. To
gain further insight into the basis for the Z3-3401 phenotype, we analyzed anti-ModC
staining patterns throughout prophase I, prior to chromosome condensation.
The anti-ModC staining pattern in wild-type spermatocytes exhibited a complex
temporal pattern (Fig. 5). In very young primary spermatocytes (stage S1 (CENCI et al.,
2004)) strong anti-ModC staining was concentrated at the nuclear periphery although
the chromosomes were distributed generally throughout the nucleus. However, by
stage S3 when the three major bivalents have mostly separated from one another and
throughout the remainder of prophase I, anti-ModC staining was largely confined to the
nucleolus and to the DAPI-stained chromosomes. Within chromosomes, anti-ModC
staining was not uniform but rather enriched in particular regions. The anti-ModC

25
staining pattern in mid and late prophase I is similar to that of MNM-GFP (THOMAS et
al., 2005), albeit somewhat less punctate. Thus, unlike condensed autosomal bivalents
at prometaphase I or metaphase I, uncondensed autosomes stain robustly with the anti-
ModC antibody throughout prophase I.
In Z3-5578 and Z3-3298 spermatocytes, early prophase I staining appeared to
be relatively normal both in intensity and localization, with the bulk of the signal
distributed around the nuclear periphery (Fig. 5A). However, by mid-prophase I (not
shown) and more clearly at late prophase I (Fig. 5B), diffuse anti-ModC staining was
distributed throughout the nucleus. Some of this staining overlapped with the DAPI-
stained chromosome territories and might represent other chromosomal Mod(mdg4)
isoforms present in spermatocytes in addition to MNM; however, the staining failed to
show the prominent enrichment on chromosomes and in nucleoli that we observed in
wild-type. These findings suggest that the mutant MNM proteins encoded by Z3-5578
and Z3-3298 are stable and able to gain entry to the nucleus but are unable to localize
properly either to chromosomes or to the nucleolus.
Z3-3401 spermatocytes exhibited a different pattern from the other two mutants.
Strong anti-ModC staining was apparent throughout prophase I but was restricted
primarily to the cytoplasm at all stages (Fig. 5A, B). This contrasts sharply with the
staining patterns in wild-type and mnm spermatocytes, in which little or no cytoplasmic
anti-ModC staining was detected at any stage. These results suggest that the Z3-3401
mutation might disrupt a nuclear localization or retention signal essential for nuclear
localization of MNM. Late prophase I nuclei from Z3-3401 males did exhibit faint
granular staining in the nucleus (Fig. 5B), consistent with the presence of small amounts

26
of mutant MNM protein or of other, less abundant, Mod(mdg4) isoforms in the nucleus.
We conclude that the mutant MNM proteins encoded by all three Z3 alleles are stable
throughout prophase I but improperly localized, to the nucleoplasm in Z3-5578 and Z3-
3298 mutants and to the cytoplasm in Z3-3401.
Is Z3-3401 specific for male meiosis? Z3-3401 differs from most other
mutations in the CR in that it does not affect viability. This could either be because it is
hypomorphic or because it disrupts a domain that is required only for meiotic homolog
conjunction and not for other functions carried out by other Mod(mdg4) proteins. To
assess the specificity of Z3-3401, we evaluated its effects on two other processes
previously shown to be affected by mod(mdg4) mutations: female fertility and chromatin
insulator function.
Effects on female fertility have been previously described for certain viable and
semi-lethal mod(mdg4) genotypes (BUCHNER et al. 2000). To determine if Z3-3401
affects female fertility, sibling Z3-3401/Df and Z3-3401/+ females were crossed singly to
two wild-type males. For comparison, similar crosses involving Z3-3298 and Z3-5578
were also carried out. The results (Table 7) show that Z3-3401 dramatically reduces
female fertility: Z3-3401/Df females produced, on average, only 2.8 progeny each
whereas their heterozygous siblings produced, on average, 42.1 progeny. This fertility
reduction must reflect effects of Z3-3401 on functioning of other isoforms besides MNM
because females hemizygous for either of the MNM-specific alleles exhibited fertility
levels comparable to those of their heterozygous sisters (Table 7) and because ectopic
expression of MNM-GFP in Z3-3401/Df females failed to signficantly improve their
fertility (data not shown).

27
Since mutations in some genes required for female meiosis cause semi-sterility,
we assayed for effects of hemizygosity for Z3-3401 on several meiotic parameters,
including the frequency of X chromosome NDJ and the frequency of recombination in
large intervals on the X, 2nd and 3rd chromosomes (Table 7). Similar assays were
conducted for Z3-3298/Df, Z3-5578/Df and mod(mdg4)T6/Df females. No significant
effects of any of the four tested mod(mdg4) alleles on chromosome segregation or
recombination were observed. Moreover, the eggs laid by Z3-3401/Df females did not
exhibit the “spindle” phenotype caused by incomplete development of the dorsal-ventral
axis that is exhibited by females carrying mutations that disrupt meiotic double-strand
break repair (data not shown; GHABRIAL and SCHUPBACH 1999). Finally,
synaptonemal complex formation, as assayed by incorporation of the central element
protein C(3)G (PAGE and HAWLEY 2001) into linear structures in pachytene oocytes
was found to be normal in Z3-3401/Df females (data not shown). Thus, the fertility
reduction caused by Z3-3401 does not result from disruption of female meiosis. No
further analysis of the female semi-sterility of Z3-3401 has been carried out as yet and it
is not known which Mod(mdg4) protein(s) is/are required for normal female fertility.
Some mod(mdg4) mutations act as enhancers or suppressors of mutations
caused by insertions of gypsy transposable elements into regulatory DNA of certain loci
such as yellow and cut (GEORGIEV and GERASIMOVA 1989; GEORGIEV and
KOZYCINA 1996; GAUSE et al. 2001). These modifier effects are thought to be due to
disruption of the chromatin insulator protein Mod(mdg4)67.2, which localizes, along with
its partner Suppressor of Hairy Wing (Su(Hw)), to gypsy sequences and blocks
activation of nearby promoters by enhancers on the opposite side of the gypsy insertion

28
(GERASIMOVA et al. 1995; GAUSE et al. 2001). To determine whether Z3-3401
disrupts functioning of the Mod(mdg4)67.2 protein, the effects of Z3-3401 on the
pigmentation pattern of y2 flies was examined. y2 flies have pigmented (black) bristles
but unpigmented (yellow) bodies and wings because of a gypsy insertion in the
upstream regulatory region of the yellow locus, which is required for production of the
major pigment in the integument, wings and bristles of adult flies. The gypsy insertion
lies between the yellow promoter and upstream enhancers specific for the body and
wings and therefore blocks activation of the yellow promoter in the body and wings.
However, the bristle-specific enhancer is located in an intron downstream of both the
yellow promoter and the gypsy insertion in y2 and is therefore not prevented from
activating the promoter (GEYER et al. 1986; GEYER and CORCES 1987). The
mod(mdg4)T6 mutation, a nonsense mutation in the 67.2-specific exon, specifically
disrupts functioning of the Mod(mdg4)67.2 protein, leading to loss of insulator function.
The result is that all of the yellow enhancers are silenced by Su(Hw), resulting in a y-
phenotype (yellow bristles, wings and bodies) (GERASIMOVA et al. 1995;
MONGELARD et al. 2002).
To determine if Z3-3401 modifies the y2 phenotype in a similar fashion, we
examined the colors of wings, abdomens and bristles of y2/Y; Z3-3401/mod(mdg4)T16
flies and compared them to y2/Y; mod(mdg4)T6/mod(mdg4)T16, y2/Y; mod(mdg4)T16/+
and y2; Z3-5578/mod(mdg4)T16 flies, scoring at least 20 flies of each genotype. The
result was that all y2; Z3-3401/mod(mdg4)T16 flies were y- in phenotype and
indistinguishable from y2; mod(mdg4)T6/mod(mdg4)T16 flies, whereas the heterozygous
control mod(mdg4)T16/+ and the y2; Z3-5578/mod(mdg4)T16 flies exhibited the y2

29
pigmentation pattern featuring black bristles (data not shown). We conclude that Z3-
3401 acts as an enhancer of y2, and infer that it disrupts the function of the
Mod(mdg4)67.2 isoform. Taken together with the MNM-independent effect of Z3-3401
on female fertility, these data provide strong evidence that the Z3-3401 mutation
disrupts functioning of other Mod(mdg4) isoforms besides MNM.
Does the mod(mdg4) common region have a general role in male homolog
segregation? Our analyses of Z3-3401 indicate that the Mod(mdg4) CR is required for
an indirect step in meiotic homolog conjunction, stable nuclear localization of MNM. If
the CR plays additional roles in homolog conjunction, then other mutations in the CR
should also disrupt homolog conjunction. Although there are numerous extant
mod(mdg4) alleles (FLYBASE, 2006), the great majority are recessive lethals that
cannot be tested directly for meiotic phenotypes. Moreover, clones homozygous for two
such mutations in the germline of mod(mdg4)/+ males were inviable (B. D. McKee,
unpublished data), precluding analysis of meiosis in such clones.
To address the role of the mod(mdg4) CR in meiotic conjunction, we conducted
complementation tests between Z3-3401 and a collection of pre-existing mod(mdg4)
alleles that included 9 small deletions, 3 transposon insertions and 6 EMS-induced
mutations (Table 1 and Fig. 1A). Except for mod(mdg4)T6, which results from a base
substitution in the C-terminal exon specific for the non-essential Mod(mdg4)67.2 protein
(GERASIMOVA et al. 1995; GAUSE et al. 2001), all of the tested mod(mdg4) mutations
and deletions are recessive lethals or semi-lethals, and nearly all have been mapped to
the CR, either by direct molecular identification of lesions within the CR or by failing to
complement mutations with molecular lesions in the CR (GERASIMOVA et al. 1995;

30
BUCHNER et al. 2000; GAUSE et al. 2001). Five of the lethal alleles carry previously
identified lesions within the mod(mdg4) transcription unit: 142∆15, a 1Kb deletion that
removes the promoter and first exon of mod(mdg4); 142∆32, a deletion that removes
upstream regulatory sequences of mod(mdg4) (AZPIAZU and FRASCH 1993) and the
three transposon insertions (2, 3 and neo129), all of which have been molecularly
mapped to sites within the mod(mdg4) common region (Fig. 1A; BUCHNER et al. 2000).
We used a SNP-mapping technique to map the breakpoints of the remaining 7 small
deletions (Table 2) and found that three of the deletions (T16, 142∆33 and 142∆10) are
deficient for the entire mod(mdg4) locus, two (142∆29 and 142∆49) are deficient for part
or all the common region but not deficient for the variable region, and two (eGP4 and
B2) have fully intact mod(mdg4) common regions but are deficient for most of the
variable exons, including the MNM-specific exon (Fig. 1A). In addition, we sequenced
two of the lethal EMS alleles, mod(mdg4)324 and mod(mdg4)340, and found that both
contain base substitutions within the common region. These mutations are predicted to
result in an amino acid substitution, G92D, within the BTB domain of mod(mdg4)324,
and a nonsense mutation at Q177, a site downstream of the BTB domain, in
mod(mdg4)340 (Fig. 1B).
Z3-3401 fully complemented the lethality of all of the lethal and semi-lethal alleles
but the trans-heterozygous males exhibited a complex complementation pattern with
respect to X-Y NDJ (Table 1). Z3-3401 failed to complement most mod(mdg4) alleles,
yielding NDJ frequencies comparable to those of Z3-3401 hemizygotes. However, it
partially or fully complemented the two deletions, B2 and eGP4, which are confined to
the VR, and fully complemented the viable mod(mdg4)T6 allele. Moreover, as

31
described above, it strongly (but not completely) complemented the mnm alleles Z3-
5578 and Z3-3298, the trans-heterozygotes yielding only about 1% X-Y NDJ (Table 3).
The finding that Z3-3401 and mod(mdg4)T6 complement with respect to X-Y NDJ was
expected as no X-Y or 4th chromosome NDJ was observed in mod(mdg4)T6 hemizygous
males (data not shown), indicating that the Mod(mdg4)67.2 isoform is not required for
chromosome segregation in male meiosis.
Parallel complementation tests between the mnm alleles, Z3-5578 and Z3-3298,
and the mod(mdg4) mutations (Table 1) yielded results that were in many cases
opposite to those of Z3-3401. Both mnm alleles fully complemented the lethality of all
mutations and deletions confined to the CR and partially or fully complemented the
same mutations with respect to X-Y NDJ. However, both mnm alleles failed to
complement all mutations that disrupt the MNM-specific exon, including the two
deletions, eGP4 and B2, that remove most of the VR exons (including the MNM-specific
exon) and the three deletions that encompass the entire mod(mdg4) locus (T16,
142∆10 and 142∆33). In addition, both mnm alleles fully complemented mod(mdg4)T6,
which affects only the Mod(mdg4)67.2-specific exon.
Thus, all of the complementation results are consistent with a pattern in which
mutations or deletions confined to the CR complement (partially or fully) mutations or
deletions confined to the VR. Moreover, mutations in two different variable exons
complement each other. Notably, deletions and point mutations exhibited similar
complementation behavior, thus ruling out the possibility that intragenic
complementation results from formation of functional protein homo-dimers from two
monomers with mutations in different domains.

32
MNM transcripts are generated by trans-splicing: Intragenic
complementation between mutations in the CR and in the C-terminal exon specific for
the Mod(mdg4)67.2 isoform has been previously reported (MONGELARD et al. 2002).
Complementation in that case has been shown to be due trans-splicing between
separate precursor transcripts for the CR and VR components of Mod(mdg4)67.2
expressed from opposite homologs (LABRADOR et al. 2001; MONGELARD et al.
2002). As the common and variable regions of Mod(mdg4)67.2 are encoded on
opposite genomic strands, the mature Mod(mdg4)67.2 transcript can presumably be
generated only by trans-splicing. Since all MNM exons are encoded on the same
strand, generation of MNM transcripts by conventional cis-splicing should be possible
(see Fig. 1A). However, our complementation data suggest that at least some MNM
transcripts are trans-spliced and, moreover, that trans-homolog trans-splicing (THTS)
(Fig. 6) makes a significant contribution to the pool of MNM transcripts.
To test for MNM transcripts derived from THTS, we sequenced a sample of
cloned testis cDNAs from Z3-3401/Z3-5578 (a complementing genotype, Table 2)
prepared by RT-PCR amplification of a segment of MNM that encompasses the sites of
both mutant lesions. If splicing occurs exclusively in the cis mode, all cDNAs from these
males should contain either the Z3-3401 mutation or the Z3-5578 mutation but never
both (Fig. 6B). However, if THTS occurs at a significant frequency, cDNAs that are
wild-type at both sites or mutant at both sites should be recovered. Of 137 cDNAs from
Z3-3401/Z3-5578 males, 55 (40%) were either wild-type at both sites or mutant at both
sites, suggesting that THTS does occur at a substantial frequency (Fig. 6C).

33
A technical complication in experiments of this type is that wild-type and doubly
mutant cDNAs can arise in vitro from singly mutant transcripts by template switching
during the PCR reaction (TASIC et al. 2002). To assess the frequency of such artifacts
under the conditions of our experiments, we carried out a control experiment in which
the RNA templates consisted of an equimolar mixture of singly mutant RNAs prepared
from testes of males hemizygous for Z3-3401 or Z3-5578. In this control, wild-type or
doubly mutant cDNAs can be generated only by template switching during the PCR
reaction. Of 130 control cDNAs, 21 (16%) were either wild-type at both sites or mutant
at both sites (Fig. 6C). Thus, template switching does occur at a significant frequency in
this experiment and we cannot conclude that the wild-type and doubly mutant cDNAs in
the trans-heterozygous sample all resulted from in vivo trans-splicing events. However,
a statistical analysis shows that the difference in frequency of wild-type and doubly
mutant cDNAs between experimental and control samples is highly significant (χ2 =
18.8, 1d.f., p<<.001), thus supporting the THTS hypothesis. Taking into account the
error introduced by template switching in both directions, we estimate that 35% (95%
confidence interval = 27-43%) of MNM mRNAs from Z3-3401/Z3-5578 males are either
wild-type at both sites or doubly mutant at both sites, presumably due to THTS.
As a further test of the trans-splicing hypothesis, we evaluated the ability of a
transgene [CR-7.5], located on chromosome 2, that carries a 7.5kb genomic fragment
encompassing the entire CR of mod(mdg4) but that lacks MNM-specific sequences (see
Fig. 1A), for ability to rescue the meiotic phenotypes of Z3-3401. This transgene
partially rescues the lethality of some mutations in the CR (BUCHNER et al. 2000),
presumably via trans-splicing between precursor transcripts for the CR and VR

34
components of one or more Mod(mdg4) isoforms encoded by the transgene and by the
native mod(mdg4) locus, respectively.
To test for complementation of Z3-3401 by the [CR-7.5] transgene, Z3-3401/Z3-
3401, Z3-3401//mod(mdg4)neo129 or Z3-3401/Df(3R)T16 males (as well as control Z3-
5578/Df males) with and without the transgene were tested for X-Y NDJ. In all three
sets of crosses involving Z3-3401, NDJ frequencies were significantly lower in the
presence of the transgene than in its absence (Table 8), whereas the transgene had no
effect on NDJ frequencies in Z3-5578 males (data not shown). These results indicate
that partial MNM precursor transcripts generated at distant genomic locations can trans-
splice at a high enough frequency in spermatocytes to significantly ameliorate the
phenotype of a strong meiotic mutant. Presumably, the relatively weak rescue afforded
by the transgene reflects decreased efficiency of trans-splicing due to spatial separation
of the coding sequences for the CR and VR portions of the MNM transcript.
DISCUSSION
Z3-3401 is an allele of mod(mdg4) that disrupts X-Y segregation more
severely than autosomal segregation. Our previous results demonstrated that
mutations in a mod(mdg4) exon presumed to be specific for the C-terminus of the MNM
isoform disrupt stable conjunction and regular segregation of all four homolog pairs in
male meiosis I. Here we have shown that a mutation located in the CR of mod(mdg4)
and that, therefore, should be present in all Mod(mdg4) isoforms, causes a similar
spectrum of meiotic defects. Like the mnm-specific mutations, Z3-3401 disrupts the
maintenance of homolog associations from mid-prophase I through M I but has no

35
apparent effect on the initiation of homolog pairing in early meiosis, on the stability of
sister chromatid cohesion or on any aspect of female meiosis.
However, the effects of Z3-3401 on autosomal segregation are much milder than
its effects on autosomal segregaton. In Z3-3401 hemizygous males, X-Y NDJ occurs at
frequencies in the 40-50% range, consistent with nearly random assortment, but 2nd and
4th chromosome NDJ frequencies are in the 10-20% range. This is not because MNM
plays a less critical role in autosomal than in X-Y segregation since both mnm-specific
alleles disrupt X-Y and autosomal segregation equally severely. The basis for this
partial X-Y specificity is not known.
One possible explanation is that the X-Y and autosomal pairs exhibit different
thresholds of sensitivity to reduction in amount of MNM protein in the nucleus, either
because X-Y pairing sites need more MNM protein than autosomal pairing sites to
associate stably, or because MNM is loaded more efficiently on autosomal than X-Y
pairing sites when MNM is present in limiting amounts. Some functional MNM protein
must be present on both autosomes and the X-Y pair in Z3-3401 spermatocytes, as
indicated both by the mild autosomal segregation defect and by the fact that Z3-3401 is
hypomorphic for both autosomal and X-Y segregation. Moreover, late prophase I nuclei
appear to contain small amounts of Mod(mdg4) proteins, some of which may represent
functional, chromosome-associated MNM. The suggestion that MNM might load more
efficiently on autosomes than on the X-Y pair is consistent with our observation that
MNM protein is recruited to the X-Y and autosomal pairs by different mechanisms. Both
autosomal conjunction and autosomal recruitment of MNM-GFP are dependent upon

36
tef+ function, whereas both X-Y conjunction and recruitment of MNM-GFP to the X-Y
bivalent are independent of tef+ (TOMKIEL et al. 2001; THOMAS et al., 2005).
An alternative explanation is suggested by our finding that mutant MNM protein is
present throughout prophase I in Z3-3401 spermatocytes but confined to the cytoplasm
prior to the onset of prometaphase I. Although MNM and SNM are normally present in
the nucleus and associated with the chromosomes and nucleoli from early prophase I
on, we have no direct evidence that their presence is required for stable conjunction
prior to chromosome condensation. Perhaps conjunction of at least the autosomal
homologs is not stably attained until they begin condensing just prior to prometaphase I.
In that case, mutant Z3-3401 protein might be able to gain access to and help conjoin
the condensing chromosomes at the onset of prometaphase I when the nucleus
becomes permeable to cytoplasmic proteins. The differential effect of Z3-3401 on
autosomal versus sex chromosome conjunction could reflect differential timing of the
establishment of autosomal versus X-Y conjunction or differential access of MNM and
SNM to autosomal versus X-Y pairing sites at late stages.
Do other mod(mdg4) isoforms function in meiosis?: We have suggested
that MNM might function in homolog pairing by binding specific chromosomal sites on
homologous chromosomes at its C-terminal C2H2 domain and utilizing its N-terminal
BTB/POZ domain to glue those sites together (THOMAS et al. 2005). mod(mdg4) is
thought to be capable of encoding 30 additional proteins with identical BTB/POZ
domains but different chromosome localization domains. Thus an appealing possibility
is that some of those additional proteins could play similar roles in homolog pairing in
male or female meiosis, or in somatic cells. Moreover, if Mod(mdg4) isoforms can

37
heterodimerize, which seems likely given that their BTB domains are identical, to form
proteins with unique chromosome localization patterns, mod(mdg4) might have the
potential to encode a small army of site-specific pairing proteins.
However, the data presented here provide no support for such a scenario.
Analysis of a mutation in the exon specific for the abundantly expressed
Mod(mdg4)67.2 isoform revealed no meiotic phenotypes in either sex. Moreover, Z3-
3401, which contains an R224C substitution which should be present in all Mod(mdg4)
isoforms, nevertheless exhibited no meiotic phenotypes that could not be accounted for
by effects on MNM. This is not because R224 is part of a domain required only for
MNM function. Z3-3401 flies are functionally mutant for at least two other Mod(mdg4)
isoforms besides MNM: the Mod(mdg4)67.2 insulator protein and at least one
unidentified isoform required for female fertility. Thus, the Z3-3401 mutation failed to
reveal additional meiotic proteins encoded by mod(mdg4). However, Z3-3401 has no
effect on viability whereas most mutations in the common region are recessive lethals,
suggesting that some Mod(mdg4) isoforms may be unaffected by the R224C
substitution. Thus, our findings do not rule out the existence of other meiotic
Mod(mdg4) isoforms that are functionally unaffected by the R224C substitution (e.g., if
the C-terminal exon encodes a nuclear localization sequence (NLS)). Genetic analysis
of mutations specific for other isoforms besides MNM and Mod(mdg4)67.2 will be
needed to test definitively for the existence of other meiotic proteins encoded by
mod(mdg4).
MNM is expressed predominantly by a mechanism involving trans-splicing:
At least some of the mature transcripts for Mod(mdg4) proteins are generated by an

38
unusual mechanism in which the CR and VR exons are separately transcribed and
spliced together in trans (LABRADOR and CORCES 2001; DORN et al. 2001). For
some isoforms, such as Mod(mdg4)67.2, trans-splicing is presumably essential for
expression, as the common and variable regions are encoded on opposite strands.
However, for most isoforms, including MNM, the CR and VR components are encoded
on the same strand and the relative contributions of cis- versus trans-splicing
mechanisms in their expression has been unclear.
Our data indicate that trans-splicing is the predominant mode of expression of
MNM. Complementation analysis demonstrated that mutations of all molecular types,
including deletions, in the CR partially or fully complement mutations in or deletions of
MNM, a pattern difficult to explain except by THTS. Sequence analysis revealed that a
significant fraction, 35% plus or minus 8%, of the mRNAs present in testis extracts from
trans-heterozygous Z3-5578/Z3-3401 males were either wild-type for both mutant sites
or doubly mutant at both sites. We presume that all such RNAs result from THTS.
These data allow a rough estimate of the relative frequency of trans- versus cis-
splicing in generation of MNM-encoding mRNAs. Assuming no bias for or against
splicing between precursor transcripts expressed from opposite homologs versus the
same homolog, an additional 35% of transcripts should result from trans-splicing
between precursors encoded on the same homolog, but would nevertheless be singly
mutant and therefore indistinguishable from cis-spliced transcripts. This leads to a total
estimate of the fraction of trans-spliced MNM transcripts of 70%. This figure could
substantially underestimate the frequency of trans-splicing since any significant spatial
separation between homologs would be expected to create a bias in favor of splicing

39
between precursors encoded on the same homolog. Thus we conclude that a majority
and perhaps all, of the MNM-encoding transcripts synthesized in spermatocytes are
generated by trans-splicing.
Our data provide the first example of the routine use of trans-splicing at the
native mod(mdg4) locus to synthesize an isoform whose exons are all encoded on the
same strand. They also extend the range of Drosophila cell types shown to be
competent for trans-splicing to the male germ line. Previous results have shown that
embryonic and somatic isoforms of the mod(mdg4) and lola genes can be generated by
trans-splicing and that somatic phenotypes controlled by these genes are subject to
intragenic complementation via THTS (DORN et al. 2001; MONGELARD et al. 2002;
HORIUCHI et al. 2003; GABLER et al. 2005). Our data indicate that spermatogonia
and/or young spermatocytes must also be competent to carry out trans-splicing.
It remains unclear how the expression of MNM is regulated. The function of
MNM appears to be limited to male meiosis and MNM protein has thus far been
detected only in primary spermatocytes. Our data demonstrating that MNM is
expressed predominantly by trans-splicing suggest that MNM expression could be
regulated either by testis-specific transcription of the C-terminal MNM exon or by
regulated trans-splicing. However, even when the intronless MNM-GFP transgene is
expressed from the ubiquitous hsp70 promoter under heat-shock conditions, we have
failed to detect significant amounts of MNM-GFP in somatic or gonial cells of the testis
or in larval salivary gland cells (M. SOLTANI-BEJNOOD, S.E. THOMAS and B.D.
MCKEE, unpublished data), suggesting that MNM protein may be unstable in cell types
other than spermatocytes. Moreover, MNM cDNAs have been cloned from embryonic

40
RNA by two different groups (HARVEY et al. 1997; BUCHNER et al. 2000), indicating
that transcription of the MNM-specific exon and splicing between the common region
and MNM precursor transcripts are not exclusive to male germ cells. It will be important
to determine the relative contributions of regulatory mechanisms at the level of
transcription, splicing, translation and protein stability in delimiting expression of MNM to
primary spermatocytes.
The role of the Mod(mdg4) common region in MNM function: We have
previously shown that mutations in a single VR exon of mod(mdg4) disrupt homolog
conjunction in male meiosis. In this study, we sought and obtained evidence that the
Mod(mdg4) CR is also required for homolog conjunction. The Z3-3401 allele, which
contains a missense mutation in the CR, causes meiotic phenotypes very similar to
those of the mnm alleles. Moreover, males trans-heterozygous for Z3-3401 and any of
17 lethal or semi-lethal mod(mdg4) alleles, most of which contain lesions in the CR,
exhibited similar meiotic phenotypes.
What role(s) does the mod(mdg4) CR play in homolog conjunction? We
previously suggested (THOMAS et al. 2005) that the BTB/POZ domain, which has been
shown to mediate dimerization and multimerization (GERASIMOVA and CORCES
1998; GERASIMOVA et al. 2000; GHOSH et al. 2001; GAUSE et al. 2001), might
function to stably connect MNM-containing conjunction complexes localized to allelic
sites on homologous chromosomes. Unexpectedly, immunofluoresence analysis
revealed that both MNM and SMN are undetectable on condensed X-Y bivalents in Z3-
3401 spermatocytes and that, although mutant MNM protein is detectable prior to
chromosome condensation, the great majority of it is restricted to the cytoplasm

41
throughout prophase I. This suggests that the Z3-3401 mutation does not disrupt
conjunction directly but rather disrupts a nuclear localization or nuclear retention signal
required for stable nuclear localization of MNM. Although the sequence surrounding
amino acid 224 in mod(mdg4) does not fully conform to known NLS sequences, R224 is
part of a two-residue cluster of basic amino acids (KR), which are critical components of
classical NLS sequences (MATTAJ and ENGELMEIER 1998). It will be of interest to
learn whether addition of a classical NLS to the Z3-3401 MNM protein restores nuclear
localization, and, if so, whether this alone suffices to restore wild-type function.
The fact that all of the lethal and semi-lethal mod(mdg4) alleles, including five
EMS-induced alleles, fully failed to complement Z3-3401 for meiotic chromosome
segregation suggests that the CR is likely to play additional roles in homolog
conjunction besides nuclear localization. The most interesting allele is mod(mdg4)324,
which is predicted to contain a G92D substitution. G92 lies within the BTB domain and
is conserved among 32 of 44 Drosophila BTB-domain proteins, including GAGA,
Tramtrack, Broad Complex and Bric a Brac (ZOLLMAN et al. 1994; READ et al. 2000).
By homology modeling, G92 is predicted to be important for formation of a β-sheet
crucial for homodimerization (READ et al. 2000). Although we have not established
whether the mutant MNM protein produced by mod(mdg4)324 is stable and/or localized
properly in spermatocytes, a mutant GAGA protein that contains a G to S substitution at
the equivalent residue in the BTB domain (which is highly similar to that of mod(mdg4)),
as well as a second substitution at a conserved residue in the BTB domain, is
expressed at normal levels in Drosophila S2 cells and localizes normally to
chromosomal foci (READ et al. 2000). Thus, the mod(mdg4)324 mutation may provide

42
a very useful tool for analysis of the role of the mod(mdg4) BTB domain in homolog
conjunction.
ACKNOWLEDGEMENTS
We thank C. Zuker, B. Wakimoto, R. Dorn, V. Corces, M. Frasch and the Drosophila
Stock Center at Indiana University, Bloomington for gifts of Drosophila stocks and
antibodies. Support for this work was provided by grant # R01 GM40489 from the
National Institutes of Health to B.D.M.
LITERATURE CITED
AZPIAZU, N., and M. FRASCH, 1993. tinman and bagpipe: two homeobox genes that
determine cell fates in the dorsal mesoderm of Drosophila. Genes Devel. 7:
1325-1340.
BARDWELL, V. J., and R. TREISMAN, 1994. The POZ domain: a conserved protein-
protein interaction motif. Genes Dev. 8: 1664-1677.
BUCHNER, K., P. ROTH, G. SCHOTTA, V. KRAUSS, H. SAUMWEBER, et al., 2000.
Genetic and molecular complexity of the position effect variegation modifier
mod(mdg4) in Drosophila. Genetics 155: 141-157.
CAI, H. N., and M. LEVINE, 1997. The gypsy insulator can function as a promoter-
specific silencer in the Drosophila embryo. EMBO J. 16: 1732-1741.
CARPENTER, A. T., 1994. Chiasma function. Cell 77: 957-962.
CENCI, G., S. BONACCORSI, C. PISANO, F. VERNI and M. GATTI, 1994. Chromatin
and microtubule organization during premeiotic, meiotic and early postmeiotic

43
stages of Drosophila melanogaster spermatogenesis. J. Cell Sci. 107: 3521-
3534.
CHEN, S. and V. G. CORCES, 2001. The gypsy insulator of Drosophila affects
chromatin structure in a directional manner. Genetics 159: 1649-1658.
DORN, R., and V. KRAUSS, 2003. The modifier of mdg4 locus in Drosophila: functional
complexity is resolved by trans splicing. Genetica 117: 165-177.
DORN, R., V. KRAUSS, G. REUTER and H. SAUMWEBER, 1993. The enhancer of
position-effect variegation of Drosophila, E(var)3-93D, codes for a chromatin
protein containing a conserved domain common to several transcriptional
regulators. Proc. Natl. Acad. Sci. USA 90: 11376-11380.
DORN, R., G. REUTER and A. LOEWENDORF, 2001. Transgene analysis proves
mRNA trans-splicing at the complex mod(mdg4) locus in Drosophila. Proc. Natl.
Acad. Sci. USA 98: 9724-9729.
ESPINAS, M. L., E. JIMENEZ-GARCIA, A. VAQUERO, S. CANUDAS, J. BERNUES et
al., 1999. The N-terminal POZ domain of GAGA mediates the formation of
oligomers that bind DNA with high affinity and specificity. J. Biol. Chem. 274:
16461-16469.
FLYBASE. URL: http://flybase.bio.indiana.edu/
GABLER, M., M. VOLKMAR, S. WEINLICH, A. HERBST, P. DOBBERTHIEN et al.,
2005. Trans-splicing of the mod(mdg4) complex locus is conserved between the
distantly related species Drosophila melanogaster and D. virilis. Genetics 169:
723-736.

44
GAUSE, M., P. MORCILLO and D. DORSETT, 2001. Insulation of enhancer-promoter
communication by a gypsy transposon insert in the Drosophila cut gene:
cooperation between Suppressor of Hairy-wing and Modifier of mdg4 proteins.
Mol. Cell. Biol. 21: 4807-4817.
GEORGIEV, P. G. and T. I. GERASIMOVA, 1989. Novel genes influencing the
expression of the yellow locus and mdg4 (gypsy) in Drosophila melanogaster.
Mol. Gen. Genet. 220: 121-126.
GEORGIEV, P. G. and M. KOZYCINA 1996. Interaction between mutations in the
suppressor of Hairy wing and modifier of mdg4 genes of Drosophila
melanogaster affecting the phenotype of gypsy-induced mutations. Genetics
142: 425-436.
GERASIMOVA, T. I., K. BYRD and V. G. CORCES, 2000. A chromatin insulator
determines the nuclear localization of DNA. Mol. Cell 6: 1025-1035.
GERASIMOVA, T. I., and V. G. CORCES, 1998. Polycomb and trithorax group proteins
mediate the function of a chromatin insulator. Cell 92: 511-521.
GERASIMOVA, T. I., D. A. GDULA, D. V. GERASIMOV, O. SIMINOVA and V. G.
CORCES, 1995. A Drosophila protein that imparts directionality on a chromatin
insulator is an enhancer of position-effect variegation. Cell 82: 587-597.
GEYER, P. K., C. SPANA and V. G. CORCES, 1986. On the molecular mechanism of
gypsy-induced mutations at the yellow locus of Drosophila melanogaster. EMBO
J. 5: 2657-2662.
GEYER, P. K. and V. G. CORCES, 1987. Separate regulatory elements are
responsible for the complex pattern of tissue-specific and developmental

45
transcription of the yellow locus in Drosophila melanogaster. Genes Dev. 1: 996-
1004.
GEYER, P. K., and V. G. CORCES, 1992. DNA position-specific repression of
transcription by a Drosophila zinc-finger protein. Genes Devel. 6: 1865-1873.
GHABRIAL, A. and T. SCHUPBACH, 1999. Activation of a meiotic checkpoint
regulates translation of gurken during Drosophila oogenesis. Nature Cell Biol 1:
354-357.
GHOSH, D., T. I. GERASIMOVA and V. G. CORCES, 2001. Interactions between the
Su(Hw) and Mod(mdg4) proteins required for gypsy insulator function. EMBO J.
20: 2518-2527.
GORCZYCA, M., E. POPOVA, X. X. JIA and V. BUDNICK, 1999. The gene mod(mdg4)
affects synapse specificity and structure in Drosophila. J. Neurobiol. 39: 447-
460.
GRANOK, H., B. A. LIEBOVITCH, C. D. SCHAFFER, S. C. R. ELGIN, 1995. Ga-ga
over GAGA factor. Curr. Biol. 5: 238-241.
GUNSALUS, K. C., S. BONACCORSI, E. WILLIAMS, F. VERNI, M. GATTI et al., 1995.
Mutations in twinstar, a Drosophila gene encoding a cofilin/ADF homologue,
result in defects in centrosome migration and cytokinesis. J. Cell Biol. 131: 1243-
1259.
HARVEY, A. J., A. P. BIDWAL and L. K. MILLER, 1997. Doom, a product of the
Drosophila mod(mdg4) gene induces apoptosis and binds to Baculovirus
inhibitor-of-apoptosis proteins. Mol. Cell. Biol. 17: 2835-2843.

46
HASSOLD, T., and P. HUNT, 2001. To err (meiotically) is human: the genesis of human
aneuploidy. Nat. Rev. Genet. 2: 280-291.
HAWLEY, R. S., 1988. Exchange and chromosomal segregation in eucaryotes, pp. 497-
527 in Genetic Recombination, edited by R. Kucherlapati and G. R. Smith.
American Society for Microbiology, Washington, D.C.
HORIUCHI, T., E. GINIGERT and T. AIGAKI, 2003. Alternative trans-splicing of
constant and variable exons of a Drosophila axon guidance gene, lola. Genes
Devel. 17: 2496-2501.
HOSKINS, R. A., A. C. PHAN, M. NAEEMUDDIN, F. A. MAPA, D. A. RUDDY, J. J.
RYAN, L. M. YOUNG, T. WELLS, C. KOPCZYNSKI and M. C. ELLIS, 2001.
Single nucleotide polymorphism markers for genetic mapping in Drosophila
melanogaster. Genome Res. 11: 1100-1113.
IGARISHI, K., H. HOSHINO, A. MUTO, N. SUWABE, S. NISHIKAWA et al., 1998.
Multivalent DNA binding complex generated by Small Maf and Bach1 as a
possible biochemical basis for B-Globin Locus Control Region complex. J. Biol.
Chem. 273: 11783-11790.
KATSANI, K. R., M. A. HAJIBAGHERI and C. P. VERRIJZER, 1999. Co-operative DNA
binding by GAGA transcription factor requires the conserved BTB/POZ domain
and reorganizes promoter topology. EMBO J. 18: 698-708.
KOUNDAKJIAN, E. J., D. M. COWAN, R. W. HARDY and A. H. BECKER, 2004. The
Zuker collection: a resource for the analysis of autosomal gene function in
Drosophila melanogaster. Genetics 167: 203-206.

47
LABRADOR, M., F. MONGELARD, P. PLATA-RENGIFO, E. M. BAXTER, V. G.
CORCES and T. I. GERASIMOVA, 2001. Protein encoding by both strands.
Nature 409: 1000.
LABRADOR, M., and V. G. CORCES, 2003. Extensive exon reshuffling over
evolutionary time coupled to trans-splicing in Drosophila. Genome Res. 13: 2220-
2228.
MATTAJ, I. W., ENGLMEIER, L., 1998. Nucleocytoplasmic transport: the soluble
phase. Annu. Rev. Biochem. 67: 265-306.
McKEE, B. D., 1996. The license to pair: identification of meiotic pairing sites in
Drosophila. Chromosoma 105: 135-141.
McKEE, B. D., 2004. Homologous pairing and chromosome dynamics in meiosis and mitosis.
Biochim. biophys. acta 1677: 165-180.
McKEE, B. D., and G. H. KARPEN, 1990. Drosophila ribosomal RNA genes function as
an X-Y pairing site during male meiosis. Cell 61: 61-72.
McKEE, B. D., and D. L. LINDSLEY, 1987. Inseparability of X-heterochromatic
functions responsible for X:Y pairing, meiotic drive, and male fertility in
Drosophila melanogaster males. Genetics 116: 399-407.
MELNICK, A., K. F. AHMAD, S. ARAI, A. POLINGER, H. BALL et al., 2000. In-depth
mutational analysis of the promyelocytic leukemia zinc finger BTB/POZ Domain
reveals motifs and residues required for biological and transcriptional functions.
Mol. Cell. Biol. 20: 6550-6567.
MOHLER, J., and M. L. PARDUE, 1982. Deficiency mapping of the 93D heat shock
locus in Drosophila melanogaster. Chromosoma 86: 457-467.

48
MOHLER, J., and M. L. PARDUE, 1984. Mutational analysis of the region surrounding
the 93D heat shock locus of Drosophila melanogaster. Genetics 106: 249-265.
MONGELARD, F., M. LABRADOR, E. M. BAXTER, T. I. GERASIMOVA and V. G. CORCES,
2002. Trans-splicing as a novel mechanisms to explain inter-allelic complementation in
Drosophila. Genetics 160: 1481-1487.
MULLER, S. A., T. SASAKI, P. BORK, B. WOLPENSINGER, T. SCHULTHESS et al.,
1999. Domain organization of Mac-2 binding protein and its oligomerization to
linear and ring-like structures. J. Mol. Biol. 291: 801-813.
PAGE, S. L. and R. S. HAWLEY, 2001. c(3)G encodes a Drosophila synaptonemal
complex protein. Genes Dev 15: 3130-3143.
PAGE, S. L, and R. S. HAWLEY, 2003. Chromosome choreography: the meiotic ballet.
Science 301: 785-789.
PAGE, S. L., and R. S. HAWLEY, 2004. The genetics and molecular biology of the
synaptonemal complex. Annu. Rev. Cell. Dev. Biol. 20: 525-558.
PARK, H. S., and M. T. YAMAMOTO, 1995. The centric region of the X chromosome
rDNA functions in male meiotic pairing in Drosophila melanogaster.
Chromosoma 103: 700-707.
RASOOLY, R. S., and L. G. ROBBINS, 1991. Rex and a suppressor of Rex are
repeated neophorphic loci in the Drosophila melanogaster ribosomal DNA.
Genetics 129: 119-132.
READ, D., M. J. BUTTE, A. F. DERNBURG, M. FRASCH and T. B. KORNBERG, 2000.
Functional studies of the BTB domain in the Drosophila GAGA and Mod(mdg4)
proteins. Nucl. Acids Res. 28: 3864-3870.

49
ROBINETT, C. C., A. STRAIGHT, G. LI, C. WILLHELM, G. SUDLOW et al., 1996. In
vivo localization of DNA sequences and visualization of large-scale chromatin
organization using lac operator/repressor recognition. J. Cell Biol. 135: 1685-
1700.
ROEDER, G. S., 1997. Meiotic chromosomes: it takes two to tango. Genes Dev. 11:
2600-2621.
STOGIOS, P. J., G. S. DOWNS, J. J. S. JAUHAL, S. K. NANDRA and G. G. PRIVE, 2005.
Sequence and structural analysis of BTB domain proteins. Genome Biology 6: R82.
STRAIGHT, A. F., A. S. BELMONT, C. C. ROBINETT and A. W. MURRAY, 1996. GFP
tagging of budding yeast chromosomes reveals that protein-protein interactions can
mediate sister chromatid cohesion. Curr. Biol. 6: 1599-1608.
TASIC, B., C. E. NABHOLZ, K. K. BALDWIN, Y. KIM, E. H. RUECKERT, et al., 2002.
Promoter choice determines splice site selection in protocadherin α and γ pre-mRNA
splicing. Mol. Cell 10: 21-33.
THOMAS, S. E., M. SOLTANI-BEJNOOD, P. ROTH, R. DORN, J. M. LOGSDON et al., 2005.
Identification of two proteins required for conjunction and regular segregation of
achiasmate homologs in Drosophila male meiosis. Cell 123: 555-568.
TOMKIEL, J. E., B. T. WAKIMOTO and A. BRISCOE, Jr., 2001. The teflon gene is
required for maintenance of autosomal homolog pairing at meiosis I in male
Drosophila melanogaster. Genetics 157: 273-281.
VAZQUEZ, J., A. S. BELMONT and J. W. SEDAT, 2002. The dynamics of homologous
chromosome pairing during male Drosophila meiosis. Curr. Biol. 12: 1473-1483.

50
WAKIMOTO, B. T., D. L. LINDSLEY and C. HERRERA, 2004. Toward a comprehensive
genetic analysis of male fertility in Drosophila melanogaster. Genetics 167: 207-
216.
WOLF, K. W., 1994. How meiotic cells deal with non-exchange chromosomes.
Bioessays 16: 107-114.
ZOLLMANN, S., D. GODT, G. G. PRIVE, J. L. COUDERC and F. A. LASKI, 1994. The
BTB domain, found primarily in zinc-finger proteins, defines an evolutionarily
conserved family that includes several developmentally regulated genes in
Drosophila. Proc. Natl. Acad. Sci. USA 91: 10717-10721.

51
FIGURE LEGENDS
FIGURE 1. Structure of the mod(mdg4) locus and molecular identification of Z3-3401.
(A) Map of the mod(mdg4) locus, modified from DORN et al. (2001). Rectangles
represent exons (white for untranslated exons, black for translated exons). The
common region (CR) contains four exons common to all Mod(mdg4) transcripts,
transcribed left-to-right and cis-spliced. The variable region (VR) contains C-terminal
exons specific for each isoform on the same strand (below the line) or the opposite
strand (above the line) from the CR. Diagonal lines represent splice sites. Cross-
hatched rectangles represent C-terminal exons of MNM. Mapped mutational lesions are
indicated by triangles (transposon insertions), asterisks (point mutations) or dark lines
below the exons (deletions). Endpoints with diamonds represent deletion breakpoints
mapped within mod(mdg4); endpoints with arrows represent deletion breakpoints
outside mod(mdg4). Dashed line (CR-7.5) depicts the 7.5Kb genomic rescue fragment
used in the experiments in Table 8 (BUCHNER et al. 2000). See Table 5 legend for
references for mod(mdg4) alleles. (B) Sequence of MNM protein, based on conceptual
translation of a cDNA isolated from y w testis RNA (THOMAS et al., 2005). Altered
residues in Z3 mutations and in mod(mdg4)324 and mod(mdg4)340 are indicated by
bold-face with mutant residue above wild-type residue. BTB domain is shaded;
FLYWCH domain is underlined; C2H2 motif is enlarged.
FIGURE 2. Cytological analysis of male meiosis in Z3-3401.
(A) Representative figures from various stages of meiosis in wild-type and Z3-3401/Z3-
3401 males stained with DAPI to visualize the DNA and an anti-tubulin antibody to

52
visualize the spindles. Round spermatid nuclei (rightmost) panel are unequal in size in
Z3-3401, indicative of NDJ. Prometaphase I and metaphase I cells exhibit too many
DNA clumps, suggestive of conjunction failure, and anaphase I cells exhibit unequal
poles, indicative of missegregation of homologs at meiosis I. Meiosis II chromosome
morphology and segregation appear normal.
(B) Quantification of meiosis I abnormalities in Z3-3401 spermatocytes. Data from wild-
type and Z3-3298 spermatocytes (THOMAS et al. 2005) provided for comparison.
(C) Meiosis I chromosomes from wild-type and Z3-3401 homozygous males stained
with acetic orcein to visualize DNA and observed by phase contrast microscopy. There
are four large univalent (unpaired) chromosomes (red arrows) and one bivalent at
prometaphase I and six large univalent chromosomes at metaphase I in Z3-3401
spermatocytes. Note the lagging chromosomes at anaphase I in Z3-3401.
(D) Representative meiotic figures from hemizygous Z3-3401/T16 males with or without
one copy of [hs::MNM-GFP]3. Both groups of males were heat-shocked twice during
larval development. Red arrows point to univalents in late prophase I; unpaired X and Y
chromosomes in upper right.
FIGURE 3. Homologous pairing and territory expansion in Z3-3401. Chromosomal
GFP-LacI foci were imaged in germ cell nuclei from wild-type (Z3-3401/+ or
Df(3R)T16/+) and Z3-3401/mod(mdg4)T16 males homozygous for a single lacO array
on chromosome arm 2R and hemizygous for [hsp83::GFP-LacI](2) (VAZQUEZ et al.
2002). (A) Homologous pairing in spermatogonia (gonia) and early prophase I (PI)
spermatocytes. Nuclei with one spot were scored as paired; nuclei with more than one

53
spot were scored as unpaired. (B) GFP-LacI spot dispersion in late prophase I
spermatocytes. Mean Nuc. Diam. = mean nuclear diameter (in uM) +/- standard
deviation; Mean Disp. = mean spot dispersion (in uM) +/- standard deviation. N =
number of nuclei scored.
FIGURE 4. Localization of MNM and SNM proteins to the X-Y bivalent is disrupted by
the Z3-3401 mutation. Spindles and DNA were stained with anti-tubulin (green) and
DAPI (blue), respectively, in right panels. Both anti-MNM (A) and anti-SNM (B)
antibodies stain a compact structure associated with the X-Y bivalent at prometaphase I
in wild-type spermatocytes (top rows). No staining of condensed meiotic chromosomes
with anti-MNM or anti-SNM is detected in Z3-3401 spermatocytes (bottom rows).
FIGURE 5. Mod(mdg4) proteins in early-mid (A) and late (B) prophase I spermatocytes
in wild-type (WT) and Z3 mod(mdg4) mutants. Dissected testes were stained with anti-
ModC antibody, which reacts against the common region of Mod(mdg4) proteins, anti-
tubulin to stain the spindles and DAPI to stain DNA. Nomenclature for stages follows
Cenci et al. (1994). S1-S2/early prophase I; S3-S4/mid-prophase I; S5-S6/late
prophase I. White arrows indicate nucleoli.
FIGURE 6. Trans-homolog trans-splicing in males trans-heterozygous for Z3-5578 and
Z3-3401. (A) Possible expression patterns for components of MNM. Expression of
common (black) and variable (red) exons of MNM continuously on a single precursor
transcript (above each homolog) or discontinuously on two separate transcripts (below

54
each homolog). m and + signify mutant and wild-type sites. (B) Trans- versus cis-
splicing. Both cis- and trans-splicing can generate parental-type (P) mRNAs. Only
trans-splicing can generate recombinant (R) mRNAs. (C) Estimated fraction of P and
R MNM RNAs in Z3-5578/Z3-3401 spermatocytes. Table gives # and frequency of
each type of MNM cDNA clone from trans-het. (Z3-5578/Z3-3401) male testis RNA and
control (an equimolar mixture of testis RNA from Z3-3401/Df(3R)GC14 and Z3-
5578/Df(3R)GC14 males). Clones were obtained by RT-PCR using an MNM-specific
primer pair. f(P & R) – estimated frequency of P and R RNAs (see Materials and
Methods).

55
TABLE 1
Results of complementation tests between Z3 alleles and mod(mdg4) alleles.
mod(mdg4) Complementation - %X-Y NDJ (N)a
Allele Typeb Locationc Viabilityd Z3-3401e Z3-5578e Z3-3298e
T16h,i Deletion Complete Lethal 39.6 (756) 44.3 (1018) 44.6 (884)
142∆10g Deletion Complete Lethal 49.3 (146) 46 (56) 43.2 (345)
142∆33g Deletion Complete Lethal 49.1 (505) 37.3 (663) 43.5 (726)
eGp4j,k Deletion VR only Semi-let. 2.94 (102) 50.3 (644) 49.0 (251)
B2k Deletion VR only Semi-let. 0 (1772) 37.3 (1311) 40.2 (672)
142∆15g,l Deletion CR only Lethal 42.1 (435) 0.56 (358) 2.34 (1196)
142∆32l,m Deletion CR only Lethal 28.3 (374) 0.95 (420) 1.46 (1096)
142∆29g Deletion CR only Lethal 46 (59) 1.54 (1492) 1.83 (1475)
142∆49g Deletion CR only Lethal 54.9 (304) 0 (371) 0.88 (1248)
Neo129n,o Insert. – P CR-Intron 3 Semi-let. 41.2 (1039) 4.46 (2288) 7.64 (1845)
02n Insert. – gypsy CR-Intron 3 Lethal 32.1 (1361) 1.72 (1918) 2.56 (1599)
03n Insert. – P CR-Exon 1 Lethal 34.9 (417) 0.07 (1438) 0.12 (1709)
20g,p,q NK NK Lethal 45.8 (609) 2.96 (1156) 0.28 (1081)
117g,q NK NK Lethal 47.2 (719) 0.75 (530) 1.55 (1869)
269g,q NK NK Lethal 49.5 (469) 1.44 (139) 2.59 (926)
324g,q Sub. (G92D) CR-BTB Lethal 44.4 (730) 2.84 (2081) 5.06 (1165)
340g,q Sub. (Q177 ) CR-Exon 4 Lethal 54.1 (745) 1.89 (1374) 3.18 (1194)
T6k,q,r Sub. (578 ) 67.2 exon Viable 0 (1113) 0.17 (1736) 0.13 (1492)
Z3-3401 Sub. (R224C) CR-exon 4 Viable 31.2 (724) 1.42 (2113) 1.09 (2848)

56
Z3-5578s Sub. (W449 ) MNM exon Viable 1.42 (2113) 51.2 (633) 45.2 (1836)
Z3-3298s Sub. (H511Y) MNM exon Viable 1.09 (2848) 45.2 (1836) Sterile
a+/BSYy+ males carrying the indicated combination of mod(mdg4) alleles were crossed
with y w females and % X-Y NDJ calculated as described in Materials and Methods. N
= # flies scored. bInsert. – insertion; P – P element; Sub. – base-pair substitution; -
nonsense codon; NK – not known. cCR – common region; VR – variable region; BTB –
BTB/POZ domain; dsemi-let. – semi-lethal. References: eWAKIMOTO et al. 1994;
fMOHLER and PARDUE 1984; gAZPIAZU and FRASCH 1993; hGERASIMOVA and
CORCES 1998; iCHEN and CORCES 2001; jMOHLER and PARDUE 1982;
kGERASIMOVA et al. 1995; lCAI and LEVINE 1997; mFLYBASE; nBUCHNER et al.
2000; oDORN et al. 1993; pGORCZYCA et al. 1999; qGAUSE et al. 2001; rGEYER and
CORCES 1992; sTHOMAS et al. 2005.

57
TABLE 2
Mapping deletion breakpoints in the mod(mdg4) region.
Exon(s) Deletions
Testeda GC14d,e T16f,g 142∆10e 142∆33e 142∆29e 142∆49e B2h eGP4h,i
52.0-56.3b Del. Del. Del. Del. NT NT Del. Del.
59.1 Del. Del. Del. Del. NT NT Del. Del.
59.0, 55.3 Del. Del. Del. Del. NT NT Del. Del.
65.0 Del. Del. Del. Del. NT NT Del. Del.
55.2 NT NT NT NT NT NT NT Not Del.
58.0 NT NT NT NT NT NT Del. Not Del.
Int 58.0/54.7c NT NT NT NT NT NT Del. NT
54.7, 53.6 NT NT NT NT NT NT Not Del. Not Del.
64.2 Del. Del. Del. Del. Not Del. Not Del. Not Del. Not Del.
CR-exon 4 Del. Del. Del. Del. Not Del. Del. Not Del. Not Del.
CR-exon 3 Del. Del. Del. Del. Del. Del. Not Del. Not Del.
aThe indicated mod(mdg4) exons (see DORN and KRAUSS (2003) for nomenclature
and map) were amplified from genomic DNA from deletion/CSiso and deletion/ORiso
flies and sequenced to determine whether the region was present in deletion DNA.
bThis region includes exons 52.0, 53.4, 58.6, 57.4, 59.3, 54.2 and 56.3. cIntron between
exons 58.0 and 54.7. Del. = deleted; NT = not tested. dMOHLER and PARDUE 1984;
eAZPIAZU and FRASCH 1993; fGERASIMOVA and CORCES 1998; gCHEN and
CORCES 2001; hGERASIMOVA et al. 1995; iMOHLER and PARDUE 1982.
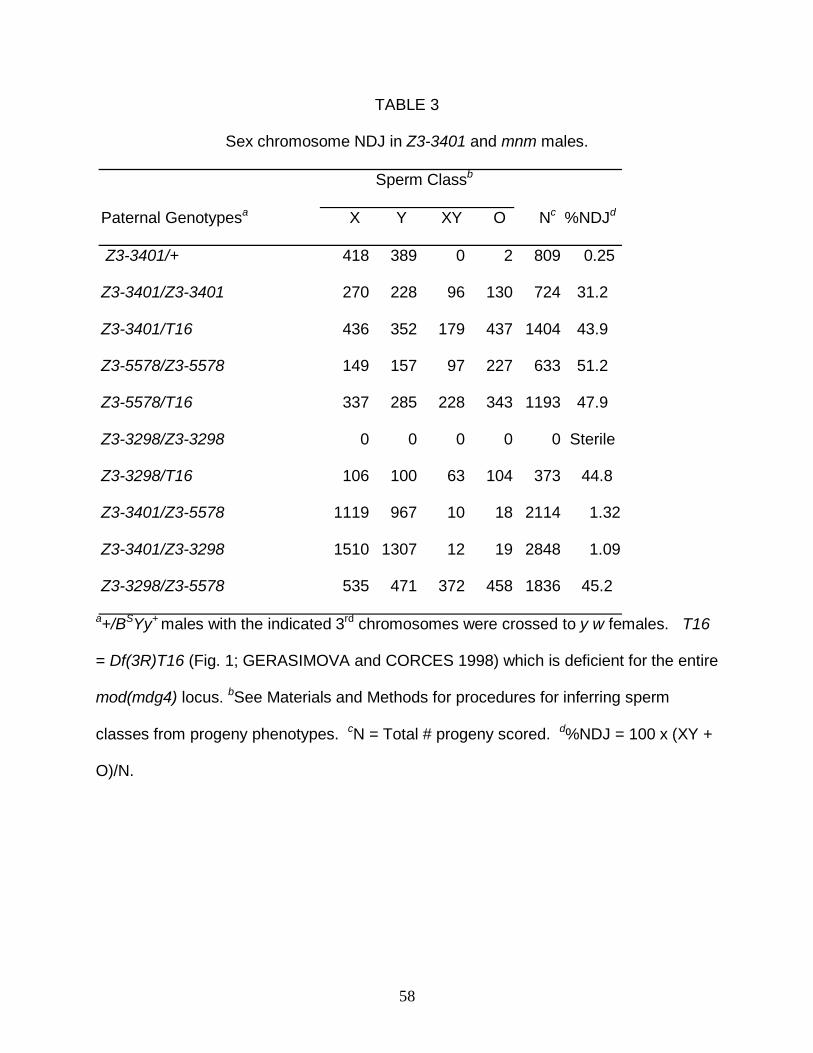
58
TABLE 3
Sex chromosome NDJ in Z3-3401 and mnm males.
Sperm Classb
Paternal Genotypesa X Y XY O Nc %NDJd
Z3-3401/+ 418 389 0 2 809 0.25
Z3-3401/Z3-3401 270 228 96 130 724 31.2
Z3-3401/T16 436 352 179 437 1404 43.9
Z3-5578/Z3-5578 149 157 97 227 633 51.2
Z3-5578/T16 337 285 228 343 1193 47.9
Z3-3298/Z3-3298 0 0 0 0 0 Sterile
Z3-3298/T16 106 100 63 104 373 44.8
Z3-3401/Z3-5578 1119 967 10 18 2114 1.32
Z3-3401/Z3-3298 1510 1307 12 19 2848 1.09
Z3-3298/Z3-5578 535 471 372 458 1836 45.2
a+/BSYy+ males with the indicated 3rd chromosomes were crossed to y w females. T16
= Df(3R)T16 (Fig. 1; GERASIMOVA and CORCES 1998) which is deficient for the entire
mod(mdg4) locus. bSee Materials and Methods for procedures for inferring sperm
classes from progeny phenotypes. cN = Total # progeny scored. d%NDJ = 100 x (XY +
O)/N.
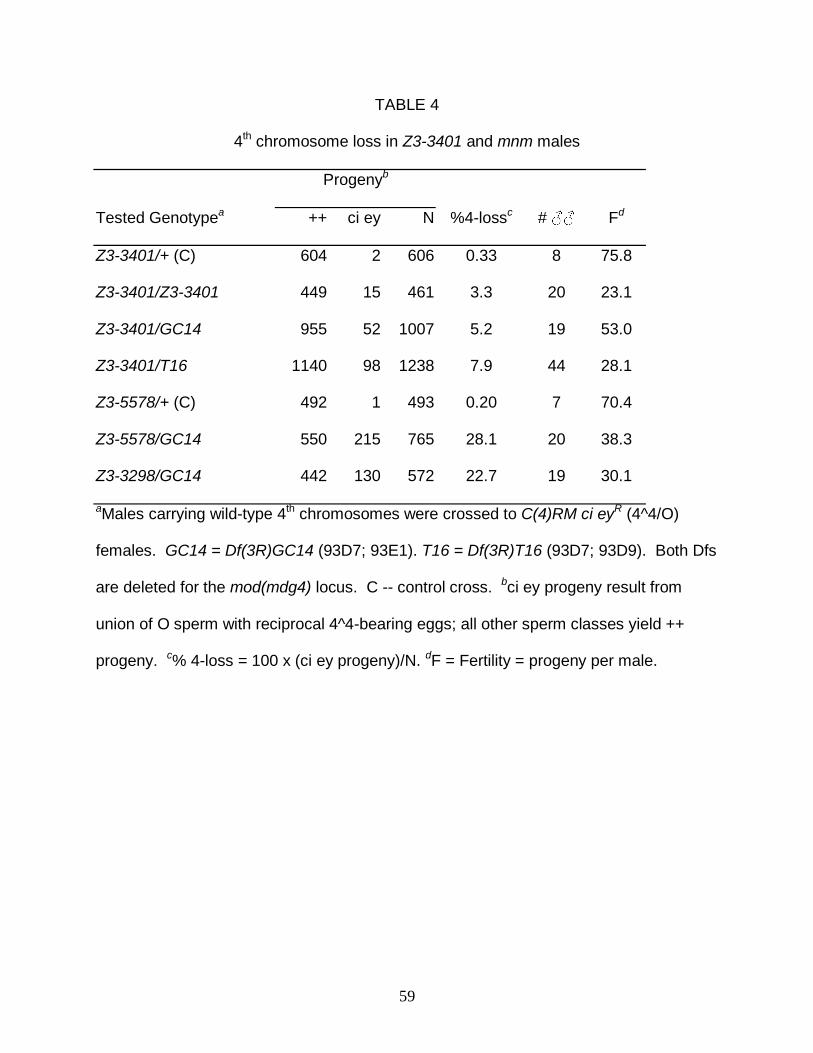
59
TABLE 4
4th chromosome loss in Z3-3401 and mnm males
Progenyb
Tested Genotypea ++ ci ey N %4-lossc # ♂♂ Fd
Z3-3401/+ (C) 604 2 606 0.33 8 75.8
Z3-3401/Z3-3401 449 15 461 3.3 20 23.1
Z3-3401/GC14 955 52 1007 5.2 19 53.0
Z3-3401/T16 1140 98 1238 7.9 44 28.1
Z3-5578/+ (C) 492 1 493 0.20 7 70.4
Z3-5578/GC14 550 215 765 28.1 20 38.3
Z3-3298/GC14 442 130 572 22.7 19 30.1
aMales carrying wild-type 4th chromosomes were crossed to C(4)RM ci eyR (4^4/O)
females. GC14 = Df(3R)GC14 (93D7; 93E1). T16 = Df(3R)T16 (93D7; 93D9). Both Dfs
are deleted for the mod(mdg4) locus. C -- control cross. bci ey progeny result from
union of O sperm with reciprocal 4^4-bearing eggs; all other sperm classes yield ++
progeny. c% 4-loss = 100 x (ci ey progeny)/N. dF = Fertility = progeny per male.

60
TABLE 5
Chromosome 2 NDJ in Z3-3401 and mnm males.
Progenyb Fd
Genotypea bw+ bw b pr Nc #♂♂ 2^2/O 2/2 %NDJe %sis-2f
Z3-5578/+ (C) 3 2 7 12 50 0.24 322.4 0.27 80.0
Z3-5578/GC14 572 1 587 1160 48 21.4 101.0 43.3 0.33
Z3-3298/GC14 434 4 591 1029 46 25.2 110.7 45.0 1.82
Z3-3401/GC14 237 1 416 664 44 15.1 267.9 16.9 0.42
a+/BSYy+; bw/+; Z3/(Df(3R)GC14 or +) males were crossed to C(2)EN, b pr (2^2/O) or y
w (2/2) females. C indicates a control cross. bListed progeny are from 2^2/O crosses.
bw+ and bw progeny result from 22 sperm and O eggs; b pr progeny result from O
sperm and 2^2 eggs. cN= # of progeny scored in 2^2 crosses. dF = progeny per male.
eSee Materials and Methods for procedure for estimating %NDJ. f%sis-2 = % sister
chromatid NDJ/total disomic sperm = 100 x 2(bw)/(bw + bw+).
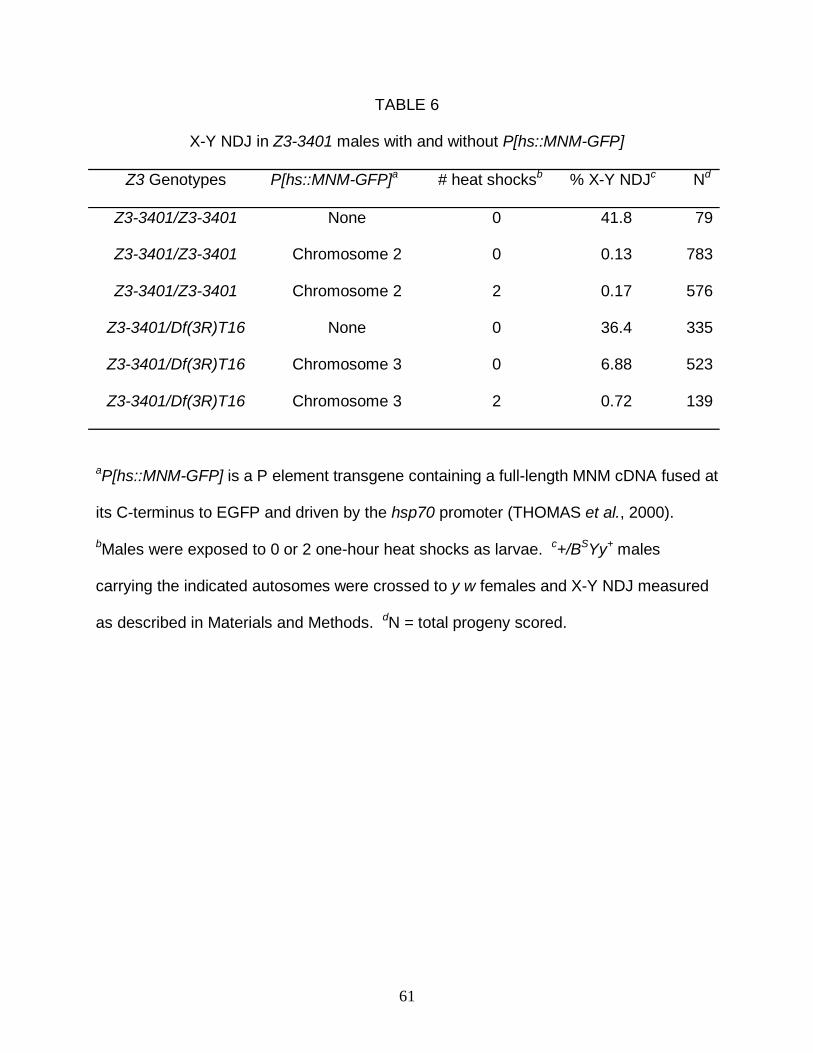
61
TABLE 6
X-Y NDJ in Z3-3401 males with and without P[hs::MNM-GFP]
Z3 Genotypes P[hs::MNM-GFP]a # heat shocksb % X-Y NDJc Nd
Z3-3401/Z3-3401 None 0 41.8 79
Z3-3401/Z3-3401 Chromosome 2 0 0.13 783
Z3-3401/Z3-3401 Chromosome 2 2 0.17 576
Z3-3401/Df(3R)T16 None 0 36.4 335
Z3-3401/Df(3R)T16 Chromosome 3 0 6.88 523
Z3-3401/Df(3R)T16 Chromosome 3 2 0.72 139
aP[hs::MNM-GFP] is a P element transgene containing a full-length MNM cDNA fused at
its C-terminus to EGFP and driven by the hsp70 promoter (THOMAS et al., 2000).
bMales were exposed to 0 or 2 one-hour heat shocks as larvae. c+/BSYy+ males
carrying the indicated autosomes were crossed to y w females and X-Y NDJ measured
as described in Materials and Methods. dN = total progeny scored.

62
TABLE 7
Fertility, X chromosome NDJ, and X, 2nd and 3rd chromosome recombination
frequencies in Z3-3401, mnm and mod(mdg4)T6 hemizygous females.
Fertility, X recombination & NDJa Map Dist. Map Dist.
Genotype #♀♀ Nb Fc %NDJd pn-y+e cn-bw (N)f st-e (N)g
Z3-3401/+ 68 2860 42.1 0 53.8 33.7(409) 24.6(678)
Z3-3401/GC14h,i 117 330 2.8 0 50.5 39.6(191) 23.3(203)
Z3-5578/+ 40 1768 44.2 0.11 57.2 ND 25.5(588)
Z3-5578/GC14h,i 40 1596 39.9 0 58.1 ND 24.6(821)
Z3-3298/+ 40 1638 40.9 0 55.8 34.8(289) 25.1(855)
Z3-3298/GC14h,i 40 1370 34.3 0 53.4 32.9(451) 22.5(672)
mod(mdg4)T6/+ 22 814 37.0 0.49 ND 33.4(511) 26.2(814)
mod(mdg4)T6/GC14h 20 558 27.9 0.72 ND 37.6(388) 30.0(558)
a y/y pn cv m f . y+ females carrying the indicated 3rd chromosomes were crossed with X-
Y, y B/Y males. bN = # progeny scored. cF = Fertility = N/#♀♀. d %NDJ = 100 x 2 x (B+
females + y B males)/(N + B+ females + y B males). eMap length of pn-y+ interval
expressed in cM. fRecombination frequency in the cn-bw interval on chromosome 2
measured in progeny of cn bw/+ females crossed with cn bw males. gRecombination
frequency in the st-e interval of chromosome 3 measured in progeny of Z3i,
st/Df(3R)T16, e females or mod(mdg4)T6 e/Df(3R)GC14, st females crossed with st e
males. hGC14 = Df(3R)GC14. iDf(3R)T16 was used instead of Df(3R)GC14 in the 3rd
chromosome recombination crosses. ND = not done.

63
TABLE 8
X-Y NDJ in Z3-3401 males with and without [CR-7.5].
Z3 Genotypes [CR-7.5]a % X-Y NDJb Nc
Z3-3401/Z3-3401 None 45.5 591
Z3-3401/Z3-3401 Chromosome 2 31.8 1021
Z3-3401/mod(mdg4)neo129 None 42.4 943
Z3-3401/mod(mdg4)neo129 Chromosome 2 33.4 1212
Z3-3401/Df(3R)T16 None 45.8 738
Z3-3401/Df(3R)T16 Chromosome 2 32.5 499
a [CR-7.5] is a P element transgene containing a 7.5Kb genomic DNA fragment
encompassing all of the common region of mod(mdg4) (BUCHNER et al., 2000).
b+/BSYy+ males carrying the indicated autosomes were crossed to y w females and X-Y
NDJ measured as described in Materials and Methods. cN = total progeny scored.

A
Soltani-Bejnood et al., Fig. 1
3’
02neo129
03 Z3-3401
5’
20 117269324340
B2142∆29142∆49
eGp4
Z3-3298
Z3-5578
T6
B T16 ,
CR-7.5
142∆33142∆10,
1 MADDEQFSLC WNNFNTNLSA GFHESLCRGD LVDVSLAAEG QIVKAHRLVL SVCSPFFRKM
61 FTQMPSNTHA IVFLNNVSHS ALKDLIQFMY CGEVNVKQDA LPAFISTAES LQIKGLTDND324 D
121 PAPQPPQESS PPPAAPHVQQ QQIPAQRVQR HEPRASARYK IETVDDGLGD EKQSTTQIVI
181 QTTAAPQATI VQQQQPQQAA QQIQSQQLQT GTTTTATLVS TNKRSAQRSS LTPASSSAGV
340 Stop
Z3-3401 C
241 KRSKTSTSAN VMDPLDSTTE TGATTTAQLV PQQITVQTSV VSAAEAKLHQ QSPQQVRQEE
301 AEYIDLPMEL PTKSEPDYSE DHGDAAGDAE GTYVEDDTYG DMRYDDSYFT ENEDAGNQTA
361 ANTSGGGVTA TTSKAVVKQQ SQNYSESSFV DTSGDQGNTE AQDLGELNPS NLADFGNESF
421 LPKTKGKRPQ NVRCGLAPDQ KCVRTLDDWD RIRYDRTRSG DVLVYDGYRY DRRANYNDIIZ3-5578 Stop
481 YWGCAKKRLS CNVYMITHKN KPTYVAISGV HNHLZ3-3298 Y
142∆15142∆32
Common Region (CR) Variable Region (VR)
GC14
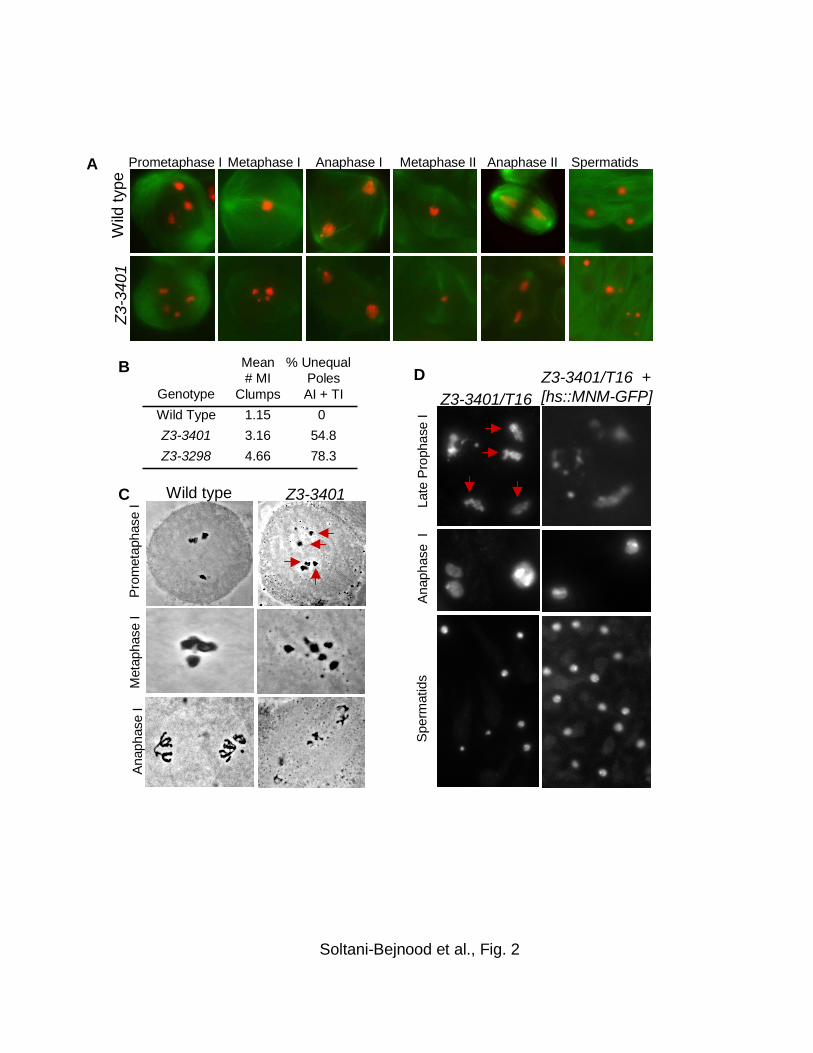
A
B
Pro
met
apha
seI
Ana
phas
e I
Met
apha
se I
Soltani-Bejnood et al., Fig. 2
C
SpermatidsMetaphase I
Z3-
3401
Wild
type
Prometaphase I Anaphase I Anaphase IIMetaphase II
Late
Pro
phas
e I
Ana
phas
e I
Spe
rmat
ids
Z3-3401/T16Z3-3401/T16 + [hs::MNM-GFP]
Z3-3401Wild type
78.34.66Z3-3298
54.83.16Z3-3401
01.15Wild Type
% Unequal PolesAI + TI
Mean # MI
ClumpsGenotypeD

% Paired (N)Gonia Early PI
Wild Type 64%(42) 74%(35)Z3-3401/T16 72%(11) 84%(59)
Wild Type
Wild Type Z3-3401/T16
Z3-3401/T16
A
B
Soltani-Bejnood et al., Fig. 3
Mean MeanNuc. Diam. Disp. N
Wild Type 17.9±2.7 2.8±0.8 24Z3-3401/T16 17.3±3.6 4.4±2.9 9
Wild Type Z3-3401/T16
Spermatogonia Early prophase I (S2a)

Z3-3401 Z3-3401
WT
WT
Z3-3401 Z3-3401
MNM
MNM
SNM
SNM
A
B
WT
WT
Soltani-Bejnood et al., Fig. 4
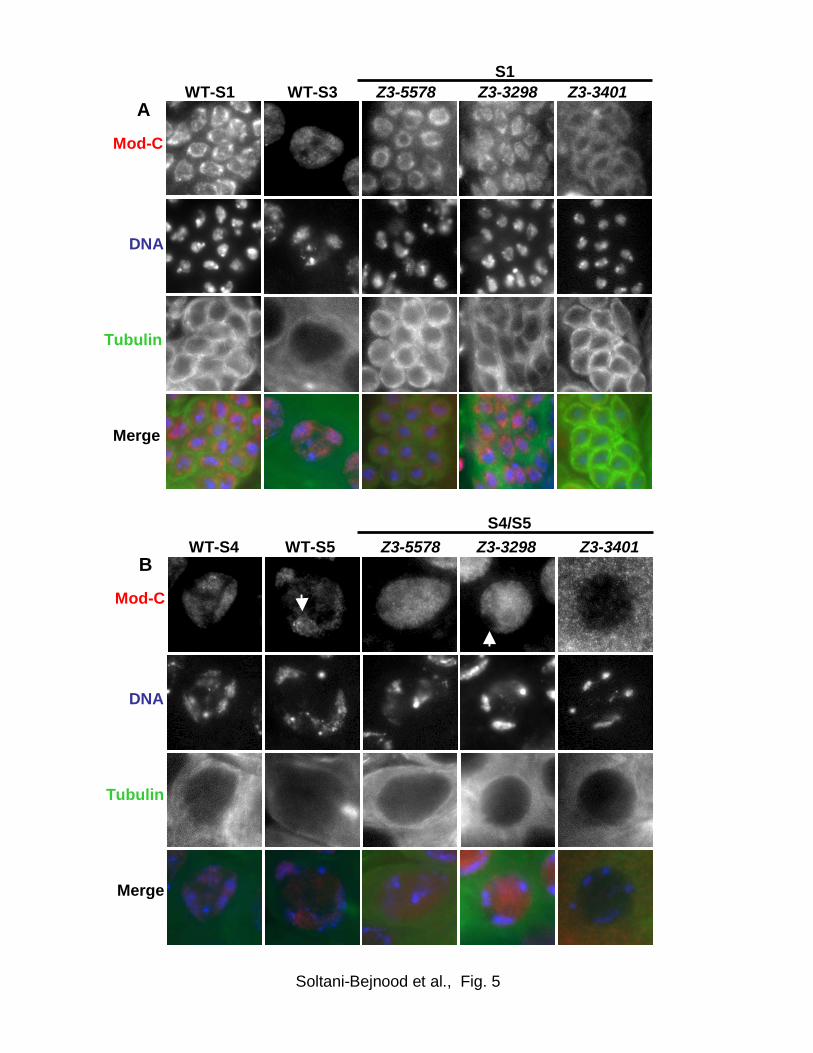
Z3-3401A
Mod-C
DNA
Z3-3298 Z3-5578
Mod-C
DNA
Z3-3298 Z3-3401
DNA
Z3-5578B
Tubulin
WT-S3 WT-S1
Tubulin
WT-S5 WT-S4
Merge
Merge
S4/S5
S1
Soltani-Bejnood et al., Fig. 5

MNM-specific exon
m
m
m
m
mm
m
m
m m
Common exons
Z3-5578
Z3-3401
m
Soltani-Bejnood et al., Fig. 6
P1
m
m
Cis-splicingTrans-splicing
Discontinuous
Discontinuous
Continuous
Continuous
P onlyP & R
B
A
m
+
++
+
+
+ +
+
+
+
+
+
130137Total
RNA source & # of cDNAs of each type
C
9 (.07)29 (.21)R2
R: 0.35+/- .08
12 (.09)26 (.19)R1
42 (.32)29 (.21)P2
P: 0.65+/- .08
67 (.52)53 (.39)P1
f(P&R)ControlTrans-het.cDNA
P2
R1
R2
+
+


















